

Abstract
Isotopic analyses of prehistoric diet have only recently reached the threshold of going beyond site-focused reports to provide regional syntheses showing larger trends. In this work we present the first regional analysis for Neolithic southeastern Italy as a whole, including both substantial original data and a review of the available published data. The results show that dietary isotopes can shed new light on a number of traditional and important questions about Neolithic foodways. First, we observe regional variations in the distribution of stable isotope values across the area, suggesting variability in the Neolithic diet. Secondly, we show that, although the plant food calorific intake was primary for these communities, animal products were also important, representing on average 40% of the total calories. Third, we note that marine fish was only minorly consumed, but that this could be an underestimation, and we observe some variability in the regions considered, suggesting differences in local human–environment interactions. People in different regions of southeastern Italy may have consumed different versions of a common Neolithic diet. Regional synthesis also allows us to take stock of gaps and new directions in the field, suggesting an agenda for Neolithic isotopic research for the 2020s.
Subject terms: Biogeochemistry, Environmental sciences, Environmental social sciences
Introduction
Stable carbon and nitrogen isotope studies of human remains in prehistoric Italy have rapidly increased in the past two decades1–17. Although the isotopic evidence has greatly improved our understanding of dietary practices throughout the peninsula, this is hardly systematic, with some geographical areas and chronological phases dramatically understudied. While this calls for increased research on prehistoric Italy, we here synthesise the existing literature for one of the areas with a higher density of human and animal collagen isotope data: Neolithic southeastern Italy. This provides the chance to have a virtually uninterrupted sequence of information for a relatively limited area, which is of great archaeological interest. Indeed, in Italy, the earliest appearance of domesticated crops and animals outside their natural climatic zone is concentrated in the southeast and corresponds to increased human occupation along the southern Adriatic coast, particularly in the first half of the 6th millennium BCE. There is a general consensus that Neolithic communities in Italy relied on domesticated crops and animals; but while farming appears to be central in the subsistence strategies of such communities, the role of herding and, more importantly, the contribution of marine environments is still to be clearly identified18.
Within a newly acquired subsistence model, could the different contribution of the same food sources to human diet help disentangle specific cultural scenarios? Further, how could local environmental phenomena participate in unravelling said complexity? Italy has a long tradition of cultural diversity. The peninsula has a varied environment, which encompasses different ecoregions, hence it is understandable that food traditions diversified. Further, besides economy and consumption, commensality may have played a role in food choices in the past: something we could associate with the concept of cuisine (sensu Robb18), by which different assortments of foods might have constituted everyday Neolithic meals, regardless of what was available on the menu.
We explore Neolithic foodways through stable carbon and nitrogen analysis of human and animal bone collagen from a large number of sites, of Early, Middle and Late phases. We do so by posing specific questions: (I) how diversified was the Neolithic diet in southeastern Italy? (II) what was the role of animal products in the diet of Neolithic communities? (III) was marine fish consumed by individuals living in proximity to the coast and was its contribution homogeneous? Part of this study stems from an earlier project, hence new data will be coupled with earlier works that were mostly focused on single case studies. Comparative analysis will be provided by published data available for prehistoric Italy.
Carbon and nitrogen stable isotope analysis in the Mediterranean
When it comes to studying dietary habits and mobility patterns of ancient human populations, stable isotope analyses are nowadays the most deployed method, as proven by the exponential increase of articles published in the last fifty years mentioning the words ‘archaeology’ and ‘stable isotopes’19. Stable carbon and nitrogen isotopes in particular are capable of unravelling ecological and social questions linked to food consumption; the working principle consists in detecting the existing differences of isotopic ratios among environments, organisms and their tissues. Isotope results are reported as δ values (δ13C and δ15N) in parts per 1000 or “per mil” (‰). Thanks to how carbon fractionates depending on different photosynthetic pathways and in different ecosystems, δ13C (‰) values can be used to discriminate between C3 (~ − 28‰), C4 (~ − 14‰) and CAM (~ − 11‰) plants and their consumers20, as well as between terrestrial (~ − 7‰) and marine (~ 0‰) or freshwater (~ − 15‰) ecosystems21. On the other hand, δ15N (‰) values are mostly affected by climatic and environmental conditions and they are mainly used as a trophic level indicator22,23.
Once plants are eaten by animals, the carbon and nitrogen atoms composing plant tissues are used by the animals to build up their own tissues through biochemical reactions. It has been observed that both carbon and nitrogen are enriched in their heavier isotope at each step of the food web and this effect is used to assess the relative contribution of different food categories to the diet of past human individuals, also using quantitative approaches. It was initially noted that δ13C (‰) values of animals are enriched by + 1‰ compared to their diet24. However, subsequent feeding experiments and controlled field studies have shown more diversified offsets from diet to consumer tissues (Δ13Ctissue−diet), ranging from + 0.5 to + 7.5‰25–31 depending on diet composition and on the fact that carbon is "scrambled" from all parts of the diet (i.e., carbohydrates, lipids and protein)32. On the contrary, the δ15N (‰) values of bone collagen are directly linked to protein consumption and therefore Δ15Ncollagen−diet offset values are not affected by total dietary composition. As a result, feeding experiments have shown much less variable Δ15Ncollagen−diet offset values compared to carbon, suggesting an overall Δ15Ncollagen−diet value of + 3.4 ± 1.0‰25,30,33–37. However, a controlled dietary study on humans38 proposed a higher Δ15Ncollagen−diet offset of ca. + 6‰.
On the basis of these principles, marine fish consumption can easily be detected using stable isotope analysis. However, the usual discriminants do not apply well in Mediterranean contexts. Indeed, archaeological marine fish samples from the Mediterranean have reported much lower δ15N (‰) values compared to those of marine fish from the Atlantic, often closer to those of terrestrial animals or manured cereals from the same area39,40, although the reasons behind this difference are still to be explored. Mean δ13C (‰) values might also be different in the Mediterranean compared to other areas of the world. It is important to note that the ancient Mediterranean coast was largely characterised by marshy lagoons (perhaps 6500 km2) and, in terms of food exploitation, they are twice as productive as the open sea41. Freshwater discharges present in coastal lagoons and wetlands determine more 13C-depleted isotope values, as observed in the fish living in their proximity42. Therefore, it is possible that more negative δ13C (‰) values of the humans from some of the coastal sites are influenced by the consumption of brackish fish with 13C-depleted isotope values caused by freshwater inputs. More negative δ13C (‰) values in the prehistoric Mediterranean were found for example in some marine fish8,43. The amount of fish consumed might be partially masked by the C3 terrestrial signal and therefore underestimated.
Results
We collected human and animal bone samples from a variety of Neolithic sites from southeastern Italy (Fig. 1). Archaeological information for each site studied here is provided in the Supplementary Information S1.
Figure 1.

Location of the archaeological sites with human individuals analysed for stable carbon and nitrogen isotope analysis. Blue crosses indicate sites from this study, red crosses indicate sites from the literature. 1: Samari; 2: Serra Cicora; 3: Carpignano Salentino; 4: Torre Castelluccia8; 5: Grotta Funeraria1; 6: Trasano; 7: Tirlecchia; 8: Malerba-Altamura; 9: Grotta delle Mura8; 10: S. Barbara; 11: Diga di Rendina; 12: Balsignano; 13: Masseria Stevanato; 14: Madonna delle Grazie; 15: Cala Colombo; 16: Cala Scizzo; 17: Masseria Maselli8; 18: Titolo-Palese; 19: Palata8; 20: Foggia; 21: Ripa Tetta; 22: Fonteviva44; 23: Passo di Corvo; 24: Diga di Occhito; 25: Masseria Candelaro45; 26: Grotta Scaloria14,45; 27: La Torretta—Poggio Imperiale; 28: Lama dei Peligni; 29: Fossacesia; 30: Catignano. Some of the sites from this study were also previously investigated by other authors and these are: Balsignano, Ripa Tetta and Samari8 and Passo di Corvo45.
This section presents the new isotopic data, which will be contextualised using existing data in the following discussion section. New data comprises 94 human samples and 16 animal samples of various wild and domesticated species (namely, Bos sp., Sus sp., Ovis vel Capra, Cervus sp. and Capreolus sp.) (Table S1). In the following analysis, only samples with C:N ratio between 2.9 and 3.6 are included46. Out of the total, this limits our results to 88 human samples and 15 animal samples, with mean δ13C values of − 19.6 ± 0.5‰ and δ15N values of 9.0 ± 1.1‰ for the humans and mean δ13C values of − 20.6 ± 0.7‰ and δ15N values of 6.4 ± 1.0‰ for the animals. Table 1 reports descriptive statistics of the data that passed the quality check presented here and of those previously published used for comparative purposes. Previously published data are reported in Table S2.
Table 1.
Descriptive statistics of the human remains analysed in this study and those from previously published work.
| Site | n | mean δ13C | median δ13C | 1SD δ13C | min δ13C | max δ13C | mean δ15N | median δ15N | 1 SD δ15N | min δ15N | max δ15N | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Balsignano | 2 | − 19.3 | − 19.3 | 0.1 | − 19.3 | − 19.2 | 9 | 9 | 1.2 | 8.1 | 9.8 | This study; Lelli et al. 20128 |
| Cala Colombo | 6 | − 19.5 | − 19.4 | 0.4 | − 20.2 | − 19.3 | 9.2 | 9.4 | 1.2 | 7 | 10.6 | This study |
| Cala Scizzo | 1 | − 19.9 | 7.6 | This study | ||||||||
| Carpignano Salentino | 1 | − 19.7 | 9.3 | This study | ||||||||
| Catignano | 3 | − 20.5 | − 20.1 | 0.9 | − 21.5 | − 19.9 | 7.3 | 7.5 | 1.5 | 5.7 | 8.7 | This study |
| Diga di Occhito | 9 | − 19.9 | − 20.2 | 0.4 | − 20.4 | − 19.3 | 9 | 8.8 | 0.7 | 8.4 | 10.8 | This study |
| Diga di Rendina | 8 | − 19.7 | − 19.7 | 0.2 | − 20 | − 19.4 | 9.4 | 9.3 | 0.4 | 9.1 | 10.2 | This study |
| Fossacesia | 1 | − 19.2 | 9.6 | This study | ||||||||
| La Torretta–Poggio Imperiale | 4 | − 19.4 | − 19.4 | 0.2 | − 19.6 | − 19.1 | 8.4 | 8.6 | 0.8 | 7.2 | 9.1 | This study |
| Lama dei Peligni | 1 | − 19.1 | 12.3 | This study | ||||||||
| Madonna delle Grazie | 4 | − 19.6 | − 19.5 | 0.4 | − 20.2 | − 19.2 | 9.5 | 9.6 | 0.7 | 8.8 | 10.1 | This study |
| Malerba–Altamura | 1 | − 19.1 | 8.7 | This study | ||||||||
| Masseria Stevanato | 2 | − 20.2 | − 20.2 | 1.3 | − 21.1 | − 19.2 | 7.3 | 7.3 | 2.8 | 5.3 | 9.3 | This study |
| Passo di Corvo | 15 | − 19.1 | − 19.1 | 0.2 | − 19.5 | − 18.8 | 8.9 | 8.7 | 0.7 | 7.9 | 10.4 | This study; Tafuri et al. 201445 |
| Ripa Tetta | 5 | − 19.9 | − 20 | 0.2 | − 20.1 | − 19.6 | 10.4 | 10 | 1.1 | 9.3 | 11.7 | This study; Lelli et al. 20128 |
| S. Barbara | 2 | − 19.2 | − 19.2 | 0.6 | − 19.6 | − 18.7 | 9 | 9 | 2.4 | 7.3 | 10.7 | This study |
| Samari | 11 | − 19.1 | − 19.1 | 0.4 | − 19.8 | − 18.5 | 9.8 | 9.8 | 0.6 | 8.4 | 10.5 | This study; Lelli et al. 20128 |
| Serra Cicora | 16 | − 19.4 | − 19.4 | 0.5 | − 20.6 | − 18.7 | 9.2 | 9.3 | 0.9 | 7.2 | 10.4 | This study |
| Tirlecchia | 4 | − 19.6 | − 19.6 | 0.1 | − 19.7 | − 19.5 | 9.1 | 9.1 | 0.2 | 8.9 | 9.3 | This study |
| Titolo-Palese | 11 | − 19.6 | − 19.5 | 0.4 | − 20.4 | − 19.1 | 8 | 8 | 0.7 | 6.3 | 9 | This study |
| Trasano | 7 | − 19.7 | − 19.8 | 0.2 | − 20 | − 19.3 | 9.2 | 9.3 | 0.5 | 8.4 | 9.9 | This study |
| Fonteviva | 2 | − 19.4 | − 19.4 | 0.1 | − 19.4 | − 19.3 | 10 | 10 | 0.4 | 9.7 | 10.2 | Parkinson and McLaughlin 202044 |
| Grotta dell'Antenato | 1 | − 19.2 | 10.8 | Arena et al. 20201 | ||||||||
| Grotta delle Mura | 2 | − 18.7 | − 18.7 | 1.3 | − 19.6 | − 17.7 | 7.9 | 7.9 | 0.1 | 7.8 | 8 | Lelli et al. 20128 |
| Grotta Funeraria | 6 | − 19.5 | − 19.5 | 0.3 | − 19.9 | − 19.1 | 9.8 | 9.8 | 0.2 | 9.5 | 10.2 | Arena et al. 20201 |
| Grotta Scaloria | 45 | − 19.3 | − 19.3 | 0.3 | − 19.9 | − 18.9 | 8.4 | 8.4 | 0.7 | 6.8 | 10.6 | Tafuri et al. 201445; Tafuri et al. 201716 |
| Masseria Candelaro | 23 | − 19.2 | − 19.2 | 0.3 | − 19.9 | − 18.2 | 9.3 | 9.2 | 0.8 | 7.9 | 11.4 | Tafuri et al. 201445 |
| Masseria Maselli | 1 | − 19.6 | 8.1 | Lelli et al. 20128 | ||||||||
| Palata | 2 | − 19.3 | − 19.3 | 0.3 | − 19.5 | − 19.1 | 9.1 | 9.1 | 0.7 | 8.6 | 9.6 | Lelli et al. 20128 |
| Torre Castelluccia | 1 | − 18.8 | 8.4 | Lelli et al. 20128 |
We explored the new data in relation to previously published domestic and wild fauna8,45, and present them below distinguishing between Early, Middle and Late Neolithic phases (Fig. 2).
Figure 2.

Early (a) Middle (b) and Late (c) Neolithic human individuals and faunal remains analysed for the present study. Early (a), Middle (b) and Late (c) Neolithic animal remains and marine fish from Italy from previous studies8,45 were included as mean values. Bars represent 1SD.
Early Neolithic (ca. 6000–5500 BCE)
In Fig. 2a are reported δ15N (‰) and δ13C (‰) values of Early Neolithic samples. 18 human individuals were successfully analysed and 1 animal sample tentatively identified as Bos from Diga di Rendina. The most 13C-depleted sample is the Early Neolithic adult female from Titolo-Palese (T9, − 19.8‰), followed by the 8 individuals from Diga di Rendina (mean δ13C = − 19.7 ± 0.2‰), the adult female from Ripa Tetta (IT27, − 19.6‰) and by the adult female from Lama dei Peligni (LdP, − 19.1‰). Finally, the individuals from Samari present the highest δ13C mean value of the Early Neolithic group (mean δ13C = − 19.1 ± 0.4‰). As for nitrogen, Lama dei Peligni exhibits the highest δ15N value (12.3‰), followed in descending order by Samari (mean δ15N = 9.7 ± 0.7‰), Ripa Tetta (IT27, 9.6‰), Diga di Rendina (mean δ15N = 9.4 ± 0.4‰) and Titolo-Palese (T9, 8‰).
Middle Neolithic (ca. 5500–4300 BCE)
The Middle Neolithic sample is the largest dataset for human individuals (n = 48) (Fig. 2b). One individual from Serra Cicora (SC23) cannot be assigned to either the Middle or the Late phase of the Neolithic due to a lack of contextualisation. This individual is here included for visualisation but excluded from the following descriptive statistics. Fourteen associated animal samples were also successfully analysed, namely one probable Bos from Catignano, 3 (Sus, Bos and Ovis vel Capra) from La Torretta—Poggio Imperiale and 10, ranging between Bos, Cervus, Sus species and Ovis vel Capra, from Serra Cicora.
The highest δ13C values are found at Passo di Corvo (n = 2, mean δ13C = − 19.2 ± 0.1‰), followed in descending order by Balsignano (BA8, − 19.2‰), the 4 individuals from La Torretta—Poggio Imperiale (mean δ13C = − 19.4 ± 0.2‰), 13 of the individuals from Serra Cicora (mean δ13C = − 19.4 ± 0.5‰), the 10 Middle Neolithic individuals from Titolo-Palese (mean δ13C = − 19.6 ± 0.4‰), the 4 individuals from Tirlecchia (mean δ13C = − 19.6 ± 0.1‰), the two individuals from Malerba-Altamura (mean δ13C = − 19.6 ± 0.7‰), by the seven individuals from Trasano (mean δ13C = − 19.7 ± 0.2‰), by the individual from Carpignano Salentino (SC28, − 19.7‰) and finally by Catignano (n = 3, mean δ13C = − 20.5 ± 0.9‰). The lowest δ15N values are found at Catignano (mean δ15N value = 7.3 ± 1.5‰) and these are followed by Malerba-Altamura (mean δ15N value = 7.8 ± 1.3‰), Titolo-Palese (mean δ15N value = 8.0 ± 0.8‰), La Torretta-Poggio Imperiale (mean δ15N value = 8.4 ± 0.8‰), Passo di Corvo (mean δ15N value = 8.6 ± 0.0‰), Tirlecchia (mean δ15N value = 9.1 ± 0.2‰), by Trasano (mean δ15N value = 9.2 ± 0.5‰), Serra Cicora (mean δ15N value = 9.3 ± 1‰), by the individual from Carpignano Salentino (SC28, 9.3‰) and finally by Balsignano (BA8, 9.8‰).
Late Neolithic (ca. 4300–3800 BCE)
From the Late Neolithic, 27 human individuals were successfully analysed (Fig. 2c). Unfortunately, no animal remains were available for this dataset and, to our knowledge, fauna dated to the Late Neolithic is also lacking in the literature. The lowest δ13C values are found at Masseria Stevanato (n = 2, mean δ13C = − 20.2 ± 1.3‰) followed in ascending order by Diga di Occhito (n = 9, mean δ13C = − 19.9 ± 0.4‰), the sub-adult from Cala Scizzo (BA21, − 19.9‰), Madonna delle Grazie (n = 4, mean δ13C = − 19.6 ± 0.4‰), Cala Colombo (n = 6, mean δ13C = − 19.5 ± 0.4‰), the female adult from Fossacesia (IT8, − 19.2‰), the two Late Neolithic individuals from Serra Cicora (mean δ13C = − 19.2 ± 0.3‰), and finally by S. Barbara (n = 2, mean δ13C = − 19.2 ± 0.6‰). The highest δ15N values exhibited by Fossacesia (9.6‰) followed in descending order by Madonna delle Grazie (mean δ15N = 9.5 ± 0.7), Serra Cicora (mean δ15N = 9.3 ± 0.1), Cala Colombo (mean δ15N = 9.2 ± 1.2‰), S. Barbara (mean δ15N = 9.0 ± 2.4‰), Diga di Occhito (mean δ15N = 9.0 ± 0.7‰), Madonna delle Grazie (mean δ15N = 8.8 ± 1.6‰), Cala Scizzo (7.6‰) and finally by Masseria Stevanato (mean δ15N = 7.3 ± 2.8‰).
Discussion
Cucina tipica in the Neolithic?
Even when people in a region share a common tradition for producing foods, and a common repertory of ingredients and techniques for preparing them, there can be strong local differences in how they eat. For example, recent historic cuisine in peninsular Italy is based on a limited, common range of resources (grains, vegetables, animal products such as cheese, pork and beef) with some regional variations (notably, the availability of fish, and the use of different grains suitable for different local climates). In spite of this, there are strong regional traditions of cucina tipica, or local cuisine, which combine and emphasise these in distinctive ways. Roman writers suggest that there were regional variations in foodways in Classical times, and as food is an important part of the habitus and the reproduction of communities, there is no reason to suppose that it was otherwise in prehistoric periods. Much of regional foodways may be invisible archaeologically, though we might perhaps see local patterns of material culture (e.g. pottery forms), animal remains (e.g. butchery patterns), or food consumption.
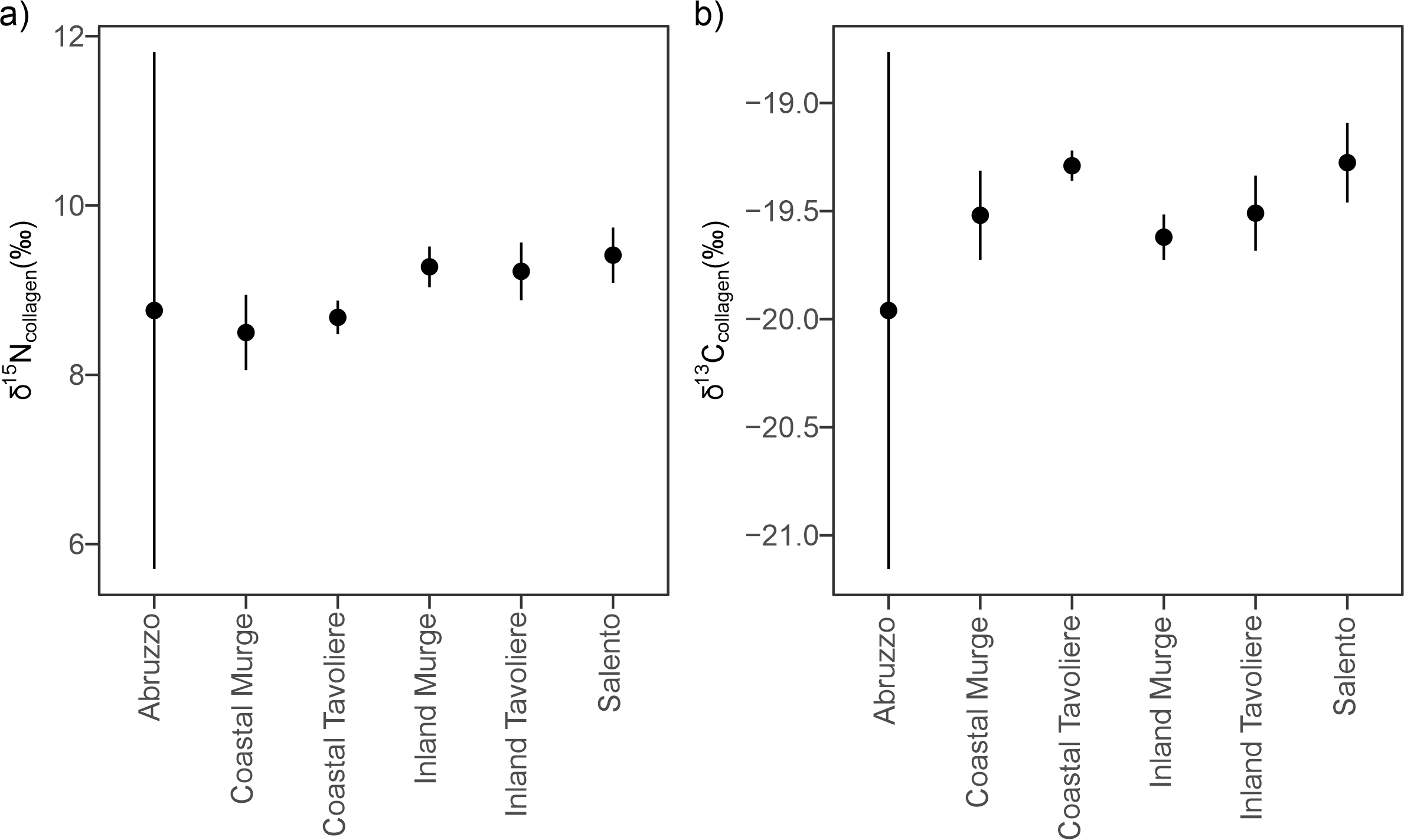
The only area of Italy where the isotopic data for the Neolithic are dense enough to examine is southeastern Italy, including Puglia and the adjacent regions of Basilicata and Abruzzo. To evaluate regional differences, we explored the variability of δ13C (‰) and δ15N (‰) of humans throughout the southeastern area of the Peninsula, coupling data reported here with those of published works. The data were elaborated using the Bayesian model AverageR (see Materials and Methods for a description47,48, and the outputs presented in Fig. 3a,b. Terrestrial animals δ13C (‰) and δ15N (‰) values were also explored as these most likely reflect the local environment and therefore can help in the discussion of the human data. Models for the animals are reported in Fig. 3c,d. Both δ15N (‰) and δ13C (‰) values in animals do not differ according to study area (Fig. 3c,d). This could suggest that the difference in climatic and environmental conditions from one area to another is not so marked to be reflected in the isotopic values of animals, also according to modern climatic data49. However, the animal assemblage is limited considering the area of investigation, therefore the evidence is not strong enough to exclude climatic or environmental variations at this stage. On the contrary, the human δ15N (‰) and δ13C (‰) values appear to vary slightly but significantly from one region to another. The differences visible from the maps were also explored using descriptive statistics (Table S3), plots with mean values and associated 95% CIs (Fig. S1) and pairwise comparisons using Wilcoxon rank sum test (Table S4)50. Inland Murge and coastal Tavoliere differ by their mean values, as well as Salento from the coastal Murge and from the coastal Tavoliere (Fig. S1, Tables S3 and S4, p-values < 0.05). Higher δ15N (‰) values in the inland Murge and in the inland Tavoliere could suggest a slightly higher consumption of animal products in these regions; in fact, a stronger reliance on animals in the uplands of the Murge has been already discussed51. The δ15N (‰) values from Salento are higher than those from the coastal areas of the Murge and the Tavoliere. This could suggest higher consumption of marine products compared to other sites, again close to the sea, but from a different area. As for the distribution of δ13C (‰) values, differences are found between the coastal Murge and the coastal Tavoliere, as well as between the coastal Tavoliere and the inland Murge and between the inland Murge and the Salento area (Fig. S1, Tables S3 and S4, p-values < 0.05). The coastal Tavoliere and the Salento areas have higher mean δ13C (‰) values, which is not surprisingly observed for the coastal Murge (Fig. 3b). Again, this suggests that although these Neolithic communities had equal access to marine resources, this did not translate into similar dietary habits.
Figure 3.

Maps with the distribution of stable nitrogen (a,c) and carbon (b,d) isotope data from Neolithic southeastern Italy, of humans (a,b) and animals (c,d), created using the app IsoMemo, tool AverageR (https://isomemoapp.com/app/iso-memo-app). Data comes from this study and from Lelli et al.8, Tafuri et al.45, Arena et al.1 and Parkinson and McLaughlin44.
As expected, more negative values are found inland, supporting the lack of marine fish consumption. An explanation for the differences in δ13C (‰) values in the three different coastal areas, namely the Tavoliere, the Murgian coast and Salento could be found in the natural environment. The area of the Tavoliere and of Salento are characterised by extensive wetlands and coastal lagoons52–54. As already mentioned, wetlands are more productive than the open sea41 and they are also more easily accessible, therefore representing an important potential source of subsistence for those living in these landscapes. On the contrary, the Bari coast (coastal Murge) comprised a more limited marsh area52, which probably resulted in the open sea being the only viable option for the procurement of aquatic resources. Curiously, the coastal area of the Tavoliere which reports among the highest δ13C (‰) values among the areas explored (Fig. 3b), where extensive marshes are present, is also one of those that show the lowest δ15N (‰) values (Fig. 3a). This seems to suggest that the fish consumed by these local communities is most likely composed of low trophic level organisms, such as benthic bivalves, which are abundant in estuaries and lagoons55–57.
The isotopic evidence suggests small variation of a common Neolithic diet. One possible reason might include proximity to the sea, estuaries and lagoons. Another might include the social role of food in different kinds of communities, for instance among the large groups of the ditched villages of the Tavoliere and of the inland Murge that may have held large social gatherings involving meat consumption (e.g. feasts) more often than more dispersed, smaller communities did. This is a preliminary hypothesis; we would suggest validating it by further investigation, including new isotope data from animals that could confirm that the differences we are observing are not related to environmental and climatic factors, and by integration with archaeological evidence of other kinds (e.g. pottery repertories; animal bones including fish and shellfish; botanical remains; food cooking facilities on sites; lipid analysis).
The role of animal products in the diet
One of the main issues in the discussion of the Neolithic diet is the role of animals in the food economy. Neolithic people in Italy, and throughout much of Europe, had a suite of domesticated animals, including cattle, pigs, sheep, goats, and dogs (though dogs seem to have been eaten rarely). They also had access to wild game, including deer, wild pig, and small animals. But how much did these contribute to their diet? Did they eat large quantities of animal products, or were they mostly subsisting on grains and vegetables, with meat reserved mostly for social occasions? Moreover, recent studies are suggesting that secondary animal products were important in the Neolithic Mediterranean basin58,59.
To estimate in calorific terms the contribution of animal products to the diet of Neolithic southeastern Italian groups, we applied a Bayesian mixing model to the isotopic data (see Materials and Methods for the description of the model). It was necessary to exclude marine fish contribution from the estimations on inland sites since there is no evidence of fish consumption from these sites (i.e., no access to marine resources), therefore results are here divided in two groups, reported in Fig. 4a,b for inland and coastal sites, respectively. On average, animal products contributed to around 40% of the total calories in the diet of Neolithic individuals from inland and coastal sites (Table S5 and Fig. 4). The results suggest that animal products were not only available but also easily accessible during the Neolithic in southeastern Italy. Therefore, cereals, although contributing significantly, were not the only source of energy for these early farmers, as our models show. Our estimates do not differ from the conclusions of other studies that explore the Neolithic diet in other parts of Europe. For example, using stable isotope analysis of human bone collagen, it was observed that the majority of the diet of Neolithic communities in England and Scotland have an animal origin60–62 and considerable consumption of animal proteins was suggested in Neolithic Switzerland63, Germany64–66, Hungary67, north and central France68–70, Spain71,72, Greece73 and Croatia74. Recently, animal products have been estimated to contribute more than 50% of the diet in Neolithic Malta, probably due to a high reliance on dairy75.
Figure 4.

Calorific contribution (%) of terrestrial animal products in Neolithic individuals from southeastern Italy divided by sites, inland (a) and coastal (b). Boxes represent a 68% credible interval (corresponding to the 16th and 84th percentiles) while the whiskers represent a 95% credible interval (corresponding to the 2.5th and 97.5th percentiles). The horizontal continuous line represents the median (50th percentile) while the horizontal discontinuous line represents the mean.
Unfortunately stable isotopes of bone collagen cannot be used to discriminate between different animal products (in this case, meat and dairy). To answer this question other types of analyses and other kinds of material need to be studied, for example the carbon isotopes of lipids from ceramics, and age at death profiles of animal remains at each site. Differences can be outlined between sites in some cases. For example, the diet of a female from Lama dei Peligni, one of the earliest Neolithic individuals recovered in Italy, is composed of ~ 55% of calories of animal origin, probably still influenced by the earlier Mesolithic tradition (Fig. 4a). This signifies that the typical ‘Neolithic diet’ was not spread uniformly in the area and that cultural and/or social variables might have influenced these communities, as also suggested by the regional analysis. We should acknowledge however the large uncertainties associated with these estimates since the two sources used in the model, C3 cereals and C3-feeding animals, show very similar isotopic values (Table S5 and Fig. 4). Analyses at a higher resolution, such as Compound Specific Stable Isotope Analysis of Amino Acids (CSIA-AA) are needed in order to confirm and better define this scenario76. We should also remember that by analysing δ13C (‰) and δ15N (‰) values we are mainly exploring the protein component of diet77. This limits our understanding of plant consumption (notably cereals and legumes) where proteins are low, but they represent the staple for Neolithic people. To explore plant consumption, different types of analysis are needed, for example carbon isotope analysis of apatite78–80.
Use of marine resources?
A major puzzle for prehistoric food use is whether Neolithic people were using marine sources. An increasing number of isotopic studies from around Europe show that, even when they lived coastally in rich marine environments, Neolithic people lived principally on terrestrial resources62,74,81–85. This was a major change from Mesolithic lifeways: overall, isotopic studies have demonstrated that a gradual decline in the exploitation of marine resources at the transition to the Neolithic is observed for the Mediterranean82,86–88, with fishing becoming more sporadic and mostly opportunistic89. It cannot be ignored that marine protein consumption is believed to be limited in the Mediterranean already in the Mesolithic compared to their Atlantic and Baltic counterparts9,10,86,90. However, a recent study using a higher resolution approach (CSIA-AA) has shown that aquatic resources were indeed exploited91. This gradual decline at the transition to the Neolithic may be ascribed to cultural practices or organisational factors83,85,92. Food procurement from an open sea environment clearly entailed greater risk and might have resulted in limited exploitation. As discussed by Galili et al.93, it might be that “fishing was a low preference mode of production, to which Neolithic communities turned only once the quantity and/or quality of terrestrial resources were reduced or impaired”. To explore the question in Neolithic southeastern Italy, we plotted all the data according to proximity to the coast, since both δ13C (‰) and δ15N (‰) would increase values according to the accessibility of marine resources (Fig. 5). What we observe is no substantial differences when we explore data in a chronological perspective (Fig. 5a) and a lack of correlation between both δ13C (‰) and δ15N (‰) and distance to the coast (km) (Fig. 5b,c, R2 = 0.16 and R2 = 0.089, respectively) indicating that marine environments were not exploited substantially and equally by all the communities living in their proximity. The Bayesian mixing model suggests an overall low marine contribution, with some nuance at an inter-site level (Fig. S2 and Table S5). We should consider however that the marine fish consumed in those sites characterised by a lagoon environment could present more 13C-depleted δ13C (‰) values compared to the marine fish specimens used in the model (which were the only available for the area of investigation and with a different chronology), and therefore the marine contribution is possibly underestimated. In general, what emerges is a certain degree of variability in the consumption of marine resources at coastal sites from Neolithic southeastern Italy.
Figure 5.

Scatterplot of δ13C (‰) and δ15N (‰) values of Early (circle), Middle (square) and Late (diamond) individuals also differentiating between coastal (orange) and inland (light blue) sites; terrestrial animals from this study and the literature8,45 are also plotted (crosses) as well as marine fish from the Adriatic8 (a). Correlation between δ13C (‰) and δ15N (‰) and distance to the coast (km) is explored in (b) and (c), respectively. The grey bands represent the 95% confidence interval bands. Data plotted comes from this study and from1,8,44,45.
Conclusion
This paper has shown the distinctive value of the analysis of a large isotopic dataset, combining new and previously published results, to evaluate regional patterns of food consumption. First, we observed regional variations throughout southeastern Neolithic Italy that are most likely not directly caused by climatic or environmental conditions but rather by how the humans adapted according to the local environment and climate. This suggests the possibility of regional variations in the Neolithic diet, even within a relatively small region with a homogeneous cultural background94. Second, Bayesian statistics was used to evaluate the contribution of animal products in the diet of Neolithic people from the area. The results suggest a certain degree of variability across the assemblage and that animal products were certainly not of minor importance in the diet (40% on average of the total calories have an animal origin), implying that cereals and legumes were not the only source of energy for Neolithic communities from southeastern Italy. Finally, we explored marine fish consumption in the area. Our dataset suggests that marine resources were only minorly exploited but that this is likely an underestimation. Some variability in marine fish consumption was observed, and this might be related to differences in the environment (the presence of lagoons makes it easier to access fish) or cultural preferences.
These results are preliminary and need to be reinforced by new material (plant and fish remains in particular, but also more terrestrial animals) and by higher resolution analyses, such as CSIA-AA, which has the potential to better define the differences observed, discriminate and quantify the contribution of terrestrial animals, plants and marine fish in the diet of Neolithic communities76. Nonetheless, a general picture of ‘local’ traditions within a larger model appears to be confirmed8,45. People adopted diversified cuisines, according to a shared habitus, that might have been specific to a network of sites in small areas (e.g., the Murge region or the Tavoliere and Salento). The heterogeneity of dietary habits between communities living in similar environments suggests that socio-cultural factors might have played a key role in food choices and/or farming and herding practices, regardless of local availability.
Materials and methods
Collagen extraction
For the majority of the samples, collagen extraction for isotopic analysis followed the Longin's95 method. In brief, cortical bone (ca. 0.5 g) was cleaned by sand abrasion and demineralized in 0.5 M aq. HCl at 4 °C for several days. The samples were then rinsed to neutral pH. Due to the poor preservation of the human remains from Titolo-Palese, ethylenediaminetetraacetic acid (EDTA) was used instead of HCl following Tuross96. These samples were decalcified in 50 mL 0.5 M EDTA at pH 7.4 for one to four weeks, changing the solution every 4 days. Once decalcified, samples were washed with deionised water 14 times, and left in water overnight. All samples were gelatinized in pH 3 water at 75 °C for 48 h. The collagen solution was filtered off with 5–8 μm Ezee filters, then frozen, and freeze dried. Stable carbon and nitrogen isotope ratios were measured using an automated elemental analyser coupled in continuous-flow mode to an isotope-ratio-monitoring mass-spectrometer. The majority of the analyses were carried out at the Godwin Laboratory, University of Cambridge. Instrument used was a Costech Elemental Analyzer coupled to a Thermo Finnigan MAT253 Mass Spectrometer. Triplicate reproducibility is less than 0.2‰ for both isotopes. The isotopic standards used are: International Atomic Energy Agency (IAEA) standards of caffeine and glutamic acid for carbon and nitrogen; in-house laboratory standards of nylon, alanine, and bovine liver standard for carbon, nitrogen, and atomic C:N ratios. The samples from Titolo-Palese were analysed at SUERC. Here, stable carbon and nitrogen isotopic compositions were determined on a Delta V Advantage continuous-flow isotope ratio mass spectrometer coupled via a ConfloIV to an IsoLink elemental analyser (Thermo Scientific, Bremen) following Sayle et al.97. The International Atomic Energy Agency (IAEA) reference materials USGS40 (L-glutamic acid, δ13CV-PDB = − 26.4 ± 0.0‰, δ15NAIR = − 4.5 ± 0.1‰) and USGS41a (L-glutamic acid, δ13CV-PDB = 36.6 ± 0.1‰, δ15NAIR = 47.6 ± 0.2‰) were used to normalise δ13C and δ15N values. Normalisation was checked using the marine collagen USGS88 (δ13CV-PDB = − 16.1 ± 0.1‰ and δ15NAIR = 15.0 ± 0.1‰) and the porcine collagen USGS89 (δ13CV-PDB = − 18.1 ± 0.1‰ and δ15NAIR = 6.3 ± 0.1‰), which gave the values: USGS88, δ13CV-PDB = − 16.3 ± 0.1‰ and δ15NAIR = 15.2 ± 0.1‰ and USGS89, δ13CV-PDB = − 18.2 ± 0.1‰ and δ15NAIR = 6.4 ± 0.2‰. Stable isotope concentrations are measured as the ratio of the heavier isotope to the lighter isotope relative to an internationally defined scale, Vienna Pee Dee Belemnite (VPDB) for carbon and Ambient Inhalable Reservoir (AIR) for nitrogen (Hoefs & Hoefs, 1997). Isotope results are reported as δ values (δ13C and δ15N) in parts per 1000 or “per mil” (‰). Results are reported in Table S1. Previous isotopic measurements on Neolithic humans and animals are reported in Table S2.
Chronological and geographical assessment
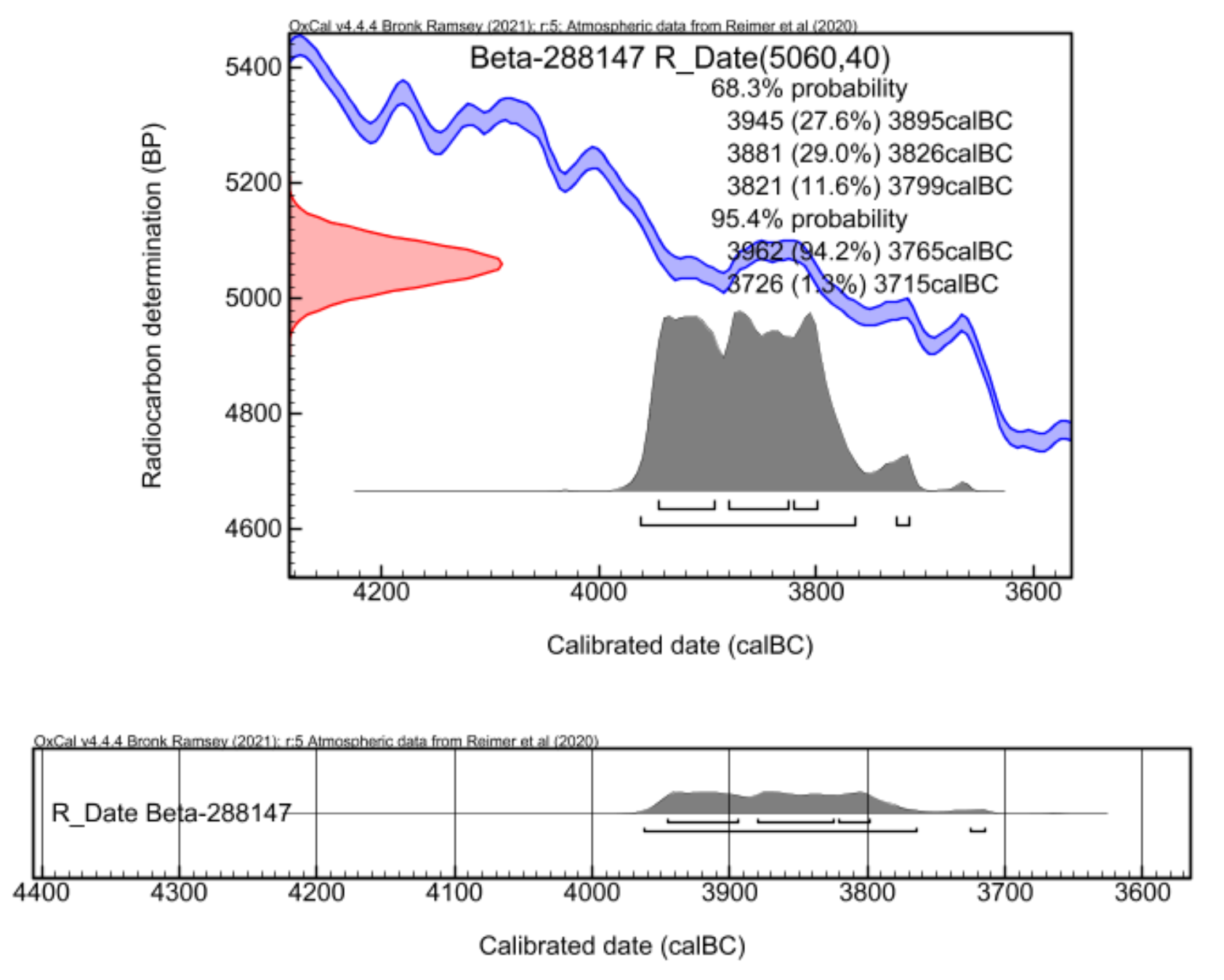
Humans and animals were classified as ‘Early’ ‘Middle’ or ‘Late’ Neolithic following radiocarbon dates when available, and material culture when they were not (Table S1)98. The limited availability of radiocarbon dates for many prehistoric Italian sites is in part a result of well-developed material culture typologies; combined, they provide a good estimation of phasing and allow us to explore dietary variability throughout time. We also provide a new radiocarbon date of one sample of human bone (OCH2) from Diga di Occhito, dated to 5060 ± 40 BP (Beta-288147: 3962–3715 cal BC, 95.4%), placing the site within the Late Neolithic99 (Fig. S3).
Coordinates of the sites were provided by the archaeologists or derived from previous publications.
Where these were not available we used the coordinates of the closest town/village or landmark from the site.
Plotting and statistical analysis
Scatter plots, descriptive statistics and statistical tests were carried out using RStudio, R version 4.0.3 using the packages dplyr, tidyr, Hmisc, and ggplot2.
Regional analysis and mixing models
To answer the archaeological questions, we used two different tools from the open access application IsoMemo (https://isomemoapp.com/app/iso-memo-app). AverageR was used to explore regional variability of δ13C (‰) and δ15N (‰) values as previously described47,48. We used the default options with the exception of the smooth type (planar), extrapolation behaviour (constant) and we used the Bayesian model with the number of MCMC iterations set to 5000 for the analysis of the data. ReSources was used instead to quantitatively evaluate the diet of Neolithic individuals using a routed and concentration-dependent model100,101. We used terrestrial animal products, C3 cereals and marine fish as possible food sources for those individuals from archaeological sites located in proximity to the coast (km from the coast ≤ 15) while we only relied on terrestrial animal products and C3 cereals for those considered inland sites (km from the coast > 15). δ13C (‰) and δ15N (‰) values of the animals from this and previous studies were used to represent the “animal products” category after having checked that the values were uniform across southeastern Italy using regional analysis. As for the ‘C3 cereals’ source, since these are dramatically lacking from the area of investigation, we relied on published wheat and barley δ13C (‰) and δ15N (‰) values from a Greek Neolithic context102. The analysis of carbon and nitrogen stable isotopes of local plant remains is essential103, and here we urge for this. δ13C (‰) and δ15N (‰) values of local marine fish specimens from Lelli et al.8 were used to represent the ‘marine fish’ category. Unfortunately, the marine fish used belongs to an older chronology but it represents the only opportunity to use a local baseline for this food category. The sources δ13C (‰) and δ15N (‰) values were corrected to represent the actual values in the tissues consumed using the recently updated offsets from Soncin et al.76. Uncertainties associated with the source values are 1SD. The offset Δ15Ncollagen-diet was set at + 5.5 ± 0.5‰, with 100% contribution from protein, and the offset Δ13Ccollagen-diet was set to + 4.8 ± 0.5‰, with 74 ± 4% contribution from protein and 26 ± 4% from lipids and carbohydrates104. Concentrations expressed as dry weight (%) are the same of those reported in Soncin et al.76 using the USDA National Nutrient Database for Standard Reference (available at https://fdc.nal.usda.gov/) with uncertainties being the standard error. As for model parameters, we used the default options with the exception of the following: model type: Individual targets (no shared info); Source contribution distribution: deselected “Optimal objective prior”; Covariates model: fixed intercept (cat. vars), fixed slope (num. vars).
Supplementary Information
{kind=link}
{kind=link}
{kind=link}
Acknowledgements
This work was partly supported by a Mary Curie grant (FOOD—Food and Society in Mediterranean Prehistory—235966) awarded to M. A. Tafuri and by the European Union through the European Research Council Advanced Grant “Making Ancestors: the Politics of Death in European Prehistory”, No. 885137 (John Robb, Mary Anne Tafuri, Silvia Soncin, Sofia Panella, Jess Thompson). We would like to thank Giorgio Manzi, Fulvio Bartoli, Giovanna Belcastro, Paola Iacumin, Cecilia Conati Barbaro, Alessandro Vanzetti, Norma Lonoce, Sandro Sublimi Saponetti, for granting access to the samples, Tamsin O’Connell for sharing some of the data presented here and Carlo Cocozza and the IsoMemo team for their help with the IsoMemo app. We would also like to thank the MAReA team for useful discussion on the paper. Please note that for the purpose of open access, the authors have applied a Creative Commons Attribution (CC BY) licence to any Author Accepted Manuscript version arising from this submission.
Author contributions
M.A.T.: conceptualization (lead); data curation (equal); formal analysis (equal); funding acquisition (lead); investigation (lead); methodology (equal); supervision (lead); writing—original draft preparation (equal). S.S.: data curation (equal); formal analysis (equal); methodology (equal); visualisation (lead); writing—original draft preparation (equal). S.P.: investigation (supporting); writing—review and editing (supporting). J.E.T.: investigation (supporting); writing—review and editing (supporting). I.T.: resources (supporting); writing—review and editing (supporting). P.F.F.: resources (supporting); writing—review and editing (supporting). S.S.: resources (supporting); writing—review and editing (supporting). F.R.: resources (supporting); writing—review and editing (supporting). S.M.: resources (supporting); writing—review and editing (supporting). I.M.M.: writing—review and editing (supporting). G.F.: writing—review and editing (supporting). J.R.: investigation (supporting); Writing—review and editing (supporting).
Data availability
The authors confirm that the data supporting the findings of this study are available within the article and/or its supplementary materials.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
M. A. Tafuri, Email: maryanne.tafuri@uniroma1.it
S. Soncin, Email: silvia.soncin@uniroma1.it
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-023-34771-y.
References
- 1.Arena F, Gualdi-Russo E, Olsen J, Philippsen B, Mannino MA. New data on agro-pastoral diets in southern Italy from the Neolithic to the Bronze Age. Archaeol. Anthropol. Sci. 2020;12:245. doi: 10.1007/s12520-020-01209-9. [DOI] [Google Scholar]
- 2.Bernardini, S., Coppa, A. & Moggi-Cecchi, J. Social dynamics and resource management strategies in copper age Italy: insights from archaeological and isotopic data. Environmentalist (2021).
- 3.Cortese, F., De Angelis, F. & Achino, K. F. Isotopic reconstruction of the subsistence strategy for a Central Italian Bronze Age community (Pastena cave, 2nd millennium BCE). Archaeol. Anthropol. Sci.14, (2022).
- 4.Craig OE, et al. Stable isotope analysis of Late Upper Palaeolithic human and faunal remains from Grotta del Romito (Cosenza) Italy. J. Archaeol. Sci. 2010;37:2504–2512. doi: 10.1016/j.jas.2010.05.010. [DOI] [Google Scholar]
- 5.De Angelis F, et al. Eneolithic subsistence economy in Central Italy: First dietary reconstructions through stable isotopes. Archaeol. Anthropol. Sci. 2019;11:4171–4186. doi: 10.1007/s12520-019-00789-5. [DOI] [Google Scholar]
- 6.Gazzoni V, Goude G, Herrscher E, Guerreschi A. Late Upper Palaeolithic human diet: first stable isotope evidence from Riparo Tagliente (Verona, Italy) Bulletins et mémoires de la Société d’anthropologie de Paris. 2013;25:103–117. [Google Scholar]
- 7.Goude G, Dori I, Sparacello VS, Starnini E, Varalli A. Multi-proxy stable isotope analyses of dentine microsections reveal diachronic changes in life history adaptations, mobility, and tuberculosis-induced wasting in prehistoric Liguria (Finale Ligure, Italy, northwestern Mediterranean) Int. J. Paleopathol. 2020;28:99–111. doi: 10.1016/j.ijpp.2019.12.007. [DOI] [PubMed] [Google Scholar]
- 8.Lelli R, et al. Examining dietary variability of the earliest farmers of south-eastern Italy. Am. J. Phys. Anthropol. 2012;149:380–390. doi: 10.1002/ajpa.22134. [DOI] [PubMed] [Google Scholar]
- 9.Mannino MA, Thomas KD, Leng MJ, Di Salvo R, Richards MP. Stuck to the shore? Investigating prehistoric hunter-gatherer subsistence, mobility and territoriality in a Mediterranean coastal landscape through isotope analyses on marine mollusc shell carbonates and human bone collagen. Quat. Int. 2011;244:88–104. doi: 10.1016/j.quaint.2011.05.044. [DOI] [Google Scholar]
- 10.Mannino, M. A. et al. Origin and diet of the prehistoric hunter-gatherers on the mediterranean island of Favignana (Ègadi Islands, Sicily). PLoS One7 (2012). [DOI] [PMC free article] [PubMed]
- 11.Rumolo A, Forstenpointner G, Rumolo P, Jung R. Palaeodiet reconstruction inferred by stable isotopes analysis of faunal and human remains at Bronze Age Punta di Zambrone (Calabria, Italy) Int. J. Osteoarchaeol. 2020;30:90–98. doi: 10.1002/oa.2836. [DOI] [Google Scholar]
- 12.Scorrano G, Baldoni M, Brilli M, Rolfo MF. Effect of Neolithic transition on an Italian community: Mora Cavorso (Jenne, Rome) Archaeol. Anthropol. Sci. 2019;11:1443–1459. doi: 10.1007/s12520-018-0615-9. [DOI] [Google Scholar]
- 13.Tafuri MA, Craig OE, Canci A. Stable isotope evidence for the consumption of millet and other plants in Bronze Age Italy. Am. J. Phys. Anthropol. 2009;139:146–153. doi: 10.1002/ajpa.20955. [DOI] [PubMed] [Google Scholar]
- 14.Tafuri, M, O’Connell, T, Souter, E, Libianchi, N, Robb, J. Diet during life: paleoeconomic studies of human diet using stable carbon and nitrogen isotopes. in The Archaeology of Grotta Scaloria: Ritual in Neolithic Southeast Italy (ed. Ernestine S. Elster, John E. Robb, Eugenia Isetti, Antonella Traverso) 131–138 (Cotsen Institute of Archaeology Press, 2016).
- 15.Tykot et al. Osservazioni sull’alimentazione della comunità preistorica di contrada Scintilia di Favara (AG) sulla base di analisi isotopiche. in Preistoria del cibo. L’alimentazione nella preistoria e nella protostoria (eds. Damiani, I., Cazzella, A. & Copat, V.) vol. 6 65–74 (Istituto Italiano di Preistoria e Protostoria., 2021).
- 16.Tafuri MA, Rottoli M, Cupitò M. Estimating C4 plant consumption in Bronze Age Northeastern Italy through stable carbon and nitrogen isotopes in bone collagen. Aquat. Microb. Ecol. 2018;28:131–142. [Google Scholar]
- 17.Varalli, A. et al. Dietary continuity vs. discontinuity in Bronze Age Italy. The isotopic evidence from Arano di Cellore (Illasi, Verona, Italy). J. Archaeol. Sci. Rep.7, 104–113 (2016).
- 18.Robb, J. The early Mediterranean village: agency, material culture, and social change in Neolithic Italy. (Cambridge University Press, 2007).
- 19.Roberts P, et al. Calling all archaeologists: guidelines for terminology, methodology, data handling, and reporting when undertaking and reviewing stable isotope applications in archaeology. Rapid Commun. Mass Spectrom. 2018;32:361–372. doi: 10.1002/rcm.8044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.O’Leary MH. Carbon isotopes in photosynthesis. Bioscience. 1988;38:328–336. doi: 10.2307/1310735. [DOI] [Google Scholar]
- 21.Peterson BJ, Fry B. Stable isotopes in ecosystem studies. Annu. Rev. Ecol. Syst. 1987;18:293–320. doi: 10.1146/annurev.es.18.110187.001453. [DOI] [Google Scholar]
- 22.Heaton THE, Vogel JC. Climatic influence on the isotopic composition of bone nitrogen. Nature. 1986;322:822–823. doi: 10.1038/322822a0. [DOI] [Google Scholar]
- 23.Van Klinken, G. J., Richards, M. P. & Hedges, B. E. M. An Overview of Causes for Stable Isotopic Variations in Past European Human Populations: Environmental, Ecophysiological, and Cultural Effects. in Biogeochemical Approaches to Paleodietary Analysis (eds. Ambrose, S. H. & Katzenberg, M. A.) 39–63 (Springer US, 2002).
- 24.DeNiro MJ, Epstein S. Influence of diet on the distribution of carbon isotopes in animals. Geochim. Cosmochim. Acta. 1978;42:495–506. doi: 10.1016/0016-7037(78)90199-0. [DOI] [Google Scholar]
- 25.Edgar Hare, P., Fogel, M. L., Stafford, T. W., Mitchell, A. D. & Hoering, T. C. The isotopic composition of carbon and nitrogen in individual amino acids isolated from modern and fossil proteins. J. Archaeol. Sci.18, 277–292 (1991).
- 26.Ambrose, S. H. & Norr, L. Experimental Evidence for the Relationship of the Carbon Isotope Ratios of Whole Diet and Dietary Protein to Those of Bone Collagen and Carbonate. in Prehistoric Human Bone: Archaeology at the Molecular Level (eds. Lambert, J. B. & Grupe, G.) 1–37 (Springer Berlin Heidelberg, 1993).
- 27.Tieszen, L. L. & Fagre, T. Effect of Diet Quality and Composition on the Isotopic Composition of Respiratory CO2, Bone Collagen, Bioapatite, and Soft Tissues. in Prehistoric Human Bone: Archaeology at the Molecular Level (eds. Lambert, J. B. & Grupe, G.) 121–155 (Springer Berlin Heidelberg, 1993).
- 28.Howland MR, et al. Expression of the dietary isotope signal in the compound-specific δ13C values of pig bone lipids and amino acids. Int. J. Osteoarchaeol. 2003;13:54–65. doi: 10.1002/oa.658. [DOI] [Google Scholar]
- 29.Jim S, Ambrose SH, Evershed RP. Stable carbon isotopic evidence for differences in the dietary origin of bone cholesterol, collagen and apatite: Implications for their use in palaeodietary reconstruction. Geochim. Cosmochim. Acta. 2004;68:61–72. doi: 10.1016/S0016-7037(03)00216-3. [DOI] [Google Scholar]
- 30.Warinner C, Tuross N. Alkaline cooking and stable isotope tissue-diet spacing in swine: Archaeological implications. J. Archaeol. Sci. 2009;36:1690–1697. doi: 10.1016/j.jas.2009.03.034. [DOI] [Google Scholar]
- 31.Webb EC, et al. The influence of varying proportions of terrestrial and marine dietary protein on the stable carbon-isotope compositions of pig tissues from a controlled feeding experiment. Sci. Technol. Archaeol. Res. 2017;3:28–44. [Google Scholar]
- 32.van der Merwe NJ. Carbon isotopes, photosynthesis, and archaeology: Different pathways of photosynthesis cause characteristic changes in carbon isotope ratios that make possible the study of prehistoric human diets. Am. Sci. 1982;70:596–606. [Google Scholar]
- 33.DeNiro MJ, Epstein S. Influence of diet on the distribution of nitrogen isotopes in animals. Geochim. Cosmochim. Acta. 1981;45:341–351. doi: 10.1016/0016-7037(81)90244-1. [DOI] [Google Scholar]
- 34.Ambrose, S. H. Controlled Diet and Climate Experiments on Nitrogen Isotope Ratios of Rats. in Biogeochemical Approaches to Paleodietary Analysis (eds. Ambrose, S. H. & Katzenberg, M. A.) 243–259 (Springer US, 2002).
- 35.Young, S. M. Metabolic mechanisms and the isotopic investigation of ancient diets with an application to human remains from Cuello, Belize. (Harvard University, 2003).
- 36.Webb EC, Stewart A, Miller B, Tarlton J, Evershed RP. Age effects and the influence of varying proportions of terrestrial and marine dietary protein on the stable nitrogen-isotope compositions of pig bone collagen and soft tissues from a controlled feeding experiment. Sci. Technol. Archaeol. Res. 2016;2:54–66. [Google Scholar]
- 37.Kendall, I. P., Lee, M. R. F. & Evershed, R. P. The effect of trophic level on individual amino acid δ15N values in a terrestrial ruminant food web. STAR: Sci. Technol. Archaeol. Res.3, 135–145 (2017).
- 38.O’Connell TC, Kneale CJ, Tasevska N, Kuhnle GGC. The diet-body offset in human nitrogen isotopic values: A controlled dietary study. Am. J. Phys. Anthropol. 2012;149:426–434. doi: 10.1002/ajpa.22140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Craig OE, Bondioli L, Fattore L, Higham T, Hedges R. Evaluating marine diets through radiocarbon dating and stable isotope analysis of victims of the AD79 eruption of Vesuvius. Am. J. Phys. Anthropol. 2013;152:345–352. doi: 10.1002/ajpa.22352. [DOI] [PubMed] [Google Scholar]
- 40.Cubas M, et al. Long-term dietary change in Atlantic and Mediterranean Iberia with the introduction of agriculture: A stable isotope perspective. Archaeol. Anthropol. Sci. 2019;11:3825–3836. doi: 10.1007/s12520-018-0752-1. [DOI] [Google Scholar]
- 41.Horden, P. & Purcell, N. The Corrupting Sea. (Blackwell Publishing, 2000).
- 42.Vizzini S, Savona B, Chi TD, Mazzola A. Spatial variability of stable carbon and nitrogen isotope ratios in a Mediterranean coastal lagoon. Hydrobiologia. 2005;550:73–82. doi: 10.1007/s10750-005-4364-2. [DOI] [Google Scholar]
- 43.Vika E, Theodoropoulou T. Re-investigating fish consumption in Greek antiquity: results from δ13C and δ15N analysis from fish bone collagen. J. Archaeol. Sci. 2012;39:1618–1627. doi: 10.1016/j.jas.2012.01.016. [DOI] [Google Scholar]
- 44.Parkinson EW, McLaughlin TR. Lifeways at the acme of the south Italian Neolithic: New chronological and bioarchaeological data from Fonteviva, Apulia. J. Archaeol. Sci. Rep. 2020;34:102589. [Google Scholar]
- 45.Tafuri, M. A. et al. Herding Practices in the Ditched Villages of the Neolithic Tavoliere (Apulia, South-east Italy): A Vicious Circle? The Isotopic Evidence. in Early farmers (British Academy, 2014).
- 46.Ambrose SH. Preparation and characterization of bone and tooth collagen for isotopic analysis. J. Archaeol. Sci. 1990;17:431–451. doi: 10.1016/0305-4403(90)90007-R. [DOI] [Google Scholar]
- 47.Cubas M, et al. Latitudinal gradient in dairy production with the introduction of farming in Atlantic Europe. Nat. Commun. 2020;11:2036. doi: 10.1038/s41467-020-15907-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Izdebski A, et al. Palaeoecological data indicates land-use changes across Europe linked to spatial heterogeneity in mortality during the Black Death pandemic. Nat. Ecol. Evol. 2022;6:297–306. doi: 10.1038/s41559-021-01652-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Desiato, F., Fioravanti, G., Fraschetti, P., Perconti, W. & Piervitali, E. Valori climatici normali di temperatura e precipitazione in Italia. (2015).
- 50.Vaiglova P, Lazar NA, Stroud EA, Loftus E, Makarewicz CA. Best practices for selecting samples, analyzing data, and publishing results in isotope archaeology. Quat. Int. 2022;650:86–100. doi: 10.1016/j.quaint.2022.02.027. [DOI] [Google Scholar]
- 51.Monaco, A. A simulation of farming and breeding activities: comparing the economic strategies in South East Italy Neolithic communities. in Origini (ed. Manfredini, A.) 61–82 (Gangemi Editore, 2011).
- 52.Boenzi, F., Caldara, M., Pennetta, L. & Simone, O. Environmental Aspects Related to the Physical Evolution of Some Wetlands Along the Adriatic Coast of Apulia (Southern Italy): A Review. J. Coast. Res. 170–175 (2006).
- 53.Caldara, M., Muntoni, I. M., Fiorentino, G., Primavera, M. & Radina, F. Hidden Neolithic landscapes in Apulian Region. in Hidden Landscapes of Mediterranean Europe Cultural and methodological biases in pre- and protohistoric landscape studies. Proceedings of the international meeting Siena (eds. van Leusen, G., Pizziolo, L. & Sarti, M.) 2320, 183–191 (Archeopress, 2011).
- 54.Fiorentino G, et al. Climate changes and human–environment interactions in the Apulia region of southeastern Italy during the Neolithic period. Holocene. 2013;23:1297–1316. doi: 10.1177/0959683613486942. [DOI] [Google Scholar]
- 55.Cresson P, et al. Primary production and depth drive different trophic structure and functioning of fish assemblages in French marine ecosystems. Prog. Oceanogr. 2020;186:102343. doi: 10.1016/j.pocean.2020.102343. [DOI] [Google Scholar]
- 56.Graniero, L. E., Grossman, E. L. & O’Dea, A. Stable isotopes in bivalves as indicators of nutrient source in coastal waters in the Bocas del Toro Archipelago, Panama. PeerJ4 (2016). [DOI] [PMC free article] [PubMed]
- 57.Mathieu-Resuge, M. et al. Different particle sources in a bivalve species of a coastal lagoon: evidence from stable isotopes, fatty acids, and compound-specific stable isotopes. Mar. Biol.166, (2019).
- 58.Spiteri CD, et al. Regional asynchronicity in dairy production and processing in early farming communities of the northern Mediterranean. Proc. Natl. Acad. Sci. 2016;113:13594–13599. doi: 10.1073/pnas.1607810113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Quagliariello, A. et al. Ancient oral microbiomes support gradual Neolithic dietary shifts towards agriculture. Nat. Commun.13 (2022). [DOI] [PMC free article] [PubMed]
- 60.Hedges R, Saville A, O’Connell TC. Characterizing the diet of individuals at the neolithic chambered tomb of Hazleton north, Gloucestershire, England, using stable isotopic analysis. Archaeometry. 2008;50:114–128. doi: 10.1111/j.1475-4754.2007.00379.x. [DOI] [Google Scholar]
- 61.Bownes JM, Ascough PL, Cook GT, Murray I, Bonsall C. Using stable isotopes and a bayesian mixing model (FRUITS) to investigate diet at the early neolithic site of carding Mill Bay, Scotland. Radiocarbon. 2017;59:1275–1294. doi: 10.1017/RDC.2017.39. [DOI] [Google Scholar]
- 62.Schulting RJ, MacDonald R, Richards MP. FRUITS of the sea? A cautionary tale regarding Bayesian modelling of palaeodiets using stable isotope data. Quat. Int. 2022;650:52–61. doi: 10.1016/j.quaint.2022.02.012. [DOI] [Google Scholar]
- 63.Siebke, I. et al. Crops vs. animals: regional differences in subsistence strategies of Swiss Neolithic farmers revealed by stable isotopes. Archaeol. Anthropol. Sci.12 (2020).
- 64.Dürrwächter C, Craig OE, Collins MJ, Burger J, Alt KW. Beyond the grave: Variability in Neolithic diets in Southern Germany? J. Archaeol. Sci. 2006;33:39–48. doi: 10.1016/j.jas.2005.06.009. [DOI] [Google Scholar]
- 65.Oelze VM, et al. Early Neolithic diet and animal husbandry: stable isotope evidence from three Linearbandkeramik (LBK) sites in Central Germany. J. Archaeol. Sci. 2011;38:270–279. doi: 10.1016/j.jas.2010.08.027. [DOI] [Google Scholar]
- 66.Münster, A. et al. 4000 years of human dietary evolution in central Germany, from the first farmers to the first elites. PLoS One13 (2018). [DOI] [PMC free article] [PubMed]
- 67.Hoekman-Sites HA, Giblin JI. Prehistoric animal use on the Great Hungarian Plain: A synthesis of isotope and residue analyses from the Neolithic and Copper Age. J. Anthropol. Archaeol. 2012;31:515–527. doi: 10.1016/j.jaa.2012.05.002. [DOI] [Google Scholar]
- 68.Goude G, et al. A multidisciplinary approach to neolithic life reconstruction. J. Archaeol. Method Theory. 2019;26:537–560. doi: 10.1007/s10816-018-9379-x. [DOI] [Google Scholar]
- 69.Rey L, et al. A multi-isotope analysis of Neolithic human groups in the Yonne valley, Northern France: Insights into dietary patterns and social structure. Archaeol. Anthropol. Sci. 2019;11:5591–5616. doi: 10.1007/s12520-019-00885-6. [DOI] [Google Scholar]
- 70.Rey L, et al. Specifying subsistence strategies of early farmers: New results from compound-specific isotopic analysis of amino acids. Int. J. Osteoarchaeol. 2022;32:654–668. doi: 10.1002/oa.3093. [DOI] [Google Scholar]
- 71.Gibaja JF, et al. Human diet and the chronology of neolithic societies in the north-east of the Iberian Peninsula: The necropolises of Puig d’en Roca and Can Gelats (Girona, Spain) Archaeol. Anthropol. Sci. 2017;9:903–913. doi: 10.1007/s12520-015-0311-y. [DOI] [Google Scholar]
- 72.Fernández-Crespo T, et al. Isotopic evidence of strong reliance on animal foods and dietary heterogeneity among Early-Middle Neolithic communities of Iberia. Archaeol. Anthropol. Sci. 2019;11:5463–5481. doi: 10.1007/s12520-019-00889-2. [DOI] [Google Scholar]
- 73.Papathanasiou A. Stable isotope analyses in neolithic and bronze age greece: An overview. Hesperia Suppl. 2015;49:25–55. [Google Scholar]
- 74.Guiry E, et al. Stable isotope palaeodietary and radiocarbon evidence from the early neolithic site of Zemunica, Dalmatia Croatia. Eur. J. Archaeol. 2017;20:235–256. doi: 10.1017/eaa.2016.24. [DOI] [Google Scholar]
- 75.McLaughlin, R. et al. An isotopic study of palaeodiet at the Circle and the Xemxija tombs. in Temple people: Bioarchaeology, resilience and culture in prehistoric Malta (eds. Stoddart, S. et al.) 295–302 (McDonald Institute for Archaeological Research, 2022).
- 76.Soncin, S. et al. High-resolution dietary reconstruction of victims of the AD79 Vesuvius eruption at Herculaneum by compound specific isotope analysis. Sci. Adv.7 (2021). [DOI] [PMC free article] [PubMed]
- 77.Fernandes R, Nadeau M-J, Grootes PM. Macronutrient-based model for dietary carbon routing in bone collagen and bioapatite. Archaeol. Anthropol. Sci. 2012;4:291–301. doi: 10.1007/s12520-012-0102-7. [DOI] [Google Scholar]
- 78.Killgrove K, Tykot RH. Food for Rome: A stable isotope investigation of diet in the Imperial period (1st–3rd centuries AD) J. Anthropol. Archaeol. 2013;32:28–38. doi: 10.1016/j.jaa.2012.08.002. [DOI] [Google Scholar]
- 79.Prowse T, Schwarcz HP, Saunders S, Macchiarelli R, Bondioli L. Isotopic paleodiet studies of skeletons from the Imperial Roman-age cemetery of Isola Sacra, Rome. Italy. J. Archaeol. Sci. 2004;31:259–272. doi: 10.1016/j.jas.2003.08.008. [DOI] [Google Scholar]
- 80.Tanasi, D., Tykot, R. H., Vianello, A. & Hassam, S. Stable isotope analysis of the dietary habits of a Greek community in Archaic Syracuse (Sicily): A pilot study. STAR Sci. Technol. Archaeol. Res.3, 466–477 (2017).
- 81.Fontanals-Coll, M., Eulàlia Subirà, M., Díaz-Zorita Bonilla, M. & Gibaja, J. F. First insight into the Neolithic subsistence economy in the north-east Iberian Peninsula: paleodietary reconstruction through stable isotopes. Am. J. Phys. Anthropol.162, 36–50 (2017). [DOI] [PubMed]
- 82.Lightfoot E, Boneva B, Miracle PT, Šlaus M, O’Connell TC. Exploring the mesolithic and neolithic transition in croatia through isotopic investigations. Antiquity. 2011;85:73–86. doi: 10.1017/S0003598X00067442. [DOI] [Google Scholar]
- 83.Richards MP, Schulting RJ. Touch not the fish: The Mesolithic-Neolithic change of diet and its significance. Antiquity. 2006;80:444–456. doi: 10.1017/S0003598X00093765. [DOI] [Google Scholar]
- 84.Salazar-García, Fontanals-Coll & Goude. ‘To “seafood”or not to “seafood”?’ An isotopic perspective on dietary preferences at the Mesolithic-Neolithic transition in the Western Mediterranean. Quat.470, 497–510 (2018).
- 85.Schulting, R. Dietary shifts at the Mesolithic-Neolithic transition in Europe: an overview of the stable isotope data. in The Oxford Handbook of the Archaeology of Diet (eds. Lee-Thorp, J. & Katzenberg, M. A.) (Oxford University Press, 2015).
- 86.Garcia Guixé, E., Richards, M. P. & Subirà, M. E. Palaeodiets of humans and fauna at the Spanish Mesolithic site of El Collado. Curr. Anthropol.47, 549–557 (2006).
- 87.Lubell D, Jackes M, Schwarcz H, Knyf M, Meiklejohn C. The mesolithic-neolithic transition in Portugal: Isotopic and dental evidence of diet. J. Archaeol. Sci. 1994;21:201–216. doi: 10.1006/jasc.1994.1022. [DOI] [Google Scholar]
- 88.Papathanasiou A. Stable isotope analysis in Neolithic Greece and possible implications on human health. Int. J. Osteoarchaeol. 2003;13:314–324. doi: 10.1002/oa.705. [DOI] [Google Scholar]
- 89.Rainsford C, O’Connor T, Miracle P. Fishing in the Adriatic at the Mesolithic-Neolithic transition: Evidence from Vela Spila. Croatia. Environ. Archaeol. 2014;19:311–320. doi: 10.1179/1749631414Y.0000000018. [DOI] [Google Scholar]
- 90.Fontanals-Coll M, Subirà ME, Marín-Moratalla N, Ruiz J, Gibaja JF. From Sado Valley to Europe: Mesolithic dietary practices through different geographic distributions. J. Archaeol. Sci. 2014;50:539–550. doi: 10.1016/j.jas.2014.07.028. [DOI] [Google Scholar]
- 91.Fontanals-Coll, M. et al. Stable isotope analyses of amino acids reveal the importance of aquatic resources to Mediterranean coastal hunter–gatherers. Proc. Royal Soc. B290 (2023). [DOI] [PMC free article] [PubMed]
- 92.Bickle P. Stable isotopes and dynamic diets: The Mesolithic-Neolithic dietary transition in terrestrial central Europe. J. Archaeol. Sci. Rep. 2018;22:444–451. [Google Scholar]
- 93.Galili, E., Gopher, A., Rosen, B. & Horwitz, L. K. The emergence of the Mediterranean Fishing Village in the Levant and the anomaly of Neolithic Cyprus. in Neolithic Revolution: New perspectives on Southwest Asia in light of recent discoveries on Cyprus (eds. Peltenburg, E. & Wasse, A.) 2, 91–101 (Oxbow Books, 2004).
- 94.Pessina, A. & Tiné, V. Archeologia del Neolitico - L’Italia tra sesto e quarto millennio. (Carocci editore, 2020).
- 95.Longin R. New method of collagen extraction for radiocarbon dating. Nature. 1971;230:241–242. doi: 10.1038/230241a0. [DOI] [PubMed] [Google Scholar]
- 96.Tuross N. Comparative decalcification methods, radiocarbon dates, and stable isotopes of the VIRI bones. Radiocarbon. 2012;54:837–844. doi: 10.1017/S0033822200047482. [DOI] [Google Scholar]
- 97.Sayle KL, Brodie CR, Cook GT, Hamilton WD. Sequential measurement of δ15 N, δ13 C and δ34 S values in archaeological bone collagen at the Scottish Universities Environmental Research Centre (SUERC): A new analytical frontier. Rapid Commun. Mass Spectrom. 2019;33:1258–1266. doi: 10.1002/rcm.8462. [DOI] [PubMed] [Google Scholar]
- 98.Binder, D. et al. Le complexe de la Céramique Imprimée en Méditerranée centrale et nord-occidentale: une synthèse chronoculturelle (VIIe et VIe millénaires AEC). in Céramiques imprimées de Méditerranée occidentale (VIe millénaire AEC): données, approches et enjeux nouveaux, Actes de la séance de la Société préhistorique française de Nice (eds. Binder, D. & Manen, C.) 18, 27–124 (Société préhistorique française, 2022).
- 99.Reimer PJ, et al. The IntCal20 northern hemisphere radiocarbon age calibration curve (0–55 cal kBP) Radiocarbon. 2020;62:725–757. doi: 10.1017/RDC.2020.41. [DOI] [Google Scholar]
- 100.Sołtysiak, A. & Fernandes, R. Much ado about nothing: assessing the impact of the 4.2 kya event on human subsistence patterns in northern Mesopotamia using stable isotope analysis. Antiquity95, 1145–1160 (2021).
- 101.Cocozza C, Cirelli E, Groß M, Teegen W-R, Fernandes R. Presenting the Compendium Isotoporum Medii Aevi, a Multi-Isotope Database for Medieval Europe. Sci. Data. 2022;9:354. doi: 10.1038/s41597-022-01462-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Vaiglova P, et al. An integrated stable isotope study of plants and animals from Kouphovouno, southern Greece: A new look at Neolithic farming. J. Archaeol. Sci. 2014;42:201–215. doi: 10.1016/j.jas.2013.10.023. [DOI] [Google Scholar]
- 103.Fiorentino G, Ferrio JP, Bogaard A, Araus JL, Riehl S. Stable isotopes in archaeobotanical research. Veg. Hist. Archaeobot. 2015;24:215–227. doi: 10.1007/s00334-014-0492-9. [DOI] [Google Scholar]
- 104.Fernandes R. A simple(R) model to predict the source of dietary carbon in individual consumers. Archaeometry. 2016;58:500–512. doi: 10.1111/arcm.12193. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The authors confirm that the data supporting the findings of this study are available within the article and/or its supplementary materials.