

Abstract
Net photosynthetic CO2 assimilation rate (An) decreases at leaf temperatures above a relatively mild optimum (Topt) in most higher plants. This decline is often attributed to reduced CO2 conductance, increased CO2 loss from photorespiration and respiration, reduced chloroplast electron transport rate (J), or deactivation of Ribulose-1,5-bisphosphate Carboxylase Oxygenase (Rubisco). However, it is unclear which of these factors can best predict species independent declines in An at high temperature. We show that independent of species, and on a global scale, the observed decline in An with rising temperatures can be effectively accounted for by Rubisco deactivation and declines in J. Our finding that An declines with Rubisco deactivation and J supports a coordinated down-regulation of Rubisco and chloroplast electron transport rates to heat stress. We provide a model that, in the absence of CO2 supply limitations, can predict the response of photosynthesis to short-term increases in leaf temperature.
Subject terms: Rubisco, C3 photosynthesis, Enzymes, Carbon cycle
Photosynthesis declines at mild temperatures in terrestrial plants. Here, the authors use published data to show that decline in photosynthetic CO2 assimilation rate with rising temperatures can be accounted for by Rubisco deactivation and declines in chloroplast electron transport rate.
Introduction
The rapid rise in leaf temperature during a heatwave has detrimental impacts on plant performance1. Photosynthesis (net CO2 assimilation—An) is particularly susceptible to heat stress, and the temperatures at which An decreases are well below those for comparable leaf metabolic processes like respiration1–3. The Farquhar, von Caemmerer, and Berry C3 photosynthesis model (FvCB model)4 is a powerful tool for predicting the response of An to environmental perturbations, and for determining what aspects of biochemistry limit photosynthetic rate and capacity. The FvCB model predicts An based on the minimal rate generated from Rubisco carboxylation reactions (Ac), Ribulose-1,5-bisphosphate (RuBP) regeneration associated with chloroplast electron transport rate (Ar), and triose phosphate utilisation (Ap)4,5. The model accounts for declines in An above the temperature optimum of photosynthesis (Topt) based on concomitant declines in chloroplast electron transport rates (J)4, which has been linked to heat damage of thylakoid membranes6,7. However, other studies suggest that Ac can be a greater contributor than Ar to the loss of An when leaf temperatures exceed Topt8. Deactivation of Rubisco and its impact on Ac has long been suspected of contributing to An inhibition above Topt9. Indeed, an analysis by Crafts-Brandner and Salvucci10 noted that declines in An with leaf heating occur well before expectations based on Rubisco kinetics, and are instead consistent with the temperature dependence of Rubisco deactivation. Further experiments and modelling have identified Ac as the rate limiting step in some instances, while other studies have implicated Ar due to declines in J with rising temperatures6,8,11,12. Whether Ac or Ar determines An above the Topt is often attributed to interspecific differences or environmental factors such as nitrogen availability, growth temperature, and ambient CO2 concentration6,12,13. An alternative possibility is that Ac and Ar are both regulated to be co-limiting. For example, Sage14 proposed and observed15 synchronised Ac and Ar biochemical adjustments within minutes of altering irradiation and CO2 concentrations.
Not only can the capacity of Rubisco to fix CO2 and the light dependent generation of RuBP be impaired by heat, but the availability of CO2 substrate at the site of assimilation can fall and become limiting. Reduced An due to falling intercellular and chloroplast CO2 concentrations following heat-associated rises in vapor pressure differences between leaves and air have been observed16–18. Additionally, foliar CO2 loss from photorespiration and respiration in the light (RL) may contribute substantially to declining An under high temperature19, as both processes rise sharply with warming20,21. From a modelling perspective, this means that for each new temperature, several parameters must be treated independently and updated. Despite, or perhaps because of, the above numerous aspects of photosynthetic metabolism that are impaired by heat, it is difficult to establish a general predictor for the decline in An at relatively moderate temperature applicable across many higher plants.
The maximum carboxylation capacity of Rubisco (Vcmax) is a key parameter in the FvCB model22. Gas-exchange estimates of Vcmax increase exponentially with temperature before peaking and then declining at higher temperatures; the point of decline is influenced by acclimation to growth temperature23,24. This decline in apparent Vcmax is not explained by susceptibility of Rubisco to high temperature. Rubisco is a relatively thermally stable protein, and in vitro thermal characterisation of Rubisco, in the absence of phosphorylated compounds, demonstrates that it has an exponentially rising carboxylation rate constant (kcat) and remains functional at temperatures far exceeding the in vivo deactivation point25–27. Rather, the deactivation of Rubisco is due to the heat sensitivity of Rubisco activase (Rca), the accessory protein of Rubisco that removes tightly bound sugar phosphate inhibitors from the Rubisco active site. Rubisco is prone to decarbamylation, where a Mg2+ ion and CO2 molecule is not bound to the active site prior to RuBP substrate binding, leading to deactivation and the need for Rca to remove bound RuBP from the active site28. Loss of Rca function leads to a reduction in the proportion of Rubisco catalytic sites that are activated and to concomitant declines in photosynthesis25,26. Accurately modelling An at temperatures above the Topt therefore requires knowledge about the activation state of Rubisco catalytic sites.
A central assumption of the FvCB model is that all Rubisco catalytic sites in a leaf are functional and invariant29. However, this assumption is inconsistent with the observed decline in apparent Vcmax values (calculated from gas-exchange data) and the number of functional sites under rising leaf temperature. One work-around is to regard Vcmax as a variable in time, like temperature itself. Alternatively, a more satisfying reconciliation of this inconsistency is to calculate Vcmax based on the kcat of Rubisco and its deactivation based on biochemical observations14,30. We hypothesise that we can accurately predict An above the Topt by allowing the number of functional Rubisco catalytic sites to vary with temperature. We explore the extent to which this Rubisco deactivation-based Ac corresponds to a previous model that predicts J-dependent Ar declines in An. We tested the scalability of these models against published temperature response curves and a global composite response curve of An measured over a wide range of leaf temperatures, biomes, and plant functional types.
Results
Accounting for Rubisco deactivation in C3 photosynthesis models
To capture Vcmax when accounting for the biochemically reported deactivation of Rubisco with rising temperature, the Sharpe-Schoolfield equation for enzyme deactivation at high temperatures31 was used:
| 1 |
The expected number of Rubisco catalytic sites (n), and the kcat of Rubisco at a given temperature were based on reported values from more than 70 higher plant species as described in the methods and presented in Supplementary Table 1 and Supplementary Fig. 1. We iteratively solved the deactivation energy (Ed) and the temperature at which enzyme activity was halved (T0.5)31 through a non-linear least-squares regression fit of Eq. 1, with the numerator set to unity, to published biochemical responses of the Rubisco activation state to leaf temperature (Fig. 1a). Heat susceptibility of Rca varies depending on acclimation to growth temperature and the thermal environment to which a plant is adapted32,33. Therefore, as with Rubisco kinetic parameters, we separated activation data based on whether a species grew at a day temperature below (cool) or above (warm) 25 °C, respectively (Supplementary Table 2). Our analysis included 17 species, consisting of seven cool and 10 warm grown species. Twelve of these species were herbaceous, while only three were temperate trees and zero were tropical trees, highlighting the current lack of knowledge of Rubisco activation across plant functional types. Non-linear least-squares iteration found a cool growth Ed of 199 kJ mol−1 and T0.5 of 39.0 °C, and a warm growth Ed of 212 kJ mol−1 and T0.5 of 42.4 °C. The decline in Vcmax derived from this mechanistic link to Rca functional control of Rubisco activation closely matched the peaked Arrhenius equation with an acclimation term, a model derived from empirical gas-exchange based apparent Vcmax data23 (Fig. 1b).
Fig. 1. Deactivation of Rubisco and its implications for Vcmax and An.
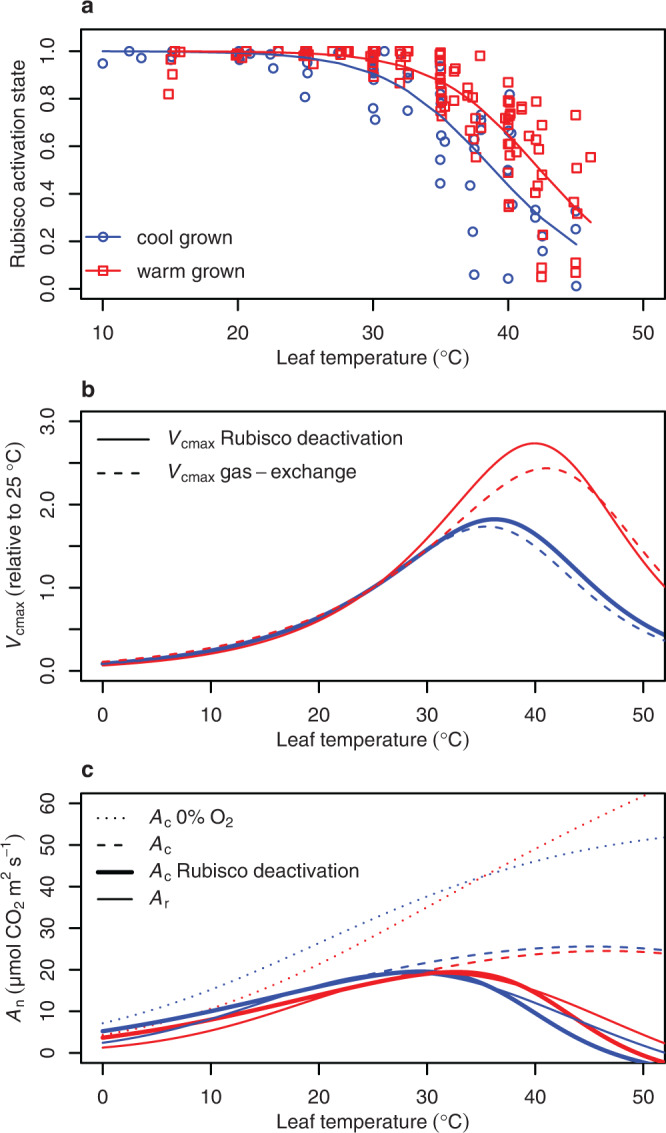
a A collection of published fractions of total functional Rubisco sites in a leaf (Rubisco activation state—points) at a given temperature (refer to Supplementary Table 2 for metadata). A Sharpe-Schoolfield equation (solid lines) accounting for enzyme deactivation at high temperature (Eq. 1) with the numerator set to unity was fit through non-linear least squares regression for both the cool (grown at <25 °C; blue circles) and warm (grown at >25 °C; red squares) growth datasets. b The temperature responses of Vcmax that we derived from Rubisco deactivation (solid lines) and the apparent Vcmax derived from gas-exchange estimates and an Arrhenius peaked model (dashed lines) with an acclimation parameter set at 24 and 36 °C based on Kattge and Knorr23. Cool (blue) or warm (red) grown species dependent on their day growth temperature being below or above 25 °C, respectively. c The net photosynthesis CO2 assimilation rate (An) predicted from carboxylation limited photosynthesis (Ac) modelled with no O2 (i.e. no photorespiration) and assuming Rubisco is totally active (dotted lines); or at 21% O2 but assuming Rubisco is totally active (dashed lines), accounting for Rubisco deactivation (bold solid lines), or assuming RuBP regeneration limited photosynthesis (Ar) based on chloroplast electron transport (J) and its response to temperature (solid lines).
Without a Rubisco deactivation term and in the absence of photorespiration (i.e. O2 parameterised to zero), modelled Ac did not reach a Topt below 50 °C for either cool or warm grown plants (Fig. 1c). When accounting for photorespiration (i.e. O2 parameterised to atmospheric concentrations of 21%) but not Rubisco deactivation, Ac reached a Topt at the relatively hot temperatures of 45.1 °C for cool and 46.6 °C for warm grown plants. With both photorespiration and Rubisco deactivation accounted for, the Topt of Ac was 29.4 °C for cool and 32.7 °C for warm grown plants and Ac declined sharply as temperatures exceeded these Topt (Fig. 1c). The use of Rubisco kinetics from a variety of studies led to variation in predicted rates of Ac, but the general pattern of decline due to Rubisco deactivation remained prominent (Supplementary Fig. 2). We further compared Ac to Ar predictions based on temperature response of J (Eq. 8 and Supplementary Table 1). J and its temperature response were derived from published observations comprising 26 species, 23 of which were either herb/grasses or temperate trees (Supplementary Table 3). Ar which factored in the temperature response of J declined at relatively mild temperatures, with a Topt of 28.1 °C and 31.9 °C for cool and warm grown plants, less than 1.5 °C difference from the corresponding Topt of Ac which included Rubisco deactivation (Fig. 1c).
Rubisco deactivation and electron transport can predict individual species and the global pattern of declines in An
To test the accuracy of the model which included Rubisco deactivation, Ac and Ar predictions were compared to observed An from 75 published temperature response curves comprising 49 C3 species (Supplementary Table 4). Observations and Ac modelled curves for species representing six major plant functional types demonstrated that high temperature-induced declines in An can be accurately predicted when Rubisco deactivation is accounted for (Fig. 2). However, significant inaccuracy was observed when modelling extremophiles. Deschampsia antarctica, an Antarctic grass that was grown at 12 °C, had a mean root mean squared error (RMSE) of 6.6 μmol CO2 m−2 s−1 due to it maintaining stable An to temperatures below 10 °C (Fig. 2a). A similar lack of predictability occurred for Larrea divaricata, an arid shrub that was grown at 44 °C, which had a mean RMSE of 8.9 μmol CO2 m−2 s−1 due to the modelled decline in An occurring ~8 °C before the observed decline (Fig. 2b). The lack of predictive power for these extremophiles occurred irrespective of whether predictions were derived from Ac with Rubisco deactivation or Ar models. There was limited difference in predictive power between cool and warm grown species or among plant functional types (Fig. 2c, d). The RMSE between predicted Ac and observed An was 2.1 μmol CO2 m−2 s−1 in cool grown species and 2.7 μmol CO2 m−2 s−1 in warm grown species. There was no significant difference (Welch Two Sample t test; df = 1124, t = −0.092, p = 0.93) in model predictions based on Ac that included Rubisco deactivation or based on Ar that included temperature dependence of J (Fig. 3 and Supplementary Fig. 3).
Fig. 2. Observations, modelled simulations, and the predictability of the leaf temperature response of net CO2 assimilation (An) for individual species and plant functional types.

a The An temperature response of four representative cool grown species: Triticum aestivum (herb/grass), Quercus pubescens (temperate tree), Picea mariana (needle-leaf tree), and Deschampsia antarctica (herb/grass). For D. antarctica, an extremophile that was grown at 12 °C, the broken line between observations highlights the deviation from the model prediction below 30 °C for this species. Points are observations and the curves are the Rubisco carboxylation limited assimilation rates (Ac) with Rubisco deactivation included parameterised to cool grown plants (solid blue lines), or RuBP regeneration limited CO2 assimilation rates (Ar) parameterised to cool grown plants (dotted blue lines). b Four representative warm grown species: Oryza sativa (grass), Cryptocarya laevigata (tropical tree), Eucalyptus camaldulensis (temperate tree), and Larrea divaricata (shrub). For L. divaricata, an extremophile that was grown at 44 °C, the broken line between observations highlights the deviation from the model prediction above 30 °C for this species. Points are observations and the curves are the Ac with Rubisco deactivation included for warm grown plants (solid red lines), or Ar for warm grown plants (dotted red lines). c, d Predictions of Ac which included Rubisco deactivation were plotted against corresponding observations for cool (c) and warm (d) grown plants. The coefficient of determination (r2), a 1:1 ratio (solid line), the root mean squared error (RMSE) between observed and predicted values (μmol m−2 s−1), and the bias in observations being greater than predictions (μmol m−2 s–1) for each growth environment are provided. Plant functional types are indicated by differing symbols and colours.
Fig. 3. Comparison of net photosynthetic CO2 assimilation predictions based on Rubisco carboxylation that included Rubisco deactivation (Ac) and rates of RuBP regeneration (Ar).
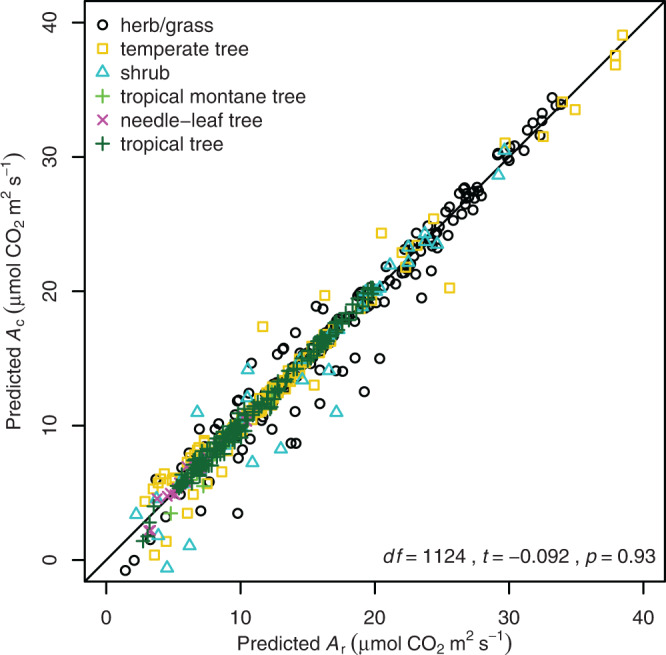
Values correspond to temperature and species observation predictions for the temperature response curves of 49 species previously published (Supplementary Table 4). Plant functional types are indicated by different symbols and colours. The degrees of freedom (df), t value (t), and p value (p) of a Welch Two Sample t test comparing Ac and Ar are presented in the graph. The solid line is the 1:1 ratio.
To further test the predictability of the model on a global scale that was independent of species, we compared model predictions with a composite curve comprised of the previously reported An temperature response observations relativised to their Topt (Fig. 4a). The predicted fall in Ac with rising temperature and the differences between cool and warm grown plants both closely aligned with observations. A similar prediction resulted based on Ar. We further compared the model incorporating Rubisco deactivation with a global composite temperature response curve generated from the mean An rates of a dataset totalling 13,876 individual gas-exchange observations from 311 species, representing a wide range of plant functional types34. The curve was developed by binning and averaging the observations for each degree of measured leaf temperature. The number of observations per degree was normally distributed, with a peak at 30 °C (Supplementary Fig. 4). Again, we found a close relationship between the observed temperature response of An and the model predictions, with a RMSE of 1.7 and 2.0 μmol CO2 m−2 s−1 for Ac and Ar warm grown predictions applied, respectively (Fig. 4b). Rubisco deactivation and the temperature response of J thus effectively predicted the peak and decline in An that occurs with rising leaf temperature on an interspecific level.
Fig. 4. The implications of Rubisco deactivation on net CO2 assimilation predictions.
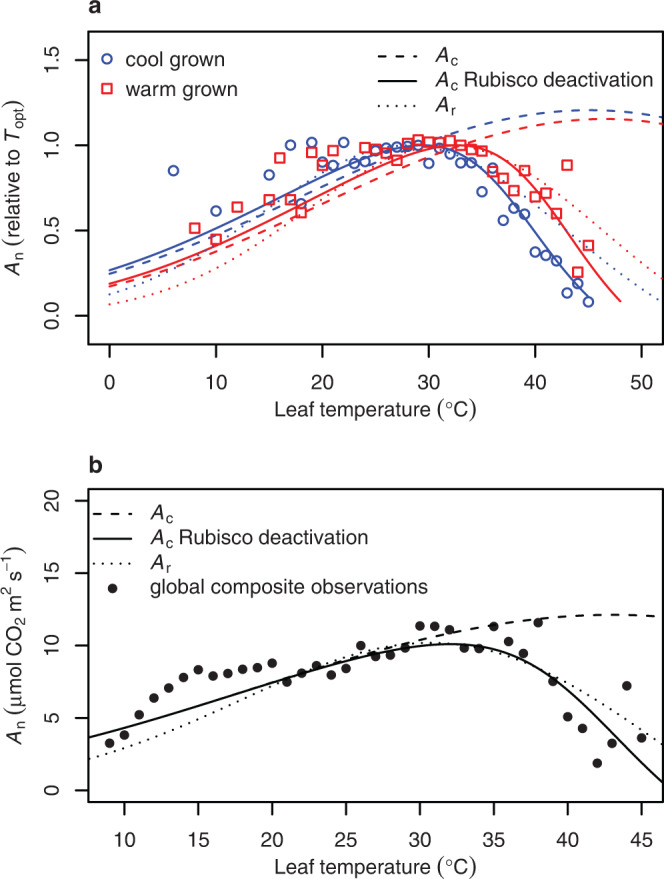
a Net CO2 assimilation rates (An) from temperature response curves representing 49 C3 plant species (Supplementary Table 4). Individual temperature response curves were relativised to the maximum rate achieved at the temperature optimum (Topt), and points represent the means of relativised values that were binned by degree Celsius. Observed points were plotted against Rubisco carboxylation limited assimilation (Ac) with Rubisco totally active (dashed lines), accounting for Rubisco deactivation (solid lines), or assuming RuBP regeneration limited photosynthesis (Ar) based on chloroplast electron transport (J) and its response to temperature (dotted lines). Rubisco kinetics were categorised as cool (blue) or warm (red) grown species dependent on their day growth temperature being below or above 25 °C, respectively. b A composite temperature response curve (closed circles) based on a global dataset comprising of 311 species and 13,876 observations34. Mean An values, independent of species and experiment, were binned by degree Celsius. Observed points were plotted against Rubisco carboxylation limited assimilation (Ac) with Rubisco totally active (dashed lines), accounting for Rubisco deactivation (solid lines), or assuming RuBP regeneration limited photosynthesis (Ar) based on chloroplast electron transport (J) and its response to temperature (dotted lines).
Discussion
Improved understanding of why photosynthesis is impaired by even moderate heat stress is needed if we are to accurately account for the influence of rising atmospheric temperatures on global vegetation. By accounting for the temperature-dependent change in the activation state of Rubisco, we were able to accurately predict warming-induced declines in An on an individual species, biome, and global interspecific level. The declines in An that were predicted based on Ac and by accounting for Rubisco deactivation were not significantly different from declines predicted by Ar when accounting for the interspecific temperature dependence of J. It has recently become apparent that Ac and Ar are optimised to co-limit An at a given growth temperature across a wide-range of species35, 36. Our results support a continuation of Ac and Ar co-limitation on shorter timeframes of one hour or less when leaf temperatures rise above the Topt.
Recent modelling on a broad range of higher plants has pointed towards Topt adapting and acclimating to growth temperature due to photosynthetic biochemistry rather than CO2 conductance limitations37. Similarly, by assuming a constant CO2 conductance in our model and thus removing the effect of water stress, we demonstrated that heat stress per se can explain declines in An beyond Topt. The study by Lin et al.34, from which we obtained the global composite An temperature response curve (Fig. 4b), demonstrated that stomatal conductance is regulated to maximise woody tissue development, including the allowance of greater water loss at warmer growth temperatures in wet environments. There is growing evidence that many plants with access to water keep their stomata open despite high air temperature and vapour pressure deficit as a means of transpirational cooling38–40. Therefore, there appear to be instances when plants prioritise thermoregulation over managing drought risks, although whether transpiration can reduce leaf temperatures below that of the surrounding air may be limited in natural sunlit canopies due to biophysical factors such as radiative heating41. Although we removed the confounding influence of water stress in this study, water availability remains influential in reducing An in species that close their stomata to preserve water, particularly during hot and dry conditions when vapour pressure deficit is high and soil moisture is low16,39. Indeed, our amended Ac model overestimated the temperature at which An began to decline when compared to tropical tree and lianas species in Panama (Supplementary Fig. 5). This was consistent with the published declines in stomatal conductance and intercellular CO2 concentrations in response to leaf heating for these same species17. Another aspect of CO2 conductance that can influence An is the rate of CO2 diffusion from intercellular airspaces to the site of chloroplasts, termed mesophyll conductance. Mesophyll conductance appears to either increase or remain constant with short-term rises in leaf temperature across many species42. This will contribute to intercellular CO2 drawdown and may exacerbate Rubisco CO2 substrate limitations when water is limited and temperatures hot.
Rubisco specificity for CO2 significantly shapes An under moderate, sustained warming, and photorespiration becomes a greater contributor to CO2 loss as leaf temperature rises (Fig. 1c). However, photorespiration cannot account for the extent of An decline that occurs as leaf temperature exceeds Topt (Fig. 1c). Indeed, model predictions of An that account for photorespiration but not for Rubisco deactivation or J limitations far exceed observed An, estimating a Topt of 45 °C in cool grown plants (Fig. 1c). It is therefore unlikely that Rubisco deactivation and J are downregulated as a mechanism to limit photorespiratory CO2 loss considering both contribute far more to declines in An above Topt than photorespiration does, assuming there is no other cost of greater photorespiratory 2-phosphoglycolate metabolism apart from CO2 release. Rather, unavoidable heat damage to membranes and proteins likely set the thermal limits in Rubisco and J capacity.
There is extensive literature linking Rubisco deactivation to the thermolability of Rca43. Rubisco deactivation with rising temperature is attributed to loss of Rca, due to the role of Rca in maintaining functional Rubisco catalytic sites and it being characteristically susceptible to degradation under relatively mild heat44. Our findings are consistent with the link between Rubisco deactivation and Rca, as our cool grown T0.5 of 39.0 °C is close to the 35-38 °C range within which isolated Rca from temperate wheat (Triticum aestivum, L.) loses 50% of its functionality33,45. Similarly, the warm grown T0.5 of 42.4 °C is within the 40 to 43 °C range in which isolated Rca from warm grown rice (Oryza sativa, L.) loses 50% of its ability to activate Rubisco33,46. The difference in T0.5 between cool and warm grown observations in the global activation state data (Fig. 1a) reflects the previously documented pattern of Rca acclimating and adapting to the prevailing growth temperature, including becoming more thermally stable in hotter environments32,33.
The dynamic and reversible decline in J at high temperatures47 has been linked to heat susceptibility of thylakoid membranes and their constituents7,11,48. The oxygen evolving complex of photosystem II (PSII) and the cytochrome b6/f complex (Cyt b6/f ) seem particularly important in setting dynamic temperature-effected rates of J49–51. There are four Mn atoms per PSII reaction centre responsible for oxidation of H2O. Mn is held by 33 kDa D1 proteins. Prolonged heat stress can dislodge D1 proteins and subsequently Mn2+ ions from the oxygen evolving complex of higher plants, resulting in a decline in J52–54. The disruption in electron accepting ability of PSII leads to the reaction centre being overly oxidised (P680+) and conducive to ROS formation, which can impair D1 protein synthesis and further diminishes PSII functionality55. The heat sensitivity of the PSII oxygen evolving complex makes it a key reason for why J declines under high leaf temperatures. Heat damage to thylakoid membranes is not confined to PSII. Moderate temperature of 40 °C is shown to disrupt the balance of electron flow between PSII and PSI which is controlled by Cyt b6/f, and through damage or regulation, the flow of electrons through Cyt b6/f leads to an overreduction of PSI upon heat exposure56. With an overly reduced PSI, cyclic electron flow is upregulated as a means of dissipating electrons and preventing irreversibly damage to the stroma7,56,57. However, cyclic electron flow is insufficient to maintain An in the absence of linear electron flow since it does not produce the necessary NADPH to run the Calvin-Benson cycle57. This is a strong indicator that the imbalance in electron flow as temperatures exceed the Topt contributes to declining J and subsequently Ar.
The alignment of Rubisco deactivation and declines in J suggest a closely aligned temperature limitation on the functionality of both, a tight temperature dependent regulation of one to a limitation in the other, or a combination of the two. Rca activity is modulated by ATP and inhibited by competitive binding of ADP58–60. Declines in J with rising temperature due to electron transport imbalance, leakiness of thylakoid membranes, or other damage likely reduce stromal ATP concentrations. Lower stromal ATP concentrations reduce the active state of Rubisco61, presumably through reduced Rca activity. Conversely, a lack of CO2 fixation by Rubisco due to heat instability of Rca may lead to an accumulation of RuBP, reductant, and ATP. Recent analysis suggests that Cyt b6/f tightly controls the dynamic flow of electrons between PSII and PSI, thus an accumulation of reductant and ATP would quickly downregulate Cyt b6/f electron transfer51. Interestingly, Rca has previously been found to associate with thylakoid membranes under heat stress in spinach (Spinacia oleracea, L.)62, and a recent report in rice noted a reduction in the quantum yield of photosystem I with overexpression of Rca63. Whether Rca and components of the electron transport chain interact directly during heat perturbation to coordinate downregulation of photosynthesis with rising temperature requires further exploration.
Model predictions based on Ac that included Rubisco deactivation and Ar were accurate across a wide range of cool and warm grown higher plant species from a range of plant functional biomes (Figs. 2 and 4). However, the model fits were poor in relation to predicting An of plants adapted to extreme cold and heat. The An temperature responses of Deschampsia antarctica, a small grass native to antarctica, and Larrea divaricata, a desert shrub, diverged from model predictions (Fig. 2). This suggests that extremophiles like D. antarctica and L. divaricata may have unusually cold or heat stable photosynthetic constituents. For example, L. divaricata may have a variant of Rca similar to that of the CAM plant Agave tequilana which has an Rca isoform that remains active up to 50 °C32. Thus, while our model may not be applicable to extremophiles, it may provide a novel means of identifying species with superior thermal stability of photosynthetic components, as indicated by observed An of a species far exceeding the model predictions.
In conclusion, we have demonstrated the importance of accounting for Rubisco deactivation when modelling photosynthesis above the Topt. By doing so, the model we presented more accurately predicted previously observed declines in An with rising leaf temperature across a broad range of higher plant species and functional types. Our predictions of An based on Rubisco deactivation are in close agreement with An predicted from the temperature dependence of J, suggesting both are likely highly coordinated and co-limit photosynthesis as temperatures rise. Attempts to engineer improvement in photosynthesis at high temperature should thus focus on both Rubisco and electron transport characteristics, as a benefit to one without a benefit to the other is likely to result in only incremental improvements in heat tolerance. Although Topt is known to shift with growth temperature at a finer scale than simply below or above 25 °C, the limited number of published Rubisco activation state and J temperature curves prohibits model parameterisation that would allow predictions of finer scale adjustments in An to changing growth temperature. Further studies that characterise the temperature dependence of Rubisco deactivation and temperature dependence of J – ideally from a wide spread of plant functional types – will improve the accuracy of the models we present. Finally, we demonstrated that neither CO2 substrate supply limitation nor photorespiratory CO2 loss was needed to explain high temperature-induced decreases in An. However, many future heatwaves are likely to coincide with drought, and drought will reduce CO2 conductance and increase photorespiratory CO2 loss, exacerbating the stress caused by Rubisco deactivation and declines in J.
Methods
Acquisition of Rubisco activation state, electron transport, and CO2 assimilation rate data
Published temperature response curves of Rubisco activation state, chloroplast electron transport rate ( J ), and net CO2 assimilation rate per unit leaf area (An) for individual species were collated through searching the published scientific literature. Metadata relating to the publication and conditions of measurement are provided in Supplementary Tables 2, 3 and 4. Data were either extracted directly from tables or by recording values from graphs using the data extraction package “digitize-package” in R statistical software64 following package instructions. For Rubisco activation data, papers that biochemically characterised the Rubisco activation state to increasing leaf measuring temperature were collated. Values were normalised to the maximum recorded value in each temperature curve. Data were separately analysed based on the mean day growth temperature being below (cool) or above (warm) 25 °C. Where plants were grown at 25 °C, cool or warm grown distinctions were made based on whether the peak in photosynthesis was below or above 30 °C, respectively. For An observational data, studies that measured An on a leaf area basis equal or less than one hour after increasing leaf measuring temperature were included. Where light, CO2 concentrations, or nitrogen were varied, we only used curves with the greatest light or nitrogen application and CO2 concentrations corresponding to ambient concentrations of 400 ± 50 μmol mol−1. Only curves with more than four temperature data points were included.
The interspecific temperature response curve comprising observations obtained from 310 species (Fig. 4b) was based on a global set of gas-exchange data as presented in Lin et al. (2015)34. An values and the leaf temperature at which they were recorded were binned per °C and the mean used to generate a global temperature response curve. Observations relating to Panama tropical species were those presented by Slot and Winter (2017)17. The global gas-exchange data and that of Panama tropical species were accessed through the TRY database65.
Modelling of CO2 assimilation
Net photosynthetic CO2 fixation (Ac) was calculated using the FvCB C3 photosynthesis model4,29 using the equation:
| 2 |
The CO2 partial pressure at the site of fixation (C) was assumed to be an ambient atmospheric CO2 partial pressure (40 Pa for current projections or as stated otherwise) multiplied by 0.7 to account for assumed intercellular CO2 drawdown. We further assumed an infinite mesophyll conductance and no CO2 conductance response to temperature. The partial pressure of oxygen in the atmosphere (O) was either set to ambient atmospheric partial pressure (21 kPa) or zero when assessing effects of photorespiration on An. Respiration in the light (RL) was assumed to be 70% of dark respiration which was 1.29 μmol m−2 s−1 at 25 °C calculated from the relationship between nitrogen and respiration as presented in Atkin et al.66, and its response to rising temperature calculated using a global quadratic model67. The CO2 compensation point in the absence of RL (Γ*; Pa), the Michaelis–Menten constant of Rubisco for CO2 (Kc; Pa) and O2 (Ko; kPa), and Rubisco kcat (CO2 s−1) at a given temperature were calculated using the Arrhenius equation:
| 3 |
where R is the gas constant (8.314 J K−1 mol−1), T is the leaf temperature in degrees Celsius and TK the leaf temperature in Kelvin. P25 is the parameter value at 25 °C and Ea the activation energy in J mol−1. The P25 and Ea values were based on multiple species both in vitro and in vivo presented in Galmés et al.68 and Orr et al.69 (Supplementary Table 1 and Supplementary Fig. 1). In regards to Orr et al.69, cool was separated from warm growth kinetics by selecting species with a maximum temperature of the warmest quartile of below 25 °C or ≥ 25 °C, respectively. In regards to the Galmés et al.68 dataset, cool was separated from warm growth kinetics by whether the maximum daily growth temperature was below 25 °C or ≥ 25 °C, respectively. Rubisco kinetics measured in vitro were converted from molar concentrations to partial pressures using Henry’s law for solubilities and their temperature dependence:
| 4 |
Where H° is the solubility at 25 °C (0.034 and 0.0013 mol L−1 atm−1 for CO2 and O2, respectively), -∆H describes the temperature dependency of solubility (2400 and 1700 for CO2 and O2, respectively).
Based on the expected number of Rubisco catalytic sites (n), and the kcat of Rubisco at a given temperature determined above, we calculated Vcmax as:
| 5 |
When models were fitted to individual species or combined species photosynthesis observations, the n was obtained by solving the Eqs. 2 and 5 using parameters and the An measured at 22 °C or closest observation above (temperatures where no Rubisco deactivation was observed):
| 6 |
For a species independent estimate, n was 26 μmol m−2 based on 1.8 g m−2 of leaf Rubisco content across a range of species70 and a molecular weight of 70,000 g per active site71. To model the impact of RuBP regeneration-dependent assimilation (Ar)—the other commonly attributed limitation on An at high temperatures29—we used the equation:
| 7 |
with the photosynthetic electron transport rate (J) and its temperature response modelled and parameterised using the equation of June et al.47:
| 8 |
Where J(To) is the electron transport rate (μmol e m−2 s−1) at its temperature optimum (To), and Ω is the difference in temperature from To at which J declines to e−1 (0.37)J(To). J(To), To and Ω were the means of multiple species reported in June et al. (2004)47 or derived from non-linear least squares fits of Eq. 8 to other published temperature response curves of J listed in Supplementary Table 3. Additionally, To and Ω were supplemented with relativised temperature response curves of the quantum efficiency of PSII (ϕPSII). We considered the relative changes in ϕPSII to short-term changes in leaf temperature as equivalent to relative changes in J. Again, we separated the analysis into cool and warm grown plants based on mean day growth temperatures as outlined above. When fitting Ar to individual species or combined species photosynthesis observations, we solved for J(To) using Eqs. 7 and 8 using parameters and the An measured at 22 °C or closest observation above:
| 9 |
Vcmax based on kcat and Rubisco deactivation (Eq. 1) were compared with the peaked Arrhenius model of Vcmax which is based on the temperature response of apparent Vcmax derived from gas-exchange measurements and adjusted for growth temperature as described by Kattge and Knorr23:
| 10 |
Where Vcmax25 was the value of Vcmax at a reference temperature (Tref) of 25 °C and set to 1. Hd was the deactivation enthalpy and was set as 200 kJ mol−1. The activation enthalpy (Ha) in J mol−1 was calculated as 82,992-632×Tgrowth. The entropy factor (∆S) in J mol−1 was calculated as 668.39-1.07×Tgrowth. Growth temperature (Tgrowth) was set to either 24 or 36 °C which provided the closest fit to Vcmax derived from Eq. 1 for cool and warm grown plants.
All modelling and analysis were performed using R v.4.1.2 (2021-11-01) statistical software72 with Rstudio graphical interface73. The R package readxl (v.1.3.1)74 (https://CRAN.R-project.org/package=readxl) was used to analyse excel data files, and the package Metrics (v.0.1.4)75 (https://CRAN.R-project.org/package=Metrics) was used to calculate root mean squared error (RMSE) and the bias between observed and predicted values.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Supplementary information
Acknowledgements
This study was supported by an Australian Research Council DP22 grant (DP220101882 provided to A.P.S. and O.K.A.). We thank Assoc. Prof. Danielle Way for providing valuable feedback on the manuscript.
Author contributions
A.P.S. conceived and drafted the initial manuscript. B.C.P., J.R.E., G.D.F., and O.K.A. provided substantive revisions and guidance in interpretation of the data.
Peer review
Peer review information
Nature Communications thanks Elizabete Carmo-Silva, Thomas Sharkey and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Data availability
The author and DOI information relating to the published articles from which all Rubisco kinetic, Rubisco activation state, net CO2 assimilation, and electron transport rate temperature response curve data where analysed are presented in Supplementary Tables 1, 2, 3, and 4. The collated data are provided at 10.6084/m9.figshare.22661989 and with the model code (10.5281/zenodo.7683559). The global individual An vs leaf temperature34 and Panama species dataset17 are available from the TRY plant trait database http://www.try-db.org/TryWeb/Data.php under the dataset names “Global Leaf Gas Exchange Database (I)” and “Photosynthesis Temperature Response Panama”.
Code availability
A model script and An data are accessible through GitHub (10.5281/zenodo.7683559). The script provides model fits to An temperature response curves using relevant model parameters based on whether the plant was grown below or above 25 °C. The script will generate graphs and provide indices of how well observed and model data match for each curve fit, comparable to the analysis presented in this study.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
The online version contains supplementary material available at 10.1038/s41467-023-38496-4.
References
- 1.Teskey R, et al. Responses of tree species to heat waves and extreme heat events. Plant Cell Environ. 2015;38:1699–1712. doi: 10.1111/pce.12417. [DOI] [PubMed] [Google Scholar]
- 2.Berry J, Björkman O. Photosynthetic response and adaptation to temperature in higher plants. Annu. Rev. Plant Physiol. 1980;31:491–543. doi: 10.1146/annurev.pp.31.060180.002423. [DOI] [Google Scholar]
- 3.Yamori W, Hikosaka K, Way D. Temperature response of photosynthesis in C3, C4, and CAM plants: temperature acclimation and temperature adaptation. Photosynth. Res. 2014;119:101–117. doi: 10.1007/s11120-013-9874-6. [DOI] [PubMed] [Google Scholar]
- 4.Farquhar GD, von Caemmerer S, Berry JA. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta. 1980;149:78–90. doi: 10.1007/BF00386231. [DOI] [PubMed] [Google Scholar]
- 5.Sharkey TD, Bernacchi CJ, Farquhar GD, Singsaas EL. Fitting photosynthetic carbon dioxide response curves for C3 leaves. Plant Cell Environ. 2007;30:1035–1040. doi: 10.1111/j.1365-3040.2007.01710.x. [DOI] [PubMed] [Google Scholar]
- 6.Sage RF, Kubien DS. The temperature response of C3 and C4 photosynthesis. Plant Cell Environ. 2007;30:1086–1106. doi: 10.1111/j.1365-3040.2007.01682.x. [DOI] [PubMed] [Google Scholar]
- 7.Sharkey TD. Effects of moderate heat stress on photosynthesis: importance of thylakoid reactions, rubisco deactivation, reactive oxygen species, and thermotolerance provided by isoprene. Plant Cell Environ. 2005;28:269–277. doi: 10.1111/j.1365-3040.2005.01324.x. [DOI] [Google Scholar]
- 8.Busch FA, Sage RF. The sensitivity of photosynthesis to O2 and CO2 concentration identifies strong Rubisco control above the thermal optimum. N. Phytol. 2017;213:1036–1051. doi: 10.1111/nph.14258. [DOI] [PubMed] [Google Scholar]
- 9.Salvucci ME, Crafts-Brandner SJ. Inhibition of photosynthesis by heat stress: the activation state of Rubisco as a limiting factor in photosynthesis. Physiol. Plant. 2004;120:179–186. doi: 10.1111/j.0031-9317.2004.0173.x. [DOI] [PubMed] [Google Scholar]
- 10.Crafts-Brandner SJ, Salvucci ME. Rubisco activase constrains the photosynthetic potential of leaves at high temperature and CO2. Proc. Natl Acad. Sci. USA. 2000;97:13430–13435. doi: 10.1073/pnas.230451497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cen Y-P, Sage RF. The regulation of rubisco activity in response to variation in temperature and atmospheric CO2 partial pressure in sweet potato. Plant Physiol. 2005;139:979–990. doi: 10.1104/pp.105.066233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Yamori W, Nagai T, Makino A. The rate-limiting step for CO2 assimilation at different temperatures is influenced by the leaf nitrogen content in several C3 crop species. Plant Cell Environ. 2011;34:764–777. doi: 10.1111/j.1365-3040.2011.02280.x. [DOI] [PubMed] [Google Scholar]
- 13.Yamori W, Noguchi K, Hikosaka K, Terashima I. Phenotypic plasticity in photosynthetic temperature acclimation among crop species with different cold tolerances. Plant Physiol. 2010;152:388–399. doi: 10.1104/pp.109.145862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sage RF. A model describing the regulation of ribulose-1,5-bisphosphate carboxylase, electron transport, and triose phosphate use in response to light intensity and CO2 in C3 plants 1. Plant Physiol. 1990;94:1728–1734. doi: 10.1104/pp.94.4.1728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Sage RF, Sharkey TD, Seemann JR. Regulation of ribulose-1,5-bisphosphate carboxylase activity in response to light intensity and CO2 in the C3 annuals Chenopodium album L. and Phaseolus vulgarisem L. Plant Physiol. 1990;94:1735–1742. doi: 10.1104/pp.94.4.1735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Grossiord C, et al. Plant responses to rising vapor pressure deficit. N. Phytol. 2020;226:1550–1566. doi: 10.1111/nph.16485. [DOI] [PubMed] [Google Scholar]
- 17.Slot M, Winter K. In situ temperature response of photosynthesis of 42 tree and liana species in the canopy of two Panamanian lowland tropical forests with contrasting rainfall regimes. N. Phytol. 2017;214:1103–1117. doi: 10.1111/nph.14469. [DOI] [PubMed] [Google Scholar]
- 18.Wong SC, et al. Humidity gradients in the air spaces of leaves. Nat. Plants. 2022;8:971–978. doi: 10.1038/s41477-022-01202-1. [DOI] [PubMed] [Google Scholar]
- 19.Dusenge ME, Duarte AG, Way DA. Plant carbon metabolism and climate change: elevated CO2 and temperature impacts on photosynthesis, photorespiration and respiration. N. Phytol. 2019;221:32–49. doi: 10.1111/nph.15283. [DOI] [PubMed] [Google Scholar]
- 20.Scafaro AP, et al. Responses of leaf respiration to heatwaves. Plant Cell Environ. 2021;44:2090–2101. doi: 10.1111/pce.14018. [DOI] [PubMed] [Google Scholar]
- 21.Badger MR, Collatz GJ. Studies on the kinetic mechanism of Ribulose-1,5-bisphosphate carboxylase and oxygenase reactions, with particular reference to the effect of temperature on kinetic parameters. Carnegie Inst. Wash. Yearb. 1978;76:355–362. [Google Scholar]
- 22.Rogers A. The use and misuse of Vc,max in Earth System Models. Photosynth. Res. 2013;119:15–29. doi: 10.1007/s11120-013-9818-1. [DOI] [PubMed] [Google Scholar]
- 23.Kattge J, Knorr W. Temperature acclimation in a biochemical model of photosynthesis: a reanalysis of data from 36 species. Plant Cell Environ. 2007;30:1176–1190. doi: 10.1111/j.1365-3040.2007.01690.x. [DOI] [PubMed] [Google Scholar]
- 24.Medlyn BE, et al. Temperature response of parameters of a biochemically based model of photosynthesis. II. A review of experimental data. Plant Cell Environ. 2002;25:1167–1179. doi: 10.1046/j.1365-3040.2002.00891.x. [DOI] [Google Scholar]
- 25.Salvucci ME, Osteryoung KW, Crafts-Brandner SJ, Vierling E. Exceptional sensitivity of rubisco activase to thermal denaturation in vitro and in vivo. Plant Physiol. 2001;127:1053–1064. doi: 10.1104/pp.010357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Salvucci ME, Crafts-Brandner SJ. Relationship between the heat tolerance of photosynthesis and the thermal stability of rubisco activase in plants from contrasting thermal environments. Plant Physiol. 2004;134:1460–1470. doi: 10.1104/pp.103.038323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Yamori W, Suzuki K, Noguchi KO, Nakai M, Terashima I. Effects of Rubisco kinetics and Rubisco activation state on the temperature dependence of the photosynthetic rate in spinach leaves from contrasting growth temperatures. Plant Cell Environ. 2006;29:1659–1670. doi: 10.1111/j.1365-3040.2006.01550.x. [DOI] [PubMed] [Google Scholar]
- 28.Portis A. Rubisco activase – rubisco’s catalytic chaperone. Photosynth. Res. 2003;75:11–27. doi: 10.1023/A:1022458108678. [DOI] [PubMed] [Google Scholar]
- 29.von Caemmerer S. Biochemical Models of Leaf Photosynthesis (CSIRO Publishing, 2000).
- 30.Sage RF, Way DA, Kubien DS. Rubisco, Rubisco activase, and global climate change. J. Exp. Bot. 2008;59:1581–1595. doi: 10.1093/jxb/ern053. [DOI] [PubMed] [Google Scholar]
- 31.Schoolfield RM, Sharpe PJH, Magnuson CE. Non-linear regression of biological temperature-dependent rate models based on absolute reaction-rate theory. J. Theor. Biol. 1981;88:719–731. doi: 10.1016/0022-5193(81)90246-0. [DOI] [PubMed] [Google Scholar]
- 32.Shivhare, D. & Mueller-Cajar, O. In vitro characterization of thermostable CAM Rubisco activase reveals a Rubisco interacting surface loop. Plant Physiol.174, 1505–1516 (2017). [DOI] [PMC free article] [PubMed]
- 33.Scafaro AP, Bautsoens N, den Boer B, Van Rie J, Gallé A. A conserved sequence from heat-adapted species improves rubisco activase thermostability in wheat. Plant Physiol. 2019;181:43. doi: 10.1104/pp.19.00425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lin Y-S, et al. Optimal stomatal behaviour around the world. Nat. Clim. Chang. 2015;5:459–464. doi: 10.1038/nclimate2550. [DOI] [Google Scholar]
- 35.Chen J-L, Reynolds JF, Harley PC, Tenhunen JD. Coordination theory of leaf nitrogen distribution in a canopy. Oecologia. 1993;93:63–69. doi: 10.1007/BF00321192. [DOI] [PubMed] [Google Scholar]
- 36.Wang H, et al. Towards a universal model for carbon dioxide uptake by plants. Nat. Plants. 2017;3:734–741. doi: 10.1038/s41477-017-0006-8. [DOI] [PubMed] [Google Scholar]
- 37.Kumarathunge DP, et al. Acclimation and adaptation components of the temperature dependence of plant photosynthesis at the global scale. N. Phytol. 2019;222:768–784. doi: 10.1111/nph.15668. [DOI] [PubMed] [Google Scholar]
- 38.Drake JE, et al. Trees tolerate an extreme heatwave via sustained transpirational cooling and increased leaf thermal tolerance. Glob. Change Biol. 2018;24:2390–2402. doi: 10.1111/gcb.14037. [DOI] [PubMed] [Google Scholar]
- 39.Lin H, Chen Y, Zhang H, Fu P, Fan Z. Stronger cooling effects of transpiration and leaf physical traits of plants from a hot dry habitat than from a hot wet habitat. Funct. Ecol. 2017;31:2202–2211. doi: 10.1111/1365-2435.12923. [DOI] [Google Scholar]
- 40.Aparecido LM, Woo S, Suazo C, Hultine KR, Blonder B. High water use in desert plants exposed to extreme heat. Ecol. Lett. 2020;23:1189–1200. doi: 10.1111/ele.13516. [DOI] [PubMed] [Google Scholar]
- 41.Still CJ, et al. No evidence of canopy-scale leaf thermoregulation to cool leaves below air temperature across a range of forest ecosystems. Proc. Natl Acad. Sci. USA. 2022;119:e2205682119. doi: 10.1073/pnas.2205682119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.von Caemmerer S, Evans JR. Temperature responses of mesophyll conductance differ greatly between species. Plant Cell Environ. 2015;38:629–637. doi: 10.1111/pce.12449. [DOI] [PubMed] [Google Scholar]
- 43.Carmo-Silva E, Scales JC, Madgwick PJ, Parry MA. Optimizing Rubisco and its regulation for greater resource use efficiency. Plant Cell Environ. 2015;38:1817–1832. doi: 10.1111/pce.12425. [DOI] [PubMed] [Google Scholar]
- 44.Salvucci ME, Crafts-Brandner SJ. Mechanism for deactivation of Rubisco under moderate heat stress. Physiol. Plant. 2004;122:513–519. doi: 10.1111/j.1399-3054.2004.00419.x. [DOI] [Google Scholar]
- 45.Degen GE, Worrall D, Carmo-Silva E. An isoleucine residue acts as a thermal and regulatory switch in wheat Rubisco activase. Plant J. 2020;103:742–751. doi: 10.1111/tpj.14766. [DOI] [PubMed] [Google Scholar]
- 46.Shivhare, D., Ng, J., Tsai, Y.-C. C. & Mueller-Cajar, O. Probing the rice Rubisco–Rubisco activase interaction via subunit heterooligomerization. Proc. Natl Acad. Sci. USA116, 24041–24048 (2019). [DOI] [PMC free article] [PubMed]
- 47.June T, Evans JR, Farquhar GD. A simple new equation for the reversible temperature dependence of photosynthetic electron transport: a study on soybean leaf. Funct. Plant Biol. 2004;31:275–283. doi: 10.1071/FP03250. [DOI] [PubMed] [Google Scholar]
- 48.Allakhverdiev S, et al. Heat stress: an overview of molecular responses in photosynthesis. Photosynth. Res. 2008;98:541–550. doi: 10.1007/s11120-008-9331-0. [DOI] [PubMed] [Google Scholar]
- 49.Takahashi S, Badger MR. Photoprotection in plants: a new light on photosystem II damage. Trends Plant Sci. 2011;16:53–60. doi: 10.1016/j.tplants.2010.10.001. [DOI] [PubMed] [Google Scholar]
- 50.Yamori W, et al. The roles of ATP synthase and the cytochrome b6/f complexes in limiting chloroplast electron transport and determining photosynthetic capacity. Plant Physiol. 2011;155:956–962. doi: 10.1104/pp.110.168435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Johnson JE, Berry JA. The role of Cytochrome b6f in the control of steady-state photosynthesis: a conceptual and quantitative model. Photosynth. Res. 2021;148:101–136. doi: 10.1007/s11120-021-00840-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Enami I, et al. Is the primary cause of thermal inactivation of oxygen evolution in spinach PS II membranes release of the extrinsic 33 kDa protein or of Mn? Biochim. Biophys. Acta. 1994;1186:52–58. doi: 10.1016/0005-2728(94)90134-1. [DOI] [Google Scholar]
- 53.Yoshioka M, et al. Quality control of photosystem II: cleavage of reaction center D1 protein in spinach thylakoids by ftsh protease under moderate heat stress. J. Biol. Chem. 2006;281:21660–21669. doi: 10.1074/jbc.M602896200. [DOI] [PubMed] [Google Scholar]
- 54.Vinyard DJ, Ananyev GM, Dismukes GC. Photosystem II: the reaction center of oxygenic photosynthesis. Annu. Rev. Biochem. 2013;82:577–606. doi: 10.1146/annurev-biochem-070511-100425. [DOI] [PubMed] [Google Scholar]
- 55.Takahashi S, Murata N. How do environmental stresses accelerate photoinhibition? Trends Plant Sci. 2008;13:178–182. doi: 10.1016/j.tplants.2008.01.005. [DOI] [PubMed] [Google Scholar]
- 56.Zhang R, Sharkey TD. Photosynthetic electron transport and proton flux under moderate heat stress. Photosynth. Res. 2009;100:29–43. doi: 10.1007/s11120-009-9420-8. [DOI] [PubMed] [Google Scholar]
- 57.Yamori W, Shikanai T. Physiological functions of cyclic electron transport around photosystem I in sustaining photosynthesis and plant growth. Annu. Rev. Plant Biol. 2016;67:81–106. doi: 10.1146/annurev-arplant-043015-112002. [DOI] [PubMed] [Google Scholar]
- 58.Scafaro AP, et al. A single point mutation in the C-terminal extension of wheat Rubisco activase dramatically reduces ADP inhibition via enhanced ATP binding affinity. J. Biol. Chem. 2019;294:17931–17940. doi: 10.1074/jbc.RA119.010684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Zhang N, Portis AR. Mechanism of light regulation of Rubisco: a specific role for the larger Rubisco activase isoform involving reductive activation by thioredoxin-f. Proc. Natl Acad. Sci. USA. 1999;96:9438–9443. doi: 10.1073/pnas.96.16.9438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Zhang N, Schürmann P, Portis A., Jr. Characterization of the regulatory function of the 46-kDa isoform of Rubisco activase from Arabidopsis. Photosynth. Res. 2001;68:29–37. doi: 10.1023/A:1011845506196. [DOI] [PubMed] [Google Scholar]
- 61.Robinson SP, Portis AR., Jr. Involvement of stromal ATP in the light activation of ribulose-1,5-bisphosphate carboxylase/oxygenase in intact isolated chloroplasts. Plant Physiol. 1988;86:293–298. doi: 10.1104/pp.86.1.293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Rokka A, Zhang L, Aro E-M. Rubisco activase: an enzyme with a temperature-dependent dual function? Plant J. 2001;25:463–471. doi: 10.1046/j.1365-313x.2001.00981.x. [DOI] [PubMed] [Google Scholar]
- 63.Suganami M, et al. Expression of flavodiiron protein rescues defects in electron transport around PSI resulting from overproduction of Rubisco activase in rice. J. Exp. Bot. 2022;73:2589–2600. doi: 10.1093/jxb/erac035. [DOI] [PubMed] [Google Scholar]
- 64.Poisot T. The digitize package: extracting numerical data from scatterplots. R. J. 2011;3:25–26. doi: 10.32614/RJ-2011-004. [DOI] [Google Scholar]
- 65.Kattge J, et al. TRY – a global database of plant traits. Glob. Chang. Biol. 2011;17:2905–2935. doi: 10.1111/j.1365-2486.2011.02451.x. [DOI] [Google Scholar]
- 66.Atkin, O. K. et al. in Plant Respiration: Metabolic Fluxes and Carbon Balance (eds Tcherkez, G. & Ghashghaie, J.) 107–142 (Springer International Publishing, 2017).
- 67.Heskel MA, et al. Convergence in the temperature response of leaf respiration across biomes and plant functional types. Proc. Natl Acad. Sci. USA. 2016;113:3832–3837. doi: 10.1073/pnas.1520282113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Galmés J, Hermida-Carrera C, Laanisto L, Niinemets Ü. A compendium of temperature responses of Rubisco kinetic traits: variability among and within photosynthetic groups and impacts on photosynthesis modeling. J. Exp. Bot. 2016;67:5067–5091. doi: 10.1093/jxb/erw267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Orr DJ, et al. Surveying rubisco diversity and temperature response to improve crop photosynthetic efficiency. Plant Physiol. 2016;172:707–717. doi: 10.1104/pp.16.00750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Onoda Y, et al. Physiological and structural tradeoffs underlying the leaf economics spectrum. N. Phytol. 2017;214:1447–1463. doi: 10.1111/nph.14496. [DOI] [PubMed] [Google Scholar]
- 71.Spreitzer RJ, Salvucci ME. Rubisco: structure, regulatory interactions, and possibilities for a better enzyme. Annu. Rev. Plant Biol. 2002;53:449. doi: 10.1146/annurev.arplant.53.100301.135233. [DOI] [PubMed] [Google Scholar]
- 72.R Core Team. A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, 2018).
- 73.R Studio Team. RStudio: Integrated Development for R (RStudio, Inc., 2018).
- 74.Wickham, H. & Bryan, J. readxl: Read Excel Files. R package version 1.3.1 (2019).
- 75.Hamner, B. & Frasco, M. Metrics: evaluation metrics for machine learning. R package version 0.1.4 (2018).
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The author and DOI information relating to the published articles from which all Rubisco kinetic, Rubisco activation state, net CO2 assimilation, and electron transport rate temperature response curve data where analysed are presented in Supplementary Tables 1, 2, 3, and 4. The collated data are provided at 10.6084/m9.figshare.22661989 and with the model code (10.5281/zenodo.7683559). The global individual An vs leaf temperature34 and Panama species dataset17 are available from the TRY plant trait database http://www.try-db.org/TryWeb/Data.php under the dataset names “Global Leaf Gas Exchange Database (I)” and “Photosynthesis Temperature Response Panama”.
A model script and An data are accessible through GitHub (10.5281/zenodo.7683559). The script provides model fits to An temperature response curves using relevant model parameters based on whether the plant was grown below or above 25 °C. The script will generate graphs and provide indices of how well observed and model data match for each curve fit, comparable to the analysis presented in this study.