

Abstract
Rabies is an ancient disease. Two centuries since Pasteur, fundamental progress occurred in virology, vaccinology, and diagnostics—and an understanding of pathobiology and epizootiology of rabies in testament to One Health—before common terminological coinage. Prevention, control, selective elimination, and even the unthinkable—occasional treatment—of this zoonosis dawned by the twenty-first century. However, in contrast to smallpox and rinderpest, eradication is a wishful misnomer applied to rabies, particularly post-COVID-19 pandemic. Reasons are minion. Polyhostality encompasses bats and mesocarnivores, but other mammals represent a diverse spectrum of potential hosts. While rabies virus is the classical member of the genus, other species of lyssaviruses also cause the disease. Some reservoirs remain cryptic. Although global, this viral encephalitis is untreatable and often ignored. As with other neglected diseases, laboratory-based surveillance falls short of the notifiable ideal, especially in lower- and middle-income countries. Calculation of actual burden defaults to a flux within broad health economic models. Competing priorities, lack of defined, long-term international donors, and shrinking local champions challenge human prophylaxis and mass dog vaccination toward targets of 2030 for even canine rabies impacts. For prevention, all licensed vaccines are delivered to the individual, whether parenteral or oral–essentially ‘one and done’. Exploiting mammalian social behaviors, future ‘spreadable vaccines’ might increase the proportion of immunized hosts per unit effort. However, the release of replication-competent, genetically modified organisms selectively engineered to spread intentionally throughout a population raises significant biological, ethical, and regulatory issues in need of broader, transdisciplinary discourse. How this rather curious idea will evolve toward actual unconventional prevention, control, or elimination in the near term remains debatable. In the interim, more precise terminology and realistic expectations serve as the norm for diverse, collective constituents to maintain progress in the field.
Keywords: Diagnosis, Epidemiology, Lyssavirus, Neglected Tropical Diseases, Pathogenesis, Prophylaxis, Rabies, Surveillance, Vaccination, Zoonosis
Introduction
Rabies is a quintessential disease of nature, predating the domestication of Canis spp., but is historically associated with dogs in the minds of humankind (Rupprecht et al. 2020). Technical advances throughout the twentieth century provided pivotal insights into the etiology, diagnosis, pathobiology, prevention, control and potential treatment of this acute, progressive encephalitis (Tarantola 2017). Such progress tempted multiple authors to apply the term ‘eradication’ in relation to rabies (Brochier et al. 1991; Flamand et al. 1992; Terré et al. 1996; Grimm 2002; Larghi 2004; Kasempimolporn et al. 2008; Pastoret et al. 2014; Thomson and Penrith 2017; Sartore et al. 2018; Reece 2019; Mubashir and Hussain 2021; Lojkić et al. 2021; Aréchiga Ceballos et al. 2022; Shafaati et al. 2023). Whether due to linguistic license or singular optimism, to muse upon the fantastic and apply such terminology to this zoonosis globally seems in conflict with prior infectious disease concepts, related in no small part to limitations in ‘…feasibility, infrastructure, funding and political will…’ (Dowdle and Cochi 2011). In retrospect, the COVID-19 pandemic chastised our collective epidemiological tomfoolery and blunted global confidence over basic conventional prevention and control (Warrell 2021; Gongal et al. 2022; Nadal et al. 2022a, b; Tidman et al. 2022; Goel et al. 2023).
Not unexpectedly, neglected tropical diseases (NTDs) were impacted disproportionately by the COVID-19 pandemic. Rabies was no exception. For example, in the USA, an uptick of five human deaths occurred (including unprecedented human prophylaxis failure), and dog importation was restricted (Pieracci et al. 2021; Ma et al. 2023). In South America, Peru grappled once again with canine rabies (Raynor et al. 2021). Despite preliminary progress, basic animal control requires a major reset in Haiti (Kunkel et al. 2021). In Sri Lanka, where a prepandemic prevention and control program made significant headway, elimination no longer appears imminent (Kanda et al. 2021). Indirectly, the enactment of COVID-19-associated global lockdowns might have temporarily reduced the incidence of dog bites, but supply chain interruptions also affected the availability of biologics and quality of medical care, including the threat of counterfeit vaccines (Henson et al. 2020; Saleem et al. 2021; Siddiqui et al. 2021). The broader controversy on human vaccine hesitancy extended to pet owners with reluctance to receive rabies vaccines (Kogan and Rishniw 2022). Rabies reports rose throughout sub-Saharan Africa (Goel et al. 2023). On the subcontinent, the burden of rabies in India was unsurpassed by any other country, even before the pandemic. Within the Indian state of Kerala, nearly 2 million dog-bite incidents were recorded during 2022, including rabies occurrence despite vaccination (Thiagarajan 2022). The death of at least six persons who began rabies prophylaxis triggered public anxiety and a major investigation into the relative quality of available biologics (Bajeli-Datt 2022).
Major repercussions from the global pandemic continue to ripple throughout the rabies profession, raising significant issues. Irrespective of these relatively recent events, why is rabies not a candidate for eradication? What is the true status of rabies prevention in the near term? Emerging technologies should bolster conventional disease control efforts, but will new biologics or concepts under development recoup contemporary setbacks and jumpstart elimination? The objectives of this communication are to present a brief update on current global efforts on rabies and its management based upon recent contributions to the literature and to provide a prospective rationale as to the realistic limitations of these actions within the larger historical context of the field.
Viral systematics and ramifications of an evolving taxonomy
Reflections on disease prevention, control, elimination, and eradication benefit from introspection on how various pathogens arose, emerged and perpetuated. However, despite the historical record of rabies and its widespread occurrence in humans, domestic animals and wild mammals, lyssavirus origins and evolution remain puzzling. Genetic distances within the Lyssavirus genus (Fig. 1) are much shorter than those in other rhabdovirus genera (Walker et al. 2018). Some investigators have suggested that such genetic homogeneity may reflect a relatively younger age of lyssaviruses (Bourhy et al. 2005).
Fig. 1.

Phylogenetic placement of lyssaviruses within the subfamily Alpharhabdovirinae, family Rhabdoviridae. The evolutionary history was inferred from an alignment of the complete L protein sequences of animal rhabdoviruses, using the Maximum Likelihood method. Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Joining and BioNJ algorithms to a matrix of pairwise distances estimated using a JTT model. Statistical significance was assessed via 100 bootstrap replicates (values > 70 are shown for key nodes)
In addition, for decades, the Lyssavirus genus was an “orphan” within the larger rhabdovirus phylogeny, without any major clues on progenitors. Only in 2019 was a lyssavirus-related rhabdovirus identified in a tree frog, Dryophytes cinereus, and another lyssavirus-related virus was detected in a lizard, Anolis allogus (Horie et al. 2021). These viruses were not monophyletic to each other (Fig. 1). Therefore, they were classified into two separate genera, with one species each, Replylivirus and Amplylivirus, by the International Committee on Taxonomy of Viruses (ICTV). Indeed, the question remains, did lyssaviruses originate from amphibian or reptilian rhabdoviruses (which sounds rather unlikely from an ecological perspective, given that the majority of lyssaviruses circulate in bats)?
Approximately 65 million years ago, at the end-Cretaceous mass extinction, most mammals were small, nocturnal, and insectivorous before their substantial ecological release and ensuing adaptive radiation of body sizes and niche occupations from presumed lessened interspecific competition (Grossnickle and Newham 2016). Perhaps an insect-borne lyssa-like virus (yet to be discovered) was a progenitor for related viruses infecting terrestrial insectivorous animals, such as frogs, lizards, and later, mammals, including bats (Shope 1982). In any scenario, the remarkable genetic similarity between the replylivirus, amplylivirus, and lyssaviruses points to an ancient origin of all these viruses. Biogeographically, both the amplylivirus and replylivirus were detected in the New World (North America and Cuba, respectively), whereas the greatest lyssavirus diversity is seen in the Old World. Africa (Nel and Rupprecht 2007) or Eurasia (Hayman et al. 2016) were suggested as the most likely areas of lyssavirus origination. However, we still do not have an instrument for retrospective estimation of age and evolutionary traits of viral genomes subjected to strong purifying selection, especially when only a limited set of recent samples is available (Wertheim and Kosakovsky Pond 2011; Wertheim et al. 2013).
When did rabies arise? Most molecular clock estimates for lyssaviruses were performed on sequences collected during recent decades. Evolutionary rates in a set of sequences are time dependent, increasing toward the present, because the transient mutations have yet to be removed by purifying selection. When the rate of decay dynamics is considered, ‘viral age’ increases by several orders of magnitude (Ho et al. 2011; Membrebe et al. 2019). We consider the discovery of lyssa-like rhabdoviruses in amphibians and reptiles as a confirmation of opinions expressed earlier that lyssaviruses are not recent over thousands of years but rather evolved over tens of millions of years (Shope 1982; Rupprecht et al. 2017). Lyssaviruses may have descended and disseminated from ancient Gondwana via now extinct mammals and later proto-bats in the Eocene. Given the susceptibility of birds and the presence of amphibian and reptile lyssa-like viruses confined thus far to the New World, such ancestral viruses may have already been present broadly by the time of ongoing continental drift during the Late Cretaceous, while mammals were in transition.
Although relative ecological compartmentalization was realized, rabies was thought to be caused by a single entity until the detection of ‘rabies-related’ viruses during the 1950s (Shope 1982). At present, ICTV recognizes 17 lyssavirus species (Fig. 2). Most of these viral species are associated with bats. The only “exception” from that rule was suggested to be rabies virus (from a taxonomic perspective, the virus lineages included in the species Lyssavirus rabies), which circulates among bats, mesocarnivores, potentially in New World nonhuman primates, such as marmosets, and perhaps even temporarily in certain ungulates under ideal ecological conditions, such as kudu antelopes (Marston et al. 2018; Ness et al. 2023). Such an assessment may be correct only as long as we consider that the assignment of lyssaviruses to the host species is correct.
Fig. 2.
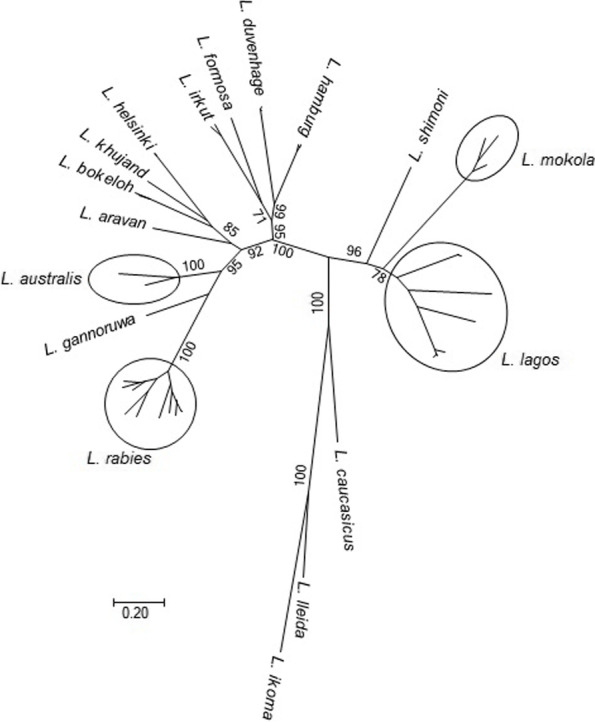
Phylogenetic structure of the Lyssavirus genus based on the complete N gene sequences of representatives from each currently recognized species. The topology was inferred using the Maximum Likelihood method. Initial trees for the heuristic search were obtained automatically by applying Neighbor-Joining and BioNJ algorithms to a matrix of pairwise distances estimated using a JTT model. Statistical significance was assessed via 100 bootstrap replicates (values > 70 are shown for key nodes)
In general, taxonomy is only a tool that humans use to break the continuity of living forms in an attempt to classify them by specific characteristics. This demarcation is neither absolute nor ideal and may vary depending on the set of criteria used. The present demarcation of lyssaviruses includes genetic sequence identity, phylogenetic tree topology, serologic cross-reactivity, and ecological traits, such as host range, geographic distribution, and pathobiology (Rhabdoviridae. Virus Taxonomy: 2021 Release. EC 53, Online, July 2021). Not all of these criteria are met for every lyssavirus species identified to date (Fig. 2). For example, “Phylogroup I” lyssaviruses cross-neutralize each other, as do “Phylogroup II” lyssaviruses (Badrane et al. 2001; Hanlon et al. 2005; Horton et al. 2010). Genetic distances between representatives of Lyssavirus gannoruwa and L. australis, as well as between related Eurasian bat lyssaviruses such as L. helsinki, L. bokeloh, and L. khujand, are very close to the distances within L. rabies (80–81% nucleotide identity for the nucleoprotein gene). In contrast, genetic distances between lineages of L. lagos (approximately 79% nucleotide identity for the nucleoprotein gene) are longer than the intraspecies demarcation limits set by ICTV (Markotter et al. 2008).
Taxonomic changes may be cogent in both extremes. For example, all “Phylogroup I” lyssaviruses may be considered members of one species (e.g., L. rabies), but the same would be hard to reconcile with “Phylogroup II” lyssaviruses, given that L. lagos and L. mokola are certainly different viruses by various characteristics, and L. shimoni, in some phylogenetic constructions, is monophyletic to L. lagos but in others is ancestral to both L. lagos and L. mokola (Kuzmin et al. 2010).
As an alternative extreme, one may consider that some of the L. rabies lineages that demonstrate strict association with a specific host and geographic area are separate lyssavirus species, at least from ecological perspectives. Indeed, even for L. rabies lineages, most cross-species transmission events from the reservoir usually lead to dead-end infections or only short-term outbreaks (Streicker et al. 2010; Kuzmin et al. 2012). The present taxonomy balances such extremes, trying to accommodate the multidimensional demarcation of lyssavirus species. However, what one is facing now is that the ICTV is attempting to classify multiple viral genomes identified during metagenomic studies, rather than entire viruses. In such settings, neither serologic, pathobiologic, nor ecologic properties of viruses are taken into account. With the entire taxonomy moving toward “genotyping”, genetic criteria are going to become overarching. As such, the present structure of the Lyssavirus genus will probably be reassessed in the near future.
Another empirical caveat associated with modern viral taxonomy is the frequent changing of species names. This alteration occurred several times over recent years, along with attempts to establish a binomial system. This binomial system, although resembling that of “larger” biology, is not exactly Linnaean but operates with “free-form” species epithets (Zerbini et al. 2022).
Species epithets are not uniform. They are used either in English, Latin (sometimes in improper form), or via historical terms. Considering the long iterative process of virus species renaming, we may expect that it will continue. Importantly, however, the concept of ‘viruses’ (biologic entities) is not the same as ‘virus species’ (taxonomic entities). Viral names are not a subject of ICTV policies and remain unchanged (and can be abbreviated, unlike species names). For example, rabies virus (RABV) is still rabies virus, whereas the species name was changed to Rabies lyssavirus and then to Lyssavirus rabies. Similarly, European bat lyssavirus, type 1 (EBLV-1) is still the same biologically, even though the species name was changed to European bat 1 lyssavirus and then to Lyssavirus hamburg (https://pubmed.ncbi.nlm.nih.gov/36437428/).
Regardless of the varying complexities of taxonomic ruminations, new lyssaviruses await detection and characterization and are particularly relevant regarding global disease emergence and public health implications (Fooks et al. 2021). To date, there are only a handful of primary human deaths attributed to ‘rabies-related’ lyssaviruses in Africa, Australia and Eurasia (van Thiel et al. 2009; Markotter and Coertse 2018; Merritt et al. 2018; Poleshchuk et al. 2023). No documented human postexposure prophylaxis (PEP) failures are yet known due to disparate lyssaviruses, despite the lack of veterinary vaccine breadth and occasional breakthrough infections from case reports in domestic animals (Coertse et al. 2021). Often, such incidents are viewed as biological curiosities, divorced from the reality that most human rabies cases are never confirmed by laboratory testing. A recent note of the identification of West Caucasian Bat Virus from a cat in Italy, > 2,000 km and 18 years from the first isolation in the Caucasus mountains, should provide some pause for concern over surveillance limitations and biosecurity, as well as the objectivity of a truly ‘rabies-free’ Europe (Leopardi et al. 2021; Vega et al. 2021). Irrespective of the existence of bat lyssaviruses, rabies in mesocarnivores perpetuates in the eastern portion of the continent and may become more exacerbated given ongoing strife.
At a minimum, taxonomic recognition of antigenically distinct lyssaviruses beyond RABV, without enhanced laboratory-based surveillance and safe and cross-reactive biologics, challenges any notion of simplistic rabies eradication. Conversely, viral taxonomy may be used by some for fleeting alterations in operational definitions (e.g., that rabies is caused by RABV alone, that wildlife rabies is comparatively unimportant, or that non-RABV causes only a ‘rabies-like’ disease) and could provide more palatable, short-term public health solutions to seemingly bend nature to the ephemeral biopolitical will.
Polyhostality: resilient wildlife and dogs
Considering the taxonomic breadth of extant warm-blooded vertebrates, more than 16,000 potential species are considered susceptible to lyssavirus infection (Paarmann 1955; Baby et al. 2015; Gilbert 2018; Rohde and Rupprecht 2020). This wide span reflects in part both viral plasticity and various host conserved factors over evolutionary time, including structural and biochemical features of receptors, replication dynamics, excretion mechanisms, and host innate/adaptive immunity (Conselheiro et al. 2022; Gérard et al. 2022; Lian et al. 2022). The capability of infection for all hosts is dissimilar from perpetuation, as birds have been toothless for millions of years, removing one major anatomical assist for viral entry, notwithstanding the potential capabilities of beaks to create lesions from modern avianized dinosaurs (Rohde and Rupprecht 2020). Any resulting productive infection is impacted at the outset by a time-space continuum overlap, requiring simultaneous engagement of host and pathogen (Jacquot et al. 2022). For example, on certain islands, such as Hawaii, susceptible hosts abound, but splendid isolation in this case of a distant, insular environment is devoid of a pathogen source, unless introduced (Sasaki et al. 1992). Greater proximity to a source also complicates the probability of relative sustainability (Morgan et al. 2020). In addition, such appreciation also implies a relevant, decentralized laboratory-based surveillance system, with more limited bias for the inclusion of wildlife (Wallace et al. 2014). For example, without considering nondomestic reservoirs, Taiwan would not have detected naturally infected bats and ferret badgers (Hsu et al. 2019; Zhao et al. 2019).
Wildlife rabies began long before the origin of domestic species, with lyssaviruses in bats evolving prior to carnivores (Badrane and Tordo 2001). Among mammalian hosts, the recorded host gamut spans from anteaters to zebras (Obonyo et al. 2014; Grome et al. 2022). Although no marsupials have been reported from Australia yet, even relatively resistant nonplacental taxa, such as New World opossums, are known (Diana et al. 2015). Most ‘spillover’ cases, such as among human victims, are considered dead-end infections unless tissues and organs are used inadvertently for transplantation (Mrzljak et al. 2020). Otherwise, when applied to rabies, intraspecific perpetuation is predictable from mammalian sociobiology, particularly among canids. Arctic foxes are key drivers of infection in circumpolar environments (Ørpetveit et al. 2022). Other fox species serve this role in temperate and tropical environments (Caraballo et al. 2021; Ma et al. 2023; Matulis et al. 2022). Coyotes and jackals are ecological and epizootiological equivalents within both the New and Old Worlds (Shwiff et al. 2008; Ngoepe et al. 2022). Raccoon dogs acquired dog and fox RABV throughout Eurasia (Yang et al. 2017; Shulpin et al. 2018). Direct cross-species transmission (CST) is also inevitable among noncanid carnivores, such as ferret badgers and mongooses. due to a suite of ecological imperatives, such as predator–prey relationships and interspecific antagonistic interactions (Fig. 3). The applicable impacts of this polyhostality suggest that oral vaccination of wildlife may be necessary within the dual context of long-term canine rabies management, as exemplified at the Mexico-USA border and elsewhere (Sidwa et al. 2005; Malan et al. 2022; Ma et al. 2023).
Fig. 3.

Phylogenetic lineages of rabies viruses (RABV) circulating in wild carnivores that are related to canine rabies viruses. The tree was generated from full-length nucleoprotein (N) gene sequences by the neighbor-joining method supported by 1,000 bootstrap replicates. GenBank accession numbers are shown at tree tips, host species and geographic region are indicated after brackets, and conventional names for RABV lineages are shown on the right
Similarly, bat to nonbat CST is evident among carnivores and other species, independent of canine lineages (Fig. 4). Compared to domestic dog-wild carnivore CST events, the frequency and mechanisms of host shifts for longer-term perpetuation from bats to other mammals is poorly understood (Kuzmin et al. 2012). Such realities not only demonstrate that rabies will perpetuate even if conventional canine rabies is controlled but also provide evidence of maintenance among carnivores with the opportunity to return from the wild to the domestic animal cycle. Moreover, the complexities of such overt polyhostality not only complicate current plans for prevention, control, and selective elimination but also nullify the suggestions of eradication of this disease, even if RABV remains the only significant lyssavirus responsible for CST, disease emergence, and perpetuation (Marston et al. 2017, 2018).
Fig. 4.

New World bat rabies virus (RABV) diversity (black) in association with infections in carnivores (red). Asterisks indicate viruses recovered during known outbreaks with sustained cross-species transmission of bat RABV among carnivores. Bat species and genera that serve as lineage hosts are indicated on the right. The neighbor-joining tree is based on RABV nucleoprotein (N) gene sequences with 1000 bootstrap replicates (shown as % for key nodes) and rooted with Mexican skunk, South-Central skunk, and raccoon RABV lineages (also related to American bat viruses within the "Indigenous American" RABV lineage)
Resolute burden estimates
How many people die of rabies? No one knows. Why? In part, explanations are similar for the lack of laboratory-based surveillance related to NTD in general (Ghai and Hemachudha 2022). Others are more pathogen specific (Table 1). As a disease related to poverty, individuals, such as children, may not fundamentally understand how rabies is acquired or may not appreciate its severity (Laorujisawat et al. 2022; Mapatse et al. 2022; Zucca et al. 2021). Even if a bite is reflected as a risk factor for disease acquisition, less serious or nonbite exposures may be ignored or not be recognized, such as related to bats, even in highly developed countries (Holzbauer et al. 2023; Ma et al. 2023). Recall bias could also be an issue if exposure occurred months to years previously (Amoako et al. 2021). Considering the lack of therapy, rather than being refused care or even locked away and ignored in a medical ward as a pariah, families may prefer patients to die at home with a modicum of empathy and simple palliation (Warrell et al. 2017). Although rabies has been described clinically and characterized by its often unique manifestations, such as hydrophobia, practitioners may not appreciate more subtle or atypical signs, such as in the paralytic form (Hemachudha and Hemachudha 2021). Similarly, the disease may be masked by acute trauma or misdiagnosed as related to other forms of encephalitis (Mallewa et al. 2007; Mudiyanselage et al. 2016; Bhat et al. 2021). Despite a number of highly sensitive and specific diagnostic tests recognized by WOAH and WHO, there may be no suitable laboratory facility in the locality where human deaths occur, such as in rural areas (Rana et al. 2020). Families traumatized by the sudden death of a family member may not consent to invasive procedures postmortem. Certain religious practices, such as requiring relatively rapid internment, may preclude desired sample collection or accurate clinical case reporting.
Table 1.
Factors related to an underreporting of rabies cases
| Number | Factors |
|---|---|
| 1 | Ignorance of causality |
| 2 | Nonrecognition of exposure |
| 3 | Longer incubation periods |
| 4 | Futility of treatment |
| 5 | Atypical clinical signs |
| 6 | Burial practices |
| 7 | Lack of local testing facilities |
| 8 | Fear of contamination |
| 9 | Misdiagnosis |
| 10 | Decreasing autopsies |
| 11 | Nonnotifiability |
| 12 | Self-perpetuating cycle of neglect |
A few inferences of the extent of global human rabies appear, which of course are not inviolated. Epidemiologically, values should vary spatiotemporally. For example, the annual number of human rabies deaths in Africa and Asia was estimated at between 24,000 and 93,000 cases and to cost more than $583 million (Knobel et al. 2005). Another often quoted global model calculated 25,000 to 159,000 human deaths, over 3.7 million disability-adjusted life years, and $8.6 billion economic losses annually (Hampson et al. 2015). More recently, an investigation extrapolating from a 2019 Global Burden of Disease Study suggested that overall fatalities declined over a decade, with a range of 6,019 to 17,939 human deaths (Gan et al. 2023). Clearly, such data are important for appropriate planning, to generate resources, to measure success, etc., but quite difficult to generate (Taylor et al. 2017; Minhaj et al. 2023).
Progress and attempted quick returns in the face of recurrent setbacks in the global elimination of human rabies by dogs
For thousands of years to the present, most human rabies fatalities occurred after dog bites. Hence, the current focus on the prevention, control, and selective elimination of canine rabies is an integrated reality built upon centuries of prior experience, unfortunately often driven by fear and indiscriminate culling of the pariah dog, as represented by the effigy and art of everyday life (Figs. 5 and 6). Large parts of Europe accomplished this task long before the advent of vaccines, largely by population reduction, leashing, and use of muzzles. Other locales, usually insular, apparently never reported canine rabies or had an abbreviated interlude, relying upon geographic isolation and quarantine. For example, in addition to an early introduction event during 1866-67, canine rabies did not become established in Australia, and an active program to prevent its reoccurrence prevailed (Sparkes et al. 2015).
Fig. 5.

Hundefängerin (dog chasers), Ernst Barlach, woodcut, ca. ~ 1919. From: Die Kunst. Monatshefte für freie und angewandte Kunst, vol. 4 https://commons.wikimedia.org/wiki/File:Hundef%C3%A4nger,_Ernst_Barlach.jpg3, 1921, p143
Fig. 6.

‘Aguador mata-perros ‘ (waterseller dog-killer), watercolor painting by Francisco ‘Pancho’ Fierro Palas (Lima, 1850). https://commons.wikimedia.org/wiki/File:Aguador_mata-perros_(1850).jpg
Building upon Pasteur’s experimental work, mass dog vaccination (MDV) began during the 1920s in Japan (Umeno and Doi 1921). Rabies was eliminated there by 1957 through coordinated canine vaccination and dog management (Kamata et al. 2023). Elsewhere, twentieth century progress occurred throughout the Americas after the original introduction of canine rabies via European colonization. In addition to culling, leash laws, etc., Canada and the USA implemented dog vaccination following World War II (Rosatte 1988; Ma et al. 2023). Both countries successfully controlled canine-mediated rabies with a regional public health strategy. Licensing of owned dogs supported animal control activities, and the outright killing of free-ranging dogs moved gradually toward a policy for their capture and holding at ‘pounds’ for adoption or euthanasia (Fig. 6). Comparatively, by the end of the twentieth century, few human cases appeared after elimination of canine rabies, acquired through imported human cases (exposed to dogs abroad) or after indigenous wildlife exposures (Daniels et al. 2020; Kunkel et al. 2022; Ma et al. 2023).
After 1993, Mexico's national program implemented massive MDV, eliminating canine-mediated human rabies deaths (González-Roldán et al. 2021). Estimates suggest that the program prevented approximately 13,000 human rabies deaths by investing more than $300 million in surveillance, prevention, and control. Mexico received validation from the WHO as a country free of canine-mediated human rabies deaths, in line with the regional program for elimination (Del Rio Vilas et al. 2017). However, because of enzootic wildlife rabies, even after curtailment of canine rabies, the challenges of human prevention remain, especially within remote communities. For example, recently, a seven-year-old boy died after being bitten by a bat in an impoverished rural community of Oaxaca (News Desk 2022). The child only reported to the hospital 20 days after exposure, already exhibiting symptoms consistent with rabies. This exposure occurred 100 km from the main city, with no rabies vaccines. Another ill child was also suspected to be infected and died. Authorities coordinated a public health response comprised of renewed educational outreach and local animal vaccination. While canine rabies elimination alleviates a significant burden, such success does not erase the public health and veterinary responsibilities associated with the disease and its ongoing prevention due to infected wildlife.
In addition to Mexico, multiple countries within Latin America (e.g., Argentina, Chile, etc.) achieved significant progress toward eliminating canine-mediated human rabies deaths through MDV programs, which began via coordinated efforts with PAHO during 1983, first targeting major urban centers in excess of 100,000 human inhabitants (Freire de Carvalho et al. 2018). Many countries within the Americas (except Bolivia, Haiti, etc.) are considered canine rabies-free, including the Caribbean region. Whether most countries will participate in a similar process of validation as accomplished by Mexico remains speculative. Thus, over the past 200 years, multiple localities have attained freedom from canine rabies and prevented its re-establishment, including western Europe, most of the Americas and the Caribbean, Japan, Australia and Pacific Oceania, even though other lyssaviruses or RABV variants of wild mesocarnivores and bats perpetuate (Rupprecht et al. 2018). Today, the major burden lies within the affected tropical portions of Asia and Africa (Rupprecht et al. 2022).
Recognizing progress in the Americas and the major needs within the Old World, in 2010, with support from the Bill and Melinda Gates Foundation (BMGF), proof-of-concept canine rabies elimination projects were initiated at sites in the Philippines, South Africa and Tanzania (Elser et al. 2018). These projects demonstrated the feasibility of local prevention and control in support of broader disease elimination plans (Shwiff et al. 2016; Hatch et al. 2017; Miranda et al. 2017; Mpolya et al. 2017; LeRoux et al. 2018). In addition, the building blocks erected from these projects via BMGF support continued to bear weight. For example, in Tanzania, on 14 February 2022, a 12 year-old village boy was bitten by an unfamiliar dog in the Ngorongoro Conservation Area. Weeks later, he died within two hours of presentation at a hospital in Karatu after displaying signs on 6 March, consistent with rabies (i.e., known locally as kichaa cha umbwa, Swahili for “madness of dogs”). Although this child was not saved, quick-acting hospital staff, after learning of other exposed children in the same village, notified public health authorities, such that 10 other at-risk persons were able to be vaccinated, exemplifying rapid risk communications and integrated preventative actions by a diverse biomedical team (Resolve To Save Lives (RTSL), 2022). Such an effective local response epitomizes a larger national plan for rabies management (Tanzania 2019). Hence, while the country is still enzootic for canine rabies, long-standing public – private partnerships and initiated local efforts continue to make major inroads toward prevention and control (Lushasi et al. 2020).
In the KwaZulu-Natal (KZN) province of South Africa, the immediate benefits of the BMGF project were appreciable in limiting human deaths. However, progress appeared rather short-lived and not readily extrapolated to the rest of the country. Overall, national human rabies cases reported from 2008 to 2018 remained largely unchanged (Weyer et al. 2020). Within KZN province, a cross-sectional observational investigation showed that less than 70% of dogs were vaccinated. Potential factors for less-than-ideal canine vaccination coverage included animal age, population turnover, the number of free roaming dogs, and husbandry practices in certain high-risk areas (Hergert et al. 2016). A qualitative study in the province during 2017-18 found a lack of broad public knowledge on rabies, economic constraints for vaccination, and the unavailability of PEP at local health facilities (Hadebe and Sibiya 2020). This was especially alarming, given the zero human deaths recorded in KZN during 2014 from the BMGF supported project (LeRoux et al. 2018). In addition, although dogs are the primary RABV reservoir in South Africa, jackals are also associated with the disease (Mogano et al. 2022; Ngoepe et al. 2022). With the maintenance of canine rabies and in the absence of broader MDV, the impacts for proper health care delivery and PEP after animal exposures remain substantial (Whitbread et al. 2022). Recently, in the popular tourist area of Nelson Mandela Bay Municipality, an unprecedented rabies outbreak began. Between 1st January 2021 and 13th February 2022, 436 rabid dogs and five human fatalities were reported during a concomitant outbreak of foot and mouth disease (Hill 2022; Ravensberg et al. 2022).
In the third BMGF-supported project, in the Philippines, from 2010 to 2017, the number of rabies-free provinces and municipalities increased from three to 49 (Leonardo et al. 2020). The overall progress was associated with greater veterinary services involvement, broad public support, communal advocacy, increased health communications, focused community education, and One Health linkages (Medina et al. 2016; Valenzuela et al. 2017; Barroga et al. 2018; Deray et al. 2018; Amparo et al. 2019). However, in the Metro Manila area, between 2006 and 2015, there was no decline in deaths or in the catchment area from which human rabies cases occurred (Guzman et al. 2022). The COVID-19 pandemic also played a prominent role in setbacks. During 2022, nationally, more than 320 human fatalities were reported, compared to less than 250 in 2021. In response, some legislative members called for an investigation of the failure of the national rabies prevention and control program to reach intended targets to eliminate human rabies by 2020 and the unmet declaration of the country to be rabies-free by 2022 (Cruz 2022). In lieu of canine rabies elimination throughout the archipelago by MDV, in addition to appropriate PEP, other strategies to reduce human deaths may include focused PrEP considerations for at-risk groups, such as pediatric populations (Quiambao et al. 2020, 2022).
The documented progress throughout the Americas and the focused attention via the BMGF projects helped spur additional initiatives. During 2015, a Pan-African Rabies Control Network (PARACON) meeting was held in South Africa to provide tools and coordination toward the elimination of canine-mediated human rabies deaths throughout the continent (Scott et al. 2015). Concomitantly with the PARACON outcome and based upon evidence acquired largely outside the region, the leadership of the WHO, WOAH, FAO, and other partners targeted an ambitious goal of global elimination of human rabies mediated via dogs (GEHRD) by 2030, focusing on LMICs in Africa and Asia (Anonymous 2015). Human prophylaxis, MDV, public education, and community engagement were considered crucial elements to attaining this 'Zero by 30' (ZBT) goal (WHO Rabies Modeling Consortium., 2020).
By the 21 century, epidemiological evidence showed that canine-mediated rabies was eliminated from all highly developed countries and increasingly controlled in middle-income countries through large-scale MDV campaigns and modern prophylaxis. To be clear, the GEHRD could be accomplished without actual canine rabies elimination by minimizing animal bites, preventing human rabies after exposure by PEP, and enhancing responsible dog ownership, but the risk of transmission of RABV from free-ranging dogs would remain (Mindekem et al. 2017). Such a first step is admirable and necessary but not economically sustainable. The long-term ultimate goal is complete elimination of canine rabies. Otherwise, the risk of human rabies acquisition from infected dogs remains, as seen in multiple situations.
Despite the wide availability of basic tools for such prevention and control, most LMICs, particularly throughout sub-Saharan Africa, still grapple with the first steps of the GEHRD (Zinsstag et al. 2017; Mbilo et al. 2021). Sub-Saharan Africa is second only to Asia in terms of the estimated global burden (Knobel et al. 2005; Hampson et al. 2015). After the announcement of the ZBT goal during 2015, few countries made substantive progress (Kanda et al. 2022). The reasons are multiple and share similarities with other NTDs. For example, in Chad, major barriers included economic barriers (e.g., vaccine costs, expense in transporting dogs, etc.), sociocultural (e.g., perceptions of the vaccine tainting dog meat for consumption, religious prohibitions on handling dogs, etc.) and institutional (unavailability of biologics, absence of training, etc.) aspects (Mbaipago et al. 2022). Such transdisciplinary concerns are relevant throughout many LMICs, as discussed in detail (Rupprecht and Salahuddin 2019; Tantawichien and Rupprecht 2020; Swedberg et al. 2022).
Since the 2015 GEHRD goal and ensuing COVID-19 pandemic, within tropical Old World hot spots, no country has eliminated canine-mediated human rabies deaths (Rupprecht et al. 2022). National surveillance databases and routine reporting mechanisms through PAHO/WHO and WOAH channels, together with regional disease conferences, confirm this reality. As one other aid to gauge progress, the Stepwise Approach Toward Rabies Elimination (SARE) uses monitoring tools for data-driven evaluation of strengths and weaknesses of rabies prevention and control programmes (Coetzer et al. 2016). With SARE scores between zero (for canine rabies endemic countries with no control) and five (signifying freedoms from canine rabies), current evidence indicates that canine-mediated rabies remains uncontrolled in much of Asia and Africa, with scores unavailable or between zero and one (Global Alliance and for Rabies Control (GARC) 2022). Optimistically, during 2018–2019, a few countries (e.g., Bhutan, the Philippines, Thailand, Vietnam, etc.) achieved SARE scores of ~ 3.5, approaching the ZBT goal (Global Alliance and for Rabies Control (GARC) 2022). Such progress resulted from long-standing free access to human PEP and dog vaccination (Tenzin 2012; Valenzuela et al. 2017; Anothaisintawee et al. 2019; Nguyen et al. 2019). Unfortunately, postpandemic COVID-19 disruptions in One Health implementation, with the detection of new rabies cases among dogs and humans, created setbacks throughout the region (Acharya et al. 2021; Lhendup and Dorji 2021; Ghai and Hemachudha 2022; Pham-Thanh et al. 2022).
While there have been significant attempts to manage canine-mediated human rabies in some parts of southern Asia through investment in millions of dollars by local authorities and international organizations, most countries still report human rabies deaths. For example, in Afghanistan, between 2017 and 2019, 36,959 animal rabies cases were recorded, with an estimated human rabies incidence of 5.7 per 100,000 population (Abdul-Jalil 2020). A national rabies prevention and control plan was developed but remains unimplemented. There are large numbers of free roaming dogs in the country and erratic dog vaccinations (Acharya et al. 2021). To date, Afghanistan has not undertaken a SARE assessment (Global Alliance and for Rabies Control (GARC) 2022).
China, second only to India with regard to the burden of human rabies, has decreased cases in most urban centers over the past decade. (Yue et al. 2022). Of 39 notifiable diseases, rabies ranked 3rd in 2015. The country had a SARE score of 1.5 during 2019 (Global Alliance and for Rabies Control (GARC) 2022). Chongqing, China, is the world's largest municipality, with a population of ~ 32 million and an estimated ~ 1.3 million dogs, of which 200,000 are considered free roaming (Mission Rabies (MR) 2023). Between 2007 and 2016, 809 human rabies deaths were recorded (Qi et al. 2018). From information on 548 human cases, only 88% sought medical care, and none received PEP. Laboratory confirmation was only possible for a few of the offending dogs. During 2008, local authorities enhanced surveillance for dog rabies, improved access to PEP, and made canine vaccination compulsory. These measures reduced human deaths to an incidence of ~ 0.1/100,000 by 2013 and improved overall data quality (Qi et al. 2018; Mission Rabies (MR) 2023). Elsewhere in a bordering province, 59 human rabies deaths were recorded in Hunan during 2022 due to failures in access to proper wound care, RIG, and vaccine after exposure (Yang et al. 2022). Between 2010 and 2020, Shandong Province recorded 414 canine-mediated human rabies cases, with the majority in farmers (Zhang et al. 2022). Hence, while China has shown progress in reducing human fatalities in some large population centers, such as Beijing, PEP is not free for exposed persons, and health disparities exist in many rural areas.
At one time, Sri Lanka seemed poised to meet the ZBT. The country demonstrated support for multiple One Health applications, including enhanced surveillance in humans, domestic animals and wildlife; the feasibility of MDV and oral vaccination of free-ranging dogs; educational outreach; and support for human PEP (Perera et al. 2000; Perera et al. 2007; Matibag et al. 2009; Matsumoto et al. 2011; Matsumoto et al. 2013; Häsler et al. 2014; Karunanayake et al. 2014; Kanda et al. 2015; Harischandra et al. 2016; Kularatne et al. 2016; De Silva et al. 2017; Gamalathge et al. 2019). Moreover, thanks to the NGO Mission Rabies (MR), in collaboration with the local authorities, Sri Lanka vaccinated millions of dogs and sterilized at least 40,000 individual dogs (Sánchez-Soriano et al. 2019; Mission Rabies (MR) 2023). These efforts have resulted in a decline in human deaths from hundreds in the 1970s to 31 in 2021. However, the impacts of the COVID-19 pandemic, an unprecedented financial crisis in 2021, and major sociopolitical strife have impacted the public health budgetary allocation for rabies, leading to a shortage in PEP for the foreseeable future (Kanda et al. 2021). At least 28 people succumbed to rabies last year.
In Vietnam, based on surveys from 2020 to 2021, rabies was the most serious of five priority zoonoses (Pham-Thanh et al. 2022). Between January and August 2022, at least 40 human rabies deaths mediated via dogs were recorded. This was an increase compared to the previous years (NewsDesk 2022). The department of preventive medicine recently recommended vaccinations for dogs and cats, confinement of dogs, and basic preventative measures, such as washing bite wounds with soap and water and seeking care in the event of exposure. Regardless, 20 provinces witnessed increased human rabies deaths in recent years.
In Bangladesh, rabies is endemic and accounts for 2,000–2,500 human deaths annually (Mondal and Yamage 2014; South Asian Association for Regional Cooperation (SAARC), 2019). The country has recently prioritized rabies, with a SARE score of 2.5 in 2019 (Global Alliance and for Rabies Control (GARC) 2022). A coordinated multisectoral One Health effort, with the support of international partners and a substantial investment of $33 million in 2017–2022 across 65 district municipalities, resulted in a decline in annual rabies incidence (Li et al. 2019). The country also trained dog catchers and vaccinators in 2019 to support MDV (SAARC 2019). However, the presence of many free-roaming dogs, poor access to PEP, and limited surveillance capacity hamper efforts to attain the ZBT target.
In contrast to the few examples in Asia, no African countries achieved a SARE score above 2.5 (Global Alliance and for Rabies Control (GARC) 2022). One factor to claim true freedom from rabies is an effective laboratory-based surveillance system to detect both human and animal cases. However, the weak capacity for routine surveillance in most countries throughout Africa provides insufficient data to inform investment in rabies prevention and control (Mbilo et al. 2021; Rupprecht et al. 2022). Moreover, the COVID-19 pandemic refocused available funding, personnel, and diagnostics (Nadal et al. 2022a, b). Other major limitations include: a shortage of trained local champions; lack of an evidence-based national strategic plan to prioritize scarce resources; limited One Health prioritization; and poor healthcare systems, that do not allow for prompt interventions in the event of RABV exposure (Mshelbwala et al. 2021). Moreover, such health care limitations lead many dog-bite victims to resort to traditional healers involving the use of herbs, potions and other concoctions (Mshelbwala and Weese 2017; Audu et al. 2019; Ghosh et al. 2020; Rana et al. 2020).
While some national governments in Africa have been supported in developing a national strategic plan to gain political support locally, implementing such a plan has been a major challenge. Unlike in Latin America, many efforts are driven by NGOs and investigators from Western countries working in prior European colonies, rather than any internal bootstrap approach, without the prospect of any long-term funding from external investors. For example, in collaboration with FAO, the Global Alliance for Rabies Control (GARC) delivered capacity-building workshops and assessments and workshops across Africa. In addition, GARC complimented the long-term MDV effort by vaccinating over 37,000 dogs in the northern region of Malawi in 2022 and supported Zanzibar, an island that has made significant progress in enhancing surveillance toward ZBT, with a SARE score of 2.5 during 2018 (Global Alliance and for Rabies Control (GARC) 2022).
During 2022, the NGO Mission Rabies (MR) reached a global milestone of vaccinating over two million dogs against rabies since its inception in 2013 (Mission Rabies 2023). In addition, MR supported community education and enhanced surveillance in multiple hot spot locations, including Cambodia, Ghana, India, Malawi, Mozambique, Sri Lanka, Thailand, Uganda and Zambia, delivering over 200,000 vaccines annually to dogs. The success of MR interventions has been due in part to the recruitment of international and local volunteers. Unlike the experience of MR, which helps deliver MDV using tools that allow monitoring of teams, personnel, supplies and impact, most interventions rolled out by local authorities in Africa remain short term and uncoordinated, without proper data documentation to measure impact.
In response to such regional needs, the FAO has been training veterinarians and para-veterinary professionals under an In-Service Applied Veterinary Epidemiology Training (ISAVET) program (Fasina et al. 2021). This program aims to strengthen local and para veterinarians' capacity through earlier detection and response to outbreaks in a timely manner through access to modern epidemiological tools. This can alleviate the current drawbacks in rabies management for Africa. There is an overdue need to actively involve private veterinarians and academics in planning and implementing MDV and recruiting students as volunteers, similar to the model of MR. University laboratories could also be better utilized for improved rabies surveillance. Thus, in principle, the GEHRD is feasible, with long-term investments in canine rabies surveillance, large-scale, data-driven MDV, public awareness creation, and application of WHO recommendations on human PEP (Cleaveland and Hampson 2017; Wallace et al. 2020).
For example, a recent MR project in the state of Goa, India, reported the elimination of human rabies and a greater than 90% reduction in monthly canine rabies (Gibson et al. 2022). Three predictable items formed the pillar of this program: MDV, targeting owned, confined, and free-roaming dogs, using a GIS-aided mobile phone app, guiding vaccination teams via assigned polygonal areas; the delivery of rabies education to school children and teachers on basic related knowledge, including how to avoid dog bites and what to do if bitten by a dog to receive medical care such as PEP; and enhanced surveillance through real-time reporting, using dedicated hotlines centrally managed to request dog vaccination and report unhealthy dogs. Based on reports, a full-scale investigation commenced to assess risk, including diagnosis, using the DFAT, rapid test kits for screening, and RABV sequencing to explore the utility of applied molecular epidemiology in disease forecasting management (Gibson et al. 2022). Plans are underway to repeat the Goa experience in other locales throughout India (Fig. 8).
Fig. 8.

Street poster, emulating responsible dog ownership, downtown Ranchi, India
Given such recent local success on the subcontinent, how would other national authorities with more limited resources be convinced to prioritize such a venture? An evidence-based evaluation of the national rabies burden (canine and human rabies) is critical as a prerequisite to driving such government investment. This would also include a plan to estimate dog populations, acquire adequate vaccines, supplies and staff, develop a program to vaccinate at least 70% dogs, and enhance laboratory capacity for canine rabies surveillance to assess success, including geographical risk assessments to better guide evidence-based interventions and public awareness creation to break the transmission cycle, consistent with long-standing recommendations (Lembo et al. 2012). As one example from West Africa, recent investigations in Nigeria began in an effort to provide multiple lines of evidence to contribute to more informed investment in national rabies prevention and control toward ZBT (Mshelbwala et al. 2021, 2022). Similar efforts are crucial elsewhere in Africa to drive evidence-based interventions toward the ZBT target.
However, with the approaching deadline of 2030 and with the evidence at hand, the ZBT target does not appear entirely feasible. Dogs thrive wherever there are humans, from the Arctic to the tropics. Livestock predation, bites and infectious diseases such as rabies are just a few of the recognized conflicts. Historically, dog ‘whippers’ or ‘wardens’ were employed to harm, catch, and kill dogs (Figs. 5 and 6). In Western countries, concomitant with the era of vaccination in the early twentieth century, with an increasing concern for general welfare, free-ranging animal management consisted largely of local removal, with captured individuals held in shelters for adoption or euthanasia (Fig. 7). Parallel industries in LMICs are much more limited in scope and operation, but gradual improvements in animal welfare have begun. Local education campaigns toward the promotion of vaccination, contraception and more holistic practices have had a major impact in some parts of Africa and Asia (Fig. 8). However, animal birth control strategies that spay/neuter and vaccinate dogs, returning them to the streets, do not alleviate all societal concerns. In turn, residents may demand the removal of dogs from their neighborhoods or result in the recidivism of local culling. Without further appreciation of such ignored facets as the local socioeconomic dynamics for broader support of sustainable animal management and responsible pet ownership, together with relevant operational research on the community ecology of dogs in line with vaccination, the likely success of the ZBT ‘…is slight and hardly realistic…’ (Subedi et al. 2022).
Fig. 7.

Dog catchers, 1924. Library of Congress, Prints and Photographs Division, Washington, D.C. 20,540 USA http://hdl.loc.gov/loc.pnp/pp.print
Riposte for therapy of clinical cases
In contrast to reasonably clear disease prevention and control considerations, one of the major, unresolved ‘holy grail’ issues in the field concerns the actual treatment of clinical rabies. Ever since the recognition of Matthew Winkler as the first documented human recovery during the 1970s, the hope continued in the quest for other survivors, but without a clear path or certainty of protocol, beyond excellent clinical support (Hattwick et al. 1972). Notably, there are many hurdles in considering such human treatment (Table 2). Clearly, pathobiology remains a fertile area, as many basic ‘infection and immunity’ features, such as cell signaling pathways (especially innate immune response), including the upregulation of TLRs and chemokine receptors for the recruitment of leukocytes and cytokines in the CNS, among others, remain an enigma.
Table 2.
Challenges in the implementation of human rabies treatment
| Number | Challenges |
|---|---|
| 1 | Historical fear and ignorance of a virulent pathogen |
| 2 | Extensive CNS involvement by time of diagnostic suspicion |
| 3 | Not an outpatient option |
| 4 | Unvaccinated healthcare workers |
| 5 | Need for a ‘dream team’ of intensive care specialists |
| 6 | Dedicated intensive care facility |
| 7 | No licensed antiviral drugs, requiring heroic use |
| 8 | Uncertain pathobiological targets and mechanisms |
| 9 | Delivery of biologics across the BBB |
| 10 | Clinical and diagnostic difficulties in monitoring patient status |
| 11 | Costs of therapy |
| 12 | Extensive rehabilitation |
| 13 | Quality of life in the face of neurological deficits |
| 14 | Religious, cultural, and ethical concerns on ‘experimentation’ |
| 15 | High probability of failure |
Recognizing such formidable drawbacks and that previous survivors had received PEP before the onset of illness, recommendations were made for aggressive considerations to management, if attempted, and the imperative need for research advances in pathobiological understanding and improvement in available tools (Jackson et al. 2003). During 2004, Jeanna Giese, a 15-year-old from Wisconsin, USA, was exposed to a bat but did not receive PEP. Approximately 1 month later, Jeanna developed signs compatible with rabies. She was hospitalized and treated with therapeutic coma and N-methyl-d-aspartate receptor antagonist therapy, becoming the first documented survivor without a history of vaccination (Willoughby et al. 2005). Afterwards, adaptations occurred of this initial regimen, termed the Milwaukee Protocol (MP). Recognizing a relationship between a favorable clinical outcome and the induction of RABV antibodies in the CSF (such as in Jeanna’s situation), critical care suggestions were made to especially target rabies patient selection for treatment toward those with early VNA onset (Wilde et al. 2008). Multiple additional treatments with MP were attempted, with many failures, while Kaplan–Meier survival curves after initiation appeared significantly longer compared to contemporary controls (Medical College of Wisconsin 2022). Controversy over the MP persists (Jackson 2018). Nevertheless, Jeanna’s dramatic recovery provoked a veritable renaissance on the topic that rabies is not invariably fatal. This reinvigorated a drive for new evidence-based tools for administration during attempted treatment (not just via in vitro models), such as effective antiviral drugs and the delivery of efficacious antibodies across the blood–brain barrier (Du Pont et al. 2019; de Melo et al. 2022; Kimitsuki et al. 2022). Another method may be in situ production using recombinant viruses (Huang et al. 2021). For example, after IV administration, RABV antibodies are expressed within the brains of animals within days following adeno-associated viral vector antibody gene therapy (Baker et al. 2022).
As opposed to the obvious drawbacks of ad hoc nature human cases and experimental treatment, is there an alternative to other species in the interim? For example, given the prevalence of naturally occurring rabies in dogs, adoption of a treatment protocol would not be wanting for a shortage of cases or restrictions due to locale. In addition, there is a built-in welfare component to ensure that such animals are removed as a public health measure from the streets, where they are otherwise considered pariahs and killed (not euthanized) outright. Veterinarians are much more likely to receive PrEP in advance, rather than as an afterthought, due to occupational exposures and have a professional familiarity with zoonoses such as rabies. Moreover, the comparative costs (i.e., facility, equipment, drugs, staff, etc.) of operation should be considerably less expensive than in human medicine. As such, a canine model offers multiple advances to understanding the basic pathobiological mechanisms in a natural reservoir that have thwarted progress in humans and is also relevant for modern medical interventions that have been otherwise difficult (e.g., intubation, long-term anesthesia, imaging, sample acquisition, etc.) in some smaller-bodied taxa (i.e., rodents) for practical application (Knobel et al. 2022).
Robust biologics
Practical rabies management is not possible without the availability of biologics, as recognized for more than a century. In contrast to many emerging infectious diseases, vaccines for rabies are broadly pure, potent, safe and efficacious (Banyard et al. 2018). Rabies vaccines for domestic animals have a minimum duration of immunity claims of three years or longer (NASPHV 2016).
For humans receiving PrEP, no failures were reported over more than four decades of surveillance (Rao et al. 2022). In addition to the use of PrEP for individuals under occupational risk (e.g., veterinarians, laboratory workers, wildlife biologists, etc.) or travelers, administration to communities at risk is also being reevaluated (Royal et al. 2022; Taylor et al. 2022). Considering more than 12 million exposures per year, human PEP is also highly effective when administered appropriately and in a timely manner, with only 122 failures identified between 1980 and 2022 (Whitehouse et al. 2022). The institution of shorter schedules and dose-sparing strategies for both PrEP and PEP provides critical biologics to those most in need (World Health Organization (WHO), 2018). In addition to humans and domestic animals, oral vaccination of wildlife has led to widespread prevention and control of rabies in western Europe and North America. Oral vaccines, proven in concept and action for wildlife, may be perceived as a game changer if applied toward free-ranging community dogs (Bobe et al. 2023). However, this is not a new concept or a panacea and will only augment success where at least a rudimentary prevention and control program already exists, including MDV and laboratory-based surveillance (Cliquet et al. 2018). All of these applications were in effect during the twentieth century.
Additional technological applications have broadened the spectrum even further for prevention and control with regard to new biologics in use or under development (Ertl 2019). For example, after initial design in 1978, monoclonal antibodies (MAbs) were finally being used in human PEP as an alternative to RIG (de Melo et al. 2022). This was in no small way prompted by a major collaboration among the WHO Collaborating Centers for such technology transfer and realization postconcept (Müller et al. 2009; Phoolcharoen et al. 2019). Besides prophylaxis, MAbs are also under consideration for treatment (de Melo et al. 2020). Although the use of virus-like particles (VLPs) was considered for veterinary applications for several years, during 2022, VLP nanoparticles were used for a 1-week, 3-dose human application (Ravish et al. 2021). Most human vaccines are not adjuvanted, but a polyinosinic-polycytidylic acid-based adjuvant stabilized with kanamycin and calcium (acting as a TLR3 agonist) was well tolerated and immunogenic when tested in healthy volunteers (Wijaya et al. 2017). Highly purified, serum-free human vaccines are on the horizon (Pichon et al. 2022). Using a very different application via nucleic acid delivery, after two administered doses of a RABV glycoprotein mRNA encapsulated in lipid nanoparticles, all recipients developed VNA (Aldrich et al. 2021). To reduce the need for a return visit to the clinic for a booster dose and consider only a single dose of vaccine for PrEP, clinical trials have begun to evaluate a simian adeno-RABV product (Jenkin et al. 2022). Many of these new and existing biologics, whether administered to humans or animals, are intended for a single application of one dose per individual. In contrast, recently, the idea of ‘self-spreading’ vaccines has re-emerged into the foreground (Griffiths et al. 2023).
A role for ‘self-spreading’ vaccines?
Vaccines represent one of the most successful biomedical technologies in history. In a One Health context, human rabies vaccines were created during the late nineteenth century, dog vaccines were applied a century ago, and wildlife vaccines were developed over 50 years ago. Paradoxically, regardless of the pathogen, such success brings a risk of failing to adequately appreciate all the diverse challenges that have been overcome in the vaccine field to date (Messinger Cayetano and Crandall 2020). One example is the concept of ‘self-spreading’ vaccines (Nuismer et al. 2016, 2018; Scudellari 2016; Bull et al. 2018; Bakker et al. 2019; Cogley 2020; Nuismer and Bull 2020). In contrast to all licensed vaccines to date that are delivered to the individual by parenteral or mucosal routes, self-spreading vaccines are replication competent and intended to spread autonomously in the environment between individuals, often with epidemic-like dynamics. This outwardly attractive idea would enable a small fraction of a target population to be inoculated, with an expectation of achieving a population-wide herd immunity effect, which may extend to as yet unborn members of the same population at the time of initiation.
The basic concept of self-spreading vaccines is not new and occurred to generations of virologists and vaccine developers (Browne 1991; Ross 1998; Angulo and Bárcena 2007; Angulo and Gilna 2008; Lentzos et al. 2022). For example, the tools necessary to assemble such genetically modified biologics were adequate in the 1990s to generate a self-spreading vaccine aimed at protecting wild native Spanish rabbits from myxomatosis and rabbit hemorrhagic disease (Bárcena et al. 2000; Torres et al. 2001; Angulo and Bárcena 2007). This product was tested in a field release on a Spanish island, although efforts to license the vaccine (under the name LAPINVAC) with the European Medical Agency were ultimately abandoned by 2007.
Self-spreading vaccines are genetically modified organisms, typically tested in biosafety level 3 or 4 facilities (Bárcena et al. 2000; Tsuda et al. 2011). Somewhat confusingly, self-spreading vaccines exist under a multitude of equivalent terms that may include transmissible, contagious, horizontally transferable, self-disseminating, or founder-based. A hypothetical term, “transferable vaccine”, has also been introduced to denote self-spreading vaccines, where intended transmission only occurs to those in direct contact with the originally inoculated individuals (Nuismer and Bull 2020; Technology Networks 2022). However, we are unaware of any evidence that such a class of licensed agents exists, particularly as viral transmissibility is a dynamic parameter in complex environmental situations.
Currently, there are several proposals for self-spreading vaccines:
1. Vaccination of African nonhuman primates to inhibit their infection by Ebola virus, with the aim of limiting their capacity to act as a wildlife reservoir for transmission to humans (PREEMPT 2018, 2022; TVG 2021).
2. Vaccination of rodent species in West Africa to inhibit their infection by Lassa fever virus, with the aim of limiting their capacity to act as a wildlife reservoir for transmission to humans (PREEMPT 2018, 2022; Regulatory News Service 2022).
3. Vaccination of North American bat species to reduce their susceptibility to an emergent fungal infection for the purposes of bat conservation against White Nose Syndrome (Rocke et al. 2019; USDA-APHIS 2019).
4. Vaccination of vampire bats is currently restricted to Central and South America to inhibit their infection by RABV, with the aim of limiting their capacity to act as a wildlife reservoir for transmission to humans and other animals (Bakker et al. 2019; Streicker et al. 2022).
In this communication concentrated on rabies, we focus on the fourth proposal, involving vampire bats (although such a concept could be applicable to other reservoirs). Vampire bats are distributed throughout Central and South America, from Mexico to Argentina (Johnson et al. 2014). As obligate parasites of vertebrates, RABV transmission occurs during feeding by these hematophagous species. Ideally, population reduction is used proactively to manage vampire populations. In bite victims, rabies prevention consists of individual vaccination of humans and domestic animals. Even allowing for the undoubted unevenness of regional disease monitoring and control efforts, the incidence of human rabies has declined to the point that the vast majority of cases now occur outside of the Americas (WHO 2023). Due unambiguously to the collective success of MDV and PEP efforts, most of the residual ‘vectoring’ of rabies in Central and South America is now due to vampire bats. This success using traditional biologics has contributed to the proposal to use self-spreading vaccines to limit the capacity of vampire bats to transmit RABV, in part by exploiting their roosting and grooming behaviors to facilitate viral vaccine spreading (Bakker et al. 2019; Nuismer and Bull 2020).
Recent modeling has suggested the intuitive finding that self-spreading vaccines are more resource efficient than conventionally deployed biologics (Nuismer et al. 2016, 2018, 2019). The question of whether self-spreading viral vaccines are likely to be operationally useful in their target remains largely unaddressed. For example, will they simultaneously be ‘transmissible enough’ to be successful and still be perceived as ‘sufficiently controllable’ by regulators and generally acceptable to the public? Given the variability in continuously dynamic model parameters, such as viral transmissibility, and the stability of genetic modification(s) introduced to the vaccine virus genome, it is difficult to discern the extent to which human communities could be reasonably encouraged to rely on such management interventions for infectious spillover protection. For example, such applications would be contrasted otherwise to well-resourced and coordinated application of available and conventionally deployed public health and veterinary technologies to date. As such, the literature promoting self-spreading vaccines in the context of zoonotic disease control frequently focuses on the exact goal of local elimination of pathogens from wildlife, rather than their stable suppression, per se. Rather lamentably, there is a conspicuous and repeated effort to rebrand current One Health methods as somehow “reactive”, whereas self-spreading vaccines are presented as innovatively “proactive” (Kerlin 2019). Such trivialization of the achievements of successive generations of researchers and local community control practitioners could be viewed as falling squarely within the paradox of success (Messinger Cayetano and Crandall 2020).
During the 1980s to 2000s, other programs considering self-spreading vaccines collapsed (Lentzos et al. 2022; Reeves 2022). Regardless of whether current proposals progress to the point that they could be argued to be not only safe but also trustworthy, useful, and effective, it is self-evident that in the current media environment, such concepts for self-spreading vaccines may construe a significant misinformation threat. This should be viewed in light of the recent erosion of confidence and vaccine hesitancy in some, but not all, parts of the globe (de Figueiredo et al. 2020). Given the importance of public trust to not only vaccine deployment but also the strategically essential capacity to innovate and develop new conventionally deployed biologics, caution by both proposers and funders of self-spreading speculations would appear to be warranted. In the absence of the need for technological breakthroughs, the speed at which self-spreading vaccines could be developed is quite short. Over the past three years, self-spreading vaccines for Lassa fever virus in rodents and Ebola virus in primates have been developed in Germany, the USA, and the UK (PREEMPT 2022; Regulatory News Service 2022; Technology Networks 2022). Furthermore, the first open field trial of the rabbit vaccine during 1999 predated any scientific publications on self-spreading vaccines (Torres et al. 2001). Such precedence appears problematic in the absence of international notifications for cross-border export of self-spreading vaccines or experimental field releases of veterinary self-spreading vaccines. As the broader vaccine community focuses on moving away from higher risk approaches where effective alternatives can be developed (i.e., replacing live attenuated vaccines generated by selection), there is considerable potential for the whole field to be represented by a small group of funders and investigators (mostly evolutionary biologists), moving rapidly in the direction of increasing not only the risk profile of vaccines but also the potential for misunderstanding or misinformation (transmissiblevaccines.org 2022). In the context of rabies prevention and control, it is reasonable to ask to whom self-spreading vaccine proposals would appear attractive, particularly if licensed biologics were being promoted and used more widely in at-risk populations. For example, how would success be measured? In the case of rabies, would exposed individuals still receive PEP? What techniques could manage vampire bat populations and their obligatory feeding behavior upon humans, domestic animals, and wildlife? At the very least, self-spreading RABV vaccines are no magic bullet, and such proposals involve multiple ethical, regulatory, scientific, and social issues, in need of much broader discussion.
Conclusions
Recognizing the 200th anniversary of his birth, aficionados of Pasteur would likely agree that ‘chance favors the prepared mind’, as applied to the considerable contributions that flowed thereafter, following his celebratory accomplishments on rabies vaccination (Tarantola 2017). When compared to the past century, major scientific and operational advances occurred in nearly all aspects of the field, providing a major example of One Health in action (Table 3). By comparison, given the underlying technical prerequisites needed for laboratory-based surveillance, vaccine-driven herd immunity, and basic epidemiological understanding that was central to the true eradication of smallpox and rinderpest, one might understand the tendency to view such significant zoonoses as rabies in the same light. Unfortunately, these are different realities. Smallpox was highly communicable, restricted to one host, killed millions annually and was eradicated globally by affordable vaccination for the public good under the auspices of the WHO, with ongoing issues related to other orthopoxviruses today (Gieryńska et al. 2023). In contrast, although rinderpest was not zoonotic and involved more than a single reservoir, its economic consequences upon ruminants (and the farming communities dependent upon them) were devastating to LMICs, prompting its eradication via vaccination (producing life-long immunity) under the auspices of the FAO (Jeggo and Roeder 2021). By simple comparison to these only two eradicated diseases to date (and similarly with polio, now more than two decades long overdue), such terminology is not applicable to rabies, promising much more than could ever be achieved.
Table 3.
Paradigm shifts in rabies detection, prevention, control & treatment in the 21st century
| Topic | Change | Reference |
|---|---|---|
| Etiology | Classical rabies virus recognized as the most important member of the genus, but > 17 putative or recognized lyssaviruses, including highly divergent viral spp. | Fooks et al. 2021 |
| Diagnosis | Since the early twentieth century, microscopic recognition of intracytoplasmic ‘viral factories’ within neurons, now supplemented by highly sensitive, specific, and economical tests for rapid ante- and postmortem laboratory confirmation and enhanced point-of-care field surveillance development | World Health Organization (WHO), 2018 |
| Pathobiology | In addition to exposure via transdermal, mucosal and aerosol routes, transmission via solid organ and tissue transplantation | Srinivasan et al. 2005 |
| Treatment | Survivors, without a history of prior vaccination | Willoughby et al. 2005 |
| Safety | Abandonment of nerve tissue-based vaccines by safer alternatives | Gongal & Sampath 2019 |
| Vaccine potency | In vitro alternates to the use of animal testing in the NIH test | Stokes et al. 2012 |
| Passive immunity | Monoclonal antibodies used increasingly in humans | Sparrow et al. 2019 |
| Improvement in biologics | Multiple novel biologics licensed or in human clinical trials | Ertl 2019 |
| Prophylaxis | Efficacious and economical dose-sparing and 1-week strategies | WHO Rabies Modeling Consortium 2019 |
| Wildlife rabies vaccination | Elimination of rabies in red foxes in western Europe, and Texas gray fox and coyote rabies virus variants in USA | Mähl et al. 2014; Maki et al. 2017; Vos et al. 1999 |
| One Health | Global program for elimination of human rabies caused via dogs by 2030 | Abela-Ridder et al. 2016 |
| Advocacy | Inauguration of World Rabies Day | Centers for Disease Control Prevention (CDC), 2007 |
Whereas progress in rabies prevention and control is inarguable, eradication does not apply as a term in the objective light of considerable viral and host diversity and the subjective socioeconomic bias of neglect and disparity. In contrast to eradication, based upon the major regional success in the Americas, the GEHRD might seem achievable, conducted with twentieth century tools (although even here limitations are obvious and continuing). Additionally, the ZBT appeared more reasonable at its forecast during 2015, especially with the added promise of global collaboration and the prescient reliance upon the technical shape of things to come (Abela-Ridder et al. 2016). Compared to other diseases, rabies is a rather low bar to achieve reasonable prevention and control by conservative standards, overshadowed only by the even lower bar of political commitment and resources enacted to date. If two of the largest growing global economies (i.e., China and India) would more actively advocate for and financially support rabies elimination campaigns among dogs in ASEAN/Far East and the Indian subcontinent, more than 50% human rabies deaths would be prevented.
The COVID-19 pandemic refocused time, talent, and resources from all NTD. Progress in rabies surveillance, prevention, and control has suffered, especially in LMICs. In retrospect, PAHO set at least four different target dates for elimination in the Americas over the past 30 years, but the region is still not free of canine rabies (Del Rio Vilas et al. 2017). As such, in light of such major setbacks and the minimal SARE scores representative today as one bellwether in most canine rabies enzootic countries, a global reappraisal is warranted unless substantive global support materializes (such as from GAVI), together with a prioritized political will for the application of modern human PEP and appropriate MDV (Thumbi et al. 2022). Before elimination can accrue tomorrow, practical control needs to begin today with available tools and techniques.
Acknowledgements
We thank the editors for their kind invitation to contribute to this special issue on zoonoses, the three anonymous reviewers for their helpful comments and our colleagues around the world for their continued investment of time, talent and treasure toward global rabies prevention and control, regardless of drawbacks and deadlines.
Abbreviations
- BMGF
Bill & Melinda Gates Foundation
- CDC
Centers for Disease Control & Prevention
- CST
Cross species transmission
- DFAT
Direct fluorescent antibody test
- FAO
Food and Agriculture Organization of the United Nations
- GARC
Global Alliance for Rabies Control
- GAVI
The vaccines alliance
- GEHRD
Global elimination of human rabies from dogs
- ICTV
International Committee for the Taxonomy of Viruses
- ISAVET
In-Service Applied Veterinary Epidemiology Training
- KZN
KwaZulu-Natal
- LMIC
Lower- and middle-income countries
- MAb
Monoclonal antibodies
- MDV
Mass dog vaccination
- MR
Mission Rabies
- NASPHV
National Association of State Public Health Veterinarians
- NTD
Neglected Tropical Diseases
- OIE/WOAH
World Organization for Animal Health
- PAHO
Pan American Health Organization
- PARACON
Pan-African Rabies Control Network
- PEP
Postexposure prophylaxis
- PrEP
Preexposure prophylaxis
- RABV
Rabies virus
- RIG
Rabies immune globulin
- RFFIT
Rapid fluorescent antibody test
- RTSL
Resolve To Save Lives
- SAARC
South Asian Association for Regional Cooperation
- SARE
Stepwise approach to rabies elimination
- VLP
Virus-like particle
- VNA
Virus neutralizing antibody
- WHO
World Health Organization
- WRD
World Rabies Day
- ZBT
‘Zero by thirty
Authors’ contributions
CER conceived the concept for the review. CER, PM, RGR, and IVK wrote, reviewed, and approved the manuscript.
Funding
Not applicable.
Availability of data and materials
Not applicable.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- Abdul-Jalil, M. 2020. Working to remove the fear of rabies from Kabul, Afghanistan. 10.2307/j.ctv10crcv7.93. https://endrabiesnow.org/stories/view/working-to-remove-the-fear-of-rabies-from-kabul-afghanistan
- Abela-Ridder B, Knopf L, Martin S, Taylor L, Torres G, De Balogh K. 2016: The beginning of the end of rabies? The Lancet Global Health. 2016;4(11):e780–e781. doi: 10.1016/S2214-109X(16)30245-5. [DOI] [PubMed] [Google Scholar]
- Acharya KP, Subedi D, Wilson RT. Rabies control in South Asia requires a One Health approach. One Health. 2021;12:100215. doi: 10.1016/j.onehlt.2021.100215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aldrich C, Leroux-Roels I, Huang KB, Bica MA, Loeliger E, Schoenborn-Kellenberger O, Walz L, Leroux-Roels G, von Sonnenburg F, Oostvogels L. Proof-of-concept of a low-dose unmodified mRNA-based rabies vaccine formulated with lipid nanoparticles in human volunteers: A phase 1 trial. Vaccine. 2021;39:1310–1318. doi: 10.1016/j.vaccine.2020.12.070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amoako YA, El-Duah P, Sylverken AA, Owusu M, Yeboah R, Gorman R, Adade T, Bonney J, Tasiame W, Nyarko-Jectey K, et al. Rabies is still a fatal but neglected disease: a case report. Journal of Medical Case Reports. 2021;15(1):575. doi: 10.1186/s13256-021-03164-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amparo, A.C.B., E.C.B. Mendoza, D.A. Licuan, L.M. Valenzuela, J.D. Madalipay, S.I. Jayme, and L.H. Taylor. 2019. Impact of integrating rabies education Into the curriculum of public elementary schools in Ilocos Norte, Philippines on rabies knowledge, and animal bite incidence. Frontiers in Public Health 7: 119. 10.3389/fpubh.2019.00119. [DOI] [PMC free article] [PubMed]
- Angulo E, Bárcena J. Toward a unique and transmissible vaccine against myxomatosis and rabbit hemorrhagic disease for rabbit populations. Wildlife Research. 2007;34:567. doi: 10.1071/WR06160. [DOI] [Google Scholar]
- Angulo E, Gilna B. When biotech crosses borders– international governance of self-dispersive GMOs purposefully released for public health, controlling invasive species and pests, and treating wildlife. Nature Biotechnology. 2008 doi: 10.1038/nbt0308-277. [DOI] [PubMed] [Google Scholar]
- Anonymous Time to eliminate rabies. Lancet. 2015;386:2446. doi: 10.1016/S0140-6736(15)01287-8. [DOI] [PubMed] [Google Scholar]
- Anothaisintawee T, Julienne Genuino A, Thavorncharoensap M, Youngkong S, Rattanavipapong W, Meeyai A, Chaikledkaew U. Cost-effectiveness modeling studies of all preventive measures against rabies: A systematic review. Vaccine. 2019;37(Suppl 1):A146–A153. doi: 10.1016/j.vaccine.2018.11.071. [DOI] [PubMed] [Google Scholar]
- Aréchiga Ceballos, N., P. Puebla Rodríguez, and S.Á. Aguilar. 2022. The new face of human rabies in Mexico, what’s next after eradicating rabies in dogs. Vector Borne and Zoonotic Diseases 22: 69–75. 10.1089/vbz.2021.0051. [DOI] [PubMed]
- Audu SW, Mshelbwala PP, Jahun BM, Bouaddi K, Weese JS. Two fatal cases of rabies in humans who did not receive rabies postexposure prophylaxis in Nigeria. Clinical Case Report. 2019;7:749–752. doi: 10.1002/ccr3.1972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baby, J., R.S. Mani, S.S. Abraham, A.T. Thankappan, P.M. Pillai, A.M. Anand, S.N. Madhusudana, J. Ramachandran, and S. Sreekumar. 2015. Natural rabies infection in a domestic fowl (Gallus domesticus): A report from India. PLoS Neglected Tropical Diseases 9 (7): e0003942. 10.1371/journal.pntd.0003942. [DOI] [PMC free article] [PubMed]
- Badrane, H., and N. Tordo. 2001. Host switching in Lyssavirus history from the Chiroptera to the Carnivora orders. Journal of Virology 75 (17): 8096–8104. 10.1128/jvi.75.17.8096-8104.2001. [DOI] [PMC free article] [PubMed]
- Badrane, H., C. Bahloul, P. Perrin, and N. Tordo. 2001. Evidence of two Lyssavirus phylogroups with distinct pathogenicity and immunogenicity. Journal of Virology 75 (7): 3268–3276. 10.1128/JVI.75.7.3268-3276.2001. [DOI] [PMC free article] [PubMed]
- Bajeli-Datt, K., 2022. Two hundred rabies deaths in eighteen states, Karnataka tops with thirty-two. The New Indian Express, 10 December. https://www.newindianexpress.com/nation/2022/dec/10/250-rabies-deaths-in-18-states-karnataka-tops-with-32-2526778.html
- Baker HJ, Martin DR, Gross AL, Chamorro MF, Naskou MC, Johnson AK, Brock KV, Van Kampen KR, Willoughby RE. Rabies: Who should care? Journal of the American Veterinary Medical Association. 2022;7:1–5. doi: 10.2460/javma.22.09.0420. [DOI] [PubMed] [Google Scholar]
- Bakker KM, Rocke TE, Osorio JE, Abbott RC, Tello C, Carrera JE, Valderrama W, Shiva C, Falcon N, Streicker DG. Fluorescent biomarkers demonstrate prospects for spreadable vaccines to control disease transmission in wild bats. Nature Ecology Evolution. 2019;3:1697–1704. doi: 10.1038/s41559-019-1032-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banyard AC, Mcelhinney LM, Johnson N, Fooks AR. History of rabies control by vaccination. Revue Scientifique et Technique. 2018;37:305–322. doi: 10.20506/rst.37.2.2804. [DOI] [PubMed] [Google Scholar]
- Bárcena, J., M. Morales, B. Vázquez, J.A. Boga, F. Parra, J. Lucientes, A. Pagès-Manté, J.M. Sánchez-Vizcaíno, R. Blasco, and J.M. Torres. 2000. Horizontal transmissible protection against myxomatosis and rabbit hemorrhagic disease by using a recombinant myxoma virus. Journal of Virology. 74: 1114–1123. 10.1128/jvi.74.3.1114-1123.2000https://www.ncbi.nlm.nih.gov/pmc/articles/PMC111445/. [DOI] [PMC free article] [PubMed]
- Barroga TR, Gordoncillo MJ, Lagayan MG, Bernales R, Caniban M, Lopez E, Abila R. Practical intersectoral linking: Tool to rabies One Health coordination to the grass-roots level. Zoonoses and Public Health. 2018;65:805–814. doi: 10.1111/zph.12502. [DOI] [PubMed] [Google Scholar]
- Bhat, M.D., P. Priyadarshini, C. Prasad, and K. Kulanthaivelu. 2021. Neuroimaging findings in rabies encephalitis. Journal of Neuroimaging 31 (3): 609–614. 10.1111/jon.12833. [DOI] [PubMed]
- Bobe K, Ortmann S, Kaiser C, Perez-Bravo D, Gethmann J, Kliemt J, Körner S, Theuß T, Lindner T, Freuling C, et al. Efficacy of oral rabies vaccine baits containing SPBN GASGAS in domestic dogs according to international standards. Vaccines (Basel). 2023;11:307. doi: 10.3390/vaccines11020307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bourhy, H., J.A. Cowley, F. Larrous, E.C. Holmes, and P.J. Walker. 2005. Phylogenetic relationships among rhabdoviruses inferred using the L polymerase gene. Journal of General Virology 86 (Pt 10): 2849–2858. 10.1099/vir.0.81128-0. [DOI] [PubMed]
- Brochier B, Kieny MP, Costy F, Coppens P, Bauduin B, Lecocq JP, Languet B, Chappuis G, Desmettre P, Afiademanyo K, et al. Large-scale eradication of rabies using recombinant vaccinia-rabies vaccine. Nature. 1991;354:520–522. doi: 10.1038/354520a0. [DOI] [PubMed] [Google Scholar]
- Browne, M. W., 1991. New animal vaccines spread like diseases. The New York Times. https://www.nytimes.com/1991/11/26/science/new-animal-vaccines-spread-like-diseases.html
- Bull, J.J., M.W. Smithson, and S.L. Nuismer. 2018. Transmissible viral vaccines. Trends in Microbiology. 26: 6–15. 10.1016/j.tim.2017.09.007. [DOI] [PMC free article] [PubMed]
- Caraballo DA, Lema C, Novaro L, Gury-Dohmen F, Russo S, Beltrán FJ, Palacios G, Cisterna DM. A novel terrestrial rabies virus lineage occurring in South America: origin, diversification, and evidence of contact between wild and domestic cycles. Viruses. 2021;13(12):2484. doi: 10.3390/v13122484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Centers for Disease Control & Prevention (CDC) Notice to readers. World Rabies Day – September 8, 2007. MMWR. 2007;56(35):915. [Google Scholar]
- Cleaveland S, Hampson K. Rabies elimination research: Juxtaposing optimism, pragmatism and realism. Proceedings of the Biological Sciences. 2017;284:20171880. doi: 10.1098/rspb.2017.1880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cliquet F, Guiot AL, Aubert M, Robardet E, Rupprecht CE, Meslin FX. Oral vaccination of dogs: A well-studied and undervalued tool for achieving human and dog rabies elimination. Veterinary Research. 2018;49:61. doi: 10.1186/s13567-018-0554-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coertse J, Geldenhuys M, le Roux K, Markotter W. Lagos Bat Virus, an Under-Reported Rabies-Related Lyssavirus. Viruses. 2021;13(4):576. doi: 10.3390/v13040576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coetzer A, Kidane AH, Bekele M, Hundera AD, Pieracci EG, Shiferaw ML, Wallace R, Nel LH. The SARE tool for rabies control: Current experience in Ethiopia. Antiviral Research. 2016;135:74–80. doi: 10.1016/j.antiviral.2016.09.011. [DOI] [PubMed] [Google Scholar]
- Cogley, M., 2020. Could self-spreading vaccines stop a coronavirus pandemic? The Telegraph (UK). https://www.telegraph.co.uk/technology/2020/01/28/could-self-spreading-vaccines-stop-global-coronavirus-pandemic/
- Conselheiro JA, Barone GT, Miyagi SAT, de Souza Silva SO, Agostinho WC, Aguiar J, Brandão PE. Evolution of rabies virus isolates: virulence signatures and effects of modulation by neutralizing antibodies. Pathogens. 2022;11(12):1556. doi: 10.3390/pathogens11121556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cruz M., 2022. Anti-rabies program a failure–lawmaker. The Manila Times, 29 November. https://www.manilatimes.net/2022/11/29/news/national/anti-rabies-program-a-failure-lawmaker/1868242
- Daniels SA, King EM, Olivier CJ, Harding JP, Fehlner-Gardiner C, Nadin-Davis S, Murray MC. Rabies virus infection in a 21-year-old male presenting with ascending paralysis after a bat scratch. Journal of the Association of Medical Microbiology and Infectious Disease Canada. 2020;5:201–208. doi: 10.3138/jammi-2020-0007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Figueiredo A, Simas C, Karafillakis E, Paterson P, Larson HJ. Mapping global trends in vaccine confidence and investigating barriers to vaccine uptake: A large-scale retrospective temporal modeling study. Lancet. 2020;396:898–908. doi: 10.1016/S0140-6736(20)31558-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Melo GD, Sonthonnax F, Lepousez G, Jouvion G, Minola A, Zatta F, Larrous F, Kergoat L, Mazo C, Moigneu C, et al. A combination of two human monoclonal antibodies cures symptomatic rabies. EMBO Molecular Medicine. 2020;12:e12628. doi: 10.15252/emmm.202012628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Melo GD, Hellert J, Gupta R, Corti D, Bourhy H. Monoclonal antibodies against rabies: current uses in prophylaxis and in therapy. Current Opinion in Virology. 2022;53:101204. doi: 10.1016/j.coviro.2022.101204. [DOI] [PubMed] [Google Scholar]
- De Silva AP, Harischandra PL, Beane A, Rathnayaka S, Pimburage R, Wijesiriwardana W, Gamage D, Jayasinghe D, Sigera C, Gunasekara A, et al. A data platform to improve rabies prevention Sri Lanka. Bulletin of the World Health Organization. 2017;95:646–651. doi: 10.2471/BLT.16.188060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Del Rio Vilas VJ, Freire de Carvalho MJ, Vigilato MA, Rocha F, Vokaty A, Pompei JA, Molina Flores B, Fenelon N, Cosivi O. Tribulations of the last mile: sides from a regional program. Frontiers in Veterinary Science. 2017;4:4. doi: 10.3389/fvets.2017.00004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deray R, Rivera C, Gripon S, Ulanday C, Roces MC, Amparo AC, Attlan M, Demont C, Kieffer A, Miranda ME. Protecting children from rabies with education and preexposure prophylaxis: A school-based campaign in El Nido, Palawan, Philippines. PLoS One. 2018;13:e0189596. doi: 10.1371/journal.pone.0189596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diana, N.K., K.C. Mitchell, and K.A. Feldman. 2015. Shedding light on rabies in opossums. Journal of the American Veterinary Medical Association 247 (11): 1229. PMID: 26925485. [PubMed]
- Dowdle WR, Cochi SL. The principles and feasibility of disease eradication. Vaccine. 2011;29(Suppl 4):D70–D73. doi: 10.1016/j.vaccine.2011.04.006. [DOI] [PubMed] [Google Scholar]
- Du Pont, V., R.K. Plemper, and M.J. Schnell. 2019. Status of antiviral therapeutics against rabies virus and related emerging lyssaviruses. Current Opinion in Virology 35: 1–13. 10.1016/j.coviro.2018.12.009. [DOI] [PMC free article] [PubMed]
- Elser JL, Hatch BG, Taylor LH, Nel LH, Shwiff SA. Toward canine rabies elimination: Economic comparisons of three project sites. Transboundary and Emerging Diseases. 2018;65(1):135–145. doi: 10.1111/tbed.12637. [DOI] [PubMed] [Google Scholar]
- Ertl, H.C.J. 2019. New rabies vaccines for use in humans. Vaccines (basel). 7 (2): 54. 10.3390/vaccines7020054. [DOI] [PMC free article] [PubMed]
- Fasina FO, Fasanmi OG, Makonnen YJ, Bebay C, Bett B, Roesel K. The one health landscape in Sub-Saharan African countries. One Health. 2021;13:100325. doi: 10.1016/j.onehlt.2021.100325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flamand A, Coulon P, Lafay F, Kappeler A, Artois M, Aubert M, Blancou J, Wandeler AJ. Eradication of rabies in Europe. Nature. 1992;360:115–116. doi: 10.1038/360115a0. [DOI] [PubMed] [Google Scholar]
- Fooks, A.R., R. Shipley, W. Markotter, N. Tordo, C.M. Freuling, T. Müller, L.M. McElhinney, A.C. Banyard, and C.E. Rupprecht. 2021. Renewed public health threat from emerging Lyssaviruses. Viruses 13 (9): 1769. 10.3390/v13091769. [DOI] [PMC free article] [PubMed]
- Freire de Carvalho, M., M.A.N. Vigilato, J.A. Pompei, F. Rocha, A. Vokaty, B. Molina-Flores, O. Cosivi, and Vilas VJ.. Del Rio. 2018. Rabies in the Americas: 1998–2014. PLoS Neglected Tropical Diseases. 12: e0006271. 10.1371/journal.pntd.0006271. [DOI] [PMC free article] [PubMed]
- Gamalathge, P.U., H.E. Carter, O. Wimalaratne, and S. Kularatne. 2019. Developing and evaluating an intervention to improve care and reduce costs of rabies postexposure therapy: An observational quality improvement initiative in selected vaccination units in Srilanka. International Journal of Evidence-Based Healthcare 17: 157–163. 10.1097/XEB.0000000000000172. [DOI] [PubMed]
- Gan H, Hou X, Wang Y, Xu G, Huang Z, Zhang T, Lin R, Xue M, Hu H, Liu M, et al. Global burden of rabies in 204 countries and territories, from 1990 to 2019: results from the Global Burden of Disease Study 2019. International Journal of Infectious Diseases. 2023;2023(126):136–144. doi: 10.1016/j.ijid.2022.10.046. [DOI] [PubMed] [Google Scholar]
- Gérard FCA, Bourhis JM, Mas C, Branchard A, Vu DD, Varhoshkova S, Leyrat C, Jamin M. Structure and dynamics of the unassembled nucleoprotein of rabies virus in complex with its phosphoprotein chaperone module. Viruses. 2022;14(12):2813. doi: 10.3390/v14122813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghai S, Hemachudha T. Continued failure of rabies elimination-consideration of challenges in applying the one health approach. Frontiers in Veterinary Science. 2022;9:847659. doi: 10.3389/fvets.2022.847659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghosh S, Rana MS, Islam MK, Chowdhury S, Haider N, Kafi MAH, Ullah SM, Shah MRA, Jahan AA, Mursalin HS, et al. Trends and clinico-epidemiological features of human rabies cases in Bangladesh 2006–2018. Science and Reports. 2020;10:2410. doi: 10.1038/s41598-020-59109-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibson AD, Yale G, Corfmat J, Appupillai M, Gigante CM, Lopes M, Betodkar U, Costa NC, Fernandes KA, Mathapati P, et al. Elimination of human rabies in Goa, India through an integrated One Health approach. Nature Communications. 2022;13(1):2788. doi: 10.1038/s41467-022-30371-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gieryńska M, Szulc-Dąbrowska L, Struzik J, Gregorczyk-Zboroch KP, Mielcarska MB, Toka FN, Schollenberger A, Biernacka Z. Orthopoxvirus Zoonoses-do we still remember and are ready to fight? Pathogens. 2023;12:363. doi: 10.3390/pathogens12030363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilbert AT. Rabies virus vectors and reservoir species. Revue Scientifique et Technique. 2018;37(2):371–384. doi: 10.20506/rst.37.2.2808. [DOI] [PubMed] [Google Scholar]
- Global Alliance for Rabies Control (GARC). 2022. https://rabiesalliance.org/
- Goel K, Sen A, Satapathy P, Kumar P, Aggarwal AK, Sah R, Padhi BK., 2023. Human rabies control in the era of post-COVID-19: a call for action. Journal of Travel Medicine. taad009. 10.1093/jtm/taad009. [DOI] [PubMed]
- Gongal G, Sampath G. Introduction of intradermal rabies vaccination - A paradigm shift in improving postexposure prophylaxis in Asia. Vaccine. 2019;37(Suppl 1):A94–A98. doi: 10.1016/j.vaccine.2018.08.034. [DOI] [PubMed] [Google Scholar]
- Gongal G, Sampath G, Kishore J, Bastola A, Punrin S, Gunesekera A. The impact of COVID-19 pandemic on rabies postexposure prophylaxis services in Asia. Human Vaccines & Immunotherapeutics. 2022;18(5):2064174. doi: 10.1080/21645515.2022.2064174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- González-Roldán JF, Undurraga EA, Meltzer MI, Atkins C, Vargas-Pino F, Gutiérrez-Cedillo V, Hernández-Pérez JR., 2021. Cost-effectiveness of the national dog rabies prevention and control program in Mexico, 1990–2015. PLoS Neglected Tropical Diseases. 15:e0009130. 10.1371/journal.pntd.0009130. [DOI] [PMC free article] [PubMed]
- Griffiths, M.E., D.K. Meza, D.T. Haydon, and D.G. Streicker. 2023. Inferring the disruption of rabies circulation in vampire bat populations using a betaherpesvirus-vectored transmissible vaccine. Proceedings. National Academy of Sciences. United States of America 120: e2216667120. 10.1073/pnas.2216667120. [DOI] [PMC free article] [PubMed]
- Grimm, R. 2002. The history of the eradication of rabies in most European countries. Historia Medicinae Veterinariae 27: 295–301. PMID: 12508823. [PubMed]
- Grome, H.N., J. Yackley, D. Goonewardene, A. Cushing, M. Souza, A. Carlson, L. Craig, B. Cranmore, R. Wallace, L. Orciari, et al. 2022. Translocation of an Anteater (Tamandua tetradactyla) infected with rabies from virginia to tennessee resulting in multiple human exposures, 2021. MMWR. 71 (15): 533–537. 10.15585/mmwr.mm7115a1. [DOI] [PMC free article] [PubMed]
- Grossnickle DM, Newham E. Therian mammals experienced ecomorphological radiation during the Late Cretaceous and selective extinction at the K-Pg boundary. Proceedings of the Royal Society B. 2016;283:20160256. doi: 10.1098/rspb.2016.0256. [DOI] [Google Scholar]
- Guzman FD, Iwamoto Y, Saito N, Salva EP, Dimaano EM, Nishizono A, Suzuki M, Oloko O, Ariyoshi K, Smith C, et al. Clinical, epidemiological, and spatial features of human rabies cases in Metro Manila, the Philippines from 2006 to 2015. PLoS Neglected Tropical Diseases. 2022;16(7):e0010595. doi: 10.1371/journal.pntd.0010595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hadebe JM, Sibiya MN. Experiences of people affected by rabies in Ethekwini district in the province of Kwazulu-Natal, South Africa. African Health Sciences. 2020;20:150–157. doi: 10.4314/ahs.v20i1.20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hampson K, Coudeville L, Lembo T, Sambo M, Kieffer A, Attlan M, Barrat J, Blanton JD, Briggs DJ, Cleaveland S, et al. Estimating the global burden of endemic canine rabies. PLoS Neglected Tropical Diseases. 2015;9:e0003709. doi: 10.1371/journal.pntd.0003709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanlon CA, Kuzmin IV, Blanton JD, Weldon WC, Manangan JS, Rupprecht CE. Efficacy of rabies biologics against new lyssaviruses from Eurasia. Virus Research. 2005;111(1):44–54. doi: 10.1016/j.virusres.2005.03.009. [DOI] [PubMed] [Google Scholar]
- Harischandra PA, Gunesekera A, Janakan N, Gongal G, Abela-Ridder B. Sri Lanka takes action toward a target of zero rabies deaths by 2020. WHO South East Asia J Public Health. 2016;5:113–116. doi: 10.4103/2224-3151.206247. [DOI] [PubMed] [Google Scholar]
- Häsler B, Hiby E, Gilbert W, Obeyesekere N, Bennani H, Rushton J. A one health framework for the evaluation of rabies control programmes: a case study from Colombo City Sri Lanka. PLoS Neglected Tropical Diseases. 2014;8:e3270. doi: 10.1371/journal.pntd.0003270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hatch B, Anderson A, Sambo M, Maziku M, Mchau G, Mbunda E, Mtema Z, Rupprecht CE, Shwiff SA, Nel L. Toward canine rabies elimination in South-Eastern Tanzania: assessment of health economic data. Transboundary and Emerging Diseases. 2017;64:951–958. doi: 10.1111/tbed.12463. [DOI] [PubMed] [Google Scholar]
- Hattwick MA, Weis TT, Stechschulte CJ, Baer GM, Gregg MB. Recovery from rabies. A case report. Annals of Internal Medicine. 1972;76:931–942. doi: 10.7326/0003-4819-76-6-931. [DOI] [PubMed] [Google Scholar]
- Hayman, D.T.S., A.R. Fooks, D.A. Marston, and J.C. Garcia-R. 2016. The global phylogeography of lyssaviruses - challenging the “out of Africa” hypothesis. PLoS Neglected Tropical Diseases. 10 (12): e0005266. 10.1371/journal.pntd.0005266. [DOI] [PMC free article] [PubMed]
- Henson KER, Santiago AAC, Namqui SS. Counterfeit rabies vaccines: the Philippine experience. Open Forum Infectious Diseases. 2020;7(8):ofaa313. doi: 10.1093/ofid/ofaa313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hergert M, le Roux K, Nel LH. Risk factors associated with nonvaccination rabies status of dogs in KwaZulu-Natal, South Africa. Veterinary Medicine. 2016;7:75–83. doi: 10.2147/VMRR.S103859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hill G., 2022. Deaths trigger warning of rabies in South Africa. Washington Times, 22 December. https://www.washingtontimes.com/news/2022/dec/22/deaths-trigger-warning-rabies-south-africa/
- Ho SYW, Lanfear R, Bromham L, Phillips MJ, Soubrier J, Rodrigo AG, Cooper A. Time-dependent rates of molecular evolution. Molecular Ecology. 2011;20(15):3087–3101. doi: 10.1111/j.1365-294X.2011.05178.x. [DOI] [PubMed] [Google Scholar]
- Holzbauer SM, Schrodt CA, Prabhu RM, Asch-Kendrick RJ, Ireland M, Klumb C, Firestone MJ, Liu G, Harry K, Ritter JM, et al. Fatal human rabies infection with suspected host-mediated failure of PostExposure prophylaxis following a recognized zoonotic exposure-Minnesota, 2021. Clinical Infectious Diseases. 2023;29:ciad098. doi: 10.1093/cid/ciad098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horie M, Akashi H, Kawata M, Tomonaga K. Identification of a reptile lyssavirus in Anolis allogus provided novel insights into lyssavirus evolution. Virus Genes. 2021;57(1):40–49. doi: 10.1007/s11262-020-01803-y. [DOI] [PubMed] [Google Scholar]
- Horton DL, McElhinney LM, Marston DA, Wood JL, Russell CA, Lewis N, Kuzmin IV, Fouchier RA, Osterhaus AD, Fooks AR, Smith DJ. Quantifying antigenic relationships among the lyssaviruses. Journal of Virology. 2010;84(22):11841–11848. doi: 10.1128/JVI.01153-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsu WC, Hsu CL, Tu YC, Chang JC, Tsai KR, Lee F, Hu SC., 2019. Standard operating procedure for Lyssavirus surveillance of the bat population in Taiwan. Journal of Visualized Experiments. 27;(150). 10.3791/59421. [DOI] [PubMed]
- Huang, F., M. Ren, J. Pei, H. Mei, B. Sui, Q. Wu, B. Chai, R. Yang, M. Zhou, Z.F. Fu, et al. 2021. Preexposure and postexposure prophylaxis of rabies with adeno-associated virus expressing virus-neutralizing antibody in rodent models. Frontiers in Microbiology. 12: 702273. 10.3389/fmicb.2021.702273. [DOI] [PMC free article] [PubMed]
- Jackson AC. Rabies: a medical perspective. Revue Scientifique et Technique. 2018;37(2):569–580. doi: 10.20506/rst.37.2.2825. [DOI] [PubMed] [Google Scholar]
- Jackson, A.C., M.J. Warrell, C.E. Rupprecht, H.C. Ertl, B. Dietzschold, M. O’Reilly, R.P. Leach, Z.F. Fu, W.H. Wunner, et al. 2003. Management of rabies in humans. Clinical Infectious Diseases 36 (1): 60–63. 10.1086/344905. [DOI] [PubMed]
- Jacquot M, Wallace MA, Streicker DG, Biek R. Geographic range overlap rather than phylogenetic distance explains rabies virus transmission among closely related bat species. Viruses. 2022;14(11):2399. doi: 10.3390/v14112399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeggo M, Roeder P. Lessons from Rinderpest Eradication with reference to COVID-19 pandemic management. EcoHealth. 2021;18:400–402. doi: 10.1007/s10393-021-01552-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jenkin D, Ritchie AJ, Aboagye J, Fedosyuk S, Thorley L, Provstgaad-Morys S, Sanders H, Bellamy D, Makinson R, Xiang ZQ, et al. Safety and immunogenicity of a simian-adenovirus-vectored rabies vaccine: an open-label, nonrandomized, dose-escalation, first-in-human, single-center, phase 1 clinical trial. Lancet Microbe. 2022;3(9):e663–e671. doi: 10.1016/S2666-5247(22)00126-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson N, Aréchiga-Ceballos N, Aguilar-Setien A. Vampire bat rabies: ecology, epidemiology and control. Viruses. 2014;6:1911–1928. doi: 10.3390/v6051911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kamata Y, Tojinbara K, Hampson K, Makita K. The final stages of dog rabies elimination from Japan. Zoonoses and Public Health. 2023;70:1–12. doi: 10.1111/zph.12989. [DOI] [PubMed] [Google Scholar]
- Kanda K, Obayashi Y, Jayasinghe A, Gunawardena GS, Delpitiya NY, Priyadarshani NG, Gamage CD, Arai A, Tamashiro H. Outcomes of a school-based intervention on rabies prevention among school children in rural Sri Lanka. International Health. 2015;7:348–353. doi: 10.1093/inthealth/ihu098. [DOI] [PubMed] [Google Scholar]
- Kanda K, Jayasinghe A, Jayasinghe C, Yoshida T. Public health implication toward rabies elimination in Sri Lanka: A systematic review. Acta Tropica. 2021;223:106080. doi: 10.1016/j.actatropica.2021.106080. [DOI] [PubMed] [Google Scholar]
- Kanda, K., A. Jayasinghe, C. Jayasinghe, and T. Yoshida. 2022. A regional analysis of the progress of current dog-mediated rabies control and prevention. Pathogens. 11: 1130. 10.3390/pathogens11101130. [DOI] [PMC free article] [PubMed]
- Karunanayake D, Matsumoto T, Wimalaratne O, Nanayakkara S, Perera D, Nishizono A, Ahmed K. Twelve years of rabies surveillance in Sri Lanka, 1999–2010. PLoS Neglected Tropical Diseases. 2014;8:e3205. doi: 10.1371/journal.pntd.0003205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kasempimolporn S, Jitapunkul S, Sitprija V. Moving toward the elimination of rabies in Thailand. Journal of the Medical Association of Thailand. 2008;91:433–437. [PubMed] [Google Scholar]
- Kerlin, K. E., 2019. $9 M to preempt zoonotic spillover threats, protect military and local communities. UC Davis. https://www.ucdavis.edu/news/9m-preempt-zoonotic-spillover-threats-protect-military-and-local-communities
- Kimitsuki K, Khan S, Kaimori R, Yahiro T, Saito N, Yamada K, Nakajima N, Komeno T, Furuta Y, Quiambao BP, et al. Implications of the antiviral drug favipiravir on rabies immunoglobulin for postexposure prophylaxis of rabies in mice model with category III-like exposures. Antiviral Research. 2022;209:105489. doi: 10.1016/j.antiviral.2022.105489. [DOI] [PubMed] [Google Scholar]
- Knobel DL, Cleaveland S, Coleman PG, Fèvre EM, Meltzer MI, Miranda ME, Shaw A, Zinsstag J, Meslin FX. Re-evaluating the burden of rabies in Africa and Asia. Bulletin of the World Health Organization. 2005;83:360–368. [PMC free article] [PubMed] [Google Scholar]
- Knobel DL, Jackson AC, Bingham J, Ertl HCJ, Gibson AD, Hughes D, Joubert K, Mani RS, Mohr BJ, Moore SM, et al. A one medicine mission for effective rabies therapy. Frontiers in Veterinary Science. 2022;9:867382. doi: 10.3389/fvets.2022.867382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kogan, L.R., and M. Rishniw. 2022. Canine and feline core vaccinations: US veterinarians’ concerns and perceived impact of COVID-19 antivaccination views on veterinary medicine. Journal of the American Veterinary Medical Association 260: 1482–1488. 10.2460/javma.22.03.0109. [DOI] [PubMed]
- Kularatne SA, Ralapanawa DM, Weerakoon K, Bokalamulla UK, Abagaspitiya N. Pattern of animal bites and post exposure prophylaxis in rabies: A five year study in a tertiary care unit in Sri Lanka. BMC Infectious Diseases. 2016;16:62. doi: 10.1186/s12879-016-1394-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kunkel A, Jeon S, Joseph HC, Dilius P, Crowdis K, Meltzer MI, Wallace R. The urgency of resuming disrupted dog rabies vaccination campaigns: a modeling and cost-effectiveness analysis. Science and Reports. 2021;11:12476. doi: 10.1038/s41598-021-92067-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kunkel A, Minhaj FS, Whitehill F, Austin C, Hahn C, Kieffer AJ, Mendez L, Miller J, Tengelsen LA, Gigante CM, et al. Three human rabies deaths attributed to bat exposures - United States, August 2021. Morbidity and Mortality Weekly Report. 2022;71:31–32. doi: 10.15585/mmwr.mm7101a5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuzmin IV, Mayer AE, Niezgoda M, Markotter W, Agwanda B, Breiman RF, Rupprecht CE. Shimoni bat virus, a new representative of the Lyssavirus genus. Virus Research. 2010;149(2):197–210. doi: 10.1016/j.virusres.2010.01.018. [DOI] [PubMed] [Google Scholar]
- Kuzmin IV, Shi M, Orciari LA, Yager PA, Velasco-Villa A, Kuzmina NA, Streicker DG, Bergman DL, Rupprecht CE. Molecular inferences suggest multiple host shifts of rabies viruses from bats to mesocarnivores in Arizona during 2001–2009. PLoS Pathogens. 2012;8(6):e1002786. doi: 10.1371/journal.ppat.1002786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laorujisawat M, Wattanaburanon A, Abdullakasim P, Maharachpong N. Rabies-related knowledge, attitudes, and practices among primary school students in Chonburi Province Thailand. Inquiry. 2022;59:469580221087881. doi: 10.1177/00469580221087881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Larghi OP. Perspectives for rabies control and eradication from domestic species in developing countries. Developments in Biologicals. 2004;119:205–212. [PubMed] [Google Scholar]
- Lembo T, Partners for Rabies Prevention The blueprint for rabies prevention and control: a novel operational toolkit for rabies elimination. PLoS Neglected Tropical Diseases. 2012;6:e1388. doi: 10.1371/journal.pntd.0001388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lentzos F, Rybicki EP, Engelhard M, Paterson P, Sandholtz WA, Reeves RG. Eroding norms over release of self-spreading viruses. Science. 2022;375:31–33. doi: 10.1126/science.abj5593. [DOI] [PubMed] [Google Scholar]
- Leonardo L, Hernandez L, Magturo TC, Palasi W, Rubite JM, de Cadiz A, Moendeg K, Fornillos RJ, Tabios IK, et al. Current status of neglected tropical diseases (NTDs) in the Philippines. Acta Tropica. 2020;203:105284. doi: 10.1016/j.actatropica.2019.105284. [DOI] [PubMed] [Google Scholar]
- Leopardi, S., E. Barneschi, G. Manna, B. Zecchin, P. Priori, P. Drzewnioková, F. Festa, A. Lombardo, F. Parca, D. Scaravelli, A. Maroni Ponti, and P. De Benedictis. 2021. Spillover of west caucasian bat Lyssavirus (WCBV) in a domestic cat and westward expansion in the Palearctic Region. Viruses 13 (10): 2064. 10.3390/v13102064. [DOI] [PMC free article] [PubMed]
- LeRoux K, Stewart D, Perrett KD, Nel LH, Kessels JA, Abela-Ridder B. Rabies control in KwaZulu-Natal, South Africa. Bulletin of the World Health Organization. 2018;96:360–365. doi: 10.2471/BLT.17.194886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lhendup K, Dorji T. Probable rabies in a child in a Bhutanese town bordering India, 2020. SAGE Open Medical Case Reports. 2021;9:2050313X211019786. doi: 10.1177/2050313X211019786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li AJ, Sreenivasan N, Siddiqi UR, Tahmina S, Penjor K, Sovann L, Gunesekera A, Blanton JD, Knopf L, Hyde TB. Descriptive assessment of rabies postexposure prophylaxis procurement, distribution, monitoring, and reporting in four Asian countries: Bangladesh, Bhutan, Cambodia, and Sri Lanka, 2017–2018. Vaccine. 2019;371 Suppl 1(Suppl 1):A14–A19. doi: 10.1016/j.vaccine.2018.10.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lian M, Hueffer K, Weltzin MM. Interactions between the rabies virus and nicotinic acetylcholine receptors: A potential role in rabies virus induced behavior modifications. Heliyon. 2022;8(9):e10434. doi: 10.1016/j.heliyon.2022.e10434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lojkić, I., I. Šimić, T. Bedeković, and N. Krešić. 2021. Current Status of Rabies and its eradication in Eastern and Southeastern Europe. Pathogens. 10: 742. 10.3390/pathogens10060742. [DOI] [PMC free article] [PubMed]
- Lushasi K, Steenson R, Bernard J, Changalucha JJ, Govella NJ, Haydon DT, Hoffu H, Lankester F, Magoti F, Mpolya EA, et al. One health in practice: using integrated bite case management to increase detection of rabid animals in Tanzania. Frontiers in Public Health. 2020;8:13. doi: 10.3389/fpubh.2020.00013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma X, Bonaparte S, Corbett P, Orciari LA, Gigante CM, Kirby JD, Chipman RB, Fehlner-Gardiner C, Thang C, Cedillo VG, et al. Rabies surveillance in the United States during 2021. Journal of the American Veterinary Medical Association. 2023;8:1–9. doi: 10.2460/javma.23.02.0081. [DOI] [PubMed] [Google Scholar]
- Mähl P, Cliquet F, Guiot AL, Niin E, Fournials E, Saint-Jean N, Aubert M, Rupprecht CE, Gueguen S. Twenty year experience of the oral rabies vaccine SAG2 in wildlife: a global review. Veterinary Research. 2014;45:77. doi: 10.1186/s13567-014-0077-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maki J, Guiot AL, Aubert M, Brochier B, Cliquet F, Hanlon CA, King R, Oertli EH, Rupprecht CE, Schumacher C, et al. Oral vaccination of wildlife using a vaccinia-rabies-glycoprotein recombinant virus vaccine (RABORAL V-RG®): A global review. Veterinary Research. 2017;48(1):57. doi: 10.1186/s13567-017-0459-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malan, A.J., A. Coetzer, C.T. Sabeta, and L.H. Nel. 2022. Epidemiological interface of sylvatic and dog rabies in the North West Province of South Africa. Tropical Medicine and Infectious Disease. 7 (6): 90. 10.3390/tropicalmed7060090. [DOI] [PMC free article] [PubMed]
- Mallewa M, Fooks AR, Banda D, Chikungwa P, Mankhambo L, Molyneux E, Molyneux ME, Solomon T. Rabies encephalitis in malaria-endemic area, Malawi, Africa. Emerging Infectious Diseases. 2007;13:136–139. doi: 10.3201/eid1301.060810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mapatse M, Sabeta C, Fafetine J, Abernethy D. Knowledge, attitudes, practices (KAP) and control of rabies among community households and health practitioners at the human–wildlife interface in Limpopo National Park, Massingir District, Mozambique. PLoS Neglected Tropical Diseases. 2022;16(3):e0010202. doi: 10.1371/journal.pntd.0010202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Markotter W, Coertse J. Bat lyssaviruses. Revue Scientifique et Technique. 2018;37:385–400. doi: 10.20506/rst.37.2.2809. [DOI] [PubMed] [Google Scholar]
- Markotter, W., I. Kuzmin, C.E. Rupprecht, and L.H. Nel. 2008. Phylogeny of Lagos bat virus: Challenges for lyssavirus taxonomy. Virus Research 135 (1): 10–21. 10.1016/j.virusres.2008.02.001. [DOI] [PubMed]
- Marston DA, Horton DL, Nunez J, Ellis RJ, Orton RJ, Johnson N, Banyard AC, McElhinney LM, Freuling CM, Fırat M, et al. Genetic analysis of a rabies virus host shift event reveals within-host viral dynamics in a new host. Virus Evolution. 2017;3:vex038. doi: 10.1093/ve/vex038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marston DA, Banyard AC, McElhinney LM, Freuling CM, Finke S, De Lamballerie X, Müller T, Fooks AR. The lyssavirus host-specificity conundrum - rabies virus - the exception not the rule. Current Opinion in Virology. 2018;28:68–73. doi: 10.1016/j.coviro.2017.11.007. [DOI] [PubMed] [Google Scholar]
- Matibag GC, Ohbayashi Y, Kanda K, Yamashina H, Kumara WR, Perera IN, De Silva DD, De S Gunawardena GS, Jayasinghe A, et al. A pilot study on the usefulness of information and education campaign materials in enhancing the knowledge, attitude and practice on rabies in rural Sri Lanka. Journal of Infection in Developing Countries. 2009;3:55–64. doi: 10.3855/jidc.106. [DOI] [PubMed] [Google Scholar]
- Matsumoto T, Ahmed K, Wimalaratne O, Nanayakkara S, Perera D, Karunanayake D, Nishizono A. Novel sylvatic rabies virus variant in endangered golden palm civet, Sri Lanka. Emerging Infectious Diseases. 2011;17:2346–2349. doi: 10.3201/eid1712.110811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsumoto T, Ahmed K, Karunanayake D, Wimalaratne O, Nanayakkara S, Perera D, Kobayashi Y, Nishizono A. Molecular epidemiology of human rabies viruses in Sri Lanka. Infection, Genetics and Evolution. 2013;18:160–7. doi: 10.1016/j.meegid.2013.05.018. [DOI] [PubMed] [Google Scholar]
- Matulis GA, Altantogtokh D, Lantos PM, Jones JH, Wofford RN, Janko M, Tsogbadrakh N, Bayar T, Ganzorig S, Boldbaatar B, et al. Hotspots in cold land-reported cases of rabies in wildlife and livestock in Mongolia from 2012–2018. Zoonoses and Public Health. 2022;69(6):655–662. doi: 10.1111/zph.12954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mbaipago N, Madjadinan A, Amalaman DM, Andrée Ndour P, Zinsstag J, Heitz-Tokpa K, Lechenne M. General insights on obstacles to dog vaccination in Chad on community and institutional level. Frontiers in Veterinary Science. 2022;9:866755. doi: 10.3389/fvets.2022.866755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mbilo C, Coetzer A, Bonfoh B, Angot A, Bebay C, Cassama B, De Benedictis P, Ebou MH, Gnanvi C, Kallo V, et al. Dog rabies control in West and Central Africa: A review. Acta Tropica. 2021;224:105459. doi: 10.1016/j.actatropica.2020.105459. [DOI] [PubMed] [Google Scholar]
- Medical College of Wisconsin. 2022. Rabies registry website. https://www.mcw.edu/departments/pediatrics/divisions/infectiousdiseases/rabies-registry-website
- Medina DJO, Jayme SI, Amparo ACB, Cresencio RO, Lopez EL, Baquilod MS, Hernandez LM, Villalon EES, 3rd, Nel LD. World rabies day campaign in the Philippines. Tropical Diseases, Travel Medicine and Vaccines. 2016;2:22. doi: 10.1186/s40794-016-0036-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Membrebe JV, Suchard MA, Rambaut A, Baele G, Lemey P. Bayesian inference of evolutionary histories under time-dependent substitution rates. Molecular Biology and Evolution. 2019;36(8):1793–1803. doi: 10.1093/molbev/msz094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Merritt, T., K. Taylor, K. Cox-Witton, H. Field, K. Wingett, D. Mendez, M. Power, and D. Durrheim. 2018. Australian bat lyssavirus. Australian Journal of General Practice. 47: 93–96. 10.31128/AFP-08-17-4314. [DOI] [PubMed]
- Messinger Cayetano, S., L. Crandall., 2020. The paradox of success and public perspective: COVID-19 and the perennial problem of prevention. Journal of Epidemiology and Community Health. jech-2020–214518. 10.1136/jech-2020-214518. [DOI] [PMC free article] [PubMed]
- Mindekem, R., M.S. Lechenne, K.S. Naissengar, A. Oussiguéré, B. Kebkiba, D.D. Moto, I.O. Alfaroukh, L.T. Ouedraogo, S. Salifou, and J. Zinsstag. 2017. Cost description and comparative cost efficiency of post exposure prophylaxis and canine mass vaccination against rabies in N’Djamena, Chad. Frontiers in Veterinary Science. 4: 38. 10.3389/fvets.2017.00038. [DOI] [PMC free article] [PubMed]
- Minhaj FS, Bonaparte SC, Boutelle C, Wallace RM. Analysis of available animal testing data to propose peer-derived quantitative thresholds for determining adequate surveillance capacity for rabies. Science and Reports. 2023;13:3986. doi: 10.1038/s41598-023-30984-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miranda LM, Miranda ME, Hatch B, Deray R, Shwiff S, Roces MC, Rupprecht CE. Toward canine rabies elimination in Cebu, Philippines: assessment of health economic data. Transboundary and Emerging Diseases. 2017;64:121–129. doi: 10.1111/tbed.12350. [DOI] [PubMed] [Google Scholar]
- Mission Rabies (MR). 2023. https://missionrabies.com/
- Mogano K, Suzuki T, Mohale D, Phahladira B, Ngoepe E, Kamata Y, Chirima G, Sabeta C, Makita K. Spatiotemporal epidemiology of animal and human rabies in northern South Africa between 1998 and 2017. PLoS Neglected Tropical Diseases. 2022;16:e0010464. doi: 10.1371/journal.pntd.0010464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morgan CN, Wallace RM, Vokaty A, Seetahal JFR, Nakazawa YJ. Risk modeling of bat rabies in the Caribbean Islands. Tropical Medicine and Infectious Disease. 2020;5(1):35. doi: 10.3390/tropicalmed5010035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mpolya EA, Lembo T, Lushasi K, Mancy R, Mbunda EM, Makungu S, Maziku M, Sikana L, Jaswant G, Townsend S, et al. Toward elimination of dog-mediated human rabies: experiences from implementing a large-scale demonstration project in Southern Tanzania. Frontiers in Veterinary Science. 2017;4:21. doi: 10.3389/fvets.2017.00021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mrzljak A, Novak R, Pandak N, Tabain I, Franusic L, Barbic L, Bogdanic M, Savic V, Mikulic D, Pavicic-Saric J, et al. Emerging and neglected zoonoses in transplant population. World Journal of Transplantation. 2020;10(3):47–63. doi: 10.5500/wjt.v10.i3.47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mshelbwala PP, Weese JS. Rabies in the developing world: challenges & prospects. Clinicians Brief. 2017;15:16–18. [Google Scholar]
- Mshelbwala PP, Weese JS, Sanni-Adeniyi OA, Chakma S, Okeme SS, Mamun AA, Rupprecht CE, Magalhaes RJS. Rabies epidemiology, prevention and control in Nigeria: Scoping progress toward elimination. PLoS Neglected Tropical Diseases. 2021;15:e0009617. doi: 10.1371/journal.pntd.0009617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mshelbwala, P.P., J.S. Weese, N.J. Clark, I. Tekki, S. Chakma, D. Shamaki, A.A. Mamun, C.E. Rupprecht, and R.J. Soares Magalhães. 2022. Spatiotemporal heterogeneity and determinants of canine rabies evidence at local government area level in Nigeria: Implications for rabies prevention and control. One Health. 14: 100378. 10.1016/j.onehlt.2022.100378. [DOI] [PMC free article] [PubMed]
- Mubashir A, Hussain SA. Is Pakistan doing enough to eradicate rabies by 2030? Journal of the College of Physicians and Surgeons–Pakistan. 2021;30:614. doi: 10.29271/jcpsp.2021.05.614. [DOI] [PubMed] [Google Scholar]
- Mudiyanselage MH, Weerasinghe NP, Pathirana K, Dias H. Misdiagnosis of cerebral malaria initially as acute psychotic disorder and later as human rabies: A case report. BMC Research Notes. 2016;9(1):400. doi: 10.1186/s13104-016-2211-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Müller T, Dietzschold B, Ertl H, Fooks AR, Freuling C, Fehlner-Gardiner C, Kliemt J, Meslin FX, Franka R, Rupprecht CE, et al. Development of a mouse monoclonal antibody cocktail for postexposure rabies prophylaxis in humans. PLoS Neglected Tropical Diseases. 2009;3:e542. doi: 10.1371/journal.pntd.0000542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nadal D, Beeching S, Cleaveland S, Cronin K, Hampson K, Steenson R, Abela-Ridder B. Rabies and the pandemic: lessons for one health. Transactions of the Royal Society of Tropical Medicine and Hygiene. 2022;116(3):197–200. doi: 10.1093/trstmh/trab123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nadal D, Hampson K, Lembo T, Rodrigues R, Vanak AT, Cleaveland S. Where rabies is not a disease. Bridging healthworlds to improve mutual understanding and prevention of rabies. Frontiers in Veterinary Science. 2022;9:867266. doi: 10.3389/fvets.2022.867266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- NASPHV. 2016. Compendium of animal rabies prevention and control. Journal of the American Veterinary Medical Association 248: 505–517. 10.2460/javma.248.5.505. [DOI] [PubMed]
- Nel LH, Rupprecht CE. Emergence of lyssaviruses in the Old World: The case of Africa. Current Topics in Microbiology and Immunology. 2007;315:161–193. doi: 10.1007/978-3-540-70962-6_8. [DOI] [PubMed] [Google Scholar]
- Ness A, Aiken J, McKenzie D. Sheep scrapie and deer rabies in England prior to 1800. Prion. 2023;17:7–15. doi: 10.1080/19336896.2023.2166749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- NewsDesk., 2022. Rabies in Mexico: Human cases reported in Oaxaca and Nayarit. Outbreak News Today. December 31, 2022. https://outbreaknewstoday.com/rabies-in-mexico-human-cases-reported-in-oaxaca-and-nayarit-85193/
- Ngoepe E, Chirima JG, Mohale D, Mogano K, Suzuki T, Makita K, Sabeta CT. Rabies outbreak in black-backed jackals (Canis mesomelas), South Africa, 2016. Epidemiology and Infection. 2022;150:e137. doi: 10.1017/S0950268821002685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nguyen HTT, Afriyie DO, Tran CH, Dang AD, Tran DN, Dang TQ, Otsu S, Urabe MI, Nguyen HT, Pham TN, et al. Progress toward rabies control and elimination in Vietnam. Revue Scientifique et Technique. 2019;38:199–212. doi: 10.20506/rst.38.1.2953. [DOI] [PubMed] [Google Scholar]
- Nuismer SL, Bull JJ. Self-disseminating vaccines to suppress zoonoses. Nature Ecology Evolution. 2020 doi: 10.1038/s41559-020-1254-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nuismer SL, Althouse BM, May R, Bull JJ, Stromberg SP, Antia R. Eradicating infectious disease using weakly transmissible vaccines. Proceedings of the Royal Society B. 2016;283:20161903. doi: 10.1098/rspb.2016.1903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nuismer SL, May R, Basinski A, Remien CH. Controlling epidemics with transmissible vaccines. PLoS One. 2018;13:e0196978. doi: 10.1371/journal.pone.0196978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nuismer SL, Basinski A, Bull JJ. Evolution and containment of transmissible recombinant vector vaccines. Evolutionary Applications. 2019 doi: 10.1111/eva.12806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Obonyo M, Arvelo W, Kadivane S, Orundu M, Lankau E, Gakuya F, Munyua P, Githinji J, Marano N, Njenga K, et al. Investigation to determine staff exposure and describe animal bite surveillance after detection of a rabid zebra in a safari lodge in Kenya, 2011. The Pan African Medical Journal. 2014;19:10. doi: 10.11604/pamj.2014.19.10.3434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ørpetveit I, Reiten MR, Benestad SL, Ropstad E, Strandbygaard B, Madslien K, Stokke E, Ytrehus B. Rabies in Arctic fox (Vulpes lagopus) and reindeer (Rangifer tarandus platyrhynchus) during an outbreak on Svalbard, Norway, 2011–12. Journal of Wildlife Diseases. 2022;58(3):550–561. doi: 10.7589/JWD-D-21-00112. [DOI] [PubMed] [Google Scholar]
- Paarmann E. Ein Beitrag zur Lyssa der Vögel. Zeitschrift Für Hygiene Und Infektionskrankheiten. 1955;141:103–109. doi: 10.1007/bf02162126. [DOI] [PubMed] [Google Scholar]
- Pastoret PP, Van Gucht S, Brochier B. Eradicating rabies at source. Revue Scientifique Et Technique. 2014;33(509–19):497–508. doi: 10.20506/rst.33.2.2250. [DOI] [PubMed] [Google Scholar]
- Perera MA, Harischandra PA, Wimalaratne O, Damboragama SN. Feasibility of canine oral rabies vaccination in Sri Lanka–a preliminary report. Ceylon Medical Journal. 2000;45:61–64. doi: 10.4038/cmj.v45i2.8002. [DOI] [PubMed] [Google Scholar]
- Perera IN, Pinidiyapathirage MJ, Wickremasinghe AR. Rabies vaccination of domesticated dogs in the Central Province of Sri Lanka. Ceylon Medical Journal. 2007;52:69–70. doi: 10.4038/cmj.v52i2.1031. [DOI] [PubMed] [Google Scholar]
- Pham-Thanh L, Nhu TV, Nguyen TV, Tran KV, Nguyen KC, Nguyen HT, Ngo Thi H, Padungtod P. Zoonotic pathogens and diseases detected in Vietnam, 2020–2021. One Health. 2022;14:100398. doi: 10.1016/j.onehlt.2022.100398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phoolcharoen W, Banyard AC, Prehaud C, Selden D, Wu G, Birch CPD, Szeto TH, Lafon M, Fooks AR, Ma JK. In vitro and in vivo evaluation of a single chain antibody fragment generated in planta with potent rabies neutralization activity. Vaccine. 2019;37:4673–4680. doi: 10.1016/j.vaccine.2018.02.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pichon, S., A. Moureau, C. Petit, L. Chu, B. Essink, D. Muse, J. Saleh, F. Guinet-Morlot, and A.M. Minutello. 2022. Safety and immunogenicity of a serum-free purified Vero rabies vaccine in healthy adults: A randomized phase II preexposure prophylaxis study. Vaccine 28: S0264-410X(22)00806–4. 10.1016/j.vaccine.2022.06.040. [DOI] [PubMed]
- Pieracci EG, Williams CE, Wallace RM, Kalapura CR, Brown CM. U.S. dog importations during the COVID-19 pandemic: Do we have an erupting problem? PLoS One. 2021;16:e0254287. doi: 10.1371/journal.pone.0254287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poleshchuk, E.M., D.N. Tagakova, G.N. Sidorov, T.S. Orlova, N.S. Gordeiko, and A.Z. Kaisarov. 2023. Lethal cases of lyssavirus encephalitis in humans after contact with bats in the Russian Far East in 2019–2021. Voprosy Virusologii. 68: 45–58. 10.36233/0507-4088-156. [DOI] [PubMed]
- PREEMPT., 2018. Big win new countermeasures to eliminate pandemic risk. https://static1.squarespace.com/static/5d927ef93f54836cb17542c1/t/5e74072e6f3a42255b9573fe/1584662333204/BIG+WIN-New+Countermeasures.pdf
- PREEMPT., 2022. About-PREEMPT. https://www.preemptproject.org/about
- Qi L, Su K, Shen T, Tang W, Xiao B, Long J, Zhao H, Chen X, Xia Y, Xiong Y, et al. Epidemiological characteristics and postexposure prophylaxis of human rabies in Chongqing, China, 2007–2016. BMC Infectious Diseases. 2018;18:6. doi: 10.1186/s12879-017-2830-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quiambao B, Varghese L, Demarteau N, Sengson RF, Javier J, Mukherjee P, Manio LC, Preiss S. Health economic assessment of a rabies preexposure prophylaxis program compared with postexposure prophylaxis alone in high-risk age groups in the Philippines. International Journal of Infectious Diseases. 2020;97:38–46. doi: 10.1016/j.ijid.2020.05.062. [DOI] [PubMed] [Google Scholar]
- Quiambao B, Montalban C, Minutello AM, Guinet-Morlot F, Moureau A, Petit C, Pichon S. Serum-free purified Vero rabies vaccine is safe and immunogenic in children: Results of a randomized phase II preexposure prophylaxis regimen study. Vaccine. 2022;40(35):5170–5178. doi: 10.1016/j.vaccine.2022.06.061. [DOI] [PubMed] [Google Scholar]
- Rana MS, Siddiqi UR, Ghosh S, Jahan AA, Islam MK, Ali Shah MR, Ullah SM, Ali SME, Ahmed BN, Shamsuzzaman AKM. Epidemiological study of human rabies cases in Bangladesh through verbal autopsy. Heliyon. 2020;6(11):e05521. doi: 10.1016/j.heliyon.2020.e05521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rao, A.K., D. Briggs, S.M. Moore, F. Whitehill, D. Campos-Outcalt, R.L. Morgan, R.M. Wallace, J.R. Romero, L. Bahta, et al. 2022. Use of a modified preexposure prophylaxis vaccination schedule to prevent human rabies: Recommendations of the advisory vommittee on Immunization Practices - United States, 2022. Morbidity and Mortality Weekly Report. 71: 619–627. 10.15585/mmwr.mm7118a2. [DOI] [PMC free article] [PubMed]
- Ravish, H.S., A. Khobragade, D. Satapathy, M. Gupta, S. Kumar, V. Bhomia, and VR, Desai M, Agrawal AD.,. 2021. Safety and Immunogenicity of a novel three-dose recombinant nanoparticle rabies G protein vaccine administered as simulated post exposure immunization: A randomized, comparator controlled, multicenter, phase III clinical study. Human Vaccines & Immunotherapeutics. 17: 4239–4245. 10.1080/21645515.2021.1957413. [DOI] [PMC free article] [PubMed]
- Ravensberg MF, Fanoy EB, Whelan J, Embregts CW, GeurtsvanKessel CH, Strydom JB., 2022. Ongoing rabies outbreak in dogs of unprecedented scale and human cases in Nelson Mandela Bay Municipality, South Africa, up to 13 February 2022. Euro Surveillance. 27(16). 10.2807/1560-7917.ES.2022.27.16.2200252. [DOI] [PMC free article] [PubMed]
- Raynor B, Díaz EW, Shinnick J, Zegarra E, Monroy Y, Mena C, De la Puente-León M, Levy MZ, Castillo-Neyra R. The impact of the COVID-19 pandemic on rabies reemergence in Latin America: The case of Arequipa, Peru. PLoS Negl Trop Dis. 2021;15(5):e0009414. doi: 10.1371/journal.pntd.0009414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reece J. Working together within the profession to eradicate rabies. The Veterinary Record. 2019;185:513–514. doi: 10.1136/vr.l6126. [DOI] [PubMed] [Google Scholar]
- Reeves R. G. 2022. Self-spreading viruses information. http://web.evolbio.mpg.de/HEVIMAs/
- Regulatory News Service., 2022. The vaccine group completes lassa fever project. https://www.londonstockexchange.com/news-article/FIPP/the-vaccine-group-completes-lassa-fever-project/about:blank
- Resolve To Save Lives (RTSL)., 2022. Epidemics that didn’t happen: rabies in Tanzania. https://preventepidemics.org/epidemics-that-didnt-happen/rabies/
- Rocke TE, Kingstad-Bakke B, Wüthrich M, Stading B, Abbott RC, Isidoro-Ayza M, Dobson HE, dos Santos Dias L, Galles K, Lankton JS, et al. Virally vectored vaccine candidates against white-nose syndrome induce an anti-fungal immune response in little brown bats (Myotis lucifugus) Science and Reports. 2019;9:6788. doi: 10.1038/s41598-019-43210-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rohde RE, Rupprecht CE. Update on lyssaviruses and rabies: will past progress play as prologue in the near term toward future elimination? Faculty Reviews. 2020;9:9. doi: 10.12703/b/9-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosatte, R.C. 1988. Rabies in Canada history, epidemiology and control. Canadian Veterinary Journal 29: 362–365. [PMC free article] [PubMed]
- Ross DW. Infectious vaccines. Archives of Pathology and Laboratory Medicine. 1998;122(1):100–101. [PubMed] [Google Scholar]
- Royal A, John D, Bharti O, Tanwar R, Bhagat DK, Padmawati RS, Chaudhary V, Umapathi R, Bhadola P, Utarini A. A cost-effectiveness analysis of pre-exposure prophylaxis to avert rabies deaths in school-aged children in India. Vaccines (basel). 2022;11:88. doi: 10.3390/vaccines11010088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rupprecht CE, Salahuddin N. Current status of human rabies prevention: remaining barriers to global biologics accessibility and disease elimination. Expert Review of Vaccines. 2019;18:629–640. doi: 10.1080/14760584.2019.1627205. [DOI] [PubMed] [Google Scholar]
- Rupprecht C, Kuzmin I, Meslin F. Lyssaviruses and rabies: current conundrums, concerns, contradictions and controversies. F1000 Research. 2017;6:184. doi: 10.12688/f1000research.10416.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rupprecht CE, Bannazadeh Baghi H, Del Rio Vilas VJ, Gibson AD, Lohr F, Meslin FX, Seetahal JFR, Shervell K, Gamble L. Historical, current and expected future occurrence of rabies in enzootic regions. Revue Scientifique et Technique. 2018;37:729–739. doi: 10.20506/rst.37.2.2836. [DOI] [PubMed] [Google Scholar]
- Rupprecht, C.E., C.M. Freuling, R.S. Mani, C. Palacios, C.T. Sabeta, and M. Ward. 2020. A history of rabies – the foundation for global canine rabies elimination. In Rabies, 4th ed., ed. A.R. Fooks and A.C. Jackson, 1–42. San Diego: Acad Press. 10.1016/b978-0-12-818705-0.00001-7.
- Rupprecht CE, Mani RS, Mshelbwala PP, Recuenco SE, Ward MP. Rabies in the tropics. Current Tropical Medicine Reports. 2022;9:28–39. doi: 10.1007/s40475-022-00257-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saleem SM, Quansar R, Haq I, Salim Khan SM. Imposing COVID-19 lockdown and reported dog bite cases: An experience from a tertiary antirabies center of North India. Journal of Veterinary Behavior. 2021;42:48–52. doi: 10.1016/j.jveb.2021.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sánchez-Soriano, C., A.D. Gibson, L. Gamble, J.L. Burdon Bailey, S. Green, M. Green, B.M.D. Bronsvoort, I.G. Handel, R.J. Mellanby, and S. Mazeri. 2019. Development of a high number, high coverage dog rabies vaccination programme in Srilanka. BMC Infectious Diseases 19: 977. 10.1186/s12879-019-4585-z. [DOI] [PMC free article] [PubMed]
- Sartore S, Mulatti P, Trestini S, Lorenzetto M, Gagliazzo L, Marangon S, Bonfanti L. The economic implications of sylvatic rabies eradication in Italy. Zoonoses and Public Health. 2018;65:147–157. doi: 10.1111/zph.12383. [DOI] [PubMed] [Google Scholar]
- Sasaki DM, Middleton CR, Sawa TR, Christensen CC, Kobayashi GY. Rabid bat diagnosed in Hawaii. Hawaii Medical Journal. 1992;51(7):181–185. [PubMed] [Google Scholar]
- Scott TP, Coetzer A, de Balogh K, Wright N, Nel LH. The Pan-African Rabies Control Network (PARACON): A unified approach to eliminating canine rabies in Africa. Antiviral Research. 2015;124:93–100. doi: 10.1016/j.antiviral.2015.10.002. [DOI] [PubMed] [Google Scholar]
- Scudellari, M., 2016. Journal Club: Can transmissible vaccines have a major role in eradicating disease? | National Academy of Sciences. https://blog.pnas.org/2016/11/journal-club-can-transmissible-vaccines-have-a-major-role-in-eradicating-disease/
- Shafaati M, Akbarpour S, Priyanka Saied AA, Choudhary OP. Tackling rabies by one health approach: Pitfalls on the road to global eradication. New Microbes and New Infections. 2023;52:101098. doi: 10.1016/j.nmni.2023.101098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shope RE. Rabies-related viruses. The Yale Journal of Biology and Medicine. 1982;55:271–275. [PMC free article] [PubMed] [Google Scholar]
- Shulpin MI, Nazarov NA, Chupin SA, Korennoy FI, Metlin AY, Mischenko AV. Rabies surveillance in the Russian Federation. Revue Scientifique et Technique. 2018;37:483–495. doi: 10.20506/rst.37.2.2817. [DOI] [PubMed] [Google Scholar]
- Shwiff SA, Kirkpatrick KN, Sterner RT. Economic evaluation of an oral rabies vaccination program for control of a domestic dog-coyote rabies epizootic: 1995–2006. Journal of the American Veterinary Medical Association. 2008;233(11):1736–1741. doi: 10.2460/javma.233.11.1736. [DOI] [PubMed] [Google Scholar]
- Shwiff SA, Hatch B, Anderson A, Nel LH, Leroux K, Stewart D, de Scally M, Govender P, Rupprecht CE. Toward canine rabies elimination in KwaZulu-Natal, South Africa: assessment of health economic data. Transboundary and Emerging Diseases. 2016;63:408–415. doi: 10.1111/tbed.12283. [DOI] [PubMed] [Google Scholar]
- Siddiqui, A., A. Ahmed, M. Tanveer, and A. Arshad. 2021. The crux of Pakistan’s prolonged rabies vaccine shortage: A rising mortal threat in the COVID-19 pandemic. Journal of Medical Virology 93 (9): 5221–5222. 10.1002/jmv.27024. [DOI] [PMC free article] [PubMed]
- Sidwa TJ, Wilson PJ, Moore GM, Oertli EH, Hicks BN, Rohde RE, Johnston DH. Evaluation of oral rabies vaccination programs for control of rabies epizootics in coyotes and gray foxes: 1995–2003. Journal of the American Veterinary Medical Association. 2005;227:785–792. doi: 10.2460/javma.2005.227.785. [DOI] [PubMed] [Google Scholar]
- South Asian Association for Regional Cooperation (SAARC)., 2019. 10.1093/law:epil/9780199231690/e678. https://www.saarc-sec.org/index.php/about-saarc/about-saarc
- Sparkes J, Fleming PJ, Ballard G, Scott-Orr H, Durr S, Ward MP. Canine rabies in Australia: A review of preparedness and research needs. Zoonoses and Public Health. 2015;62:237–253. doi: 10.1111/zph.12142. [DOI] [PubMed] [Google Scholar]
- Sparrow E, Torvaldsen S, Newall AT, Wood JG, Sheikh M, Kieny MP, Abela-Ridder B. Recent advances in the development of monoclonal antibodies for rabies post exposure prophylaxis: A review of the current status of the clinical development pipeline. Vaccine. 2019;37 Suppl 1:A132–A139. doi: 10.1016/j.vaccine.2018.11.004. [DOI] [PubMed] [Google Scholar]
- Srinivasan A, Burton EC, Kuehnert MJ, Rupprecht C, Sutker WL, Ksiazek TG, Paddock CD, Guarner J, Shieh WJ, Goldsmith C, et al. Transmission of rabies virus from an organ donor to four transplant recipients. New England Journal of Medicine. 2005;352(11):1103–1111. doi: 10.1056/NEJMoa043018. [DOI] [PubMed] [Google Scholar]
- Stokes W, McFarland R, Kulpa-Eddy J, Gatewood D, Levis R, Halder M, Pulle G, Kojima H, Casey W, Gaydamaka A, et al. Report on the international workshop on alternative methods for human and veterinary rabies vaccine testing: State of the science and planning the way forward. Biologicals. 2012;40(5):369–381. doi: 10.1016/j.biologicals.2012.07.005. [DOI] [PubMed] [Google Scholar]
- Streicker DG, Turmelle AS, Vonhof MJ, Kuzmin IV, McCracken GF, Rupprecht CE. Host phylogeny constrains cross-species emergence and establishment of rabies virus in bats. Science. 2010;329(5992):676–679. doi: 10.1126/science.1188836. [DOI] [PubMed] [Google Scholar]
- Streicker DG, Bull JJ, Nuismer SL. Self-spreading vaccines: Base policy on evidence. Science. 2022;375:1362–1363. doi: 10.1126/science.abo0238. [DOI] [PubMed] [Google Scholar]
- Subedi D, Chandran D, Subedi S, Acharya KP. Ecological and socioeconomic factors in the occurrence of rabies: a forgotten scenario. Infectious Disease Reports. 2022;14(6):979–986. doi: 10.3390/idr14060097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swedberg, C., S. Mazeri, R.J. Mellanby, K. Hampson, and N.R. Chng. 2022. Implementing a One Health approach to rabies surveillance: Lessons from integrated bite case management. Frontiers in Tropical Diseases. 3: 829132. 10.3389/fitd.2022.829132. [DOI] [PMC free article] [PubMed]
- Tantawichien T, Rupprecht CE. Modern biologics for rabies prophylaxis and the elimination of human cases mediated by dogs. Expert Opinion on Biological Therapy. 2020;20:1347–1359. doi: 10.1080/14712598.2020.1766021. [DOI] [PubMed] [Google Scholar]
- Tanzania., 2019. The United Republic of Tanzania, National Rabies Control Strategy, July. https://static1.squarespace.com/static/60ae10230c660f2b45453582/t/61237d549a97d543daf6728b/1629715811493/National+Rabies+Control+Strategy+2019.pdf
- Tarantola A. Four thousand years of concepts relating to rabies in animals and humans: its prevention and cure. Tropical Medicine and Infectious Disease. 2017;2:5. doi: 10.3390/tropicalmed2020005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor LH, Wallace RM, Balaram D, Lindenmayer JM, Eckery DC, Mutonono-Watkiss B, Parravani E, Nel LH. The role of dog population management in rabies elimination-a review of current approaches and future opportunities. Frontiers in Veterinary Science. 2017;4:109. doi: 10.3389/fvets.2017.00109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor E, Aguilar-Ancori EG, Banyard AC, Abel I, Mantini-Briggs C, Briggs CL, Carrillo C, Gavidia CM, Castillo-Neyra R, Parola AD, et al. The Amazonian Tropical Bites Research Initiative, a hope for resolving zoonotic neglected tropical diseases in the One Health era. International Health. 2022;27:ihac048. doi: 10.1093/inthealth/ihac048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Technology Networks., 2022. Transmissible and Transferable Vaccines. Vaccines from Technology Networks. http://www.technologynetworks.com/vaccines/articles/transmissible-and-transferable-vaccines-361204
- Tenzin, Ward MP. Review of rabies epidemiology and control in South, South East and East Asia: Past, present and prospects for elimination. Zoonoses and Public Health. 2012;59:451–467. doi: 10.1111/j.1863-2378.2012.01489.x. [DOI] [PubMed] [Google Scholar]
- Terré J, Chappuis G, Lombard M, Desmettre P. Eradication of rabies, using a rec-DNA vaccine. Journal of Biotechnology. 1996;46:156–157. doi: 10.1016/0168-1656(96)80467-6. [DOI] [PubMed] [Google Scholar]
- Thiagarajan K. Why did rabies control fail in India? BMJ. 2022;378:2284. doi: 10.1136/bmj.o2284. [DOI] [PubMed] [Google Scholar]
- Thomson GR, Penrith ML. Eradication of transboundary animal diseases: can the rinderpest success story be repeated? Transboundary and Emerging Diseases. 2017;64:459–475. doi: 10.1111/tbed.12385. [DOI] [PubMed] [Google Scholar]
- Thumbi SM, Blumberg L, le Roux K, Salahuddin N, Abela B. A call to accelerate an end to human rabies deaths. Lancet. 2022;400:2261–2264. doi: 10.1016/S0140-6736(22)02487-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tidman R, Thumbi SM, Wallace R, de Balogh K, Iwar V, Dieuzy-Labaye I, Song J, Shadomy S, Qiu Y, Torres G, et al. United against rabies forum: the one health concept at work. Frontiers in Public Health. 2022;10:854419. doi: 10.3389/fpubh.2022.854419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torres JM, Sánchez C, Ramírez MA, Morales M, Bárcena J, Ferrer J, Espuña E, Pagès-Manté A, Sánchez-Vizcaíno JM. First field trial of a transmissible recombinant vaccine against myxomatosis and rabbit hemorrhagic disease. Vaccine. 2001;19:4536–4543. doi: 10.1016/S0264-410X(01)00184-0. [DOI] [PubMed] [Google Scholar]
- Transmissiblevaccines.org., 2022. Workshop: Development of self-disseminating vaccines – transmissible vaccines. https://transmissiblevaccines.org/workshop-dev-vaccines/
- Tsuda, Y., P. Caposio, C.J. Parkins, S. Botto, I. Messaoudi, L. Cicin-Sain, H. Feldmann, and M.A. Jarvis. 2011. A replicating cytomegalovirus-based vaccine encoding a dingle Ebola virus nucleoprotein CTL epitope confers protection against Ebola Virus. PLoS Neglected Tropical Diseases. 5: e1275. 10.1371/journal.pntd.0001275. [DOI] [PMC free article] [PubMed]
- TVG., 2021. Animal vaccines to address zoonotic and livestock diseases. https://thevaccinegroup.com/pipeline/
- Umeno S, Doi Y. A study of the anti-rabic inoculation of dogs and the results of its practical application. The Kitasato Archives of Experimental Medicine. 1921;4:89–108. [Google Scholar]
- USDA-APHIS., 2019. Availability of an environmental assessment for field testing of a Pseudogymnoascus destructans Vaccine, Live Raccoon Poxvirus Vector (RCN-CAL/SP). https://www.regulations.gov/docket/APHIS-2019-0043/document
- Valenzuela, L.M., S.I. Jayme, A.C.B. Amparo, L.H. Taylor, M.P.Z. Dela Cruz, D.A. Licuan, R. Gamal-Bitao, and L.H. Nel. 2017. The Ilocos Norte Communities against rabies exposure elimination project in the Philippines: Epidemiological and economic aspects. Frontiers in Veterinary Science. 4: 54. 10.3389/fvets.2017.00054. [DOI] [PMC free article] [PubMed]
- van Thiel PP, de Bie RM, Eftimov F, Tepaske R, Zaaijer HL, van Doornum GJ, Schutten M, Osterhaus AD, Majoie CB, Aronica E, et al. Fatal human rabies due to Duvenhage virus from a bat in Kenya: failure of treatment with coma-induction, ketamine, and antiviral drugs. PLoS Neglected Tropical Diseases. 2009;3:e428. doi: 10.1371/journal.pntd.0000428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vega, S., L. Lorenzo-Rebenaque, C. Marin, R. Domingo, and F. Fariñas. 2021. Tackling the threat of rabies reintroduction in Europe. Frontiers in Veterinary Science. 7: 613712. 10.3389/fvets.2020.613712. [DOI] [PMC free article] [PubMed]
- Vos A, Neubert A, Aylan O, Schuster P, Pommerening E, Müller T, Chivatsi DC. An update on safety studies of SAD B19 rabies virus vaccine in target and nontarget species. Epidemiology and Infection. 1999;123:165–175. doi: 10.1017/s0950268899002666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walker, P.J., K.R. Blasdell, C.H. Calisher, R.G. Dietzgen, H. Kondo, G. Kurath, B. Longdon, D.M. Stone, R.B. Tesh, N. Tordo, et al. 2018. ICTV Report Consortium. ICTV virus taxonomy profile: Rhabdoviridae. The Journal of General Virology. 99 (4): 447–448. 10.1099/jgv.0.001020. [DOI] [PubMed]
- Wallace RM, Gilbert A, Slate D, Chipman R, Singh A, Wedd Cassie, Blanton JD. Right place, wrong species: a 20-year review of rabies virus cross species transmission among terrestrial mammals in the United States. PLoS One. 2014;9(10):e107539. doi: 10.1371/journal.pone.0107539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wallace RM, Cliquet F, Fehlner-Gardiner C, Fooks AR, Sabeta CT, Setien AA, Tu C, Vuta V, Yakobson B, Yang DK, et al. Role of oral rabies vaccines in the elimination of dog-mediated human rabies deaths. Emerging Infectious Diseases. 2020;26:1–9. doi: 10.3201/eid2612.201266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warrell MJ. Time to revise the strategy for Gavi funding of rabies vaccine? Lancet. 2021;397(10273):471–472. doi: 10.1016/S0140-6736(21)00220-8. [DOI] [PubMed] [Google Scholar]
- Warrell, M., D.A. Warrell, and A. Tarantola. 2017. The imperative of palliation in the management of rabies encephalomyelitis. Tropical Medicine and Infectious Disease. 2 (4): 52. 10.3390/tropicalmed2040052. [DOI] [PMC free article] [PubMed]
- Wertheim JO, Kosakovsky Pond SL. Purifying selection can obscure the ancient age of viral lineages. Molecular Biology and Evolution. 2011;28(12):3355–3365. doi: 10.1093/molbev/msr170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wertheim JO, Chu DKW, Peiris JSM, Kosakovsky Pond SL, Poon LLM. A case for the ancient origin of coronaviruses. Journal of Virology. 2013;87(12):7039–7045. doi: 10.1128/JVI.03273-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weyer J, Dermaux-Msimang V, Grobbelaar A, Le Roux C, Moolla N, Paweska J, Blumberg L. Epidemiology of human rabies in South Africa, 2008–2018. South African Medical Journal . 2020;110:877–881. doi: 10.7196/SAMJ.2020.v110i9.14324. [DOI] [PubMed] [Google Scholar]
- Whitbread TA, Kabuya KJ, Naran N, Juggernath AM, Mathews MA, Blumberg LH, Weyer J, Essel V. A retrospective review of rabies postexposure prophylaxis queries, South Africa, 2016–2019. Southern African Journal of Infectious Diseases. 2022;37:354. doi: 10.4102/sajid.v37i1.354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitehouse, E.R., A. Mandra, J. Bonwitt, E.A. Beasley, J. Taliano, and A.K. Rao. 2022. Human rabies despite postexposure prophylaxis: A systematic review of fatal breakthrough infections after zoonotic exposures. The Lancet Infectious Diseases S1473–3099 (22): 00641–00647. 10.1016/S1473-3099(22)00641-7. [DOI] [PMC free article] [PubMed]
- WHO., 2023. Health topics-Rabies. https://www.who.int/health-topics/rabies
- WHO Rabies Modeling Consortium The potential effect of improved provision of rabies postexposure prophylaxis in Gavi-eligible countries: A modeling study. The Lancet Infectious Diseases. 2019;19(1):102–111. doi: 10.1016/S1473-3099(18)30512-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WHO Rabies Modeling Consortium Zero human deaths from dog-mediated rabies by 2030: perspectives from quantitative and mathematical modeling. Gates Open Res. 2020;3:1564. doi: 10.12688/gatesopenres.13074.2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wijaya L, Tham CYL, Chan YFZ, Wong AWL, Li LT, Wang LF, Bertoletti A, Low JG. An accelerated rabies vaccine schedule based on toll-like receptor 3 (TLR3) agonist PIKA adjuvant augments rabies virus specific antibody and T-cell response in healthy adult volunteers. Vaccine. 2017;35:1175–1183. doi: 10.1016/j.vaccine.2016.12.031. [DOI] [PubMed] [Google Scholar]
- Wilde H, Hemachudha T, Jackson AC. Viewpoint: Management of human rabies. Transactions of the Royal Society of Tropical Medicine and Hygiene. 2008;102:979–982. doi: 10.1016/j.trstmh.2008.04.008. [DOI] [PubMed] [Google Scholar]
- Willoughby RE, Jr, Tieves KS, Hoffman GM, Ghanayem NS, Amlie-Lefond CM, Schwabe MJ, Chusid MJ, Rupprecht CE. Survival after treatment of rabies with induction of coma. New England Journal of Medicine. 2005;352(24):2508–2514. doi: 10.1056/NEJMoa050382. [DOI] [PubMed] [Google Scholar]
- World Health Organization (WHO)., 2018. Laboratory techniques in rabies, 5th ed. Geneva
- Yang DK, Kim HH, Lee KK, Yoo JY, Seomun H, Cho IS. Mass vaccination has led to the elimination of rabies since 2014 in South Korea. Clinical and Experimental Vaccine Research. 2017;6:111–119. doi: 10.7774/cevr.2017.6.2.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang, H., S. Chen, Z. Dai, L. Luo, Q. Sun, S. Zhao, G. Zeng, Z. Liu, and S. Hu. 2022. Integrated rabies surveillance - Hunan Province, China, 2020. China CDC Weekly. 4: 986–989. 10.46234/ccdcw2022.200. [DOI] [PMC free article] [PubMed]
- Yue, Y., Q. Chen, D. Mu, Y. Li, and W. Yin. 2022. A descriptive analysis of human rabies in mainland China, 2005–2020. International Journal of Environmental Research and Public Health 20: 380. 10.3390/ijerph20010380. [DOI] [PMC free article] [PubMed]
- Zerbini FM, Siddell SG, Mushegian AR, Walker PJ, Lefkowitz EJ, Adriaenssens EM, Alfenas-Zerbini P, Dutilh BE, García ML, Junglen S, et al. Differentiating between viruses and virus species by writing their names correctly. Archives of Virology. 2022;167(4):1231–1234. doi: 10.1007/s00705-021-05323-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, J., N. Shi, G. Li, L. Li, Y. Bai, L. Yang, W. Zhao, J. Gao, J. Wei, W. Zhao, et al. 2022. Population pharmacodynamic analyses of human anti-rabies virus monoclonal antibody (Ormutivimab) in healthy adult subjects. Vaccines (basel). 10 (8): 1218. 10.3390/vaccines10081218. [DOI] [PMC free article] [PubMed]
- Zhao JH, Zhao LF, Liu F, Jiang HY, Yang JL. Ferret badger rabies in Zhejiang, Jiangxi and Taiwan China. Archives of Virology. 2019;164(2):579–584. doi: 10.1007/s00705-018-4082-5. [DOI] [PubMed] [Google Scholar]
- Zinsstag J, Lechenne M, Laager M, Mindekem R, Naissengar S, Oussiguere A, Bidjeh K, Rives G, Tessier J, Madjaninan S, et al. Vaccination of dogs in an African city interrupts rabies transmission and reduces human exposure. Science Translational Medicine. 2017;9:eaaf6984. doi: 10.1126/scitranslmed.aaf6984. [DOI] [PubMed] [Google Scholar]
- Zucca, P., M.C. Rossmann, M. Dodic, Y. Ramma, T. Matsushima, S. Seet, S. Holtze, A. Bremini, I. Fischinger, G. Morosetti, et al. 2021. What do adolescents know about One Health and zoonotic risks? A school-based survey in Italy, Austria, Germany, Slovenia, Mauritius, and Japan. Front Public Health. 9: 658876. 10.3389/fpubh.2021.658876. [DOI] [PMC free article] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.