

Abstract
The isopod crustaceans reported from or expected to occur in littoral and sublittoral marine habitats of the Southern California Bight (SCB) in the northeastern Pacific Ocean are reviewed. A total of 190 species, representing 105 genera in 42 families and six suborders are covered. Approximately 84% of these isopods represent described species with the remaining 16% comprising well-documented “provisional” but undescribed species. Cymothoida and Asellota are the most diverse of the six suborders, accounting for ca. 36% and 29% of the species, respectively. Valvifera and Sphaeromatidea are the next most speciose suborders with between 13–15% of the species each, while the suborder Limnorioidea represents fewer than 2% of the SCB isopod fauna. Finally, the mostly terrestrial suborder Oniscidea accounts for ca. 5% of the species treated herein, each which occurs at or above the high tide mark in intertidal habitats. A key to the suborders and superfamilies is presented followed by nine keys to the SCB species within each of the resultant groups. Figures are provided for most species. Bathymetric range, geographic distribution, type locality, habitat, body size, and a comprehensive list of references are included for most species.
Keywords: Baja California, intertidal, isopod crustaceans, keys to species, northeastern Pacific, southern California, subtidal
Introduction
The Southern California Bight (SCB) is an important ecological region and economic resource that extends more than 600 km from Point Conception, California, USA to Cabo Colonet, Baja California, Mexico in the northeastern Pacific Ocean (Fig. 1). Because of its diverse and productive coastal ecosystems (e.g., rocky and sandy beach intertidal habitats, marshes, bays, lagoons, and estuaries, nearshore kelp forests and reefs, and offshore soft-bottom and hard-bottom benthic habitats of the continental shelf, slope, basins, and submarine canyons), and its proximity to dense human populations and associated pollutant inputs, the SCB is the focus of some of the largest and most comprehensive ocean monitoring programs in the world (Schiff et al. 2002, 2016, 2019). A key component of these programs is documenting changes in bottom dwelling invertebrate communities over space and time. In the SCB these include long-term localized programs conducted by four major wastewater discharge agencies (see City of San Diego 2018; Los Angeles County Sanitation Districts 2020; City of Los Angeles 2021; Orange County Sanitation District 2022), as well as broader regional programs that have been conducted every 4 or 5 years since 1994 (e.g., Allen et al. 1998, 2002, 2007, 2011; Bergen et al. 1998; Ranasinghe et al. 2003, 2007, 2012; Gillett et al. 2017, 2022; Walther et al. 2017; Wisenbaker et al. 2021). Key to the success of these and other programs and surveys is assuring taxonomic standardization of the resultant macroinvertebrate data sets. Fortunately, the SCB has also been home to the Southern California Association of Marine Invertebrate Taxonomists (SCAMIT) since 1982, whose members have ensured the production of accurate and reliable information for the region’s macrobenthic invertebrate fauna since that time.
Figure 1.

Southern California Bight region from Point Conception, California, USA to Punta Colonet, Baja California, Mexico, including the eight Channel Islands and Islas Coronado.
Isopoda (Crustacea: Peracarida) is a diverse and ancient order of crustaceans comprising more than 10,600 living marine, freshwater and terrestrial species known worldwide (see Boyko et al. 2008 onwards). Several recent reviews have focused on the diversity and distribution of the major groups of isopods throughout the world. These include global reviews of the biodiversity of freshwater isopods (Wilson 2008a), the marine isopods exclusive of the epicarideans and asellotes (Poore and Bruce 2012), the bopyrid and cryptoniscid isopods that are ectoparasitic on other crustaceans (McDermott et al. 2010; Williams and Boyko 2012), and the cymothoid isopods that are parasites of marine and freshwater fishes (Smit et al. 2014). Also see Smit et al. (2019) for a review of the current state of knowledge for the various parasitic crustacean taxa, including the isopods. Overall, the isopods are well represented in the SCB with more than 130 species listed in SCAMIT (2021).
There are a number of general monographs, natural history guides, taxonomic keys, and other works relevant to the coastal marine invertebrates of the northeastern Pacific Ocean that contain useful information on SCB isopods even though many are focused on regions further to the north or south and in the Gulf of California (e.g., Richardson 1905a; Ricketts and Calvin 1952, 1968; Menzies and Barnard 1959; Johnson and Snook 1967; Schultz 1969; Kozloff 1974, 1983, 1996; Miller 1975; Allen 1976; Brusca and Brusca 1978; Brusca 1980; Lee and Miller 1980; Ricketts et al. 1985; Hinton 1987; Wetzer and Brusca 1997; Wilson 1997; O’Clair and O’Clair 1998; Sept 2002, 2019; Brusca et al. 2004, 2007; Kerstitch and Bertsch 2007). Additional information is included in several species checklists (e.g., Leistikow and Wägele 1999; Espinosa-Pérez and Hendrickx 2001a, 2006; Schmalfuss 2003; Campos and Villarreal 2008). However, except for treatments of the southern California coastal shelf and submarine canyons from more than 50 years ago (Menzies and Barnard 1959; Schultz 1966), a more recent survey of the offshore benthos of the Santa Maria Basin and Western Santa Barbara Channel in the northern SCB (Wetzer and Brusca 1997; Wilson 1997), and a guide to the intertidal and supralittoral species of the California and Oregon coasts (Brusca et al. 2007), most information on the coastal isopods of the region remains scattered amongst various taxonomic or ecological contributions. Consequently, there is presently no single comprehensive treatment of the SCB coastal isopod fauna.
The purpose of this guide is to review all macrobenthic species of isopods known or expected to occur in littoral or sublittoral marine habitats of the SCB. Most of these isopods are free-living species that inhabit soft or hard bottom habitats ranging from the upper intertidal to offshore continental shelf and upper slope (depths < 500 m), as well as inland bays and estuaries. Additional species that occur in deeper waters of the lower continental slope, nearshore basins, submarine canyons, and around oceanic islands in the region are also included. Some species are typically associated with more specific microhabitats or niches. For example, these include isopods living on or within sponges (e.g., some sphaeromatids and asellotes), species living commensally with other isopods or echinoderms (e.g., some asellotes and idoteids), species living on or closely associated with kelp or other marine algae (e.g., many idoteids), species that burrow into wood, algal holdfasts or other substrates (e.g., limnoriids and some sphaeromatids), species that are micropredators or temporary parasites of fishes (e.g., aegids, cirolanids, corallanids, gnathiids), and species that are obligate parasites of other crustaceans (epicarideans) or fishes (cymothoids). Finally, although the focus of this review is on marine isopods, halophilic or semi-terrestrial species within the suborder Oniscidea that occur at or just above the high tide line in many SCB intertidal areas are also included.
A key to the suborders and superfamilies of marine isopods occurring in the SCB is provided, which is followed by nine subsequent keys that identify the local isopod fauna to species. Some representative body types for the major groups are illustrated in Fig. 2. Dorsal whole-body illustrations are included for most species. Unless otherwise noted, these figures were reproduced or modified after original species descriptions or other sources as credited in their respective captions and the list of references. Additional figures illustrating enlarged views of specific diagnostic features referred to in the keys are provided where possible. For provisional (undescribed) species where no figures exist, an image of a representative congener is included if available. A series of endnotes to these keys is provided that includes comprehensive additional information useful in the identification or classification of specific taxa, but which does not fit neatly in the annotated list of species. This list follows the endnote section and includes bathymetric and geographic distribution information, type locality, brief habitat notes, maximum body size, and the primary taxonomic, biogeographical, and ecological literature references for each species. Our effort focuses on summarizing and organizing current biodiversity data for the SCB marine isopods and is our best attempt at bringing together their morphological, habitat, and distributional data. Although by no means exhaustive, we recognize that available illustrations in many instances are less than ideal. Presenting primarily dorsal whole-body figures is a means of standardizing taxonomic diversity and aiding in the identification of different species, but we acknowledge it is not always the best depiction of a given taxon. Other views are often readily available, and we encourage the reader to refer to the cited and original descriptions for greater detail.
Figure 2.

Examples of representative family-level body types for each of the six suborders of marine isopods occurring in the SCB. A suborder CymothoidaA1Anthuridae (Haliophasmageminatum) A2Cirolanidae (Cirolanaharfordi) A3Gnathiidae (Gnathiatridens, male and female) A4Bopyridae (Munidionpleuroncodis) B suborder Limnorioidea, Limnoriidae (Limnoriaquadripunctata) C suborder SphaeromatideaC1Serolidae (Heteroseroliscarinata) C2Sphaeromatidae (Dynoideselegans) D suborder Valvifera, Idoteidae (Synidoteamagnifica) E suborder AsellotaE1Janiridae (Ianiropsisanaloga) E2Joeropsididae (Joeropsisconcava) E3Paramunnidae (Pleurogoniumcaliforniense) F suborder Oniscidea, Alloniscidae (Alloniscusperconvexus).
Materials and methods
Species review
All species currently recognized by the Southern California Association of Marine Invertebrate Taxonomists (SCAMIT 2021), as well as previously by the organization (SCAMIT 1994–2018) were reviewed and changes (e.g., additions, deletions, synonymies) verified. Additional species were added based on reviews of original species descriptions, taxonomic monographs, species or habitat guides, ecological studies, and published reports of site-specific or regional surveys conducted in the Southern California Bight (SCB) or elsewhere, as well as by personal observations and examination of field or museum specimens by the authors.
Classification and terminology
The higher-level classification of crustaceans and isopods has evolved over the past two decades, with 11 isopod suborders currently recognized, of which eight include marine species (e.g., see Martin and Davis 2001; Brandt and Poore 2003; Ahyong et al. 2011; Boyko et al. 2013; Wetzer et al. 2013; Martin et al. 2014; Wetzer 2015). The treatment followed herein for the SCB isopods reflects that presently accepted by the World Register of Marine Species (WoRMS: Boyko et al. 2008 onwards) and SCAMIT (2021), with any differences between these lists addressed in the endnotes or annotated species list following the keys.
The Isopoda Latreille, 1817 can be distinguished from the other peracarid orders, and crustaceans in general, by the following combination of characters (after Brusca and Iverson 1985; Wetzer et al. 1997; Brusca et al. 2007).
Body usually dorsoventrally depressed but may be cylindrical or tubular in some suborders (e.g., Anthuridea and Phreatoicidea).
Body without a carapace but with a cephalic shield.
Head (cephalon) compact, typically with compound eyes, two pairs of antennae (first pair minute in Oniscidea), and mouthparts comprising one pair of mandibles, two pairs of maxillae, and one pair of maxillipeds.
Compound eyes sessile (not stalked) but may be on ocular lobes in some Asellota, Cymothoida (i.e., Gnathiidae), and Valvifera.
First and second antennae (antennules and antennae, respectively) uniramous, but with minute scales in a few taxa.
Mandible usually with a 1- to 3-articulate palp and a multidentate incisor process, left and right lacinia mobilis often differ, molar process highly variable.
First and second maxillae (maxillules and maxillae, respectively) without palps.
First thoracic appendages (thoracopods) modified as maxillipeds comprising fifth pair of mouthparts.
Long thorax of eight segments (thoracomeres), the first (and second in Gnathiidae) fused to the cephalon and bearing the maxillipeds, the remaining seven segments (pereonites) being free and collectively comprising a region called the pereon.
Pereon with seven pairs of uniramous legs (pereopods), all generally alike; the exception being the Gnathiidae in which only five pairs of walking legs are present.
Abdomen (pleon) relatively short comprising six somites (pleonites), at least one of which is always fused to the telson to form a pleotelson.
Six pairs of biramous pleonal appendages, including five pairs of plate-like pleopods specialized for respiration and/or swimming, and one pair of fan-like or stick-like, uniarticulate uropods.
Heart located primarily in the pleon.
Young isopods develop within a brood pouch (female’s marsupium) and emerge as a manca before appearance of the last pair of pereopods. Mancas have six pairs of pereopods.
Biphasic molting in which the posterior half of the body molts before the anterior half.
In terms of reproductive status, isopods can be sexed based on the presence or absence of male or female secondary sex characters. If oostegites (or a marsupium) are present with or without eggs or developing embryos, an individual is obviously female. If oostegites are absent, males can be distinguished by the presence of paired penes (may be fused) on the sternum of pereonite 7 (or pleonite 1) and/or appendices masculinae on the endopods of the second pleopods. Absence of all these characters indicates that the individual is either a juvenile, an immature female, or an immature male that has not yet developed secondary sexual features.
Terminology in the keys follows that which is typical for isopods in general (see Brusca and Iverson 1985; Kensley and Schotte 1989; Brusca and Wilson 1991; Wetzer et al. 1997; Brusca et al. 2007). The generalized isopod body plan characteristic of most groups is diagrammed in Fig. 3A, while Fig. 4 shows the specialized body plan characteristic of an adult female bopyrid (i.e., representative epicaridean). See Cohen and Poore (1994: fig. 1) for a detailed diagram of a stylized male gnathiid isopod.
Figure 3.

A Generalized isopod body plan (modified after Kensley and Schotte 1989: fig. 2) B ventral view of valviferan pleon and pleotelson showing opercular uropods (after Poore and Lew Ton 1993: fig. 25) C example of ambulatory pereopod (after Brusca 1983a: fig. 11G) D example of prehensile pereopod (after Brusca 1983a: fig. 11F).
Figure 4.

Bopyrid isopod body plan (adult female). A dorsal view B ventral view. Modified after Markham (1985a: fig. 2).
In general, the isopod body is divided into three regions: the cephalon (head), pereon, and pleon. The cephalic region is referred to as “head” throughout the following keys. The segments of the pereon are referred to as pereonites 1–7 for most taxa or as pereomeres 1–7 for the epicarideans. Likewise, the segments of the pleon are referred to as pleonites 1–5 and the pleotelson (incorporating the fused pleonite 6 and telson) for most taxa (unless fused into fewer segments) or as pleomeres 1–6 plus telson for the epicarideans. The appendages of the pereon are numbered as pereopods 1–7, while the appendages of the pleon are referred to as pleopods 1–5 plus the uropods for all taxa. The first antennae (antenna 1) and second antennae (antenna 2) are referred to as antennules and antennae, respectively. Additional terminology is defined in glossaries of technical terms provided in Martin et al. (2022). Other useful glossaries are included in Kensley and Schotte (1989) and Wetzer et al. (1997), while Brusca and Wilson (1991) provide an extensive detailed description of isopod morphology. Key references regarding taxonomy, systematics, biogeography, and ecology of various isopod taxa are listed below for each suborder represented in this review, while specific references for each species are provided in the annotated species list.
Suborder Cymothoida: Primary references for the superfamily Anthuroidea include Menzies and Barnard (1959), Poore (1975, 1980, 1984a, 1984b, 2001a), Wägele (1984a), Poore and Lew Ton (1986, 1988a, 1988b, 1988c, 1988d), Cadien and Brusca (1993), and Annisaqois and Wägele (2021). Key references for the superfamily Cymothooidea are available for the families Aegidae (e.g., Brusca 1983a; Brusca and France 1992; Bruce 2009a), Cirolanidae (e.g., Bruce 1981; Brusca et al. 1995), Corallanidae (e.g., Delaney 1984, 1989), Cymothoidae (e.g., Brusca 1981; Bruce 1990; Smit et al. 2014; Hata et al. 2017), Gnathiidae (e.g., Cohen and Poore 1994; Smit and Davies 2004; Haney 2006; Tanaka 2007; Wilson et al. 2011), and Tridentellidae (e.g., Bruce 1984; Delaney and Brusca 1985). Within the infraorder Epicaridea, the family Bopyridae has been treated in detail in a long series of papers by Markham since the 1970s (e.g., Markham 1974a, 1975, 1977a, 1985a, 1986, 1992, 2001, 2003, 2004, 2008, 2016, 2020), while Boyko and Williams (2021a, 2021b) recently covered the two known local genera of the family Dajidae. Other valuable recent references for bopyrid and dajid isopods include Boyko et al. (2017), Williams et al. (2019), and Williams and Boyko (2021).
Suborder Limnoriidea: Key references for the family Limnoriidae include Menzies (1951a, 1957, 1958), Cookson (1991), and Borges et al. (2014).
Suborder Sphaeromatidea: Key references for the family Serolidae include Sheppard (1933), Harrison and Poore (1984), Brandt (1988), Wägele (1994), and Bruce (2009b), while important references for the family Ancinidae include Holmes and Gay (1909), Menzies and Barnard (1959), Schultz (1973), Glynn and Glynn (1974), Bruce (1993), and Shimomura (2008). Key references for the Sphaeromatidae include Menzies (1954), Harrison and Holdich (1982, 1984), Harrison and Ellis (1991), Bruce (1993), Schotte (2005), Wetzer et al. (2013, 2018, 2021), Wall et al. (2015), and Wetzer and Mowery (2017).
Suborder Valvifera: Key references for the valviferans include Menzies (1950a, 1951b), Sheppard (1957), Menzies and Barnard (1959), Menzies and Miller (1972), Brusca and Wallerstein (1977), Brusca (1983b, 1984), Brandt (1990), Poore and Lew Ton (1990, 1993), Rafi and Laubitz (1990), Wägele (1991), Poore (2001b, 2015), and King (2003).
Suborder Asellota: Key references for the asellotes include Menzies (1951b, 1952), Menzies and Barnard (1959), George and Strömberg (1968), Hessler et al. (1979), Wilson and Hessler (1980, 1981), Poore (1984c), Wilson (1987, 1994, 1997, 2008b), Wilson and Wägele (1994), Serov and Wilson (1995), Just and Wilson (2004, 2007, 2021), Doti and Wilson (2010), and Brix et al. (2021).
Suborder Oniscidea: Useful references for the oniscids include Van Name (1936, 1940, 1942), Garthwaite et al. (1985, 1992), Garthwaite and Lawson (1992), Leistikow and Wägele (1999), Schmalfuss (2003), Schmidt and Leistikow (2004), Schmidt (2008), and Sfenthourakis and Taiti (2015).
Results
Biodiversity
A total of 190 species of isopods are covered in this review of the Southern California Bight (SCB) isopod fauna, representing 105 genera, 42 families, and six suborders (Appendix 1). These include all but two of the 132 species-level taxa currently recognized by SCAMIT (2021)Endnote 1. Approximately 84% of these isopods represent described species (see Boyko et al. 2008 onwards), while the remaining 16% represent well-documented “provisional” but undescribed species.
Of the six isopod suborders occurring in SCB coastal and offshore waters, the Cymothoida and Asellota are the most diverse, accounting for ~ 36% and 29% of the species, respectively (Table 1). The Valvifera and Sphaeromatidea are the next most speciose suborders, accounting for ~ 13–15% of species, while the suborder Limnorioidea is represented by only three species (< 2%) in SCB waters. Finally, the suborder Oniscidea is represented by nine species (~ 5%) that occur at the terrestrial-marine interface in several intertidal areas of the SCB. See Brusca et al. (2007) or contact R Wetzer for more complete lists of California oniscid isopods.
Table 1.
Number of families, genera, and species for each of the six isopod suborders occurring in littoral and sublittoral marine habitats of the Southern California Bight. For suborders represented by more than one superfamily, the breakdown per superfamily is indicated.
| Suborder/Superfamily | Number taxa per group | ||
|---|---|---|---|
| Families | Genera | Species | |
| Suborder Cymothoida | 15 | 47 | 69 |
| Superfamily Anthuroidea | 5 | 10 | 11 |
| Superfamily Cymothooidea | 6 | 18 | 38 |
| Infraorder Epicaridea | |||
| Superfamily Bopyroidea | 2 | 16 | 17 |
| Superfamily Cryptoniscoidea | 2 | 3 | 3 |
| Suborder Limnoriidea | 1 | 1 | 3 |
| Suborder Sphaeromatidea | 4 | 12 | 25 |
| Superfamily Seroloidea | 1 | 1 | 1 |
| Superfamily Sphaeromatoidea | 3 | 12 | 24 |
| Suborder Valvifera | 4 | 12 | 29 |
| Suborder Asellota | 12 | 26 | 55 |
| Superfamily Janiroidea | 11 | 25 | 54 |
| Superfamily Stenetrioidea | 1 | 1 | 1 |
| Suborder Oniscoidea | 6 | 6 | 9 |
| Total taxa | 42 | 105 | 190 |
The suborder Cymothoida is represented by 69 species distributed amongst four superfamilies (Anthuroidea, Cymothooidea, Bopyroidea, Cryptnoniscoidea) and 15 families. The superfamily Anthuroidea, characterized by long, thin, cylindrical bodies usually at least 6 × longer than wide, includes 11 species in five families. Most anthuroids are thought to feed on other small invertebrates and occur in the SCB amongst fouling communities in marinas, bays, and harbors, and in both littoral and sublittoral habitats on the outer coast from the low intertidal to soft and hard-bottom benthos of the continental shelf, slope, and submarine canyons (0–1300 m depths).
The superfamily Cymothooidea is represented by six families with 38 species. Five of these families (Aegidae, Cirolanidae, Corallanidae, Cymothoidae, Tridentellidae) are generally similar in body form with sleek symmetrical bodies usually ~ 2–6 × longer than wide, the uropods and pleotelson forming a distinct tail fan, and the pereopods modified from ambulatory to prehensile reflecting their different lifestyles (e.g., Fig. 3C, D). For example, the family Cirolanidae is represented by seven species with mostly ambulatory pereopods, although pereopods 1–3 tend to have well-developed dactyli modified for grasping. In the SCB, these species occur intertidally on both sandy and rocky beaches, as well as in shallow to deep water habitats of the continental shelf, slope, and basins (0–1250 m depths). Corallanidae and Tridentellidae, represented by two species each in the SCB, are similar in shape to the cirolanids, but with the mouthparts and anterior pereopods modified for a predatory lifestyle. For example, the dactyli of pereopods 1–3 are usually as long or longer than the propodi and thus adapted for grasping prey. The two SCB corallanids occur from the intertidal to 138 m on the continental shelf, while the two SCB tridentellids have been reported from offshore depths of 53–360 m. The family Aegidae is represented by seven species in the SCB, all of which are temporary parasites of marine fishes and are characterized by prehensile pereopods 1–3 (dactylus strongly recurved and as long or longer than propodus). These species have been reported from the intertidal to deep waters of the continental shelf, slope, basins, and oceanic islands (0–2534 m depths). Although somewhat similar in general body form to the above four families, species of Cymothoidae are even more highly modified for a parasitic lifestyle with all seven pairs of pereopods being strongly prehensile. All cymothoids are obligate parasites of marine or freshwater fishes, most commonly being found attached in the gill chamber or buccal region of their host. Nine species of cymothoids are recognized herein as parasitizing a wide range of marine fishes in nearshore to offshore coastal waters of the SCB.
The sixth family of Cymothooidea occurring in SCB waters is Gnathiidae of which the males are highly modified and characterized by only six free pereonites and five pairs of pereopods. Eleven species of gnathiids are reported in the SCB, including three that are awaiting formal description. These 11 species occur in a wide range of habitats from the intertidal to shallow subtidal, and from the benthos of the offshore continental shelf, slopes, and submarine canyons to depths of ~ 1400 m.
Bopyroidea and Cryptoniscoidea comprise the remaining two superfamilies of Cymothoida, both of which are highly modified obligate parasites of other crustaceans. Bopyroidea is represented by 17 species in two families (Bopyridae and Ionidae) in shallow to deep SCB waters. Sixteen of these species are branchial parasites of a wide range of decapod crustaceans (e.g., hermit crabs, shrimp, mud shrimp, ghost shrimp, galatheid crabs, squat lobsters, porcelain crabs, grapsid crabs) while one species is an abdominal parasite of mud shrimp. In contrast, Cryptoniscoidea is represented by only three species in two families. These include Dajidae represented herein by at least two species that are ectoparasites on the dorsal carapace of several species of shrimp, and Hemioniscidae represented by a single species that is an ectoparasite of barnacles.
The suborder Limnoriidea is represented by three species of the family Limnoriidae in SCB waters. All three species occur in shallow waters (0–30 m depth) where they burrow into either wood (2 species) or algal holdfasts (1 species).
The suborder Sphaeromatidea is represented by 25 species in SCB waters distributed between two superfamilies (Seroloidea and Sphaeromatoidea) and four families. The serolids (family Serolidae) presently include only a single recognized species in the SCB, Heteroseroliscarinata, which burrows just beneath the sediment surface from shallow waters in bays and harbors, and offshore to depths of ~ 100 m. However, it is possible that shallow vs. deep water populations in the region represent two distinct species (TDS, pers. obs.). In contrast, the superfamily Sphaeromatoidea includes 24 species in three families. Of these, the family Sphaeromatidae is the most diverse, represented herein by a total of 20 species. Most of these species occur from intertidal to shallow subtidal habitats < 30 m depth, although two species, Discerceisgranulosa and Paracerceisgilliana, have been reported from slightly deeper waters between 37–73 m. The other two sphaeromatoid families, Ancinidae and Tecticipitidae, are represented by only three and one species, respectively. All four of these species occur in intertidal or shallow subtidal habitats (< 30 m depths).
The suborder Valvifera can be distinguished from all other local isopods by the possession of hinged opercular uropods that cover the ventral surface of the pleon and pleotelson enclosing the pleopods (see Fig. 3B). Four families and 29 species of valviferans are represented in SCB waters. Idoteidae is the most diverse of these families, comprising 23 species in the study area. Many of these species, especially within the genera Colidotea, Erichsonella, Eusymmerus, Idotea, Pentidotea, and Stenosoma are most common in intertidal and shallow subtidal habitats associated with various species of kelp or algae. In contrast, the two local species of Edotia occur in soft-bottom habitats of the continental shelf between depths of ~ 14–64 m, while three of the four SCB species of Synidotea (except S.harfordi) occur on the shelf or slope at depths of ~ 30–800 m. The remaining three families of SCB valviferans comprise a total of six species. The family Arcturidae is represented by four species that occur in the low intertidal (1 species) or offshore shelf benthos (3 species) at depths < 100 m. The last two families are each represented by a single species. These include Cleantioidesoccidentalis of the Holognathidae in relatively shallow waters (intertidal to ~ 50 m), and Califarcturustannerensis of the Thermoarcturidae in deep waters (~ 1200–1300 m).
The suborder Asellota is represented by 55 species in SCB waters distributed between two superfamilies (Janiroidea and Stenetrioidea) and 12 families. However, only ~ 64% of these species are formally described, with the remaining 36% representing provisional species (see Appendix 1). Stenetrioidea includes a single species (Stenetrium sp. A) reported from 90–131 m in the Santa Maria Basin. In contrast, Janiroidea is represented by a diverse group of 11 families that occur in a wide range of habitats and depths from the intertidal to nearly 4000 m. Janiridae is the most diverse of these families, comprising 18 species; ca. two-thirds of the janirids occur in shallow waters from the intertidal to depths of ~ 30 m, while the remaining third occur on the continental shelf at depths between ~ 30–200 m. Paramunnidae is the next most diverse family with nine species, two of which occur in the shallow subtidal (9–20 m) and seven on the continental shelf (75–197 m). Munnidae is the third most diverse family with eight species, of which five species occur in intertidal to shallow subtidal habitats, two species occur at mostly shelf depths (12–237 m), and one species occurs in deeper slope waters (500 m). The Munnopsidae is represented by six species that occur at depths between 73–1118 m. The Joeropsididae is represented by five species that occur at depths from the intertidal to 161 m, while the Desmosomatidae includes three species that range in depth from ~ 100–3000 m. The remaining five families are each represented by a single species. These include Dendrotionidae (Acanthomunnatannerensis at ~ 600–800 m), Haplomunnidae (Haplomunnacaeca at ~ 4000 m), Lepidocharontidae (Microcharon sp. A at 75 m), Nannoniscidae (Nannonisconuslatipleonus at ~ 300–500 m), and Pleurocopidae (Pleurocope sp. A at < 1 m).
The suborder Oniscidea is represented in this review by nine species distributed between six families. Each of these species typically occurs at or above the high tide mark in its respective habitat. The families Alloniscidae (2 species) and Tylidae (1 species) occur on sandy beaches in the SCB. The Detonidae (3 species) and Halophiloscidae (1 species) both occur in marshes, bays, and estuaries. The Platyarthridae (1 species) is reported to occur on both sandy beaches and at the edges of marshes. The Ligiidae is represented herein by a single species that typically occurs in the spray zone on rocky intertidal shores.
Keys to species
Ten keys were constructed to facilitate identification of the 190 species of isopods included in this guide. Key A represents a key to the suborders and main superfamilies, which are then identified to species in Keys B–J. Keys B–E cover the suborder Cymothoida (superfamilies Anthuroidea, Cymothooidea, Bopyroidea, and Cryptoniscoidea). Key B covers the Anthuroidea (5 families, 11 species). Key C covers the families Aegidae, Cirolanidae, Corallanidae, Cymothoidae, and Tridentellidae of Cymothooidea (27 species). Key D covers the remaining cymothooidean family, Gnathiidae (11 species). Key E covers the epicaridean superfamilies Bopyroidea and Cryptoniscoidea (4 families, 20 species). Key F covers the suborder Limnoriidea (1 family, 3 species). Key G covers the superfamilies Seroloidea and Sphaeromatoidea of the suborder Sphaeromatidea (4 families, 25 species). Key H covers the suborder Valvifera (4 families, 29 species). Key I covers the superfamilies Janiroidea and Stenetrioidea of the suborder Asellota (12 families, 55 species). Key J covers the suborder Oniscidea (6 families, 9 species).
Key A. Suborders and Superfamilies of SCB Marine Isopods
| 1 | Adult isopods free-living, but may live commensally with other isopods or invertebrates; females and males with clear bilateral symmetry, typically similar in size; antennae well developed, never vestigial; antennules variable | 2 |
| – | Adult isopods obligate parasites of other crustaceans; females with slightly to highly distorted or reduced bilateral symmetry; male minute, bilaterally symmetrical, living on body of adult female; antennae vestigial in female; antennules reduced to ≤ 3 articles [Suborder Cymothoida: Superfamilies Bopyroidea and Cryptoniscoidea] | Key E |
| 2 | Adult isopods with 5 pairs of pereopods (thoracomere 2 fused to cephalon with its appendages modified into pylopods; thoracomere 8 reduced, without legs); adult males with enlarged, forward projecting forceps-like mandibles; females without mandibles [Suborder Cymothoida: Superfamily Cymothooidea (in part), Family Gnathiidae] | Key D |
| – | Adult isopods usually with 7 pairs of distinct pereopods (7th pair may be small and folded against ventral body wall in juveniles); newly released isopods (mancas) from the marsupium with only 6 pairs of pereopods; males without projecting forceps-like mandibles; females with mandibles | 3 |
| 3 | Species terrestrial or halophilic, mostly restricted to upper littoral (e.g., high tide line, spray zone) or brackish water habitats along coast; antennules vestigial, minute; pleon always composed of 5 free pleonites plus pleotelson [Suborder Oniscidea: Superfamily Oniscoidea] | Key J |
| – | Species fully marine, occurring in littoral or sublittoral habitats; antennules normal, or not minute if reduced; pleon variable, with or without fused pleonites | 4 |
| 4 | Uropods operculate, modified into pair of ventral covers (opercula) enclosing the pleopods (Fig. 3B), but do not confuse with operculate pleopods [Suborder Valvifera] | Key H |
| – | Uropods not modified as ventral opercula, hinged laterally or terminally on pleotelson | 5 |
| 5 | Uropods typically flattened and hinged on anterolateral margins of pleotelson (may be greatly reduced) | 6 |
| – | Uropods styliform and hinged terminally or nearly so on posterior margins of pleotelson [Suborder Asellota: Superfamilies Janiroidea and Stenetrioidea] | Key I |
| 6 | Adult body elongated, usually > 6 × longer than wide [Suborder Cymothoida: Superfamily Anthuroidea] | Key B |
| – | Adult body not elongated, < 4 × longer than wide (not elongated) | 7 |
| 7 | Uropods greatly reduced with small claw-like exopods, generally not visible dorsally; species burrow in wood or algal holdfasts [Suborder Limnoriidea: Superfamily Limnorioidea] | Key F |
| – | Uropods not as above, clearly visible dorsally as expanded, flattened “tail fan” or long caudal processes | 8 |
| 8 | Pleon composed of 4 or 5 free pleonites plus pleotelson [Suborder Cymothoida: Superfamily Cymothooidea (in part), Families Aegidae, Cirolanidae, Corallanidae, Cymothoidae, and Tridentellidae] | Key C |
| – | Pleon composed of ≤ 3 free pleonites plus pleotelson [Suborder Sphaeromatidea: Superfamilies Seroloidea and Sphaeromatoidea] | Key G |
Key B. Suborder Cymothoida, Superfamily Anthuroidea
Figure 5.

Isopoda, Cymothoida, Anthuroidea, Paranthuridae: AParanthuraelegansA1 lateral view of head (piercing mouthparts) A2 dorsal view of pereonite 7, free pleonites 1–5, pleotelson and uropods (after Menzies 1951b; Wetzer and Brusca 1997) BParanthurajaponicaB1 dorsal close-up view of head B2 dorsal view of fused pleonites 1–5, pleotelson and uropods (after Richardson 1909; Brusca et al. 2007) CCalifanthurasquamosissima (male with antenna loaded with aesthetascs) C1 lateral view of head (piercing mouthparts) C2 dorsal view of pereonites 6 and 7, dorsally fused pleonites 1–5, pleotelson and uropods (after Menzies 1951b) DColanthurabruscai, dorsal view of pereonites 6 and 7, free pleonites 1–5, pleotelson and uropods (after G. Poore, personal contribution).
Figure 6.

Isopoda, Cymothoida, Anthuroidea, Anthuridae: AAmakusanthuracaliforniensisA1 dorsal close-up view of head A2 dorsal view of pereonite 7, dorsally fused pleonites 1–5, pleotelson and uropods (after Wetzer and Brusca 1997) BCyathuramundaB1 lateral view of head (biting mouthparts) B2 dorsal view of pereonite 7, dorsally fused pleonites 1–5, pleotelson and uropods (after Menzies 1951b) CHaliophasmageminatumC1 dorsal view of head and pereonites 1 and 2 of male C2 dorsal view of pereonite 7, dorsally fused pleonites 1–5, pleotelson and uropods (after Wetzer and Brusca 1997) DMesanthuraoccidentalisD1 dorsal view of pereonite 7, dorsally fused pleonites 1–5, pleotelson and right uropod (after Allen 1976). Antheluridae: EAnanthuralunaE1 dorsal close-up view of head E2 dorsal view of pereonite 7, free pleonites 1–5, pleotelson and uropods (after Schultz 1966). Hyssuridae: FKupellonura sp. A F1 dorsal close-up view of head F2 dorsal view of pereonite 7, free pleonites 1–5, pleotelson and uropods (after Wetzer and Brusca 1997).
| 1 | Eyes present | 2 |
| – | Eyes absent | 10 |
| 2 | Mouthparts form forward directed cone-like structure under the head, adapted for piercing and sucking | 3 [Paranthuridae] |
| – | Mouthparts not forming ventral cone-like structure, adapted for biting and chewing | 6 |
| 3 | Pereon composed of 7 distinct, well-developed pereonites with 7 pairs of pereopods; pereonite 7 ca. half as long as pereonite 6 and visible laterally | 4 |
| – | Pereon composed of 6 distinct pereonites with 6 pairs of pereopods (7th pereopods absent); pereonite 7 very short, < 20% as long as pereonite 6, not visible laterally | 5 |
| 4 | Pleonites 1–5 free, not fused; pleonite 5 ca. 3 × longer than other pleonites (Fig. 5A) | Paranthuraelegans |
| – | Pleonites 1–5 fused mid-dorsally, but distinct laterally; all pleonites of similar length (Fig. 5B) | Paranthurajaponica |
| 5 | Pleonites 1–5 free, separated from each other by dorsal integumental folds; pleonite 1 ca. 2 × longer than pleonite 2 (Fig. 5D) | Colanthurabruscai |
| – | Pleonites 1–5 dorsally fused, all pleonites of similar length (Fig. 5C) | Califanthurasquamosissima |
| 6 | Dorsal surface of pleotelson with median row of spines; uropodal and pleotelsonic margins serratedEndnote 2 | Eisothistos sp. A [Expanathuridae] |
| – | Dorsal surface of pleotelson smooth or ridged, without spines; uropodal and pleotelsonic margins not serrated | 7 [Anthuridae] |
| 7 | Pleotelson with 3 raised dorsal longitudinal ridges or carinae; uropodal exopods curve up and over base of pleotelson (Fig. 6C) | Haliophasmageminatum |
| – | Pleotelson without dorsal ridges or carinae; uropodal exopods may or may not curve up over pleotelson | 8 |
| 8 | Pleonites 1–5 fused only along dorsal midline, segments free laterally and visible dorsally; uropodal exopods ca. half length of endopods and pleotelson, curving up and partially over pleotelson base; uropodal endopods narrow, ca. half as wide as pleotelson (Fig. 6A) | Amakusanthuracaliforniensis |
| – | Pleonites 1–5 completely fused dorsally; uropodal exopods > 50% length of endopods and pleotelson, may or may not cover the pleotelson dorsally; uropodal endopods broad, subequal in width to pleotelson | 9 |
| 9 | Dorsal surface of pereon pigmented, with complete or nearly complete dark rings on pereonites 2–6 and posterior transverse band on pereonite 7; uropodal exopods partially cover dorsal surface of pleotelson (Fig. 6D) | Mesanthuraoccidentalis |
| – | Dorsal surface of pereon covered with diffuse pigment splotches, but without pigment rings; uropodal exopods do not cover base of pleotelson (Fig. 6B) | Cyathuramunda |
| 10 | Uropodal exopods with distinct lateral lobes, exopods overlapping broadly to cover almost entire dorsal surface of pleotelson; uropodal rami nearly reach, but do not exceed, posterior margin of pleotelson; tips of uropodal rami without tufts of stiff setae (Fig. 6F) | Kupellonura sp. A [Hyssuridae] |
| – | Uropodal exopods without lateral lobes, overlapping only base of pleotelson; uropodal endopod distinctly longer than pleotelson; tips of uropodal rami with tufts of long stiff setae (Fig. 6E) | Ananthuraluna [Antheluridae] |
Key C. Suborder Cymothoida, Superfamily Cymothooidea (in part): Families Aegidae, Cirolanidae, Corallanidae, Cymothoidae, Tridentellidae
Figure 7.

Isopoda, Cymothoida, Cymothooidea, Cirolanidae: ACirolanadiminuta (after Brusca et al. 1995) BCirolanaharfordi (after Brusca et al. 1995) CEurydicecaudata (after Brusca et al. 1995) DExcirolanachiltoni (after Schultz 1969) EExcirolanalinguifrons (after Schultz 1969) FMetacirolanajoanneae (after Schultz 1966) GNatatolanacaliforniensis (after Brusca et al. 1995).
Figure 11.

Isopoda, Cymothoida, Cymothooidea, Cymothoidae (in part): ACeratothoagaudichaudiiA1 pereopod 4 with arrow indicating carina (after Brusca 1981) BCeratothoagilbertiB1 pereopod 4 (after Brusca 1981) CElthusacalifornicaC1 pereopod 4 (after Brusca 1981) DElthusamenziesiD1 pereopod 4 (after Menzies 1962a; Brusca 1981) EElthusavulgarisE1 pereopod 4 with arrow indicating carina (after Brusca 1981).
| 1 | Pereopods 1–3 ambulatory with dactylus shorter than propodus (e.g., Fig. 3C) | 2 [Cirolanidae] |
| – | Pereopods 1–3 prehensile or sub-prehensile with dactylus generally as long as, or longer than propodus and strongly curved (e.g., Fig. 3D) | 8 |
| 2 | Eyes absent; head immersed in pereonite 1 with posterior margin appearing deeply concave; pereon with coxae 4–7 produced beyond posterior margins of their respective pereonites, at least 2 or more visible in dorsal view; lateral margins of pleonite 5 obscured by pleonite 4 (Fig. 7G) | Natatolanacaliforniensis |
| – | Eyes present; posterior margin of head not appearing distinctly concave; with or without dorsally visible coxae on pereonites; lateral margins of pleonite 5 may or may not be obscured by pleonite 4 | 3 |
| 3 | Coxae of pereonites 2–7 well-developed, typically visible in dorsal view and expanded laterally with acute posterior angles; epimeres of pleonites 2–5 well-developed, expanded laterally, with acute posterior angles; pleotelson with squarish to pointed posterior margin and a strong, middorsal longitudinal ridge; margins of pleotelson and uropodal rami notched (Fig. 7F) | Metacirolanajoanneae |
| – | Pereonites, pleonites, pleotelson and uropods not as above | 4 |
| 4 | Head with prominent spatulate rostral process separating left and right antennules (Fig. 7D, E) | 5 |
| – | Head without a prominent rostral process between antennules | 6 |
| 5 | Posterior margin of pleotelson broadly rounded and crenulate; antennular peduncle articles 2 and 3 subequal in length (Fig. 7E) | Excirolanalinguifrons |
| – | Posterior margin of pleotelson obtusely rounded and acuminate; antennular peduncle article 3 longer than article 2 (Fig. 7D) | Excirolanachiltoni |
| 6 | Antennules geniculated, with peduncle article 1 longer than articles 2 or 3, and article 2 arising at right angles to article 1; peduncle of antennae with 4 articles, antennae long and extending beyond pereonite 7; lateral margins of pleonite 5 not obscured by pleonite 4; uropodal rami truncate distally, exopod does not extend to posterior margin of pleotelson (Fig. 7C) | Eurydicecaudata |
| – | Antennules not geniculated; peduncle of antennae with 5 articles; lateral margins of pleonite 5 obscured by pleonite 4; uropodal rami distally rounded or acuminate, extending beyond posterior of pleotelson | 7 |
| 7 | Uropodal rami with apical notches and not distally rounded; peduncle articles 1 and 2 of antennules fused; coxae of pereonites 5–7 visible dorsally; pereonites, pleonites and pleotelson without dorsal tubercles, carina, or setae (Fig. 7A) | Cirolanadiminuta |
| – | Uropodal rami rounded distally, without notches; peduncle articles 1 and 2 of antennules not fused; coxae visible dorsally on pereonites 2–7; pleonites 3–5 with row of small tubercles on posterior margins; pleotelson of adult males with two large dorsal submedian tubercles or carinae (Fig. 7B) | Cirolanaharfordi |
| 8 | Pereopods 4–7 ambulatory (dactylus shorter than propodus) | 9 |
| – | Pereopods 4–7 prehensile (dactylus generally as long as, or longer than propodus and strongly curved); adults parasitic on fishes | 19 [Cymothoidae] |
| 9 | Dorsal surface of pleon tuberculate, with small to medium tubercles present on posterior margins of at least pleonites 3–5 | 10 |
| – | Dorsal surface of pleon without tubercles | 13 [Aegidae] |
| 10 | Pleotelson dorsally setose, lateral margins with single incision | 11 [Corallanidae] |
| – | Pleotelson not dorsally setose, lateral margins without incisions | 12 [Tridentellidae] |
| 11 | Male head with 3 large horns or tubercles, including 1 rostral and 2 posterolateral between the eyes (female without tubercles); pereonites 2–7 without dorsal setae or tubercles; pleotelson subtriangular with rounded apex, dorsal surface setose except for median longitudinal area (Fig. 8A) | Excorallanatricornisoccidentalis |
| – | Head of both males and females without horns or tubercles; pereonites 4–7 with dorsal setae and row of small tubercles on posterior margin; pleotelson triangular with subacute apex, entire dorsal surface densely covered with bifid golden setae (Fig. 8B) | Excorallanatruncata |
| 12 | Body dorsal surface sculptured with low or small tubercles; head of male with 5 low tubercles, including 1 rostral, 1 pair near anterior margin, and 1 pair near posterior margin; male pereonite 1 with 2 small, median tubercles near anterior margin; female lacking tubercles on head and pereon; pleonites 3–5 with small tubercles on posterior margins; pleotelson minutely tuberculate dorsally with widely rounded, slightly crenulate posterior margin (Fig. 8D) | Tridentellaquinicornis |
| – | Body dorsal surface sculptured with large processes and numerous tubercles; male head with 2 dorsal posterolateral horns, frontal margin produced into large, upturned process and smaller ventrally projecting rostrum; pereonite 1 with 3 large dorsal processes; all pereonites and pleonites with numerous dorsal tubercles that increase in size and become more spine-like posteriorly; pleotelson triangular with subtruncate apex, dorsally covered with longitudinal rows of large, spine-like tubercles; females much less spinose than males, lacking large processes on head and pereonite 1 (Fig. 8C) | Tridentellaglutacantha |
| 13 | Peduncular articles 1 and 2 of antennules greatly expanded (dilated), article 2 with gradual distal process extending 25–50% the distance into article 3; posterior margin of pleotelson truncate, crenulated and fringed with setae (Fig. 8E) | Aegalecontii |
| – | Peduncular articles of antennules not dilated, article 2 without distal process; posterior margin of pleotelson rounded or subacuminate | 14 |
| 14 | Eyes large, close-set, nearly touching at midline; pleotelson shield-shaped with subacuminate apex and weekly serrated (notched) posterolateral margins; uropodal rami ovate with subacuminate apices (Fig. 8F)Endnote 3 | Aegiochusplebeia |
| – | Eyes medium to large, but distinctly separated and not nearly touching medially; posterior margin of pleotelson rounded; uropodal rami with broadly rounded to truncate apices | 15 |
| 15 | Medial process of uropodal peduncle very long, extending at least 75% of length of endopod | 16 |
| – | Medial process of uropodal peduncle extends 50% or less of length of endopod | 17 |
| 16 | Propodi of pereopods 1–3 with large, broad, spine-bearing medial lobe; dactyli of pereopods 1–3 longer than propodi; frontal lamina broadly expanded anteriorly, arrowhead or spatulate shaped (Fig. 9B) | Rocinelabelliceps |
| – | Propodi of pereopods 1–3 without expanded medial lobe; dactyli of pereopods 1–3 subequal in length to propodi; frontal lamina thin and narrow (Fig. 9E) | Rocinelasignata |
| 17 | Medial process of uropodal peduncle extends < 40% of length of endopod; propodi of pereopods 1–3 with 4 stout, recurved acute spines; merus of pereopods 1–3 with 5–8 acute spines (3–5 distal, 2 or 3 proximal) (Fig. 9A) | Rocinelaangustata |
| – | Medial process of uropodal peduncle extends ~ 50% of length of endopod; propodi of pereopods 1–3 with 4–6 acute spines; merus of pereopods 1–3 with 4 acute spines (3 distal, 1 proximal) | 18 |
| 18 | Propodi of pereopods 1–3 with 5 thin, straight acute spines; apical article of maxillipedal palp with thin, nearly straight, acute spines (Fig. 9C) | Rocinelalaticauda |
| – | Propodi of pereopods 1–3 with 4–6 stout and recurved acute spines; apical article of maxillipedal palp with stout, recurved acute spines (Fig. 9D) | Rocinelamurilloi |
| 19 | Pleopods and uropods not setose | 20 |
| – | Pleopods and uropods heavily setose, adapted for swimming (juvenile cymothoids)Endnote 4 | unidentified Cymothoidae |
| 20 | Body very broad and darkly pigmented; pereon at least 2× as wide as pleon with strongly convex lateral margins (widest at pereonite 5); parasite of barspot cardinalfish and Panamic fanged blenny in Eastern Pacific (Fig. 10A)Endnote 5 | Renocilathresherorum |
| – | Body not as above | 21 |
| 21 | Posterior margin of head weakly to strongly trisinuate; pleon not immersed in pereon | 22 |
| – | Posterior margin of head not trisinuate; pleon partially immersed in pereon | 23 |
| 22 | Head not immersed in pereonite 1, posterior border distinctly trisinuate; coxal margins of all or just posterior pereonites with acute or subacute posterolateral angles, coxae may be held close to body (acuminata form) or greatly expanded laterally (aster form); uropods visible dorsally, extending clearly beyond posterior border of pleotelson; parasite of ~ 40 different species of fishes (Fig. 10B) | Nerocilaacuminata |
| – | Head somewhat immersed in pereonite 1, subquadrate anteriorly with weakly trisinuate posterior border; uropods not visible in dorsal view, typically held concealed under pleotelson and not extending beyond posterior border; parasite of Pacific bumper, pompanos, serranos, carangids, and other fishes (Fig. 10C) | Smenispaconvexa |
| 23 | Basal articles of antennules expanded and touching or nearly touching | 24 |
| – | Basal articles of antennules not expanded and touching | 25 |
| 24 | Pereopods 4–7 carinate; posterior margin of pleonite 5 trisinuate except in occasional males; parasite of pelagic fishes, including striped mullet off southern California and pompanos and herring off Baja California (Fig. 11A)Endnote 6 | Ceratothoagaudichaudii |
| – | Pereopods 4–7 not carinate; posterior margin of pleonite 5 smooth, not trisinuate; parasite of mullets and flatfish (Fig. 11B) | Ceratothoagilberti |
| 25 | Antennules longer than antennae; parasite of California and skipper halfbeaks (Fig. 10D) | Mothocyarosea |
| – | Antennules shorter than antennae | 26 |
| 26 | Frontal margin of head broadly rounded or truncate (not produced); bases of pereopods 4–7 with distinct carinae; coxae of pereonites 6 and 7 extending to and usually beyond posterior edge of respective pereonites; pleotelson in adult females nearly 2× as wide as long; parasite of at least 30 species of fishes (Fig. 11E) | Elthusavulgaris |
| – | Frontal margin of head produced; bases of posterior pereopods of females without distinct carinae; coxae of pereonites 6 and 7 not reaching posterior margins of respective pereonites; pleotelson in adult females either as wide as or wider than long | 27 |
| 27 | Merus and carpus of pereopod 4 expanded; bases of pleopods with well-developed accessory lamellae; pleotelson in adult females broadly rounded, ~ 1.5–2.0 × wider than long; males with coxal carinae on pereopods 4–7; parasite of wooly sculpin, northern clingfish, and reef finspot (Fig. 11D) | Elthusamenziesi |
| – | Merus and carpus of pereopod 4 not expanded; accessory lamellae of pleopodal bases not well developed; pleotelson in adult females ca. as wide as long; males without carinae on posterior pereopods; parasite of surfperch, smelt, gobies, killifish, and grunion (Fig. 11C) | Elthusacalifornica |
Figure 8.

Isopoda, Cymothoida, Cymothooidea, Corallanidae: AExcorallanatricornisoccidentalis (after Delaney 1984) BExcorallanatruncata (after Delaney 1984). Tridentellidae: CTridentellaglutacantha (after Delaney and Brusca 1985) DTridentellaquinicornis (after Delaney and Brusca 1985). Aegidae (in part): EAegalecontii (after Schultz 1969) FAegiochusplebeia (after Brusca 1983a).
Figure 9.

Isopoda, Cymothoida, Cymothooidea, Aegidae (in part): ARocinelaangustata (after Brusca and France 1992) BRocinelabelliceps (after Brusca and France 1992) CRocinelalaticauda (after Brusca and France 1992) DRocinelamurilloi (after Brusca and Iverson 1985; Brusca and France 1992) ERocinelasignata (after Brusca and Iverson 1985; Brusca and France 1992) A1–E1 = pereopods 3 A2–E2 = uropods with medial process of peduncle indicated by arrows.
Figure 10.

Isopoda, Cymothoida, Cymothooidea, Cymothoidae (in part): ARenocilathresherorum (after Brusca 1981) BNerocilaacuminata (after Brusca and Iverson 1985) CSmenispaconvexa (after Brusca 1981) DMothocyarosea (after Brusca et al. 2007).
Key D. Suborder Cymothoida, Superfamily Cymothooidea (in part): Family Gnathiidae
Fig. 12
Figure 12.

Isopoda, Cymothoida, Cymothooidea, Gnathiidae: ACaecognathiacrenulatifrons (after Wetzer and Brusca 1997) BCaecognathiasanctaecrucis (after Wetzer and Brusca 1997) CGnathiaclementensis (after Schultz 1966) DGnathiacoronadoensis (after Schultz 1966) EGnathiaproductatridens (after Wetzer and Brusca 1997) FGnathiasteveni (after Menzies 1962a) GGnathiatridens (after Wetzer and Brusca 1997) HGnathiatrilobata (after Schultz 1966).
| 1 | Head with enlarged forceps-like mandibles projecting anteriorly (male gnathiids) | 2 |
| – | Head not as above, without projecting mandibles; body often sac-like (female and juvenile gnathiids)Endnote 7 | Gnathiidae spp. |
| 2 | Eyes present, with or without pigment; telson variable in shape | 3 |
| – | Eyes absent; frontal margin of head (frons) with 3 central processes, laterals larger than middle process; epimeres single, dorsal, laterally projected; telson distinctly triangular (Fig. 12D) | Gnathiacoronadoensis |
| 3 | Pleotelson distinctly triangular | 4 |
| – | Pleotelson arrowhead or T-shaped with base expanded | 8 |
| 4 | Pleonal epimeres laterally expanded, highly visible; body with few to numerous setae | 5 |
| – | Pleonal epimeres not laterally expanded, barely visible; body with relatively few setae | 7 |
| 5 | Frontal margin of head forming broad, transverse, minutely crenulated plate (best viewed ventrally); eyes brown to reddish brown; body setosity light, without numerous setae; pleopods without setae (Fig. 12A) | Caecognathiacrenulatifrons |
| – | Frontal margin of head not transverse (lobes or processes present); eyes reddish brown; body not hirsute, but with numerous setae; pleopods with setae | 6 |
| 6 | Body mottled with brown pigment; frontal margin of head forming centrally extended narrow lobe with crenulations; mandibles split into 2 articlesEndnote 8 | Caecognathia sp. A |
| – | Body without pigment; frontal margin of head trilobed with 3 central subequal processes; mandibles of single article only (not split into 2 articles) (Fig. 12G) | Gnathiatridens |
| 7 | Eyes dark brown, body mottled with brown pigment; frontal margin of head with 3 processes, median process largest and shaped as stepwise pyramid; head with setae and tuberculations (Fig. 12F) | Gnathiasteveni |
| – | Eyes reddish brown; body without pigmentation; frontal margin of head with central 3-dimensional expansion in shape of box, with 2 large setae extending outward centrally; head with setae, but lacking tuberculationsEndnote 8 | Gnathia sp. MBC1 |
| 8 | Pleotelson distinctly T-shaped | 9 |
| – | Pleotelson arrowhead-shaped | 10 |
| 9 | Eyes sessile, dark brown, lens with tuberculations; body speckled with tiny black dots; frontal margin of head produced into single large lobe; pleopods ovate, paddle-like; body hirsute (Fig. 12B) | Caecognathiasanctaecrucis |
| – | Eyes on distinct ocular peduncles, unpigmented, lens without tuberculations; frontal margin of head with 2 medium–large lateral processes and 4 central subequal processes; pleopods long and narrow; body hirsute (Fig. 12C) | Gnathiaclementensis |
| 10 | Pleonal epimeres double (dorsal and ventral); eyes golden/amber in color; frontal margin of head with 3 central, subequal processes; body with numerous scattered setae, but not hirsute (Fig. 12H) | Gnathiatrilobata |
| – | Pleonal epimeres single (dorsal only) | 11 |
| 11 | Eyes golden or amber in color; head with dorsal carina present; frontal margin of head as 1 broad truncate lobe with medial carina; body hirsuteEndnote 8 | Caecognathia sp. SD1 |
| – | Eyes with red and white checkerboard pattern; head without dorsal carina; frontal margin of head with 3 central subequal processes; body not hirsute, but with numerous setae (Fig. 12E) | Gnathiaproductatridens |
Key E. Suborder Cymothoida, Superfamilies Bopyroidea and Cryptoniscoidea
Figure 13.

Isopoda, Cymothoida, Epicaridea, Bopyroidea, Ionidae: AIonecornuta (after Richardson 1905a). Bopyridae (in part): BBathygygegrandis (after Markham 2016) CAnathelgeshyphalus (after Markham 1974a) DLeidyainfelix (after Markham 2002) EMunidionpleuroncodis (after Markham 1975); FPhyllodurusabdominalis (after Richardson 1905a).
Figure 16.

Isopoda, Cymothoida, Epicaridea, Cryptoniscoidea, Dajidae: AHolophryxusalaskensis (adult female) A1 dorsal view A2 ventral view (after Richardson 1905b) BZonophryxusprobisowa (adult female) B1 dorsal view B2 ventral view (Peruvian species = representative for unidentified SCB species of Zonophryxus; after Boyko and Williams 2021a). Hemioniscidae: CHemioniscusbalaniC1 adult female C2 juvenile female (after Nierstrasz and Brender à Brandis 1926; Williams and Boyko 2012).
| 1 | Body of adult female distinctly segmented with clear division of head (cephalon), thoracic (pereon), and abdominal (pleon) regions | 2 [Bopyridae and Ionidae] |
| – | Body of adult female sac-like, with at most weak segmentation visible | 18 [Dajidae and Hemioniscidae] |
| 2 | Adult female body broadly oval; pleon strongly torsioned and reflexed forward over pereon; head without eyes, deeply embedded in pereon, anterior margin covered by oostegites; pleomeres 3–6 fused medially; branchial parasite of deep-water crangonid shrimps of genus Glyphocrangon (Fig. 13B) | Bathygygegrandis |
| – | Female body not as above, pleon not strongly reflexed over pereon | 3 |
| 3 | Female pleon with pleopods and/or elongated lateral plates (epimeres or pleural lamellae) clearly noticeable in dorsal view | 4 |
| – | Female pleon without pleopods or elongated lateral plates noticeable in dorsal view | 12 |
| 4 | Lateral plates of pleon and/or pleopods conspicuously elongate, foliaceous, or lanceolate, with or without digitated or crenulated margins | 5 |
| – | Lateral plates of pleon and pleopods relatively short, distal margins of pleopods mostly rounded | 10 |
| 5 | Adult female with elongated lateral plates fringed with long, branched processes; branchial parasite of ghost shrimps of genus Neotrypaea (Fig. 13A) | Ionecornuta |
| – | Adult female with or without lateral plates, but lateral plates not as above if present | 6 |
| 6 | Adult female body nearly oval, with oostegites conspicuously visible dorsally arching over margins of head and pereon; head longer than wide, deeply immersed in pereon, eyes absent; pereomere 6 much longer than other pereomeres, pereomeres 1–5 concave anteriorly, pereomeres 6 and 7 concave posteriorly; pleon with “triramous appendages” in appearance, but each one consists of a foliaceous lanceolate lateral plate and identical biramous pleopod; branchial parasite of hermit crabs Parapagurodeslaurentae and P.makarovi (Fig. 13C) | Anathelgeshyphalus |
| – | Adult female body not as above, pereomere 6 subequal in length to other pereomeres; pleon with lateral plates and pleopods not appearing like triramous appendages | 7 |
| 7 | Adult females with pair of dorsolateral papillae on pleomere 1; pleon with long, narrow biramous pleopods arising from thin peduncle or stem on each segment, without lateral plates; head slightly wider than long, bilobate, eyes absent; abdominal parasite of mud shrimps of genus Upogebia (Fig. 13F) | Phyllodurusabdominalis |
| – | Adult females without dorsolateral papillae on pleomere 1; pleomeres with both elongated lateral plates and pleopods, but morphology of pleopods not as above | 8 |
| 8 | Pleon of adult females enclosed by tentacular-like, elongated lanceolate lateral plates and pleopodal exopods, each with deeply digitate margins, ventral surface covered partially by smaller foliate pleopodal endopods with crenulated margins; head completely embedded in pereon, dorsal surface divided into two large sub-oval lobes, eyes absent; branchial parasite of grapsid crab Pachygrapsuscrassipes (Fig. 13D) | Leidyainfelix |
| – | Pleon of adult females with lateral plates and pleopods not as above; head not bilobate | 9 |
| 9 | Adult female body ~ 2 × longer than wide, pyriform in shape; head slightly wider than long, eyes absent; coxal plates very large, overlapping, and extending well beyond lateral margins of pereomeres; pleon obscured by prominent ovate lateral plates and long, biramous lanceolate pleopods; branchial parasite of pelagic galatheid “red crab” Pleuroncodesplanipes (Fig. 13E) | Munidionpleuroncodis |
| – | Adult female body slightly longer than wide; head subcircular, eyes absent; coxal plates prominent but not widely extended on pereomeres 1–4; lateral edges on convex side of pereomeres 5–7 produced into slender points reflexed back over dorsum; pleomeres 1–5 with dentate-margined lanceolate lateral plates and similar biramous pleopods; branchial parasite of hermit crab Isochelespilosus (Fig. 14D) | Asymmetrioneambodistorta |
| 10 | Adult female head somewhat bilobate, eyes absent; pereomeres 1–6 with sharply pointed tergal projections on longer side of body; pleomeres 1–5 with short lateral plates and distally rounded uniramous pleopods; branchial parasite of several genera of crangonid and hippolytid shrimps (e.g., Crangon, Eualus) (Fig. 14A) | Argeiapugettensis |
| – | Adult females not as above; head not bilobate, with or without eyes; pereon without sharp tergal projections; pleopods biramous | 11 |
| 11 | Adult female head wider than long with rounded anterior and posterior margins, eyes absent; narrow and rudimentary coxal plates on pereomeres 3 and 4 on both sides of body; branchial parasite of porcelain crab Pachychelespubescens (Fig. 14C) | Aporobopyrusoviformis |
| – | Adult female head subtriangular in shape, anterolateral margins produced into small obtuse projections, eyes present; rudimentary coxal plates of pereomeres 3 and 4 only on longer side of body; branchial parasite of porcelain crabs Pachycheles spp. (Fig. 14B) | Aporobopyrusmuguensis |
| 12 | Adult female body oval to broadly oval, pereon and pleon subequal in width; branchial parasite of mud shrimps of genus Upogebia | 13 |
| – | Adult female body not oval, pleon tapering to much narrower than widest pereomeres | 14 |
| 13 | Adult female body broadly oval, almost as wide as long (L:W ratio ~ 1.2); head almost square, deeply embedded in pereomere 1, eyes absent; ventral surface of pleomeres covered by overlapping lanceolate lateral plates and uniramous uropods, middle region covered by similar sized biramous pleopods (Fig. 14E) | Orthionegriffenis |
| – | Adult female body oval, distinctly longer than wide (L:W ratio ~ 1.5); head slightly wider anteriorly than posteriorly, deeply embedded in pereomere 1, eyes present; ventral surface of pleomeres covered by numerous ridges and lanceolate biramous, marginally tuberculate pleopods (Fig. 14F) | Progebiophilusbruscai |
| 14 | Pleon of adult females composed of 5 medially fused pleomeres and pleotelson, the latter deeply arcuate and embedded in pleomere 5; head roughly triangular, separated from pereon by deep groove, eyes absent; branchial parasite of hippolytid shrimps Hippolytecaliforniensis and Thoralgicola (Fig. 15E) | Schizobopyrinastriata |
| – | Adult female body not as above; pleomeres distinctly separate, not medially fused; head with or without eyes | 15 |
| 15 | Head of adult female partially fused with pereomere 1 and separated by only short lateral notches; frontal margin of head slightly sinuated with anterolateral process usually on just short side of body, small eyes present; pleotelson entirely set within curves of pleomere 5; branchial parasite of snapping shrimps Alpheopsisequidactylus and Synalpheuslockingtoni (Fig. 15A) | Bopyrellacalmani |
| – | Female head distinctly separate and not fused with pereomere 1, eyes absent; pleotelson not completely set within curves of pleomere 5 | 16 |
| 16 | Head of adult female squarish with distinct anterolateral horns and a small anteromedial indentation; pleon with 6 distinct pleomeres separated by deep lateral notches; pleopods 1–4 biramous (5th pair absent), uropods absent; branchial parasite of alpheid shrimp Automate sp. A (Fig. 15B)Endnote 9 | Capitetragonia sp. A |
| – | Female head wider anteriorly than posteriorly, without anterolateral horns; pleon with 5 pleomeres not separated by deep notches and very small 6th segment, 5 pairs of biramous pleopods, and terminal pair of lanceolate uropods | 17 |
| 17 | Endopods of pleopods 1–5 much larger than exopods, elongate and pointed, surface rough with irregular rugae; coxal plates of pereomeres 5–7 not developed as lamellae; branchial parasite of hermit crabs of genus Pagurus (Fig. 15C)Endnote 10 | Eremitionegiardi |
| – | Endopods of pleopods 1–5 only slightly larger than exopods, triangular or ovate, surface smooth; coxal plates of pereomeres 5–7 developed as lamellae; branchial parasite of squat lobsters Galacanthadiomedeae and Munidaquadrispina in the East Pacific (Fig. 15D) | Pseudionegalacanthae |
| 18 | Adult female body simply an egg sac, without evidence of segmentation; antennae and mouthparts absent; parasite on barnacles of genera Balanus and Chthamalus (Fig. 16C) | Hemioniscusbalani [Hemioniscidae] |
| – | Adult female body with weak evidence of segmentation visible dorsally or laterally; antennae and mouthparts present; parasitic on caridean shrimp of families Pasiphaeidae and Pandalidae | 19 [Dajidae] |
| 19 | Adult female body typically elongate and symmetrical, but may be irregular with deeper, stouter body in some specimens; head separate, typically hemispherical, visible dorsally or ventrally; segmentation of pereon usually visible laterally by 4 coxal plates; pleonal region posteriorly projected as unsegmented conical prominence; parasite on carapace of pasiphaeid shrimp Pasiphaeapacifica (Fig. 16A)Endnote 11 | Holophryxusalaskensis |
| – | Adult female body ovate, all regions fused and indistinct dorsally; posterior margin of pleon appears notched with row of triangularly shaped processes; parasite on carapace of pandalid shrimps Pantomusaffinis and Plesionikatrispinus (Fig. 16B)Endnote 12 | Zonophryxus sp. |
Figure 14.

Isopoda, Cymothoida, Epicaridea, Bopyroidea, Bopyridae (in part): AArgeiapugettensis (after Richardson 1905a) BAporobopyrusmuguensis (after Markham 2008) CAporobopyrusoviformis (after Shiino 1934) DAsymmetrioneambodistorta (after Markham 1985b) EOrthionegriffenisE1 ventral view of pleon and pleopods (after Markham 2004) FProgebiophilusbruscaiF1 ventral view of pleon and pleopods (after Campos and de Campos 1998).
Figure 15.

Isopoda, Cymothoida, Epicaridea, Bopyroidea, Bopyridae (in part): ABopyrellacalmani (after Sassaman et al. 1984) BCapitetragoniaalphei (Caribbean species = representative for Capitetragonia sp. A; after Markham 1985a) CEremitionegiardi (after Richardson 1905a) DPseudionegalacanthae (after Richardson 1905a) ESchizobopyrinastriata (after Campos and de Campos 1990).
Key F. Suborder Limnoriidea, Superfamily Limnorioidea: Family Limnoriidae
Fig. 17
Figure 17.

Isopoda, Limnoriidea, Limnorioidea, Limnoriidae: ALimnoriaalgarum (after Menzies 1957) BLimnoriaquadripunctataB1 inner surface of left mandible showing “rasp” (after Menzies 1957) CLimnoriatripunctata (after Allen 1976).
| 1 | Dorsal surface of pleotelson without symmetrically arranged tubercles; left mandible without rasp or file-like ridges; burrowing in algal holdfasts (Fig. 17A) | Limnoriaalgarum |
| – | Dorsal surface of pleotelson with 3 or 4 symmetrically arranged anterior tubercles; left mandible with rasp or file-like ridges (see Fig. 17B); burrowing in wood | 2 |
| 2 | Dorsal surface of pleotelson with 3 anterior tubercles arranged in triangle; lateral and posterior margins of pleotelson tuberculate (Fig. 17C) | Limnoriatripunctata |
| – | Dorsal surface of pleotelson with 2 pairs of anterior tubercles; margins of pleotelson not tuberculate (Fig. 17B) | Limnoriaquadripunctata |
Key G. Suborder Sphaeromatidea, Superfamilies Seroloidea and Sphaeromatoidea
Figure 18.
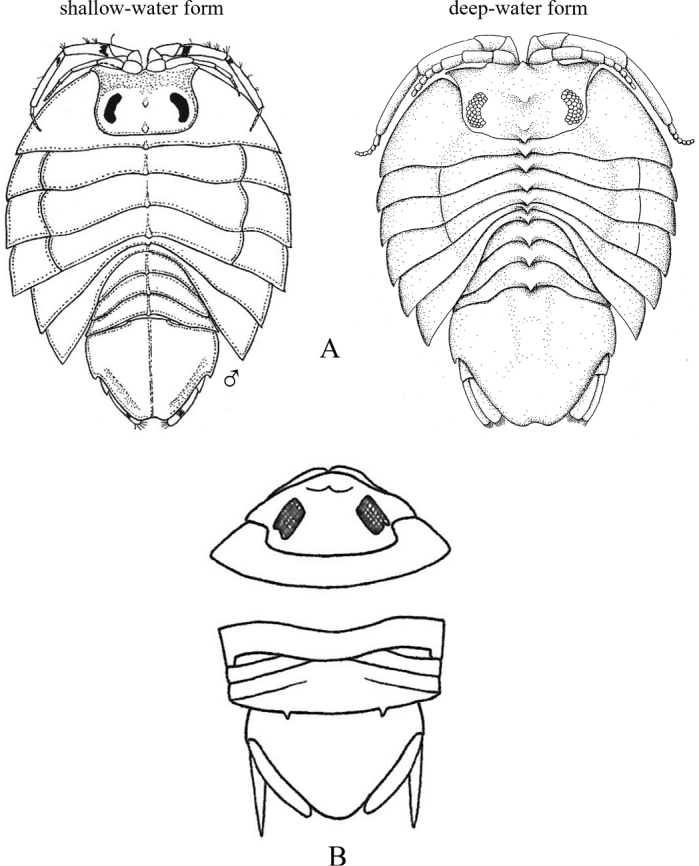
Isopoda, Sphaeromatidea, Seroloidea, Serolidae: AHeteroseroliscarinata (shallow-water form after Menzies and Barnard 1959; deep-water form after Wetzer and Brusca 1997). Tecticipitidae: BTecticepsconvexus (after Richardson 1905a; Schultz 1969).
Figure 23.

Isopoda, Sphaeromatidea, Sphaeromatoidea, Sphaeromatidae (in part): AExosphaeromaamplicauda (after Wall et al. 2015) BExosphaeromaaphrodita (after Wall et al. 2015) CExosphaeromainornata (after Iverson 1982) DExosphaeromapentcheffi (after Wall et al. 2015) EExosphaeromarhomburum (after Richardson 1899, 1905a).
| 1 | Pleon composed of 3 free pleonites plus pleotelson; body broad, depressed and platter-like; dorsum with distinct medial carinae (Fig. 18A)Endnote 13 | Heteroseroliscarinata [Serolidae] |
| – | Pleon composed of 1 or 2 dorsally visible free pleonites plus pleotelson | 2 |
| 2 | Pereopod 1 subchelate in both sexes, with broadly expanded propodus and prehensile dactylus (propodus at least 5 × wider than dactylus); pereopod 2 prehensile only in males | 3 |
| – | Pereopod 1 ambulatory or only weakly prehensile, propodus narrow (propodus < 2 × as wide as dactylus) | 6 [Sphaeromatidae] |
| 3 | Head medially fused to pereonite 1; pleopod 5 with both rami lacking transverse pleats or folds; uropods uniramous, lacking exopods | 4 [Ancindae] |
| – | Head and pereonite 1 not fused; both rami of pleopod 5 with transverse pleats or folds; uropods biramous, exopods slender and spine-like; first segment of pleon with 3 suture lines (fused pleonites 1–4) and 2 small triangular processes on posterior margin; pleotelson with rounded posterior border (Fig. 18B) | Tecticepsconvexus [Tecticipitidae] |
| 4 | Lateral margins of head strongly produced; pleon with 2 short posterior projections overhanging anterior margin of pleotelson; pleopod 1 biramous; uropodal ramus narrow proximally, then expands at least twofold for ~ 80% of length before tapering to acute point (Fig. 19C) | Bathycopeadaltonae |
| – | Lateral margins of head weakly produced; pleon without posterior projections; pleopod 1 uniramous; uropodal ramus styliform, widest proximally and tapering to an acute point | 5 |
| 5 | Body broad (L:W ratio ~ 1.7) with densely granulated surfaces; pereonite 1 narrower than pereonites 2–7; pleotelson inflated, distinctly wider than long with truncate apex; eyes slightly elevated on swellings; uropods strongly recurved distally (Fig. 19A)Endnote 14 | Ancinusgranulatus |
| – | Body relatively narrow (L:W ratio > 2.0) with smooth surfaces; pereonite 1 wider than pereonites 2–7; pleotelson not inflated, longer than wide with nearly acute or narrowly rounded apex; eyes not elevated; distal tips of uropods only slightly recurved (Fig. 19B)Endnote 14 | Ancinusseticomvus |
| 6 | Endopods of pleopods 4 and 5 without branchial pleats or folds | 7 |
| – | Endopods of pleopods 4 and 5 with branchial pleats or folds | 8 |
| 7 | In lateral view, anterior margins of coxal plates of pereonites 2–4 appear raised and posterior margins not raised, giving the coxae a somewhat S-shaped appearance; species is fully marine (Fig. 20B)Endnote 15 | Gnorimosphaeromaoregonense |
| – | In lateral view, anterior margins of coxal plates 2–4 are not raised, and coxae do not appear S-shaped; species occurs in brackish or freshwater habitats (Fig. 20A) | Gnorimosphaeromanoblei |
| 8 | Pleopods 4 and 5 with branchial pleats on both rami | 9 |
| – | Pleopods 4 and 5 with branchial pleats only on endopods | 19 |
| 9 | Uropods of male highly modified, each composed of elongated, cylindrical, or flattened exopod and very short endopod fused to the protopod; both uropodal rami lamellar in females; ovigerous females with 4 pairs of oostegites on pereonites 1–4 | 10 |
| – | Uropods lamellar in both sexes; ovigerous females with 3 pairs, 1 pair, or no pairs of oostegites | 14 |
| 10 | Pleotelson of males with pronounced posteromedial tooth that completely fills apical notch and extends posteriorly beyond level of notch opening; dorsum of pleotelson with 3 transverse elevations at base, median elevation terminating in a spine; body surface densely granulated; sexual dimorphism pronounced with female body smooth, lacking ornamentation or setae (Fig. 20C) | Discerceisgranulosa |
| – | Pleotelson of males with highly complex and open apical notch, internal lateral margins of notch with various numbers of teeth forming sinuses of different shapes; body surface not densely granulated; sexual dimorphism pronounced, females without complex pleotelsonic notch, terminal notch either very short and simple or absent; dorsum of pleotelson sculptured with various types of tubercles | 11 |
| 11 | Uropods of male with ventrolateral spines on exopods; female pleotelson stout, with 4 dorsal tubercles and wide, but shallow apical notch (Fig. 21A) | Paracerceiscordata |
| – | Uropods of male without spines; female pleotelson not as above | 12 |
| 12 | Pleotelson of males long, subequal in length to pereon, with complex medial sinus formed by 2 pairs of teeth, pleotelsonic sinus expanded basally into round foramen overhung by basal knob bearing tall acute spine (spine length ≥ 4 × diameter), sinus then narrowing distally to long thin channel; female pleotelson with small distal notch and 5 dorsal tubercles (1 large medial, 4 smaller lateral); female uropodal exopods with sharply toothed posteromedial margins (Fig. 21D) | Paracerceis sp. A |
| – | Pleotelson of males relatively short, ca. half length of pereon; pleotelsonic sinus not as above, without round basal foramen, with or without short spine overhanging base of notch; female uropodal exopods without sharply toothed margins | 13 |
| 13 | Male pleotelson with short acute spine (i.e., length subequal to diameter) on basal knob overhanging sinus, interior margins of sinus with 2 pairs of lateral teeth; pleotelson of female elongate, acuminate posteriorly, with 3 dorsal tubercles (Fig. 21C)Endnote 16 | Paracerceissculpta |
| – | Male pleotelson with large dorsal tubercle at base of sinus, interior margins of sinus with 3 pairs of sharp teeth; female pleotelson not as above (Fig. 21B) | Paracerceisgilliana |
| 14 | Pleotelson apex entire, upturned; dorsal surface of pleotelson with 2 pairs of prominent tubercles and numerous scattered small tubercles; female sculpturing less pronounced than males (see Bruce and Wetzer 2008: fig. 1) | Pseudosphaeroma sp. |
| – | Pleotelson without upturned apex, sculpturing not as above | 15 |
| 15 | Pleotelson of males with deeply slit apical notch or expanded foramen, notch may be reduced to small depression or dorsally visible slit in females; uropodal rami subequal in length, typically extending beyond posterior margin of pleotelson; ovigerous females with 1 or 3 pairs of oostegites | 16 |
| – | Pleotelson of males and females with shallow terminal notch; uropodal exopod shorter than endopod, rami not extending beyond posterior border of pleotelson; ovigerous females without oostegites | 17 |
| 16 | Pleotelsonic sinus (foramen) distinctly heart shaped with median point in males, but greatly reduced in females; dorsum of pleon with 2 rounded submedian tubercles just lateral to midline on pleonite 5, and 4 similarly spaced tubercles on pleotelson; uropodal rami with crenulated margins (at least in males) (Fig. 22B) | Paradelladianae |
| – | Pleotelsonic sinus long and narrow, with prominent rounded tubercle barely overhanging anterior base, sinus walls straight sided and finely crenulate; dorsal surface of pleotelson covered with small tubercles; uropodal rami without crenulated margins (Fig. 22A) | Dynoideselegans |
| 17 | Frontal margin of head produced as a quadrangular process; antennular articles 1 and 2 dilated; dorsal surface of pleotelson sculptured with 3 longitudinal ridges (Fig. 22C)Endnote 17 | Dynamenelladilatata |
| – | Frontal margin of head not produced; antennular articles not dilated; dorsal surface of pleotelson smooth or tuberculate | 18 |
| 18 | Dorsal surface of pleotelson with many tubercles (Fig. 22E) | Dynamenellasheareri |
| – | Dorsal surface of pleotelson smooth, without tubercles or ridges (Fig. 22D)Endnote 17 | Dynamenellaglabra |
| 19 | Uropodal exopods with distinctly serrated outer margins | 20 |
| – | Uropodal exopods with smooth or lightly crenulated outer margins | 21 |
| 20 | Dorsal surface of pleotelson lightly sculptured with 2 longitudinal rows of small tubercles, posterior margin with prominent transverse elevation (Fig. 20D) | Sphaeromaquoianum |
| – | Dorsal surface of pleotelson heavily sculptured with many longitudinal rows of small to large heavy tubercles, posterior margin without prominent transverse elevation (Fig. 20E) | Sphaeromawalkeri |
| 21 | Dorsal surface of pereonites, pleonites and pleotelson smooth, without tubercles or other ornamentation; apex of pleotelson rounded and truncate (Fig. 23C)Endnote 18 | Exosphaeromainornata |
| – | Dorsal surface of body with ornamentation; apex of pleotelson acuminateEndnote 18 | 22 |
| 22 | Pleotelson with produced rhomboid-shaped apical process and concave lateral margins, sides folded inward, forming kind of funnel-like opening when viewed ventrally; dorsum of pleotelson sculptured with 2 basal tubercles, which are continuous with 2 longitudinal ridges that converge posteriorly into a single median ridge; uropods distinctly shorter than pleotelson (Fig. 23E)Endnote 19 | Exosphaeromarhomburum |
| – | Pleotelson triangular, dorsum without parallel ridges; uropods subequal in length to pleotelson | 23 |
| 23 | Dorsal sculpturing subtle, starting on pereonites 6 and 7; pereonite 6 with row of 7 weak tubercles along posterior margin; pereonite 7 with posteriorly directed median process and weak lateral tubercle on each side along posterior margin; pleon with 2 medium tubercles, 1 on each side of midline; pleotelson with 1 strong anteromedial tubercle and 2 weak tubercles lateral to midline (Fig. 23B) | Exosphaeromaaphrodita |
| – | Dorsal sculpturing distinct on most or all pereonites, comprising either low, weak tubercles or taller, more pronounced tubercles | 24 |
| 24 | Dorsal surface of pereonites with low, weak tubercles along posterior margins; pereonite 1 with 1 very weak median tubercle; pereonites 2–4 with 3 tubercles, 1 median and 2 laterals (1 per side); pereonites 5 and 6 with row of 7 tubercles, 1 at midline and 3 laterals per side; pereonite 7 with weak median process and paired lateral tubercles; pleotelson with 2 small anterior tubercles (Fig. 23A) | Exosphaeromaamplicauda |
| – | Dorsal surface of all pereonites sculptured with heavy, strong tubercles giving body appearance of 7 prominent longitudinal rows of tubercles; pereonite 1 with 2 transverse rows of tubercles; pereonites 2–7 each with 1 transverse row of 7 tubercles; pereonite 7 with strong median process and 3 lateral tubercles per side; pleotelson with 3 strong medial tubercles on either side of midline, plus numerous smaller tubercles scattered over dorsal surface (Fig. 23D) | Exosphaeromapentcheffi |
Figure 19.

Isopoda, Sphaeromatidea, Sphaeromatoidea, Ancinidae: AAncinusgranulatus (after Holmes and Gay 1909) BAncinusseticomvus (after Iverson 1982) CBathycopeadaltonae (after Menzies and Barnard 1959).
Figure 20.

Isopoda, Sphaeromatidea, Sphaeromatoidea, Sphaeromatidae (in part): AGnorimosphaeromanoblei (after Menzies 1954) BGnorimosphaeromaoregonenseB1 dorsal view (after Menzies 1954) B2 lateral view of pereonites 1–7 showing S-shaped coxae of anterior pereonites (e.g., arrow) CDiscerceisgranulosa (after Richardson 1899, 1905a) DSphaeromaquoianum (after Harrison and Holdich 1984) ESphaeromawalkeri (after Kensley and Schotte 1989).
Figure 21.

Isopoda, Sphaeromatidea, Sphaeromatoidea, Sphaeromatidae (in part): AParacerceiscordata (after Schultz 1969) BParacerceisgilliana (after Allen 1976) CParacerceissculpta (after Brusca 1980) DParacerceis sp. A (after Brusca 1980).
Figure 22.

Isopoda, Sphaeromatidea, Sphaeromatoidea, Sphaeromatidae (in part): ADynoideselegans (after Wetzer and Mowery 2017) BParadelladianae (after Harrison and Holdich 1982) CDynamenelladilatata (after Richardson 1899, 1905a) DDynamenellaglabra (after Richardson 1899, 1905a) EDynamenellasheareri (after Brusca et al. 2007).
Key H. Suborder Valvifera
Figure 24.

Comparison of maxillipeds and number of articles in maxillipedal palps (3–5) between genera and families of SCB valviferan isopods: A 3-article maxillipedal palps B 4-article maxillipedal palps C 5-article maxillipedal palps.
Figure 30.

Isopoda, Valvifera, Idoteidae (in part): APentidoteaaculeata (after Stafford 1913b) BPentidoteakirchanskii (after Miller and Lee 1970) CPentidoteamontereyensis (after Menzies 1950a) DPentidotearesecata (after Menzies 1950a) EPentidoteaschmitti (after Menzies 1950a) FPentidoteastenops (after Brusca and Brusca 1978) GPentidoteawosnesenskii (after Menzies 1950a).
| 1 | Eyes absent; head, pereonites and pleonites with transverse rows of submedian, sublateral and lateral tubercles bearing bead-like spheres; pleotelson with short oblique lateral tubercles; maxillipedal palp 4-segmented (Figs 24B, 25D) | Califarcturustannerensis [Thermoarcturidae] |
| – | Eyes present; body not as above | 2 |
| 2 | Head fused with pereonite 1; pereonite 4 much longer (> 2 ×) than other pereonites; pereonite 4 much broader anteriorly than posteriorly in females; pereopods 1–4 slender, fringed with setae, directed anteriorly against ventral body wall (modified for filter feeding); pereopods 5–7 stout, prehensile; maxillipedal palp 5-segmented (Figs 24C, 25A–C) | 3 [Arcturidae] |
| – | Head distinct from pereonite 1; length of pereonite 4 subequal to other pereonites; anterior pereopods not modified for filter feeding, mostly ambulatory and similar in size | 6 |
| 3 | Head incompletely fused with pereonite 1, indicated by distinct lateral incision posterior and ventral to eye (must be viewed laterally); flagellum of antennae with ventral blade-like setae; dorsum of pereonite 4 smooth in males or with medial, bilobed swelling or tubercle in females; dorsum of other pereonites mostly smooth; pleon composed of 3 segments, including 2 pleonites plus the pleotelson (Fig. 25C) | Neastacillacalifornica |
| – | Head completely fused with pereonite 1, lateral margins entire; flagellum of antennae without blade-like setae; dorsum of pereonites 2–7 typically with 2–4 small to large dorsal spines in females or smooth in males; pleon composed of a single fused pleotelson with 2 anterodorsal median spines | 4 |
| 4 | Fused head and pereonite 1 with shallow dorsal transverse groove (fusion line) posterior to eyes; antennae robust with peduncular articles 3 and 4 each ~ 5 × longer than wide; pereonites 2–7 with large, broadly triangulate anterolateral or lateral extensions and a transverse row of 4 large posterodorsal spines; pereonites 4–6 also with medium to large posterolateral spines (Fig. 25B)Endnote 20 | Idarcturushedgpethi |
| – | Fused head and pereonite 1 without dorsal transverse groove; antennae relatively long and narrow with peduncular articles 3 and 4 each > 10 × longer than wide; lateral extensions of pereonites 2–7 not as above, and each one with pair of medial posterodorsal spines (size variable); pereonites 5–7 also with 2 smaller distinct lateral spines | 5 |
| 5 | Body color typically golden; pereonites 2–4 anterolateral extensions medium in size, broad at base and distally blunt or obtusely rounded; pereonites 5–7 lateral extensions broadly triangular; anterolateral margins of head and antennal peduncular article 1 acute, but not produced forward as recurved spines (Fig. 25A)Endnote 20 | Idarcturusallelomorphus |
| – | Body color whitish; pereonites 2–7 anterolateral or lateral extensions narrowly triangular and sharply acute; anterolateral margins of head and peduncular article 1 of antennae distinctly produced forward as sharply acute recurved spinesEndnote 20 | Idarcturus sp. A |
| 6 | Pereopod 4 greatly reduced, non-ambulatory; pleon composed of 4 segments with pair of elevated dorsal humps near posterior end; lateral margins of body with dense tufts of setae (Fig. 25E) | Cleantioidesoccidentalis [Holognathidae] |
| – | Pereopod 4 not reduced, all pereopods ambulatory; pleon composed of 1 or 3 segments; dorsal surface of pleon and lateral margins of body not as above | 7 [Idoteidae] |
| 7 | Pleon composed of single fused pleotelson, with or without partial suture lines or indentations | 8 |
| – | Pleon composed of 3 to 4 segments, including 1 pair of partial sutures | 19 |
| 8 | Pleotelson without suture lines | 9 |
| – | Pleotelson with 1 pair of partial suture lines or indentations | 10 |
| 9 | Flagellum of antennae multiarticulate, head with distinct, slightly elongated dorsal tubercle; pereon dorsally smooth (Fig. 26A) | Stenosomawetzerae |
| – | Flagellum of antennae uniarticulate, flagellar article large and clavate; head with large, multilobed or conical tubercle or elevation; pereonites 1–5 with (female) or without (male) middorsal spines (Fig. 26B) | Erichsonellacrenulata |
| 10 | Antennae shorter than or subequal to antennules, antennal flagellum reduced to single vestigial article; maxillipedal palp of 3 articles; pleonal fusion indicated by 1 pair of lateral grooves instead of distinct incisions | 11 |
| – | Antennae much longer than antennules, antennal flagellum multiarticulate or comprising a single, large clavate article; maxillipedal palp of 3 or 4 articles; pleotelson with distinct lateral incisions | 12 |
| 11 | Pleotelson with dorsal transverse ridge ca. midway between lateral insertions and posterior border, without dorsal swellings; lateral margins of pereonite 4 in females angular, but not forming acute posterolateral projections; propodi of pereopods with 2 large proximal spines along inferior margins (Fig. 27B)Endnote 21 | Edotia sp. B |
| – | Pleotelson inflated or bulbous with large dorsal swellings, without a dorsal transverse ridge; lateral margins of pereonite 4 in females form acute posterolateral projections; propodi of pereopods without large proximal spines along inferior margins (Fig. 27A) | Edotiasublittoralis |
| 12 | Flagellum of antennae uniarticulate, flagellar article large and clavate; head with large, median bifid tubercle projecting anteriorly over frontal margin; anterolateral margins of pereonite 1 produced into bilobed processes (Fig. 26C) | Eusymmeruspseudoculata |
| – | Antennal flagellum multiarticulate; head and pereonite 1 not as above | 13 |
| 13 | Maxillipedal palp 4-segmented (Fig. 24B) | 14 |
| – | Maxillipedal palp 3-segmented (Fig. 24A) | 16 |
| 14 | Eyes large, oval to subpyriform in shape, situated at mid-lateral margins of head; lateral margins of pleotelson flare slightly outward to form obtuse posterolateral angles, after which they taper inwards to strongly acuminate apex; coxal plates of pereonites not visible dorsally (Fig. 27C) | Colidoteafindleyi |
| – | Eyes small or large, situated laterally on head adjacent to anterolateral angles of pereonite 1; pleotelson not as above, with rounded or obtuse apex; coxal plates of some posterior pereonites visible dorsally | 15 |
| 15 | Eyes large; head not deeply immersed in pereonite 1, dorsal surface with a large, medial, apically rounded rostrum-like process; lateral margins of pereon convex, widest around pereonite 4; coxal plates visible dorsally on pereonites 5–7 (or just pereonites 6 and 7); commensal on sea urchins Strongylocentrotuspurpuratus and Mesocentrotusfranciscanus with body of live isopods matching purple or dark red color of host urchin, but fading to bluish gray in alcohol (Fig. 27D) | Colidotearostrata |
| – | Eyes very small; head deeply immersed in pereonite 1, without a large dorsal process; lateral margins of pereon not convex, generally straight-sided from pereonites 1–4 and then becoming slightly wider from pereonites 5–7; coxal plates visible dorsally on pereonites 4–7 (Fig. 27E) | Colidoteawallersteini |
| 16 | Body smooth, but with triangulate anterior median dorsal pattern on pereonites 2–4; lateral margins of pereonites rounded to straight; posterior margin of pleotelson emarginate (Fig. 28B) | Synidoteaharfordi |
| – | Body sculptured with conspicuous tubercles or longitudinal rugae; lateral margins of pereonites angular; pleotelson spatulate, rounded posteriorlyEndnote 22 | 17 |
| 17 | Lateral margins of adult body roughly parallel, widest part of pereon subequal in width to pleon; pleotelson widest at midpoint; dorsal sculpturing generally reduced to low, conical tubercles on head and medial row of tubercles along pereonites; eyes small and lightly pigmented (Fig. 28A) | Synidoteacalcarea |
| – | Lateral margins of adult body not parallel, generally widest at pereonites 3 and 4; pleotelson widest anteriorly; dorsal sculpturing variable, pereonites with 3 to 4 longitudinal rugae on lateral areas; eyes large and heavily pigmented | 18 |
| 18 | Flange present on basis of pereopods 2–6 in adults, but tiny and difficult to see or not visible in juveniles; anteromedial tubercles of head generally large, highly variable with size, often asymmetrical, becoming broad, flattened, and forward projecting in large specimens; coxae of pereonite 1 not notched laterally; body surface appearing rough, color brownish (Fig. 28C) | Synidoteamagnifica |
| – | Flange absent on basis of pereopods; anteromedial tubercles not as above, typically smaller, narrowly rounded or conical; lateral margins of pereonite 1 deeply notched in lateral view, separating coxal margins into upper and lower lobes; body surface generally smooth and whitish (Fig. 28D) | Synidoteamedia |
| 19 | Maxillipedal palp 4-segmented in adults, article 4 much larger than article 3 (Fig. 24B)Endnote 23 | 20 |
| – | Maxillipedal palp 5-segmented in adults, article 5 much smaller than article 4 (article 5 may not be developed in juveniles) (Fig. 24C) | 23 |
| 20 | Eyes large, occupying almost entire lateral margins of head; dorsum of head with curved, sharply defined posterior groove; coxal plates of pereonites 2–7 all wide and extending entire length of respective pereonites; pleotelson broadest anteriorly, tapering to truncate posterior border or apex (Fig. 29B) | Idoteametallica |
| – | Eyes small, situated mid-laterally on head; coxal plates of pereonites narrow; pleotelson with concave or acuminate posterior border or apex | 21 |
| 21 | Posterior margin of pleotelson concave with rounded posterolateral corners; coxae 5–7 reach posterior edges of their respective pereonites (Fig. 29C) | Idotearufescens |
| – | Posterior margin of pleotelson acuminate, with or without distinct median projection; only coxae 6 and 7 or just 7 reach posterior edges of respective pereonites | 22 |
| 22 | Posterior margin of pleotelson triangular, converging to obtuse point; coxae 6 and 7 reach posterior edges of their pereonites; head with apically blunt frontal process (Fig. 29D) | Idoteaurotoma |
| – | Posterior margin of pleotelson with an elongate median projection; only coxae 7 reach posterior edge of pereonite 7; head with thin, pointed frontal process (Fig. 29A) | Idoteafewkesi |
| 23 | Posterior margin of pleotelson deeply concave with acute posterolateral corners (Fig. 30D) | Pentidotearesecata |
| – | Posterior margin of pleotelson not concave | 24 |
| 24 | Coxae visible dorsally on pereonites 5–7, and occasionally on pereonite 4; posterior margin of pleotelson convex, without median projection; body narrow, with head, pereon and pleotelson subequal in width; apex of frontal process with median notch (Fig. 30B) | Pentidoteakirchanskii |
| – | Coxae visible dorsally on pereonites 2–7; posterior margin of pleotelson with short (sometimes poorly developed) to elongate median projection; frontal process with or without apical notch | 25 |
| 25 | Pleonite 1 lateral margins curve and taper posteriorly to form narrow, acutely pointed posterolateral angles (margins not parallel) | 26 |
| – | Pleonite 1 lateral margins truncate or convex, more-or-less parallel, not curving posteriorly to form acute points | 27 |
| 26 | Pleotelson lateral margins concave, posterior margin with median projection; pereonites widely separated laterally, coxae 2–7 reaching posterior edges of pereonites, anterior margins of pereonite 1 separated from head; eyes rectangular or pyriform; apex of frontal process entire, without notch (Fig. 30E) | Pentidoteaschmitti |
| – | Pleotelson lateral margins convex, broadly rounded, posterior margin with small median projection; pereonites not widely separated laterally, only coxae 5–7 reach posterior edges of pereonites; anterior margins of pereonite 1 flush with head; eyes reniform; apex of frontal process entire, without notch (Fig. 30G) | Pentidoteawosnesenskii |
| 27 | Eyes elongated and thin dorsoventrally; maxilliped with 1–3 coupling hooks; coxae of pereonites 2–7 contiguous with each other (Fig. 30F) | Pentidoteastenops |
| – | Eyes not elongated; maxilliped with 1 coupling hook; coxae of at least some pereonites not contiguous with each other | 28 |
| 28 | Pleotelson posterior margin with long median projection; lateral margins of pereonite 1 convex; eyes circular to oval; apex of frontal process notched (Fig. 30A) | Pentidoteaaculeata |
| – | Pleotelson posterior margin with small or poorly developed median projection, sometimes appearing slightly truncate; lateral margins of pereonite 1 barely rounded, almost parallel; eyes with straight anterior and convex posterior borders; apex of frontal process entire, without notch (Fig. 30C) | Pentidoteamontereyensis |
Figure 25.

Isopoda, Valvifera, Arcturidae: AIdarcturusallelomorphus (after Menzies and Barnard 1959) BIdarcturushedgpethi (after Menzies 1951b) CNeastacillacalifornica (after Menzies and Barnard 1959). Thermoarcturidae: DCalifarcturustannerensis (after Schultz 1966). Holognathidae: ECleantioidesoccidentalisE1 dorsal view (after Brusca and Wallerstein 1979a) E2 lateral view of C.verecundus from Florida showing reduced size of pereopod 4 characteristic of Cleantioides (after Kensley and Clark 1998).
Figure 26.

Isopoda, Valvifera, Idoteidae (in part): AStenosomawetzerae (after Ormsby 1991) BErichsonellacrenulata (after Menzies 1950b) CEusymmeruspseudoculata (after Menzies and Bowman 1956).
Figure 27.

Isopoda, Valvifera, Idoteidae (in part): AEdotiasublittoralis (after Menzies and Barnard 1959) BEdotia sp. B (after Stebbins 2012c) CColidoteafindleyi (after Brusca and Wallerstein 1977) DColidotearostrata (after Allen 1976) EColidoteawallersteini (after Brusca 1983b).
Figure 28.

Isopoda, Valvifera, Idoteidae (in part): ASynidoteacalcarea (after Schultz 1966) BSynidoteaharfordi (after Menzies and Miller 1972) CSynidoteamagnifica (after Menzies and Barnard 1959) DSynidoteamedia (after Iverson 1972).
Figure 29.

Isopoda, Valvifera, Idoteidae (in part): AIdoteafewkesi (after Menzies 1950a) BIdoteametallica (after Brusca and Wilson 1991) CIdotearufescens (after Menzies 1950a) DIdoteaurotoma (after Menzies 1950a).
Key I. Suborder Asellota, Superfamilies Janiroidea and Stenetrioidea
Figure 31.

Isopoda, Asellota, Janiroidea, Dendrotionidae: AAcanthomunnatannerensis (after Schultz 1966). Janiridae (in part): BIaiscalifornica (after Menzies and Barnard 1951). Stenetrioidea, Stenetriidae: CStenetrium sp. A C1 pereopod 1 (after Wilson 1997).
Figure 41.

Isopoda, Asellota, Janiroidea, Paramunnidae (in part): AParamunnaquadratifrons (after Iverson and Wilson 1981) BParamunna sp. A (after Cadien 1996) CPleurogoniumcalifornienseC1 antennule (after Wilson 1997) DPleurogonium sp. A D1 antennule (after Wilson 1997).
| 1 | Eyes present | 2 |
| – | Eyes absent | 36 |
| 2 | Body (except head) covered with large, jointed-appearing spines; uropods very long if not broken off, with basal segments and rami each ca. as long as width of pleotelson (Fig. 31A) | Acanthomunnatannerensis [Dendrotionidae] |
| – | Body and uropods not as above | 3 |
| 3 | Eyes sessile, situated dorsolaterally on head but not reaching lateral margins; pleotelson subequal in width to cephalon and most pereonites | 4 |
| – | Eyes situated laterally on head, bulging outward on short to elongated ocular lobes or peduncle-like projections; pleotelson distinctly narrower than cephalon and widest pereonites | 24 |
| 4 | Head with rounded anterolateral margins and prominent rounded median lobe between antennae; body widest at ca. pereonite 6; adult body length < 3 mm; commensal on or in burrows of sphaeromatid Sphaeromaquoianum in bays and estuaries (Fig. 31B) | Iaiscalifornica [Janiridae, in part] |
| – | Head not as above; body lateral margins parallel or gently convex and slightly widest around mid-pereon; maximum adult length varies between ~ 1.3–7.3 mm | 5 |
| 5 | Body elongated and unpigmented, lateral margins parallel and covered with long simple setae; head with acute anterolateral angles and lightly pigmented eyes; pleotelson shield-like with small posterolateral spines; male pereopod 1 strongly subchelate with thick projection on inferior margin of propodus, and many long simple setae on dactylus, propodus, carpus, and merus (Fig. 31C) | Stenetrium sp. A [Stenetriidae] |
| – | Body, head, and male pereopod 1 not as above | 6 |
| 6 | Uropods short, inserted in subterminal excavations of pleotelson and not extending much beyond its posterior margin; both antennules and antennae small, flagella lacking or rudimentary; peduncle articles of antennae dilated | 7 [Joeropsididae] |
| – | Uropods well-developed, extending well beyond posterior margin of pleotelson; both antennules and antennae relatively long with multiarticulate flagella, although flagella often broken off in preserved specimens; peduncle articles of antennae not dilated | 11 [Janiridae, in part] |
| 7 | Body without pigment, covered with numerous fine simple setae | 8 |
| – | Body typically pigmented, not covered with fine setae | 9 |
| 8 | Body elongated and parallel-sided (L:W ratio ~ 4.3); head with distinct indentation on lateral margins below eyes; basal article of antennules with large, outwardly produced distolateral projection bearing ~ 6 sharp, flat spines curving toward article 2; pleotelson lateral margins with 7 large posteriorly curved spines, rest of body with small denticulations on lateral margins (Fig. 32A) | Joeropsis concava |
| – | Body broad and laterally curved (L:W ratio ~ 2.6); margins of head produced forming large lateral plate, with ~ 9 anteriorly curved spines, eyes not reaching lateral margins; basal article of antennules with small distolateral process bearing several lateral curving flat spines; pleotelson with 8 posteriorly curved spines (Fig. 32E) | Joeropsis sp. A |
| 9 | Lateral margins of pleotelson without spines; body banded in appearance, with dark brown pigment typically covering head, pereonites 2–4 and 7, and pleotelson; maxillipedal palp article 2 not produced medially; anterior margin of frontal lamina entire, without fringe of small scales (Fig. 32D) | Joeropsislobata |
| – | Lateral margins of pleotelson with 3–7 spines; body typically with diffuse pigment, which may be most pronounced on head and pereonite 4; maxillipedal palp article 2 produced medially; anterior margin of frontal lamina with fringe of small scales | 10 |
| 10 | Lateral margins of pleotelson with 5–7 spines; head typically heavily pigmented, pereon and pleon lightly pigmented or without pigments (Fig. 32B) | Joeropsisdubiadubia |
| – | Lateral margins of pleotelson with 2 or 3 spines; head and pereonite 4 both typically heavily pigmented but compare pigment pattern of head with that of J.dubiadubia above (Fig. 32C) | Joeropsisdubiapaucispinis |
| 11 | Pereopod 1 with conspicuous serrations or denticles on proximal third of inferior margin of propodus; maxillipedal palp articles 1–3 as wide as endite | 12 |
| – | Pereopod 1 without denticles on proximal third of inferior margin of propodus; maxillipedal palp articles 2 and 3 much wider than endite | 19 |
| 12 | Pleotelson with distinct distolateral angles or spines; head with pointed anterolateral margins and distinct rostrum | 13 |
| – | Pleotelson without distolateral spines, posterolateral margins rounded; head with pointed or rounded anterolateral margins, with or without rostrum | 16 |
| 13 | Adults with acutely pointed rostrum extending well beyond anterior limits of anterolateral margins of head; body completely devoid of pigment (Fig. 33F) | Janiralata sp. B |
| – | Adults with rostrum not extending anteriorly beyond anterolateral margins of head; body pigmented | 14 |
| 14 | Head with acute rostral point; posterior margin of pleotelson concave laterally with convex median lobe that does not extend beyond posterior limits of distolateral spines; species lives commensally on sun stars (Solaster spp.), and its occurrence in southern California is questionable (Fig. 33D) | Janiralatasolasteri |
| – | Head with obtuse rostral point; posterior margin of pleotelson with convex lateral areas and obtusely rounded median point that extends posteriorly distinctly beyond limits of distolateral spines | 15 |
| 15 | Body covered with diffuse, anastomosing chromatophores, with pigment extending into region anterior to eyes (Fig. 33E) | Janiralata sp. A |
| – | Body covered with dense, non-anastomosing chromatophores that are absent from region anterior to eyes (Fig. 33B) | Janiralataoccidentalis |
| 16 | Head with angular anterolateral margins and short, acute median lobe or slightly rounded median point; eyes medium to large, dorsal, clearly separated from lateral margins of head; body relatively short and broad (L:W ratio < 2.5); pleotelson nearly 2 × as wide as long | 17 |
| – | Head with rounded anterolateral margins, without distinct median lobe or rostrum; eyes large, almost touching lateral margins of head; body long and narrow (L:W ratio > 3.0); pleotelson ca. as wide as long | 18 |
| 17 | Dorsal surface with numerous black chromatophores; pereonites 1–3 with distinct lateral lobes or lappets, each pair separated by narrow V-shaped slit; all lateral margins with conspicuous setal fringe; posterolateral margins of pereonites 5–7 angular; pereonite 5 not shorter medially than other pereonites (Fig. 33A) | Janiralatadavisi |
| – | Dorsal surface without pigment, but with many fine setae; pereonites 2 and 3 with short anterior lappets and indistinct posterior lappets; lateral margins without conspicuous setal fringe; posterolateral margins of pereonites 5–7 rounded; pereonite 5 distinctly shorter medially than other pereonites (Fig. 33G) | Janiralata sp. C |
| 18 | Body whitish, without distinct chromatophores; pereopod 1 propodus with ~ 12 or 13 denticles on proximoventral border (Fig. 33C)Endnote 24 | Janiralatarajata |
| – | Body pigmented with numerous dense chromatophores; pereopod 1 propodus with ~ 18 denticles on proximoventral border (Fig. 33H)Endnote 24 | Janiralata sp. D |
| 19 | Lateral margins of pleotelson with spine-like serrations | 20 |
| – | Lateral margins of pleotelson without serrations, although setae may be present | 21 |
| 20 | Pleotelson with 4–7 spine-like serrations on each side (Fig. 34A) | Ianiropsisanaloga |
| – | Pleotelson with 3 spine-like serrations on each side (Fig. 34F) | Ianiropsistridens |
| 21 | Pleotelson with distinct posterolateral angles extended beyond uropodal insertions (Fig. 34B) | Ianiropsisderjugini |
| – | Pleotelson without posterolateral angles, posterior margins rounded | 22 |
| 22 | Uropods relatively short, not exceeding half-length of pleotelson; frontal margin of head truncate (Fig. 34D) | Ianiropsisminuta |
| – | Uropods much longer than half-length of pleotelson; frontal margin of head with short pointed or blunt medial lobe | 23 |
| 23 | Uropods very long, exceeding length of pleotelson; lateral apices of male pleopod 1 bifurcate (Fig. 34E) | Ianiropsismontereyensis |
| – | Uropods not exceeding length of pleotelson; lateral apices of male pleopod 1 not bifurcate (Fig. 34C) | Ianiropsiskincaidi |
| 24 | Pereonites 1–3, 5 and 6 with long, finger-like lateral processes, each with 1 or 2 terminal setae; pleotelson bulbous anteriorly, then tapers posteriorly to a narrow truncate end; uropods inserted dorsolaterally on anterior half of pleotelson, each uropod ca. as long as pleotelson; eyes tiny, each composed of two lenses at tip of very long and thin lateral peduncle or lobe (Fig. 35E)Endnote 25 | Pleurocope sp. A [Pleurocopidae] |
| – | Pereonites, pleotelson and uropods not as above; eyes situated at tips of short to elongated ocular lobes | 25 |
| 25 | Antennules and antennae both long, subequal in length; frontal margin of head broadly rounded | 26 [Paramunnidae, in part] |
| – | Antennae > 2 × longer than antennules; frontal margin of head broadly truncate, with or without medial concavity | 29 [Munnidae] |
| 26 | Pereonites 1–3 with dorsal posterolateral setal tufts; pleotelson as wide or wider than pereonite 7, lateral margins of pleotelson serrate or denticulate anterior to uropodal insertions, denticulations with embedded compound setae in cusps (Fig. 35B) | Boreosignum sp. A |
| – | Pereonites without dorsal setal tufts; pleotelson narrower than pereonite 7, lateral margins convex without lateral serrations or denticulations | 27 |
| 27 | Body broadly ovate with pereonites 2 and 3 distinctly wider than pereonites 4 and 5 (body L:W ratio ~ 1.6); head deeply immersed in pereon with pereonite 1 much longer (male) or subequal (female) in length to pereonite 2; eyestalk (ocular lobe) much longer than article 1 of antennules; coxae visible in dorsal view only on pereonites 5–7 (Fig. 35A) | Austrosignumlatum |
| – | Body narrower than above with pereonites 2–5 subequal in width (body L:W ratio > 2.0); head not deeply immersed in pereon, with pereonite 1 shorter than pereonite 2 in both sexes; ocular lobes much shorter than article 1 of antennules; coxae visible in dorsal view on at least pereonites 2–7 | 28 |
| 28 | Article 1 of antennules shorter than article 2; pereopod 2 in males with robust subtriangular projection on distoventral corner of basis, and proximal projection on ischium; pereopod 2 ventral margins of propodi with short robust setae (Fig. 35C)Endnote 26 | Munnogoniumerratum |
| – | Article 1 of antennules subequal in length or longer than article 2; pereopod 2 in males not as above, without distinct projections on basis and ischium; pereopod 2 ventral margins of propodi with simple setae only (Fig. 35D)Endnote 26 | Munnogoniumtillerae |
| 29 | Pleotelson with 4–6 spines or serrations on ventrolateral margins; male pleopod 1 apically pointed (Fig. 36H) | Uromunnaubiquita |
| – | Pleotelson without spines on lateral margins; male pleopod 1 with apices laterally expanded | 30 |
| 30 | Pleotelson with concave posterolateral margins; posterior border of pereonite 7 convex (Fig. 36D) | Munnamagnifica |
| – | Posterolateral margins of pleotelson not concave, although small indentations may be present around uropodal insertions; pereonite 7 with deeply concave posterior border wrapping around anterior portion of pleotelson | 31 |
| 31 | Uropods with large acute spine-like protuberances on distal margins; pleotelson posterior margin without subanal channel or shelf visible in dorsal view | 32 |
| – | Uropods without spine-like protuberances on distal margins; pleotelson with or without subanal shelf visible in dorsal view | 33 |
| 32 | Lateral margins of pleotelson with 2 or 3 backwards-curved robust sensory setae on each side; pleotelson broad medially, nearly as wide as long (Fig. 36F) | Munnastephenseni |
| – | Lateral margins of pleotelson without robust sensory setae; pleotelson narrow, much longer than wide (Fig. 36A) | Munnachromatocephala |
| 33 | Frontal margin of head with ~ 8 stout, 2-pointed spines; lateral margins of pereonites 1–6 with stout spines; pleotelson without subanal shelf (Fig. 36E) | Munnaspinifrons |
| – | Margins of head and pereonites without spines, although setae may be present; pleotelson with subanal shelf visible in dorsal view | 34 |
| 34 | Eyes small, occupying ca. half of short ocular lobes; body surfaces smooth, not covered with setae; uropods tiny, hidden within short, broad subanal shelf (Fig. 36C) | Munnahalei |
| – | Eyes large, occupying most of prominent ocular lobes; all dorsal surfaces covered with fine simple setae; uropods larger and clearly visible, not hidden by subanal shelf | 35 |
| 35 | Subanal shelf short and broad, reaching but not obscuring uropods; male pleopod 1 with large sensillate setae on ventral surface (Fig. 36B) | Munnafernaldi |
| – | Subanal shelf elongated posteriorly, but no wider than anus and not reaching or extending under uropods; male pleopod 1 with short simple setae on ventral surface (Fig. 36G) | Munna sp. A |
| 36 | Surface of body covered with many small spines and fewer large spines; dorsal surface of pleotelson with pair of large stout posterior protuberances, each with 3 large distal spines (Fig. 37) | Haplomunnacaeca [Haplomunnidae] |
| – | Surface of body and pleotelson not as above | 37 |
| 37 | Pereopods 5–7 expanded and natatory, with enlarged, paddle-like carpi and propodi; posterior body arranged as distinctive natasome, consisting of partially or completely fused pereonites 5–7 and the pleotelson | 38 [Munnopsidae] |
| – | Pereopods 5–7 without enlarged, paddle-like carpi and propodi; posterior part of body without distinct natasome | 43 |
| 38 | Head with bilobed or pointed rostrum projecting anteriorly between antennules | 39 |
| – | Head without rostrum extending between antennules | 40 |
| 39 | Head with broad, indented (bilobed) rostrum; natasome flattened, tapering; pereonites 5 and 6 fused dorsally; pereonite 7 smaller and narrower than pereonites 5 and 6 (Fig. 38A) | Belonectes sp. A |
| – | Head with narrow, roundly pointed rostrum, not bilobed; natasome robust and deep; pereonites 5 and 6 not fused dorsally; pereonite 7 larger than pereonites 5 and 6 (Fig. 38B) | Eurycopecaliforniensis |
| 40 | Natasome not triangular, pleotelson inflated dorsally with relatively straight lateral margins; pereonites 5–7 subequal in size; uropodal protopod small, tubular | 41 |
| – | Natasome triangular, lateral margins of pleotelson taper to pointed apex; pereonite 7 distinctly narrower than pereonites 5 and 6; uropodal protopod large, flattened | 42 |
| 41 | Head subequal in width to pereonite 1; small species, with body length of adults < 2 mm (Fig. 38E) | Munnopsurus sp. A |
| – | Head very large, distinctly broader than pereonite 1; much larger species than above, with body length of adults reaching 7 mm (Fig. 38F) | Munnopsurus sp. B |
| 42 | Anterior margins of pereonites 1–4 with large pedestal setae (Fig. 38C) | Ilyarachnaacarina |
| – | Anterior margins of pereonites 1–4 with small, non-pedestal setae (Fig. 38D) | Ilyarachnaprofunda |
| 43 | Body with conspicuous setal fringe on lateral margins; head trilobate anteriorly; pleotelson wider than long with concave posterior margins at uropodal insertions and blunt median apex; basal article of antennules greatly expanded; article 3 of antennae with elongate, setiferous scale; small intertidal isopods that live in burrows in wood excavated by limnoriid isopods (Fig. 39A) | Caecijaerahorvathi [Janiridae, in part] |
| – | Body and habitat not as above | 44 |
| 44 | Body greatly elongated, > 6 × longer than wide; lateral margins generally parallel with head, pereonal, and pleonal segments subequal in width | 45 |
| – | Body not as long as above, ≤ 4.5 × longer than wide; lateral margins distinctly not parallel | 47 |
| 45 | Antennae with strong scale on peduncular article 3; uropods long and distinct, protopod subequal in length to pleotelson and half as wide, sides parallel until pinching off slightly towards distal medial tip; uropodal endopod ca. one-third length of protopod and attached subapically; uropodal exopod ca. half-length of endopod and attached more laterally near where protopod begins to curve inwards (Fig. 39C)Endnote 27 | Microcharon sp. A [Lepidocharontidae] |
| – | Antennae without scale on peduncle; uropods styliform, not as above (Fig. 39B) [Janiridae, in part] | 46 |
| 46 | Integument of body with complete and prominent scaling; peduncular article 2 of antennules with ≥ 3 strong longitudinal ridges; mandibles whitish, not appearing sclerotized; setal comb present on posteroventral margin of basis of anterior pereopodsEndnote 28 | Caecianiropsis sp. LA1 |
| – | Body with very light and intermittent scaling; peduncle of antennules without longitudinal ridges; mandibles brownish, heavily sclerotized; setal comb absent on posteroventral margins of basis of anterior pereopodsEndnote 28 | Caecianiropsis sp. LA2 |
| 47 | Pereonite 7 fused dorsally with pleotelson; body widest at posterior third of pleotelson; posterior margins of pleotelson with large bilobed medial extension and large tooth on each lateral edge (Fig. 40D) | Nannonisconuslatipleonus [Nannoniscidae] |
| – | All pereonites dorsally free; body widest at anterior half of pereon; pleotelson not as above | 48 |
| 48 | Body long and narrow (L:W ratio ~ 4.5); pereonites 1–3 only slightly wider than head and pleotelson | 49 [Desmosomatidae] |
| – | Body short and broad (L:W ratio ≤ 2.0); pereonite 1 and usually pereonites 2–6 much wider than head and pleotelson | 51 [Paramunnidae, in part] |
| 49 | Pereonite 1 distinctly larger than pereonite 2, with large ventral median spine; posteriorly curving ventral median spines also present on pereonites 3 and 7; pleotelson posteriorly rounded, with large subdistal ventrolateral spines (Fig. 40C) | Prochelator sp. A |
| – | Pereonite 1 subequal or smaller than pereonite 2; ventral median spines absent on all pereonites | 50 |
| 50 | Pereonite 1 subequal to pereonite 2; head with anterolateral spines; pleotelson with posterolateral spines or angles and a rounded posterior medial border; uropods biramous (Fig. 40B) | Momedossasymmetrica |
| – | Pereonite 1 smaller than pereonite 2; head without anterolateral spines; pleotelson without posterolateral spines; uropods uniramous, lacking exopods (Fig. 40A) | Desmosoma sp. A |
| 51 | Lateral margins of pleotelson serrate with numerous denticulations or spines; coxae of pereonites not visible dorsally | 52 |
| – | Lateral margins of pleotelson smooth, without denticulations or spines; coxae visible dorsally on pereonites 2–7, most or all with single large projecting spine | 54 |
| 52 | Anterior margin of head broadly quadrate with rounded anterolateral angles; lateral margins of pereonites heavily denticulate; pleotelson ca. as wide as long, lateral margins completely serrate; uropods inserted on posterolateral margins of pleotelson (Fig. 41A) | Paramunnaquadratifrons |
| – | Anterior margin of head with strongly produced and rounded anterior margin; lateral margins of pereonites with or without denticulations or spines; uropods inserted slightly dorsally on pleotelson | 53 |
| 53 | Head with 2 granulate tubercles above antennal bases; body widest at pereonite 1; lateral margins of pereonites serrate, anterior-most tooth enlarged and pointed forward on pereonites 1–3; pleotelson dentate along basal two-thirds of lateral margins, with 2 strong teeth located on rounded end where uropods would typically be inserted (Fig. 41B) | Paramunna sp. A |
| – | Head without granulate tubercles; body widest at pereonites 2 and 3; lateral margins of pereonites without denticulations or spines; pleotelson with serrated lateral margins, although the details are undocumentedEndnote 29 | Paramunna sp. SD1 |
| 54 | Basal articles of antennules with large curved anterolateral projections; body surface rough with apparent microscales; projecting spine on each coxa of pereonite 7 typically large and conspicuous (Fig. 41D) | Pleurogonium sp. A |
| – | Antennules without anterolateral projections on basal articles; surface of body smooth to slightly scaled; coxal spines of pereonite 7 reduced or absent (Fig. 41C) | Pleurogoniumcaliforniense |
Figure 32.

Isopoda, Asellota, Janiroidea, Joeropsididae: AJoeropsisconcavaA1 = antennule (after Wilson 1997) BJoeropsisdubiadubiaB1 antennule B2 lateral margin of pleotelson (after Menzies 1951b) CJoeropsisdubiapaucispinisC1 head C2 pleotelson (after Menzies 1951b) DJoeropsislobata (after Richardson 1905a) EJoeropsis sp. A E1 antennule (after Wilson 1997).
Figure 33.

Isopoda, Asellota, Janiroidea, Janiridae (in part): AJaniralatadavisi (after Menzies 1951b) BJaniralataoccidentalis (after Menzies 1951b) CJaniralatarajata (after Menzies 1951b) DJaniralatasolasteri (after Menzies 1951b) EJaniralata sp. A (after Wilson 1997) FJaniralata sp. B (after Wilson 1997) GJaniralata sp. C (after Wilson 1997) HJaniralata sp. D (after Wilson 1997).
Figure 34.

Isopoda, Asellota, Janiroidea, Janiridae (in part): AIaniropsisanalogaA1 enlarged dorsal view of pleotelson showing lateral spines (after Menzies 1952) BIaniropsisderjugini (after Menzies 1952) CIaniropsiskincaidi (after Menzies 1952) DIaniropsisminuta (after Menzies 1952) EIaniropsismontereyensis (after Menzies 1952) FIaniropsistridensF1 enlarged dorsal view of pleotelson showing lateral spines (after Menzies 1952).
Figure 35.

Isopoda, Asellota, Janiroidea, Paramunnidae (in part): AAustrosignumlatum (after Just and Wilson (2021)BBoreosignum sp. A (after Cadien 2008) CMunnogoniumerratum, C1 male pereopod 2 (after Wilson 1997) DMunnogoniumtilleraeD1 male pereopod 2 (after Just and Wilson (2021). Pleurocopidae: EPleurocopefloridensis (Caribbean species = representative for Pleurocope sp. A; after Kensley and Schotte 1989).
Figure 36.

Isopoda, Asellota, Janiroidea, Munnidae: AMunnachromatocephala (after Menzies 1952) BMunnafernaldi (after George and Strömberg 1968) CMunnahalei (after Menzies 1952) DMunnamagnifica (after Schultz 1964) EMunnaspinifrons (after Menzies and Barnard 1959) FMunnastephenseniF1 close-up of pleotelson, arrow indicating sensory setae on left lateral margin (after Menzies 1952) GMunna sp. A (after Wilson 1997) HUromunnaubiquitaH1 close-up of pleotelson, arrow indicating spines or serrations on right lateral margin (after Menzies 1952).
Figure 37.
Isopoda, Asellota, Janiroidea, Haplomunnidae: AHaplomunnacaecaA1 dorsal view of brooding female A2 right lateral view of brooding female A3 left lateral view of pleotelson tip and 3-spined tubercle (after Wilson 1976).
Figure 38.
Isopoda, Asellota, Janiroidea, Munnopsidae: ABelonectes sp. A (after Wilson 1997) BEurycopecaliforniensis (after Schultz 1966) CIlyarachnaacarina (after Thistle 1979) DIlyarachnaprofunda (after Thistle 1979) EMunnopsurus sp. A (after Wilson 1997) FMunnopsurus sp. B (after Wilson 1997).
Figure 39.

Isopoda, Asellota, Janiroidea, Janiridae (in part): ACaecijaerahorvathi (after Menzies 1951c) BCaecianiropsispsammophila (northern California species = representative for Caecianiropsis spp. LA1 and LA2; after Menzies and Pettit 1956). Lepidocharontidae: CMicrocharonsabulumC1 close-up of uropods (Caribbean species = representative for Microcharon sp. A; after Kensley 1984).
Figure 40.

Isopoda, Asellota, Janiroidea, Desmosomatidae: ADesmosoma sp. A (after Wilson 1997) BMomedossasymmetrica (after Schultz 1966) CProchelator sp. A (after Wilson (1997); Nannoniscidae: DNannonisconuslatipleonus (after Schultz 1966).
Key J. Suborder Oniscidea, Superfamily Oniscoidea
Figure 42.

Isopoda, Oniscidea, Oniscoidea, Ligiidae: ALigiaoccidentalis (representative species for Ligiacf.occidentalis; after Allen 1976). Halophilosciidae: BLittorophilosciarichardsonae (after Brusca et al. 2007). Platyarthridae: CNiambiacapensis (after Brusca et al. 2007). Tylidae: DTylospunctatus, lateral view of whole body with posterior view of pleonites 4 and 5 plus pleotelson (after Brusca et al. 2007).
Figure 43.

Isopoda, Oniscidea, Oniscoidea, Alloniscidae: AAlloniscusmirabilis (after Brusca et al. 2007) BAlloniscusperconvexus (after Richardson 1905a). Detonidae: CArmadilloniscuscoronacapitalis (after Menzies 1950c) DArmadilloniscusholmesi (after Menzies 1950c) EArmadilloniscuslindahli (after Menzies 1950c).
| 1 | Uropods ventral and operculate covering pleopods, not visible in dorsal view; body able to roll up into a ball (conglobate) (Fig. 42D) | Tylospunctatus |
| – | Uropods terminal, visible in dorsal view; body may or may not be able to conglobate | 2 |
| 2 | Flagellum of antenna with > 10 articles; eyes large with > 50 ommatidia each, and wrapping around lateral margins of head, distance between eyes equal to length of 1 eye; isopods unable to conglobate (Fig. 42A)Endnote 30 | Ligiacf.occidentalis |
| – | Flagellum of antenna with 2–4 articles; eyes not as above, much smaller (< 30 ommatidia) and not wrapping around lateral margins of head; isopods may or may not be able to conglobate | 3 |
| 3 | Flagellum of antenna with 2 or 3 articles; head without large anterolateral lobes | 4 |
| – | Flagellum of antenna with 4 articles; head with very large, distinctly produced anterolateral lobes, each lobe broad and truncate at tip | 7 |
| 4 | Flagellum of antenna with 2 articles; dorsal surface of body covered with fine, but distinct scales (Fig. 42C) | Niambiacapensis |
| – | Flagellum of antenna with 3 articles; dorsum without distinctly visible scales | 5 |
| 5 | Head with small, forward protruding cone-shaped lobes situated just anterior to eyes; frontal margin of head produced into blunt median lobe; pleon not abruptly narrower than pereon | 6 |
| – | Head without cone-shape lateral lobes anterior to eyes; frontal margin of head slightly sinuous, but not produced into median lobe; pleon abruptly narrower than pereon (Fig. 42B) | Littorophilosciarichardsonae |
| 6 | Uropodal protopod with rounded and expanded posterolateral margins (Fig. 43A) | Alloniscusmirabilis |
| – | Uropodal protopod with oblique posterolateral margins, not produced (Fig. 43B) | Alloniscusperconvexus |
| 7 | Anterior margin of head produced into strong, truncate median lobe; body markedly convex and capable of conglobation (Fig. 43E) | Armadilloniscuslindahli |
| – | Anterior margin of head produced into sharply acute median lobe; body not markedly convex and not capable of conglobation | 8 |
| 8 | Dorsal surface of body covered with large, elevated tubercles best developed on head; posterior border of all body segments except pleotelson appear beaded with minute, evenly spaced tubercles; antennal peduncle article 4 with spur-like process on lateral margin (Fig. 43C) | Armadilloniscuscoronacapitalis |
| – | Dorsal surface of body rough, with low, rounded tubercles; posterior border of all body segments smooth; antennal peduncle article 4 without spur-like process on lateral margin (Fig. 43D) | Armadilloniscusholmesi |
Endnotes
The provisional species Sphaeromatidae sp. A listed in SCAMIT (2021) was reported based on immature female specimens collected off Catalina Island at a depth of 12–16 m. Although these isopods co-occurred with two other sphaeromatids, Dynamenellaglabra and Paracerceissculpta, they differed from females of both species as well as other known members of those two genera (but see Endnote 17 below regarding the correct generic assignment for some SCB Dynamenella). Although the voucher sheet developed for Sphaeromatidae sp. A included a note that these isopods could be immature Paracerceisgilliana, this hypothesis has not been confirmed. Consequently, this provisional species is not included in Key G because its correct generic assignment is not known. The second species listed in SCAMIT (2021) not included in this review is the oniscid Armadillidiumvulgare, which is a strictly terrestrial species with a worldwide distribution.
No illustrations exist for the provisional anthuroid Eisothistos sp. A collected from Santa Barbara to Catalina Island at depths of 5–77 m, although Brusca and Cadien (1993) stated that the species is similar in gross morphology to Eisothistosantarcticus (see Wägele 1984a: fig. 2). However, neither E.antarcticus nor any of the other 30 species of Eisothistos currently recognized by WoRMS (Boyko et al. 2008 onwards) are known to occur near the SCB. For additional information on the genus see Wägele (1979, 1984a), Müller (1990), Poore (2001a), and Poore and Lew Ton (2002).
The aegid Aegiochusplebeia has not yet been reported from the SCB. However, the species is included in this guide since its known geographic range spans the region. See Brusca (1983a) and Bruce (2009a) for additional information.
It is currently unclear how to clearly distinguish juveniles of most cymothoid species from each other. Although Brusca (1978a, 1978b) provided detailed descriptions of the life histories of Elthusavulgaris (as Lironecavulgaris) and Nerocilaacuminata (as N.californica), such information is not readily available for other local cymothoid species. Consequently, all juvenile cymothoids are treated as unidentified Cymothoidae in Key C.
Since its description in 1980, the cymothoid Renocilathresherorum has been considered primarily a Gulf of California species with only a few known records from elsewhere in the Eastern Pacific (see Brusca 1981). These include one SCB record of a female and an associated juvenile collected off Corona del Mar in southern California plus a couple of records from Magdalena Bay on the southwest coast of Baja California. The species, however, has also been reported recently from the Suez Canal area of Egypt (e.g., Eman et al. 2014; Mahmoud et al. 2016; Ali and Aboyadak 2018). Given this disjunct distribution, it would be useful to perform a comprehensive morphological and molecular comparison of Eastern Pacific vs. Egyptian specimens to confirm whether they represent the same or perhaps two similar looking species of Renocila.
The cymothoid Ceratothoagaudichaudii is included in this guide based on historical distribution records (e.g., Brusca 1981). However, the species is listed as “taxon inquirendum” in WoRMS (Boyko et al. 2008 onwards) based on the review by Martin et al. (2015). Also see Hadfield et al. (2016) for a current list of accepted species in Ceratothoa Dana, 1852, and for information regarding taxonomic uncertainties within the genus.
Female and juvenile gnathiids key only to family in Key D. However, Haney (2006) provides a comprehensive discussion and guide to pairing unidentifiable females and juveniles with their male counterparts for many of the northeast Pacific species of Gnathia Leach, 1814 and Caecognathia Dollfus, 1901.
No illustrations exist for the three provisional species of gnathiids included in Key D (Caecognathia sp. A, Caecognathia sp. SD1, Gnathia sp. MBC1). However, photographs of these and all other known species of gnathiids that occur in SCB waters are available in Haney (2006).
The provisional species Capitetragonia sp. A was originally reported in 2011 as Probopyria sp. A (species voucher sheet available from SCAMIT Toolbox at https://www.scamit.org; T. D. Stebbins, 4 September 2011). However, the genus Probopyria Markham, 1985 was synonymized with Capitetragonia Pearse, 1953 by An et al. (2015). Because no illustrations exist for this provisional species, an image of the type species of Capitetragonia, C.alphei (Richardson, 1900), is provided as an example in Fig. 15B.
The bopyrid Eremitionegiardi is listed in SCAMIT (2021) as Pseudionegiardi, although this and eight other species previously included in Pseudione were transferred to the newly erected genus Eremitione by Williams et al. (2019). This species was originally added to the SCAMIT species list in 2011 (Edition 6) based on a trawl record from off the coast of Palos Verdes (D. Cadien, pers. comm.). However, the record may be questionable because E.giardi was not previously known from California waters (see Markham 1974b) except possibly for a report of an unconfirmed species identified as Pseudione sp. found infesting the hermit crab Pagurushirsutiusculus in central California (J. Markham, pers. comm.). The record of E.giardi off Palos Verdes is likely valid if the host was a hermit crab, but not if the host was another type of crustacean (J. Markham, pers. comm.).
Two different adult body morphologies have been described for the dajid isopod Holophryxusalaskensis (see Richardson 1905a, 1905b: Butler 1964; Boyko and Williams 2021b), while Coyle and Mueller (1981) provided complete descriptions of the larval and juvenile stages of the species. In the original species description based on three adult female specimens collected off Alaska without any host information, Richardson (1905b) described and illustrated a stout and somewhat irregular body form, the dorsal and ventral views which are reproduced in Fig. 16A herein. Richardson (1908) later described a second species of the genus, H.californiensis, based on one adult female collected off Alaska attached to the shrimp Pasiphaeapacifica, plus one female and one male collected off Santa Barbara, southern California. In his subsequent redescription of H.alaskensis based on Richardson’s type material and additional specimens of both sexes from British Columbia, Butler (1964: figs 1, 3) synonymized H.californiensis with H.alaskensis and described a more typical elongate and symmetrical body form for adult females. Additionally, Boyko and Williams (2021b) recently provided a thorough review of the above with new records that extended the southern distribution of H.alaskensis to deep waters off the US-Mexico border. These authors also provided color photographs of typical female and male H.alaskensis, as well as an in-situ photo of an adult female attached to its host shrimp (Boyko and Williams 2021b: fig. 1).
Specimens of unidentified dajid isopods listed as Zonophryxus sp. in SCAMIT (2021) were collected in SCB waters during the 1990s parasitizing two species of caridean shrimp, Pantomusaffinis and Plesionikatrispinus (see Montagne and Cadien 2001; Boyko and Williams 2021a). Whether these isopods represent one or more species awaits further investigation. Because no illustrations exist for Zonophryxus sp., images of a recently described species from Peruvian waters, Zonophryxusprobisowa Boyko & Williams, 2021, are provided in Fig. 16B as an example of the genus. Although it is not clear how closely this species resembles the SCB specimens, these images clearly show the notched appearance of the posterior end that is characteristic of species of Zonophryxus. In addition to describing Z.probisowa, Boyko and Williams (2021a) reviewed and provided a key to all seven species of Zonophryxus based on female characters. Finally, a couple of other eastern Pacific specimens of Zonophryxus recently discovered from mis-identified Gulf of California samples collected during the Talud VIII cruises in 2005 (TDS, pers. obs.) may represent an additional undescribed species of this genus (C. Boyko, pers. comm.).
There may be a new species of Heteroserolis in deeper waters of the SCB. Preliminary comparisons of specimens collected from depths > 100 m off San Diego with those from shallower waters in San Diego Bay (depth ≤ 15 m) reveal several differences in morphology. Whether these differences merit a new species requires further investigation. The differences between the shallow-water and deep-water specimens include:
Deep-water specimens have a much larger dorsal carina than do shallow-water animals. This is like differences described by Hessler (1972) between H.mgrayi (like deep-water specimens) and H.carinata (like San Diego Bay specimens).
Deep-water specimens have a rather shallow lateral notch on the pleotelson that does not form a well-defined tooth (like described for H.tropica and H.mgrayi), while shallow-water animals have the distinct tooth or deep notch characteristic of H.carinata.
Pereonites 5 and 6 are subequal in width to pereonite 4 in the deep-water animals (the lateral margins are nearly continuous with a gradual tapering to the pleotelson). In contrast, there is a distinct narrowing of the pereon between pereonites 4 and 5 in the shallow-water animals. This difference in shape appears distinct even in juveniles.
Two species of the sphaeromatoid genus Ancinus H. Milne Edwards, 1840 are reported herein as occurring in the SCB, including A.granulatus Holmes & Gay, 1909 and A.seticomvus Trask, 1971. Although Schultz (1973) considered A.seticomvus a junior synonym of A.granulatus as it has been listed by SCAMIT (1998–2021) and Brusca et al. (2007), Glynn and Glynn (1974) disputed Schultz’s assessment and provided a detailed suite of characters by which to distinguish the two species. Consequently, most subsequent workers have recognized A.seticomvus as a valid species (e.g., Dexter 1976; Iverson 1982; Pires 1987; McLaughlin et al. 2005; Rocha-Ramírez et al. 2010) as currently accepted by WoRMS (i.e., Boyko et al. 2008 onwards).
Although the sphaeromatid Gnorimosphaeromaoregonense is included in the SCAMIT (2021) list of SCB species, Wetzer et al. (2021), in their recent redescription and assessment of the species and genus, could only verify its range from Alaska to San Francisco, California. Thus, the presence of G. oregonense south of central California is questionable, and any specimens encountered should be carefully examined. While G. oregonense is a fully marine species, the other species of the genus in SCB waters included in this review, G. noblei, occurs in brackish or freshwater habitats. In addition to habitat differences, these two species can best be distinguished by the shape of their coxal plates as indicated in the key. See Wetzer et al. (2021: fig. 15) for SEM comparisons of the coxae of G. oregonense and G. noblei. Finally, although Gnorimosphaeromainsulare (Van Name, 1940) has been reported from the SCB, it is not included in this guide since it is only known from a freshwater pond on San Nicolas Island and is therefore not a marine species (see Wetzer et al. 2021 for additional details).
Even though the couplet in Key G identifying the sphaeromatid Paracerceissculpta distinguishes between male and female characteristics, it does not account for differences between the three discrete male morphs of the species described in Shuster (1987). The male traits specified in the key represent those of the larger and more elaborate alpha-males that attract females to the spongocoels of calcareous sponges where mating and brooding of young occurs. Alpha-males are characterized by elongated uropods and highly sculptured pleotelsons as indicated in the key. Smaller beta-males lack such ornamentation and resemble females of P.sculpta, while tiny gamma-males resemble immature juveniles. All three male morphs, however, possess well-developed secondary sex characters (penes and appendix masculina) and are sexually mature. See Shuster (1987, 1992), Shuster and Wade (1991a, 1991b), and Shuster and Guthrie (1999) for additional information on the reproductive biology and ecology of P.sculpta, and the relationship between alpha, beta, and gamma males.
The sphaeromatids Dynamenelladilatata and Dynamenellaglabra as treated herein and in SCAMIT (2021) are instead classified in the genus Dynamene Leach, 1814 in both WoRMS (Boyko et al. 2008 onwards) and Wall et al. (2015), although listed as “taxon inquirendum” or “correct generic placement uncertain,” respectively. Thus, the correct generic assignment for these two species is currently in question. Based on molecular data, Wetzer et al. (2018) demonstrated that Dynamene and Dynamenella are not closely related (i.e., occurring in Cerceis and Dynamenella clades, respectively). Morphological revisions based on adult males are needed to be able to correctly place these species.
Species of the sphaeromatid genus Exosphaeroma Stebbing, 1900 have either a 1:1 male to female ratio in which males are observed to mate guard an individual female or a highly skewed sex ratio where adult males guard harems and are very rare relative to females and subadults. In the SCB, E.inornata exhibits the first of these mating patterns with individual male/female pairs. Both sexes in this species are similar in general appearance, are unornamented, and have uropodal endopods in which the posterior margin is evenly rounded. Alternatively, in the E.amplicauda-like species (E.amplicauda, E.aphrodita, and E.pentcheffi) treated herein in which males guard harems of females, the adult males are characterized by a pleotelson to overall body length ratio of 0.30 or greater, a pleotelson with an acuminate posterior border, and uropodal endopods in which the posterior margins are falcate. A review of Exosphaeroma from the north-eastern Pacific evaluated the status of E.amplicauda and E.aphrodita and described an additional three new species (Wall et al. 2015). A neotype of E.amplicauda was designated and the species redescribed based on the neotype and topotypic specimens. As a result of this study, the distribution of E.amplicauda was restricted to northern California along the coasts of Marin, Sonoma, and San Mateo Counties. In contrast, E.aphrodita originally described from La Jolla (San Diego County) and previously considered a nomen dubium was taken out of synonymy and revalidated by Wall et al. (2015) as a SCB species. Additionally, the new species E.pentcheffi was described from intertidal samples collected at Point Fermin (Los Angeles County). These authors are also aware that there are likely other undescribed species of Exosphaeroma occurring in the Channel Islands within the SCB, with specimens from Santa Catalina Island being genetically closely related to E.pentcheffi (RW, pers. obs.). The E.amplicauda-like species along the western coast of North America have been previously lumped and unrecognized, in part because original descriptions were not detailed and the best drawings available were those of Kussakin (1979). However, Kussakin’s material was based on specimens collected from Amchitka Island, Alaska, some 2000 km north of the type locality of E.amplicauda near San Francisco. Consequently, Kussakin’s specimens probably also represent an undescribed species.
Even though the sphaeromatid Exosphaeromarhomburum is an accepted species in WoRMS (Boyko et al. 2008 onwards) and is reported in SCAMIT (2021), Wall et al. (2015) designated the species incertae sedis in their recent treatment of north-eastern Pacific Exosphaeroma. This was due in part to the brief original description provided by Richardson (1899: as Sphaeromarhomburum), in which she only described and figured the pleon, pleotelson, and uropods in any detail. Although a figure of the maxilliped was also presented, Richardson (1899) provided no accompanying description of the structure. Richardson (1899) also did not specify the sex of either of the two specimens she examined, both of which were collected in central California (Monterey Bay). Because of the above and the lack of confirmed males known to them at the time, Wall et al. (2015) were unable to further evaluate the status of the species, thus resulting in their incertae sedis designation. As part of the present study, however, we were able to examine specimens of both male and female isopods attributed to E.rhomburum and confirm its validity as a distinct species without any significant sexual dimorphism. On the other hand, we question whether the species actually belongs in Exosphaeroma because it appears to differ noticeably in morphology from other species of the genus. In her original description and subsequent work, Richardson (1899, 1905a) also noted that the species was “similar to S.egregium Chilton” (= Sphaeromaegregia Chilton, 1892), a species now placed in the southern hemisphere genus Cymodocella Pfeffer, 1887 (see Harrison and Holdich 1982; Bruce 1995). Based on a cursory review of some Cymodocella specimens available to us, we agree that E.rhomburum appears to resemble that genus more closely than it does Exosphaeroma. However, it will require further morphological and preferably molecular studies to confirm whether the species should be reassigned to Cymodocella or another existing or perhaps new genus. Finally, we extend the southern distribution of E.rhomburum to NW Baja California based on our examination of specimens collected by the City of San Diego’s Ocean Monitoring Program, while the northern range limit is extended herein to Washington State, USA based on observations reported by Lafferty and Suchanek (2016).
Three species of the valviferan genus Idarcturus Barnard, 1914 are reported herein as occurring in the SCB. Of these, I.allelomorphus is the most common, occurring in soft-bottom continental shelf sediments at depths < 100 m from central California to Baja California Norte (see Menzies and Barnard 1959; Wetzer and Brusca 1997; Campos and Villarreal 2008). A second species, I.hedgpethi, was long known from only northern California where it was collected in 1948 living on hydroids in the low intertidal (see Menzies 1951b). Although I.hedgpethi was added to the SCAMIT species list in 2008 (Edition 5), we have been unable to confirm the source of that record. One record of I.hedgpethi was reported from Port Hueneme (Ventura County, southern California) during the 2000 and 2001 surveys of California bays and harbors (Fairey et al. 2002), but it is unknown whether that record was the source of the listing in SCAMIT (2008). The species was also reported from much further north in the Strait of Georgia, British Columbia by Macdonald et al. (2010). If accurate, these last two records expand the geographic range of I.hedgpethi from the northern California type locality to span ~ 1700 km from British Columbia to the SCB. Although I.hedgpethi was also recently reported from soft-bottom benthic sites located near the northern Channel Islands during the Bight 2013 regional monitoring project (Gillett et al. 2017), those specimens likely represent an undescribed species of Idarcturus that was first observed in the region during the previous Bight 2008 regional project (TDS, pers. obs.), but which was lumped together with I.allelomorphus in the project’s final report (i.e., Ranasinghe et al. 2012). Although the Bight 2008 specimens were initially thought to resemble I.hedgpethi in terms of their pronounced spination pattern, subsequent observations demonstrated that they clearly do not represent that species. Although it is possible that the Bight 2008 and Bight 2013 specimens of Idarcturus collected near the Channel Islands may turn out to be a more spinous form of I.allelomorphus, this appears unlikely based on preliminary results of ongoing investigations. Consequently, we have designated these specimens as Idarcturus sp. A in this review.
Edotia sp. B is currently being described by TD Stebbins and R Wetzer (unpublished results, manuscript in preparation). This paper also includes a redescription of E.sublittoralis with a global review of the distribution and ecology of the genus.
The three species of Synidotea characterized by an apically rounded or spatulate pleotelson (S.calcarea, S.magnifica, and S.media) that occur in SCB waters are difficult to distinguish using the existing literature (e.g., Menzies and Barnard 1959; Schultz 1966; Iverson 1972; Menzies and Miller 1972; Wetzer and Brusca 1997). The problem is primarily due to an incomplete original description of S.magnifica. For a more detailed discussion of the issue and its resolution, see Stebbins (2012a).
The idoteid isopods Idoteafewkesi, I.rufescens, and I.urotoma included in the SCB fauna were listed in the genus Pentidotea in SCAMIT (2021). Although Poore and Lew Ton (1993) provided a more restrictive diagnosis of the genus Idotea while also raising Pentidotea to full generic status, they did not recommend that species previously assigned to Idotea be transferred to Pentidotea or vice versa. Consequently, species formerly assigned to the subgenera Idotea and Pentidotea were placed in the genera Idotea and Pentidotea, respectively, as is currently accepted in WoRMS (see Boyko et al. 2008 onwards). Although the proper affinities of some species of Idotea such as I.fewkesi and I.urotoma are uncertain at this time due to the more restrictive generic diagnosis as pointed out by Poore and Lew Ton (1993), reassignment of any of these species to Pentidotea or another existing or new genus awaits further investigation.
Janiralatarajata and Janiralata sp. D may be difficult to distinguish unless comparing actual specimens of both species. For example, Wilson (1997) stated that Janiralata sp. D differs from J.rajata as follows: “the eyes are smaller, the body has numerous dense chromatophores, the antennula is longer, and the propodus of pereopod I has more proximoventral denticles (18 instead of 12 or 13).” Another possible difference is that Janiralata sp. D has only been reported from continental shelf waters deeper than 90 m in the Santa Maria Basin, while J.rajata is reported from shallower waters < 40 m deep from central to southern California.
Although no illustrations exist of the provisional asellote species Pleurocope sp. A, the SCAMIT voucher sheet for the species compares it to another provisional species of the same name in manuscript (i.e., Pleurocope sp. A Wilson MS), as well as to three of the other four recognized species of the genus (voucher sheet for Pleurocope sp. IS1 available from SCAMIT Toolbox at https://www.scamit.org; D. B. Cadien, 4 March 2012). These three species include P.dasyura from the Mediterranean, P.floridensis from the tropical western Atlantic, and P.wilsoni from Thailand and Aldabra. The fourth species, P.iriomotensis, is from Japan. An image of P.floridensis modified after Kensley and Schotte (1989: fig. 43B) is provided in Fig. 35E herein as an example of the genus.
Just and Wilson (2021) recently redescribed the SCB asellote Munnogoniumtillerae (Menzies & Barnard, 1959) in their review of several species in the ‘ Austrosignum-’ complex sensu Just and Wilson (2007). In doing so, these authors compared M.tillerae collected off San Diego to Munnogoniumerratum (Schultz, 1964) known only from the type locality off Santa Barbara County, as well as to specimens identified as Munnogonium cf. tillerae in Wilson (1997) that were also collected off Santa Barbara in the Santa Maria Basin. As a result of their comparisons, Just and Wilson (2021) concluded that the San Diego and Santa Barbara specimens represent distinct species, and they therefore restricted M.tillerae to occurring just off San Diego. Although these authors reported that “the type of M.erratum has been destroyed,” they also stated that the specimens of Munnogonium cf. tillerae studied by Wilson (1997) were collected near that type locality and at around the same depth. Thus, Just and Wilson (2021) used the features of M. cf. tillerae described in Wilson (1997) as M.erratum in their accompanying key to species of Munnogonium (G Wilson, pers. comm.). This treatment is followed in this paper, with M.tillerae restricted to the San Diego region and M.erratum considered synonymous with M. cf. tillerae from the Santa Maria Basin (i.e., Fig. 35C herein was reproduced from Wilson 1997: fig. 1.28).
The provisional asellote species Microcharon sp. A was originally assigned to the family Microparasellidae as listed in SCAMIT (2021). However, this genus was transferred to the family Lepidocharontidae by Galassi et al. (2016). Although no complete illustrations exist for Microcharon sp. A, the species is similar in general appearance to the Caribbean species Microcharonsabulum Kensley, 1984. Thus, an image of M.sabulum is provided in Fig. 39C as an example of the genus.
No illustrations exist for the SCB asellotes Caecianiropsis sp. LA1 and Caecianiropsis sp. LA2. However, these two provisional species are similar in general morphology to the type species of the genus from northern California, Caecianiropsispsammophila Menzies & Petit, 1956. Thus, an image of C.psammophila is provided in Fig. 39B herein as an example of the genus.
No illustrations exist for the provisional species Paramunna sp. SD1 collected during the Bight 2003 regional monitoring project (see Ranasinghe et al. 2007). Thus, the species can presently be distinguished from the other SCB species of Paramunna only by the characters in Key I.
Hurtado et al. (2010) and Eberl et al. (2013) presented fine-scale phylogeographic studies of Ligia populations distributed from central California to central Mexico, including the Gulf of California, describing several highly divergent lineages. Their conclusions suggest the presence of multiple Ligia species spanning this region, including at least one unnamed species that ranges throughout the SCB from south of Point Conception to NW Baja California, including the Channel Islands, as well as perhaps further south along the Baja peninsula. This species corresponds to the “Baja Pacific-Southern California” clade of Hurtado et al. (2010) and is referred to in this guide as Ligia cf. occidentalis.
Annotated list of species
Suborder Cymothoida
Superfamily Anthuroidea
Family Antheluridae
Ananthuraluna (Schultz, 1966) [Fig. 6E] Continental shelf, slope, and submarine canyon benthos (78–1298 m); Santa Monica Bay, Tanner Canyon, La Jolla, and Coronado canyons, southern California, USA. Type locality: USA, California, Tanner Canyon (R/V Velero IV Station No. 6832-60; 32°33'36"N, 118°55'40"W). Body length to ~ 21 mm. See Schultz (1966 original description as Bathuraluna; 1969, 1977), Kensley (1978), Poore and Lew Ton (1988d), Wetzer et al. (1991), Cadien and Brusca (1993), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Ranasinghe et al. (2007), SCAMIT (2008–2021), Espinosa-Pérez et al. (2009), and Gillett et al. (2019).
Family Anthuridae
Amakusanthuracaliforniensis (Schultz, 1964) [Fig. 6A] Continental shelf (depth to at least 80 m); Santa Maria Basin/Western Santa Barbara Channel, southern California, USA to Guadalupe Island off western Baja California Norte, Mexico. Type locality: USA, California, Los Angeles County, 9.9 mi SSW of Santa Monica Pier (R/V Velero IV Station No. 2998-55; 33°53'22"N, 118°34'40"W). Body length to ~ 11 mm. See Schultz (1964 original description as Apanthuracaliforniensis; 1969), Lissner et al. (1986), Poore and Lew Ton (1988b), Wetzer et al. (1991), Cadien and Brusca (1993), SCAMIT (1994–2021), Wetzer and Brusca (1997), Ranasinghe et al. (2003, 2007, 2012), McLaughlin et al. (2005), and Espinosa-Pérez and Hendrickx (2006).
Cyathuramunda Menzies, 1951 [Fig. 6B] Low intertidal to continental shelf, on kelp holdfasts to deep-water rocks (0–132 m); Marin County, northern California, USA to NW Baja California, Mexico, and the Gulf of California. Type locality: USA, California, Marin County, Tomales Point, Tomales Bluff (reef). Body length to ~ 6 mm. See Menzies (1951b), Menzies and Barnard (1959), Schultz (1964, 1969), Miller (1975), Kussakin (1982), Lissner et al. (1986), Wetzer et al. (1991), Cadien and Brusca (1993), SCAMIT (1994–2021), Chess and Hobson (1997), Wetzer and Brusca (1997), Espinosa-Pérez and Hendrickx (2001a, 2006), McLaughlin et al. (2005), and Gillett et al. (2017).
Haliophasmageminatum Menzies & Barnard, 1959 [Fig. 6C] Continental shelf, slope, and submarine canyon benthos (9–512 m); Strait of Georgia, British Columbia, Canada to San Quintin Bay, Baja California Norte, Mexico. Type locality: USA, California, off San Mateo Point (R/V Velero IV Station No. 4771-56; 33°21'40"N, 117°35'50"W). Body length to ~ 8 mm. See Menzies and Barnard (1959 original description as Haliophasmageminata; gender of specific name corrected to geminatum in Boyko et al. 2008 onwards), Menzies (1962a), Schultz (1964, 1966, 1969), Lie (1968), Iverson (1974), Poore (1975), Gillard (1978), Lissner et al. (1986), Poore and Lew Ton (1988a), Wetzer et al. (1991), Cadien and Brusca (1993), Thompson et al. (1993), SCAMIT (1994–2021), Wetzer and Brusca (1997), Bergen et al. (1998, 2001), Smith et al. (2001, 2003), Ranasinghe et al. (2003, 2007, 2012), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Campos and Villarreal (2008), Macdonald et al. (2010), Puget Sound Institute (2012–2023), Gillett et al. (2017, 2019, 2022), and Henkel et al. (2020).
Mesanthuraoccidentalis Menzies & Barnard, 1959 [Fig. 6D] Low intertidal to shallow water, on kelp and rocks (0–20 m); Point Conception, southern California, USA to Baja California and the Gulf of California, Mexico, and to the Gulf of Nicoya, Costa Rica. Type locality: California, 11 miles east of Point Conception (R/V Velero IV Station No. 4822-57; 34°27'15"N, 120°14'45"W). Body length to ~ 7 mm. See Menzies and Barnard (1959), Schultz (1964, 1969), Allen (1976), Wägele (1984b), Wetzer et al. (1991), Cadien and Brusca (1993), Chess and Hobson (1997), Espinosa-Pérez and Hendrickx (2001a), McLaughlin et al. (2005), Campos and Villarreal (2008), and SCAMIT (2012–2021).
Family Expanathuridae
Eisothistos sp. A [No figure] Inner to mid shelf offshore habitats collected off rocks and the coral Coenocyathusbowersi (5–77 m); southern California, USA from Tajiguas, Santa Barbara County and Catalina Island. Body length of juveniles to ~ 2.5 mm (i.e., adults not known). See Cadien and Brusca (1993) and SCAMIT (2013–2021). Note: See Endnote 2.
Family Hyssuridae
Kupellonura sp. A [Fig. 6F] Continental shelf, hard bottom benthos (105–117 m); Santa Maria Basin offshore of Purisima Point, Santa Barbara County, California, USA (~ 35 km north of SCB northern boundary). Body length = no information available. See Wetzer and Brusca (1997: 19–20, fig. 1.7, description and figure) and SCAMIT (2008–2021).
Family Paranthuridae
Califanthurasquamosissima (Menzies, 1951) [Fig. 5C] Intertidal to offshore continental shelf and bays, in kelp and eelgrass beds, and on muddy or sandy sediments (0–142 m); Dillon Beach, northern California, USA to Oaxaca, Mexico, and the Gulf of California. Type locality: USA, California, Marin County, Tomales Point, Tomales Bluff. Body length to ~ 5.6 mm. See Menzies (1951b original description as Colanthurasquamosissima), Menzies and Barnard (1959), Schultz (1969), Miller (1975), Brusca (1980), Kussakin (1982), Poore (1984b), Wetzer et al. (1991), Cadien and Brusca (1993), SCAMIT (1994–2021), Espinosa-Pérez and Hendrickx (2001a, 2006), McLaughlin et al. (2005), Brusca et al. (2007), Moore and Hovel (2010), Ranasinghe et al. (2012), Bastida-Zavala et al. (2013), Cruz-García et al. (2013), Gillett et al. (2017), and García-Madrigal et al. (2022).
Colanthurabruscai Poore, 1984 [Fig. 5D] Intertidal to shallow subtidal (0–27 m); San Clemente, southern California, USA to Salinas Bay, Costa Rica. Type locality: Mexico, Sonora, Puerto Peñasco, Station Beach Reef (on Sargassum; 31°20'N, 113°35'W). Body length to ~ 5.4 mm. See Poore (1984b), Wetzer et al. (1991), Cadien and Brusca (1993), Espinosa-Pérez and Hendrickx (2001a, 2006), McLaughlin et al. (2005), and Brusca et al. (2007).
Paranthuraelegans Menzies, 1951 [Fig. 5A] Intertidal to subtidal, in low rocky intertidal, eelgrass beds, algal mats, muddy sediments, and pier pilings (0–55 m); Marin County, northern California, USA to San Quintin Bay, Baja California Norte, Mexico, and the Gulf of California. Type locality: USA, California, Marin County, Tomales Point, Tomales Bluff (reef). Body length to ~ 9.1 mm. See Menzies (1951b, 1962a), Menzies and Barnard (1959), Miller (1975), Brusca (1980), Kussakin (1982), Lissner et al. (1986), Wetzer et al. (1991), Cadien and Brusca (1993), SCAMIT (1994–2021), Bergen et al. (1998), Espinosa-Pérez and Hendrickx (2001a, 2006), Ranasinghe et al. (2003, 2007, 2012), Smith et al. (2003), Healey and Hovel (2004), McLaughlin et al. (2005), Reed and Hovel (2006), Sirota and Hovel (2006), Brusca et al. (2007), and Melwani and Kim (2008).
Paranthurajaponica Richardson, 1909 [Fig. 5B] Introduced Asian species that occurs in fouling communities and shallow-water marinas; worldwide distribution, including northern and southern Japan, eastern Russia, eastern China, southwestern France, and from northern to southern California, USA in North America. Type locality: Japan, Mororan. Body length to ~ 8.5 mm. See Richardson (1909), Kussakin (1982), Cohen and Carlton (1995), Cohen et al. (2005), Brusca et al. (2007), Ruiz et al. (2011), Lavesque et al. (2013), SCAMIT (2013–2021), and Gillett et al. (2017, 2022).
Superfamily Cymothooidea
Family Aegidae
Aegalecontii (Dana, 1854) [Fig. 8E] Continental shelf and submarine canyons (depth to at least 218 m), collected from fishes or soft-bottom habitats; central to southern California, USA. Type locality: USA, California. Body length to ~ 20 mm. See Dana (1854 original description as Aegacyllalecontii), Stimpson (1857), Richardson (1899, 1900b, 1905a), Hewatt (1946), Schultz (1966, 1969), Miller (1975), Kussakin (1979), Brusca (1983a), SCAMIT (2001–2021), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), and Bruce (2009a). Note: Although Dana (1854) did not designate a specific type specimen or locality for A.lecontii, the specimen on which his original description was based is believed to be in the invertebrate zoology collections of the Yale Peabody Museum of Natural History (Catalog No. YPM IZ 000303.CR).
Aegiochusplebeia (Hansen, 1897) [Fig. 8F] Deep water off oceanic islands (688–2534 m); eastern Pacific (Aleutian Islands to Peru), western Pacific (Indonesia to Japan), and New Caledonia. Type locality (syntypes): Eastern Pacific off Cocos Island and Panama (Albatross Station No. 3363). Body length to ~ 37 mm. See Hansen (1897 original description as Aegaplebeia), Richardson (1909 as Aegamagnoculis), Birstein (1973), Brusca (1983a), Brusca and Iverson (1985), Bruce (2009a), Azofeifa-Solano and Cortés (2020), and Zúñiga Delgado et al. (2021). Note: See Endnote 3.
Rocinelaangustata Richardson, 1904 [Fig. 9A] Continental shelf, slope, and basins (30–2534 m), collected from fishes or soft-bottom habitats; Bering Sea, Alaska, USA to Baja California, Mexico, and the Gulf of California. Type locality (lectotype): Bering Sea, USA, Alaska, NW of Unimak Island (54°48'50"N, 165°42'W). Body length to ~ 37.4 mm. See Richardson (1904a, 1905a), Hatch (1947), George and Strömberg (1968), Schultz (1969), Birstein (1973), Miller (1975), Lissner et al. (1986), Brusca and France (1992), SCAMIT (1994–2021), Wetzer and Brusca (1997), Smith et al. (2001), Espinosa-Pérez and Hendrickx (2002, 2006), McLaughlin et al. (2005), Moles (2007), Ranasinghe et al. (2007), Bruce (2009a), Macdonald et al. (2010), Gillett et al. (2017, 2019), and Henkel et al. (2020).
Rocinelabelliceps (Stimpson, 1864) [Fig. 9B] Continental shelf and slope (shallow subtidal to 284 m), collected from fishes or soft-bottom habitats; Alaska, USA to western Baja California, Mexico, and the Gulf of California. Type locality (neotype): USA, Washington, San Juan Island. Body length to ~ 29.1 mm. See Stimpson (1864 original description as Aegabelliceps), Richardson (1898, 1899, 1905a), Fee (1927), Hatch (1947), Schultz (1966, 1969), George and Strömberg (1968), Miller (1975), Allen (1976), Kussakin (1979), Brusca and France (1992), SCAMIT (1994–2021), Chess and Hobson (1997), Espinosa-Pérez and Hendrickx (2001a, 2002, 2006), McLaughlin et al. (2005), Moles (2007), Macdonald et al. (2010), Puget Sound Institute (2012–2023), and Ranasinghe et al. (2012).
Rocinelalaticauda Hansen, 1897 [Fig. 9C] Continental shelf and slope (120–906 m), collected from fishes or soft bottom habitats; San Simeon, California, USA and Acapulco, Mexico. Type locality (lectotype): Mexico, off Acapulco. Body length to ~ 41.4 mm. See Hansen (1897), Richardson (1898, 1899, 1900b, 1905a), Schultz (1969), Brusca and France (1992), Espinosa-Pérez and Hendrickx (2001a, 2006), Bruce (2009a), and García-Madrigal et al. (2022).
Rocinelamurilloi Brusca & Iverson, 1985 [Fig. 9D] Continental slopes and basins (700–1866 m), collected from fishes or soft-bottom habitats; Point Sur, California, USA to Arica, Chile. Type locality: Costa Rica, 14 miles 233°T from Point Guiones at the mouth of Golfo de Nicoya (9°45'N, 85°55'W). Body length to ~ 35 mm. See Brusca and Iverson (1985), Wetzer et al. (1991), Brusca and France (1992), Espinosa-Pérez and Hendrickx (2001a), Hendrickx (2008, 2018a), Bruce (2009a), and Azofeifa-Solano and Cortés (2020). Note: Although R.murilloi has also been listed as occurring in mangrove lagoons or estuaries in Baja California Sur (table 15.1 in Whitmore et al. 2005), we suspect that record may be erroneous as the species is otherwise known only from offshore waters of 700 m or deeper.
Rocinelasignata Schioedte & Meinert, 1879 [Fig. 9E] Mostly intertidal to shallow subtidal in embayments and on the offshore continental shelf (0–68 m), collected from fishes or soft-bottom habitats; Humboldt Bay, northern California, USA to Baja California and the Gulf of California, Mexico, and south to Costa Rica, Panama, Colombia and Ecuador, including the Galapagos Islands in the eastern Pacific, and in the western Atlantic, Caribbean, and Gulf of Mexico from Florida, USA to northeastern Brazil. Type locality (lectotype): West Indies, St. Croix Island. Body length to ~ 20.6 mm. See Schioedte and Meinert (1879), Richardson (1898, 1905a), Menzies and Glynn (1968), Bowman (1977), Brusca and Iverson (1985), Garzón-Ferreira (1990), Brusca and France (1992), Espinosa-Pérez and Hendrickx (2001a, 2002, 2006), Brusca et al. (2004), McLaughlin et al. (2005), Bunkley-Williams et al. (2006), Lazarus-Agudelo and Cantera-Kintz (2007), Bruce (2009a), Cavalcanti et al. (2012), Bastida-Zavala et al. (2013), Pires-Vanin et al. (2014), Fogg et al. (2016), Cardoso et al. (2017), Shaughnessy et al. (2017), Aguilar-Perera et al. (2018), Ortiz and Lalana (2018), Alves et al. (2019), Silva et al. (2019), Aguilar-Perera (2022), Aguilar-Perera and Nóh-Quiñones (2022), and García-Madrigal et al. (2022).
Family Cirolanidae
Cirolanadiminuta Menzies, 1962 [Fig. 7A] Intertidal to subtidal, on rocks, sandy silt sediments, algae, and member of the demersal zooplankton community (0–50 m); Point Conception, southern California, USA to western Baja California and Gulf of California, Mexico, and the Galapagos Islands. Type locality: Mexico, Baja California, Bahia de San Quintin. Body length to ~ 10.5 mm. See Menzies (1962a), Schultz (1969), Bruce and Bowman (1982), Stepien and Brusca (1985), Bruce (1986a), SCAMIT (1994–2021), Brusca et al. (1995, 2007), Chess and Hobson (1997), Espinosa-Pérez and Hendrickx (2001a), and McLaughlin et al. (2005).
Cirolanaharfordi (Lockington, 1877) [Fig. 7B] Intertidal to shallow subtidal, abundant in mussel beds on rocky shores, also occurs in mangrove lagoons or estuaries at southern end of the range; Vancouver Island, Canada to Magdalena Bay, Baja California Sur, Mexico. Type locality (probable syntypes; see Brusca et al. 1995): USA, California, Santa Rosa Island. Body length to ~ 20 mm. See Lockington (1877b original description as Aegaharfordii), Richardson (1899, 1900b, 1905a), Stafford (1913a), Shearer (1942), Hewatt (1946), Hatch (1947), George and Strömberg (1968), Schultz (1969), Miller (1975), Allen (1976), Johnson (1976a, 1976b), Gillard (1978), Kussakin (1979), Lee and Miller (1980), Bruce (1981), Ricketts et al. (1985), Brusca et al. (1995, 2007), Espinosa-Pérez and Hendrickx (2001a, 2002, 2006), McLaughlin et al. (2005), Whitmore et al. (2005), Campos and Villarreal (2008), SCAMIT (2011–2021), Roletto et al. (2014), Bugnot et al. (2015), Lafferty and Suchanek (2016), and Morales-Zarate et al. (2016).
Eurydicecaudata Richardson, 1899 [Fig. 7C] Intertidal to continental shelf (0–160 m), and occasionally on fishes (e.g., California scorpionfish and longfin sanddab); San Diego, southern California, USA to western Baja California Sur and the Gulf of California, Mexico, and south to Ecuador, including the Galapagos and other offshore islands of the tropical eastern Pacific. Type locality: USA, California, Catalina Island, Isthmus Cove. Body length to ~ 9 mm. See Richardson (1899, 1900b, 1905a), Menzies and Barnard (1959 as E.branchuropus), Schultz (1966 as E.branchuropus; 1969 as both E.branchuropus and E.caudata), Bowman (1977), Brusca and Iverson (1985), Bruce (1986a), Hobson and Chess (1986), SCAMIT (1994–2021), Brusca et al. (1995, 2004, 2007), Espinosa-Pérez and Hendrickx (2001a, 2002, 2006), Smith et al. (2001, 2003), Ranasinghe et al. (2003, 2007, 2012), McLaughlin et al. (2005), Kalman (2006), Campos and Villarreal (2008), and Gillett et al. (2017, 2019, 2022). Note: If confirmed, the Gillett et al. (2019) record extends the maximum depth range of this species slightly from 160 m to the upper slope between 200–500 m.
Excirolanachiltoni (Richardson, 1905) [Fig. 7D] Intertidal on sandy beaches; British Columbia, Canada to southern California, USA, as well as Japan, Taiwan, and Hong Kong. Type locality: USA, California, San Francisco. Body length to ~ 13.2 mm. See Richardson (1905a original description as Cirolanachiltoni), Richardson (1912), Schultz (1969), Klapow (1972), Iverson (1974), Miller (1975), Kussakin (1979), Bruce and Jones (1981), Dexter (1983), Bruce (1986a), Brusca et al. (1995, 2007), Kaneko and Omori (2005), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Ranasinghe et al. (2007, 2012), SCAMIT (2011–2021), Gillett et al. (2017), and Shaughnessy et al. (2017).
Excirolanalinguifrons (Richardson, 1899) [Fig. 7E] Intertidal on sandy beaches; Coos Bay, Oregon to southern California, USA. Type locality: USA, California, Monterey Bay. Body length to ~ 3.8 mm. See Richardson (1899 original description as Cirolanalinguifrons; 1900b, 1905a, 1912), Shearer (1942), Schultz (1969), Miller (1975), Kussakin (1979), Bruce (1986a), Brusca et al. (1995, 2007), SCAMIT (2001–2021), Ranasinghe et al. (2003), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), and Campos and Villarreal (2008).
Metacirolanajoanneae (Schultz, 1966) [Fig. 7F] Offshore submarine canyons and basins (218–500 m); Strait of Georgia, British Columbia, Canada to southern California, USA, including Santa Cruz Canyon. Type locality: USA, California, Santa Cruz Canyon (R/V Velero IV Station No. 6805-59; 33°35'59"N, 119°15'11"W). Body length to ~ 5 mm. See Schultz (1966 original description as Cirolanajoanneae, but with prior listing in Schultz 1964 designated a nomen nudum; 1969), Bruce (1981, 1986a), Lissner et al. (1986), Wetzer et al. (1991), Brusca et al. (1995), Wetzer and Brusca (1997), McLaughlin et al. (2005), Macdonald et al. (2010), and SCAMIT (2011–2021).
Natatolanacaliforniensis (Schultz, 1966) [Fig. 7G] Offshore submarine canyons and basins, on fine sand and muddy bottoms (792–1250 m); primarily southern California, USA (e.g., Coronado Canyon, Tanner Canyon), with a few records from the Gulf of California, Mexico and further south (Costa Rica and Peru-Chile Trench). Type locality: USA, California, Coronado Canyon offshore southern San Diego (R/V Velero IV Station No. 6851-60; 32°37'54"N, 118°55'40"W). Body length to ~ 13.4 mm. See Schultz (1966 original description as Cirolanacaliforniensis; 1969), Brusca and Ninos (1978), Bruce (1981, 1986a), Brusca and Iverson (1985), Wetzer et al. (1991), Brusca et al. (1995), Espinosa-Pérez and Hendrickx (2001a, 2006), McLaughlin et al. (2005), Keable (2006), Espinosa-Pérez et al. (2009), SCAMIT (2011–2021), Ranasinghe et al. (2012), Gillett et al. (2019), and Zúñiga Delgado et al. (2021).
Family Corallanidae
Excorallanatricornisoccidentalis Richardson, 1905 [Fig. 8A] Intertidal to continental shelf, on rocks, sandy beaches, in mangrove habitats, and as a commensal in sponges or temporary fish parasite (0–138 m); Catalina Island, southern California, USA to western Baja California Sur and the Gulf of California, Mexico, and south to Costa Rica and Panama. Type locality: Mexico, Gulf of California. Body length to ~ 11 mm. See Richardson (1905a), Bowman (1977), Delaney (1984, 1989, 1993), Brusca and Iverson (1985), Guzman et al. (1988), Espinosa-Pérez and Hendrickx (2001a, 2006), and Brusca et al. (2007).
Excorallanatruncata (Richardson, 1899) [Fig. 8B] Intertidal to continental shelf (0–183 m); Point Conception, southern California, USA to Galapagos Islands, including Gulf of California (Bahia Concepcion). Type locality: USA, California, Catalina Island. Body length to ~ 21.5 mm. See Richardson (1899 original description as Corallanatruncata; 1900b, 1905a), Delaney (1982, 1984, 1989), SCAMIT (1994–2021), Espinosa-Pérez and Hendrickx (2001a, 2002), McLaughlin et al. (2005), Brusca et al. (2007), Melwani and Kim (2008), and Gillett et al. (2019). Note: If confirmed, the Gillett et al. (2019) record extends the maximum depth range of this species slightly from 183 m to the upper slope between 200 and 500 m.
Family Cymothoidae
Ceratothoagaudichaudii (H. Milne Edwards, 1840) [Fig. 11A] Parasite of many species of pelagic fishes, including striped mullet off California, and pompano and herring from Baja California; southern California, USA to western Baja California, Mexico, and southward to Cape Horn and around to southern Patagonia, South America. Type locality: Chile, Coquimbo. Body length to ~ 55 mm. See H. Milne Edwards (1840 original description as Cymothoagaudichaudii), Richardson (1899, 1905a as Meinertiagaudichaudii), Menzies (1962b as Meinertiagaudichaudi), Schultz (1969), Brusca (1977, 1981), Brusca and Iverson (1985), Espinosa-Pérez and Hendrickx (2001a, 2002, 2006), McLaughlin et al. (2005), Whitmore et al. (2005), Brusca et al. (2007), and Martin et al. (2015). Note: Species listed as species inquirenda by Martin et al. (2015), Hadfield and Smit (2020), and Zúñiga Delgado et al. (2021). Note: See Endnote 6.
Ceratothoagilberti (Richardson, 1904) [Fig. 11B] Parasite in mouths of mullets and an unidentified flatfish; Newport Bay, Orange County, southern California, USA to western Baja California, Mexico, and the Gulf of California to Mazatlán. Type locality: Mexico, Mazatlán. Body length to ~ 29 mm. See Richardson (1904a original description as Meinertiagilberti; 1905a), Schultz (1969), Brusca (1977, 1981), Espinosa-Pérez and Hendrickx (2001a, 2006), Brusca et al. (2004), McLaughlin et al. (2005), Whitmore et al. (2005), Martin et al. (2015), Hadfield et al. (2016), and Hadfield and Smit (2020).
Elthusacalifornica (Schioedte & Meinert, 1884) [Fig. 11C] Parasite of dwarf surfperch, shiner surfperch, surf smelt, topsmelt, arrow goby, California killifish, and California grunion (nearshore to 90 m), common in bays and lagoons; Alaska, USA to Punta Eugenio, Baja California Sur, Mexico, and off Peru (but uncommon south of San Diego, California and north of Washington State). Type locality: USA, California, near San Francisco. Body length to ~ 15 mm. See Schioedte and Meinert (1884 original description as Livonecacalifornica, but genus misspelled as Lironeca), (Richardson 1899, 1900b, 1905a), Fee (1927), Hatch (1947), Schultz (1969), Iverson (1974), Miller (1975), Brusca (1977, 1981), Kussakin (1979), Bruce (1990), SCAMIT (1998–2021), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Espinosa-Pérez et al. (2009), Shaughnessy et al. (2017), van der Wal et al. (2019), and Zúñiga Delgado et al. (2021).
Elthusamenziesi (Brusca, 1981) [Fig. 11D] Parasite in gill chambers of the wooly sculpin, northern clingfish and reef finspot (tidepools to 30 m); Baja California Norte (e.g., Coronado Islands, Todos Santos Bay, San Quintin Bay, and Guadalupe Island) to Baja California Sur (e.g., Alijos Rocks) and the Gulf of California, Mexico. Type locality: Mexico, Baja California Norte, Guadalupe Island, Melpomene Cove. Body length to 21 mm. See Menzies (1962a as Lironeca sp. nov., but undescribed), Brusca (1981 original description as Lironecamenziesi), Campos-González et al. (1986), Bruce (1990), Espinosa-Pérez and Hendrickx (2001a, 2002, 2006), McLaughlin et al. (2005), Whitmore et al. (2005), and van der Wal et al. (2019).
Elthusavulgaris (Stimpson, 1857) [Fig. 11E] Parasite in gill chambers of at least 30 species of fishes (1–311 m); Coos Bay, Oregon, USA to Colombia, South America, and common from southern California, USA and the Gulf of California, Mexico to Costa Rica. Type locality: USA, California, San Francisco Bay. Body length to ~ 43 mm. See Stimpson (1857 original description as Livonecavulgaris), Richardson (1899, 1900b, 1905a), Shearer (1942), Hatch (1947), Schultz (1969), Miller (1975), Brusca (1978b, 1981), Kussakin (1979), Lee and Miller (1980), Brusca and Iverson (1985), Bruce (1990), Thompson et al. (1993), SCAMIT (1994–2021), Espinosa-Pérez and Hendrickx (2001a, 2006), Brusca et al. (2004, 2007), McLaughlin et al. (2005), Whitmore et al. (2005), Kalman (2006), Ranasinghe et al. (2007), Shaughnessy et al. (2017), van der Wal et al. (2019), and García-Madrigal et al. (2022).
Mothocyarosea Bruce, 1986 [Fig. 10D] Parasite of California halfbeak and skipper halfbeak; San Diego, southern California, USA to Magdalena Bay, Baja California Sur, Mexico, and Nicaragua. Type locality: Mexico, Baja California Sur, San Ignacio Lagoon (~ 26°25'N, 113°13'W). Body length to ~ 8.4 mm. See Bruce (1986b), Wetzer et al. (1991), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), and Espinosa-Pérez et al. (2009).
Nerocilaacuminata Schioedte & Meinert, 1881 [Fig. 10B] Parasite of ~ 40 species of fishes, especially common in shallow coastal lagoons and bays; southern California, USA to Peru, including the west coast of Baja California, Mexico, the Gulf of California, Costa Rica, Peru, the offshore islands of Las Tres Marias and the Galapagos, Hawaii, and in the southern Atlantic Ocean. Type locality: not specified, but the types for N.acuminata and its junior synonym N.californica as designated by Brusca (1981) are deposited at Muséum National d’Histoire Naturelle, Paris. Body length to ~ 25 mm. See Schioedte and Meinert (1881), Richardson (1900b, 1905a), Schultz (1969), Miller (1975), Brusca (1977, 1978a, 1981), Kussakin (1979), Brusca and Iverson (1985), SCAMIT (1994–2021), Espinosa-Pérez and Hendrickx (2001a, 2002, 2006), Brusca et al. (2004, 2007), McLaughlin et al. (2005), Whitmore et al. (2005), Fogg et al. (2016), Esteves-Silva et al. (2020), Zúñiga Delgado et al. (2021), Aguilar-Perera (2022), Bueno et al. (2022), and García-Madrigal et al. (2022). Note: See Brusca (1981: 157–158) for descriptions and discussion of the acuminata and aster forms of N.acuminata. Additionally, although Brusca (1981) synonymized N.acuminata with N.californica Schioedte & Meinert, 1881, both are currently accepted as separate species in WoRMS (Boyko et al. 2008 onwards). Also see Williams and Bunkley-Williams (2003) for additional information. However, until a rigorous morphological and molecular comparison can be made, we are continuing to treat these as the single synonymized species N.acuminata in agreement with Brusca (1981).
Renocilathresherorum Williams & Bunkley-Williams, 1980 [Fig. 10A] Parasite of barspot cardinalfish and Panamic fanged blenny in the Eastern Pacific (i.e., Corona del Mar, southern California, USA; Magdalena Bay, Baja California Sur and the central to southern Gulf of California, Mexico), and of Egyptian sole, European bass, redbelly tilapia, and thinlip grey mullet in the Suez Canal region of Egypt (e.g., Lake Qaroun). Type locality: Mexico, Baja California Sur, Loreto. Body length to ~ 30.5 mm. See Williams and Williams (1980), Brusca (1981), Williams and Williams (1987), Espinosa-Pérez and Hendrickx (2006), Kerstitch and Bertsch (2007), Eman et al. (2014), Mahmoud et al. (2016), Ali and Aboyadak (2018), and Aneesh et al. (2020, 2021). Note: See Endnote 5.
Smenispaconvexa (Richardson, 1905) [Fig. 10C] Parasite in mouths or gill chambers of Pacific bumper, pompanos, serranos, and unidentified carangids; southern California, USA to the Gulf of Guayaquil, Ecuador, and Peru. Type locality (neotypes; see Brusca 1981): Mexico, Nayarit, Playa Noviella, west of Tecuelao. Body length to ~ 15 mm. See Richardson (1905c original description as Livonecaconvexa), Menzies et al. (1955), Schultz (1969), Brusca (1977, 1981), Brusca and Iverson (1985), Wetzer et al. (1991), SCAMIT (1998–2021), Espinosa-Pérez and Hendrickx (2001a, 2006), McLaughlin et al. (2005), Whitmore et al. (2005), Kalman (2006), Brusca et al. (2007), Özdikem (2009), and Zúñiga Delgado et al. (2021).
Family Gnathiidae
Caecognathiacrenulatifrons (Monod, 1926) [Fig. 12A] Continental shelf, slope, and submarine canyons, benthic sediments (9–1376 m); Strait of Georgia, British Columbia, Canada to Todos Santos Bay, Baja California Norte, Mexico. Type locality: USA, California, La Jolla. Male body length to ~ 4.3 mm. See Monod (1926 original description as Gnathiacrenulatifrons), Menzies and Barnard (1959), Schultz (1964, 1966, 1969), Iverson (1974), Miller (1975), Lissner et al. (1986), Thompson et al. (1993), SCAMIT (1994–2021), Wetzer and Brusca (1997), Bergen et al. (1998), Smit and Davies (2004), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Haney (2006), Ranasinghe et al. (2007, 2012), Campos and Villarreal (2008), Espinosa-Pérez et al. (2009), Macdonald et al. (2010), and Gillett et al. (2017, 2019, 2022).
Caecognathiasanctaecrucis (Schultz, 1972) [Fig. 12B] Continental shelf, slope, and submarine canyons, benthic sediments (76–218 m); Strait of Georgia, British Columbia, Canada to Todos Santos Bay, Baja California Norte, Mexico. Type locality: USA, California, Santa Cruz Canyon (R/V Velero IV Station No. 6805-59; 33°56'03"N, 119°52'03"W). Male body length ~ 2.6 mm. See Schultz (1966 original description as Gnathiahirsuta; 1972 for “substitution for preoccupied name”), Lissner et al. (1986), Wetzer et al. (1991), SCAMIT (1994–2021), Wetzer and Brusca (1997), Bergen et al. (1998), Smit and Davies (2004), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Haney (2006), Campos and Villarreal (2008), Macdonald et al. (2010), and Gillett et al. (2017).
Caecognathia sp. A [No figure] Continental shelf (110–126 m); Western Santa Barbara Channel off Point Conception, southern California, USA. Male body length to ~ 4.3 mm. See Haney (2006) and SCAMIT (2008–2021). Note: See Endnote 8.
Caecognathia sp. SD1 [No figure] Continental shelf, soft-bottom benthos (116–153 m); San Diego, southern California, USA. Male body length ~ 3.4 mm. See Haney (2006), SCAMIT (2008–2021), Ranasinghe et al. (2012), and Gillett et al. (2022). Note: See Endnote 8.
Gnathiaclementensis Schultz, 1966 [Fig. 12C] Continental shelf, benthic sediments with manganese nodules (162 m); San Clemente Canyon, southern California, USA (known only from type locality). Type locality: USA, California, San Clemente Canyon (R/V Velero IV Station No. 6840-60; 32°44'35"N, 118°12'45"W). Male body length to ~ 8.5 mm. See Schultz (1966, 1969), Wetzer et al. (1991), Smit and Davies (2004), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), and Haney (2006).
Gnathiacoronadoensis Schultz, 1966 [Fig. 12D] Offshore submarine canyons, benthic sediments (344–812 m); Coronado Canyon, southern California, USA. Type locality: USA, California, Coronado Canyon (R/V Velero IV Station No. 6851-60; 32°30'42"N, 117°21'37"W). Male body length to ~ 3.5 mm. See Schultz (1966, 1969), Wetzer et al. (1991), Smit and Davies (2004), McLaughlin et al. (2005), and Haney (2006).
Gnathiaproductatridens Menzies & Barnard, 1959 [Fig. 12E] Continental shelf, benthic sediments (20–164 m); Point Conception, Santa Barbara County, southern California, USA. Type locality: USA, California, off Summerland (R/V Velero IV Station No. 5173-57; 34°14'50"N, 119°32'25"W). Male body length to ~ 3.2 mm. See Menzies and Barnard (1959), Schultz (1969), Lissner et al. (1986), Wetzer et al. (1991), SCAMIT (1996–2021), Wetzer and Brusca (1997), Smit and Davies (2004), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Haney (2006), Ranasinghe et al. (2012), and Gillett et al. (2019). Note: If confirmed, the Gillett et al. (2019) record extends the maximum depth range of this species slightly from 164 m to the upper slope between 200–500 m.
Gnathiasteveni Menzies, 1962 [Fig. 12F] Intertidal; Strait of Georgia, British Columbia, Canada to San Quintin Bay, Baja California Norte, Mexico. Type locality: Mexico, Baja California, Bahia de San Quintin. Male body length to ~ 2.3 mm. See Menzies (1962a), Schultz (1969), Wetzer et al. (1991), Fairey et al. (2002), Smit and Davies (2004), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Haney (2006), Brusca et al. (2007), SCAMIT (2008–2021), and Macdonald et al. (2010).
Gnathiatridens Menzies & Barnard, 1959 [Fig. 12G] Shallow subtidal (11–27 m); Strait of Georgia, British Columbia, Canada to San Clemente Island, southern California, USA. Type locality: USA, California, 11 miles east of Point Conception (R/V Velero IV Station No. 4822-57; 34°27'15"N, 120°14'45"W). Male body length to ~ 3 mm. See Menzies and Barnard (1959), Schultz (1969), Lissner et al. (1986), Wetzer et al. (1991), SCAMIT (1994–2021), Wetzer and Brusca (1997), Smit and Davies (2004), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Haney (2006), Espinosa-Pérez et al. (2009), Macdonald et al. (2010), and Henkel et al. (2020).
Gnathiatrilobata Schultz, 1966 [Fig. 12H] Continental shelf and submarine canyons, collected from polychaete tubes and benthic sediments (98–976 m); Strait of Georgia, British Columbia, Canada to Point Loma, San Diego, southern California, USA. Type locality: USA, California, Coronado Canyon (R/V Velero IV Station No. 6851-60; 32°30'42"N, 117°21'37"W). Male body length to ~ 5 mm. See Schultz (1966, 1969), Wetzer et al. (1991), SCAMIT (1994–2021), Bergen et al. (1998), Smit and Davies (2004), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Haney (2006), Ranasinghe et al. (2007), and Macdonald et al. (2010).
Gnathia sp. MBC1 [No figure] Shallow rocky reef, collected from a white sponge (~ 11 m); Pequenot’s Reef, Corona del Mar, southern California, USA. Male body length ~ 2.2 mm. See Haney (2006) and SCAMIT (2008–2021). Note: See Endnote 8.
Family Tridentellidae
Tridentellaglutacantha Delaney & Brusca, 1985 [Fig. 8C] Continental shelf and upper slope, on green muds or rocky bottoms (128–360 m); Farallon Islands, central California to Catalina Island and Los Angeles, southern California, USA. Type locality: USA, California, west of North Farallon Islands (37°43.8'N; 123°11.3'W). Body length to ~ 18 mm. See Delaney and Brusca (1985), Wetzer et al. (1991), Kensley and Heard (1997), Wetzer and Brusca (1997), Brandt and Poore (2001), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Espinosa-Pérez et al. (2009), and SCAMIT (2013–2021).
Tridentellaquinicornis Delaney & Brusca, 1985 [Fig. 8D] Continental shelf, on “rock” substrates (depth of at least 53 m); Santa Barbara Island to near Catalina Island, southern California, USA. Type locality: USA, California, Los Angeles County, Farnsworth Bank near Catalina Island (R/V Velero IV Station No. 1903-49). Body length to ~ 11 mm. See Delaney and Brusca (1985), Wetzer et al. (1991), SCAMIT (1996–2021), Kensley and Heard (1997), Brandt and Poore (2001), McLaughlin et al. (2005), and Espinosa-Pérez and Hendrickx (2006).
Infraorder Epicaridea
Superfamily Bopyroidea
Family Bopyridae
Anathelgeshyphalus (Markham, 1974) [Fig. 13C] Branchial parasite of hermit crabs Parapaguroideslaurentae and P.makarovi, deep water (69–319 m); Carmel, California, USA to western Baja California Norte, Mexico. Type locality: USA, California, Santa Barbara, Channel Islands (R/V Velero III Station No. 1026-39; 33°00'N; 118°38'W) infesting P.makarovi. Female body length to ~ 6.1 mm (male ~ 2.6 mm). See Markham (1974a original description as Stegophryxushyphalus; 1992, 2003, 2020), Wetzer et al. (1991), SCAMIT (2001–2021),Boyko and Williams (2003), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), and Stebbins (2012b).
Aporobopyrusmuguensis Shiino, 1964 [Fig. 14B] Branchial parasite of the porcelain crabs Pachychelesholosericus, P.pubescens, and P.rudis, shallow water (10–12 m); Bodega Bay, northern California, USA to central western Baja California, Mexico. Type locality: USA, California, Point Mugu, off Mugu Pier infesting P.rudis. Female body length to ~ 4 mm (male ~ 2.7 mm). See Shiino (1964), Miller (1975), Lee and Miller (1980), Campos-González and Campoy-Favela (1987), Markham (1992, 2008), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Stebbins (2012b), and Aguilar-Perera (2022).
Aporobopyrusoviformis Shiino, 1934 [Fig. 14C] Branchial parasite of the porcelain crabs Pachychelespubescens and Petrolisthespubescens, shallow water (10–12 m); Point Mugu, California, USA, and Seto, Japan. Type locality: Japan, Seto, Yusaki infesting Petrolisthespubescens. Female body length to ~ 5.8 mm (male ~ 2.5 mm). See Shiino (1934), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), and Stebbins (2012b).
Argeiapugettensis Dana, 1853 [Fig. 14A] Branchial parasite of multiple species of crangonid and hippolytid shrimp (intertidal to 188 m); Bering Sea to southern California, and the US-Mexico border, Japan, and Korea. Type locality: USA, Washington, Puget Sound infesting Metacrangonmunitus. Female body length to ~ 14 mm (male ~ 5.5 mm). See Dana (1853), Richardson (1899, 1900c, 1905a), Fee (1927), Shearer (1942), Hatch (1947), George and Strömberg (1968), Miller (1975), Markham (1977b, 1992), Higley and Holton (1978), Ricketts et al. (1985), Jay (1989), SCAMIT (2001–2021), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Espinosa-Pérez et al. (2009), Puget Sound Institute (2012–2023), and Stebbins (2012b).
Asymmetrioneambodistorta Markham, 1985 [Fig. 14D] Branchial parasite of the hermit crab Isochelespilosus, shallow water (3 m): southern California, USA. Type locality: USA, California, Newport Beach, Corona del Mar State Beach. Female body length to ~ 5.4 mm (male ~ 2.8 mm). See Markham (1985b, 2003), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al, (2007), and Stebbins (2012b).
Bathygygegrandis Hansen, 1897 [Fig. 13B] Branchial parasite of deep-water shrimp of the genus Glyphocrangon (1792–1896 m in SCB); worldwide distribution, including San Clemente Basin, southern California, USA. Type locality: Mexico, off Acapulco (21°15'N, 106°23'W) infesting G. spinulosa. Female body length to ~ 12.7 mm (male ~ 6.8 mm). See Hansen (1897), Richardson (1899, 1905a), Espinosa-Pérez and Hendrickx (2001a), Stebbins (2012b), Markham (2016, 2020), and García-Madrigal et al. (2022).
Bopyrellacalmani (Richardson, 1905) [Fig. 15A] Branchial parasite of alpheid snapping shrimps Alpheopsisequidactylus and Synalpheuslockingtoni, (intertidal to 9 m); Central to southern California, USA. Type locality: USA, California, between Santa Barbara and San Nicolas Island. Female body length to ~ 8 mm (male ~ 2.2 mm). See Richardson (1905a original description as Bopyriscuscalmani), Sassaman et al. (1984), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), and Stebbins (2012b).
Capitetragonia sp. A [Fig. 15B, image of representative species C.alphei] Branchial parasite of the undescribed alpheid shrimp Automate sp. A (30 m depth); collected offshore of Palos Verdes, Los Angeles County, southern California, USA. No body size information available, but total length for the type species of the genus, Capitetragoniaalphei (Richardson, 1900a), measured 5.1 mm (see Markham 1985a, p. 43). See SCAMIT (2012–2021) and Stebbins (2012b). Note: Provisional species originally described as Probopyria sp. A (see Endnote 9).
Eremitionegiardi (Calman, 1898) [Fig. 15C] Branchial parasite of hermit crabs of the genus Pagurus; Bering Sea to Palos Verdes, southern California, USA. Type locality: USA, Washington, Puget Sound infesting P.armatus. Female body length to ~ 12 mm (male ~ 3 mm). See Calman (1898 original description as Pseudionegiardi), Richardson (1899, 1900c, 1905a), Hatch (1947), George and Strömberg (1968), Markham (1974b), Boyko and Williams (2004), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), SCAMIT (2011–2021), Stebbins (2012b), and Williams et al. (2019). Note: Record of species in California waters questionable (see Endnote 10).
Leidyainfelix Markham, 2002 [Fig. 13D] Branchial parasite of the grapsid crab Pachygrapsuscrassipes (intertidal); southern California, USA to Baja California Norte, Mexico. Type locality: Mexico, Baja California Norte, San Quintín (30°28'N; 115°58'W). Female body length to ~ 7.9 mm (male ~ 1 mm). See Markham (2002), Boyko (2003), and Stebbins (2012b).
Munidionpleuroncodis Markham, 1975 [Fig. 13E] Branchial parasite of the pelagic galatheid crab Pleuroncodesplanipes; Central California, USA to central Mexico, typically occurring in southern California during warm water years (e.g., El Niños). Type locality: Mexico, Baja California (26°22'N, 115°05'W). Female body length to ~ 9.8 mm (male ~ 4.4 mm). See Markham (1975), Lissner et al. (1986), Wetzer and Brusca (1997), SCAMIT (2001–2021), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), and Stebbins (2012b).
Orthionegriffenis Markham, 2004 [Fig. 14E] Branchial parasite of mud shrimp Upogebiapugettensis and U.macginitieorum; introduced species from Asia (China, Japan), occurring from British Columbia, Canada to southern California, USA along west coast of North America. Type locality: USA, Oregon, Yaquina Bay, Idaho Inlet mudflats (44°35.4'N, 124°01.5'W) infesting U.pugettensis. Female body length to ~ 24 mm (male to ~ 10.3 mm). See Markham (2004), Williams and An (2009), Dumbauld et al. (2011), Chapman et al. (2012), Puget Sound Institute (2012–2023), Stebbins (2012b), SCAMIT (2013–2021), Hong et al. (2015), and Whalen et al. (2020).
Phyllodurusabdominalis Stimpson, 1857 [Fig. 13F] Abdominal parasite of the mud shrimp Upogebialepta, U.macginitieorum, and U.pugettensis (intertidal): British Columbia, Canada to Baja California Norte, Mexico. Type locality (see Markham 1977a): USA, Washington, Puget Sound and California, Tomales Bay. Female body length to ~ 14 mm (male ~ 6 mm). See Stimpson (1857), Richardson (1899, 1900c, 1905a), Fee (1927), Hatch (1947), George and Strömberg (1968), Miller (1975), Markham (1977a, 1992), Lee and Miller (1980), Campos-González and Campoy-Favela (1987), SCAMIT (1994–2021), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Stebbins (2012b), and Aguilar-Perera (2022).
Progebiophilusbruscai Salazar-Vallejo and Leija-Tristán, 1990 [Fig. 14F] Branchial parasite of the mud shrimp Upogebiadawsoni, U.spinigera, U.macginitieorum, and Pomatogebiarugosa; Todos Santos Bay, Ensenada, Baja California Norte, Mexico to Tortugas Bay, Baja California Sur, the Gulf of California, and Nicaragua. Type locality: Mexico, Baja California Sur, Bahía de la Paz, Laguna de La Paz, El Comitán infesting U.dawsoni. Female body length to ~ 12 mm (male ~ 3.9 mm). See Salazar-Vallejo and Leija-Tristán (1990), Campos and de Campos (1998), Espinosa-Pérez and Hendrickx (2001a, 2006), Markham (2005), Boyko et al. (2017), and Aguilar-Perera (2022).
Pseudionegalacanthae Hansen, 1897 [Fig. 15D] Branchial parasite of the squat lobsters Galacanthadiomedeae and Munidaquadrispina, deep water (> 50 m); British Columbia, Canada to Gulf of California, Mexico. Type locality: Mexico, central Gulf of California (Albatross Station No. 3435; 26°48'00"N, 110°45'20"W) infesting Galacanthadiomedeae. Female body length to ~ 11 mm (male ~ 4.8 mm). See Hansen (1897), Richardson (1899, 1905a), Fee (1927), Hatch (1947), George and Strömberg (1968), Markham (1974b), Espinosa-Pérez and Hendrickx (2001a, 2006), Boyko and Williams (2004), McLaughlin et al. (2005), and Stebbins (2012b). Note: Record of species in California waters questionable (J Markham, pers. comm.), nor is this species listed within SCB waters in Markham (2020).
Schizobopyrinastriata (Nierstrasz & Brender à Brandis, 1929) [Fig. 15E] Branchial parasite of the alpheoid shrimps Hippolytecaliforniensis in San Diego Bay and Thoralgicola in the Gulf of California, shallow water; California, USA to Baja California, Mexico, and the Gulf of California. Type locality: USA, California, San Diego, San Diego Bay. Female body length to ~ 4 mm. See Nierstrasz and Brender à Brandis (1929 original description as Bopyrinastriata), Ricketts et al. (1985), Campos and de Campos (1990), Markham (1992), Espinosa-Pérez and Hendrickx (2001a, 2006), McLaughlin et al. (2005), Brusca et al. (2007), and Stebbins (2012b).
Family Ionidae
Ionecornuta Spence Bate, 1863 [Fig. 13A] Branchial parasite of ghost shrimps of the genera Callianassa and Neotrypaea in the eastern Pacific, and Nihonotrypaea and Upogebia in Asia, intertidal to shallow water; British Columbia, Canada to California, USA, the Gulf of California, Mexico, Japan, Korea, and China. Type locality: Canada, British Columbia, Vancouver Island, infesting Neotrypaeagigas (originally as Callianassalongimana). Female body length to ~ 12 mm. See Bate (1863), Richardson (1899, 1900c, 1905a), Hatch (1947), Miller (1975), Markham (1992), Nobuhiro and Kyoko (2004), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), An et al. (2009), Espinosa-Pérez et al. (2009), Stebbins (2012b), and Boyko et al. (2017).
Superfamily Cryptoniscoidea
Family Dajidae
Holophryxusalaskensis Richardson, 1905 [Fig. 16A] Ectoparasite on carapace of the Pacific glass shrimp Pasiphaeapacifica, mostly deep waters (44–1200 m); Prince William Sound, Alaska, USA to offshore of the US-Mexico border. Type locality: USA, Alaska, Behm Canal, vicinity of Yes Bay (~ 55°51'05"N, 131°49'43"W). Adult female body length to ~ 13.6 mm (male ~ 2.0 mm). See Richardson (1905a, 1905b, 1908), Butler (1964), Coyle and Mueller (1981), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), SCAMIT (2012–2021), Stebbins (2012b), Markham (2020), and Boyko and Williams (2021b). Note: See Endnote 11.
Zonophryxus sp. [Fig. 16B, image of representative species Z.probisowa] Ectoparasite on carapace of the caridean shrimps Pantomusaffinis and Plesionikatrispinus, offshore (98–305 m); southern California, USA between Point Dume and Imperial Beach. Body length = no information available. See Montagne and Cadien (2001), SCAMIT (2012–2021), Markham (2020), and Boyko and Williams (2021a). Note: See Endnote 12.
Family Hemioniscidae
Hemioniscusbalani Buchholz, 1866 [Fig. 16C] Parasite on barnacles of genera Balanus and Chthamalus, intertidal; worldwide distribution, including from at least British Columbia, Canada to Baja California, Mexico along the west coast of North America. Type locality: Norway, Kristiansand. See Buchholz (1866), Nierstrasz and Brender à Brandis (1926), Crisp (1968), Campos-González and Campoy-Favela (1987), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), SCAMIT (2012–2021), and Stebbins (2012b).
Suborder Limnoriidea
Superfamily Limnorioidea
Family Limnoriidae
Limnoriaalgarum Menzies, 1957 [Fig. 17A] Burrowing into algal holdfasts, intertidal to subtidal (0–15 m); Strait of Georgia, British Columbia, Canada to San Diego, southern California, USA. Type locality: USA, California, Marin County, Dillon Beach. Body length to ~ 5.5 mm. See Menzies (1957), Menzies and Barnard (1959), Schultz (1969), Miller (1975), Kussakin (1979), Lee and Miller (1980), Ricketts et al. (1985), Cookson (1991), SCAMIT (1996–2021), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Espinosa-Pérez et al. (2009), Macdonald et al. (2010), and Roletto et al. (2014).
Limnoriaquadripunctata Holthuis, 1949 [Fig. 17B] Burrowing into wood, intertidal to subtidal (0–30 m); worldwide cool temperate distribution, including Humboldt Bay, northern California to San Diego, southern California, USA. Type locality: North Sea coast of Dutch province of Zuid-Holland between villages of Katwijk and Noordwijk, not far from Leiden. Body length to ~ 3.4 mm. See Holthuis (1949), Menzies and Mohr (1952), Menzies (1957), Schultz (1969), Miller (1975), Kussakin (1979), Lee and Miller (1980), Ricketts et al. (1985), Cookson (1991, 1997), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Espinosa-Pérez et al. (2009), SCAMIT (2012–2021), Borges and Costa (2014), and Borges et al. (2014).
Limnoriatripunctata Menzies, 1951 [Fig. 17C] Burrowing into wood, intertidal to subtidal (0–7 m); temperate and tropical locations worldwide, including Puget Sound, Washington, USA to Mazatlán, Mexico, and the Gulf of California in the eastern Pacific. Type locality: USA, California, San Diego County, Mission Bay. Body length to ~ 4 mm. See Menzies (1951a, 1957), Menzies and Glynn (1968), Schultz (1969), Bastida and Torti (1972), Miller (1975), Allen (1976), Lee and Miller (1980), Brusca and Iverson (1985), Ricketts et al. (1985), Cookson (1991), Wetzer et al. (1991), Espinosa-Pérez and Hendrickx (2001a, 2006), Cohen et al. (2005), McLaughlin et al. (2005), Brusca et al. (2007), SCAMIT (2008–2021), Puget Sound Institute (2012–2023), Borges and Costa (2014), Borges et al. (2014), Shaughnessy et al. (2017), Ortiz and Lalana (2018), and Abdelsalam and Ramadan (2019).
Suborder Sphaeromatidea
Superfamily Seroloidea
Family Serolidae
Heteroseroliscarinata (Lockington, 1877) [Fig. 18A] Intertidal to continental shelf, soft bottom benthos (0–98 m); Santa Monica Bay, southern California, USA to San Quintin Bay, Baja California Norte, Mexico, and the Gulf of California. Type locality: USA, California, San Diego. Body length to ~ 6.4 mm. See Lockington (1877a original description as Seroliscarinatus), Richardson (1899, 1900b, 1905a), Menzies and Barnard (1959), Menzies (1962a), Schultz (1969), Hessler (1972), Glynn (1976), Dexter (1983), Lissner et al. (1986), SCAMIT (1994–2021), Bergen et al. (1998), Espinosa-Pérez and Hendrickx (2001a, 2006), Smith et al. (2001, 2003), Ranasinghe et al. (2003, 2007, 2012), McLaughlin et al. (2005), Reed and Hovel (2006), Sirota and Hovel (2006), Brusca et al. (2007), Campos and Villarreal (2008), Melwani and Kim (2008), Bruce (2009b), and Gillett et al. (2017). Note: See Endnote 13.
Superfamily Sphaeromatoidea
Family Ancinidae
Ancinusgranulatus Holmes & Gay, 1909 [Fig. 19A] Intertidal to shallow subtidal (0–10 m); Santa Barbara, southern California, USA to the Gulf of California, Mexico. Type locality: USA, California, near Coronado Island from depth of 3 fathoms. Body length to ~ 9.5 mm. See Holmes and Gay (1909), Loyola e Silva (1971), Schultz (1973), Glynn and Glynn (1974), Pires (1987), SCAMIT (1998–2021), Espinosa-Pérez and Hendrickx (2001a, 2006), Ranasinghe et al. (2003, 2007), Brusca et al. (2004, 2007), McLaughlin et al. (2005), Campos and Villarreal (2008), and Rocha-Ramírez et al. (2010). Note: See Endnote 14.
Ancinusseticomvus Trask, 1971 [Fig. 19B] Low intertidal on sandy beaches; Santa Barbara, southern California, USA to Mazatlán, Mexico, and the Gulf of California. Type locality: USA, California, Santa Barbara County, Coal Oil Beach (~ 34°28'N; 119°50'W). Body length to 10.5 mm. See Trask (1971), Schultz (1973), Glynn and Glynn (1974), Dexter (1976), Iverson (1982), Iverson and Chivers (1984), Pires (1987), Wetzer et al. (1991), McLaughlin et al. (2005), and Rocha-Ramírez et al. (2010). Note: Ancinusseticomvus is listed as a junior synonym of A.granulatus in SCAMIT (1998–2021) and Brusca et al. (2007). See Endnote 14 for a summary of the taxonomic history of A.seticomvus.
Bathycopeadaltonae (Menzies & Barnard, 1959) [Fig. 19C] Sandy subtidal sediments (17–137 m); Columbia River Estuary, Washington State to San Miguel Island and White’s Point, Los Angeles County, southern California. USA. Type locality: USA, California, off Point Conception (R/V Velero IV Station No. 4819-57; 34°26'30"N, 120°28'10"W). Body length to ~ 4 mm. See Menzies and Barnard (1959 original description as Ancinusdaltonae), Loyola e Silva (1971), Schultz (1973), Iverson and Chivers (1984), Miller (1975), Higley and Holton (1978), Kussakin (1979), Wetzer et al. (1991), SCAMIT (2001–2021), Ranasinghe et al. (2003), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Melwani and Kim (2008), Shimomura (2008), and Henkel et al. (2020).
Family Sphaeromatidae
Discerceisgranulosa (Richardson, 1899) [Fig. 20C] Subtidal (37 m); southern California, USA to Cedros Island, Baja California Norte, Mexico. Type locality: Mexico, Baja California, Cedros Island. Body length to ~ 9 mm. See Richardson (1899 original description as Cilicaeagranulosa; 1905a, 1906), Schultz (1969), SCAMIT (1994–2021), McLaughlin et al. (2005), Schotte (2005), and Espinosa-Pérez and Hendrickx (2006).
Dynamenelladilatata (Richardson, 1899) [Fig. 22C] Intertidal; Tatoosh Island, Washington to southern California, USA. Type locality: USA, California, Monterey Bay. Body length to ~ 3 mm. See Richardson (1899 original description as Dynamenedilatata; 1900b, 1905a), Shearer (1942), Hewatt (1946), Hatch (1947), Schultz (1969), Miller (1975), Kussakin (1979), Lissner et al. (1986), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Espinosa-Pérez et al. (2009), SCAMIT (2011–2021), and Lafferty and Suchanek (2016). Note: See Endnote 17 regarding generic assignment of this species.
Dynamenellaglabra (Richardson, 1899) [Fig. 22D] Intertidal; Coos Bay, Oregon to San Diego, southern California, USA. Type locality: USA, California, Monterey Bay. Body length to ~ 3 mm. See Richardson (1899 original description as Dynameneglabra; 1900b, 1905a), Shearer (1942), Hewatt (1946), Hatch (1947), Schultz (1969), Miller (1975), Kussakin (1979), Chess and Hobson (1997), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Espinosa-Pérez et al. (2009), and SCAMIT (2011–2021). Note: See Endnote 17 regarding generic assignment of this species.
Dynamenellasheareri (Hatch, 1947) [Fig. 22E] Intertidal to shallow subtidal; San Juan Archipelago, Washington to southern California, USA. Type locality: USA, Oregon, Coos Bay. Body length to ~ 3.3 mm. See Hatch (1947 original description as Dynamenesheareri), George and Strömberg (1968), Schultz (1969), Miller (1975), Kussakin (1979), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Toft and Cordell (2006), Brusca et al. (2007), Toft et al. 2008, 2010), Espinosa-Pérez et al. (2009), SCAMIT (2011–2021), and Lafferty and Suchanek (2016).
Dynoideselegans (Boone, 1923) [Fig. 22A] Intertidal; southern California, USA from Santa Barbara County to San Diego, and possibly further south to Cedros Island, Baja California, Mexico. Type locality: USA, California, San Diego County, La Jolla, Scripps Institution of Oceanography (~ 32.27°N, 117.61°W). Body length to ~ 7.4 mm. See Boone (1923 original description as Clianellaelegans), Hewatt (1946), Li (2000), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Wetzer and Mowery (2017), SCAMIT (2018–2021), and Khalaji-Pirbalouty and Gagnon (2021). Note: SCB records of Dynoideselegans identified as D.saldani in SCAMIT (2012–2016).
Exosphaeromaamplicauda (Stimpson, 1857) [Fig. 23A] Intertidal; Marin, Sonoma, and San Mateo Counties, northern California, USA (as redefined by Wall et al. 2015). Type locality (neotype): USA, California, Marin County, Tomales Bay (38.201°N, 122.922°W). Body length to ~ 8.4 mm. See Stimpson (1857 original description as Sphaeromaamplicauda), Richardson (1900b, 1905a), Hewatt (1946), George and Strömberg (1968), Miller (1975), Allen (1976), Kussakin (1979), Lee and Miller (1980), Bruce (2003), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Espinosa-Pérez et al. (2009), SCAMIT (2011–2021), Wall et al. (2015), Lafferty and Suchanek (2016), and Gillett et al. (2022). Note: See Endnote 18 regarding restricted range of E.amplicauda based on review by Wall et al. (2015).
Exosphaeromaaphrodita Boone, 1923 [Fig. 23B] Intertidal to shallow subtidal, on pier pilings and amongst detritus at the base of pilings; La Jolla, southern California, USA. Type locality (lectotype): USA, California, San Diego County, La Jolla, Scripps Institution of Oceanography pier pilings. Body length to ~ 8.7 mm. See Boone (1923), Bruce (2003), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Wall et al. (2015), and SCAMIT (2016–2021). Note: See Endnote 18.
Exosphaeromainornata Dow, 1958 [Fig. 23C] Intertidal to shallow subtidal, in holdfasts of the kelp Macrocystispyrifera; Puget Sound, Washington, USA to Todos Santos Bay, Baja California Norte, Mexico. Type locality: USA, California, Los Angeles County, San Pedro, Point Fermin (~ 33°40'N, 118°20'W). Body length to ~ 6 mm. See Dow (1958), Schultz (1969), Iverson (1974, 1978, 1982), Miller (1975), Wetzer et al. (1991), Bruce (2003), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Campos and Villarreal (2008), Espinosa-Pérez et al. (2009), SCAMIT (2011–2021), Puget Sound Institute (2012–2023), Roletto et al. (2014), and Wall et al. (2015).
Exosphaeromapentcheffi Wall, Bruce & Wetzer, 2015 [Fig. 23D] High to low rocky intertidal, amongst barnacles, algal turf, and Phragmatopoma worm tubes on undersides of rocks; Palos Verdes, southern California, USA. Type locality: USA, California, Los Angeles County, Palos Verdes Peninsula, Point Fermin (33.71°N, 118.3°W). Body length to ~ 6.8 mm. See Wall et al. (2015) and SCAMIT (2015–2021). Note: Records of E.pentcheffi reported as provisional species Exosphaeroma sp. A in SCAMIT (2012–2014).
Exosphaeromarhomburum (Richardson, 1899) [Fig. 23E] Intertidal to 26 m; Tatoosh Island, Washington, USA to Baja California Norte, Mexico (~ 4.3–6.7 km south of the US-Mexico border). Type locality: USA, California, Monterey Bay. Body length to ~ 4 mm. See Richardson (1899 original description as Sphaeromarhomburum; 1900b, 1905a), Shearer (1942), Schultz (1969), Miller (1975), Kussakin (1979), Hobson and Chess (1986), SCAMIT (1994–2021), Chess and Hobson (1997), Bruce (2003), Ranasinghe et al. (2003), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Wall et al. (2015), Lafferty and Suchanek (2016), and Looby and Ginsburg (2021). Note: See Endnote 19.
Gnorimosphaeromanoblei Menzies, 1954 [Fig. 20A] High intertidal, brackish to fully freshwater habitats; Del Norte County, northern California to Palos Verdes, Los Angeles County, southern California, USA. Type locality: USA, California, Marin County, Tomales Bay, Marshall (38.162°N, 122.89°W). Body length to ~ 2.9 mm. See Menzies (1954), Schultz (1969), Iverson (1974), Miller (1975), Kussakin (1979), Ricketts et al. (1985), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Espinosa-Pérez et al. (2009), and Wetzer et al. (2021).
Gnorimosphaeromaoregonense (Dana, 1853) [Fig. 20B] Intertidal to subtidal (0–21 m); Vancouver, British Columbia, Canada to San Francisco, California, USA. Type locality (neotype; see Wetzer et al. 2021): Canada, British Columbia, Stanley Park (49.294°N, 123.155°W). Body length to ~ 8.5 mm. See Dana (1853 original description as Spheromaoregonensis), Richardson (1899, 1900b, 1905a), Menzies (1954), George and Strömberg (1968), Miller (1968), Schultz (1969), Miller (1975), Higley and Holton (1978), Kussakin (1979), Ricketts et al. (1985), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Ranasinghe et al. (2007), Espinosa-Pérez et al. (2009), SCAMIT (2011–2021), Puget Sound Institute (2012–2023), Lafferty and Suchanek (2016), Gillett et al. (2017), and Wetzer et al. (2021). Note: See Endnote 15 regarding distribution of G. oregonense.
Paracerceiscordata (Richardson, 1899) [Fig. 21A] Intertidal to shallow subtidal, on coralline algae and kelp holdfasts; Aleutian Islands to southern California, USA. Type locality: Aleutian Islands, Popoff Island. Body length to ~ 7.1 mm. See Richardson (1899 original description as Cilicaeacordata; 1900b, 1905a), Hewatt (1946), Schultz (1969), Miller (1975), Kussakin (1979), Lee and Miller (1980), SCAMIT (1994–2021), Chess and Hobson (1997), O’Clair and O’Clair (1998), Fairey et al. (2002), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Espinosa-Pérez et al. (2009), and Roletto et al. (2014).
Paracerceisgilliana (Richardson, 1899) [Fig. 21B] Subtidal, in sandy mud, and in bays on pilings, among mussel beds, and in association with hydroids, algae and ectoprocts (shallow to 73 m); Mendocino County, northern California, USA to Catalina Island, southern California, and south to Todos Santos Bay, Baja California Norte, Mexico. Type locality: USA, California, Catalina Island. Body length to ~ 4.8 mm. See Richardson (1899 original description as Cilicaeacaudatagilliana; 1905a), Schultz (1969), Allen (1976), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Campos and Villarreal (2008), and Espinosa-Pérez et al. (2009).
Paracerceissculpta (Holmes, 1904) [Fig. 21C] Intertidal to shallow subtidal; Morro Bay, central California to San Diego, southern California, USA, and south to San Juan Alima, Michoacan, Mexico, including the entire Gulf of California, and also distributed throughout the world via oceanic shipping. Type locality: USA, California, from pieces of sponge dredged in shallow water at San Clemente Island. Body length to ~ 7.5 mm. See Holmes (1904 original description as Dynamenesculpta), Richardson (1905a), Menzies (1962a), Miller (1968), Schultz (1969), Brusca (1980), Shuster (1987, 1992), Shuster and Wade (1991a, 1991b), Rodríguez et al. (1992), SCAMIT (1994–2021), Shuster and Guthrie (1999), Espinosa-Pérez and Hendrickx (2001a, 2001b, 2006), Hewitt and Campbell (2001), Fairey et al. (2002), Smith et al. (2003), Ariyama and Otani (2004), Ranasinghe et al. (2003, 2005, 2007, 2012), Brusca et al. (2004, 2007), Healey and Hovel (2004), McLaughlin et al. (2005), Whitmore et al. (2005), Reed and Hovel (2006), Sirota and Hovel (2006), Montelli and Lewis (2008), Cruz-García et al. (2013), Munguia and Shuster (2013), Morales-Zarate et al. (2016), Gillett et al. (2017, 2022), Ramalhosa et al. (2017), Marchini et al. (2018), Rumbold et al. (2018), and Wetzer et al. (2018).
Paracerceis sp. A [Fig. 21D] Intertidal; La Jolla, southern California, USA to the central Gulf of California, Mexico. Body length = no information available. See Brusca (1980: species listed as Paracerceis sp.), SCAMIT (2008–2021), and Gillett et al. (2022). Note: Provisional species voucher sheet available at https://www.scamit.org (SCAMIT Newsletter, Vol. 15, No. 8, December 1996).
Paradelladianae (Menzies, 1962) [Fig. 22B] Intertidal to shallow subtidal; Ventura County, southern California, USA to San Juan de Alima, Michoacan, Mexico, including the Gulf of California, and widely introduced to ports around the world by international shipping. Type locality: Mexico, Baja California, Bahia de San Quintin. Body length to ~ 4 mm. See Menzies (1962a original description as Dynamenopsisdianae), Menzies and Glynn (1968), Schultz (1969), Iverson (1974), Allen (1976), Pires (1980), Kensley and Schotte (1989), Rodríguez et al. (1992), Espinosa-Pérez and Hendrickx (2001a, 2006), Bruce and Wetzer (2004), McLaughlin et al. (2005), Brusca et al. (2007), Wetzer and Bruce (2007), Campos and Villarreal (2008), and SCAMIT (2008–2021).
Pseudosphaeroma sp. [No figure] Intertidal, bays and harbors; San Francisco and central California Coast regions, and Salinas de San Pedro, Los Angeles County, southern California, USA. Body length to 5.2 mm. See Bruce and Wetzer (2008) and references therein (e.g., Hurley and Jansen 1977; Harrison 1984), and additional information available from R Wetzer (unpublished results). Note: Species probably translocated to the SCB from the southern hemisphere (e.g., likely New Zealand). The most similar species is P.campbellense, but the present species concept includes more than one species (see Bruce and Wetzer 2008).
Sphaeromaquoianum H. Milne Edwards, 1840 [Fig. 20D] Intertidal to shallow subtidal, boring in wood, mud, and soft rocks; Humboldt County, northern California, USA to San Quintin Bay, Baja California Norte, Mexico, and in Australia and New Zealand (i.e., probably introduced to western North America in the late 1800’s on ships from Australia). Type locality: Uncertain (i.e., not indicated in original description), but listed as Tasmania, Australia in Boyko et al. (2008 onwards). Body length to ~ 9.9 mm. See H. Milne Edwards (1840), Menzies (1962a, as Sphaeromapentodon), Rotramel (1972), Iverson (1974), Carlton (1979), Kussakin (1979), Lee and Miller (1980), Carlton and Iverson (1981), Ricketts et al. (1985), Talley et al. (2001), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Davidson and de Rivera (2010), Davidson et al. (2010), SCAMIT (2011–2021), and Brown et al. (2022).
Sphaeromawalkeri Stebbing, 1905 [Fig. 20E] Intertidal to shallow water, common in fouling communities in warm to warm-temperate ports and harbors, and boring into wood or soft rocks; introduced species with a worldwide distribution, but occurring in at least San Diego Bay, southern California, USA. Type locality: Sri Lanka. Body length to ~ 9.5 mm. See Stebbing (1905), Menzies and Glynn (1968), Carlton and Iverson (1981), Mak et al. (1985), Kensley and Schotte (1989), Cohen et al. (2005), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Galil (2008), Montelli and Lewis (2008), Khalaji-Pirbalouty and Wägele (2010), Ramalhosa et al. (2017), and Ortiz and Lalana (2018).
Family Tecticipitidae
Tecticepsconvexus Richardson, 1899 [Fig. 18B] Low intertidal to shallow subtidal, on sandy, rocky bottoms; southern Washington State (northwest of Columbia River mouth) to Catalina Island, southern California, USA. Type locality: USA, California, Monterey Bay. Body length to ~ 11.5 mm. See Richardson (1899, 1900b, 1905a, 1906), Iwasa (1934), Schultz (1969), Iverson (1974), Miller (1975), Kuris and Carlton (1977), Higley and Holton (1978), Kussakin (1979), Lee and Miller (1980), Hinton and Emmett (1994, 1996), Hunt et al. (1999), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Espinosa-Pérez et al. (2009), and Looby and Ginsburg (2021).
Suborder Valvifera
Family Arcturidae
Idarcturusallelomorphus Menzies & Barnard, 1959 [Fig. 25A] Continental shelf, soft-bottom substrates (12–94 m); Monterey, California, USA to Todos Santos Bay, Baja California Norte, Mexico. Type locality: USA, California, off Goleta (R/V Velero IV Station No. 4938-57; 34°27'25"N, 120°12'55"W). Body length to ~ 5.2 mm. See Menzies and Barnard (1959), Schultz (1969), Iverson (1974), Gillard (1978), Kussakin (1982), Dexter (1983), Lissner et al. (1986), Wetzer et al. (1991), SCAMIT (1994–2021), Wetzer and Brusca (1997), Bergen et al. (1998), Smith et al. (2001, 2003), Ranasinghe et al. (2003, 2007, 2012), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Campos and Villarreal (2008), Espinosa-Pérez et al. (2009), Stebbins (2012a, 2012c), Gillett et al. (2017, 2022), and Kim and Yoon (2021).
Idarcturushedgpethi Menzies, 1951 [Fig. 25B] Intertidal, on hydroids in laminarian zone at type locality; Strait of Georgia, British Columbia, Canada to Port Hueneme, southern California, USA. Type locality: USA, California, Marin County, Tomales Point, Tomales Bluff. Body length to ~ 4.2 mm. See Menzies (1951b), Schultz (1969), Miller (1975), Kussakin (1982), Wetzer and Brusca (1997), Fairey et al. (2002), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), SCAMIT (2008–2021), Macdonald et al. (2010), Stebbins (2012a, 2012c), Gillett et al. (2017), and Kim and Yoon (2021). Note: See Endnote 20.
Idarcturus sp. A [No figure] Continental shelf, soft bottom substrates (18–95 m); northern Channel Islands and Western Santa Barbara Channel, southern California, USA. Body length to ~ 5.5 mm. Note: See Endnote 20.
Neastacillacalifornica (Boone, 1918) [Fig. 25C] Continental shelf, on seaweed and soft bottom benthos (subtidal to 99 m); Point Conception, California, USA to Guerrero Negro Lagoon, Baja California Sur, Mexico, and in the Gulf of California. Type locality: USA, California, Venice. Body length to ~ 7.4 mm. See Boone (1918 original description as Astacillacalifornica), Menzies and Barnard (1959), Schultz (1969), Kussakin (1982), Thompson et al. (1993), SCAMIT (1994–2021), Hendrickx et al. (1997), Bergen et al. (1998), Espinosa-Pérez and Hendrickx (2001a, 2006), Smith et al. (2001, 2003), King (2003), Ranasinghe et al. (2003, 2007, 2012), McLaughlin et al. (2005), Melwani and Kim (2008), Stebbins (2012a, 2012c), Morales-Zarate et al. (2016), Gillett et al. (2017, 2022), and Mejía-Rebollo et al. (2018).
Family Holognathidae
Cleantioidesoccidentalis (Richardson, 1899) [Fig. 25E] Low intertidal to subtidal, typically on sand and muddy-sand substrates (0–50 m); southern California, USA to Baja California Sur, Mexico, throughout the Gulf of California, and south to Columbia and Ecuador, including the Galapagos Islands. Type locality: Mexico, Baja California Sur, Magdalena Bay. Body length to ~ 12.5 mm. See Richardson (1899 original description as Cleantisoccidentalis; 1905a), Schultz (1969), Kensley and Kaufman (1978), Brusca and Wallerstein (1979a, 1979b), Brusca and Iverson (1985), Poore and Lew Ton (1990), Espinosa-Pérez and Hendrickx (2001a, 2002, 2006), McLaughlin et al. (2005), Brusca et al. (2007), Lazarus-Agudelo and Cantera-Kintz (2007), SCAMIT (2011–2021), Ranasinghe et al. (2012), Liu and Poore (2013), and Negromonte et al. (2022).
Family Idoteidae
Colidoteafindleyi Brusca & Wallerstein, 1977 [Fig. 27C] Low intertidal to shallow subtidal, common on fronds of the brown algae Sargassum; San Diego, California, USA (rare) to Guadalupe Island, Baja California Norte, and the Gulf of California, Mexico. Type locality: Mexico, Sonora, 5 miles north of Cabo Tepoca (Puerto Lobos) (~ 30°17'N; 112°50'W). Body length to ~ 24.5 mm. See Brusca and Wallerstein (1977, 1979b, 1982), Brusca (1980, 1983b), Wetzer et al. (1991), Espinosa-Pérez and Hendrickx (2001a, 2002, 2006), Brusca et al. (2004, 2007), Campos and Villarreal (2008), and Stebbins (2012a, 2012c).
Colidotearostrata (Benedict, 1898) [Fig. 27D] Low intertidal to shallow subtidal, living as obligate commensal on sea urchins Strongylocentrotuspurpuratus and Mesocentrotusfranciscanus; Coos Bay, Oregon, USA to Baja California Norte, Mexico, but rare north of southern California. Type locality: USA, California, San Pedro. Body length to ~ 13.7 mm. See Benedict (1898 original description as Idotearostrata), Richardson (1899, 1900b, 1905a), Shearer (1942), Hewatt (1946), Johnson and Snook (1967), Schultz (1969), Allen (1976), Brusca and Wallerstein (1979b), Lee and Miller (1980); Ricketts et al. (1985), Stebbins (1988a, 1988b, 1989, 2012a, 2012c), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), and SCAMIT (2012–2021).
Colidoteawallersteini Brusca, 1983 [Fig. 27E] Intertidal; known only from two localities off the coast of Baja California Norte (i.e., Punta Clara and Guadalupe Island), Mexico. Type locality: Mexico, Baja California Norte, Punta Clara, south Rio Santo Tomás. Body length to ~ 16.5 mm. See Brusca (1983b), Wetzer et al. (1991), Espinosa-Pérez and Hendrickx (2002, 2006), McLaughlin et al. (2005), and Stebbins (2012a, 2012c).
Edotiasublittoralis Menzies & Barnard, 1959 [Fig. 27A] Continental shelf, typically occurring in sandy-silt benthic sediments (10–46 m, and one record > 100 m off San Diego); Vancouver Island, British Columbia to western Baja California Norte, Mexico, the Gulf of California (Bahia Concepcion), and one record off Costa Rica. Type locality: USA, California, off Newport (R/V Velero IV Station No. 4720-56; 33°37'39"N, 117°58'16"W). Body length to ~ 5.8 mm. See Menzies and Barnard (1959 original description as Edoteasublittoralis), Schultz (1969), Iverson (1974), Miller (1975), Brusca and Wallerstein (1979b), Kussakin (1982), Vargas-Zamora et al. (1985), Rafi and Laubitz (1990), Wetzer et al. (1991), Thompson et al. (1993), Hinton and Emmett (1994, 1996), SCAMIT (1994–2021), Espinosa-Pérez and Hendrickx (2001a, 2006), Ranasinghe et al. (2003, 2007, 2012), McLaughlin et al. (2005), Campos and Villarreal (2008), Melwani and Kim (2008), Stebbins (2012a, 2012c), Lafferty and Suchanek (2016), Gillett et al. (2017, 2022), and Henkel et al. (2020). Note: See Endnote 21.
Edotia sp. B [Fig. 27B] Continental shelf, typically occurring in sandy-silt benthic sediments (24–62 m); southern California, USA to Baja California Norte, Mexico. Body length to ~ 4.1 mm. See Menzies and Barnard (1959, as Edoteasublittoralis in part), SCAMIT (1994–2021), Ranasinghe et al. (2003, 2007, 2012), Stebbins (2012a, 2012c), and Gillett et al. (2017). Note: See Endnote 21.
Erichsonellacrenulata Menzies, 1950 [Fig. 26B] Intertidal to shallow subtidal, typically occurring on the eelgrass Zosteramarina: Newport Bay, southern California to San Quintin Bay, Baja California Norte, Mexico. Type locality: USA, California, Orange County, Newport Bay. Body length to ~ 23 mm. See Menzies (1950b, 1962a), Schultz (1969), Allen (1976), Pires (1984), Wetzer et al. (1991), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Jorgensen et al. (2007), Ranasinghe et al. (2007, 2012), Campos and Villarreal (2008), SCAMIT (2008–2021), Stebbins (2012a, 2012c), and Gillett et al. (2017, 2022).
Eusymmeruspseudoculata (Boone, 1923) [Fig. 26C] Intertidal to subtidal (0–18 m); Point Conception, California to US-Mexico border. Type locality: USA, California, San Pedro. Body length to ~ 9 mm. See Boone (1923 original description as Erichsonellapseudoculata), Menzies and Bowman (1956 redescription as Ronaleapseudoculata), Menzies and Barnard (1959), Schultz (1969), Kussakin (1982), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Espinosa-Pérez et al. (2009), SCAMIT (2012–2021), and Stebbins (2012a, 2012c).
Idoteafewkesi Richardson, 1905 [Fig. 29A] Intertidal to shallow water (0–6 m); Alaska, USA to Guerrero Negro, Baja California Sur, Mexico. Type locality: USA, California, Santa Barbara. Body length to ~ 42 mm. See Richardson (1905a), Fee (1927), Shearer (1942), Hatch (1947), Menzies (1950a), Schultz (1969), Miller (1975), Brusca and Wallerstein (1979b), Lee and Miller (1980), Kussakin (1982), Rafi and Laubitz (1990), O’Clair and O’Clair (1998), SCAMIT (2001–2021), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Campos and Villarreal (2008), Espinosa-Pérez et al. (2009), Puget Sound Institute (2012–2023), Stebbins (2012a, 2012c), Roletto et al. (2014), Morales-Zarate et al. (2016), and Shaughnessy et al. (2017).
Idoteametallica Bosc, 1801 [Fig. 29B] Cosmopolitan pelagic species found attached to drifting seaweed, occasionally occurring in southern California and Gulf of California during warm water years. Type locality: Unknown to us (not indicated in original description). Body length to ~ 29 mm. See Bosc (1801), Miers (1881), Richardson (1900b, 1905a, 1909), Naylor (1957), Schultz (1969), Kussakin (1982), Poore and Lew Ton (1993), Franke et al. (1998), Abelló et al. (2004), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), and Riera (2014). Note: Poore and Lew Ton (1993) provide a comprehensive synonymy and figures for I.metallica.
Idotearufescens Fee, 1927 [Fig. 29C] Low intertidal to subtidal (0–82 m), occurring mostly on algae; Prince Williams Sound, Alaska, USA to western Baja California, Mexico, including Catalina and the Coronado Islands. Type locality: Canada, British Columbia, Gabriola Passage. Body length to ~ 20.6 mm. See Fee (1927), Hatch (1947), Menzies (1950a), Schultz (1969), Iverson (1974), Miller (1975), Kussakin (1982), Rafi and Laubitz (1990), Wetzer and Brusca (1997), O’Clair and O’Clair (1998), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), SCAMIT (2008–2021), Macdonald et al. (2010), Stebbins (2012a, 2012c), and Shaughnessy et al. (2017).
Idoteaurotoma Stimpson, 1864 [Fig. 29D] Intertidal to shallow subtidal (0–14 m); Alaska, USA to Todos Santos Bay, Baja California Norte, Mexico, and the Gulf of California. Type locality: USA, Washington, Puget Sound. Body length to ~ 26.5 mm. See Stimpson (1864), Richardson (1899, 1900b, 1905a), Shearer (1942), Hewatt (1946), Hatch (1947), Menzies (1950a), George and Strömberg (1968), Schultz (1969), Miller (1975), Allen (1976), Brusca and Wallerstein (1977, 1979b, 1982), Brusca (1980), Lee and Miller (1980), Kussakin (1982), Dexter (1983), Ricketts et al. (1985), Rafi and Laubitz (1990), O’Clair and O’Clair (1998), Espinosa-Pérez and Hendrickx (2001a, 2006), SCAMIT (2001–2021), McLaughlin et al. (2005), Brusca et al. (2007), Campos and Villarreal (2008), Macdonald et al. (2010), Stebbins (2012a, 2012c), and Roletto et al. (2014).
Pentidoteaaculeata Stafford, 1913 [Fig. 30A] Intertidal; occurring in various habitats, including encrusting red algae (e.g., Melobesia) on the surfgrass Phyllospadix: British Columbia, Canada to Cedros Island, Baja California Norte, Mexico, and the Gulf of California. Type locality: USA, California, Laguna Beach, low tide. Body length to ~ 23 mm. See Stafford (1913b), Hewatt (1946), Menzies (1950a), George and Strömberg (1968), Schultz (1969), Miller (1975), Allen (1976), Brusca and Wallerstein (1977, 1979b, 1982), Brusca (1980), Lee and Miller (1980), Kussakin (1982), Espinosa-Pérez and Hendrickx (2001a, 2006), McLaughlin et al. (2005), Brusca et al. (2007), Campos and Villarreal (2008), SCAMIT (2011–2021), and Stebbins (2012a, 2012c).
Pentidoteakirchanskii (Miller & Lee, 1970) [Fig. 30B] Intertidal, commonly on surfgrass Phyllospadix; Oregon to southern California, USA. Type locality: USA, California, Monterey County, Pebble Beach, 17 Mile Drive at Seal Rock. Body length to ~ 21 mm. See Miller and Lee (1970 original description as Idothea (Pentidotea) kirchanskii), Miller (1975), Brusca and Wallerstein (1979b), Lee and Miller (1980), Kussakin (1982), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), and Stebbins (2012a, 2012c).
Pentidoteamontereyensis Maloney, 1933 [Fig. 30C] Intertidal to subtidal (0–109 m), common on surfgrass (Phyllospadix spp.) in shallow waters; Alaska, USA to western Baja California, Mexico. Type locality: USA, California, Monterey Bay. Body length to ~ 28.3 mm. See Maloney (1933), Menzies (1950a), Schultz (1969), Iverson (1974), Miller (1975), Brusca and Wallerstein (1979b), Lee and Miller (1980), Kussakin (1982), Ricketts et al. (1985), Rafi and Laubitz (1990), SCAMIT (1994–2021), O’Clair and O’Clair (1998), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), and Stebbins (2012a, 2012c).
Pentidotearesecata (Stimpson, 1857) [Fig. 30D] Intertidal to subtidal, frequently occurring in kelp and eelgrass beds; Alaska, USA to western Baja California Sur, Mexico, and to Mazatlán in the Gulf of California. Type locality: USA, Washington, Straits of Juan de Fuca, opposite Fort Townsend. Body length to ~ 57 mm. See Stimpson (1857 original description as Idotearesecata), Richardson (1899, 1900b, 1905a), Fee (1927), Hewatt (1946), Hatch (1947), Menzies and Waidzunas (1948), Menzies (1950a), George and Strömberg (1968), Schultz (1969), Iverson (1974), Miller (1975), Allen (1976), Brusca and Wallerstein (1977, 1979b, 1982), Brusca (1980), Lee and Miller (1980), Kussakin (1982), Ricketts et al. (1985), Rafi and Laubitz (1990), SCAMIT (1994–2021), Chess and Hobson (1997), O’Clair and O’Clair (1998), Espinosa-Pérez and Hendrickx (2001a, 2002, 2006), McLaughlin et al. (2005), Brusca et al. (2007), Campos and Villarreal (2008), Stebbins (2012a, 2012c), Roletto et al. (2014), and Shaughnessy et al. (2017).
Pentidoteaschmitti (Menzies, 1950) [Fig. 30E] Intertidal to subtidal (0–73 m); Bering Sea to Punta Banda, Baja California Norte, Mexico. Type locality: USA, California, Marin County, Dillon Beach, Second Sled Road. Body length to ~ 35 mm. See Menzies (1950a original description as Idothea (Pentidotea) schmitti), Schultz (1969), Iverson (1974), Miller (1975), Brusca and Wallerstein (1979b), Kussakin (1982), Ricketts et al. (1985), Rafi and Laubitz (1990), Wetzer et al. (1991), O’Clair and O’Clair (1998), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Stebbins (2012a, 2012c), Roletto et al. (2014), and Lafferty and Suchanek (2016).
Pentidoteastenops (Benedict, 1898) [Fig. 30F] Intertidal to shallow subtidal, often occurring on the feather-boa kelp Egregiamenziesii, surfgrass (Phyllospadix spp.), and algae-covered rocks; Alaska, USA to San Eugenio Point, Baja California Sur, Mexico, and the Gulf of California. Type locality: USA, California, Monterey. Body length to 60 mm. See Benedict (1898 original description as Idoteastenops), Richardson (1899, 1900b, 1905a), Shearer (1942), Hatch (1947), Menzies (1950a), George and Strömberg (1968), Schultz (1969), Iverson (1974), Miller (1975), Allen (1976), Brusca and Wallerstein (1977, 1979b, 1982), Brusca and Brusca (1978), Brusca (1980), Lee and Miller (1980), Kussakin (1982), Ricketts et al. (1985), O’Clair and O’Clair (1998), Espinosa-Pérez and Hendrickx (2001a, 2006), McLaughlin et al. (2005), Brusca et al. (2007), Campos and Villarreal (2008), Stebbins (2012a, 2012c), and Roletto et al. (2014).
Pentidoteawosnesenskii (Brandt, 1851) [Fig. 30G] Intertidal to subtidal (0–16 m); Aleutian Islands, Alaska, USA to southern California, plus a single record from off La Paz, Mexico in the Gulf of California. Type locality: Listed in Brandt (1851) as more than one site, including Sea of Okhotsk, Attu, Atcha, St. Paul, Kadjak, Sitka and the northern California coast. Body length to ~ 40.2 mm. See Brandt (1851 original description as Idoteawosnesenskii), Richardson (1899, 1900b, 1905a), Fee (1927), Shearer (1942), Hatch (1947), Menzies (1950a), George and Strömberg (1968), Schultz (1969), Miller (1975), Brusca and Wallerstein (1977, 1979b, 1982), Brusca (1980), Lee and Miller (1980), Kussakin (1982), Ricketts et al. (1985), Rafi and Laubitz (1990), O’Clair and O’Clair (1998), Espinosa-Pérez and Hendrickx (2001a, 2006), McLaughlin et al. (2005), Brusca et al. (2007), Toft et al. (2008, 2010), Stebbins (2012a, 2012c), Roletto et al. (2014), and Lafferty and Suchanek (2016).
Stenosomawetzerae (Ormsby, 1991) [Fig. 26A] Subtidal, occurring on brown algae Sargassumpalmeri and Cystoseiraneglecta (13 m); Catalina Island, California, USA, and Guaymas, Mexico in the Gulf of California (unpublished data). Type locality: USA, California, Catalina Island, Isthmus Reef (33°25.4'N; 118°30.8'W). Body length to ~ 10.6 mm. See Ormsby (1991 original description as Synisomawetzerae), Hedo and Junoy (1999), Espinosa-Pérez and Hendrickx (2001a, 2006), Castellanos and Junoy (2005), McLaughlin et al. (2005), Santos et al. (2011), Stebbins (2012a, 2012c), Xavier et al. (2012), and Artüz and Kubanç (2015).
Synidoteacalcarea Schultz, 1966 [Fig. 28A] Continental shelf, slope, and submarine canyons (54–1816 m); Tanner and Santa Rosa Canyons, southern California, USA. Type locality: USA, California, Tanner Canyon (R/V Velero IV Station No. 6833-60; 32°37'54"N, 118°58'40"W). Body length to ~ 6 mm. See Schultz (1966 original description, but with prior listing in Schultz 1964 a nomen nudum; 1969), Iverson (1972), Menzies and Miller (1972), Lissner et al. (1986), Wetzer et al. (1991), SCAMIT (1994–2021), Poore (1996), Wetzer and Brusca (1997), Moore (2004), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Ranasinghe et al. (2007), Stebbins (2012a, 2012c), and Gillett et al. (2019). Note: See Endnote 22.
Synidoteaharfordi Benedict, 1897 [Fig. 28B] Intertidal to shallow subtidal (0–12 m); Oregon, USA to western Baja California Sur, Mexico, the Gulf of California, one record from Costa Rica, and introduced to Japan. Type locality: Mexico, Baja California Sur, Magdalena Bay. Body length to ~ 17 mm. See Benedict (1897), Richardson (1899, 1905a), Schultz (1969), Menzies and Miller (1972), Miller (1975), Brusca and Wallerstein (1979b, 1982), Kussakin (1982), Vargas-Zamora at al. (1985), Poore (1996), Espinosa-Pérez and Hendrickx (2001a, 2006), SCAMIT (2001–2021), Moore (2004), McLaughlin et al. (2005), Brusca et al. (2007), Stebbins (2012a, 2012c), and Hendrickx (2018b).
Synidoteamagnifica Menzies & Barnard, 1959 [Fig. 28C] Continental shelf (29–98 m); San Luis Obispo, California, USA to western Baja California Norte, Mexico. Type locality: USA, California, off Oceanside (R/V Velero IV Station No. 5108-57; 33°10'30"N, 117°25'25"W). Body length to ~ 6 mm. See Menzies and Barnard (1959), Schultz (1969), Iverson (1972, 1974), Menzies and Miller (1972), Kussakin (1982), Wetzer et al. (1991), SCAMIT (1994–2021), Poore (1996), Ranasinghe et al. (2003, 2007, 2012), Moore (2004), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Campos and Villarreal (2008), Stebbins (2012a, 2012c), and Gillett et al. (2017, 2022). Note: See Endnote 22.
Synidoteamedia Iverson, 1972 [Fig. 28D] Continental shelf (depths to at least 183 m): Strait of Georgia, British Columbia, Canada to Santa Maria Basin, California, USA. Type locality: USA, California, Monterey County, southwest of Point Soberanes (between 36°25'7"N–36°26'23"N). Body length to ~ 8.7 mm. See Iverson (1972), Kussakin (1982), Lissner et al. (1986), SCAMIT (1994–2021), Poore (1996), Wetzer and Brusca (1997), Ranasinghe et al. (2003, 2007), Moore (2004), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Macdonald et al. (2010), Stebbins (2012a, 2012c), and Henkel et al. (2020). Note: See Endnote 22.
Family Thermoarcturidae
Califarcturustannerensis (Schultz, 1966) [Fig. 25D] Continental basin and canyon benthos, green mud (1197–1335 m); Tanner Basin and Tanner Canyon, southern California, USA. Type locality: USA, California, Tanner Canyon (R/V Velero IV Station No. 6832-60; 32°33'36"N, 118°55'40"W). Body length to ~ 5.5 mm. See Schultz (1966 original description as Microarcturustannerensis; 1969), Wetzer et al. (1991), Stebbins (2012a, 2012c), and Poore (2015: supplementary redescription). Note: The only previously known record for this species was the holotype collected in 1960 from the Tanner Canyon at depths between 1298–1320 m (see Schultz 1966 and Poore 2015). Depth range is herein slightly expanded based on additional specimens collected in 1971 and 1976, which were recently rediscovered in the Crustacea Collection of the Natural History Museum of Los Angeles County (Specimen Nos. 28303 and 28313; TDS, pers. obs.).
Suborder Asellota
Superfamily Janiroidea
Family Dendrotionidae
Acanthomunnatannerensis Schultz, 1966 [Fig. 31A] Continental slope and submarine canyon benthos (570–813 m); southeast of Catalina Island and Tanner Canyon, southern California, USA. Type locality: USA, California, Tanner Canyon (R/V Velero IV Station No. 6833-60; 32°37'54"N, 118°58'40"W). Body length to ~ 3.5 mm. See Schultz (1966, 1969), Wetzer et al. (1991), Cohen (1998), McLaughlin et al. (2005), SCAMIT (2011–2021), Ranasinghe et al. (2012), and Golovan et al. (2018).
Family Desmosomatidae
Desmosoma sp. A [Fig. 40A] Continental slope, soft bottom benthos (290 m); Santa Maria Basin off Point Buchon and Point Estero, San Luis Obispo County, California, USA (~ 89–110 km north of SCB northern boundary). Body length to ~ 2.2 mm. See Wilson (1997: 106–107, fig. 1.44, description and figures).
Momedossasymmetrica (Schultz, 1966) [Fig. 40B] Continental slope and submarine canyons (469–2955 m); Gulf of the Farallones, San Francisco, northern California to Tanner Canyon, southern California, USA. Type locality: USA, California, Tanner Canyon (R/V Velero IV Station No. 6836-60; 32°36'00"N, 119°05'18"W). Body length to ~ 3.2 mm. See Schultz (1966 original description as Desmosomasymmetrica), Lissner et al. (1986), Wetzer et al. (1991), Wilson (1997), Blake et al. (2009), and SCAMIT (2011–2021).
Prochelator sp. A [Fig. 40C] Continental shelf, slope, and basins (99–2955 m); Gulf of the Farallones, San Francisco, northern California to Santa Maria Basin off Purisima Point, Santa Barbara County, California, USA (~ 35 km north of SCB northern boundary). Body length to ~ 2.5 mm. See Wilson (1997: 108–109, fig. 1.45, description and figures), Blake et al. (2009), and SCAMIT (2001–2021). Note: Also see Lissner et al. (1986) for previous report of likely same species from the region.
Family Haplomunnidae
Haplomunnacaeca (Richardson, 1905) [Fig. 37] Abyssal seafloor (3998 m); ~ 210 km southwest of Santa Catalina Island, southern California, USA. Type locality: USA, southern California (Albatross Station No. 4390; 33°02'15"N, 120°42'W). Body length to ~ 7.7 mm (estimated). See Richardson (1905a original description as Munnacaeca; 1908), Menzies (1962b), Wolff (1962), Wilson (1976), Poore (1984c), Cunha and Wilson (2003).
Family Janiridae
Caecianiropsis sp. LA1 [Fig. 39B, image of representative species C.psammophila] Continental shelf benthos (95 m): off west end of San Miguel Island, southern California, USA. No body size information available, but a similar species from northern California (C.psammophila Menzies & Pettit, 1956) measured 1.8 mm in length. See Menzies and Pettit (1956: description of C.psammophila), SCAMIT (2001–2021), and Gillett et al. (2017). Note: See Endnote 28.
Caecianiropsis sp. LA2 [Fig. 39B, image of representative species C.psammophila] Continental shelf benthos, coarse sediments (27 m): offshore Palos Verdes, southern California, USA. No body size information available other than similar in size to Caecianiropsis sp. LA1 (see preceding species). See Menzies and Pettit (1956: description of C.psammophila), SCAMIT (2001–2021), Ranasinghe et al. (2012), and Gillett et al. (2017, 2022). Note: See Endnote 28.
Caecijaerahorvathi Menzies, 1951 [Fig. 39A] Intertidal, in burrows excavated in wood by the isopods Limnoria spp.; southern California, USA, and Hawaii. Type locality: USA, California, Los Angeles-Long Beach Harbor, Terminal Island. Body length to ~ 1.7 mm. See Menzies (1951c), Schultz (1969), Cooke (1977), Wetzer et al. (1991), Wilson and Wägele (1994), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), and Ruiz et al. (2011).
Iaiscalifornica (Richardson, 1904) [Fig. 31B] Shallow water bays and estuaries, commensal with the wood-boring isopod Sphaeromaquoianum; Humboldt Bay, northern California to San Diego, southern California, USA, introduced from Australia or New Zealand, also in Singapore. Type locality: USA, California, Sausalito. Body length to ~ 2.9 mm. See Richardson (1904a original description as Janiropsiscalifornica), Menzies and Barnard (1951), Schultz (1969), Rotramel (1972, 1975), Iverson (1974), Miller (1975), Kussakin (1988), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Espinosa-Pérez et al. (2009), and Ruiz et al. (2011).
Ianiropsisanaloga Menzies, 1952 [Fig. 34A] Intertidal, under rocks or amongst Laminaria holdfasts; San Juan Archipelago, Washington to Los Angeles Harbor, southern California, USA. Type locality: USA, California, Marin County, Tomales Point (bay side). Body length to ~ 3.7 mm. See Menzies (1952), George and Strömberg (1968), Schultz (1969), Miller (1975), Kussakin (1988), Wetzer et al. (1991), Fairey et al. (2002), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), SCAMIT (2008–2021), Doti and Wilson (2010), Lafferty and Suchanek (2016), and Gillett et al. (2017, 2022).
Ianiropsisderjugini Gurjanova, 1933 [Fig. 34B] Intertidal, under rocks covered by algae; Bering Sea to southern California, USA, and Korea (Korea Strait and East Sea). Type locality: Bering Sea. Body length to ~ 4.8 mm. See Gurjanova (1933 original description as Janiropsisderjugini), Miller (1975), Kussakin (1988), Jang and Kwon (1990), Wilson (1994), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Doti and Wilson (2010), and SCAMIT (2011–2021).
Ianiropsiskincaidi Richardson, 1904 [Fig. 34C] Intertidal; Bering Sea to Catalina Island and Los Angeles Harbor, southern California, USA. Type locality: USA, Alaska, Yakutat Bay. Body length to ~ 3.8 mm. See Richardson (1904b), George and Strömberg (1968), Schultz (1969), Miller (1975), Kussakin (1988), Jang and Kwon (1990), Wilson (1994), Fairey et al. (2002), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Espinosa-Pérez et al. (2009), Doti and Wilson (2010), Macdonald et al. (2010), Roletto et al. (2014), Lafferty and Suchanek (2016), and Looby and Ginsburg (2021).
Ianiropsisminuta Menzies, 1952 [Fig. 34D] Intertidal, under rocks or sand; Marin County, northern California to southern California, USA. Type locality: USA, California, Marin County, Dillon Beach, Second Sled Road. Body length to ~ 1.3 mm. See Menzies (1952), Schultz (1969), Miller (1975), Lissner et al. (1986), Kussakin (1988), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Doti and Wilson (2010), and SCAMIT (2011–2021).
Ianiropsismontereyensis Menzies, 1952 [Fig. 34E] Intertidal to shallow subtidal, under rocks or in Macrocystis holdfasts; Marin County, northern California to southern California, USA. Type locality: USA, California, Monterey County, Pescadero Point. Body length to ~ 3.6 mm. See Menzies (1952), Schultz (1969), Miller (1975), Kussakin (1988), Wetzer et al. (1991), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Espinosa-Pérez et al. (2009), Doti and Wilson (2010), and SCAMIT (2011–2021).
Ianiropsistridens Menzies, 1952 [Fig. 34F] Intertidal, on algae or occasionally in sponges; Strait of Georgia, British Columbia, Canada to Oceanside, southern California, USA, Peru, northern Chile, and Korea (Korea Strait and East Sea). Type locality: USA, California, Marin County, Tomales Point (bay side). Body length to ~ 3 mm. See Menzies (1952, 1962b), George and Strömberg (1968), Schultz (1969), Miller (1975), Kussakin (1988), Jang and Kwon (1990), Wetzer et al. (1991), Fairey et al. (2002), Cohen et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), SCAMIT (2008–2021), Doti and Wilson (2010), Macdonald et al. (2010), Gillett et al. (2017), and Zúñiga Delgado et al. (2021).
Janiralatadavisi Menzies, 1951 [Fig. 33A] Low intertidal, under rocks; Monterey County, California, USA to Todos Santos Bay, Baja California Norte, Mexico. Type locality: USA, California, Monterey County, Carmel Cove. Body length to ~ 4.7 mm. See Menzies (1951b), Miller (1975), Kussakin (1988), Wetzer et al. (1991), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), and Campos and Villarreal (2008).
Janiralataoccidentalis (Walker, 1898) [Fig. 33B] Intertidal, under rocks; Strait of Georgia, British Columbia, Canada to Orange County, southern California, USA. Type locality: USA, Washington, Puget Sound. Body length to ~ 6 mm. See Walker (1898 original description as Janiraoccidentalis), Richardson (1899, 1905a), Stafford (1913b), Hatch (1947), Menzies and Barnard (1959), George and Strömberg (1968), Schultz (1969), Miller (1975), Kussakin (1988), SCAMIT (1994–2021), Bergen et al. (1998), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Ranasinghe et al. (2007), and Macdonald et al. (2010).
Janiralatarajata Menzies, 1951 [Fig. 33C] Subtidal (1–36 m); Monterey Bay, California to Anacapa and Catalina Islands, southern California, USA. Type locality: USA, California, Monterey Bay (from eggcase of the skate Rajabinoculata). Body length to ~ 4 mm. See Menzies (1951b), Miller (1968), Schultz (1969), Miller (1975), Kussakin (1988), and Espinosa-Pérez and Hendrickx (2006). Note: See Endnote 24.
Janiralatasolasteri (Hatch, 1947) [Fig. 33D] Subtidal (50–218 m): Dall Island, Gulf of Alaska to southern California, USA. Type locality: USA, Washington, Hood Canal, on the sea star Solasterstimpsoni. Body length to ~ 5 mm. See Hatch (1947 original description as Janirasolasteri), Menzies and Barnard (1959), Schultz (1966, 1969), George and Strömberg (1968), Miller (1975), Lissner et al. (1986), Kussakin (1988), SCAMIT (1994–2021), Bergen et al. (1998), Espinosa-Pérez and Hendrickx (2006), Ranasinghe et al. (2007), Espinosa-Pérez et al. (2009), Macdonald et al. (2010), and Puget Sound Institute (2012–2023).
Janiralata sp. A [Fig. 33E] Continental shelf (160–168 m); Santa Maria Basin, southern California, USA. Body length ~ 4.4 mm. See Wilson (1997: 80–82, fig. 1.31, description and figures), SCAMIT (2011–2021), and Ranasinghe et al. (2012).
Janiralata sp. B [Fig. 33F] Continental shelf and upper slope (168–237 m); Santa Maria Basin, southern California, USA. Body length ~ 4.8 mm. See Wilson (1997: 82–84, fig. 1.32, description and figures), Ranasinghe et al. (2007), and SCAMIT (2008–2021).
Janiralata sp. C [Fig. 33G] Continental shelf, hard bottom benthos (91–123 m); Santa Maria Basin offshore of Purisima Point, Santa Barbara County, California, USA (~ 35 km north of SCB northern boundary). Body length ~ 2.4 mm. See Wilson (1997: 84–85, fig. 1.33, description and figures).
Janiralata sp. D [Fig. 33H] Continental shelf, hard bottom benthos (91–123 m); Santa Maria Basin offshore of Purisima Point, Santa Barbara County, California, USA (~ 35 km north of SCB northern boundary). Body length ~ 3.5 mm. See Wilson (1997: 86–87, fig. 1.34, description and figures) and SCAMIT (2008–2021). Note: See Endnote 24.
Family Joeropsididae
Joeropsisconcava Schultz, 1966 [Fig. 32A] Continental shelf and submarine canyons, coarse sediments (60–221 m); Central California to San Diego, southern California, USA. Type locality: USA, California, Santa Cruz Canyon (R/V Velero IV Station No. 6806-59; 33°56'06"N, 118°52'17"W). Body length to ~ 4.2 mm. See Schultz (1966, 1969), Wetzer et al. (1991), Wilson (1997), SCAMIT (1994–2021), Ranasinghe et al. (2003, 2007, 2012), Espinosa-Pérez and Hendrickx (2006), Espinosa-Pérez et al. (2009), and Gillett et al. (2017, 2022).
Joeropsisdubiadubia Menzies, 1951 [Fig. 32B] Low intertidal to offshore shelf and submarine canyons, often occurring under rocks, in algal holdfasts, and on bryozoans, tunicates, hydroids and barnacles (0–116 m); Strait of Georgia, British Columbia, Canada to San Quintin Bay, Baja California Norte, the Gulf of California, and western Mexico (i.e., Bahia de Banderas, Nayarit). Type locality: USA, California, Marin County, Dillon Beach, First Sled Road north of Pacific Marine Station. Body length to ~ 2.8 mm. See Menzies (1951b original description as Jaeropsisdubia; 1962a), Menzies and Barnard (1959), Schultz (1966, 1969), Miller (1968, 1975), Carvacho (1983), Lissner et al. (1986), Wetzer et al. (1991), Thompson et al. (1993), SCAMIT (1994–2021 as J.dubia), Kussakin (1999), Espinosa-Pérez and Hendrickx (2001a, 2006), Smith et al. (2001, 2003), Ranasinghe et al. (2003, 2007, 2012), Brusca et al. (2007), Campos and Villarreal (2008), Macdonald et al. (2010), Puget Sound Institute (2012–2023), Cruz-García et al. (2013), Lafferty and Suchanek (2016), and Gillett et al. (2017, 2019, 2022). Note: If confirmed as J.dubiadubia, the Gillett et al. (2019) record extends the maximum depth range of this species from 116 m to the lower slope between 500–1000 m.
Joeropsisdubiapaucispinis Menzies, 1951 [Fig. 32C] Intertidal to offshore shelf and submarine canyons (0–116 m); Marin County, northern California to Santa Monica Canyon, southern California, USA. Type locality: USA, California, Marin County, Dillon Beach. Body length < 3 mm. See Menzies (1951b original description as Jaeropsisdubiapaucispinis; 1962a), Menzies and Barnard (1959), Schultz (1966, 1969), Miller (1968, 1975), Kussakin (1999), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), and Espinosa-Pérez et al. (2009).
Joeropsislobata Richardson, 1899 [Fig. 32D] Intertidal to shallow water; Coos Bay, Oregon to southern California, USA, plus one recent record from Namhyeongjeseom Islet, Busan, Republic of Korea (see Hwang et al. 2014) that should be further studied to verify. Type locality: USA, California, Monterey Bay. Body length to ~ 3.5 mm. See Richardson (1899, 1900c, 1905a), Shearer (1942), Hatch (1947), Schultz (1969), Miller (1975), Bergen et al. (1998), Kussakin (1999), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), SCAMIT (2008–2021), and Espinosa-Pérez et al. (2009).
Joeropsis sp. A [Fig. 32E] Continental shelf, hard bottom benthos (54–131 m); Santa Maria Basin offshore of Point Arguello and Purisima Point, Santa Barbara County, California, USA (~ 30–35 km north of SCB northern boundary). Body length ~ 2.8 mm. See Wilson (1997: 90–91, fig. 1.36, description and figures).
Family Lepidocharontidae
Microcharon sp. A [Fig. 39C, image of representative species M.sabulum] Continental shelf benthos (75 m); Channel Islands, southern California, USA. No body size information available, but a similar species from the Caribbean listed in Kensley and Schotte (1989), M.sabulum, measured ~ 1.5 mm in length. See Kensley and Schotte (1989), Ranasinghe et al. (2012), and SCAMIT (2012–2021). Note: See Endnote 27.
Family Munnidae
Munnachromatocephala Menzies, 1952 [Fig. 36A] Intertidal, on red algae and encrusting organisms on rocks; Strait of Georgia, British Columbia, Canada to central and southern California, USA. Type locality: USA, California, Marin County, Dillon Beach, First Sled Road. Body length to ~ 2.2 mm. See Menzies (1952), George and Strömberg (1968), Schultz (1969), Miller (1975), Poore (1984c), Kussakin (1988), Wetzer et al. (1991), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), SCAMIT (2008–2021), Espinosa-Pérez et al. (2009), Macdonald et al. (2010), and Lafferty and Suchanek (2016).
Munnafernaldi George & Strömberg, 1968 [Fig. 36B] Intertidal, from stones and algae; San Juan Archipelago, Washington to southern California, USA. Type locality: USA, Washington, San Juan Archipelago, San Juan Island, Friday Harbor. Body length to ~ 1.5 mm. See George and Strömberg (1968), Schultz (1969), Poore (1984c), Kussakin (1988), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Puget Sound Institute (2012–2023), and SCAMIT (2012–2021).
Munnahalei Menzies, 1952 [Fig. 36C] Intertidal to shallow subtidal, under rocks, in Macrocystis holdfasts, and among spines of the purple sea urchin Strongylocentrotuspurpuratus; Tomales Bay, northern California to southern California, USA. Type locality: USA, California, Marin County, Tomales Point, Tomales Bluff. Body length to ~ 1.5 mm. See Menzies (1952), Schultz (1969), Iverson (1974), Miller (1975), Harty (1979), Poore (1984c), Kussakin (1988), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), and SCAMIT (2011–2021).
Munnamagnifica Schultz, 1964 [Fig. 36D] Offshore continental slope, black mud, nodules, and flat shaley rocks (500 m); southern California, USA. Known only from the type material. Type locality: USA, California, 13.1 miles WNW of Santa Barbara Island (R/V Velero IV Station No. 2969-54; 33°35'59"N, 119°15'11"W). Body length to ~ 2 mm. See Schultz (1964, 1969), Wilson (1980), Poore (1984c), Wetzer et al. (1991), and McLaughlin et al. (2005).
Munnaspinifrons Menzies & Barnard, 1959 [Fig. 36E] Offshore continental shelf; 12–218 m: Point Conception to Point Loma, San Diego, southern California, USA. Type locality: USA, California, 11 miles east of Point Conception (R/V Velero IV Station No. 4822-57; 34°27'15"N, 120°14'45"W). Body length to ~ 1.5 mm. See Menzies and Barnard (1959), Schultz (1966, 1969), Poore (1984c), Kussakin (1988), Wetzer et al. (1991), SCAMIT (1998–2021), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), and Espinosa-Pérez et al. (2009).
Munnastephenseni Gurjanova, 1933 [Fig. 36F] Intertidal to shallow subtidal (0–18 m); Bering Sea to central and southern California, USA, and Japan. Type locality: Bering Sea, Komador Islands. Body length to ~ 3 mm. See Gurjanova (1933), Schultz (1969), Miller (1975), Poore (1984c), Kussakin (1988), SCAMIT (1994–2021), Shimomura and Mawatari (2001), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), and Brusca et al. (2007). Note: See Shimomura and Mawatari (2001) for a brief discussion of differences between California and Japanese specimens of M.stephenseni.
Munna sp. A [Fig. 36G] Continental shelf and upper slope, hard substrates (105–237 m); Santa Maria Basin, southern California, USA. Body length to ~ 1.5 mm. See Wilson (1997: 71–72, fig. 1.27, description and figures), Espinosa-Pérez and Hendrickx (2006), Ranasinghe et al. (2007), and SCAMIT (2008–2021). Note: See Lissner et al. (1986) for previous report of likely same species from the region.
Uromunnaubiquita (Menzies, 1952) [Fig. 36H] Intertidal to shallow subtidal; Strait of Georgia, British Columbia, Canada to San Quintin Bay, Baja California Norte, and the Gulf of California, Mexico. Type locality: USA, California, Marin County, Tomales Bay. Body length to ~ 1.2 mm. See Menzies (1952 original description as Munnaubiquita; 1962a), Menzies and Barnard (1959), George and Strömberg (1968), Schultz (1969), Miller (1975), Carvacho (1983), Poore (1984c), Kussakin (1988), Wetzer et al. (1991), SCAMIT (1994–2021), Chess and Hobson (1997), Espinosa-Pérez and Hendrickx (2001a, 2006), Ranasinghe et al. (2003, 2012), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Macdonald et al. (2010), Puget Sound Institute (2012–2023), Esquete and Wilson (2016), and Gillett et al. (2017, 2022).
Family Munnopsidae
Belonectes sp. A [Fig. 38A] Continental slope and submarine canyons (305–401 m); Santa Maria Basin to La Jolla Canyon, southern California, USA. Body length to ~ 2.1 mm. See SCAMIT (1994–2021), Wilson (1997: 94–95, fig. 1.38, description and figures), and Gillett et al. (2017, 2019). Note: If confirmed, the Gillett et al. (2019) record extends the maximum depth range of this species slightly from 401 m to the lower slope between 500 and 1000 m.
Eurycopecaliforniensis Schultz, 1966 [Fig. 38B] Continental slope (478–930 m); Santa Maria Basin to Newport Canyon, southern California, USA. Type locality: USA, California, Newport Canyon (R/V Velero IV Station No. 7032-60; 33°31'28"N, 117°54'58"W). Body length to ~ 3.5 mm. See Schultz (1966, 1969), Gillard (1978), Lissner et al. (1986), Wetzer et al. (1991), Wilson (1997), McLaughlin et al. (2005), Ranasinghe et al. (2007, 2012), SCAMIT (2008–2021), and Gillett et al. (2017, 2019, 2022).
Ilyarachnaacarina Menzies & Barnard, 1959 [Fig. 38C] Continental shelf, slope, and submarine canyons (73–1118 m, and possibly > 2100 m per ‘Note’ below); San Francisco, northern California to San Diego, southern California, USA (including Santa Maria Basin, and Coronado, Redondo, San Pedro, Santa Cruz, and Santa Monica canyons). Type locality: USA, California, off Santa Barbara (R/V Velero IV Station No. 4980-57; 34°15'50"N, 119°34'28"W). Body length to ~ 4.3 mm. See Menzies and Barnard (1959), Schultz (1964, 1966, 1969), Hessler and Thistle (1975), Gillard (1978), Thistle (1979), Lissner et al. (1986), Wetzer et al. (1991), SCAMIT (1994–2021), Wilson 1997), Bergen et al. (1998), Smith et al. (2001, 2003), Kussakin (2003), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Ranasinghe et al. (2007, 2012), Blake et al. (2009), Espinosa-Pérez et al. (2009), and Gillett et al. (2017, 2019, 2022). Note: San Francisco specimens identified as Ilyarachnacf.acarina were collected from a subset of lower slope stations ranging between 2160–3140 m in depth (see Blake et al. 2009).
Ilyarachnaprofunda Schultz, 1966 [Fig. 38D] Continental slope, basins, and submarine canyons (461–1670 m, and possibly > 2100 m per ‘Note’ below), extremely abundant on whale falls (skeletons); San Francisco, northern California to San Diego, southern California, USA (including Santa Cruz Basin, La Jolla, San Pedro, Santa Catalina and Tanner canyons). Type locality: USA, California, La Jolla Canyon (R/V Velero IV Station No. 7047-60; 32°54'21"N, 117°29'53"W). Body length to ~ 3.8 mm. See Schultz (1966, 1969), Hessler and Thistle (1975), Thistle (1979), Wetzer et al. (1991), Wilson (1997), Smith and Baco (2003), McLaughlin et al. (2005), Ranasinghe et al. (2007, 2012), SCAMIT (2008–2021), Blake et al. (2009), Bernardino et al. (2010), and Gillett et al. (2019, 2022). Note: San Francisco specimens identified as Ilyarachnacf.profunda were collected from a subset of lower slope stations ranging between 2160–3140 m in depth (see Blake et al. 2009).
Munnopsurus sp. A [Fig. 38E] Continental slope and submarine canyons (393–582 m); Santa Maria Basin to La Jolla Canyon, southern California, USA. Body length to ~ 1.8 mm. See Wilson (1997: 98–99, fig. 1.40, description and figures), SCAMIT (1998–2021), Ranasinghe et al. (2012), and Gillett et al. (2019). Note: See Lissner et al. (1986) for previous report of likely same species from the region.
Munnopsurus sp. B [Fig. 38F] Continental slope, soft bottom benthos (930 m); Santa Maria Basin west of Point Conception, Santa Barbara County, California, USA. Body length to ~ 7 mm. See Wilson (1997: 100–101, fig. 1.41, description and figures).
Family Nannoniscidae
Nannonisconuslatipleonus Schultz, 1966 [Fig. 40D] Continental slope and submarine canyons (294–465 m); Western Santa Barbara Channel off Point Conception to Redondo Canyon, southern California, USA. Type locality: USA, California, Redondo Canyon (R/V Velero IV Station No. 2793-54; 33°48'00"N, 118°32'00"W). Body length to ~ 2.8 mm. See Schultz (1966, 1969), Siebenaller and Hessler (1981), Lissner et al. (1986), Wetzer et al. (1991), Wilson (1997, 2008b), and McLaughlin et al. (2005). Note: Wilson (1997) briefly discusses that N.latipleonus may be conspecific with the more northern N.carinatus Mezhov, 1986. If true, this will extend the geographic range of N.latipleonus to Alaska Bay in the northeast Pacific.
Family Paramunnidae
Austrosignumlatum Just & Wilson, 2021 [Fig. 35A] Subtidal benthos (20 m); La Jolla Canyon, southern California, USA. Type locality: USA, California, La Jolla (32°52.09'N, 117°15.69'W). Body length to ~ 2 mm. See Just and Wilson (2021) for original description and comparison with other species of the genus.
Boreosignum sp. A [Fig. 35B] Shallow subtidal rocks (9–16 m); Point La Jolla to Pin Rock, Catalina Island, southern California, USA. Body length < 1 mm. See Cadien (2008: voucher sheet for Boreosignum sp. IS1 available from SCAMIT Toolbox at https://www.scamit.org; specimens examined 5 April 2008, D. B. Cadien), Maloney et al. (2008), and SCAMIT (2012–2021).
Munnogoniumerratum (Schultz, 1964) [Fig. 35C] Continental shelf (98–158 m); Morro Bay, central California to Santa Maria Basin, southern California, USA. Type locality: USA, California, Santa Barbara County, 4.6 miles 137 degrees from Gaviota Pier (R/V Velero IV Station No. 6003-58; 34°24'45"N, 120°08'40"W). Body length to ~ 1.8 mm. See Schultz (1964 original description as Austrosignumerratum; 1969), Bowman and Schultz (1974), Lissner et al. (1986), Wetzer et al. (1991), SCAMIT (1994–1996), Wilson (1997, as Munnogoniumcf.tillerae), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Just and Wilson (2021: comparison to redescribed M.tillerae), and Golovan and Malyutina (2022).
Munnogoniumtillerae (Menzies & Barnard, 1959) [Fig. 35D] Continental shelf (101–127 m); San Diego, southern California, USA. Type locality: USA, California, off Point Loma (R/V Velero IV Station No. 4753-56; 32°41'50"N, 117°20'25"W). Body length to ~ 1.3 mm. See Menzies and Barnard (1959 original description as Austrosignumtillerae), Schultz (1969), Bowman and Schultz (1974), Lissner et al. (1986), Kussakin (1988), Wetzer et al. (1991), SCAMIT (1994–2021), Wilson (1997: but not M.cf.tillerae [see M.erratum above]), Bergen et al. (1998), Smith et al. (2001, 2003), Ranasinghe et al. (2003, 2007, 2012), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Gillett et al. (2017, 2019, 2022), Just and Wilson (2021: redescription, range restriction, and comparison to M.erratum), and Golovan and Malyutina (2022).
Paramunnaquadratifrons Iverson & Wilson, 1981 [Fig. 41A] Continental shelf (197 m); southern Channel Islands and Tanner bank, southern California, USA. Type locality: USA, California, Tanner Bank, ~ 160 kilometers southwest of Los Angeles (33°53.24'N; 119°23.35'W). Body length to ~ 1.3 mm. See Iverson and Wilson (1981), Wetzer et al. (1991), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), SCAMIT (2011–2021), Ranasinghe et al. (2012), and Golovan and Malyutina (2022).
Paramunna sp. A [Fig. 41B] Continental shelf (100 m); off Del Mar, San Diego County, southern California, USA. Body length to ~ 1 mm. See Cadien (1996: voucher sheet available [in part] in SCAMIT Newsletter Vol. 15, No. 2 at https://www.scamit.org; specimens examined 30 May 1996, D. Cadien) and SCAMIT (2013–2021).
Paramunna sp. SD1 [No figure] Channel Island shelf (75 m); NE Anacapa Island, southern California, USA. No body size information available. See Ranasinghe et al. (2007) and SCAMIT (2008–2021). Note: Provisional species designation based on diagnostic notes for “Asellota sp. SD1” drafted by D. Pasko in 2004 (see Endnote 29).
Pleurogoniumcaliforniense Menzies, 1951 [Fig. 41C] Continental shelf, slope and submarine canyons, benthic sediments (93–150 m, and possibly > 2100 m as per ‘Note’ below); Strait of Georgia, British Columbia, Canada to Point Loma, San Diego, southern California, USA. Type locality: USA, California, Sonoma County, 3 miles west of mouth of Russian River. Body length to ~ 1.3 mm. See Menzies (1951b), Menzies and Barnard (1959), Schultz (1966, 1969), Lissner et al. (1986), Kussakin (1988), SCAMIT (1994–2021), Bergen et al. (1998), Smith et al. (2001, 2003), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Ranasinghe et al. (2007, 2012), Blake et al. (2009), Macdonald et al. (2010), Gillett et al. (2017, 2019, 2022), and Golovan and Malyutina (2022). Note: If confirmed, the Gillett et al. (2019) record extends the maximum depth range of this species slightly from 150 m to the upper slope between 200–500 m. Additionally, San Francisco specimens identified as Pleurogoniumcf.californiense were collected from a subset of lower slope stations ranging between 2160–3140 m in depth as reported by Blake et al. (2009), thus extending this bathymetric range into even deeper waters.
Pleurogonium sp. A [Fig. 41D] Continental shelf benthos (90–154 m); Santa Maria Basin to San Diego, southern California, USA. Body length to ~ 1.6 mm. See Wilson (1997: 78–79, fig. 1.30, description and figures), Bergen et al. (1998), SCAMIT (1994–2021), Ranasinghe et al. (2007, 2012), and Gillett et al. (2019, 2022). Note: If confirmed, the Gillett et al. (2019) record extends the maximum depth range of this species slightly from 154 m to the upper slope between 200–500 m.
Family Pleurocopidae
Pleurocope sp. A [Fig. 35E] Shallow water fouling communities (0.5 m); Central California to Avalon Harbor, Catalina Island and Point Loma, San Diego, southern California, USA. Body length to ~ 1.2 mm. See Cadien (2012: voucher sheet for original provisional species designation Pleurocope sp. IS1 available from TD Stebbins; specimens examined 4 March 2012, DB Cadien), SCAMIT (2012–2021), and prior SCB reports of P.floridensis in Maloney et al. (2006, 2008). Note: See Endnote 25.
Superfamily Stenetrioidea
Family Stenetriidae
Stenetrium sp. A [Fig. 31C] Continental shelf, hard bottom benthos (90–131 m); Santa Maria Basin offshore of Purisima Point, Santa Barbara County, California, USA (~ 35 km north of SCB northern boundary). Body length to ~ 7.3 mm. See Wilson (1997: 69–70, fig. 1.26, description and figures). Note: See Lissner et al. (1986) for previous report of likely same species from the region.
Suborder Oniscidea
Superfamily Oniscoidea
Family Alloniscidae
Alloniscusmirabilis (Stuxberg, 1875) [Fig. 43A] Littoral halophilic species common on sandy beaches, burrows into sand under driftwood above high tide line (may co-occur with A.perconvexus); San Mateo County, northern California, USA to Magdalena Bay, Baja California Sur, Mexico. Type locality: USA, California. Body length to ~ 9.4 mm. See Stuxberg (1875 original description as Rhinoryctesmirabilis), Richardson (1899, 1900c, 1905a), Stafford (1913a as A.cornutusvar.lagunae), Schultz (1984), Garthwaite and Lawson (1992), Leistikow and Wägele (1999), Jass and Klausmeier (2000, 2001), Schmalfuss (2003), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), and Campos-Filho et al. (2018).
Alloniscusperconvexus Dana, 1854 [Fig. 43B] Littoral halophilic species common on sandy beaches, burrows into sand under driftwood above most recent high tide line (may co-occur with A.mirabilis or Tylospunctatus in southern California); British Columbia, Canada to Magdalena Bay, Baja California Sur, Mexico. Type locality: USA, California. Body length to ~ 12 mm. See Dana (1854), Richardson (1899, 1900c, 1905a), Shearer (1942), Hewatt (1946), Hatch (1947), Brusca (1966), George and Strömberg (1968), Miller (1975), Lee and Miller (1980), Schultz (1984), Ricketts et al. (1985), Garthwaite and Lawson (1992), Leistikow and Wägele (1999), Jass and Klausmeier (2000, 2001), Espinosa-Pérez and Hendrickx (2001a, 2006), Schmalfuss (2003), McLaughlin et al. (2005), Brusca et al. (2007), SCAMIT (2011–2021), Hubbard et al. (2014), Campos-Filho et al. (2018), and Wright and Harris (2020).
Family Detonidae
Armadilloniscuscoronacapitalis Menzies, 1950 [Fig. 43C] Littoral halophilic species, occurring under stones, driftwood, decaying eelgrass, or other debris along high tide line of marshes, bays, and estuaries; Marin County, northern California to San Miguel and Anacapa Islands, southern California, USA. Type locality: USA, California, Marin County, Tomales Bay, Tomales Point. Body length to ~ 4.6 mm. See Menzies (1950c), Miller (1975), Taiti and Ferrara (1989), Garthwaite and Lawson (1992), Garthwaite et al. (1992), Leistikow and Wägele (1999), Jass and Klausmeier (2000, 2001), Schmalfuss (2003), McLaughlin et al. (2005), and Espinosa-Pérez and Hendrickx (2006).
Armadilloniscusholmesi Arcangeli, 1933 [Fig. 43D] Littoral halophilic species, occurring under stones, driftwood, decaying eelgrass, or other debris along high tide line of marshes, bays and estuaries, and in the spray zone on rocky beaches: British Columbia, Canada to Magdalena Bay, Baja California Sur, Mexico. Type locality: USA, California, San Diego County, San Diego. Body length to ~ 3.9 mm. See Arcangeli (1933), Menzies (1950c), Miller (1975), Taiti and Ferrara (1989), Garthwaite and Lawson (1992), Garthwaite et al. (1992), Leistikow and Wägele (1999), Jass and Klausmeier (2000, 2001), Schmalfuss (2003), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Wright and Harris (2020), and SCAMIT (2021).
Armadilloniscuslindahli (Richardson, 1905) [Fig. 43E] Littoral halophilic species, occurring under stones, driftwood, decaying eelgrass, or other debris along high tide line of marshes, bays and estuaries, and in the spray zone on rocky beaches; Tomales Bay, northern California to San Diego, southern California, USA. Type locality: USA, California, Oakland. Body length to ~ 4.5 mm. See Richardson (1905a original description as Actoniscuslindahli), Menzies (1950c), Miller (1975), Taiti and Ferrara (1989), Garthwaite and Lawson (1992), Garthwaite et al. (1992), Leistikow and Wägele (1999), Jass and Klausmeier (2000, 2001), Schmalfuss (2003), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), and Wright and Harris (2020).
Family Halophilosciidae
Littorophilosciarichardsonae (Holmes & Gay, 1909) [Fig. 42B] Littoral halophilic species common above the high tide line in marshes, bays and estuaries, and in the spray zone on rocky beaches; Vancouver Island, British Columbia, Canada to Cedros Island, Baja California Norte. Mexico. Type locality: USA, California, San Diego, on moist swampy ground. Body length to ~ 5 mm. See Holmes and Gay (1909 original description as Philosciarichardsonae), Hatch (1947), George and Strömberg (1968), Miller (1975), Taiti and Ferrara (1986), Garthwaite and Lawson (1992), Leistikow and Wägele (1999), Jass and Klausmeier (2000, 2001), Schmalfuss (2003), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Whitcraft et al. (2008), Roletto et al. (2014), Shaughnessy et al. (2017), and Wright and Harris (2020).
Family Ligiidae
Ligiacf.occidentalis (Dana, 1853) [Fig. 42A, image of representative species L.occidentalis] Spray zone of high intertidal rocky shores; Refugio (south of Point Conception), southern California, USA to Puerto San Carlos, Baja California Sur, Mexico. Type locality: not applicable, but near the Sacramento River, California, USA for L.occidentalis [overall range: Oregon, USA to Chamela Bay, Jalisco, Mexico, and Gulf of California]. Body length to ~ 25 mm (without uropods). See Dana (1853 original description as Lygiaoccidentalis), Richardson (1899, 1900c, 1905a), Hewatt (1946), Brusca (1966), Miller (1975), Allen (1976), Lee and Miller (1980), Ricketts et al. (1985), Garthwaite and Lawson (1992), Leistikow and Wägele (1999), Jass and Klausmeier (2000, 2001), Espinosa-Pérez and Hendrickx (2001a, 2002, 2006), Schmalfuss (2003), Brusca et al. (2004, 2007), McLaughlin et al. (2005), Campos and Villarreal (2008), Hurtado et al. (2010), Eberl (2012), SCAMIT (2012–2021), Eberl et al. (2013), Roletto et al. (2014), Santamaria et al. (2016), and Wright and Harris (2020). Note: References cited above cover L.occidentalis in general (including L.cf.occidentalis as recognized herein); see Endnote 30.
Family Platyarthridae
Niambiacapensis (Dollfus, 1895) [Fig. 42C] Supralittoral and riparian species, occurring on sandy beaches and the edges of marshes in beach wrack, holdfasts, and under driftwood, logs, and rocks; introduced species from Namibia and South Africa, occurring along Pacific coast of North America from southern Washington State to southern California, USA, and in continental Chile and on Saint Helena Island in the South Atlantic Ocean. Type locality: Africa, Namibia. Body length to ~ 5.4 mm. See Dollfus (1895 original description as Metoponorthuscapensis), Miller (1936 as Porcelliolittorinus), Leistikow and Wägele (1999), Jass and Klausmeier (2000, 2001), Schmalfuss (2003), McLaughlin et al. (2005), Espinosa-Pérez and Hendrickx (2006), Brusca et al. (2007), Maloney et al. (2008), Pérez-Schultheiss et al. (2018), and Pérez-Schultheiss and Urra (2020).
Family Tylidae
Tylospunctatus Holmes & Gay, 1909 [Fig. 42D] Littoral halophilic species on sandy beaches, burrows in sand above the most recent high tide line during the day and is active on the surface at night (may co-occur with Alloniscusperconvexus in southern California); San Diego, California, USA to Ensenada, Baja California Norte, Mexico, and the Gulf of California. Type locality: USA, California, San Diego, in sand near the beach. Body length to ~ 10 mm. See Holmes and Gay (1909), Stafford (1913b), Hewatt (1946), Schultz (1970), Brusca (1980), Ricketts et al. (1985), Leistikow and Wägele (1999), Jass and Klausmeier (2000, 2001), Espinosa-Pérez and Hendrickx (2001a, 2006), Schmalfuss (2003), Brusca et al. (2004, 2007), McLaughlin et al. (2005), SCAMIT (2011–2021), Hubbard et al. (2014), and Wright and Harris (2020).
Discussion
The marine habitats of the Southern California Bight (SCB) are characterized by a diverse isopod fauna with species occurring from the upper intertidal of rocky shores and sandy beaches, to the shallow waters of local bays and estuaries, and in the deeper offshore benthos of the continental shelf, slope, basins, and submarine canyons (Table 2). The offshore shelf, composed of soft and hard bottom substrates at depths to ~ 200 m, houses the greatest diversity of SCB isopods (119 species), while the various intertidal habitats have the second highest diversity (89 species). Fewer species have been reported from the shallow subtidal waters of SCB bays and estuaries (13 species) as well as deeper waters of the continental slope to ~ 1000 m (38 species) and below (15 species). However, this pattern may be due in part to less intensive sampling that has been conducted in the deeper areas compared to the shelf and intertidal habitats.
Table 2.
Diversity of isopods by habitat type in the Southern California Bight. Data are expressed as the number of species per taxon occurring in each habitat. Intertidal habitats include outer coast rocky shores and sandy beaches, as well as soft and hard substrates along the edges of marshes, bays, and estuaries. Subtidal habitats include shallow bays and estuaries, plus shallow to deep waters of the continental shelf, slope, basins, and submarine canyons.
| Taxon | Intertidal | Bays & estuaries | Shelf, slope, basins, canyons | ||
|---|---|---|---|---|---|
| < 200 m | 200–1000 m | > 1000 m | |||
| Suborder Cymothoida | 20 | 7 | 47 | 19 | 8 |
| Superfamily Anthuroidea | 4 | 2 | 10 | 2 | 1 |
| Superfamily Cymothooidea | 8 | 1 | 24 | 14 | 5 |
| Infraorder Epicaridea | |||||
| Superfamily Bopyroidea | 7 | 4 | 11 | 1 | 1 |
| Superfamily Cryptoniscoidea | 1 | – | 2 | 2 | 1 |
| Suborder Limnorioidea | 3 | – | 1 | – | – |
| Suborder Sphaeromatidea | 22 | 4 | 16 | – | – |
| Superfamily Seroloidea | 1 | 1 | 1 | – | – |
| Superfamily Sphaeromatoidea | 21 | 3 | 15 | – | – |
| Suborder Valvifera | 18 | 1 | 22 | 1 | 2 |
| Suborder Asellota | 17 | 1 | 33 | 18 | 5 |
| Superfamily Janiroidea | 17 | 1 | 32 | 18 | 5 |
| Superfamily Stenetrioidea | – | – | 1 | – | – |
| Suborder Oniscoidea | 9 | – | – | – | – |
| Total No. Species | 89 | 13 | 119 | 38 | 15 |
Overall, 190 species of isopods are recognized in this review as likely to occur in the SCB. However, seven of these species represent provisional taxa confirmed from just a few sites located ~ 30–90 km north of the SCB northern boundary (i.e., between Point Conception and San Luis Obispo) that were sampled during the benthic survey of the Santa Maria Basin and Western Santa Barbara Channel in the 1980s (see Wetzer and Brusca 1997; Wetzer et al. 1997; Wilson 1997). These species include the anthuroid Kupellonura sp. A, and the asellotes Desmosoma sp. A, Janiralata sp. C, Janiralata sp. D, Joeropsis sp. A, Prochelator sp. A, and Stenetrium sp. A that occurred at depths between ~ 50–300 m. Of these species, Prochelator sp. A has also been reported from much deeper waters (2955 m) located further north in the Gulf of the Farallones off San Francisco, northern California (Blake et al. 2009). It seems reasonable to expect that each of the above seven species may also range at least a little further south into northern SCB waters and therefore be encountered in future surveys of the region.
Approximately 22% of the isopods treated herein (41 species) appear restricted to SCB waters at this time. However, fewer than half of these isopods have been formally described, while the remainder represent distinct provisional species of which many are known from only one or a few samples. Thus, it seems likely that some or all these provisional species may have broader geographic distributions than summarized in this review. In contrast, most of the species covered in this paper (~ 76%) have broader eastern Pacific distributions. For example, 111 of the species covered (~ 58%) also occur north of the SCB ranging variously along the coasts of central and northern California to Oregon, Washington, British Columbia, Alaska, and into the Bering Sea. Additionally, 73 species (~ 38%) also range south of the SCB in the eastern Pacific, occurring along the southwestern coast of Baja California, throughout the Gulf of California, and off the western coasts of Mexico, Central and South America, and the Galapagos Islands.
Twenty-seven of the SCB isopod species also occur in other areas of the world beyond the eastern Pacific, some of which have been introduced to new regions or distributed globally by international shipping. These wide-ranging species include one paranthurid (Paranthurajaponica), two aegids (Aegiochusplebeia and Rocinelasignata), one cirolanid (Excirolanachiltoni), two cymothoids (Ceratothoagaudichaudii and Renocilathresherorum), six epicarideans (Aporobopyrusoviformis, Argeiapugettensis, Bathygygegrandis, Orthionegriffenis, Ionecornuta, and Hemioniscusbalani), two limnoriids (Limnoriaquadripunctata and L.tripunctata), five sphaeromatids (Paracerceissculpta, Paradelladianae, Pseudosphaeroma sp., Sphaeromaquoianum, and S.walkeri), one idoteid (Synidoteaharfordi), five asellotes (Caecijaerahorvathi, Iaiscalifornica, Ianiropsisderjugini, Ianiropsistridens, and Munnastephenseni), and one oniscid (Niambiacapensis). Also occasionally occurring in SCB waters during warm water years is the cosmopolitan pelagic species Idoteametallica. See the annotated species list provided in this review for additional details and references regarding each of the above species.
Some species may have been introduced multiple times, and others have become established residents in their new localities with expanding ranges. For example, based on mitochondrial data, the sphaeromatid Pseudosphaeroma sp. appears to have been introduced multiple times from the southern hemisphere (RW, pers. obs.). In Morro Bay located just north of the SCB, Pseudosphaeroma sp. is now known to be associated with dead barnacle tests found attached to dock pilings. Additionally, the species is known to co-occur with the sacoglossan sea slug Alderia sp. in San Francisco Bay on high intertidal mudflats, tolerating freshwater immersion following rain events. Most recently in 2018, a single specimen identified as Pseudosphaeroma sp. was collected in the Salinas de San Pedro (Los Angeles County), thus extending the range of this introduced species to the SCB (RW, pers. obs.; specimen in Natural History Museum of Los Angeles County collections, Catalog No. LACM:DISCO:9917).
Another sphaeromatid, Paracerceissculpta, best known for its multiple male morphs (designated alpha, beta, and gamma) in the northern Gulf of California (Shuster 1987, 1992), is expanding its range not only in SCB bays and harbors, but also on rocky outer coast habitats. In its newly adopted habitats, P.sculpta readily occurs on non-native bryozoans, algae, and assorted fouling organisms. Although originally described from pieces of a sponge dredged from shallow water off San Clemente Island in the SCB, P.sculpta is now widely distributed in coastal waters around the globe (Marchini et al. 2018). On the other hand, it is possible that P.sculpta as currently recognized represents more than one species. For example, the smaller beta and gamma males are known to co-occur with the larger alpha males only in northern Gulf of California populations, while populations outside of the Gulf include only alpha males. Unfortunately, there are no genetic barcode sequences available for northern Gulf of California specimens (RW and S Shuster, pers. comm.). In contrast, genetic sequences for P.sculpta occurring in the central and southern Gulf of California are similar to those from specimens collected in San Diego bays and harbors and northwards where beta and gamma males have never been noted. In other words, it is not out of the question that specimens from the northern Gulf of California are an unnamed Paracerceis species, and that the species name P.sculpta has been attributed incorrectly to isopods from that region. In fact, taxa attributed to broad geographic ranges are likely to be found to be different species upon closer examination unless it is a species with a propensity to be readily relocated by anthropogenic means. Another sphaeromatid example of such a pattern is the “Exosphaeromaamplicauda” clade referred to previously in this review (see Endnote 18), which in the past was reported as a single species ranging from the Aleutian Islands to northern Baja California. However, Wall et al. (2015) demonstrated that between Alaska and southern California there are at least five species comprising this group, and perhaps more, as these authors did not consider specimens from the Channel Islands where additional species are likely to occur. Because of such uncertainties, we acknowledge that unknown numbers of undescribed taxa are likely to become evident in the future as morphological taxonomic revisions are undertaken and new genetic data are added.
We also recognize the shortcomings of reproducing dorsal view line drawings for most species yet are grateful to previous authors for their efforts. Although dorsal views have long been the accepted standard depiction for isopods, such views are often not especially useful for identifying some species. For example, identification of some asellotes and arcturid valviferans would greatly benefit from lateral views. Ventral views depicting the pleotelson features of valviferans and sphaeromatids would also improve and simplify many identifications. However, such views do not exist for most taxa. High quality color photographs of live specimens would also be a vast improvement in facilitating accurate species identifications. Fortunately, with the increasing popularity of community platforms such as iNaturalist, in-situ color photographs of live animals are helping to expand our knowledge of species distributions, as well as contributing new and valuable habitat information. As videos of living specimens become more common, insights into the behavior and habitat use of species should also improve.
Several other active projects are also moving forward that should further our understanding of the biodiversity and taxonomy of many groups. For example, the National Science Foundation’s digitization efforts of natural history museum collections, such as the current 4-year effort underway at the Natural History Museum of Los Angeles County in southern California (DigIn: https://www.digin-tcn.org), involving 19 US collections, are making biological specimens that have been silently sitting on basement shelves for the past hundred years available on the web. Some collections such as the National Museum of Natural History (Smithsonian Institution) already have most of their collections digitized. Similarly, many collections in Europe, Australia, and New Zealand have all or most of their collections publicly available or are similarly working on capturing specimen metadata and publishing it. Consequently, these data are now becoming publicly available through existing portals, including the Integrated Digitized Biocollections (iDigBio.org) and Global Biodiversity Information Facility (GBIF), using standardized data formats. Genetic sequence data are also accumulating in the NIH genetic sequence database (GenBank) and the Barcode of Life Data Systems (BOLD). Electronic specimen data portals, such as InvertEBase https://invertebase.org/portal/ (based on Symbiota, open-source software for managing and mobilizing biodiversity data), are supporting a growing network of natural history collections, but will need continuing refinements in order to support regional needs and improve user accessibility and applicability. The Biodiversity Heritage Library (BHL) has made enormous amounts of previously inaccessible primary taxonomic literature readily available. Together these resources are game changers for accessibility to worldwide marine biodiversity data and our understanding of the changing marine environment around us.
The next step, just now beginning, is to knit the systems above (and others like them) into an integrated architecture of biodiversity knowledge. The goal is to realize the ideals expressed in the emerging “Extended Specimen” concept (see Hedrick et al. 2020; Lendemer et al. 2020; Miller et al. 2020; NASEM 2020; Heberling et al. 2021; Hardisty et al. 2022). In this view, specimens (e.g., type specimens) stand at the center of an interlinked cloud of digitally accessible information, enabling taxonomic understanding of biodiversity by combining local and non-local specimen-based data (Rocha et al. 2014; Meyer et al. 2021). Therefore, we hope that future versions of this guide to the SCB marine isopod fauna will function as a direct entry point to all available specimen-based data and taxonomic research relevant to the region’s taxonomy and biogeography.
Acknowledgements
The following people are gratefully acknowledged for their assistance in completing this review of the SCB coastal marine isopod fauna, including responding to numerous queries for information, answering questions and providing advice, locating and supplying specimens for examination, providing updated taxonomic information or distributional data for various species, accessing difficult to acquire literature, preparing illustrations or granting permission to reuse published figures, and reviewing or testing draft keys and more: Rick Brusca (University of Arizona); Don Cadien (Los Angeles County Sanitation Districts); Lisa Haney (Irvine Ranch Water District); Ami Latker, Ron Velarde, Kathy Langan and Wendy Enright (City of San Diego Marine Biology Laboratory); Ed Parnell (Scripps Institution of Oceanography); Dean Pasko (Pasko Taxonomic Services); Dean Pentcheff, Adam Wall and Vijay Barve (Natural History Museum of Los Angeles County); Lan Wiborg (Orange County Sanitation District); Dave Gillett and Ken Schiff (Southern California Coastal Water Research Project); Christopher Boyko (American Museum of Natural History); Niel Bruce (Queensland Museum); John Markham (Arch Cape Marine Laboratory); Gary Poore (Museums Victoria); Stefano Taiti (Istituto di Ricerca sugli Ecosistemi Terrestri, Consiglio Nazionale delle Ricerche); Buz Wilson (Saugatuck Natural History Laboratory); Bill Moser and Karen Reed (Smithsonian Institution), Shelly Moore and Dan Pondella (Southern California Academy of Sciences). Approval to reuse previously published illustrations as credited in the figure captions was acquired through various means, including many that were freely available due to being in the public domain, being published open access or under a creative commons license, or via a stated free-use policy of a specific journal, publisher, or institution. Other approvals were granted by the author(s) and/or journal/publisher of a specific paper, or via requests submitted to the Copyright Clearance Center. We thank everyone who assisted with these requests. Institutional support to TDS during the early phases of this project was provided courtesy of Alan Langworthy (City of San Diego Public Utilities Department). Additional institutional support to TDS and RW during the completion of this project was provided courtesy of the Marine Biodiversity Center and Research and Collections Branch of the Natural History Museum of Los Angeles County (NHM). We also greatly appreciate the generous financial support for publication costs provided by NHM Research and Collections, as well as from a SCAMIT Taxonomic Support Grant awarded by the Southern California Association of Marine Invertebrate Taxonomists. Finally, we thank Christopher Boyko, Emanuel Pereira, and Gary Poore for their thorough reviews and suggestions that significantly improved the final manuscript.
This is Contribution Number 14 of the NHM Diversity Initiative of the Southern California Ocean (DISCO).
Appendix 1
Table A1.
Systematic treatment and geographic distribution summary for Southern California Bight marine isopods (Crustacea: Isopoda). See annotated species list for details of geographic ranges. Distribution codes: BCN = Baja California Norte; BCS = Baja California Sur; CA = California; MX = Mexico; SCB = Southern California Bight; WW = worldwide (i.e., species also reported from regions beyond Eastern Pacific).
| Taxon | Eastern Pacific Distribution |
|---|---|
| Suborder Cymothoida | |
| Superfamily Anthuroidea | |
| Family Antheluridae | |
| Ananthuraluna (Schultz, 1966) | SCB |
| Family Anthuridae | |
| Amakusanthuracaliforniensis (Schultz, 1964) | SCB – Isla Guadalupe, BCN |
| Cyathuramunda Menzies, 1951 | Northern CA – Gulf of CA |
| Haliophasmageminatum Menzies & Barnard, 1959 | British Columbia – San Quintin Bay, BCN |
| Mesanthuraoccidentalis Menzies & Barnard, 1959 | SCB – Costa Rica |
| Family Expanathuridae | |
| Eisothistos sp. A | SCB |
| Family Hyssuridae | |
| Kupellonura sp. A | ~ 35 km north of SCB |
| Family Paranthuridae | |
| Califanthurasquamosissima (Menzies, 1951) | Northern CA – Oaxaca, MX |
| Colanthurabruscai Poore, 1984 | SCB – Costa Rica |
| Paranthuraelegans Menzies, 1951 | Northern CA – Costa Rica |
| Paranthurajaponica Richardson, 1909 | Northern CA – SCB (+ WW) |
| Superfamily Cymothooidea | |
| Family Aegidae | |
| Aegalecontii (Dana, 1854) | Central CA – SCB |
| Aegiochusplebeia (Hansen, 1897) | Aleutian Islands – Peru (+ West Pacific) |
| Rocinelaangustata Richardson, 1904 | Bering Sea – Gulf of CA |
| Rocinelabelliceps (Stimpson, 1864) | Alaska – Gulf of CA |
| Rocinelalaticauda Hansen, 1897 | Central CA – Acapulco, MX |
| Rocinelamurilloi Brusca & Iverson, 1985 | Central CA – Chile |
| Rocinelasignata Schioedte & Meinert, 1879 | Northern CA – Ecuador + Galapagos (+ WW) |
| Family Cirolanidae | |
| Cirolanadiminuta Menzies, 1962 | SCB – Gulf of CA + Galapagos |
| Cirolanaharfordi (Lockington, 1877) | British Columbia – Magdalena Bay, BCS |
| Eurydicecaudata Richardson, 1899 | SCB – Ecuador + Galapagos |
| Excirolanachiltoni (Richardson, 1905) | British Columbia – SCB (+ WW) |
| Excirolanalinguifrons (Richardson, 1899) | Oregon – SCB |
| Metacirolanajoanneae (Schultz, 1966) | British Columbia – SCB |
| Natatolanacaliforniensis (Schultz, 1966) | SCB – Peru-Chile Trench |
| Family Corallanidae | |
| Excorallanatricornisoccidentalis Richardson, 1905 | SCB – Panama |
| Excorallanatruncata (Richardson, 1899) | SCB – Galapagos |
| Family Cymothoidae | |
| Ceratothoagaudichaudii (H. Milne Edwards, 1840) | SCB – Cape Horn (+ SW Atlantic) |
| Ceratothoagilberti (Richardson, 1904) | SCB – Mazatlán, MX |
| Elthusacalifornica (Schioedte & Meinert, 1884) | Alaska – Peru |
| Elthusamenziesi (Brusca, 1981) | Coronado Islands, BCN – Gulf of CA |
| Elthusavulgaris (Stimpson, 1857) | Oregon – Columbia |
| Mothocyarosea Bruce, 1986 | SCB – Nicaragua |
| Nerocilaacuminata Schioedte & Meinert, 1881 | SCB – Peru |
| Renocilathresherorum Williams & Bunkley-Williams, 1980 | SCB – Gulf of CA (+ Egypt) |
| Smenispaconvexa (Richardson, 1905) | SCB |
| Family Gnathiidae | |
| Caecognathiacrenulatifrons (Monod, 1926) | British Columbia – BCN |
| Caecognathiasanctaecrucis (Schultz, 1972) | British Columbia – BCN |
| Caecognathia sp. A | SCB |
| Caecognathia sp. SD1 | SCB |
| Gnathiaclementensis Schultz, 1966 | SCB |
| Gnathiacoronadoensis Schultz, 1966 | SCB |
| Gnathiaproductatridens Menzies & Barnard, 1959 | SCB |
| Gnathiasteveni Menzies, 1962 | British Columbia – SCB |
| Gnathiatridens Menzies & Barnard, 1959 | British Columbia – SCB |
| Gnathiatrilobata Schultz, 1966 | British Columbia – SCB |
| Gnathia sp. MBC1 | SCB |
| Family Tridentellidae | |
| Tridentellaglutacantha Delaney & Brusca, 1985 | Central CA – SCB |
| Tridentellaquinicornis Delaney & Brusca, 1985 | SCB |
| Infraorder Epicaridea | |
| Superfamily Bopyroidea | |
| Family Bopyridae | |
| Anathelgeshyphalus (Markham, 1974) | Central CA – SCB |
| Aporobopyrusmuguensis Shiino, 1964 | Northern CA – Central Baja CA |
| Aporobopyrusoviformis Shiino, 1934 | SCB (+ Japan) |
| Argeiapugettensis Dana, 1853 | Bering Sea – SCB (+ Japan, Korea) |
| Asymmetrioneambodistorta Markham, 1985 | SCB |
| Bathygygegrandis Hansen, 1897 | SCB – MX (+ WW) |
| Bopyrellacalmani (Richardson, 1905) | Central CA – SCB |
| Capitetragonia sp. A | SCB |
| Eremitionegiardi (Calman, 1898) | Bering Sea – SCB |
| Leidyainfelix Markham, 2002 | SCB – San Quintin Bay, BCN |
| Munidionpleuroncodis Markham, 1975 | Central CA – Central MX |
| Orthionegriffenis Markham, 2004 | British Columbia – SCB (+ Asia) |
| Phyllodurusabdominalis Stimpson, 1857 | British Columbia – BCN |
| Progebiophilusbruscai Salazar-Vallejo & Leija-Tristán, 1990 | SCB – Nicaragua |
| Pseudionegalacanthae Hansen, 1897 | British Columbia – Gulf of CA |
| Schizobopyrinastriata (Nierstrasz & Brender à Brandis, 1929) | SCB – Gulf of CA |
| Family Ionidae | |
| Ionecornuta Spence Bate, 1863 | British Columbia – Gulf of CA (+ Asia) |
| Superfamily Cryptoniscoidea | |
| Family Dajidae | |
| Holophryxusalaskensis Richardson, 1905 | Alaska – SCB |
| Zonophryxus sp. | SCB |
| Family Hemioniscidae | |
| Hemioniscusbalani Buchholz, 1866 | British Columbia – MX (+ WW) |
| Suborder Limnoriidea | |
| Superfamily Limnorioidea | |
| Family Limnoriidae | |
| Limnoriaalgarum Menzies, 1957 | British Columbia – SCB |
| Limnoriaquadripunctata Holthuis, 1949 | Northern CA – SCB (+ WW) |
| Limnoriatripunctata Menzies, 1951 | Washington – Mazatlán, MX (+ WW) |
| Suborder Sphaeromatidea | |
| Superfamily Seroloidea | |
| Family Serolidae | |
| Heteroseroliscarinata (Lockington, 1877) | SCB – San Quintin Bay, BCN, + Gulf of CA |
| Superfamily Sphaeromatoidea | |
| Family Ancinidae | |
| Ancinusgranulatus Holmes & Gay, 1909 | SCB – Gulf of CA |
| Ancinusseticomvus Trask, 1971 | SCB – Mazatlán, MX |
| Bathycopeadaltonae (Menzies & Barnard, 1959) | Washington – SCB |
| Family Sphaeromatidae | |
| Discerceisgranulosa (Richardson, 1899) | SCB – Isla Cedros, BCN |
| Dynamenelladilatata (Richardson, 1899) † | Washington – SCB |
| Dynamenellaglabra (Richardson, 1899) † | Oregon – SCB |
| Dynamenellasheareri (Hatch, 1947) | Washington – SCB |
| Dynoideselegans (Boone, 1923) | SCB – Isla Cedros, BCN |
| Exosphaeromaamplicauda (Stimpson, 1857) ‡ | Northern CA |
| Exosphaeromaaphrodita Boone, 1923 | SCB |
| Exosphaeromainornata Dow, 1958 | Washington – SCB |
| Exosphaeromapentcheffi Wall, Bruce & Wetzer, 2015 | SCB |
| Exosphaeromarhomburum (Richardson, 1899) | Washington – SCB |
| Gnorimosphaeromanoblei Menzies, 1954 | Northern CA – SCB |
| Gnorimosphaeromaoregonense (Dana, 1853) ‡ | British Columbia – Central CA |
| Paracerceiscordata (Richardson, 1899) | Aleutian Islands – SCB |
| Paracerceisgilliana (Richardson, 1899) | Northern CA – SCB |
| Paracerceissculpta (Holmes, 1904) | Central CA – Gulf of CA (+ WW) |
| Paracerceis sp. A | SCB + Gulf of CA |
| Paradelladianae (Menzies, 1962) | SCB – Gulf of CA (+ WW) |
| Pseudosphaeroma sp. | Northern CA – SCB (+ WW) |
| Sphaeromaquoianum H. Milne Edwards, 1840 | Northern CA – SCB (+ WW) |
| Sphaeromawalkeri Stebbing, 1905 | SCB (+ WW) |
| Family Tecticipitidae | |
| Tecticepsconvexus Richardson, 1899 | Washington – SCB |
| Suborder Valvifera | |
| Family Arcturidae | |
| Idarcturusallelomorphus Menzies & Barnard, 1959 | Central CA – SCB |
| Idarcturushedgpethi Menzies, 1951 | British Columbia – SCB |
| Idarcturus sp. A | SCB |
| Neastacillacalifornica (Boone, 1918) | SCB – Gulf of CA |
| Family Holognathidae | |
| Cleantioidesoccidentalis (Richardson, 1899) | SCB – Gulf of CA, Ecuador + Galapagos |
| Family Idoteidae | |
| Colidoteafindleyi Brusca & Wallerstein, 1977 | SCB – Guadalupe Isla, BCN + Gulf of CA |
| Colidotearostrata (Benedict, 1898) | Oregon – SCB |
| Colidoteawallersteini Brusca, 1983 | SCB – Guadalupe Isla, BCN |
| Edotiasublittoralis Menzies & Barnard, 1959 | British Columbia – Gulf of CA + Costa Rica |
| Edotia sp. B | SCB |
| Erichsonellacrenulata Menzies, 1950 | SCB – San Quintin Bay, BCN |
| Eusymmeruspseudoculata (Boone, 1923) | SCB |
| Idoteafewkesi Richardson, 1905 | Alaska – BCS |
| Idoteametallica Bosc, 1801 | WW (pelagic), including SCB |
| Idotearufescens Fee, 1927 | Alaska – Baja CA |
| Idoteaurotoma Stimpson, 1864 | Alaska – Gulf of CA |
| Pentidoteaaculeata Stafford, 1913 | British Columbia – Gulf of CA |
| Pentidoteakirchanskii (Miller & Lee, 1970) | Oregon – SCB |
| Pentidoteamontereyensis Maloney, 1933 | Alaska – Baja CA |
| Pentidotearesecata (Stimpson, 1857) | Alaska – Gulf of CA |
| Pentidoteaschmitti (Menzies, 1950) | Bering Sea – SCB |
| Pentidoteastenops (Benedict, 1898) | Alaska – Gulf of CA |
| Pentidoteawosnesenskii (Brandt, 1851) | Aleutian Islands – Gulf of CA |
| Stenosomawetzerae (Ormsby, 1991) | SCB – Gulf of CA |
| Synidoteacalcarea Schultz, 1966 | SCB |
| Synidoteaharfordi Benedict, 1897 | Oregon – Costa Rica (+ Japan) |
| Synidoteamagnifica Menzies & Barnard, 1959 | Central CA – SCB |
| Synidoteamedia Iverson, 1972 | British Columbia – SCB |
| Family Thermoarcturidae | |
| Califarcturustannerensis (Schultz, 1966) | SCB |
| Suborder Asellota | |
| Superfamily Janiroidea | |
| Family Dendrotionidae | |
| Acanthomunnatannerensis Schultz, 1966 | SCB |
| Family Desmosomatidae | |
| Desmosoma sp. A | ~ 89 km north of SCB |
| Momedossasymmetrica (Schultz, 1966) | Northern CA – SCB |
| Prochelator sp. A | Northern CA to ~ 35 km north of SCB |
| Family Haplomunnidae | |
| Haplomunnacaeca (Richardson, 1905) | SCB |
| Family Janiridae | |
| Caecianiropsis sp. LA1 | SCB |
| Caecianiropsis sp. LA2 | SCB |
| Caecijaerahorvathi Menzies, 1951 | SCB (+ Hawaii) |
| Iaiscalifornica (Richardson, 1904) | Northern CA – SCB (+ WW) |
| Ianiropsisanaloga Menzies, 1952 | Washington – SCB |
| Ianiropsisderjugini Gurjanova, 1933 | Bering Sea – SCB (+ Korea) |
| Ianiropsiskincaidi Richardson, 1904 | Bering Sea – SCB |
| Ianiropsisminuta Menzies, 1952 | Northern CA – SCB |
| Ianiropsismontereyensis Menzies, 1952 | Northern CA – SCB |
| Ianiropsistridens Menzies, 1952 | British Columbia – Chile (+ Korea) |
| Janiralatadavisi Menzies, 1951 | Central CA – SCB |
| Janiralataoccidentalis (Walker, 1898) | British Columbia – SCB |
| Janiralatarajata Menzies, 1951 | Central CA – SCB |
| Janiralatasolasteri (Hatch, 1947) | Alaska – SCB |
| Janiralata sp. A | SCB |
| Janiralata sp. B | SCB |
| Janiralata sp. C | ~ 35 km north of SCB |
| Janiralata sp. D | ~ 35 km north of SCB |
| Family Joeropsididae | |
| Joeropsisconcava Schultz, 1966 | Central CA – SCB |
| Joeropsisdubiadubia Menzies, 1951 | British Columbia – Gulf of CA |
| Joeropsisdubiapaucispinis Menzies, 1951 | Northern CA – SCB |
| Joeropsislobata Richardson, 1899 | Oregon to SCB |
| Joeropsis sp. A | ~ 30 km north of SCB |
| Family Lepidocharontidae | |
| Microcharon sp. A | SCB |
| Family Munnidae | |
| Munnachromatocephala Menzies, 1952 | British Columbia – SCB |
| Munnafernaldi George & Strömberg, 1968 | Washington – SCB |
| Munnahalei Menzies, 1952 | Northern CA – SCB |
| Munnamagnifica Schultz, 1964 | SCB |
| Munnaspinifrons Menzies & Barnard, 1959 | SCB |
| Munnastephenseni Gurjanova, 1933 | Bering Sea – SCB (+ Japan) |
| Munna sp. A | SCB |
| Uromunnaubiquita (Menzies, 1952) | British Columbia – Gulf of CA |
| Family Munnopsidae | |
| Belonectes sp. A | SCB |
| Eurycopecaliforniensis Schultz, 1966 | SCB |
| Ilyarachnaacarina Menzies & Barnard, 1959 | Northern CA – SCB |
| Ilyarachnaprofunda Schultz, 1966 | Northern CA – SCB |
| Munnopsurus sp. A | SCB |
| Munnopsurus sp. B | SCB |
| Family Nannoniscidae | |
| Nannonisconuslatipleonus Schultz, 1966 | SCB |
| Family Paramunnidae | |
| Austrosignumlatum Just & Wilson, 2021 | SCB |
| Boreosignum sp. A | SCB |
| Munnogoniumerratum (Schultz, 1964) | Central CA – SCB |
| Munnogoniumtillerae (Menzies & Barnard, 1959) | SCB |
| Paramunnaquadratifrons Iverson & Wilson, 1981 | SCB |
| Paramunna sp. A | SCB |
| Paramunna sp. SD1 | SCB |
| Pleurogoniumcaliforniense Menzies, 1951 | British Columbia – SCB |
| Pleurogonium sp. A | SCB |
| Family Pleurocopidae | |
| Pleurocope sp. A | Central CA – SCB |
| Superfamily Stenetrioidea | |
| Family Stenetriidae | |
| Stenetrium sp. A | ~ 35 km north of SCB |
| Suborder Oniscidea | |
| Superfamily Oniscoidea | |
| Family Alloniscidae | |
| Alloniscusmirabilis (Stuxberg, 1875) | Northern CA – Magdalena Bay, BCS |
| Alloniscusperconvexus Dana, 1854 | British Columbia – Magdalena Bay, BCS |
| Family Detonidae | |
| Armadilloniscuscoronacapitalis Menzies, 1950 | Northern CA – SCB |
| Armadilloniscusholmesi Arcangeli, 1933 | British Columbia – Magdalena Bay, BCS |
| Armadilloniscuslindahli (Richardson, 1905) | Northern CA – SCB |
| Family Halophilosciidae | |
| Littorophilosciarichardsonae (Holmes & Gay, 1909) | British Columbia – Isla Cedros, BCN |
| Family Ligiidae | |
| Ligiacf.occidentalis (Dana, 1853) | SCB – BCS |
| Family Platyarthridae | |
| Niambiacapensis (Dollfus, 1895) | Washington – Chile (+ WW) |
| Family Tylidae | |
| Tylospunctatus Holmes & Gay, 1909 | SCB – Gulf of CA |
† Taxon inquirendum (genus uncertain for Dynamenelladilatata and D.glabra). ‡ No longer considered a SCB species (see Wall et al. 2015 for E.amplicauda, and Wetzer et al. 2021 for G. oregonense).
Citation
Stebbins TD, Wetzer R (2023) Review and guide to the isopods (Crustacea, Isopoda) of littoral and sublittoral marine habitats in the Southern California Bight. ZooKeys 1162: 1–167. https://doi.org/10.3897/zookeys.1162.100390
References
- Abdelsalam K, Ramadan S. (2019) First record of the wood borer Limnoriatripunctata Menzies 1951, in the Egyptian Mediterranean Sea. Rapports et procés-verbaux des reunions Commission international pour l’exploration scientifique de la Mer Méditerranée 42: 178.
- Abelló P, Guerao G, Codina M. (2004) Distribution of the neustonic isopod Idoteametallica in relation to shelf-slope frontal structures. Journal of Crustacean Biology 24(4): 558–566. 10.1651/C-2505 [DOI] [Google Scholar]
- Aguilar-Perera A. (2022) Checklist of parasitic isopods (Crustacea: Isopoda) infesting marine decapod and fishes off Mexico’s coasts. Thalassas: An International Journal of Marine Sciences 38(1): 113–121. 10.1007/s41208-021-00336-x [DOI] [Google Scholar]
- Aguilar-Perera A, Nóh-Quiñones V. (2022) Prevalence of Rocinelasignata Schioedte & Meinert, 1879 (Crustacea: Isopoda: Aegidae) in hogfish Lachnolaimusmaximus (Walbaum, 1792) (Teleostei: Labridae). Thalassas: An International Journal of Marine Sciences 38(2): 1195–1198. 10.1007/s41208-022-00459-9 [DOI] [Google Scholar]
- Aguilar-Perera A, Valle-Nava J, Williams EH, Bunkley-Williams L. (2018) Rocinelasignata (Isopoda: Aegidae) parasitizing Indo-Pacific lionfish Pteroisvolitans (Scorpaeniformes: Scorpaenidae) in the Southern Gulf of Mexico. Marine Biology Research 14(8): 883–886. 10.1080/17451000.2018.1528012 [DOI] [Google Scholar]
- Ahyong ST, Lowry JK, Alonso M, Bamber RN, Boxshall GA, Castro P, Gerken S, Karaman GS, Goy JW, Jones DS, Meland K, Rogers DC, Svavarsson J. (2011) Subphylum Crustacea Brünnich, 1772. In: Zhang Z-Q. (Ed.) Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness.Zootaxa 3148(1), 165–191. 10.11646/zootaxa.3148.1.33 [DOI]
- Ali NG, Aboyadak IM. (2018) Histopathological alterations and condition factor deterioration accompanied by isopod infestation in Tilapiazilli, Mugilcapito and Soleaaegyptiaca from Lake Qaroun. Egyptian Journal of Aquatic Research 44(1): 57–63. 10.1016/j.ejar.2018.03.001 [DOI] [Google Scholar]
- Allen RK. (1976) Common Intertidal Invertebrates of Southern California. Revised Edition. Peek Publications, Palo Alto, 316 pp. [Google Scholar]
- Allen MJ, Moore SL, Schiff KC, Weisberg SB, Diener D, Stull JK, Groce A, Mubarak J, Tang CL, Gartman R. (1998) Southern California Bight 1994 Pilot Project: Vol. V. Demersal Fishes and Megabenthic Invertebrates. Technical Report 308. Southern California Coastal Water Research Project, Westminster, 146 pp. [+ Literature Cited + appendices] https://www.sccwrp.org/publications [Google Scholar]
- Allen MJ, Groce AK, Diener D, Brown J, Steinert SA, Deets G, Noblet JA, Moore SL, Diehl D, Jarvis ET, Raco-Rands V, Thomas C, Ralph Y, Gartman R, Cadien D, Weisberg SB, Mikel T. (2002) Southern California Bight 1998 Regional Monitoring Program: Vol. V. Demersal Fishes and Megabenthic Invertebrates. Technical Report 380. Southern California Coastal Water Research Project, Westminster, 572 pp. https://www.sccwrp.org/publications [Google Scholar]
- Allen MJ, Mikel TK, Cadien DB, Kalman JE, Jarvis ET, Schiff KC, Diehl DW, Moore SL, Walther S, Deets G, Cash C, Watts S, Pondella DJ II, Raco-Rands V, Thomas C, Gartman R, Sabin L, Power W, Groce AK, Armstrong JL. (2007) Southern California Bight 2003 Regional Monitoring Program: Vol. IV. Demersal Fishes and Megabenthic Invertebrates. Technical Report 505. Southern California Coastal Water Research Project, Costa Mesa, 338 pp. [+ appendices] https://www.sccwrp.org/publications [Google Scholar]
- Allen MJ, Cadien DB, Miller E, Diehl DW, Ritter K, Moore SL, Cash C, Pondella DJ, Raco-Rands V, Thomas C, Gartman R, Power W, Latker AK, Williams J, Armstrong JL, Schiff K. (2011) Southern California Bight 2008 Regional Monitoring Program: Vol. IV. Demersal Fishes and Megabenthic Invertebrates. Technical Report 655. Southern California Coastal Water Research Project, Costa Mesa, 127 pp. [+ appendices] https://www.sccwrp.org/publications [Google Scholar]
- Alves AM, Leonardo MG, Souza GTR, Takemoto RM, de Lima FS, Tavares LER, Melo CM, Madi RR, Jeraldo VLS. (2019) Occurrence of isopods in two species of snappers (Lutjanidae) from Northeast Brazil. Journal of Parasitology Research 2019(8176283): 1–8. 10.1155/2019/8176283 [DOI] [PMC free article] [PubMed] [Google Scholar]
- An J, Williams JD, Yu H. (2009) The Bopyridae (Crustacea: Isopoda) parasitic on thalassinideans (Crustacea: Decapoda) from China. Proceedings of the Biological Society of Washington 122(2): 225–246. 10.2988/08-26.1 [DOI] [Google Scholar]
- An J, Boyko CB, Li X. (2015) A review of bopyrids (Crustacea: Isopoda: Bopyridae) parasitic on caridean shrimps (Crustacea: Decapoda: Caridea) from China. Bulletin of the American Museum of Natural History 399: 1–85. 10.1206/amnb-921-00-01.1 [DOI] [Google Scholar]
- Aneesh PT, Bruce NL, Nashad M, Bineesh K, Mohamed Hatha AA. (2020) A new species of Renocila Miers, 1880 (Crustacea: Isopoda: Cymothoidae), a fish parasitic isopod from Andaman Island, India. Marine Biology Research 16(5): 396–410. 10.1080/17451000.2020.1761028 [DOI] [Google Scholar]
- Aneesh PT, Nashad M, Bijukumar A, Bineesh K, Mohamed Hatha AA. (2021) Review of the global distribution and hosts of the fish parasitic isopod genus Renocila Miers, 1880 (Crustacea: Cymothoidae) with the description of a new species from Andaman Islands, India. Journal of Natural History 55(43–44): 2761–2785. 10.1080/00222933.2021.2019341 [DOI] [Google Scholar]
- Annisaqois M, Wägele JW. (2021) Morphology and taxonomy of IsopodaAnthuroidea (Crustacea) from Sulawesi with description of six new species. European Journal of Taxonomy 768: 1–52. 10.5852/ejt.2021.768.1501 [DOI] [Google Scholar]
- Arcangeli A. (1933) Isopodi terrestri delle isole Samoa e considerazioni sopra la distribuzione geographica del genere Armadillo Latr. emend. Verh. Bollettino dei Musei di Zoologia ed Anatomia comparata della R. Università di Torino 43: 25–33. [Google Scholar]
- Ariyama H, Otani M. (2004) Paracerceissculpta (Crustacea: Isopoda: Sphaeromatidae), a newly introduced species into Osaka Bay, central Japan. Benthos Research 59(2): 53–59. 10.5179/benthos1996.59.2_53 [DOI] [Google Scholar]
- Artüz ML, Kubanç C. (2015) Two new species of Stenosoma (Isopoda, Idoteidae) from the Sea of Marmara. Crustaceana 88(10–11): 1201–1220. 10.1163/15685403-00003481 [DOI] [Google Scholar]
- Azofeifa-Solano JC, Cortés J. (2020) Chapter 24. Diversity and biology of deep-water crustaceans in Costa Rica. In: Hendrickx ME (Ed.) Deep-Sea Pycnogonids and Crustaceans of the Americas, Springer, 633–658. 10.1007/978-3-030-58410-8_24 [DOI]
- Barnard KH. (1914) Contributions to the crustacean fauna of South Africa. 3. Additions to the marine Isopoda, with notes on some previously incompletely known species. Annals of the South African Museum 10(11): 325a–358a, 359–440. 10.5962/bhl.part.9319 [DOI]
- Bastida R, Torti MR. (1972) Organismos perforantes de las costas argentinas. II. La presencia de Limnoriatripunctata Menzies, 1951 (Isopoda, Limnoriidae) en el puerto de Mar del Plata. Physis 31(82): 143–153. [Google Scholar]
- Bastida-Zavala JR, García-Madrigal MS, Rosas-Alquicira EF, López-Pérez RA, Benítez-Villalobos F, Meraz-Hernando JF, Torres-Huerta AM, Montoya-Márquez A, Barrientos-Luján NA. (2013) Marine and coastal biodiversity of Oaxaca, Mexico. Check List 9(2): 329–390. 10.15560/9.2.329 [with erratum] [DOI] [Google Scholar]
- Bate CS. (1863) On a new species of Ione. Report of the Thirty-Third Meeting of the British Association for the Advancement of Science 33: 98.
- Benedict JE. (1897) A revision of the genus Synidotea. Proceedings of the Academy of Sciences of Philadelphia 1897: 387–404. [Google Scholar]
- Benedict JE. (1898) Two new isopods of the genus Idotea from the coast of California. Proceedings of the Biological Society of Washington 12: 53–55. [Google Scholar]
- Bergen M, Weisberg SB, Cadien D, Dalkey A, Montagne D, Smith RW, Stull JK, Velarde RG. (1998) Southern California Bight 1994 Pilot Project: IV. Benthic Infauna. Southern California Coastal Water Research Project, Westminster, 260 pp. https://www.sccwrp.org/publications [Google Scholar]
- Bergen M, Weisberg SB, Smith RW, Cadien DB, Dalkey A, Montagne DE, Stull JK, Velarde RG, Ranasinghe JA. (2001) Relationship between depth, sediment, latitude, and the structure of benthic infaunal assemblages on the mainland shelf of southern California. Marine Biology 138(3): 637–647. 10.1007/s002270000469 [DOI] [Google Scholar]
- Bernardino AF, Smith CR, Baco A, Altamira I, Sumida PYG. (2010) Macrofaunal succession in sediments around kelp and wood falls in the deep NE Pacific and community overlap with other reducing habitats. Deep-Sea Research I 57(5): 708–723. 10.1016/j.dsr.2010.03.004 [DOI] [Google Scholar]
- Birstein YA. (1973) Deep water isopods (Crustacea. Isopoda) of the north-western part of the Pacific Ocean. Translated from Russian. Published for the Smithsonian Institution and the National Science Foundation, Washington, D.C. by the Indian National Scientific Documentation Centre, New Delhi, 316 pp. [Google Scholar]
- Blake JA, Maciolek NJ, Ota A, Williams IP. (2009) Long-term benthic infaunal monitoring at a deep-ocean dredged material disposal site off Northern California. Deep-sea Research. Part II, Topical Studies in Oceanography 56(19–20): 1775–1803. 10.1016/j.dsr2.2009.05.021 [DOI] [Google Scholar]
- Boone PL. (1918) Description of ten new isopods. Proceedings of the United States National Museum 54(2253): 591–604. 10.5479/si.00963801.54-2253.591 [DOI] [Google Scholar]
- Boone PL. (1923) New tanaid and isopod Crustacea from California. Proceedings of the Biological Society of Washington 36: 147–156. [Google Scholar]
- Borges LMS, Costa FO. (2014) New records of marine wood borers (Bivalvia: Teredinidae and Isopoda: Limnoriidae) from São Miguel, Azores, with a discussion of some aspects of their biogeography. Acoreana (Supplement 10): 109–116.
- Borges LMS, Merckelbach LM, Cragg SM. (2014) Biogeography of wood-boring crustaceans (Isopoda: Limnoriidae) established in European coastal waters. PLoS ONE 9(10): e109593. 10.1371/journal.pone.0109593 [DOI] [PMC free article] [PubMed]
- Bosc LAG. (1801) Histoire naturelle des Crustacés contenant leur description et leurs moeurs; avec figures dessinées, d’après nature. Deterville, Paris 2: 1–296 [pls. 6–18]. 10.5962/bhl.title.59168 [DOI]
- Bowman TE. (1977) Isopod crustaceans (except Anthuridae) collected on the Presidential Cruise of 1938. Proceedings of the Biological Society of Washington 89(57): 653–666. [Google Scholar]
- Bowman TE, Schultz GA. (1974) The isopod crustacean genus Munnogonium George and Strömberg, 1968 (Munnidae, Asellota). Proceedings of the Biological Society of Washington 87(25): 265–272. https://www.biodiversitylibrary.org/page/34672960#page/293 [Google Scholar]
- Boyko CB. (2003) New host and distribution records for Leidyabimini Pearse, 1951 in the Gulf of Mexico, with comments on related taxa and a redescription of Cardioceponpteroides Nobili, 1906 (Crustacea: Isopoda: Bopyridae: Ioninae). Gulf and Caribbean Research 15(1): 5–11. 10.18785/gcr.1501.02 [DOI] [Google Scholar]
- Boyko CB, Williams JD. (2003) A revision of Anathelges and Stegophryxus (Isopoda: Bopyridae: Athelginae) with descriptions of two new genera and one new species. Journal of Crustacean Biology 23(4): 795–813. 10.1651/C-2398 [DOI] [Google Scholar]
- Boyko CB, Williams JD. (2004) New records of marine isopods (Crustacea: Peracarida) from the Bahamas, with descriptions of two new species of epicarideans. Bulletin of Marine Science 74(2): 353–383. [Google Scholar]
- Boyko CB, Williams JD. (2021a) A review of the genus Zonophryxus Richardson, 1903 (Crustacea: Isopoda: Dajidae) with recognition of Colypurusagassizi Richardson, 1905 (Crustacea: Isopoda: Colypuridae) as a synonym of Zonophryxussimilis Searle, 1914 and the requisite sinking of Colypuridae. Proceedings of the Biological Society of Washington 134(1): 243–264. 10.2988/0006-324X-134.1.243 [DOI] [Google Scholar]
- Boyko CB, Williams JD. (2021b) New records of species of Holophryxus Richardson, 1905 (Crustacea: Isopoda: Dajidae) from the Atlantic and Pacific Oceans with comments on taxonomic issues in the genus. Bulletin of the Peabody Museum of Natural History 62(2): 111–121. 10.3374/014.062.0203 [DOI] [Google Scholar]
- Boyko CB, Bruce NL, Hadfield KA, Merrin KA, Ota Y, Poore GCB, Taiti S. (Eds) (2008. onwards) World Marine, Freshwater and Terrestrial Isopod Crustaceans database. https://www.marinespecies.org/isopoda [Accessed on 03 Mar 2023]
- Boyko CB, Moss J, Williams JD, Shields JD. (2013) A molecular phylogeny of Bopyroidea and Cryptoniscoidea (Crustacea: Isopoda). Systematics and Biodiversity 11(4): 495–506. 10.1080/14772000.2013.865679 [DOI] [Google Scholar]
- Boyko CB, Williams JD, Shields JD. (2017) Parasites (Isopoda: Epicaridea and Nematoda) from ghost and mud shrimp (Decapoda: Axiidea and Gebiidea) with descriptions of a new genus and a new species of bopyrid isopod and clarification of Pseudione Kossmann, 1881. Zootaxa 4365(3): 251–301. 10.11646/zootaxa.4365.3.1 [DOI] [PubMed] [Google Scholar]
- Brandt JF. (1851) Krebse. In: von Middendorff AT (Ed.) Reise in den äussersten Norden und Osten Sibiriens während der Jahre 1843 und 1844 mit allerhöchster Genehmigung auf Veranstaltung der Kaiserlichen Akademie der Wissenschaften zu St. Petersburg ausgeführt und in Verbindung mit vielen Gelehrten herausgegeben 2(Part 1): 77–148 [plates 5–6]. 10.5962/bhl.title.53712 [DOI]
- Brandt A. (1988) Antarctic Serolidae and Cirolanidae (Crustacea: Isopoda). In: Fricke R (Ed.) Theses Zoologicae, Vol. 10. Koeltz Scientific Books, Koenigstein, 143 pp. [Google Scholar]
- Brandt A. (1990) Antarctic Valviferans (Crustacea, Isopoda, Valvifera): New Genera, New Species and Redescriptions. E.J. Brill, Leiden, The Netherlands and New York, 176 pp. [Google Scholar]
- Brandt A, Poore GCB. (2001) Two new species of Tridentella (Crustacea: Isopoda: Tridentellidae) from Namibia. Beaufortia 51(11): 199–212. [Google Scholar]
- Brandt A, Poore GCB. (2003) Higher classification of the flabelliferan and related Isopoda based on a reappraisal of relationships. Invertebrate Systematics 17(6): 893–923. 10.1071/IS02032 [DOI] [Google Scholar]
- Brix S, Held C, Kaiser S, Jennings RM, Driskell A, Brandt A. (2021) Evolution and phylogeny of the deep-sea isopod families Desmosomatidae Sars, 1897 and Nannoniscidae Hansen, 1916 (Isopoda: Asellota). Organisms, Diversity & Evolution 21(4): 691–717. 10.1007/s13127-021-00509-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown ME, Keller JK, Whitcraft CR. (2022) Relationship of Sphaeromaquoianum to sediment characteristics and invertebrate community. Biological Invasions 24(11): 3631–3645. 10.1007/s10530-022-02867-7 [DOI] [Google Scholar]
- Bruce NL. (1981) Cirolanidae (Crustacea: Isopoda) of Australia: diagnoses of Cirolana Leach, Metacirolana Nierstrasz, Neocirolana Hale, Anopsilana Paulian & Debouteville, and three new genera – Natatolana, Politolana and Cartetolana. Marine and Freshwater Research 32(6): 945–966. 10.1071/MF9810945 [DOI] [Google Scholar]
- Bruce NL. (1984) A new family for the isopod crustacean genus Tridentella Richardson, 1905, with description of a new species from Fiji. Zoological Journal of the Linnean Society 80(4): 447–455. 10.1111/j.1096-3642.1984.tb02555.x [DOI] [Google Scholar]
- Bruce NL. (1986a) Cirolanidae (Crustacea: Isopoda) of Australia. Records of the Australian Museum (Supplement 6): 1–239. 10.3853/j.0812-7387.6.1986.98 [DOI]
- Bruce NL. (1986b) Revision of the isopod crustacean genus Mothocya Costa, in Hope, 1851 (Cymothoidae: Flabellifera), parasitic on marine fishes. Journal of Natural History 20(5): 1089–1192. 10.1080/00222938600770781 [DOI] [Google Scholar]
- Bruce NL. (1990) The genera Catoessa, Elthusa, Enipsa, Ichthyoxenus, Idusa, Livoneca, and Norileca n. gen. (Isopoda, Cymothoidae), crustacean parasites of marine fishes, with descriptions of eastern Australian species. Records of the Australian Museum 42(3): 247–300. 10.3853/j.0067-1975.42.1990.118 [DOI] [Google Scholar]
- Bruce NL. (1993) Two new genera of marine isopod crustaceans (Flabellifera: Sphaeromatidae) from southern Australia, with a reappraisal of the Sphaeromatidae. Invertebrate Taxonomy 7(1): 151–171. 10.1071/IT9930151 [DOI] [Google Scholar]
- Bruce NL. (1995) The taxonomy and phylogeny of tube-tailed sphaeromatid isopods (Crustacea) with descriptions of new species and a new genus from southern Australia. Ophelia 43(2): 127–180. 10.1080/00785326.1995.10429829 [DOI] [Google Scholar]
- Bruce NL. (2003) New genera and species of sphaeromatid isopod crustaceans from Australian marine coastal waters. Memoirs of the Museum of Victoria 60(2): 309–369. 10.24199/j.mmv.2003.60.28 [DOI] [Google Scholar]
- Bruce NL. (2009a) The Marine Fauna of New Zealand: Isopoda, Aegidae (Crustacea). National Institute of Water and Atmospheric Research Ltd. (NIWA), Wellington. NIWA Biodiversity Memoir 122: 252.
- Bruce NL. (2009b) New genera and species of the marine isopod family Serolidae (Crustacea, Sphaeromatidea) from the southwestern Pacific. ZooKeys 18: 17–76. 10.3897/zookeys.18.96 [DOI] [Google Scholar]
- Bruce NL, Bowman TE. (1982) The status of Cirolanaparva Hansen, 1890 (Crustacea, Isopoda, Cirolanidae) with notes on its distribution. Proceedings of the Biological Society of Washington 95(2): 325–333. [Google Scholar]
- Bruce NL, Jones DA. (1981) The systematics and ecology of some cirolanid isopods from southern Japan. Journal of Natural History 15(1): 67–85. 10.1080/00222938100770061 [DOI] [Google Scholar]
- Bruce NL, Wetzer R. (2004) Paradellatiffany sp. nov., a distinctive sphaeromatid isopod (Crustacea: Isopoda: Sphaeromatidea) from Baja California, Mexico. Zootaxa 623(1): 1–12. 10.11646/zootaxa.623.1.1 [DOI] [Google Scholar]
- Bruce NL, Wetzer R. (2008) New Zealand exports: Pseudosphaeroma Chilton, 1909 (Isopoda: Sphaeromatidae), a southern hemisphere genus introduced to the Pacific coast of North America. Zootaxa 1908(1): 51–56. 10.11646/zootaxa.1908.1.4 [DOI] [Google Scholar]
- Brusca GJ. (1966) Studies on the salinity and humidity tolerances of five species of isopods in a transition from marine to terrestrial life. Bulletin of the Southern California Academy of Sciences 65(3): 146–154. [Google Scholar]
- Brusca RC. (1977) Range extensions and new host records of cymothoid isopods (Isopoda: Cymothoidae) in the Eastern Pacific Ocean. Bulletin of the Southern California Academy of Sciences 76(2): 128–131. [Google Scholar]
- Brusca RC. (1978a) Studies on the cymothoid fish symbionts of the eastern Pacific (Isopoda, Cymothoidae) I. Biology of Nerocilacalifornica. Crustaceana 34(2): 141–154. 10.1163/156854078X00718 [DOI] [Google Scholar]
- Brusca RC. (1978b) Studies on the cymothoid fish symbionts of the eastern Pacific (Crustacea: Cymothoidae) II. Systematics and biology of Lironecavulgaris Stimpson 1857. Occasional Papers of the Allan Hancock Foundation (n. ser. ) 2: 1–19. [Google Scholar]
- Brusca RC. (1980) Common Intertidal Invertebrates of the Gulf of California, 2nd edn. University of Arizona Press, Tucson, 513 pp. [Google Scholar]
- Brusca RC. (1981) A monograph on the IsopodaCymothoidae (Crustacea) of the eastern Pacific. Zoological Journal of the Linnean Society 73(2): 117–199. 10.1111/j.1096-3642.1981.tb01592.x [DOI] [Google Scholar]
- Brusca RC. (1983a) A monograph on the isopod family Aegidae in the tropical eastern Pacific I. The genus Aega. Allan Hancock Monographs in Marine Biology 12: 1–39.
- Brusca RC. (1983b) Two new isopods from Baja California and the Gulf of California (Mexico) and an analysis of the evolutionary history of the genus Colidotea (Crustacea: Isopoda: Idoteidae). Transactions of the San Diego Society of Natural History 20(4): 69–79. 10.5962/bhl.part.28998 [DOI] [Google Scholar]
- Brusca RC. (1984) Phylogeny, evolution and biogeography of the marine isopod subfamily Idoteinae (Crustacea: Isopoda: Idoteidae). Transactions of the San Diego Society of Natural History 20(7): 99–134. 10.5962/bhl.part.29001 [DOI] [Google Scholar]
- Brusca GJ, Brusca RC. (1978) A Naturalist’s Seashore Guide. Common Marine Life of the Northern California Coast and Adjacent Shores. Mad River Press, Eureka, 205 pp. [Google Scholar]
- Brusca RC, France SC. (1992) The genus Rocinela (Crustacea: Isopoda: Aegidae) in the tropical eastern Pacific. Zoological Journal of the Linnean Society 106(3): 231–275. 10.1111/j.1096-3642.1992.tb01248.x [DOI] [Google Scholar]
- Brusca RC, Iverson EW. (1985) A Guide to the Marine Isopod Crustacea of Pacific Costa Rica. Revista de Biología Tropical 33: 1–77. [Google Scholar]
- Brusca RC, Ninos M. (1978) The status of Cirolanacaliforniensis Schultz, and C.diminuta Menzies and George, with a key to the California species of Cirolana (Isopoda: Cirolanidae). Proceedings of the Biological Society of Washington 91(2): 379–385. [Google Scholar]
- Brusca RC, Wallerstein BR. (1977) The marine isopod Crustacea of the Gulf of California. I. Family Idoteidae. American Museum Novitates 2634: 1–17. http://hdl.handle.net/2246/2949 [Google Scholar]
- Brusca RC, Wallerstein BR. (1979a) The marine isopod crustaceans of the Gulf of California II. Idoteidae: New genus and species, range extensions, and comments on evolution and taxonomy within the family. Proceedings of the Biological Society of Washington 92(2): 253–271. [Google Scholar]
- Brusca RC, Wallerstein BR. (1979b) Zoogeographic patterns of idoteid isopods in the northeast Pacific, with a review of shallow water zoogeography of the area. Bulletin of the Biological Society of Washington 3: 67–105. [Google Scholar]
- Brusca RC, Wallerstein BR. (1982) Fish predation: a preliminary study of its role in the zoogeography and evolution of shallow water idoteid isopods (Crustacea: Isopoda: Idoteidae). Journal of Biogeography 9(2): 135–150. 10.2307/2844698 [DOI] [Google Scholar]
- Brusca RC, Wilson GDF. (1991) A phylogenetic analysis of the Isopoda with some classificatory recommendations. Memoirs of the Queensland Museum 31: 143–204. [Google Scholar]
- Brusca RC, Wetzer R, France SC. (1995) Cirolanidae (Crustacea: Isopoda: Flabellifera) of the tropical eastern Pacific. Proceedings of the San Diego Natural History Museum 30: 1–96. [Google Scholar]
- Brusca RC, Kimrey E, Moore W [Eds] (2004) A Seashore Guide to the Northern Gulf of California. Arizona-Sonora Desert Museum, Tucson, 203 pp. [Google Scholar]
- Brusca RC, Coelho VR, Taiti S. (2007) Isopoda. In: Carlton JT. (Ed.) The Light and Smith Manual: Intertidal Invertebrates from Central California to Oregon.4th edn., University of California Press, Berkeley, 503–542.
- Buchholz R. (1866) Ueber Hemioniscus, eine neue Gattung parasitischer Isopoden. Zeitschrift für Wissenschaftliche Zoologie 16(3): 303–327. [Google Scholar]
- Bueno LS, Souza TF, Leite JR, Hostim-Silva M, Olier BS, Sanches EG. (2022) First record of parasitic isopods Nerocilaacuminata (Schiödte & Meinert, 1881) and Nerocilaarmata (Dana, 1853) (Arthropoda, Cymothoidae) in Goliath grouper Epinephelusitajara (Lichtenstein, 1822) in southern Atlantic Ocean. Brazilian Journal of Biology 82(e237668): 1–2. 10.1590/1519-6984.237668 [DOI] [PubMed]
- Bugnot AB, Coleman RA, Figueira WF, Marzinelli EM. (2015) Community-level impacts of the invasive isopod Cirolanaharfordi. Biological Invasions 17(4): 1149–1161. 10.1007/s10530-014-0785-z [DOI] [Google Scholar]
- Bunkley-Williams L, Williams Jr EH, Bashirullah AKM. (2006) Isopods (Isopoda: Aegidae, Cymothoidae, Gnathiidae) associated with Venezuelan marine fishes (Elasmobranchii, Actinopterygii). Revista de Biología Tropical 54(Suppl. 3): 175–188.
- Butler TH. (1964) Redescription of the parasitic isopod Holophryxusalaskensis Richardson, and a note on its synonymy. Journal of the Fisheries Research Board of Canada 21(5): 971–976. 10.1139/f64-090 [DOI] [Google Scholar]
- Cadien D. (1996) SCAMIT voucher sheet for Paramunna sp A (Pleurogoniidae). Specimen examined and vouchered by D Cadien, 30 May 1996. SCAMIT Newsletter 15(2): 2. https://www.scamit.org/newsletters [Google Scholar]
- Cadien D. (2008) SCAMIT voucher sheet for Boreosignum sp IS1 (Isopoda, Family Paramunnidae). Specimens examined and vouchered by DB Cadien, 5 April 2008. https://www.scamit.org/tools
- Cadien D, Brusca RC. (1993) Anthuridean isopods (Crustacea) of California and the temperate northeast Pacific. SCAMIT Newsletter 12(6): 1–26. https://www.scamit.org/newsletters [Google Scholar]
- Calman WT. (1898) On a collection of Crustacea from Puget Sound (Puget Sound Zoology, Columbia University Contributions No. 9). Annals of the New York Academy of Sciences 11(13): 259–292. 10.1111/j.1749-6632.1898.tb54972.x [DOI] [Google Scholar]
- Campos E, de Campos AR. (1990) Taxonomic remarks on Schizobopyrina Markham, 1985, with the description of S.bruscai (Crustacea: Isopoda: Bopyridae). Proceedings of the Biological Society of Washington 103(3): 633–642. [Google Scholar]
- Campos E, de Campos AR. (1998) Taxonomy and distribution of the parasitic isopod Progebiophilusbruscai Salazar-Vallejo and Leija-Tristán, 1990 (Crustacea: Bopyridae). Proceedings of the Biological Society of Washington 111: 288–294. [Google Scholar]
- Campos E, Villarreal G. (2008) Littoral and shallow water isopods from Todos los Santos Bay, Baja California, México. Revista Mexicana de Biodiversidad 79: 347–354. 10.22201/ib.20078706e.2008.002.566 [DOI] [Google Scholar]
- Campos-Filho IS, Cardoso GM, Aguiar JO. (2018) New species and first record of Alloniscus Dana, 1854 (Isopoda: Oniscidae: Alloniscidae) from Brazil. Nauplius 26(0): e2018014. 10.1590/2358-2936e2018014 [DOI]
- Campos-González E, Campoy-Favela JR. (1987) Epicarideans from Baja California 1. First record and bioecological notes on two Bopyridae and one Cryptoniscidae (Crustacea, Isopoda) for Mexico. Ciencias Marinas 13(3): 39–48. 10.7773/cm.v13i3.543 [DOI] [Google Scholar]
- Campos-González E, Ruiz-Campos G, Macías-Chávez JL. (1986) Nuevos huespedes para Lironecamenziesi Brusca, 1981 (Crustacea, isopoda, Cymothoidae) con algunos comentarios sobre preferencia por los mismos. Ciencias Marinas 12(2): 99–104. 10.7773/cm.v12i2.505 [DOI] [Google Scholar]
- Cardoso L, Martins ML, De Aquino Golzio JES, Bomfim CNC, De Oliveira RLM, Dos Santos LBG, Lacerda ACF. (2017) Rocinelasignata (Isopoda: Aegidae) parasitizing the gills of the spotted goatfish Pseudupeneusmaculatus (Actinopterygii: Mullidae) in Northeastern Brazil. Anais da Academia Brasileira de Ciências 89(3, Supplement): 2075–2080. 10.1590/0001-3765201720150677 [DOI] [PubMed]
- Carlton JT. (1979) Introduced invertebrates of San Francisco Bay. In: Conomos TJ. (Ed.) San Francisco Bay: the Urbanized Estuary.California Academy of Sciences, San Francisco, 427–444.
- Carlton JT, Iverson EW. (1981) Biogeography and natural history of Sphaeromawalkeri Stebbing (Crustacea: Isopoda) and its introduction to San Diego Bay, California. Journal of Natural History 15(1): 31–48. 10.1080/00222938100770031 [DOI] [Google Scholar]
- Carvacho A. (1983) Asellota del golfo de California, con descripción de dos nuevos géneros y dos nuevas especies (Crustacea, Isopoda). Cahiers de Biologie Marine 24: 281–295. [Google Scholar]
- Castellanos C, Junoy J. (2005) Synisomaalbertoi, a new species from the Strait of Gibraltar (southern Spain) with a key to known species of the genus (Crustacea: Isopoda: Idoteidae). Journal of the Marine Biological Association of the United Kingdom 85(6): 1461–1466. 10.1017/S0025315405012646 [DOI] [Google Scholar]
- Cavalcanti E, Do Nascimento S, Barros N, Chellappa S. (2012) Occurrence of the isopod parasite Rocinelasignata (Isopoda: Aegidae) on marine fish Sparisomafrondosum (Osteichthyes: Scaridae). Marine Biodiversity Records 5: e66. 10.1017/S1755267212000516 [DOI]
- Chapman JW, Dumbauld BR, Itani G, Markham JC. (2012) An introduced Asian parasite threatens northeastern Pacific estuarine ecosystems. Biological Invasions 14(6): 1221–1236. 10.1007/s10530-011-0151-3 [DOI] [Google Scholar]
- Chess JR, Hobson ES. (1997) Benthic invertebrates of four southern California marine habitats prior to onset of ocean warming in 1976, with lists of fish predators. NOAA Technical Memorandum NMFS-SWFSC-243, 110 pp. https://repository.library.noaa.gov/view/noaa/3013
- Chilton C. (1892) Notes on some New Zealand Amphipoda and Isopoda. Transactions of the New Zealand Institute 24: 258–269. [Google Scholar]
- City of Los Angeles (2021) Marine Monitoring in Santa Monica Bay Biennial Assessment Report 2019-2020. Department of Public Works, Environmental Monitoring Division, LA Sanitation & Environment, Hyperion Water Reclamation Plant, Playa del Rey, 226 pp. [+ appendices] https://ww.lacitysan.org/san/sandocview?docname=cnt068942 [Google Scholar]
- City of San Diego (2018) Biennial Receiving Waters Monitoring and Assessment Report for the Point Loma and South Bay Ocean Outfalls, 2016–2017. City of San Diego, Public Utilities Department, Environmental Monitoring and Technical Services Division, San Diego, California, 210 pp. https://www.sandiego.gov/sites/default/files/2016_2017_biennial_rpt.pdf [Google Scholar]
- Cohen BF. (1998) Dendrotiidae (Crustacea: Isopoda) of the southeastern Australian continental slope. Memoirs of the Museum of Victoria 57(1): 1–38. 10.24199/j.mmv.1998.57.01 [DOI] [Google Scholar]
- Cohen AN, Carlton JT. (1995) Biological Study. Nonindigenous aquatic species in a United States estuary: A case study of the biological invasions of the San Francisco Bay and Delta. U.S. Fish and Wildlife Service, Washington, DC, 246 pp. [Google Scholar]
- Cohen BF, Poore GCB. (1994) Phylogeny and biogeography of the Gnathiidae (Crustacea, Isopoda) with descriptions of new genera and species, most from south-eastern Australia. Memoirs of the Museum of Victoria 54(2): 271–397. 10.24199/j.mmv.1994.54.13 [DOI] [Google Scholar]
- Cohen AN, Harris LH, Bingham BL, Carlton JT, Chapman JW, Lambert CC, Lambert G, Ljubenkov JC, Murray SN, Rao LC, Reardon K, Schwindt E. (2005) Rapid Assessment Survey for exotic organisms in southern California bays and harbors, and abundance in port and non-port areas. Biological Invasions 7(6): 995–1002. 10.1007/s10530-004-3121-1 [DOI] [Google Scholar]
- Cooke WJ. (1977) On the occurrence of the commensal asellote Caecijaerahorvathi Menzies, 1951, in Hawaii. Crustaceana 33(1): 105–106. 10.1163/156854077X00322 [DOI] [Google Scholar]
- Cookson LJ. (1991) Australasian species of Limnoriidae (Crustacea: Isopoda). Memoirs of the Museum of Victoria 52(2): 137–262. 10.24199/j.mmv.1991.52.02 [DOI] [Google Scholar]
- Cookson LJ. (1997) Additions to the taxonomy of the Limnoriidae. Memoirs of the Museum of Victoria 56(1): 129–143. 10.24199/j.mmv.1997.56.07 [DOI] [Google Scholar]
- Coyle KO, Mueller GJ. (1981) Larval and juvenile stages of the isopod Holophryxusalaskensis (Epicarida, Dajidae) parasitic on decapods. Canadian Journal of Fisheries and Aquatic Sciences 38(11): 1438–1443. 10.1139/f81-190 [DOI] [Google Scholar]
- Crisp DJ. (1968) Distribution of the parasitic isopod Hemioniscusbalani with special reference to the east coast of North America. Journal of the Fisheries Research Board of Canada 25(6): 1161–1167. 10.1139/f68-102 [DOI] [Google Scholar]
- Cruz-García R, Cupul-Magana AL, Hendrickx ME, Rodríguez-Troncoso AP. (2013) Abundance of three species of Isopoda (Peracarida, Isopoda) associated with a coral reef environment in Pacific Mexico. Crustaceana 86(13–14): 1664–1674. 10.1163/15685403-00003264 [DOI] [Google Scholar]
- Cunha MR, Wilson GDF. (2003) Haplomunnidae (Crustacea: Isopoda) reviewed, with a description of an intact specimen of Thylakogaster Wilson & Hessler, 1974. Zootaxa 326(1): 1–16. 10.11646/zootaxa.326.1.1 [DOI] [Google Scholar]
- Dana JD. (1852) On the classification of the CrustaceaChoristopoda or Tetradecapoda. The American Journal of Science and Arts. Second Series 14(41): 297–316. [Google Scholar]
- Dana JD. (1853) United States Exploring Expedition during the years 1838, 1839, 1840, 1841, 1842, under the command of Charles Wilkes, U.S.N. Vol. XIII. Crustacea. Part II: 692–1618. [dated 1852, but published 1853]
- Dana JD. (1854) Catalogue and descriptions of Crustacea collected in California by Dr. John L. Leconte. Proceedings of the Philadelphia Academy of Natural Sciences 7: 175–177. [Google Scholar]
- Davidson TM, de Rivera CE. (2010) Accelerated erosion of saltmarshes infested by the non-native burrowing crustacean Sphaeromaquoianum. Marine Ecology Progress Series 419: 129–136. 10.3354/meps08836 [DOI] [Google Scholar]
- Davidson TM, Shanks AL, Rumrill SS. (2010) The composition and density of fauna utilizing burrow microhabitats created by a non-native burrowing crustacean (Sphaeromaquoianum). Biological Invasions 12(5): 1403–1413. 10.1007/s10530-009-9556-7 [DOI] [Google Scholar]
- Delaney PM. (1982) The synonymy of Excorallanakathyae Menzies, 1962, with Excorallanatruncata (Richardson, 1899), with a redescription of the species (Crustacea, Isopoda, Excorallanidae). Journal of Crustacean Biology 2(2): 273–280. 10.2307/1548008 [DOI] [Google Scholar]
- Delaney PM. (1984) Isopods of the genus Excorallana Stebbing, 1904 from the Gulf of California, Mexico (Crustacea, Isopoda, Corallanidae). Bulletin of Marine Science 34: 1–20. [Google Scholar]
- Delaney PM. (1989) Phylogeny and biogeography of the marine isopod family Corallanidae (Crustacea, Isopoda, Flabellifera). Natural History Museum of Los Angeles County. Contributions in Science 409: 1–75. 10.5962/p.226815 [DOI] [Google Scholar]
- Delaney PM. (1993) X-ray microanalysis of the cuticle surface of two southern California marine isopod species. Bulletin of the Southern California Academy of Sciences 92: 64–69. [Google Scholar]
- Delaney PM, Brusca RC. (1985) Two new species of Tridentella Richardson, 1905 (Isopoda: Flabellifera: Tridentellidae) from California, with a rediagnosis and comments on the family, and a key to the genera of Tridentellidae and Corallanidae. Journal of Crustacean Biology 5(4): 728–742. 10.2307/1548249 [DOI] [Google Scholar]
- Dexter DM. (1976) The sandy-beach fauna of Mexico. The Southwestern Naturalist 20(4): 479–485. 10.2307/3669864 [DOI] [Google Scholar]
- Dexter DM. (1983) Soft bottom infaunal communities in Mission Bay. California Fish and Game 69(1): 5–17. [Google Scholar]
- Dollfus A. (1895) Voyage de M. E. Simon dans l’Afrique australe (Janvier-Avril 1893). Crustaces isopodes terrestres. Mémoires de la Société Zoologique de France 8: 345–352. [Google Scholar]
- Dollfus A. (1901) Étude préliminaire des Gnathiidae recueillis dans les campagnes de l’Hirondelle et de La Princesse-Alice. Bulletin de la Société Zoologique de France 26: 239–246. [Google Scholar]
- Doti BL, Wilson GDF. (2010) The genera Carpias Richardson, Ianiropsis Sars and Janaira Moreira & Pires (Isopoda: Asellota: Janiridae) from Australia, with description of three new species. Zootaxa 2625(1): 1–39. 10.11646/zootaxa.2625.1.1 [DOI] [Google Scholar]
- Dow TG. (1958) Description of a new isopod from California, Exosphaeromainornata. Bulletin of the Southern California Academy of Sciences 57(2): 93–97. [Google Scholar]
- Dumbauld BR, Chapman JW, Torchin ME, Kuris AM. (2011) Is the collapse of mud shrimp (Upogebiapugettensis) populations along the pacific coast of North America caused by outbreaks of a previously unknown bopyrid isopod parasite (Orthionegriffenis)? Estuaries and Coasts 34(2): 336–350. 10.1007/s12237-010-9316-z [DOI]
- Eberl R. (2012) Distribution, habitat and food preferences of sympatric high intertidal isopod species Ligiaoccidentalis and Ligiapallasii (Ligiidae: Oniscidea). Journal of Natural History 46(29–30): 1779–1797. 10.1080/00222933.2012.700334 [DOI] [Google Scholar]
- Eberl R, Mateos M, Grosberg RK, Santamaria CA, Hurtado LA. (2013) Phylogeography of the supralittoral isopod Ligiaoccidentalis around the Point Conception marine biogeographical boundary. Journal of Biogeography 40(12): 2361–2372. 10.1111/jbi.12168 [DOI] [Google Scholar]
- Eman MY, Nahla HS, Eissa IAM, Mona SZ. (2014) Parasitological studies on the Isopoda (Cymothoidae) parasites infesting some marine fishes at Suez Canal area at Ismailia Province, Egypt with a key to the cymothoid genera. Life Science Journal 11(1): 227–231. [Google Scholar]
- Espinosa-Pérez MC, Hendrickx ME. (2001a) Checklist of isopods (Crustacea: Peracarid: Isopoda) from the eastern tropical Pacific. Belgian Journal of Zoology 131(1): 43–55. [Google Scholar]
- Espinosa-Pérez MC, Hendrickx ME. (2001b) The genus Paracerceis Hansen, 1905 (Isopoda, Sphaeromatidae) in the eastern tropical Pacific, with the description of a new species. Crustaceana 74(11): 1169–1187. 10.1163/15685400152885165 [DOI] [Google Scholar]
- Espinosa-Pérez MC, Hendrickx ME. (2002) Distribution and ecology of isopods (Crustacea: Peracarida: Isopoda) of the Pacific coast of Mexico. In: Escobar-Briones E, Alvarez F. (Eds) Modern approaches to the study of Crustacea.Springer, Boston, 95–103. 10.1007/978-1-4615-0761-1_16 [DOI]
- Espinosa-Pérez MC, Hendrickx ME. (2006) A comparative analysis of biodiversity and distribution of shallow-water marine isopods (Crustacea: Isopoda) from polar and temperate waters in the East Pacific. Belgian Journal of Zoology 136(2): 219–247. [Google Scholar]
- Espinosa-Pérez MC, Hendrickx ME, Morrone JJ. (2009) Identification of generalized tracks for the species of Isopoda (Peracarida) from the Eastern Pacific. Journal of Crustacean Biology 29(2): 224–231. 10.1651/08-3074R.1 [DOI] [Google Scholar]
- Esquete P, Wilson GDF. (2016) The genus Uromunna (Crustacea: Isopoda: Munnidae) in New South Wales, with a key for all known species. Marine Biology Research 2016(online): 1–25. 10.1080/17451000.2015.1131298 [DOI]
- Esteves-Silva PH, Oliveira MSB, Gentil-Vasconcelos HC, Costa-Campos CE, Tavares-Dias M. (2020) New records of hosts for Excorallanalongicornis and Nerocilaacuminata (Crustacea: Isopoda) in brackish fish from the coast of the State of Amapá (Brazil), with an update on the geographic distribution of Nerocilaacuminata. Journal of Parasitic Diseases 44(2): 420–428. 10.1007/s12639-020-01192-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fairey R, Dunn R, Roberts C, Sigala M, Oliver J. (2002) Introduced Aquatic Species in California Coastal Waters. Final Report (July 2002). California Department of Fish and Game. Sacramento, 116 pp.
- Fee AR. (1927) The Isopoda of Departure Bay and vicinity, with descriptions of new species, variations, and colour notes. Contributions to Canadian Biology and Fisheries 3(1): 15–46. [Published 13 Jan 1927] 10.1139/f26-002 [DOI] [Google Scholar]
- Fogg AQ, Ruiz CF, Curran SS, Bullard SA. (2016) Parasites from the red lionfish, Pteroisvolitans from the Gulf of Mexico. Gulf and Caribbean Research 27(1): SC1–SC5. 10.18785/gcr.2701.07 [DOI]
- Franke HD, Gutow L, Janke M. (1998) The recent arrival of the oceanic isopod Idoteametallica Bosc off Helgoland (German Bight, North Sea): An indication of a warming trend in the North Sea? Helgoländer Meeresuntersuchungen 52(3–4): 347–357. 10.1007/BF02908908 [DOI]
- Galassi DMP, Bruce NL, Fiasca B, Dole-Oliver M-J. (2016) A new family Lepidocharontidae with description of Lepidocharon gen. n., from the Great Barrier Reef, Australia, and redefinition of the Microparasellidae (Isopoda, Asellota). ZooKeys 594: 11–50. 10.3897/zookeys.594.7539 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galil BS. (2008) Sphaeromawalkeri Stebbing, 1905 (Crustacea: Isopoda: Sphaeromatidae) established on the Mediterranean coast of Israel. Aquatic Invasions 3(4): 443–444. 10.3391/ai.2008.3.4.13 [DOI] [Google Scholar]
- García-Madrigal MS, Pérez-Enriquez VI, Jarquín-Martínez U. (2022) Crustáceos (Arthropoda: Crustacea). In: Bastida-Zavala JR, García-Madrigal MS. (Eds) Invertebrados Marinos y Costeros del Pacífico sur de México.Universidad del Mar y Geomare, Puerto Ángel, Oaxaca, México, 199–275.
- Garthwaite RL, Lawson R. (1992) Oniscidea (Isopoda) of the San Francisco Bay area. Proceedings of the California Academy of Sciences 47(11): 303–328. [Google Scholar]
- Garthwaite RL, Hochberg FG, Sassaman C. (1985) The occurrence and distribution of terrestrial isopods (Oniscoidea) on Santa Cruz Island with preliminary data for the other California islands. Bulletin of the Southern California Academy of Sciences 84: 23–37. [Google Scholar]
- Garthwaite RL, Lawson R, Taiti S. (1992) Morphological and genetic relationships among four species of Armadilloniscus Uljanin, 1875 (Isopoda: Oniscidea: Scyphacidae). Journal of Natural History 26(2): 327–338. 10.1080/00222939200770171 [DOI] [Google Scholar]
- Garzón-Ferreira J. (1990) An isopod, Rocinelasignata (Crustacea: Isopoda: Aegidae), that attacks humans. Bulletin of Marine Science 46(3): 813–815. [Google Scholar]
- George RY, Strömberg JO. (1968) Some new species and new records of marine isopods from San Juan Archipelago, Washington, U.S.A. Crustaceana 14: 225–254. 10.1163/156854068X00827 [DOI] [Google Scholar]
- Gillard RM [Ed.] (1978) POCS Reference Paper No. III. Benthic organisms and estuarine communities of the Southern California Bight. U.S. Department of the Interior, Bureau of Land Management, Pacific Outer Continental Shelf Office. Los Angeles, 221 pp. [Google Scholar]
- Gillett DJ, Lovell LL, Schiff KC. (2017) Southern California Bight 2013 Regional Monitoring Program: Vol. VI. Benthic Infauna. Technical Report 971. Southern California Coastal Water Research Project, Costa Mesa, 300 pp. https://www.sccwrp.org/publications [Google Scholar]
- Gillett DJ, Gilbane L, Schiff KC. (2019) Benthic Infauna of the Southern California Bight Continental Slope: Characterizing Community Structure for the Development of an Index of Disturbance. U.S. Department of the Interior, Bureau of Ocean Energy Management, Camarillo. OCS Study BOEM 2019-050, 157 pp. [Google Scholar]
- Gillett DJ, Enright W, Walker JB. (2022) Southern California Bight 2018 Regional Monitoring Program: Vol. III. Benthic Infauna. Technical Report 1289. Southern California Coastal Water Research Project. Costa Mesa, 61 pp. [+ appendices] https://www.sccwrp.org/publications
- Glynn PW. (1976) A new shallow-water serolid (Isopoda: Flabellifera) from the Pacific coast of Panamá. Journal of Natural History 10(1): 7–16. 10.1080/00222937600770021 [DOI] [Google Scholar]
- Glynn PW, Glynn CS. (1974) On the systematics of Ancinus (Isopoda, Sphaeromatidae), with the description of a new species from the tropical eastern Pacific. Pacific Science 28(4): 401–422. [Google Scholar]
- Golovan OA, Malyutina MV. (2022) The first record of the family Paramunnidae (Isopoda: Asellota) from the bathyal of the Bering Sea with descriptions of two new species of Munnogonium. Deep-sea Research. Part II, Topical Studies in Oceanography 200: 105095. 10.1016/j.dsr2.2022.105095 [DOI]
- Golovan OA, Malyutina MV, Brandt A. (2018) First record of the deep-sea isopod family Dendrotionidae (Isopoda: Asellota) from the Northwest Pacific with description of two new species of Dendromunna. Marine Biodiversity 48(1): 531–544. 10.1007/s12526-017-0725-0 [DOI]
- Gurjanova EF. (1933) Contribution to the Isopoda-Fauna of the Pacific Ocean. II. New species of Gnathiidea and Asellota. Issledovaniia Fauny Morei: Akademiia nauk SSSR 19: 79–91. [Google Scholar]
- Guzman HM, Obando VL, Brusca RC, Delaney PM. (1988) Aspects of the population biology of the marine isopod Excorallanatricornisoccidentalis Richardson, 1905 (Crustacea: Isopoda: Corallanidae) at Cano Island, Pacific Costa Rica. Bulletin of Marine Science 43(1): 77–87. [Google Scholar]
- Hadfield KA, Smit NJ. (2020) Review of the global distribution and hosts of the economically important fish parasitic isopod genus Ceratothoa (Isopoda: Cymothoidae), including the description of Ceratothoa springbok n. sp. from South Africa. International Journal for Parasitology 50(10–11): 899–919. 10.1016/j.ijpara.2020.07.001 [DOI] [PubMed] [Google Scholar]
- Hadfield KA, Bruce NL, Smit NJ. (2016) Redescription of poorly known species of Ceratothoa Dana, 1852 (Crustacea, Isopoda, Cymothoidae) based on original type material. ZooKeys 592: 39–91. 10.3897/zookeys.592.8098 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haney L. (2006) Pairing Northeast Pacific gnathiid isopods. Which females go with what males? SCAMIT Newsletter, Supplement to Vol. 24(6): 37. https://www.scamit.org/newsletters [Google Scholar]
- Hansen HJ. (1897) Reports on the dredging operations off the west coast of Central America to the Galapagos Islands, to the west coast of Mexico, and in the Gulf of California, in charge of Alexander Agassiz, carried on by the U.S. Fish Commission Steamer Albatross during 1891, Lieut. Commander Z.L. Tanner, U.S. Navy, commanding. XXII. The Isopods. Bulletin of the Museum of Comparative Zoology at Harvard College 31(5): 95–129. [Google Scholar]
- Hardisty AR, Ellwood ER, Nelson G, Zimkus B, Buschbom J, Addink W, Rabeler RK, Bates J, Bentley A, Fortes JAB, Hansen S, Macklin JA, Mast AR, Miller JT, Monfils AK, Paul DL, Wallis E, Webster M. (2022) Digital extended specimens: Enabling an extensible network of biodiversity data records as integrated digital objects on the Internet. Bioscience 72(10): 978–987. 10.1093/biosci/biac060 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison K. (1984) Some sphaeromatid isopods (Crustacea) from southern and south-western Australia, with the description of a new genus and two new species. Records of the Western Australian Museum 11(3): 259–286. [Google Scholar]
- Harrison K, Ellis JP. (1991) The genera of the Sphaeromatidae (Crustacea: Isopoda): a key and distribution list. Invertebrate Taxonomy 5(4): 915–952. 10.1071/IT9910915 [DOI] [Google Scholar]
- Harrison K, Holdich DM. (1982) Revision of the genera Dynamenella, Ischyromene, Dynamenopsis, and Cymodocella (Crustacea: Isopoda), including a new genus and five new species of eubranchiate sphaeromatids from Queensland waters. Journal of Crustacean Biology 1(1): 84–119. 10.2307/1548115 [DOI] [Google Scholar]
- Harrison K, Holdich DM. (1984) Hemibranchiate sphaeromatids (Crustacea: Isopoda) from Queensland, Australia, with a world-wide review of the genera discussed. Zoological Journal of the Linnean Society 81(4): 275–387. 10.1111/j.1096-3642.1984.tb01175.x [DOI] [Google Scholar]
- Harrison K, Poore GCB. (1984) Serolis (Crustacea, Isopoda, Serolidae) from Australia, with a new species from Victoria. Memoirs of the Museum of Victoria 45(1): 13–31. 10.24199/j.mmv.1984.45.03 [DOI] [Google Scholar]
- Harty R. (1979) Range extension and notes on the habitat of the isopod Munnahalei Menzies. Bulletin of the Southern California Academy of Sciences 78(3): 196–199. [Google Scholar]
- Hata H, Sogabe A, Tada S, Nishimoto R, Nakano R, Kohya N, Takeshima H, Kawanishi R. (2017) Molecular phylogeny of obligate fish parasites of the family Cymothoidae (Isopoda, Crustacea): Evolution of the attachment mode to host fish and the habitat shift from saline water to freshwater. Marine Biology 164(5): 1–15. 10.1007/s00227-017-3138-527980349 [DOI] [Google Scholar]
- Hatch MH. (1947) The Chelifera and Isopoda of Washington and adjacent regions. University of Washington Publications in Biology 10(5): 155–274. [Google Scholar]
- Healey D, Hovel KA. (2004) Seagrass bed patchiness: Effects on epifaunal communities in San Diego Bay, USA. Journal of Experimental Marine Biology and Ecology 313(1): 155–174. 10.1016/j.jembe.2004.08.002 [DOI] [Google Scholar]
- Heberling JM, Miller JT, Noesgaard D, Weingart SB, Schigel D. (2021) Data integration enables global biodiversity synthesis. Proceedings of the National Academy of Sciences of the United States of America 118(6): e2018093118. 10.1073/pnas.2018093118 [DOI] [PMC free article] [PubMed]
- Hedo G, Junoy J. (1999) A new species of Synisoma (Isopoda: Valvifera: Idoteidae) from the Strait of Gibraltar and the Alborán Sea (Spain, western Mediterranean). Cahiers de Biologie Marine 40: 87–92. [Google Scholar]
- Hedrick BP, Heberling JM, Meineke EK, Turner KG, Grassa CJ, Park DS, Kennedy J, Clarke JA, Cook JA, Blackburn DC, Edwards SV, Davis CC. (2020) Digitization and the future of natural history collections. Bioscience 70(3): 243–251. 10.1093/biosci/biz163 [DOI] [Google Scholar]
- Hendrickx ME. (2008) Rediscovery of Rocinelamurilloi Brusca & Iverson, 1985 (Isopoda, Aegidae) in the Gulf of California, Mexico, and ecological data associated with its capture. Crustaceana 81(10): 1259–1262. 10.1163/156854008X374586 [DOI] [Google Scholar]
- Hendrickx ME. (2018a) Additional distribution and ecology records of the deep-water isopod Rocinelamurilloi Brusca & Iverson, 1985 (Isopoda, Aegidae) in western Mexico. Nauplius 26: e2018036. 10.1590/2358-2936e2018036 [DOI]
- Hendrickx ME. (2018b) Notes on two shallow water isopods (Crustacea: Peracarida: Isopoda: Idoteidae) from Kino Bay, Gulf of California, Mexico. Nauplius 26: e2018027. 10.1590/2358-2936e2018027 [DOI]
- Hendrickx ME, Espinosa-Pérez MC, Salgado-Barragan J, Mendez-Ubach MN. (1997) Isópodos (Crustacea: Isopoda). In: Estación Mazatlán U, ICML (Eds) Tercer Católogo de la Colección de Referencia de Invertebrados.Estación Mazatlón, ICML, UNAM México: Comisión Nacional para el Conocimiento y Uso de la Biodiversidadd e Instituto de Ciecias del Mar y Limnología, UNAM, México, 27–49.
- Henkel SK, Gilbane L, Phillips AJ, Gillett DJ. (2020) Cross-shelf habitat suitability modeling for benthic macrofauna. Camarillo (CA): US Department of the Interior, Bureau of Ocean Energy Management. OCS Study BOEM 2020-008, 71 pp. [Google Scholar]
- Hessler RR. (1972) The relationship between Seroliscarinata Lockington and Serolismgrayi Menzies and Frankenberg (Isopoda, Flabellifera). Crustaceana (Supplement 3): 1–6. https://www.jstor.org/stable/25027403
- Hessler RR, Thistle D. (1975) On the place of origin of the deep-sea isopods. Marine Biology 32(2): 155–165. 10.1007/BF00388508 [DOI] [Google Scholar]
- Hessler RR, Wilson GD, Thistle D. (1979) The deep-sea isopods: A biogeographic and phylogenetic overview. Sarsia 64(1–2): 67–75. 10.1080/00364827.1979.10411365 [DOI] [Google Scholar]
- Hewatt WG. (1946) Marine ecological studies on Santa Cruz Island, California. Ecological Monographs 16(3): 185–210. 10.2307/1948649 [DOI] [Google Scholar]
- Hewitt C, Campbell M. (2001) The Australian distribution of the introduced sphaeromatid isopod, Paracerceissculpta. Crustaceana 74(9): 925–936. 10.1163/15685400152682674 [DOI]
- Higley DL, Holton RL. (1978) A grab-sample study of the benthic invertebrates of the Columbia River Estuary. Supplemental Data Report 1 November 1975 through 29 February 1976 [Supplement to Reference 76-3]. Submitted to Port of Astoria, Astoria, Oregon, 27 pp. [Google Scholar]
- Hinton S. (1987) Seashore Life of Southern California. Revised edn. University of California Press, Berkeley, Los Angeles, London, 217 pp. [Google Scholar]
- Hinton SA, Emmett RL. (1994) Benthic infaunal, sediment, and fish offshore from the Columbia River, July 1992. Report to the U.S. Army Corps of Engineers, Contract E96920040, 60 pp. [+ appendices] [Available from Northwest Fisheries Science Center, 2725 Montlake Boulevard East, Seattle, WA 98112] [Google Scholar]
- Hinton SA, Emmett RL. (1996) Benthic infauna and sediment characteristics offshore from the Columbia River, August 1994. Report to the U.S. Army Corps of Engineers, Contract E96930048, 30 pp. [+ appendices] [Available from Northwest Fisheries Science Center, 2725 Montlake Boulevard East, Seattle, WA 98112] [Google Scholar]
- Hobson ES, Chess JR. (1986) Relationships among fishes and their prey in a nearshore sand community off southern California. Environmental Biology of Fishes 17(3): 201–226. 10.1007/BF00002096 [DOI] [Google Scholar]
- Holmes S. (1904) Remarks on the sexes of sphaeromids, with a description of a new species of Dynamene. Proceedings of the California Academy of Sciences, Third Series Zoology 3(11): 295–306. [Google Scholar]
- Holmes S, Gay ME. (1909) Four new species of isopods from the coast of California. Proceedings of the United States National Museum 36(1670): 375–379. 10.5479/si.00963801.36-1670.375 [DOI] [Google Scholar]
- Holthuis LB. (1949) The Isopoda and Tanaidacea of the Netherlands, including the description of a new species of Limnoria. Zoologische Mededelingen Rijksmuseum Van Natuurlijke Historie te Leiden 30: 163–190. [Google Scholar]
- Hong J-S, Lee C-L, Min G-S. (2015) Occurrence of Orthionegriffenis Markham, 2004 (Isopoda: Bopyridae), parasite of the mud shrimp Upogebiamajor (De Haan, 1841) in South Korean Waters, and its implications with respect to marine bioinvasion. Journal of Crustacean Biology 35(5): 605–615. 10.1163/1937240X-00002361 [DOI] [Google Scholar]
- Hubbard DM, Dugan JE, Schooler NK, Viola SM. (2014) Local extirpations and regional declines of endemic upper beach invertebrates in southern California. Estuarine, Coastal and Shelf Science 150: 67–75. 10.1016/j.ecss.2013.06.017 [DOI] [Google Scholar]
- Hunt SL, Mulligan TJ, Komori K. (1999) Oceanic feeding habits of chinook salmon, Oncorhynchustshawytscha, off northern California. Fish Bulletin 97(3): 717–721. [Google Scholar]
- Hurley DE, Jansen KP. (1977) The marine fauna of New Zealand: Family Sphaeromatidae (Crustacea: Isopoda: Flabellifera). New Zealand Oceanographic Institute Memoir 63: 1–95. [Google Scholar]
- Hurtado LA, Mateos M, Santamaria CA. (2010) Phylogeography of supralittoral rocky intertidal Ligia isopods in the Pacific Region from Central California to Central Mexico. PLoS ONE 5(7): e11633. 10.1371/journal.pone.0011633 [DOI] [PMC free article] [PubMed]
- Hwang H, Kang J, Cho IY, Kang DW, Paek WK, Lee SH. (2014) Benthic invertebrate fauna in the islets of Namuseom and Bukhyeongjeseom off Busan. Journal of Asia-Pacific Biodiversity 7(2): e206–e212. 10.1016/j.japb.2014.03.009 [DOI]
- Iverson EW. (1972) A new subtidal Synidotea from central California (Crustacea: Isopoda). Proceedings of the Biological Society of Washington 85(47): 541–548. [Google Scholar]
- Iverson EW. (1974) Range extensions for some California marine isopod crustaceans. Bulletin of the Southern California Academy of Sciences 73(3): 164–169. [Google Scholar]
- Iverson EW. (1978) Status of Exosphaeromainornata Dow and E.media George and Stromberg (Isopoda: Sphaeromatidae) with ecological notes. Journal of the Fisheries Research Board of Canada 35(10): 1381–1384. 10.1139/f78-218 [DOI] [Google Scholar]
- Iverson EW. (1982) Revision of the isopod family Sphaeromatidae (Crustacea: Isopoda: Flabellifera) I. Subfamily names with diagnoses and key. Journal of Crustacean Biology 2(2): 248–254. 10.2307/1548005 [DOI] [Google Scholar]
- Iverson EW, Chivers DD. (1984) A new genus and species of Sphaeromatidae (Crustacea: Isopoda: Flabellifera) from the Miocene of Kern County, California. Journal of Paleontology 58(1): 104–108. http://www.jstor.org/stable/1304737 [Google Scholar]
- Iverson EW, Wilson GDF. (1981) Paramunnaquadratifrons, new species, the first record of the genus in the North Pacific Ocean (Crustacea: Isopoda: Pleurogoniidae). Proceedings of the Biological Society of Washington 85(47): 541–548. [Google Scholar]
- Iwasa M. (1934) Description of a new isopod crustacean (Sphaeromidae) from Hokkaido. Proceedings of the Imperial Academy 10(6): 370–373. 10.2183/pjab1912.10.370 [DOI] [Google Scholar]
- Jang IK, Kwon DH. (1990) Ianiropsis (Isopoda, Asellota, Ianiridae) from Korea, with description of a new species. Korean Journal of Systematic Zoology 6(2): 193–208. [Google Scholar]
- Jass J, Klausmeier B. (2000) Endemics and immigrants: North American terrestrial isopods (Isopoda, Oniscidea) north of Mexico. Crustaceana 73(7): 771–799. 10.1163/156854000504804 [DOI] [Google Scholar]
- Jass J, Klausmeier B. (2001) Terrestrial isopod (Crustacea: Isopoda) atlas for Canada, Alaska, and the contiguous United States. Milwaukee Public Museum Contributions in Biology and Geology 95: 1–105. [Google Scholar]
- Jay CV. (1989) Prevalence, size and fecundity of the parasitic isopod Argeiapugettensis on its host shrimp Crangonfrancisorum. American Midland Naturalist 121(1): 68–77. 10.2307/2425657 [DOI] [Google Scholar]
- Johnson WS. (1976a) Biology and population dynamics of the intertidal isopod Cirolanaharfordi. Marine Biology 36(4): 343–350. 10.1007/BF00389196 [DOI] [Google Scholar]
- Johnson WS. (1976b) Population energetics of the intertidal isopod Cirolanaharfordi. Marine Biology 36(4): 351–357. 10.1007/BF00389197 [DOI] [Google Scholar]
- Johnson ME, Snook HJ. (1967) Seashore Animals of the Pacific Coast. (Reprint of 1927 edn.). Dover Publications, New York, 659 pp. [Google Scholar]
- Jorgensen P, Ibarra-Obando SE, Carriquiry JD. (2007) Top-down and bottom-up stabilizing mechanisms in eelgrass meadows differentially affected by coastal upwelling. Marine Ecology Progress Series 333: 81–93. 10.3354/meps333081 [DOI] [Google Scholar]
- Just J, Wilson GDF. (2004) Revision of the Paramunna complex (Isopoda: Asellota: Paramunnidae). Invertebrate Systematics 18(4): 377–466. 10.1071/IS03027 [DOI] [Google Scholar]
- Just J, Wilson GDF. (2007) Revision of Austrosignum Hodgson and Munnogonium George & Strömberg (Paramunnidae) with descriptions of eight new genera and two new species (Crustacea: Isopoda: Asellota). Zootaxa 1515(1): 1–29. 10.11646/zootaxa.1515.1.1 [DOI] [Google Scholar]
- Just J, Wilson GDF. (2021) Redescriptions and new species in the ‘Austrosignum–Munnogonium’ complex sensu Just and Wilson (2007), mainly from the Southern Hemisphere (Crustacea: Isopoda: Paramunnidae). Zootaxa 4952(3): 401–447. 10.11646/zootaxa.4952.3.1 [DOI] [PubMed] [Google Scholar]
- Kalman JE. (2006) Ectoparasites of demersal marine fishes in Santa Monica Bay, California, U.S.A., with 31 new host records and three range extensions. Comparative Parasitology 73(2): 201–213. 10.1654/4190.1 [DOI] [Google Scholar]
- Kaneko K, Omori M. (2005) Life history and utilization of the beach face slope by the sandy beach isopod Excirolanachiltoni in Sendai Bay, northern Japan. Crustacean Research 34: 27–39. 10.18353/crustacea.34.0_27 [DOI] [Google Scholar]
- Keable SJ. (2006) Taxonomic revision of Natatolana (Crustacea: Isopoda: Cirolanidae). Records of the Australian Museum 58(2): 133–244. 10.3853/j.0067-1975.58.2006.1469 [DOI] [Google Scholar]
- Kensley B. (1978) Five new genera of anthurid isopod crustaceans. Proceedings of the Biological Society of Washington 91(3): 775–792. [Google Scholar]
- Kensley B. (1984) The Atlantic Barrier Reef Ecosystem at Carrie Bow Cay, Belize, III: New marine Isopoda. Smithsonian Contributions to Marine Sciences 24, [iv +] 81 pp. 10.5479/si.01960768.24.1 [DOI]
- Kensley B, Clark K. (1998) A new isopod species from Key Largo, Florida (Crustacea: Isopoda: Holognathidae). Proceedings of the Biological Society of Washington 111(2): 314–319. [Google Scholar]
- Kensley B, Heard RW. (1997) Tridentellaornata (Richardson 1911), new combination: records of hosts and localities (Crustacea: Isopoda: Tridentellidae). Proceedings of the Biological Society of Washington 110(3): 422–425. [Google Scholar]
- Kensley B, Kaufman HW. (1978) Cleantioides, a new idoteid isopod genus from Baja California and Panama. Proceedings of the Biological Society of Washington 91(3): 658–665. [Google Scholar]
- Kensley B, Schotte M. (1989) Guide to the Marine Isopod Crustaceans of the Caribbean. Smithsonian Institution Press, Washington, 308 pp. 10.5962/bhl.title.10375 [DOI] [Google Scholar]
- Kerstitch A, Bertsch H. (2007) Sea of Cortez Marine Invertebrates. A Guide for the Pacific Coast, Mexico to Peru. 2nd edn. Sea Challengers, Monterey, 124 pp. [Google Scholar]
- Khalaji-Pirbalouty V, Gagnon J-M. (2021) A new species of Dynoides Barnard, 1914 (Crustacea, Isopoda, Sphaeromatidae) from Canada, with notes on geographic distribution of the north-eastern Pacific Ocean species. Marine Biology Research 17(1): 12–20. 10.1080/17451000.2021.1892766 [DOI] [Google Scholar]
- Khalaji-Pirbalouty V, Wägele J-W. (2010) A new species and a new record of Sphaeroma Bosc, 1802 (Sphaeromatidae: Isopoda: Crustacea) from intertidal marine habitats of the Persian Gulf. Zootaxa 2631(1): 1–18. 10.11646/zootaxa.2631.1.1 [DOI] [Google Scholar]
- Kim SH, Yoon SM. (2021) Four new species of isopods (Crustacea, Isopoda) from South Korea. ZooKeys 1010: 133–164. 10.3897/zookeys.1010.59101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- King RA. (2003) Neastacilla Tattersall, 1921 redefined, with eight new species from Australia (Crustacea: Isopoda: Arcturidae). Memoirs of the Museum of Victoria 60(2): 371–416. 10.24199/j.mmv.2003.60.29 [DOI] [Google Scholar]
- Klapow LA. (1972) Fortnightly molting and reproductive cycles in the sand-beach isopod, Excirolanachiltoni. The Biological Bulletin 143(3): 568–591. 10.2307/1540184 [DOI] [PubMed] [Google Scholar]
- Kozloff EN. (1974) Keys to the Marine Invertebrates of Puget Sound, the San Juan Archipelago, and Adjacent Regions. University of Washington Press, Seattle and London, 226 pp. [Google Scholar]
- Kozloff EN. (1983) Seashore Life of the Northern Pacific Coast. An Illustrated Guide to Northern California, Oregon, Washington, and British Columbia. University of Washington Press, Seattle and London, 370 pp. [Google Scholar]
- Kozloff EN. (1996) Marine Invertebrates of the Pacific Northwest. Revised and expanded edition. University of Washington Press, Seattle and London, 539 pp. [Google Scholar]
- Kuris AM, Carlton JT. (1977) Description of a new species, Crangonhandi, and new genus, Lissocrangon, of crangonid shrimps (Crustacea: Caridea) from the California coast, with notes on adaptation in body shape and coloration. The Biological Bulletin 153(3): 540–559. 10.2307/1540605 [DOI] [Google Scholar]
- Kussakin OG. (1979) Marine and brackish-water Isopoda of cold and temperate (boreal) waters of the Northern Hemisphere. Flabellifera. National Academy of Sciences, Union of Soviet Socialist Republics, Zoology (Opredeliteli po Faune USSR, Akademiya Nauk USSR) 122: 1–470. [Google Scholar]
- Kussakin OG. (1982) Marine and brackish-water Isopoda of cold and temperate (boreal) waters of the Northern Hemisphere. Anthuridea, Microcerberidea, Valvifera, Tyloidea. National Academy of Sciences, Union of Soviet Socialist Republics, Zoology (Opredeliteli po Faune USSR, Akademiya Nauk USSR) 131: 1–461. [Google Scholar]
- Kussakin OG. (1988) Marine and brackish-water Isopoda of cold and temperate (boreal) waters of the Northern Hemisphere. Vol. III. Asellota, Part 1. National Academy of Sciences, Union of Soviet Socialist Republics, Zoology (Opredeliteli po Faune USSR, Akademiya Nauk USSR) 152: 1–500. [Google Scholar]
- Kussakin OG. (1999) Marine and brackish-water Isopoda of cold and temperate (boreal) waters of the Northern Hemisphere. Vol. III. Asellota, Part 2. National Academy of Sciences, Russia, Zoology (Opredeliteli po Faune, Akademiya Nauk Russia) 169: 1–383. [Google Scholar]
- Kussakin OG. (2003) Marine and brackish-water Isopoda of cold and temperate (boreal) waters of the Northern Hemisphere. Vol. III. Asellota, Part 3. National Academy of Sciences, Russia, Zoology (Opredeliteli po Faune, Akademiya Nauk Russia) 171: 1–380. [Google Scholar]
- Lafferty KD, Suchanek TH. (2016) Revisiting Paine’s 1966 sea star removal experiment, the most-cited empirical article in the American Naturalist. American Naturalist 188(4): 365–378. 10.1086/688045 [DOI] [PubMed] [Google Scholar]
- Latreille PA. (1817) Les Crustacés, les Arachnides et les Insectes, distribués en familles naturelles. In: Cuvier M leC (Ed. ) Le Règne Animal Distribué d’Après son Organisation, pour Servir de Base a l’Histoire Naturelle des Animaux et d’Introduction a l’Anatomie Comparée Paris, Deterville 3: 1–653. [Google Scholar]
- Lavesque N, Sorbe J-C, Bachelet G, Gouillieux B, de Montaudouin X, Bonifacio P, Blanchet H, Dubois S. (2013) Recent discovery of Paranthurajaponica Richardson, 1909 (Crustacea: Isopoda: Paranthuridae) in European marine waters (Arcachon Bay, Bay of Biscay). BioInvasions Records 2(3): 215–219. 10.3391/bir.2013.2.3.07 [DOI] [Google Scholar]
- Lazarus-Agudelo JF, Cantera-Kintz JR. (2007) Crustáceos (Crustacea: Sessilia, Stomatopoda, Isopoda, Amphipoda, Decapoda) de Bahía Málaga, Valle del Cauca (Pacífico colombiano). Biota Colombiana 8(2): 221–239. [Google Scholar]
- Leach WE. (1814) Crustaceology. In: Brewster D. (Ed.) The Edinburgh Encyclopedia.Balfour, Edinburgh 7(2): 385–437, 765–766.
- Lee WL, Miller MA. (1980) Isopoda and Tanaidacea: the isopods and allies. In: Morris RH, Abbott DP, Haderlie EC. (Eds) Intertidal Invertebrates of California, 3rd edn.Stanford University Press, Stanford, 536–558.
- Leistikow A, Wägele J-W. (1999) Checklist of the terrestrial isopods of the new world (Crustacea, Isopoda, Oniscidea). Revista Brasileira de Zoologia 16(1): 1–72. 10.1590/S0101-81751999000100001 [DOI] [Google Scholar]
- Lendemer J, Thiers B, Monfils AK, Zaspel J, Ellwood ER, Bentley A, LeVan K, Bates J, Jennings D, Contreras D, Lagomarsino L, Mabee P, Ford LS, Guralnick R, Gropp RE, Revelez M, Cobb N, Seltmann K, Aime MC. (2020) The extended specimen network: A strategy to enhance US biodiversity collections, promote research and education. Bioscience 70(1): 23–30. 10.1093/biosci/biz140 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li L. (2000) A new species of Dynoides (Crustacea: Isopoda: Sphaeromatidae) from the Cape d’Aguilar Marine Reserve, Hong Kong. Records of the Australian Museum 52(2): 137–149. 10.3853/j.0067-1975.52.2000.1311 [DOI] [Google Scholar]
- Lie U. (1968) A quantitative study of benthic infauna in Puget Sound, Washington, USA, in 1963–1964. Fiskeridirektoratets skrifter. Serie Havundersøkelser 14(5): 229–556. [Google Scholar]
- Lissner A, Phillips C, Cadien D, Smith R, Bernstein B, Cimberg R, Kauwling T, Anikouchine W. (1986) Assessment of long-term changes in biological communities in the Santa Maria Basin and Western Santa Barbara Channel – Phase I. Vol. II, Synthesis of Findings. Prepared for: United States Department of the Interior, Minerals Management Service, Pacific OCS Region, Los Angeles, 789 pp. [Google Scholar]
- Liu W, Poore GCB. (2013) A new record of Cleantioidesemarginata Kwon & Kim, 1992 (Crustacea, Isopoda, Valvifera) from Changjiang River estuary, China. Chinese Journal of Oceanology and Limnology 31(3): 625–631. 10.1007/s00343-013-2197-9 [DOI] [Google Scholar]
- Lockington WN. (1877a) Remarks on the Crustacea of the Pacific Coast, with descriptions of some new species. Proceedings of the California Academy of Sciences 7 (for 1876): 44–46. 10.5962/bhl.part.27534 [DOI]
- Lockington WN. (1877b) Description of seventeen new species of Crustacea. Proceedings of the California Academy of Sciences 7 (for 1876): 41–48. 10.5962/bhl.title.37841 [DOI]
- Looby A, Ginsburg DW. (2021) Nearshore species biodiversity of a marine protected area off Santa Catalina Island, California. Western North American Naturalist 81(1): 113–130. 10.3398/064.081.0110 [DOI] [Google Scholar]
- Los Angeles County Sanitation Districts [LACSD] (2020) Joint Water Pollution Control Plant Biennial Receiving Water Monitoring Report 2018–2019. LACSD, Whittier, 162 pp. https://www.lacsd.org/home/showpublisheddocument/4432/637648109065900000 [Google Scholar]
- Loyola e Silva J. (1971) Sôbre os gêneros Ancinus Milne Edwards, 1840 e Bathycopea Tattersall, 1909, da coleção U.S. Nat. Mus. (Isopoda-Crustacea). Arquivos do Museu Nacional, Rio de Janeiro 54: 209–223. [Google Scholar]
- Macdonald TA, Burd BJ, Macdonald VI, van Roodselaar A. (2010) Taxonomic and feeding guild classification for the marine benthic macroinvertebrates of the Strait of Georgia, British Columbia. Canadian Technical Report of Fisheries and Aquatic Sciences 2874: [iv +] 63 pp.
- Mahmoud NE, Fahmy MM, Abuowarda MM, Khattab MS. (2016) Parasitic cymothoid isopods and their impacts in commercially important fishes from Lake Qarun, Egypt. International Journal of Chemtech Research 9(12): 221–229. [Google Scholar]
- Mak PMS, Huang ZG, Morton BS. (1985) Sphaeromawalkeri Stebbing (Isopoda, Sphaeromatidae) introduced into and established in Hong Kong. Crustaceana 49(1): 75–81. 10.1163/156854085X00224 [DOI] [Google Scholar]
- Maloney JO. (1933) Two new species of isopod crustaceans from California. Journal of the Washington Academy of Sciences 23(3): 144–147. [Google Scholar]
- Maloney E, Fairey R, Lyman A, Reynolds K, Sigala M. (2006) Introduced Aquatic Species in California Open Coastal Waters. Final Report (December 2006). California Department of Fish and Game. Office of Spill Prevention and Response. Sacramento, 93 pp.
- Maloney E, Fairey R, Lyman A, Walton Z, Sigala M. (2008) Introduced Aquatic Species in California’s Open Coastal Waters – 2007. Final Report. California Department of Fish and Game, Sacramento, 84 pp. [Google Scholar]
- Marchini A, Costa AC, Ferrario J, Micael J. (2018) The global invader Paracerceissculpta (Isopoda: Sphaeromatidae) has extended its range to the Azores Archipelago. Marine Biodiversity 48(2): 1001–1007. 10.1007/s12526-017-0674-7 [DOI] [Google Scholar]
- Markham JC. (1974a) Parasitic bopyrid isopods of the amphi-American genus Stegophryxus Thompson with the description of a new species from California. Bulletin of the Southern California Academy of Sciences 73: 33–41. [Google Scholar]
- Markham JC. (1974b) Extension of range and new host records for the parasitic isopod Pseudionegiardi Calman in the northeastern Pacific. Wasmann Journal of Biology 32(2): 195–201. [Google Scholar]
- Markham JC. (1975) A review of the bopyrid isopod genus Munidion Hansen, 1897, parasitic on galatheid crabs in the Atlantic and Pacific oceans. Bulletin of Marine Science 25(3): 422–441. [Google Scholar]
- Markham JC. (1977a) The status and systematic position of the species of the bopyrid isopod genus Phyllodurus Stimpson, 1857. Proceedings of the Biological Society of Washington 90(4): 813–818. [Google Scholar]
- Markham JC. (1977b) Description of new western Atlantic species of Argeia Dana with a proposed new subfamily for this and related genera (Crustacea, Isopoda, Bopyridae). Zoölogische Mededeelingen 52(9): 107–123. [Google Scholar]
- Markham JC. (1985a) A review of the bopyrid isopods infesting caridean shrimps in the northwestern Atlantic Ocean, with special reference to those collected during the Hourglass cruises in the Gulf of Mexico. Memoirs of the Hourglass Cruises 7(3): 1–156. [Google Scholar]
- Markham JC. (1985b) A new species of Asymmetrione (Isopoda: Bopyridae) infesting the hermit crab Isochelespilosus (Holmes) in southern California. Bulletin of the Southern California Academy of Sciences 84: 104–108. [Google Scholar]
- Markham JC. (1986) Evolution and zoogeography of the IsopodaBopyridae, parasites of CrustaceaDecapoda. In: Gore RH, Heck KL. (Eds) Crustacean Issues Vol.4. Crustacean Biogeography. Balkema, Rotterdam, 143–164. 10.1201/9781315140674-7 [DOI]
- Markham JC. (1992) The IsopodaBopyridae of the Eastern Pacific – missing or just hiding? Proceedings of the San Diego Society of Natural History 17: 1–4.
- Markham JC. (2001) A review of the bopyrid isopods parasitic on thalassinidean decapods. In: Brusca RC, Kensley B. (Eds) Crustacean Issues Vol.13. Isopod Systematics and Evolution. Balkema, Rotterdam, 195–204.
- Markham JC. (2002) A new species of Leidya Cornalia and Panceri, 1861, and the first record of the genus Lobocepon Nobili, 1905, both from the eastern Pacific Ocean, with a review of the parasites of grapsid crabs worldwide (Isopoda, Bopyridae, Ioninae). In: Alvarez F, Escobar-Briones E. (Eds) Modern Approaches to the Study of Crustacea.Kluwer Academic/Plenum Publishers, New York, 329–338. 10.1007/978-1-4615-0761-1_46 [DOI]
- Markham JC. (2003) A worldwide list of hermit crabs and their relatives (Anomura: Paguroidea) reported as hosts of IsopodaBopyridae. In: Lemaitre R, Tudge CC. (Eds) Biology of the Anomura.Proceedings of a symposium at the Fifth International Crustacean Congress, Melbourne, Australia, 9–13 July 2001. Memoirs of Museum Victoria 60(1), 71–77. 10.24199/j.mmv.2003.60.10 [DOI]
- Markham JC. (2004) New species and records of Bopyridae (Crustacea: Isopoda) infesting species of the genus Upogebia (Crustacea: Decapoda: Upogebiidae): the genera Orthione Markham, 1988, and Gyge Cornalia and Panceri, 1861. Proceedings of the Biological Society of Washington 117(2): 186–198. [Google Scholar]
- Markham JC. (2005) New species and records of Bopyridae (Crustacea: Isopoda) infesting species of the genus Upogebia (Crustacea: Decapoda: Upogebiidae): the genus Progebiophilus Codreanu and Codreanu, 1963; and remarks on Phyllodurus Stimpson, 1857. Proceedings of the Biological Society of Washington 118(1): 84–95. 10.2988/0006-324X(2005)118[84:NSAROB]2.0.CO;2 [DOI]
- Markham JC. (2008) New records of pseudionine bopyrid isopods, including two new species and one new genus, infesting porcellanid crabs (Decapoda: Anomura) on the Pacific coast of North and Central America. Bulletin of the Southern California Academy of Sciences 107(3): 145–157. 10.3160/0038-3872-107.3.145 [DOI] [Google Scholar]
- Markham JC. (2016) Redescription of Bathygygegrandis Hansen, 1897 (Crustacea, Isopoda, Bopyridae) from Southern California with erection of a new subfamily, Bathygyginae, for it. Bulletin of the Southern California Academy of Sciences 115(1): 72–80. 10.3160/soca-115-01-72-80.1 [DOI] [Google Scholar]
- Markham JC. (2020) Chapter 6. IsopodaEpicaridea from deep water around North and Central America. In: Hendrickx ME (Ed.) Deep-Sea Pycnogonids and Crustaceans of the Americas, Springer, 143–156. 10.1007/978-3-030-58410-8_6 [DOI]
- Martin JW, Davis GE. (2001) An updated classification of the recent Crustacea. Natural History Museum of Los Angeles County Contributions in Science 39: 1–124. [Google Scholar]
- Martin JW, Olesen J, Høeg JT [Eds] (2014) Atlas of crustacean larvae. John Hopkins University Press, Baltimore, 370 pp. 10.1353/book.31448 [DOI] [Google Scholar]
- Martin MB, Bruce NL, Nowak BF. (2015) Review of the fish-parasitic genus Ceratothoa Dana, 1852 (Crustacea: Isopoda: Cymothoidae) from Australia, with description of two new species. Zootaxa 3963(3): 251–294. 10.11646/zootaxa.3963.3.1 [DOI] [PubMed] [Google Scholar]
- Martin JW, Pentcheff ND, Striley D, Perrodin C, Wall AR, Wall J, Zieba J. (Eds) (2022) Crustacea Glossary. Crustacea Department, Natural History Museum of Los Angeles County. [Accessed: 06 Sep 2022]https://research.nhm.org/glossary
- McDermott JJ, Williams JD, Boyko CB. (2010) The unwanted guests of hermits: A global review of the diversity and natural history of hermit crab parasites. Journal of Experimental Marine Biology and Ecology 394(1–2): 2–44. 10.1016/j.jembe.2010.06.022 [DOI] [Google Scholar]
- McLaughlin PA, Camp DK, Angel MV, Bousfield EL, Brunel P, Brusca RC, Cadien D, Cohen AC, Conlan K, Eldredge LG, Felder DL, Goy JW, Haney T, Hann B, Heard RW, Hendrycks EA, Hobbs III HH, Holsinger JR, Kensley B, Laubitz DR, LeCroy SE, Lemaitre R, Maddocks RF, Martin JW, Mikkelsen P, Nelson E, Newman WA, Overstreet RM, Poly WJ, Price WW, Reid JW, Robertson A, Rogers DC, Ross A, Schotte M, Schram FR, Shih C-T, Watling L, Wilson GDF, Turgeon DD. (2005) Common and Scientific Names of Aquatic Invertebrates from the United States and Canada: Crustaceans. American Fisheries Society Special Publication 31: [xiii +] 1–545 [+ CD-ROM].
- Mejía-Rebollo A, Zayas-Álvarez A, Morales-Zárate V, Salinas-Zavala CA. (2018) New record and contribution to the knowledge of Neastacillacalifornica (Boone, 1918) (Isopoda, Arcturidae) on the west coast of Baja California, Mexico. Latin American Journal of Aquatic Research 46(1): 230–233. 10.3856/vol46-issue1-fulltext-23 [DOI] [Google Scholar]
- Melwani AR, Kim SL. (2008) Benthic infaunal distributions in shallow hydrothermal vent sediments. Acta Oecologica 33(2): 162–175. 10.1016/j.actao.2007.10.008 [DOI] [Google Scholar]
- Menzies RJ. (1950a) The taxonomy, ecology, and distribution of northern California isopods of the genus Idothea with the description of a new species. Wasmann Journal of Biology 8(2): 155–195. [Google Scholar]
- Menzies RJ. (1950b) A remarkable new species of marine isopod, Erichsonellacrenulata n. sp., from Newport Bay, California. Bulletin of the Southern California Academy of Sciences 49: 29–35. 10.3160/0038-3872-49.1.29 [DOI] [Google Scholar]
- Menzies RJ. (1950c) Notes on California isopods of the genus Armadilloniscus, with the description of Armadilloniscuscoronacapitalis n. sp. Proceedings of the California Academy of Sciences 4th series 26(13): 467–481. https://biostor.org/reference/78362
- Menzies RJ. (1951a) A new species of Limnoria (Crustacea: Isopoda) from southern California. Bulletin of the Southern California Academy of Sciences 50: 86–88. [Google Scholar]
- Menzies RJ. (1951b) New marine isopods, chiefly from northern California, with notes on related forms. Proceedings of the United States National Museum 101(3273): 105–156. 10.5479/si.00963801.101-3273.105 [DOI] [Google Scholar]
- Menzies RJ. (1951c) A new genus and new species of asellote isopod, Caecijaerahorvathi, from Los Angeles-Long Beach Harbor. American Museum Novitates 1542: 1–7. http://hdl.handle.net/2246/3702 [Google Scholar]
- Menzies RJ. (1952) Some marine asellote isopods from northern California, with descriptions of nine new species. Proceedings of the United States National Museum 102(3293): 117–159. 10.5479/si.00963801.102-3293.117 [DOI] [Google Scholar]
- Menzies RJ. (1954) A review of the systematics and ecology of the genus “Exosphaeroma,” with the description of a new genus, a new species, and a new subspecies (Crustacea, Isopoda, Sphaeromatidae). American Museum Novitates 1683: 1–24. http://hdl.handle.net/2246/4909 [Google Scholar]
- Menzies RJ. (1957) The marine borer family Limnoriidae (Crustacea, Isopoda). Bulletin of Marine Science of the Gulf and Caribbean 7: 101–200. [Google Scholar]
- Menzies RJ. (1958) The distribution of wood-boring Limnoria in California. Proceedings of the California Academy of Sciences 4th series 29(7): 267–272.
- Menzies RJ. (1962a) The marine isopod fauna of Bahia de San Quintin, Baja California, Mexico. Pacific Naturalist 3(11): 337–348. [Google Scholar]
- Menzies RJ. (1962b) The zoogeography, ecology, and systematics of the Chilean marine isopods. Reports of the Lund University Chile Expedition 1948–49. 42. Lunds Universitets Arsskrifter N.F. Avd. 2, 57(11): 1–162.
- Menzies RJ, Barnard JL. (1951) The isopodan genus Iais (Crustacea). Bulletin of the Southern California Academy of Sciences 50(Part 3): 136–151.
- Menzies RJ, Barnard JL. (1959) Marine Isopoda on coastal shelf bottoms of southern California: Systematics and ecology. Pacific Naturalist 1(11): 1–35. [Google Scholar]
- Menzies RJ, Bowman TE. (1956) Emended description and assignment to the new genus Ronalea of the idotheid isopod Erichsonellapseudoculata Boone. Proceedings of the United States National Museum 106(3371): 339–343. 10.5479/si.00963801.106-3371.339 [DOI] [Google Scholar]
- Menzies RJ, Glynn PW. (1968) The common marine isopod Crustacea of Puerto Rico. A handbook for marine biologists. Studies on the fauna of Curaçao and other Caribbean Islands 27(1): 1–133. [Google Scholar]
- Menzies RJ, Miller MA. (1972) Systematics and zoogeography of the genus Synidotea (Crustacea: Isopoda) with an account of Californian species. Smithsonian Contributions to Zoology 102(102): 1–33. 10.5479/si.00810282.102 [DOI] [Google Scholar]
- Menzies RJ, Mohr JL. (1952) The occurrence of the wood-boring crustacean Limnoria and of Nebaliacea in Morro Bay, California. Wasmann Journal of Biology 10(1): 81–86. [Google Scholar]
- Menzies RJ, Pettit J. (1956) A new genus and species of marine asellote isopod, Caecianiropsispsammophila, from California. Proceedings of the United States National Museum 106(3376): 441–446. 10.5479/si.00963801.106-3376.441 [DOI] [Google Scholar]
- Menzies RJ, Waidzunas RJ. (1948) Postembryonic growth changes in the isopod Pentidotearesecata (Stimpson) with remarks on their taxonomic significance. The Biological Bulletin 95(1): 107–113. 10.2307/1538157 [DOI] [PubMed] [Google Scholar]
- Menzies RJ, Bowman TE, Alverson FG. (1955) Studies of the biology of the fish parasite Livonecaconvexa Richardson (Crustacea, Isopoda, Cymothoidae). Wasmann Journal of Biology 13(2): 277–295. [Google Scholar]
- Meyer C, Duffy E, Collins A, Paulay G, Wetzer R. (2021) The U.S. Ocean Barcode. Marine Technology Society Journal 55(3): 140–141. 10.4031/MTSJ.55.3.33 [DOI] [Google Scholar]
- Mezhov BV. (1986) Bathyal and abyssal Nannoniscidae and Desmosomatidae (Isopoda, Asellota) from Alaska Bay. Archives of the Zoological Museum, Moscow State University 24: 126–167. [In Russian] [Google Scholar]
- Miers EJ. (1881) Revision of the Idoteidae, a family of sessile-eyed Crustacea. Zoological Journal of the Linnean Society 16(89): 1–88. 10.1111/j.1096-3642.1881.tb02274a.x [DOI] [Google Scholar]
- Miller MA. (1936) California isopods of the genus Porcellio with descriptions of a new species and a new subspecies. University of California Publications in Zoology 41: 165–172. [Google Scholar]
- Miller MA. (1968) Isopoda and Tanaidacea from buoys in coastal waters of the continental United States, Hawaii, and the Bahamas (Crustacea). Proceedings of the United States National Museum 125(3652): 1–53. 10.5479/si.00963801.125-3652.1 [DOI] [Google Scholar]
- Miller MA. (1975) Phylum Arthropoda: Crustacea, Tanaidacea and Isopoda. In: Smith RI, Carlton JT. (Eds) Light’s Manual: Intertidal Invertebrates of the Central California Coast.University of California Press, Berkeley, 277–312.
- Miller MA, Lee WL. (1970) A new idoteid isopod Idothea (Pentidotea) kirchanskii, from Central California (Crustacea). Proceedings of the Biological Society of Washington 82: 789–798. [Google Scholar]
- Miller SE, Barrow LN, Ehlman SM, Goodheart JA, Greiman SE, Lutz HL, Misiewicz TM, Smith SM, Tan M, Thawley CJ, Cook JA, Light JE. (2020) Building natural history collections for the twenty-first century and beyond. Bioscience 70(8): 674–687. 10.1093/biosci/biaa069 [DOI] [Google Scholar]
- Milne Edwards H. (1840) Histoire Naturelle des Crustacés, Comprenant l’Anatomie, la Physiologie et la Classification de ces Animaux. Encyclopédique Roret, Paris III: 1–638.
- Moles A. (2007) Parasites of the fishes of Alaska and surrounding waters. Alaska Fishery Research Bulletin 12(2): 197–226. [Google Scholar]
- Monod T. (1926) Les Gnathiidae. Essai monographique (morphologie, biologie, systematique). Memoires de la Societe des Sciences Naturelles du Maroc 12: 1–667. [Google Scholar]
- Montagne DE, Cadien DB. (2001) Northern range extensions into the Southern California Bight of ten decapod Crustacea related to the 1991/92 and 1997/98 El Niño events. Bulletin of the Southern California Academy of Sciences 100(3): 199–211. [Google Scholar]
- Montelli L, Lewis J. (2008) Survey of Biofouling on Australian Navy Ships: Crustacea; Isopoda and Amphipoda; Caprellidea. Technical Report DSTO-TR-2098, Maritime Platforms Division, DSTO Defense Science and Technology Organization, Victoria, Australia, 50 pp. [Google Scholar]
- Moore W. (2004) Description of a new Synidotea species (Crustacea: Isopoda: Valvifera: Idoteidae) from Hawaii. Proceedings of the Biological Society of Washington 117(1): 76–87. [Google Scholar]
- Moore EC, Hovel KA. (2010) Relative influence of habitat complexity and proximity to patch edges on seagrass epifaunal communities. Oikos 119(8): 1299–1311. 10.1111/j.1600-0706.2009.17909.x [DOI] [Google Scholar]
- Morales-Zarate MV, Zayas-Alvarez A, Salinas-Zavala CA, Mejia-Rebollo A. (2016) Biocenosis of benthic community in Guerrero Negro Lagoon, Baja California Sur, Mexico: Spatio-temporal characterization. Latin American Journal of Aquatic Research 44(4): 726–742. 10.3856/vol44-issue4-fulltext-8 [DOI] [Google Scholar]
- Müller H. (1990) Hyssuridae from the Caribbean coast of Colombia, with descriptions of four new species (Isopoda: Anthuridea). Bijdragen tot de Dierkunde 60(2): 65–78. 10.1163/26660644-06002001 [DOI] [Google Scholar]
- Munguia P, Shuster SM. (2013) Established populations of Paracerceissculpta (Isopoda) in the northern Gulf of Mexico. Journal of Crustacean Biology 33(1): 137–139. 10.1163/1937240X-00002112 [DOI] [Google Scholar]
- NASEM [National Academies of Sciences, Engineering, and Medicine] (2020) Biological Collections: Ensuring Critical Research and Education for the 21st Century. Washington, DC: The National Academies Press, 244 pp. 10.17226/25592 [DOI] [PubMed] [Google Scholar]
- Naylor E. (1957) The occurrence of Idoteametallica Bosc in British waters. Journal of the Marine Biological Association of the United Kingdom 36(3): 599–602. 10.1017/S002531540002587X [DOI] [Google Scholar]
- Negromonte AO, Lucatelli D, Paiva RJC, Souza-Filho JF. (2022) Three new species of the genus Cleantioides Kensley and Kaufman, 1978 (Isopoda: Valvifera) from Brazil, with new record of the family Holognathidae from Brazil. Journal of Natural History 56(13–16): 923–941. 10.1080/00222933.2022.2056538 [DOI] [Google Scholar]
- Nierstrasz HF, Brender à Brandis GA. (1926) Isopoda. Epicaridea. Die Tierwelt der Nord- und Ostsee Teil X, Section E, Part 1: 1–56. 10.1163/26660644-02401001 [DOI] [Google Scholar]
- Nierstrasz HF, Brender à Brandis GA. (1929) Papers from Dr. Th. Mortensen’s Pacific Expedition 1914-16. 48. Epicaridea 1. Videnskabelige Meddelelser fra Dansk Naturhistorisk Forening I Kjobenhavn 87: 1–44. [Google Scholar]
- Nobuhiro S, Kyoko K. (2004) Prevalence of the bopyrid isopod Ionecornuta (Crustacea: Isopoda: Epicaridea) on the ghost shrimp Nihonotrypaeajaponica (Crustacea: Decapoda: Thalassinidea) on a tidal flat of Shinhama Lagoon, northern Tokyo Bay. Japanese Journal of Benthology 59: 1–7. 10.5179/benthos.59.1 [DOI] [Google Scholar]
- O’Clair RM, O’Clair CE. (1998) Southeast Alaska’s Rocky Shores. Animals. Plant Press, Auke Bay, 564 pp. [Google Scholar]
- Orange County Sanitation District [OCSD] (2022) Marine Monitoring Annual Report, Program Year 2020–2021. OCSD, Environmental Laboratory and Ocean Monitoring Division, Fountain Valley, 144 pp. https://www.ocsan.gov/home/showpublisheddocument/32585/637830212490770000 [Google Scholar]
- Ormsby B. (1991) Synisomawetzerae, a new species and the first record of Synisoma from the new world (Crustacea: Isopoda: Valvifera: Idoteidae). Proceedings of the Biological Society of Washington 104(4): 758–763. [Google Scholar]
- Ortiz M, Lalana R. (2018) Lista de especies y distribución de los isópodos (Crustacea: Peracarida) de Cuba. Novitates Caribaea (12): 102–126. 10.33800/nc.v0i12.87 [DOI]
- Özdikem H. (2009) A new name, Smenispa for the preoccupied isopod genus Enispa Schioedte & Meinert, 1884 (Isopoda: Cymothoidae). Munis Entomology & Zoology 4(2): 611–612. [Google Scholar]
- Pearse AS. (1953) Parasitic crustaceans from Alligator Harbor, Florida. Quarterly Journal of the Florida Academy of Sciences 15(4): 187–243. [Google Scholar]
- Pérez-Schultheiss J, Urra F. (2020) Isópodos terrestres (Crustacea: Isopoda: Oniscidea) del Parque Nacional Bosque Fray Jorge, Región de Coquimbo, Chile. Boletín del Museo Nacional de Historia Natural 69(2): 29–54. 10.54830/bmnhn.v69.n2.2020.4 [DOI] [Google Scholar]
- Pérez-Schultheiss J, Ayala K, Fariña JM, Coccia C. (2018) Exotic oniscideans (Crustacea: Isopoda) in coastal salt marshes: first record of the families Halophilosciidae and Platyarthridae in continental Chile. New Zealand Journal of Zoology 46(3): 225–235. 10.1080/03014223.2018.1539017 [DOI] [Google Scholar]
- Pfeffer G. (1887) Die Krebse von Süd-Georgien nach der Ausbeute der Deutschen Station 1882-83. Jahrbuch der Hamburgischen Wissenschaftlichen Anstalten 4: 43–150 [pls. 1–7]. 10.5962/bhl.title.10084 [DOI]
- Pires AMS. (1980) New record of Sphaeromatidae (Isopoda) from the Brazilian southern coast: Dynamenelladianae (Menzies, 1962). Crustaceana 39(2): 133–140. 10.1163/156854080X00030 [DOI] [Google Scholar]
- Pires AMS. (1984) Taxonomic revision and phylogeny of the genus Erichsonella with a discussion on Ronalea (Isopoda, Valvifera). Journal of Natural History 18(5): 665–683. 10.1080/00222938400770571 [DOI] [Google Scholar]
- Pires AMS. (1987) The contribution of isopods in the feeding of Sympterygia spp. (Pisces: Rajidae) with a description of Ancinusgaucho sp. n. (Isopoda: Sphaeromatidae). Boletim do Instituto Oceanografico, São Paulo 35(2): 115–122. 10.1590/S0373-55241987000200002 [DOI] [Google Scholar]
- Pires-Vanin AMS, Muniz P, Bromberg S. (2014) Inventory of the marine soft bottom macrofauna of São Sebastião Channel, southeastern Brazilian continental shelf. Check List 10(4): 795–807. 10.15560/10.4.795 [DOI] [Google Scholar]
- Poore GCB. (1975) Australian species of Haliophasma (Crustacea: Isopoda: Anthuridae). Records of the Australian Museum 29(19): 503–533. 10.3853/j.0067-1975.29.1975.211 [DOI] [Google Scholar]
- Poore GCB. (1980) A revision of the genera of the Paranthuridae (Crustacea: Isopoda: Anthuridea) with a catalogue of species. Zoological Journal of the Linnean Society 68(1): 53–67. 10.1111/j.1096-3642.1980.tb01918.x [DOI] [Google Scholar]
- Poore GCB. (1984a) Paranthura (Crustacea, Isopoda, Paranthuridae) from south-eastern Australia. Memoirs of the Museum of Victoria 45(1): 33–69. 10.24199/j.mmv.1984.45.04 [DOI] [Google Scholar]
- Poore GCB. (1984b) Colanthura, Califanthura, Cruranthura and Cruregens, related genera of the Paranthuridae (Crustacea: Isopoda). Journal of Natural History 18(5): 697–715. 10.1080/00222938400770601 [DOI] [Google Scholar]
- Poore GCB. (1984c) Redefinition of Munna and Uromunna (Crustacea: Isopoda: Munnidae), with descriptions of five species from coastal Victoria. Proceedings of the Royal Society of Victoria 96(2): 61–81. [Google Scholar]
- Poore GCB. (1996) Species differentiation in Synidotea (Isopoda: Idoteidae) and recognition of introduced marine species: a reply to Chapman and Carlton. Journal of Crustacean Biology 16(2): 384–394. 10.2307/1548894 [DOI] [Google Scholar]
- Poore GCB. (2001a) Families and genera of IsopodaAnthuridea. In: Kensley B, Brusca RC. (Eds) Isopod Systematics and Evolution.Balkema: Rotterdam, Crustacean Issues 13: 63–173.
- Poore GCB. (2001b) IsopodaValvifera: Diagnoses and relationships of the families. Journal of Crustacean Biology 21(1): 205–230. 10.1163/20021975-99990118 [DOI] [Google Scholar]
- Poore GCB. (2015) Thermoarcturidae, a new crustacean family of three genera (Isopoda: Valvifera). Zootaxa 4007(3): 409–418. 10.11646/zootaxa.4007.3.7 [DOI] [PubMed] [Google Scholar]
- Poore GCB, Bruce NL. (2012) Global diversity of marine isopods (except Asellota and crustacean symbionts). PLoS ONE 7(8): e43529. 10.1371/journal.pone.0043529 [DOI] [PMC free article] [PubMed]
- Poore GCB, Lew Ton HM. (1986) Mesanthura (Crustacea: Isopoda: Anthuridae) from southeastern Australia. Memoirs of the Museum of Victoria 47(1): 87–104. 10.24199/j.mmv.1986.47.04 [DOI] [Google Scholar]
- Poore GCB, Lew Ton HM. (1988a) More Australian species of Haliophasma (Crustacea: Isopoda: Anthuridae). Memoirs of the Museum of Victoria 49(1): 85–106. 10.24199/j.mmv.1988.49.07 [DOI] [Google Scholar]
- Poore GCB, Lew Ton HM. (1988b) Amakusanthura and Apanthura (Crustacea: Isopoda: Anthuridae) with new species from tropical Australia. Memoirs of the Museum of Victoria 49(1): 107–147. 10.24199/j.mmv.1988.49.08 [DOI] [Google Scholar]
- Poore GCB, Lew Ton HM. (1988c) A generic review of the Hysurridae (Crustacea: Isopoda). Memoirs of the Museum of Victoria 49(1): 169–193. 10.24199/j.mmv.1988.49.11 [DOI] [Google Scholar]
- Poore GCB, Lew Ton HM. (1988d) Antheluridae, a new family of Crustacea (Isopoda: Anthuridea) with new species from Australia. Journal of Natural History 22(2): 489–506. 10.1080/00222938800770341 [DOI] [Google Scholar]
- Poore GCB, Lew Ton HM. (1990) The Holognathidae (Crustacea: Isopoda: Valvifera) expanded and redefined on the basis of body-plan. Invertebrate Taxonomy 4(1): 55–80. 10.1071/IT9900055 [DOI] [Google Scholar]
- Poore GCB, Lew Ton HM. (1993) Idoteidae of Australia and New Zealand (Crustacea: Isopoda: Valvifera). Invertebrate Taxonomy 7(2): 197–278. 10.1071/IT9930197 [DOI] [Google Scholar]
- Poore GCB, Lew Ton HM. (2002) Expanathuridae (Crustacea: Isopoda) from the Australian region. Zootaxa 82(1): 1–60. 10.11646/zootaxa.82.1.1 [DOI] [Google Scholar]
- Puget Sound Institute (2012–2023) Encyclopedia of Puget Sound. University of Washington Tacoma Center for Urban Waters, Tacoma. https://www.eopugetsound.org [Accessed on 30 Jan 2023]
- Rafi F, Laubitz DR. (1990) The Idoteidae (Crustacea: Isopoda: Valvifera) of the shallow waters of the northeastern North Pacific Ocean. Canadian Journal of Zoology 68(12): 2649–2687. 10.1139/z90-370 [DOI] [Google Scholar]
- Ramalhosa P, Nebra A, Gestoso I, Canning-Clode J. (2017) First record of the non-indigenous isopods Paracerceissculpta (Holmes, 1904) and Sphaeromawalkeri Stebbing, 1905 (Isopoda, Sphaeromatidae) for Madeira Island. Crustaceana 90(14): 1747–1764. 10.1163/15685403-00003727 [DOI] [Google Scholar]
- Ranasinghe JA, Montagne DE, Smith RW, Mikel TK, Weisberg SB, Cadien D, Velarde R, Dalkey A. (2003) Southern California Bight 1998 Regional Monitoring Program: VII. Benthic Macrofauna. Technical Report 382. Southern California Coastal Water Research Project, Westminster, CA, 91 pp. [+ appendices] https://www.sccwrp.org/publications [Google Scholar]
- Ranasinghe JA, Mikel TK, Velarde RG, Weisberg SB, Montagne DE, Cadien DB, Dalkey A. (2005) The prevalence of non-indigenous species in southern California embayments and their effects on benthic macroinvertebrate communities. Biological Invasions 7(4): 679–686. 10.1007/s10530-004-5857-z [DOI] [Google Scholar]
- Ranasinghe JA, Barnett AM, Schiff K, Montagne DE, Brantley C, Beegan C, Cadien DB, Cash C, Deets GB, Diener DR, Mikel TK, Smith RW, Velarde RG, Watts SD, Weisberg SB. (2007) Southern California Bight 2003 Regional Monitoring Program: III. Benthic Macrofauna. Technical Report 529. Southern California Coastal Water Research Project, Costa Mesa, 44 pp. [+ appendices] https://www.sccwrp.org/publications [Google Scholar]
- Ranasinghe JA, Schiff KC, Brantley CA, Lovell LL, Cadien DB, Mikel TK, Velarde RG, Holt S, Johnson SC. (2012) Southern California Bight 2008 Regional Monitoring Program: VI. Benthic Macrofauna. Technical Report 665. Southern California Coastal Water Research Project, Costa Mesa, CA, 71 pp. [+ appendices] https://www.sccwrp.org/publications [Google Scholar]
- Reed BJ, Hovel KA. (2006) Seagrass habitat disturbance: How loss and fragmentation of eelgrass Zosteramarina influences epifaunal abundance and diversity. Marine Ecology Progress Series 326: 133–143. 10.3354/meps326133 [DOI] [Google Scholar]
- Richardson H. (1898) Description of four new species of Rocinela, with a synopsis of the genus. Proceedings of the American Philosophical Society 37(157): 8–17. [Google Scholar]
- Richardson H. (1899) Key to the isopods of the Pacific coast of North America, with descriptions of twenty-two new species. Proceedings of the United States National Museum 21(1175): 815–869. 10.5479/si.00963801.21-1175.815 [DOI] [Google Scholar]
- Richardson H. (1900a) Results of the Branner-Agassiz expedition to Brazil. II. The isopod Crustacea. Proceedings of the Washington Academy of Sciences 2: 157–159. 10.5962/bhl.part.5669 [DOI] [Google Scholar]
- Richardson H. (1900b) Synopses of North-American invertebrates. VIII. The Isopoda. Part I. Chelifera, Flabellifera, Valvifera. American Naturalist 34(399): 207–230. 10.1086/277593 [DOI] [Google Scholar]
- Richardson H. (1900c) Synopses of North-American invertebrates. VIII. The Isopoda. Part II. Asellota, Oniscoidea, Epicaridea. American Naturalist 34(400): 295–309. 10.1086/277619 [DOI] [Google Scholar]
- Richardson H. (1904a) Contributions to the natural history of the Isopoda. Parts I–IV. Proceedings of the United States National Museum 27(1350): 1–89. 10.5479/si.00963801.27-1350.113 [DOI] [Google Scholar]
- Richardson H. (1904b) Contributions to the natural history of the Isopoda. Parts V–VI. Proceedings of the United States National Museum 27(1369): 657–681. 10.5479/si.00963801.27-1369.657 [DOI] [Google Scholar]
- Richardson H. (1905a) A monograph of the isopods of North America. Bulletin of the United States National Museum 54(54): 1–727. 10.5479/si.03629236.54.i [DOI] [Google Scholar]
- Richardson H. (1905b) Isopods from the Alaska Salmon Investigation. Bulletin of the Bureau of Fisheries (for 1904) 24(1): 209–221.
- Richardson H. (1905c) Description of a new species of Livoneca from the Atlantic coast of Panama. Proceedings of the United States National Museum 29(1430): 445–446. 10.5479/si.00963801.29-1430.445 [DOI] [Google Scholar]
- Richardson H. (1906) Descriptions of new isopod crustaceans of the family Sphaeromidae. Proceedings of the United States National Museum 31(1479): 1–22. 10.5479/si.00963801.31-1479.1 [DOI] [Google Scholar]
- Richardson H. (1908) On some isopods of the family Dajidae from the northwest Pacific Ocean, with descriptions of a new genus and two new species. Proceedings of the United States National Museum 33(1586): 689–696. 10.5479/si.00963801.33-1586.689 [DOI] [Google Scholar]
- Richardson H. (1909) Isopods collected in the northwest Pacific by the U. S. Bureau of Fisheries Steamer “Albatross” in 1906. Proceedings of the United States National Museum 37(1701): 75–129. 10.5479/si.00963801.37-1701.75 [DOI] [Google Scholar]
- Richardson H. (1912) Descriptions of a new genus of isopod crustaceans, and of two new species from South America. Proceedings of the United States National Museum 43(1929): 201–204. 10.5479/si.00963801.43-1929.201 [DOI] [Google Scholar]
- Ricketts EF, Calvin J. (1952) Between Pacific Tides. 3rd edn., revised by JW Hedgpeth. Stanford University Press, Stanford, 502 pp. [Google Scholar]
- Ricketts EF, Calvin J. (1968) Between Pacific Tides. 4th edn., revised by JW Hedgpeth. Stanford University Press, Stanford, 614 pp. [Google Scholar]
- Ricketts EF, Calvin J, Hedgpeth JW. (1985) Between Pacific Tides. 5th edn., revised by DW Phillips. Stanford University Press, Stanford, 652 pp. 10.1515/9781503621329 [DOI] [Google Scholar]
- Riera R. (2014) Are marine drifting species cosmopolitan? The example of the isopod Idoteametallica. Thalassas. International Journal of Material Science 30(1): 75–78. [Google Scholar]
- Rocha LA, Aleixo A, Allen G, Almeda F, Baldwin CC, Barclay MVL, Bates JM, Bauer AM, Benzoni F, Berns CM, Berumen ML, Blackburn DC, Blum S, Bolaños F, Bowie RCK, Britz R, Brown RM, Cadena CD, Carpenter K, Ceríaco LM, Chakrabarty P, Chaves G, Choat JH, Clements KD, Colette BB, Collins A, Coyne J, Cracraft J, Daniel D, de Carvalho MR, de Queiroz K, Di Dario F, Drewes R, Dumbacher JP, English Jr A, Erdmann MV, Eschmeyer W, Feldman CR, Fisher BL, Fjeldså J, Fritsch PW, Fuchs J, Getahun A, Gill A, Gomon M, Gosliner T, Graves GR, Griswold CE, Guralnick R, Hartel K, Helgen KM, Ho H, Iskandar DT, Iwamoto T, Jaafar Z, James HF, Johnson D, Kavanaugh D, Knowlton N, Lacey E, Larson HK, Last P, Leis JM, Lessios H, Liebherr J, Lowman M, Mahler DL, Mamonekene V, Matsuura K, Mayer GC, Mays Jr H, McCosker J, McDiarmid RW, McGuire J, Miller MJ, Mooi R, Mooi RD, Moritz C, Myers P, Nachman MW, Nussbaum RA, Foighil DÓ, Parenti LR, Parham JF, Paul E, Paulay G, Pérez-Emán J, Pérez-Matus A, Poe S, Pogonoski J, Rabosky DL, Randall JE, Reimer JD, Robertson DR, Rödel M-O, Rodrigues MT, Roopnarine P, Rüber L, Ryan MJ, Sheldon F, Shinohara G, Short A, Simison WB, Smith-Vaniz WF, Springer VG, Stiassny M, Tello JG, Thompson CW, Trnski T, Tucker P, Valqui T, Vecchione M, Verheyen E, Wainwright PC, Wheeler TA, White WT, Will K, Williams JT, Williams G, Wilson EO, Winker K, Winterbottom R, Witt CC. (2014) Specimen collection: An essential tool. Science 344(6186): 814–815. 10.1126/science.344.6186.814 [DOI] [PubMed] [Google Scholar]
- Rocha-Ramírez A, Chávez-López R, Peláez-Rodríguez E. (2010) Ancinusjarocho (Isopoda: Sphaeromatidea: Ancinidae), a new species from the central Gulf of Mexico, Mexico. Zootaxa 2397(1): 61–68. 10.11646/zootaxa.2397.1.6 [DOI] [Google Scholar]
- Rodríguez A, Drake P, Arias AM. (1992) First records of Paracerceissculpta (Holmes, 1904) and Paradelladianae (Menzies, 1962) (Isopoda, Sphaeromatidae) at the Atlantic coast of Europe. Crustaceana 63(1): 94–97. 10.1163/156854092X00334 [DOI] [Google Scholar]
- Roletto J, Kimura S, Cosentino-Manning N, Berger R, Bradley R. (2014) Status and trends of the rocky intertidal community on the Farallon Islands. Monographs of the Western North American Naturalist 7(1): 260–275. 10.3398/042.007.0120 [DOI] [Google Scholar]
- Rotramel G. (1972) Iaiscalifornica and Sphaeromaquoyanum, two symbiotic isopods introduced to California (Isopoda, Janiridae and Sphaeromatidae). Crustaceana (Supplement 3): 193–197.
- Rotramel GL. (1975) Observations on the commensal relations of Iaiscalifornica (Richardson, 1904) and Sphaeromaquoyanum H. Milne Edwards, 1840 (Isopoda). Crustaceana 28(1): 247–256. 10.1163/156854075X01125 [DOI] [Google Scholar]
- Ruiz G, Fofonoff P, Steves B, Dahlstrom A. (2011) Marine crustacean invasions in North America: a synthesis of historical records and documented impacts. In: Galil BS, Clark PF, Carlton JT (Eds) In the Wrong Place – Alien Marine Crustaceans: Distribution, Biology and Impacts, Springer, Invading Nature 6: 215–250. 10.1007/978-94-007-0591-3_6 [DOI] [Google Scholar]
- Rumbold C, Meloni M, Doti B, Correa N, Albano M, Sylvester F, Obenat S. (2018) Two new nonindigenous isopods in the southwestern Atlantic: Simultaneous assessment of population status and shipping transport vector. Journal of Sea Research 138: 1–7. 10.1016/j.seares.2018.04.008 [DOI] [Google Scholar]
- Salazar-Vallejo SI, Leija-Tristán A. (1990) Progebiophilusbruscai n. sp., a new bopyrid isopod parasitic on the mud shrimp, Upogebiadawsoni Williams (Thalassinoidea), from the Gulf of California. Cahiers de Biologie Marine 30: 423–432. [Google Scholar]
- Santamaria CA, Mateos M, DeWitt TJ, Hurtado LA. (2016) Constrained body shape among highly genetically divergent allopatric lineages of the supralittoral isopod Ligiaoccidentalis (Oniscidea). Ecology and Evolution 6(5): 1537–1554. 10.1002/ece3.1984 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santos AM, Xavier R, Zenboudji S, Branco T, Branco M. (2011) Stenosomastephenseni sp. n. (Isopoda, Idoteidae), from the southwestern Mediterranean, with a note on the nomenclatural status of Synisoma Collinge, 1917. ZooKeys 141: 29–44. 10.3897/zookeys.141.1376 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sassaman C, Schultz GA, Garthwaite R. (1984) Host, synonymy, and parasitic incidence of Bopyrellacalmani (Richardson) from central California (Isopoda: Epicaridea: Bopyridae). Proceedings of the Biological Society of Washington 97(3): 645–654. [Google Scholar]
- SCAMIT (1994) A taxonomic listing of soft bottom macroinvertebrates from infaunal monitoring programs in the Southern California Bight, 1st edn. Southern California Association of Marine Invertebrate Taxonomists, San Pedro, 72 pp. https://www.scamit.org/publications [Google Scholar]
- SCAMIT (1996) A taxonomic listing of soft bottom macro- and megainvertebrates from infaunal and epibenthic monitoring programs in the Southern California Bight, 2nd edn. Southern California Association of Marine Invertebrate Taxonomists, San Pedro, 86 pp. https://www.scamit.org/publications [Google Scholar]
- SCAMIT (1998) A taxonomic listing of soft bottom macro- and megainvertebrates from infaunal and epibenthic monitoring programs in the Southern California Bight, 3rd edn. Southern California Association of Marine Invertebrate Taxonomists, San Pedro, 167 pp. https://www.scamit.org/publications [Google Scholar]
- SCAMIT (2001) A taxonomic listing of soft bottom macro- and megainvertebrates from infaunal and epibenthic monitoring programs in the Southern California Bight, 4th edn. Southern California Association of Marine Invertebrate Taxonomists, San Pedro, 192 pp. https://www.scamit.org/publications [Google Scholar]
- SCAMIT (2008) A taxonomic listing of benthic macro- and megainvertebrates from infaunal and epifaunal monitoring and research programs in the Southern California Bight, 5th edn. Cadien DB, Lovell LL (Eds) Southern California Association of Marine Invertebrate Taxonomists, Los Angeles, 210 pp. https://www.scamit.org/publications [Google Scholar]
- SCAMIT (2011) A taxonomic listing of benthic macro- and megainvertebrates from infaunal and epifaunal monitoring and research programs in the Southern California Bight, 6th edn. Cadien DB, Lovell LL (Eds) Southern California Association of Marine Invertebrate Taxonomists, Los Angeles, 211 pp. https://www.scamit.org/publications [Google Scholar]
- SCAMIT (2012) A taxonomic listing of benthic macro- and megainvertebrates from infaunal and epifaunal monitoring and research programs in the Southern California Bight, 7th edn. Cadien DB, Lovell LL (Eds) Southern California Association of Marine Invertebrate Taxonomists, Los Angeles, 137 pp. https://www.scamit.org/publications [Google Scholar]
- SCAMIT (2013) A taxonomic listing of benthic macro- and megainvertebrates from infaunal and epifaunal monitoring and research programs in the Southern California Bight, 8th edn. Cadien DB, Lovell LL (Eds) Southern California Association of Marine Invertebrate Taxonomists, Los Angeles, 143 pp. https://www.scamit.org/publications [Google Scholar]
- SCAMIT (2014) A taxonomic listing of benthic macro- and megainvertebrates from infaunal and epifaunal monitoring and research programs in the Southern California Bight, 9th edn. Cadien DB, Lovell LL (Eds) Southern California Association of Marine Invertebrate Taxonomists, Los Angeles, 166 pp. https://www.scamit.org/publications [Google Scholar]
- SCAMIT (2015) A taxonomic listing of benthic macro- and megainvertebrates from infaunal and epifaunal monitoring and research programs in the Southern California Bight, 10th edn. Cadien DB, Lovell LL (Eds) Southern California Association of Marine Invertebrate Taxonomists, Los Angeles, 171 pp. https://www.scamit.org/publications [Google Scholar]
- SCAMIT (2016) A taxonomic listing of benthic macro- and megainvertebrates from infaunal and epifaunal monitoring and research programs in the Southern California Bight, 11th edn. Cadien DB, Lovell LL (Eds) Southern California Association of Marine Invertebrate Taxonomists, Los Angeles, 171 pp. https://www.scamit.org/publications [Google Scholar]
- SCAMIT (2018) A taxonomic listing of benthic macro- and megainvertebrates from infaunal and epifaunal monitoring and research programs in the Southern California Bight, 12th edn. Cadien DB, Lovell LL, Barwick KL (Eds) Southern California Association of Marine Invertebrate Taxonomists, Los Angeles, 167 pp. https://www.scamit.org/publications [Google Scholar]
- SCAMIT (2021) A taxonomic listing of benthic macro- and megainvertebrates from infaunal and epifaunal monitoring and research programs in the Southern California Bight, 13th edn. Cadien DB, Lovell LL, Barwick KL, Haggin BM (Eds) Southern California Association of Marine Invertebrate Taxonomists, Los Angeles, 203 pp. https://www.scamit.org/publications [Google Scholar]
- Schiff K, Weisberg S, Raco-Rands V. (2002) Inventory of ocean monitoring in the Southern California Bight. Environmental Management 29(6): 871–876. 10.1007/s00267-001-2628-9 [DOI] [PubMed] [Google Scholar]
- Schiff K, Greenstein D, Dodder N, Gillett DJ. (2016) Southern California Bight regional monitoring. Regional Studies in Marine Science 4: 34–46. 10.1016/j.rsma.2015.09.003 [DOI] [Google Scholar]
- Schiff K, McLaughlin K, Moore S, Cao Y. (2019) Chapter 19 – Southern California Bight. In: Sheppard C. (Ed.) World Seas: An Environmental Evaluation (2nd edn.), Academic Press, 465–482. 10.1016/B978-0-12-805068-2.00023-1 [DOI]
- Schioedte JC, Meinert FW. (1879) Symbolae ad Monographiam Cymothoarum Crustaceorum Isopodum Familiae 1. Aegidae. Naturhistorisk Tidsskrift 3(12): 321–414. 10.5962/bhl.title.10300 [DOI] [Google Scholar]
- Schioedte JC, Meinert FW. (1881) Symbolae ad Monographiam Cymothoarum Crustaceorum Isopodum Familiae 2. Anilocridae. Naturhistorisk Tidsskrift 3(13): 1–166. [Google Scholar]
- Schioedte JC, Meinert FW. (1884) Symbolae ad Monographiam Cymothoarum Isopodum Familiae 4. Cymothoidae. Trib. II. Cymothoinae. Trib. III. Livonecinae. Naturhistorisk Tidsskrift 3(14): 221–454. [Google Scholar]
- Schmalfuss H. (2003) World catalog of terrestrial isopods (Isopoda: Oniscidea). Stuttgarter Beitrage zur Naturkunde, Serie A, Biologie 654: 1–341. [Google Scholar]
- Schmidt C. (2008) Phylogeny of the terrestrial Isopoda (Oniscidea): A review. Arthropod Systematics & Phylogeny 66(2): 191–226. [Google Scholar]
- Schmidt C, Leistikow A. (2004) Catalogue of genera of the terrestrial Isopoda (Crustacea: Isopoda: Oniscidea). Steenstrupia (Copenhagen) 28(1): 1–118. [Google Scholar]
- Schotte M. (2005) Discerceiskensleyi, n. sp., from Caribbean Columbia, the third species of the genus (Crustacea: Isopoda: Sphaeromatidae). Proceedings of the Biological Society of Washington 118(1): 48–54. 10.2988/0006-324X(2005)118[48:DKNSFC]2.0.CO;2 [DOI]
- Schultz GA. (1964) Some marine isopod crustaceans from off the southern California coast. Pacific Science 18(3): 307–314. http://hdl.handle.net/10125/5557 [Google Scholar]
- Schultz GA. (1966) Marine isopods of the submarine canyons of the southern California continental shelf. Allan Hancock Pacific Expeditions 27(4): 1–56. [Google Scholar]
- Schultz GA. (1969) How to Know the Marine Isopod Crustaceans. Wm. C. Brown Company Publishers, Dubuque, 359 pp. [Google Scholar]
- Schultz GA. (1970) A review of the species of the genus Tylos Latreille from the New World (Isopoda, Oniscoidea). Crustaceana 19(3): 297–305. 10.1163/156854070X00383 [DOI] [Google Scholar]
- Schultz GA. (1972) Gnathiasanctaecrucis nom. nov., substituted for the preoccupied name Gnathiahirsuta Schultz, 1966, from southern California (Isopoda, Gnathiidae). Crustaceana 23(1): 112. 10.1163/156854072X00110 [DOI] [Google Scholar]
- Schultz GA. (1973) Ancinus H. Milne Edwards in the New World (Isopoda, Flabellifera). Crustaceana 25(3): 267–275. 10.1163/156854073X00254 [DOI] [Google Scholar]
- Schultz GA. (1977) Anthurids from the west coast of North America, including a new species and three new genera (Crustacea, Isopoda). Proceedings of the Biological Society of Washington 90: 839–848. [Google Scholar]
- Schultz GA. (1984) Four species of Alloniscus Dana, 1854, from the west coast of North America and Hawaii (Isopoda, Oniscoidea). Crustaceana 47(2): 149–167. 10.1163/156854084X00388 [DOI] [Google Scholar]
- Sept DJ. (2002) The Beachcomber’s Guide to Seashore Life of California. Harbour Publishing, Madeira Park, 312 pp. [Google Scholar]
- Sept DJ. (2019) The New Beachcomber’s Guide to the Pacific Northwest. Harbour Publishing, Madeira Park, 416 pp. [Google Scholar]
- Serov PA, Wilson GDF. (1995) A review of the Stenetriidae (Crustacea: Isopoda: Asellota). Records of the Australian Museum 47(1): 39–82. 10.3853/j.0067-1975.47.1995.2 [DOI] [Google Scholar]
- Sfenthourakis S, Taiti S. (2015) Patterns of taxonomic diversity among terrestrial isopods. In: Taiti S, Hornung E, Štrus J, Bouchon D. (Eds) Trends in Terrestrial Isopod Biology.ZooKeys 515: 13–25. 10.3897/zookeys.515.9332 [DOI] [PMC free article] [PubMed]
- Shaughnessy F, Mulligan T, Kramer S, Kullmann S, Largier J. (2017) Baseline Characterization of Biodiversity and Target Species in Estuaries along the North Coast of California. Final Report (September 2017), 112 pp. 10.13140/RG.2.2.30944.94724 [DOI]
- Shearer GM. (1942) A study of marine isopods of the Coos Bay region. Master of Science Thesis, Oregon State College. Corvallis, 64 pp. [Google Scholar]
- Sheppard EM. (1933) Isopod Crustacea. Part I. The family Serolidae. Discovery Reports 7: 253–362. [Google Scholar]
- Sheppard EM. (1957) Isopod Crustacea. Part II. The suborder Valvifera. Families: Idoteidae, Pseudidotheidae and Xenarcturidae fam. n. with a supplement to isopod Crustacea, Part I. The family Serolidae. Discovery Reports 29: 141–198. 10.5962/bhl.part.12485 [DOI] [Google Scholar]
- Shiino SM. (1934) Bopyrids from Tanabe Bay II. Memoirs of the College of Science, Kyoto University, Series B 9 (4, Article 7): 257–287.
- Shiino SM. (1964) On three bopyrid isopods from California. Report of the Faculty of Fisheries. Perfectural University of Mie 5: 19–25. [Google Scholar]
- Shimomura M. (2008) Bathycopea (Isopoda: Sphaeromatidea: Ancinidae) from Japan, with descriptions of two new species and redescription of B.parallela Birstein. Zootaxa 1678(1): 25–49. 10.11646/zootaxa.1678.1.2 [DOI] [Google Scholar]
- Shimomura M, Mawatari SF. (2001) Munnidae from Japan (Crustacea: Isopoda: Asellota). Publications of the Seto Marine Biological Laboratory 39(2/3): 45–73. 10.5134/176302 [DOI]
- Shuster SM. (1987) Alternative reproductive behaviors: Three discrete male morphs in Paracerceissculpta, an intertidal isopod from the northern Gulf of California. Journal of Crustacean Biology 7(2): 318–327. 10.2307/1548612 [DOI] [Google Scholar]
- Shuster SM. (1992) The reproductive behaviour of α-, β-, and γ-male morphs in Paracerceissculpta, a marine isopod crustacean. Behaviour 121(3–4): 231–257. 10.1163/156853992X00381 [DOI] [Google Scholar]
- Shuster SM, Guthrie EE. (1999) Effects of temperature and food availability on adult body length in natural and laboratory populations of Paracerceissculpta (Holmes), a Gulf of California isopod. Journal of Experimental Marine Biology and Ecology 233(2): 269–284. 10.1016/S0022-0981(98)00133-6 [DOI] [Google Scholar]
- Shuster SM, Wade MJ. (1991a) Equal mating success among male reproductive strategies in a marine isopod. Nature 350(6319): 608–610. 10.1038/350608a0 [DOI] [Google Scholar]
- Shuster SM, Wade MJ. (1991b) Female copying and sexual selection in a marine isopod crustacean, Paracerceissculpta. Animal Behaviour 41(6): 1071–1078. 10.1016/S0003-3472(05)80645-1 [DOI] [Google Scholar]
- Siebenaller JF, Hessler RR. (1981) The genera of the Nannoniscidae (Isopoda, Asellota). Transactions of the San Diego Society of Natural History 19(16): 227–250. [Google Scholar]
- Silva CG, Gomes EL, Figueiredo NC, Silva GB. (2019) First record of the biodiversity of isopods in Katsuwonuspelamis from South America (NE Brazil). Journal of Dairy, Veterinary &. Animal Research 8(2): 102–105. [Google Scholar]
- Sirota L, Hovel KA. (2006) Simulated eelgrass Zosteramarina structural complexity: Effects of shoot length, shoot density, and surface area on the epifaunal community of San Diego Bay, California, USA. Marine Ecology Progress Series 326: 115–131. 10.3354/meps326115 [DOI] [Google Scholar]
- Smit NJ, Davies AJ. (2004) The curious life-style of the parasitic stages of gnathiid isopods. Advances in Parasitology 58: 289–391. 10.1016/S0065-308X(04)58005-3 [DOI] [PubMed] [Google Scholar]
- Smit NJ, Bruce NL, Hadfield KA. (2014) Global diversity of fish parasitic isopod crustaceans of the family Cymothoidae. International Journal for Parasitology. Parasites and Wildlife 3(2): 188–197. 10.1016/j.ijppaw.2014.03.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smit NJ, Bruce NL, Hadfield KA [Eds] (2019) Parasitic Crustacea. State of Knowledge and Future Trends. Zoological Monographs 3. Springer, Cham, 481 pp. 10.1007/978-3-030-17385-2 [DOI] [Google Scholar]
- Smith CR, Baco AR. (2003) Ecology of whale falls at the deep-sea floor. Oceanography and Marine Biology - an Annual Review 41: 311–354. [Google Scholar]
- Smith RW, Bergen M, Weisberg SB, Cadien D, Dalkey A, Montagne D, Stull JK, Velarde RG. (2001) Benthic response index for assessing infaunal communities on the mainland shelf of southern California. Ecological Applications 11(4): 1073–1087. 10.1890/1051-0761(2001)011[1073:BRIFAI]2.0.CO;2 [DOI]
- Smith RW, Ranasinghe JA, Weisberg SB, Montagne DE, Cadien DB, Mikel TK, Velarde RG, Dalkey A. (2003) Extending the southern California Benthic Response Index to assess benthic condition in bays. Technical Report 410. Southern California Coastal Water Research Project. Westminster, 36 pp. [+ 4 appendices] https://www.sccwrp.org/publications
- Stafford BE. (1913a) Studies in Laguna Beach Isopoda II. Journal of Entomology and Zoology 5(3): 161–172. [Google Scholar]
- Stafford BE. (1913b) Studies in Laguna Beach Isopoda IIB. Journal of Entomology and Zoology 5(4): 182–188. [Google Scholar]
- Stebbing TRR. (1900) On some crustaceans from the Falkland Islands, collected by Mr. Rupert Vallentin. Proceedings of the Zoological Society of London 1900: 517–568. 10.5962/bhl.title.10309 [DOI] [Google Scholar]
- Stebbing TRR. (1905) Report on the Isopoda collected by Professor Herdman, at Ceylon, in 1902. In: Herdman WA. Report to the Government of Ceylon on the Pearl Oyster Fisheries of the Gulf of Manaar, Supplemental Report 23: 1–64. [Google Scholar]
- Stebbins TD. (1988a) Observations and experiments on the natural history and behavior of the commensal isopod Colidotearostrata (Benedict, 1898) (Isopoda: Idoteidae). Journal of Crustacean Biology 8(4): 539–547. 10.2307/1548690 [DOI] [Google Scholar]
- Stebbins TD. (1988b) The role of sea urchins in mediating fish predation on a commensal isopod (Crustacea: Isopoda). Journal of Experimental Marine Biology and Ecology 124(2): 97–113. 10.1016/0022-0981(88)90114-1 [DOI] [Google Scholar]
- Stebbins TD. (1989) Population dynamics and reproductive biology of the commensal isopod Colidotearostrata (Crustacea: Isopoda: Idoteidae). Marine Biology 101(3): 329–337. 10.1007/BF00428129 [DOI] [Google Scholar]
- Stebbins TD. (2012a) Key and notes to California valviferan isopods (Crustacea, Isopoda, Valvifera). https://www.scamit.org/tools
- Stebbins TD. (2012b) California “Epicaridean” Isopods Superfamilies Bopyroidea and Cryptoniscoidea (Crustacea, Isopoda, Cymothoida). Presentation to Southern California Association of Marine Invertebrate Taxonomists, 13 February 2012. https://www.scamit.org/tools
- Stebbins TD. (2012c) California valviferan isopods (Crustacea, Isopoda, Valvifera). Presentation to Southern California Association of Marine Invertebrate Taxonomists, 13 February 2012. https://www.scamit.org/tools
- Stepien CA, Brusca RC. (1985) Nocturnal attacks on nearshore fishes in southern California by crustacean zooplankton. Marine Ecology Progress Series 25: 91–105. 10.3354/meps025091 [DOI] [Google Scholar]
- Stimpson W. (1857) The Crustacea and Echinodermata of the Pacific shores of North America. Boston Journal of Natural History 6: 503–513. 10.5962/bhl.title.59693 [DOI] [Google Scholar]
- Stimpson W. (1864) Descriptions of new species of marine invertebrates from Puget Sound. Proceedings of the Philadelphia Academy of Sciences 16: 153–161. [Google Scholar]
- Stuxberg A. (1875) Om Nord-Amerika’s oniscider. Ofversigt Kongliga Svenska Vetenskaps- Akademiens Handlingar 32(2): 43–63. [Google Scholar]
- Taiti S, Ferrara F. (1986) Taxonomic revision of the genus Littorophiloscia Hatch, 1947 (Crustacea, Isopoda, Oniscidea) with description of six new species. Journal of Natural History 20(6): 1347–1380. 10.1080/00222938600770911 [DOI] [Google Scholar]
- Taiti S, Ferrara F. (1989) New species and records of Armadilloniscus Uljanin 1875 (CrustaceaIsopodaOniscidea) from the coasts of the Indian and Pacific oceans. Tropical Zoology 2(1): 59–88. 10.1080/03946975.1989.10539427 [DOI] [Google Scholar]
- Talley TS, Crooks JA, Levin LA. (2001) Habitat utilization and alteration by the invasive burrowing isopod, Sphaeromaquoyanum, in California salt marshes. Marine Biology 138(3): 561–573. 10.1007/s002270000472 [DOI] [Google Scholar]
- Tanaka K. (2007) Life history of gnathiid isopods–current knowledge and future directions. Plankton & Benthos Research 2(1): 1–11. 10.3800/pbr.2.1 [DOI] [Google Scholar]
- Thistle D. (1979) A redescription of two species of Ilyarachna (Asellota: Janiroidea) from off California (Crustacea: Isopoda). Zoological Journal of the Linnean Society 67(4): 381–386. 10.1111/j.1096-3642.1979.tb01120.x [DOI] [Google Scholar]
- Thompson B, Tsukada D, O’Donohue D. (1993) Final Report 1990 Reference Site Survey. Technical Report 269. Southern California Coastal Water Research Project, Westminster, CA, 105 pp. https://www.sccwrp.org/publications [Google Scholar]
- Toft J, Cordell J. (2006) Olympic Sculpture Park: Results from Pre-construction Biological Monitoring of Shoreline Habitats. Technical Report SAFS-UW-0601, School of Aquatic and Fishery Sciences, University of Washington. Prepared for Seattle Public Utilities, City of Seattle, 36 pp. [Google Scholar]
- Toft J, Cordell J, Heerhartz S, Armbrust E, Ogston A, Flemer E. (2008) Olympic Sculpture Park: Results from Year 1 Post-construction Monitoring of Shoreline Habitats. Technical Report SAFS-UW-0801, School of Aquatic and Fishery Sciences, University of Washington. Prepared for Seattle Public Utilities, Seattle, 113 pp. [Google Scholar]
- Toft J, Ogston A, Heerhartz S, Cordell J, Armbrust E, Levy C. (2010) Olympic Sculpture Park: Year 3 Monitoring of Shoreline Enhancements. Technical Report SAFS-UW-1002, School of Aquatic and Fishery Sciences, University of Washington. Prepared for Seattle Public Utilities, Seattle, 110 pp. [Google Scholar]
- Trask T. (1971) Ancinusseticomvus, n. sp. (Isopoda), from Santa Barbara, California. Bulletin of the Southern California Academy of Sciences 69(3–4): 145–149. [Google Scholar]
- van der Wal S, Smit NJ, Hadfield KA. (2019) Review of the fish parasitic genus Elthusa Schioedte & Meinert, 1884 (Crustacea, Isopoda, Cymothoidae) from South Africa, including the description of three new species. ZooKeys 841: 1–37. 10.3897/zookeys.841.32364 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Name WG. (1936) The American land and fresh-water isopod Crustacea. Bulletin of the American Museum of Natural History 71: 1–535. [Google Scholar]
- Van Name WG. (1940) A supplement to the American land and fresh-water isopod Crustacea. Bulletin of the American Museum of Natural History 77: 109–142. [Google Scholar]
- Van Name WG. (1942) A second supplement to the American land and fresh-water isopod Crustacea. Bulletin of the American Museum of Natural History 80: 299–329. [Google Scholar]
- Vargas-Zamora AJ, Dean HK, Mauer D, Orellana P. (1985) Lista preliminar de invertebrados asociados a los sedimentos del Golfo de Nicoya, Costa Rica. Brenesia 24: 327–343. [Google Scholar]
- Wägele J-W. (1979) Morphologische Studien an Eisothistos mit Beschreibung von drei neuen Arten (Crustacea, Isopoda, Anthuridea). Mitteilungen aus dem Zoologischen Museum der Universitaet Kiel 1(2): 1–19. [Google Scholar]
- Wägele J-W. (1984a) Studies on Antarctic CrustaceaIsopoda. 1. Anthuridea from the Weddell Sea. Polar Biology 3: 99–117. 10.1007/BF00258154 [DOI] [Google Scholar]
- Wägele J-W. (1984b) Two new littoral Anthuridea from Baja California and redescription of Mesanthuraoccidentalis (Crustacea, Isopoda). Zoologica Scripta 13(1): 45–57. 10.1111/j.1463-6409.1984.tb00022.x [DOI] [Google Scholar]
- Wägele J-W. (1991) Antarctic IsopodaValvifera. Keys and notes for the identification of the species. In: Wägele J-W, Sieg J (Eds) Synopses of the Antarctic Benthos, Vol. 2. Koeltz Scientific Books, Koenigstein, 213 pp. [Google Scholar]
- Wägele J-W. (1994) Notes on Antarctic and South American Serolidae (Crustacea, Isopoda) with remarks on the phylogenetic biogeography and a description of new genera. Zoologische Jahrbücher. Abteilung für Systematik, Ökologie und Geographie der Tiere 121(1): 3–69. [Google Scholar]
- Walker AO. (1898) Crustacea collected by W.A. Herdman, F.R.S., in Puget Sound, Pacific coast of North America, September 1897. Transactions of the Liverpool Biological Society 12: 268–287. [Google Scholar]
- Wall A, Bruce NL, Wetzer R. (2015) Status of Exosphaeromaamplicauda (Stimpson, 1857), E.aphrodita (Boone, 1923) and description of three new species (Crustacea, Isopoda, Sphaeromatidae) from the north-eastern Pacific. ZooKeys 504: 11–58. 10.3897/zookeys.504.8049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walther SM, Williams JP, Latker AK, Cadien DB, Diehl DW, Wisenbaker K, Miller E, Gartman R, Stransky C, Schiff K. (2017) Southern California Bight 2013 Regional Monitoring Program: Vol. VII. Demersal Fishes and Megabenthic Invertebrates. Technical Report 972. Southern California Coastal Water Research Project, Costa Mesa, 58 pp [+ appendices] https://www.sccwrp.org/publications
- Wetzer R. (2015) Collecting and preserving marine and freshwater Isopoda (Crustacea: Peracarida). Biodiversity Data Journal 3: e4912. 10.3897/BDJ.3.e4912 [DOI] [PMC free article] [PubMed]
- Wetzer R, Bruce NL. (2007) A new species of Paradella Harrison & Holdich, 1982 (Crustacea: Isopoda: Sphaeromatidae) from Baja California, Mexico, with a key to East Pacific species. Zootaxa 1512(1): 39–49. 10.11646/zootaxa.1512.1.2 [DOI] [Google Scholar]
- Wetzer R, Brusca RC. (1997) Descriptions of the species of the suborders Anthuridea, Epicaridea, Flabellifera, Gnathiidea, and Valvifera. In: Blake JA, Scott PH. (Eds) Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and Western Santa Barbara Channel, Vol.11: The Crustacea Part 2: The Isopoda, Cumacea and Tanaidacea, 9–58.
- Wetzer R, Mowery G. (2017) Redescription of Dynoideselegans (Boone, 1923) (Crustacea, Isopoda, Sphaeromatidae) from the north-eastern Pacific. ZooKeys 646: 1–16. 10.3897/zookeys.646.10626 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wetzer R, Kuck HG, Baéz R, Brusca RC, Jurkevics LM. (1991) Catalog of the isopod Crustacea type collection of the Natural History Museum of Los Angeles County. Technical Reports No. 3, Natural History Museum of Los Angeles County, Los Angeles, 59 pp. [Google Scholar]
- Wetzer R, Brusca RC, Wilson GDF. (1997) Introduction to the marine Isopoda. In: Blake JA, Scott PH. (Eds) Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and Western Santa Barbara Channel, Vol.11: The Crustacea Part 2: The Isopoda, Cumacea and Tanaidacea, 1–8.
- Wetzer R, Pérez-Losada M, Bruce NL. (2013) Phylogenetic relationships of the family Sphaeromatidae Latreille, 1825 (Crustacea: Peracarida: Isopoda) within Sphaeromatidea based on 18S-rDNA molecular data. Zootaxa 3599(2): 161–177. 10.11646/zootaxa.3599.2.3 [DOI] [PubMed] [Google Scholar]
- Wetzer R, Bruce NL, Pérez-Losada M. (2018) Relationships of the Sphaeromatidae genera (Peracarida: Isopoda) based on 18SrDNA and 16SrDNA genes. Arthropod Systematics & Phylogeny 76: 1–30. http://hdl.handle.net/10394/26937 [Google Scholar]
- Wetzer R, Wall A, Bruce NL. (2021) Redescription of Gnorimosphaeromaoregonense (Dana, 1853) (Crustacea, Isopoda, Sphaeromatidae), designation of neotype, and 16s-rDNA molecular phylogeny of the north-eastern Pacific species. ZooKeys 1037: 23–56. 10.3897/zookeys.1037.63017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whalen MA, Millard-Martin BR, Cox KD, Lemay MA, Paulay G. (2020) Poleward range expansion of invasive bopyrid isopod, Orthionegriffenis Markham, 2004, confirmed by establishment in Central British Columbia, Canada. BioInvasions Records 9(3): 538–548. 10.3391/bir.2020.9.3.09 [DOI] [Google Scholar]
- Whitcraft CR, Levin LA, Talley D, Crooks JA. (2008) Utilization of invasive tamarisk by salt marsh consumers. Oecologia 158(2): 259–272. 10.1007/s00442-008-1144-5 [DOI] [PubMed] [Google Scholar]
- Whitmore RC, Brusca RC, de la Luz JLL, González-Zamorano P, Mendoza-Salgado R, Amador-Silva ES, Holguin G, Galvan-Magana F, Hastings PA, Cartron JE, Felger RS, Seminoff JA, McIvor CC. (2005) The ecological importance of mangroves in Baja California Sur: conservation implications for an endangered ecosystem. Chapter 15 In: Cartron JE, Ceballos G, Felger RS. (Eds) Biodiversity, Ecosystems, and Conservation in Northern Mexico.Oxford Press, New York, 298–333.
- Williams JD, An J. (2009) The cryptogenic parasitic isopod Orthionegriffenis Markham, 2004 from the eastern and western Pacific. Integrative and Comparative Biology 49(2): 114–126. 10.1093/icb/icp021 [DOI] [PubMed] [Google Scholar]
- Williams JD, Boyko CB. (2012) The global diversity of parasitic isopods associated with crustacean hosts (Isopoda: Bopyroidea and Cryptoniscoidea). PLoS ONE 7(4): e35350. 10.1371/journal.pone.0035350 [DOI] [PMC free article] [PubMed]
- Williams JD, Boyko CB. (2021) Out on a limb: novel morphology and position on appendages of two genera and three new species of ectoparasitic isopods (Epicaridea: Dajidae) infesting isopod and decapod hosts. Zoosystema 43(4): 79–100. 10.5252/zoosystema2021v43a4 [DOI] [Google Scholar]
- Williams Jr EH, Bunkley-Williams L. (2003) New records of fish-parasitic isopods (Cymothoidae) in the Eastern Pacific (Galápagos and Costa Rica). Noticias de Galápagos 62: 21–23. http://hdl.handle.net/1834/25255 [Google Scholar]
- Williams Jr EH, Williams LB. (1980) Four new species of Renocila (Isopoda: Cymothoidae), the first reported from the New World. Proceedings of the Biological Society of Washington 93(3): 573–592. [Google Scholar]
- Williams LB, Williams Jr EH. (1987) Three new species of Renocila (Crustacea: Isopoda: Cymothoidae), external parasites of coral reef fishes from the Ryukyu Islands of Japan. Proceedings of the Biological Society of Washington 100(3): 417–432. [Google Scholar]
- Williams JD, Boyko CB, Madad AZ. (2019) Branchial parasitic isopods (Crustacea: Isopoda: Bopyridae: Pseudioninae) of hermit crabs (Crustacea: Decapoda: Diogenidae) from the western Pacific, with descriptions of a new genus and three new species. The Raffles Bulletin of Zoology 67: 83–118. 10.1080/00222937600770451 [DOI] [Google Scholar]
- Wilson GD. (1976) The systematics and evolution of Haplomunna and its relatives (Isopoda, Haplomunnidae, New family). Journal of Natural History 10(5): 569–580. 10.1080/00222937600770451 [DOI] [Google Scholar]
- Wilson GD. (1980) New insights into the colonization of the deep sea: Systematics and zoogeography of the Munnidae and the Pleurogoniidae comb. nov. (Isopoda; Janiroidea). Journal of Natural History 14(2): 215–236. 10.1080/00222938000770201 [DOI] [Google Scholar]
- Wilson GDF. (1987) The road to the Janiroidea: Comparative morphology and evolution of the assellote isopod crustaceans. Journal of Zoological Systematics and Evolutionary Research 25(4): 257–280. 10.1111/j.1439-0469.1987.tb00608.x [DOI] [Google Scholar]
- Wilson GDF. (1994) A phylogenetic analysis of the isopod family Janiridae (Crustacea). Invertebrate Taxonomy 8(3): 749–766. 10.1071/IT9940749 [DOI] [Google Scholar]
- Wilson GDF. (1997) The suborder Asellota. In: Blake JA, Scott PH. (Eds) Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and Western Santa Barbara Channel, Vol.11: The Crustacea Part 2: The Isopoda, Cumacea and Tanaidacea, 59–120.
- Wilson GDF. (2008a) Global diversity of isopod crustaceans (Crustacea; Isopoda) in freshwater. Hydrobiologia 595(1): 231–240. 10.1007/s10750-007-9019-z [DOI] [Google Scholar]
- Wilson GDF. (2008b) A review of taxonomic concepts in the Nannoniscidae (Isopoda, Asellota), with a key to the genera and a description of Nannoniscusoblongus Sars. Zootaxa 1680(1): 1–24. 10.11646/zootaxa.1680.1.1 [DOI] [Google Scholar]
- Wilson GD, Hessler RR. (1980) Taxonomic characters in the morphology of the genus Eurycope (Isopoda, Asellota) with a redescription of Eurycopecornuta GO Sars, 1864. Cahiers de Biologie Marine 21(3): 241–263. [Google Scholar]
- Wilson GD, Hessler RR. (1981) A revision of the genus Eurycope (Isopoda, Asellota) with descriptions of three new genera. Journal of Crustacean Biology 1(3): 401–423. 10.2307/1547972 [DOI] [Google Scholar]
- Wilson GDF, Wägele J-W. (1994) Review of the family Janiridae (Crustacea: Isopoda: Asellota). Invertebrate Taxonomy 8(3): 683–747. 10.1071/IT9940683 [DOI] [Google Scholar]
- Wilson GDF, Sims CA, Grutter AS. (2011) Toward a taxonomy of the Gnathiidae (Isopoda) using juveniles. The external anatomy of Gnathiaaureamaculosa zuphea stages using scanning electron microscopy. Journal of Crustacean Biology 31(3): 509–522. 10.1651/10-3432.1 [DOI] [Google Scholar]
- Wisenbaker K, McLaughlin K, Diehl D, Latker A, Stolzenbach K, Gartman R, Schiff K. (2021) Southern California Bight 2018 Regional Monitoring Program: Vol. IV. Demersal Fishes and Megabenthic Invertebrates. Technical Report 1183. Southern California Coastal Water Research Project. Costa Mesa, 213 pp. https://www.sccwrp.org/publications
- Wolff T. (1962) The systematics and biology of bathyal and abyssal IsopodaAsellota. Galathea Report 6: 1–320. [Google Scholar]
- Wright JC, Harris ZC. (2020) Aerial and aquatic respiration in littoral Oniscidea (Isopoda) from Southern California, USA. Journal of Crustacean Biology 40(4): 462–470. 10.1093/jcbiol/ruaa023 [DOI] [Google Scholar]
- Xavier R, Santos AM, Harris DJ, Sezgin M, Machado M, Branco M. (2012) Phylogenetic analysis of the north-east Atlantic and Mediterranean species of the genus Stenosoma (Isopoda, Valvifera, Idoteidae). Zoologica Scripta 41(4): 386–399. 10.1111/j.1463-6409.2012.00537.x [DOI] [Google Scholar]
- Zúñiga Delgado JG, Torres Conde EG, Hinostroza Miranda JD. (2021) Systematic list of marine isopods of Peru, first record of Isocladusbahamondei Carvacho, 1997 (Crustacea, Peracarida) and contributions of its ecology for the Peruvian coast. Revista de Investigaciones Marinas 41(2): 22–35. http://hdl.handle.net/1834/41821 [Google Scholar]