

Abstract
Species co-existence depends on how organisms utilize their environment and resources. Little is known about the winter diet composition and sympatric co-existence of South China sika deer and its companion species in Taohongling. In this study, high-throughput sequencing and metabarcoding trnL were used to study the diet composition and interspecific relationship including sika deer, Reeve’s muntjac, and Chinese hare. Our results show that 203 genera in 90 families are contained in the diet of sika deer, 203 genera in 95 families for Reeve’s muntjac, and 163 genera in 75 families for Chinese hare. Sika deer fed on Rubuschingii, Loropetalumchinense, and Euryajaponica in winter, accounting for 75.30%; Reeve’s muntjac consumed mainly R.chingii, E.japonica, and Euonymusgrandiflorus, accounting for 68.80%, and Chinese hare mainly fed on R.chingii, Smilaxchina, and Rhuschinensis, accounting for 41.98%. The Shannon index showed no significant difference between groups (p > 0.05). The NMDS analysis found considerable overlap among three species. Sika deer and Reeve’s muntjac consumed similar forage plants but varied greatly in Chinese hare, which occupied the widest choice in winter, resulting in higher diet breadth and increased dietary divergence, thereby reducing competition and facilitating coexistence. The diet niche overlap index among them, as represented by Pianka’s index, ranging from 0.62 between sika deer and Chinese hare to 0.83 between sika deer and Reeve’s muntjac, which indicated a more similar niche and potential competition in closely related species. Our findings provide a new diet perspective of three herbivores, leading to a more comprehensive understanding of resource partitioning and species coexistence.
Keywords: Diet composition, niche breadth, niche overlap, sympatry, winter
Introduction
Sika deer (Cervusnippon Temminck, 1838), also known as the spotted deer, is a species native to much of East Asia and a national first-class protected wild animal in China (Yao et al. 2010). Its conservation status is Endangered globally and Endangered in China (Wang and Song 2013). Most wild sika deer populations in China have disappeared due to heavy hunting pressure that has existed for a very long period (Zhang et al. 2011a), and populations have also become gradually more isolated. South China sika deer (Cervusnipponkopschi Swinhoe, 1873) are mainly distributed in the Taohongling Sika deer National Nature Reserve (hereafter, TNNR) in northeastern Jiangxi Province, southern Anhui Province, and a part of northwestern Zhejiang Province. Statistics have previously shown that the population of sika deer in TNNR is 365 individuals (Gao et al. 2009; Jiang et al. 2012) which inhabit the hilly area at 300–500 m elevation (Wang 2018). Reeve’s muntjac and Chinese hare are the main companion species of sika deer, and they have co-existed in the TNNR for numerous generations. Infrared-camera detection has been used to determine that the relative abundance of the Reeve’s muntjac was 0.4160, which is significantly higher than that of the sika deer population (0.0411), and the relative abundance of the Chinese hare was 0.0138. Furthermore, the spatial distribution of both sika deer and Reeve’s muntjac is primarily concentrated in the core area (Zhou 2019). Nowadays, little is known about the diets and sympatric co-existence of these three herbivores, particularly in winter when their food resources are scarcer.
Diet analysis is one of the core contents of studying the habitat requirements of animals (Liu 2009; Hoenig et al. 2022). Food not only provides the necessary energy and nutrients for life activities but also reflects the trophic niche of the species in the biome (Lu et al. 2020). Therefore, diet analysis can serve to understand a species’ access to resources and habitat distribution to facilitate population conservation and recovery of endangered species. The study of diet mainly includes stomach contents analysis, indirect utilization methods, direct tracking observation, microscopic fecal analysis, and DNA metabarcoding analysis (Monro 1982; Zheng and Bao 2004; Lu et al. 2020). Stomach contents analysis is more accurate for identifying food resources but collecting stomach contents requires sacrificing animals (Fujii et al. 2019). Utilization methods and tracking observations are difficult to observe and may be influenced by subjective factors (Gong et al. 2022). Fecal microscopic analysis can be quantitative (Zhang et al. 2011b) but requires accurate identification of taxa from partially digested plant fragments and likely over-emphasizes less digestible components of the diet (Holechek 1982). For the endangered species, it is necessary to prioritize noninvasive sampling. Diet research must adopt proven selection methods based on actual needs and conditions.
High-throughput sequencing (HTS) has the advantages of high throughput, a large amount of data, high sensitivity, and fine classification (Pompanon et al. 2012; Deagle et al. 2019). Compared with conventional methods of diet research, the metabarcoding method based on high-throughput sequencing can improve the deficiencies of the traditional methods that do not fully reflect consumers’ diet information (Ma et al. 2021; Tabassum et al. 2022). The present technique has achieved remarkable results in diet research on filter-feeding shellfish (Kasim and Mukai 2009), small herbivores, fish, etc. (Soininen et al. 2009; Lin et al. 2018; Li et al. 2021), and this can identify species of lower taxonomic orders.
Food resources are the medium that connects the natural environment and often influence the distribution and survival of species. Species coexistence theory suggests that niche overlap and potential competition will inevitably occur when closely related species with similar ecological needs share the same area, which requires them to obtain more resources to survive by expanding the niche scale (Schaller 2000; Palmer and Truscott 2003). We predict that the three species in this study have been sympatric and have evolved together for numerous generations in the TNNR. Natural selection may have led to a separation in forage use (niche differentiation) among them (Pascual-Rico et al. 2020). Fitness may be reduced by competition; i.e., sika deer may increase their niche breadth, particularly in winter when food resources are scarcer (Schoener 1971). We explore the diet composition and dietary overlap of Taohongling Sika deer, Reeve’s muntjac, and Chinese hare and assess the extent of potential dietary competition among these species to enhance our understanding of mechanisms underlying their coexistence. Research into the diet of sika deer and its sympatric herbivores can clarify food items and explore the interspecific competition and coexistence, which is of great significance to the population conservation of sika deer and biodiversity monitoring.
Methods
Study area
This study was conducted in the TNNR, the area where South China sika deer is distributed. The TNNR is located on the south bank of the middle and lower reaches of the Yangtze River, Pengze, Jiangxi Province, China. The total area of TNNR is 12,500 hm2, the core area is 2,670 hm2, the experimental area is 1,830 hm2, and the buffer zone is 8,000 hm2. The TNNR mainly consists of low mountains and hills (Wang et al. 2021). The TNNR lies in a climatic zone transitional from tropical to middle subtropical and has transitional climate characteristics that are warm, with a humid monsoon, and four distinct seasons. The frost-free period is up to 247 days with little snow cover (Zhou 2019). The vegetation type is mainly composed of mixed evergreen–deciduous broad-leaved forest, coniferous forest, mixed coniferous–broad-leaved forest, broad-leaved forest, and bamboo (Zhou 2019). From December 2020 to February 2021, 90 fecal samples were collected from sika deer, Reeve’s muntjac, and Chinese hare in the TNNR and stored at −80 °C. Sampling sites mainly focused on Nursery bases (MP), Fir forests (SS), NieJiashan (NJS), XianLingAn (XLA), WuGuiShi (WGS), and Bamboo Garden (ZY) (Fig. 1).
Figure 1.

Fecal samples sites of three sympatric species at Taohongling Sika Deer Nature Reserve (MP: Nursery bases; SS: Fir forests; NJS: NieJiashan; XLA: XianLingAn; WGS: WuGuiShi; ZY: Bamboo Garden).
DNA extraction and PCR amplification
In our study, to minimize possible bias caused by variation in individual digestibility, five fecal pellets were randomly taken from each fecal sample and mixed to form a single composite sample. Total DNA was extracted using the DNA extraction kit (TIANGEN, Beijing) following the liquid nitrogen grinding method. The final DNA concentration and purification were determined by NanoDrop 2000 UV-vis spectrophotometer (Thermo Scientific, Wilmington, USA), and DNA quality was checked by 1% agarose gel electrophoresis. The P6 loop region of the trnL(UAA)intron region was amplified with universal primers g (5՚-GGGCAATC CTGAGCCAA-3՚) and h (5՚-CCATTGAGTCTCTGCACCTATC-3՚) by thermocycler PCR system (Gene Amp 9700, ABI, USA). PCR amplifications were carried out in a total volume of 25 μl containing 12.5 μl PCR mix (Tiangen, Beijing, China), 1 μl DNA, 1 μl of each primer, and 9.5 μl H2O. The reaction conditions were as follows: denaturation at 95 °C for 3 min followed by 35 cycles at 95 °C for 30 sec, 56 °C for 30 sec, and 72 °C for 45 sec, with a final 10 min at 72 °C and storage at 4 °C for 10 h. The PCR products were detected by Agarose gel electrophoresis and sequenced by Shanghai Personal Biotechnology Co., Ltd.
Illumina MiSeq sequencing and bioinformatics analysis
Purified amplicons were pooled in equimolar and paired end sequenced (2 × 300) on an Illumina MiSeq platform (Illumina, San Diego, USA) according to the standard protocols. The analysis was conducted by following the tutorial of QIIME2 docs along with customized program scripts (https://docs.qiime2.org/2019.1/). Briefly, raw FASTQ files were demultiplexed using the QIIME2 v. 2019.4 demux plugin based on their unique barcodes (Caporaso et al. 2010). Demultiplexed sequences from each sample were quality filtered and trimmed, denoised, and merged, and then the chimeric sequences were identified and removed using the QIIME2 dada2 plugin to obtain the feature table of operational taxonomic units (OTUs). The QIIME2 feature-classifier plugin was then used to align OTUs sequences to the National Center for Biotechnology Information (NCBI) database to generate the taxonomy table. Diversity metrics were calculated using the core-diversity plugin within QIIME2. Feature level alpha diversity indices, such as observed OTUs, Chao1 richness estimator, and Goods coverage index were calculated to estimate the diet diversity within an individual sample. A difference significance test was performed by R v. 4.1.3. Beta diversity distance measurements using Bray–Curtis were performed to investigate the structural variation of fecal plant communities across samples and then visualized via principal coordinate analysis (PCoA) and nonmetric multidimensional scaling (NMDS) (Vázquez-Baeza et al. 2013). Venn diagram analysis was performed to explore the common and special OTUs types among the three herbivores.
Data statistics
Formulas of forage plants diversity and niche analysis were conducted as follows:
The Shannon–Wiener diversity index (H′) was calculated to explore diet diversity (Shannon 1948), according to the following formula:
(1)
Where Pi is the proportion of food item i out of all foods, and n is the total number of food items.
Pielou evenness index (Pielou 1969), according to the following formula:
J′ = H′ / Hmax (2)
Hmax = ln n (3)
Where n is the number of plant species in the fecal sample, and the number of plant species is represented by the number of plant OTUs types.
The Levin index (Smith 1982) was applied to standardize the trophic niche measure, and the formula was as follows:
(4)
The niche overlap index was obtained using the Pianka index (Pianka 1973; Hou et al. 2021), and the formula was as follows:
(5)
Where Qjk is Pianka’s niche overlap index between species j and species k; Pij is the proportion of resource i out of all resources used by species j, and Pik is the proportion of resource i out of all resources used by species k. The values range from 0 (no food item in common) to 1 (complete overlap in resource use).
Results
OTUs analysis
After processing the raw reads, a total of 11,411,958 counts were obtained from 90 fecal samples. The mean OTUs length was 67.96 bp with a range from 32 bp to 189 bp. Venn diagram showed that the OTUs in the overlap were commonly shared, and those in the nonoverlapping parts were special OTUs. In total, 764 OTUs, 833 OTUs, and 843 OTUs were obtained from sika deer, Reeve’s muntjac, and Chinese hare samples, respectively. The number of OTUs among the three herbivore groups was 373, and the specific OTUs in sika deer, Reeve’s muntjac, and Chinese hare were 391, 449, and 470, respectively (Fig. 2).
Figure 2.
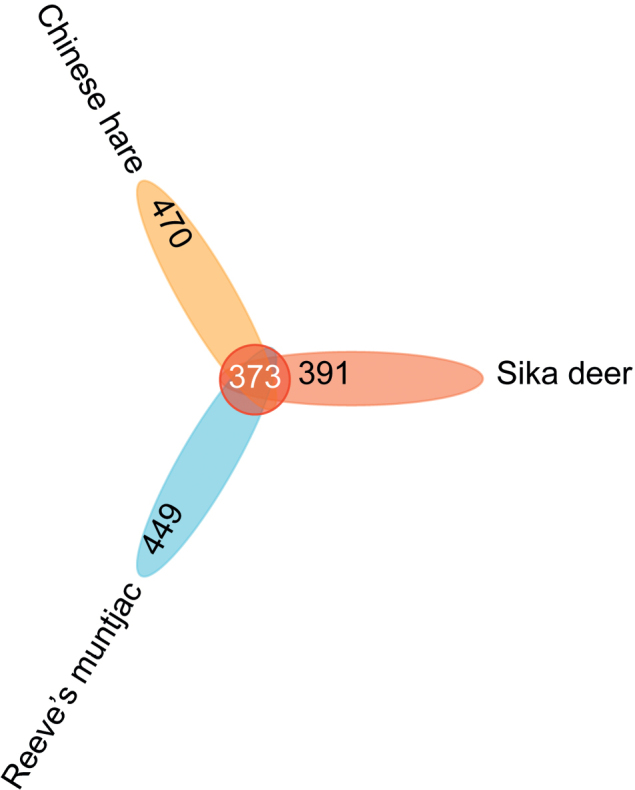
Venn analysis of OTUs in three herbivores of Taohongling nature reserve.
Based on OTUs sequences alignment in the NCBI database, the diets of sika deer, Reeve’s muntjac, and Chinese hare includes 203 genera in 90 families, 203 genera in 95 families, and 163 genera in 75 families, respectively (Editorial Committee of FRPS 2004; Jiang 2009; see Suppl. material 1). The three species consumed common and specific forage plants but varied greatly in their use of the available forages. On the whole, the 10 most abundant families in the diet of sika deer include Rosaceae (36.94%), Hamamelidaceae (25.75%), Pentaphylacaceae (13.41%), Theaceae (3.89%), Celastraceae (3.01%), Poaceae (2.98%), Ericaceae (2.43%), Moraceae (2.09%), Cupressaceae (1.68%), and Cannabaceae (1.16%). For the Reeve’s muntjac, the 10 most abundant families include Rosaceae (52.41%), Pentagliaceae (11.01%), Celastraceae (6.12%), Poaceae (4.52%), Cannabaceae (3.27%), Moraceae (2.64%), Sabiaceae (2.14%), Asteraceae (2.10%), Oleaceae (1.50%), and Smilacaceae (1.26%), and the 10 most abundant families in the Chinese hare consist of Rosaceae (16.35%), Poaceae (16.33%), Smilacaceae (15.58%), Anacardiaceae (10.71%), Fabaceae (9.74%), Asteraceae (6.77%), Rubiaceae (5.56%), and Cupressaceae (4.49%).
The dominant genera foraged by sika deer were Rubus (36.49%) and Loropetalum (25.52%), followed by Eurya (13.41%), Camellia (3.89%), Euonymus (2.94%), Phyllostachys (2.46%), Maclura (1.88%), Cunninghamia (1.67%), Rhododendron (1.45%), and Celtis (0.99%), and others (9.30%). For Reeve’s muntjac the diet was strongly dominated by Rubus (51.77%), other genera high abundance were Eurya (11.01%), Euonymus (6.11%), Celtis (3.21%), Arrhenatherum (3.02%), Sabia (2.14%), Maclura (1.67%), Ligustrum (1.49%), Phyllostachys (1.31%), and Smilax (1.26%). Chinese hare consumed almost equal proportions of Rubus (15.81%) and Smilax (15.58%), followed by Rhus (10.64%), Campylotropis (9.52%), Bidens (5.37%), Hedyotis (5.02%), Cunninghamia (4.47%), Eleusine (3.57%), Digitaria (3.09%) and Miscanthus (2.55%). To sum up, these three herbivores all mostly feed on Rubus in winter. The species composition heatmap was drawn from the species and sample levels. The 20 genera with the highest abundance were selected based on species annotation information for 90 samples of the three species. The clustering results show the differences in the relative abundances of sika deer, Chinese hare, and Reeve’s muntjac (Fig. 3).
Figure 3.

Species composition heat map at the genus level. (Ch: Chinese hare; Sd: Sika deer; Rm: Reeve’s muntjac).
High-throughput sequencing can be used to detect the diet on species levels in most samples combined with a background survey of TNNR, except for OTUs that were undetectable. Sika deer food items included Rubuschingii, Loropetalumchinense, Euryajaponica, Camelliajaponica, Euonymusgrandiflorus, etc. The forage plants of Reeve’s muntjac consisted of Rubuschingii, Euryajaponica, Euonymusgrandiflorus, Arrhenatherumelatius, Celtissinensis, etc. The diet of Chinese hares mainly focused on Rubuschingii, Smilaxchina, Rhuschinensis, Campylotropis sp., Hedyotisdiffusa, etc. The detailed top 30 forage species in three species are shown in Table 1.
Table 1.
Winter diet of sika deer, Reeve’s muntjac, and Chinese hare in Taohongling nature reserve. “+” = present in Jiangxi; “–” = not or uncertain present in Jiangxi.
| Number | Sika deer | Reeve’s muntjac | Chinese hare | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | Genus | Disrtibution | Percentage of abundance | Species | Genus | Distribution | Percentage of abundance | Species | Genus | Distribution | Percentage of abundance | |
| 1 | Rubuschingii | Rubus | + | 36.42% | Rubuschingii | Rubus | + | 51.69% | Rubuschingii | Rubus | + | 15.78% |
| 2 | Loropetalumchinense | Loropetalum | + | 25.48% | Euryajaponica | Eurya | + | 11.01% | Smilaxchina | Smilax | + | 15.56% |
| 3 | Euryajaponica | Eurya | + | 13.41% | Euonymusgrandiflorus | Euonymus | + | 6.10% | Rhuschinensis | Rhus | + | 10.64% |
| 4 | Camelliajaponica | Camellia | + | 3.88% | Arrhenatherumelatius | Arrhenatherum | + | 3.02% | Campylotropis sp. | Campylotropis | – | 9.40% |
| 5 | Euonymusgrandiflorus | Euonymus | + | 2.94% | Celtissinensis | Celtis | + | 2.57% | Bidens sp. | Bidens | – | 5.37% |
| 6 | Phyllostachysedulis | Phyllostachys | + | 2.46% | Sabiaswinhoei | Sabia | + | 2.14% | Hedyotisdiffusa | Hedyotis | + | 5.02% |
| 7 | Macluratricuspidata | Maclura | + | 1.88% | Macluratricuspidata | Maclura | + | 1.66% | Cunninghamialanceolata | Cunninghamia | + | 4.47% |
| 8 | Cunninghamialanceolata | Cunninghamia | + | 1.67% | Ligustrumlucidum | Ligustrum | + | 1.48% | Eleusineindica | Eleusine | + | 3.57% |
| 9 | Rhododendronmucronatum | Rhododendron | + | 1.45% | Phyllostachysedulis | Phyllostachys | + | 1.31% | Digitaria sp. | Digitaria | – | 2.79% |
| 10 | Vacciniumvitis-idaea | Vaccinium | + | 0.77% | Smilaxchina | Smilax | + | 1.26% | Miscanthussinensis | Miscanthus | + | 2.55% |
| 11 | Celtissinensis | Celtis | + | 0.78% | Cunninghamialanceolata | Cunninghamia | + | 1.21% | Oxaliscorniculata | Oxalis | + | 2.31% |
| 12 | Ilexcornuta | Ilex | + | 0.59% | Nyssa sp. | Nyssa | – | 1.16% | Setariaviridis | Setaria | + | 1.56% |
| 13 | Sabiaswinhoei | Sabia | + | 0.48% | Aster sp. | Aster | – | 1.08% | Panicumbisulcatum | Panicum | + | 1.49% |
| 14 | Cocculusorbiculatus | Cocculus | + | 0.37% | Loropetalumchinense | Loropetalum | + | 0.83% | Pinusmassoniana | Pinus | + | 1.46% |
| 15 | Lysimachiacongestiflora | Lysimachia | + | 0.32% | Lysimachiacongestiflora | Lysimachia | + | 0.81% | Aster sp. | Aster | – | 1.18% |
| 16 | Saxifragastolonifera | Saxifraga | + | 0.30% | Oxaliscorniculata | Oxalis | + | 0.79% | Secalecereale | Secale | + | 1.16% |
| 17 | Arrhenatherumelatius | Arrhenatherum | + | 0.27% | Ilexcornuta | Ilex | + | 0.67% | Polypogonfugax | Polypogon | + | 1.00% |
| 18 | Ehretiaacuminata | Ehretia | + | 0.27% | Celtis sp. | Celtis | – | 0.64% | Phyllostachysedulis | Phyllostachys | + | 0.96% |
| 19 | Nyssa sp. | Nyssa | – | 0.22% | Cirsiumarvense | Cirsium | + | 0.62% | Sabiaswinhoei | Sabia | + | 0.96% |
| 20 | Hedyotisdiffusa | Hedyotis | + | 0.21% | Broussonetiapapyrifera | Broussonetia | + | 0.59% | Euryajaponica | Eurya | + | 0.73% |
| 21 | Rhuschinensis | Rhus | + | 0.22% | Saxifragastolonifera | Saxifraga | + | 0.44% | Mallotusjaponicus | Mallotus | + | 0.59% |
| 22 | Celtis sp. | Celtis | – | 0.20% | Rhododendronmucronatum | Rhododendron | + | 0.40% | Liquidambarformosana | Liquidambar | + | 0.55% |
| 23 | Corylopsismultiflora | Corylopsis | + | 0.20% | Mallotusjaponicus | Mallotus | + | 0.37% | Nicotianatabacum | Nicotiana | + | 0.55% |
| 24 | Polypogonfugax | Polypogon | + | 0.19% | Tetradiumruticarpum | Tetradium | + | 0.32% | Loropetalumchinense | Loropetalum | + | 0.49% |
| 25 | Abeliaschumannii | Abelia | + | 0.18% | Galiumaparine | Galium | + | 0.32% | Prunussibirica | Prunus | – | 0.46% |
| 26 | Broussonetiapapyrifera | Broussonetia | + | 0.15% | Coreopsistinctoria | Coreopsis | + | 0.29% | Arrhenatherumelatius | Arrhenatherum | + | 0.39% |
| 27 | Vacciniumovalifolium | Vaccinium | – | 0.15% | Leptodermis sp. | Leptodermis | – | 0.25% | Ilexcornuta | Ilex | + | 0.38% |
| 28 | Pinusmassoniana | Pinus | + | 0.14% | Iryantherahostmannii | Iryanthera | – | 0.25% | Sargentodoxacuneata | Sargentodoxa | + | 0.32% |
| 29 | Pteroceltistatarinowii | Pteroceltis | + | 0.14% | Morusyunnanensis | Morus | + | 0.24% | Rhododendronmucronatum | Rhododendron | + | 0.29% |
| 30 | Oxaliscorniculata | Oxalis | + | 0.12% | Acorusgramineus | Acorus | + | 0.24% | Microstegiumvimineum | Microstegium | + | 0.28% |
| 31 | Others | 4.14% | Others | 6.24% | Others | 7.74% | ||||||
Diet diversity and interspecific niche analysis
Alpha diversity reflects the abundance and diversity of species communities. The Chao1 and Observed species indices showed the highest community richness was Reeve’s muntjac (Chao1 index; Reeve’s muntjac = 242.46, Sika deer = 236.52, Chinese hare = 192.03, on average). The Shannon and Simpson indices showed the highest community diversity was Chinese hare (Shannon index; Chinese hare = 2.36, Reeve’s muntjac = 2.21, Sika deer = 1.82, on average), with no significant differences (P > 0.05). The goods coverage of 0.998 indicated that an average of 99% of the species were annotated (Fig. 4a; Table 2). Rarefaction curves describe the increase in species diversity as the sample size increases. It is crucial to point out that the characterization of species diversity was considered very reliable since the depth of rarefaction applied (35,000) was found to be sufficiently satisfactory (e.g., rarefaction curves had already reached a plateau at ~35,000 sequences in all samples) (Fig. 4b). The rank abundance curve reflects the richness and evenness of species in the sample through the flatness. The evenness of community composition of sika deer and Reeve’s muntjac was higher, while the lowest of Chinese hare (Fig. 4c).
Figure 4.

a box-plot of the alpha diversity index. In each panel, the abscissa is the group, and the ordinate is the value of the corresponding alpha diversity index b sample rare faction curves c rank abundance curve. The abscissa is the sequence number of OTUs arranged according to the Abundance size. The ordinate is the abundance value of each OTU in this grouping by Log2 log transformation (Ch: Chinese hare; Sd: Sika deer; Rm: Reeve’s muntjac).
Table 2.
Alpha diversity index among three sympatric species including sika deer, Reeve’s muntjac, and Chinese hare.
| Sample ID | Sika deer | Reeve’s muntjac | Chinese hare | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chao1 | Goods _coverage | Observed _species | Pielou_e | Shannon | Simpson | Chao1 | Goods _coverage | Observed _species | Pielou_e | Shannon | Simpson | Chao1 | Goods _coverage | Observed _species | Pielou_e | Shannon | Simpson | |
| 001 | 280.705 | 0.998038 | 170.2 | 0.40159 | 2.97535 | 0.760442 | 221.216 | 0.998458 | 147.5 | 0.169581 | 1.22164 | 0.254821 | 234.755 | 0.998217 | 141.9 | 0.384791 | 2.75059 | 0.766172 |
| 002 | 229.404 | 0.998361 | 154.1 | 0.257889 | 1.87412 | 0.511389 | 223.198 | 0.998492 | 144.1 | 0.22161 | 1.58889 | 0.400989 | 144.924 | 0.998847 | 93.3 | 0.348889 | 2.28297 | 0.717446 |
| 003 | 226.705 | 0.998481 | 131.2 | 0.265175 | 1.86516 | 0.504481 | 148.447 | 0.998978 | 76.4 | 0.152733 | 0.955071 | 0.242638 | 122.406 | 0.999139 | 66.2 | 0.442545 | 2.67587 | 0.781497 |
| 004 | 83.0718 | 0.999378 | 47.5 | 0.04717 | 0.262596 | 0.050021 | 199.96 | 0.998608 | 139 | 0.4334 | 3.08469 | 0.822047 | 190.257 | 0.998583 | 97.2 | 0.342704 | 2.26105 | 0.678976 |
| 005 | 260.619 | 0.998344 | 140.5 | 0.41492 | 2.95988 | 0.745168 | 343.206 | 0.997691 | 213.2 | 0.565935 | 4.37752 | 0.917577 | 132.754 | 0.999174 | 64.7 | 0.442643 | 2.66008 | 0.786546 |
| 006 | 197.435 | 0.998546 | 118.2 | 0.265982 | 1.83103 | 0.542388 | 265.68 | 0.998131 | 178.1 | 0.247444 | 1.84993 | 0.420895 | 244.217 | 0.998111 | 141.9 | 0.27918 | 1.99433 | 0.632656 |
| 007 | 231.011 | 0.998546 | 109.6 | 0.321788 | 2.17979 | 0.606797 | 198.505 | 0.998824 | 126.7 | 0.148754 | 1.03889 | 0.208596 | 109.736 | 0.999253 | 59.4 | 0.34015 | 2.00346 | 0.532667 |
| 008 | 186.477 | 0.998679 | 128.3 | 0.132388 | 0.926968 | 0.19294 | 173.466 | 0.998816 | 119.8 | 0.095464 | 0.658982 | 0.127527 | 223.278 | 0.99833 | 109.7 | 0.32719 | 2.21691 | 0.679038 |
| 009 | 176.872 | 0.998759 | 121.7 | 0.227483 | 1.57562 | 0.515318 | 201.798 | 0.998665 | 123.3 | 0.357705 | 2.48433 | 0.745497 | 147.016 | 0.999049 | 62.2 | 0.32465 | 1.9334 | 0.602632 |
| 010 | 226.03 | 0.998381 | 138.1 | 0.243521 | 1.73098 | 0.492527 | 284.977 | 0.998219 | 170.6 | 0.336498 | 2.49463 | 0.590127 | 145.945 | 0.998992 | 83.5 | 0.320825 | 2.04777 | 0.5727 |
| 011 | 224.427 | 0.998378 | 150.5 | 0.177627 | 1.28473 | 0.291992 | 230.017 | 0.998401 | 151.9 | 0.307556 | 2.22853 | 0.589744 | 199.096 | 0.998449 | 125.3 | 0.333101 | 2.32102 | 0.750859 |
| 012 | 280.773 | 0.997847 | 183.2 | 0.340063 | 2.55598 | 0.716689 | 244.458 | 0.998393 | 151.8 | 0.297168 | 2.15298 | 0.502685 | 290.705 | 0.998009 | 142.6 | 0.418089 | 2.99021 | 0.757656 |
| 013 | 220.095 | 0.998438 | 135.6 | 0.242773 | 1.71921 | 0.485882 | 239.338 | 0.99831 | 135.8 | 0.292677 | 2.0731 | 0.64653 | 264.841 | 0.9981 | 152.8 | 0.411819 | 2.9874 | 0.809688 |
| 014 | 328.541 | 0.997802 | 209.9 | 0.343807 | 2.65167 | 0.673406 | 217.883 | 0.998549 | 146.6 | 0.32392 | 2.33042 | 0.568292 | 138.488 | 0.999117 | 76.9 | 0.42012 | 2.63072 | 0.749443 |
| 015 | 381.036 | 0.997089 | 237.8 | 0.336105 | 2.65258 | 0.723663 | 150.282 | 0.998963 | 80.2 | 0.218794 | 1.38347 | 0.43972 | 132.877 | 0.999191 | 76.6 | 0.198498 | 1.2422 | 0.305309 |
| 016 | 334.337 | 0.997563 | 191.8 | 0.413 | 3.13158 | 0.781254 | 277.335 | 0.997947 | 181.3 | 0.440708 | 3.30546 | 0.785065 | 120.473 | 0.999199 | 68.5 | 0.32578 | 1.9862 | 0.504604 |
| 017 | 207.833 | 0.998586 | 107.7 | 0.282636 | 1.90708 | 0.611331 | 348.937 | 0.997558 | 209 | 0.401727 | 3.09589 | 0.765458 | 87.2741 | 0.999438 | 43.5 | 0.3264 | 1.77444 | 0.551934 |
| 018 | 393.986 | 0.997237 | 210.8 | 0.292397 | 2.257 | 0.609557 | 274.614 | 0.998214 | 178.1 | 0.318902 | 2.38396 | 0.58144 | 445.677 | 0.997171 | 207.1 | 0.583434 | 4.48816 | 0.909631 |
| 019 | 233.482 | 0.998353 | 158.1 | 0.323043 | 2.35938 | 0.639344 | 260.417 | 0.998234 | 165 | 0.310687 | 2.28763 | 0.651905 | 167.976 | 0.998648 | 98.6 | 0.239998 | 1.58907 | 0.424112 |
| 020 | 170.192 | 0.998677 | 118.9 | 0.07857 | 0.541529 | 0.109537 | 210.012 | 0.9986 | 148.8 | 0.21039 | 1.51822 | 0.361145 | 155.937 | 0.998963 | 75.2 | 0.299848 | 1.86801 | 0.523298 |
| 021 | 198.276 | 0.998529 | 127.2 | 0.162252 | 1.134 | 0.299678 | 237.626 | 0.998549 | 147.7 | 0.32251 | 2.32356 | 0.61479 | 107.632 | 0.999233 | 60.8 | 0.419374 | 2.48264 | 0.744613 |
| 022 | 373.107 | 0.997535 | 211.1 | 0.400956 | 3.09591 | 0.787719 | 281.474 | 0.998185 | 143.1 | 0.355952 | 2.54866 | 0.668624 | 121.692 | 0.999142 | 64.6 | 0.25249 | 1.51731 | 0.452847 |
| 023 | 242.856 | 0.998137 | 145.9 | 0.195825 | 1.40746 | 0.387288 | 253.578 | 0.998359 | 153.8 | 0.294375 | 2.13846 | 0.59175 | 524.096 | 0.996277 | 257.2 | 0.531387 | 4.2544 | 0.873609 |
| 024 | 194.829 | 0.998489 | 123.5 | 0.202812 | 1.40898 | 0.463457 | 226.685 | 0.998458 | 137.9 | 0.414342 | 2.94376 | 0.747215 | 162.056 | 0.99889 | 105.8 | 0.138661 | 0.932367 | 0.218141 |
| 025 | 228.774 | 0.998424 | 157.8 | 0.381325 | 2.78406 | 0.712446 | 316.573 | 0.997734 | 187.1 | 0.35747 | 2.69716 | 0.728621 | 157.534 | 0.999094 | 76.6 | 0.459812 | 2.8768 | 0.781035 |
| 026 | 165.272 | 0.998938 | 108.6 | 0.256367 | 1.73316 | 0.499269 | 200.122 | 0.998572 | 119.8 | 0.269879 | 1.86281 | 0.449013 | 62.196 | 0.999548 | 45.3 | 0.047218 | 0.259645 | 0.051564 |
| 027 | 118.54 | 0.999171 | 73.7 | 0.150522 | 0.933483 | 0.232503 | 165.642 | 0.998935 | 115.2 | 0.224942 | 1.54008 | 0.352465 | 316.768 | 0.997776 | 171.4 | 0.550031 | 4.08039 | 0.905171 |
| 028 | 216.523 | 0.998398 | 133.9 | 0.089286 | 0.630779 | 0.129084 | 241.525 | 0.998131 | 156.7 | 0.35582 | 2.59425 | 0.723594 | 176.083 | 0.998725 | 101.4 | 0.270131 | 1.80004 | 0.452913 |
| 029 | 274.14 | 0.998097 | 171.2 | 0.226096 | 1.67727 | 0.435829 | 300.379 | 0.997947 | 184.8 | 0.326165 | 2.45546 | 0.660694 | 313.133 | 0.997844 | 172.6 | 0.458045 | 3.40363 | 0.799484 |
| 030 | 210.264 | 0.998441 | 152.1 | 0.089529 | 0.64893 | 0.128594 | 336.477 | 0.997683 | 197.7 | 0.353038 | 2.69256 | 0.689019 | 121.149 | 0.999105 | 81.2 | 0.424873 | 2.69476 | 0.734457 |
| Average | 236.520 | 0.998321 | 145.6 | 0.252097 | 1.82321 | 0.488000 | 242.461 | 0.998353 | 151.0 | 0.304205 | 2.21037 | 0.561616 | 192.032 | 0.998654 | 104.1 | 0.355422 | 2.36686 | 0.635023 |
We assessed the beta diversity using the Bray–Curtis distance. When the distance between the samples was smaller, the species-composition structure was more similar, and the PCoA diagram and NMDS analysis revealed the similarity of the composition of the diet between sika deer and Reeve’s muntjac (Fig. 5a, b), which was consistent with our expected results. In addition, a hierarchical clustering heat map was convenient for the intuitive identification of the species present in corresponding samples; Rubus had the highest abundance among the three species. The tree plot indicated that there was much similar distance in the samples between sika deer and Reeve’s muntjac but less similar with Chinese hare (Fig. 5c).
Figure 5.

aPCoA analysis chart, in which each point represents a sample bNMDS analysis chart. Diagram analysis with 95% confidence ellipse c hierarchical clustering diagram. Analysis of the hierarchical clustering tree diagram and the stacked bar diagram of the top 10 genera in abundance (Ch: Chinese hare; Sd: Sika deer; Rm: Reeve’s muntjac).
The intergroup difference analysis shows the difference between the intragroup and intergroup sample distances. Compared with Reeve’s muntjac, the intragroup distance of Reeve’s muntjac was smaller than the intergroup distance of sika deer and Chinese hare (Fig. 6a). While compared with sika deer, the intragroup distance of sika deer was slightly higher than the intergroup distance of Reeve’s muntjac (Fig. 6b). With different species composition, the difference between the intragroup should be smaller than the intergroup. In comparison with the NMDS, this phenomenon was speculated to be the fact that the diet composition of Reeve’s muntjac was much similar to those of sika deer, and the Reeve’s muntjac has more forage plant diversity.
Figure 6.

Intergroup difference analysis a shows the boxplots of the distances between samples in the sika deer group and the distances between samples in this group and samples in other groups b shows the boxplots of the distances between samples in Reeve’s muntjac group and the distances between samples in this group and samples in other groups.
By analyzing the niches of the three sympatric herbivorous animals, we found that the highest niche breadth was Chinese hare (~7.78), followed by sika deer (~4.53) and Reeve’s muntjac (~3.44). The niche overlap index between sika deer and Reeve’s muntjac was 0.83, sika deer and Chinese hare was 0.62, and Reeve’s muntjac and Chinese hare was 0.69. The overlap index ranges from 0 to 1, where 0 indicates that the food ranges do not overlap at all, and 1 indicates that the food ranges overlap entirely. Our results suggested that Reeve’s muntjac and sika deer have the highest diet overlap (Table 3).
Table 3.
The dietary niche overlap and Observed niche overlap index among the three sympatric species.
| Dietary niche breadth | Interspecific comparation | Observed niche overlap index | |
|---|---|---|---|
| Sika deer | 4.53 | Sika deer vs Reeve’s muntjac | 0.83 |
| Reeve’s muntjac | 3.44 | Sika deer vs Chinese hare | 0.62 |
| Chinese hare | 7.78 | Reeve’s muntjac vs Chinese hare | 0.69 |
| – | – | Sika deer vs Reeve’s muntjac vs Chinese hare | 0.68 |
Discussion
The food composition of the three herbivorous animals
Quantitative analysis is of great significance for the families, genera, and species of the herbivores’ diet. Liu (2012) investigated the diet of wild northeast sika deer in winter and found the sik deer feed on 35 plant species belonging to 25 genera in 17 families. Liu (2020) showed that 131 plants, including nine herbs, 31 shrubs, and eight trees, were foraged by South China sika deer in the Tianmu Mountains. One hundred and thirty-nine food items were identified in the feces of sika deer in our study. Comparative analysis shows that the metabarcoding method based on high-throughput sequencing provides more detailed forage plant information.
Yao et al. (2010) found that the sika deer in TNNR mainly eat Homonoiariparia, Fallopiamultiflora, Lespedezabicolor, Puerariamontana, and Viciafaba, and other species, and then especially fed on young leaves and shoots (Li et al. 2014). Compared with our study, the dominant forages in the feces of sika deer were Rubuschingii, Loropetalumchinense, Euryajaponica, Camelliajaponica, Euonymusgrandiflorus, and Phyllostachysedulis in winter; these are all Chinese medicinal herbs, which ensure the prevention of various diseases throughout the life cycle of sika deer (Yao et al. 2010; Ye 2015). The differences may be attributed to geographical and seasonal differences. The vegetation condition varied according to the different regions, which means that diverse forage plants are available for the sika deer.
In seasons when plant resources are scarce, sika deer will choose to eat non-favorable plants or the available food resources at the moment. Studies on Japanese sika deer showed that they mainly choose their favorite deciduous species from summer to autumn, such as Cornuscontroversa, and Quercus sp., but from early winter to spring, non-favored herbaceous and tree species, such as Juncusdecipiens and Cryptomeriajaponica, will be foraged (Nakahama et al. 2020). Therefore, sika deer have a diverse diet and feed on various plants in different seasons (Yao et al. 2010; Wang et al. 2019). In the TNNR, oaks included Quercusacutissima, Q.aliena, Q.chenii, Q.fabri, and Q.serrata (Jiang 2009). Significantly, oak leaves, which are rich in tannins and toxic to most mammals, including cattle (Doce et al. 2009), are conversely found to increase the reproductive rate and fawn survival rate of sika deer in captive breeding by some farmers (Xing et al. 2022). Oak leaves are essential for maintaining healthy sika deer in wild and farmed populations. However, we found a lower abundance in Quercus for three herbivorous mammals’ diets. We speculated this might be related to deciduous Quercus in winter or the gestation period of female sika deer, even though some studies claimed that tannins were not toxic to sika deer because of their rumen microbes and fermentation processes (Li et al. 2013). There are differences in the forage plants of South China sika deer in different seasons and regions.
In our study, we identified most of the forage species; however, someof the forage species that we identified were not previously known from Jiangxi Province. It may be difficult to identify all forage plants using a single gene fragment, and continuous succession of plant communities caused by invasive species adds a level complication to this. Therefore, it is necessary to add auxiliary barcodes as well as strengthen the overall investigation of potential food resources in the reserve. The construction of a database of plant species barcodes for the Taohongling Sika Deer Reserve would provide a reference and source of sequence alignments. Such as database would allow for a more accurate determination of the diets of herbivores and allow for better comparisons of the diets of sympatric herbivores.
Interspecific niches
Competition theory indicates that the greater the overlap of resources between species, the greater the competition coefficient because of the widespread use of niche overlap to estimate competition for resources (Colwell and Futuyma 1971). Tibetan red deer have a similar diet to sympatric ungulates, which inevitably leads to interspecific conflicts in food use (Lu et al. 2020). However, spatial-temporal variations in dietary consumption of the two dominant rodent species on Mount Kilimanjaro, Tanzania, have been found serve as a mechanism of resource portioning that enable these species to coexist with a niche overlap (Mulungu et al. 2011; Thomas et al. 2022); this contradicts the key assumption of competition theory. Thus, we cannot limit the research on interspecific competition in sympatric species to trophic ecological niches, and spatial-temporal dimensions should also be considered. For sika deer in the TNNR, we do not know the exact reasons for spatial variations in dietary overlaps with its two sympatric herbivores. We suggest that a cautious approach is required to interpret the high dietary overlaps and their implications for competitive interactions among the three studied herbivores in the TNNR. As pointed out by others (Belovsky 1986; Jenkins and Wright 1988; Gordon and Illius 1989), high dietary overlaps may not necessarily imply competition (Andersen et al. 2017) and may simply indicate that the food item is sufficient, permitting sympatric species to share resources. In the TNNR there are increasing populations of sympatric wild boar and Reeve’s muntjac. Sika deer face competition stress in space and food resources, especially when food is scarce in winter (Li et al. 2014). Thus, even when the determinants of competition mechanisms are uncertain, competition does exist and its important role among species cannot be denied.
Competition among sympatric species is mostly expressed as a compensatory mechanism in ecological niches when species are similar in one dimension, they differ on another. Food resources, habitat, and temporal partitioning are the most common dimension partitioned (Bagchi et al. 2003). For example, high dietary overlap among the species may result in niche differentiation (Reitz and Trumble 2002; Yin et al. 2007; Cao et al. 2009); large herbivores are forced to expand their food range to avoid competition during periods of food scarcity (Noor et al. 2013). In northeastern China, red deer tend to increase their browsing intensity to maintain their high food intake, but sika deer meet their relatively constant food intake and potential nutritional requirements by increasing their bite diameter in winter. This reflects the short-term foraging strategies by sharing similar foods with the sympatric ungulates (Zhong et al. 2020). Currently, we do not know the competition mechanism of sika deer and further studies are needed to determine the coexistence mechanism with its sympatric species.
In our study, we found the niche breadth of the sika deer was higher than the Reeve’s muntjac. Optimal forage theory suggests that preference and palatability will be selected for the animals in abundant food periods. While in a period of scarce food resources, feeding generalization will occur by selecting different forage plants (Belovsky 1978). It remains to be studied whether the higher dietary niche breadth of sika deer results from the physiological characteristics of digesting a wide range of foods or to avoid competition. Moreover, the niche breadth of Chinese hare was larger than sika deer and Reeve’s muntjac. We speculate that Chinese hare, as opportunistic feeders, have a broader range of forage plants but consume less due to their smaller body size. The diet composition of Chinese hares includes trees, shrubs, and herbs, but this may be due to a passive and random proximity foraging strategy or even the indirect ingestion from the process of grinding teeth.
Conclusions
The South China sika deer is the most endangered among the three remaining subspecies of sika deer in China. In our study, sika deer and Reeve’s muntjac showed a higher overlapping index of niche. Reeve’s muntjac may affect the survival of the sika deer due to the shortage of food resources in winter. We speculated that potential competition probably occurs in two cervid species. In addition, the growth of the secondary vegetation has accelerated in the reserve, and the decline of suitable habitats is a serious threat to the growth of the sika deer population. It is urgent to strengthen habitat management, improve habitat quality, and study forage plants. It is also necessary to provide food for sika deer and other wildlife through artificial planting during food shortages and dry seasons. Further studies need to establish local DNA databases to identify the forage plants and introduce the auxiliary barcoding to solve accurate species-level diet composition. Overall, our study determined the diet composition and interspecific niches of South China sika deer and its sympatric Reeve’s muntjac and Chinese hare. These result should be helpful to facilitate habitat improvements and artificial planting, monitor forage resources, and conserve biodiversity, and manage the reserve.
Acknowledgements
This research was supported by the Science and Technology Program of the Jiangxi Provincial Department of Education (GJJ180225) and the National Natural Science Foundation of China (31960118). We are grateful to Xiaohong Liu, Yongjiang Chen, and Yulu Chen of Taohongling Sika Deer National Nature Reserve for their help in sample collection, and Dr Hao Zang for the help with the R analysis.
Citation
Wang D, Hu X, Li M, Liu J, Tang M, Liu W, Zhan J, Xu Y, Zhang W (2023) Diet composition and interspecific niche of Taohongling Sika deer (Cervus nippon kopschi) and its sympatric Reeve’s muntjac (Muntiacus reevesi) and Chinese hare (Lepus sinensis) in winter (Animalia, Mammalia). ZooKeys 1149: 17–36. https://doi.org/10.3897/zookeys.1149.96936
Funding Statement
Science and Technology Program of the Jiangxi Provincial Department of Education and National Natural Science Foundation of China.
Contributor Information
Yongtao Xu, Email: ytxu666@jxau.edu.cn.
Weiwei Zhang, Email: zhangweiwei_nefu@163.com.
Supplementary materials
Dietary of Sika deer, Reeves՚ muntjac and Chinese hare
This dataset is made available under the Open Database License (http://opendatacommons.org/licenses/odbl/1.0/). The Open Database License (ODbL) is a license agreement intended to allow users to freely share, modify, and use this Dataset while maintaining this same freedom for others, provided that the original source and author(s) are credited.
Dandan Wang, Xiaolong Hu, Minling Li, Jie Liu, Ming Tang, Wuhua Liu, Jianwen Zhan, Yongtao Xu,Weiwei Zhang
Data type
data (excel document)
References
- Andersen GE, Johnson CN, Barmuta LA, Jones ME. (2017) Dietary partitioning of Australia’s two marsupial hypercarnivores, the Tasmanian devil and the spotted-tailed quoll, across their shared distributional range. PLoS ONE 12(11): e0188529. 10.1371/journal.pone.0188529 [DOI] [PMC free article] [PubMed]
- Bagchi S, Goyal SP, Sankar K. (2003) Niche relationships of an ungulate assemblage in a dry tropical forest. Journal of Mammalogy 84(3): 981–988. 10.1644/BBa-024 [DOI] [Google Scholar]
- Belovsky GE. (1978) Diet optimization in a generalist herbivore: The moose. Theoretical Population Biology 14(1): 105–134. 10.1016/0040-5809(78)90007-2 [DOI] [PubMed] [Google Scholar]
- Belovsky GE. (1986) Generalist herbivore foraging and its role in competitive interactions. American Zoologist 26(1): 51–69. 10.1093/icb/26.1.51 [DOI] [Google Scholar]
- Cao YF, Zhang TZ, Lian XM, Cui QH, Deng DD, Su JP. (2009) Diet overlap among selected ungulates in Kekexili region, Qinghai province. Sichuan Journal of Zoology 28: 49–54. [In Chinese] [Google Scholar]
- Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, Fierer N, Peña AG, Goodrich JK, Gordon JI, Huttley GA, Kelley ST, Knights D, Koenig JE, Ley RE, Lozupone CA, McDonald D, Muegge BD, Pirrung M, Reeder J, Sevinsky JR, Turnbaugh PJ, Walters WA, Widmann J, Yatsunenko T, Zaneveld J, Knight R. (2010) QIIME allows analysis of high-throughput community sequencing data. Nature Methods 7(5): 335–336. 10.1038/nmeth.f.303 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colwell RK, Futuyma DJ. (1971) On the measurement of niche breadth and overlap. Ecology 52(4): 567–576. 10.2307/1934144 [DOI] [PubMed] [Google Scholar]
- Deagle BE, Thomas AC, McInnes JC, Clarke LJ, Vesterinen EJ, Clare EL, Kartzinel TR, Eveson JP. (2019) Counting with DNA in metabarcoding studies: How should we convert sequence reads to dietary data? Molecular Ecology 28(2): 391–406. 10.1111/mec.14734 [DOI] [PMC free article] [PubMed]
- Doce RR, Hervás G, Belenguer A, Toral PG, Giráldez FJ, Frutos P. (2009) Effect of the administration of young oak (Quercuspyrenaica) leaves to cattle on ruminal fermentation. Animal Feed Science and Technology 150(1–2): 75–85. 10.1016/j.anifeedsci.2008.08.005 [DOI] [Google Scholar]
- Editorial Committee of FRPS (2004) Flora Reipublicae Popularis Sinicae (FRPS) (Vol. 1). Science Press, Beijing. [In Chinese]
- Fujii T, Ueno K, Minami M. (2019) Identification of food plants in the diet of Japanese ptarmigan (Lagopusmutusjaponicus) in the Japan’s Northern Japan Alps using DNA Barcoding. Wildlife and Human Society 6(2): 13–18. 10.20798/awhswhs.6.2_13 [DOI] [Google Scholar]
- Gao YM, Yu B, Wang JQ, Wu WG, Gao B. (2009) Habitat characteristics used by sika deer at Taohongling Natural Reserve. Jiangxi Science 27(6): 877–878. [+891] 10.13990/j.issn1001-3679.2009.06.012 [DOI] [Google Scholar]
- Gong S, Zhang P, Luan X, Bao H, Jiang GS. (2022) Application and comparison of Fecal microscopic analysis and DNA metabarcoding technology for dietary analysis of wild boar. Chinese Journal of Wildlife 43: 1–9. 10.12375/ysdwxb.20220308 [DOI] [Google Scholar]
- Gordon I, Illius AW. (1989) Resource partitioning by ungulates on the Isle of Rhum. Oecologia 79(3): 383–389. 10.1007/BF00384318 [DOI] [PubMed] [Google Scholar]
- Hoenig BD, Snider AM, Forsman AM, Hobson KA, Latta SC, Miller ET, Polito MJ, Powell LL, Rogers SL, Sherry TW, Toews DPL, Welch AJ, Taylor SS, Porter BA. (2022) Current methods and future directions in avian diet analysis. Ornithology 139(1): ukab077. 10.1093/ornithology/ukab077 [DOI]
- Holechek JL. (1982) Sample preparation techniques for microhistological analysis. Journal of Range Management Archives 35(2): 267–268. 10.2307/3898409 [DOI] [Google Scholar]
- Hou J, Li L, Wang YF, Wang WJ, Zhan HY, Dai NH, Lu P. (2021) Influences of submerged plant collapse on diet composition, breadth, and overlap among four crane species at Poyang Lake, China. Frontiers in Zoology 18(24): 1–17. 10.1186/s12983-021-00411-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jenkins KJ, Wright RG. (1988) Resource partitioning and competition among cervids in the northern Rocky Mountains. Journal of Applied Ecology 25(1): 11–24. 10.2307/2403606 [DOI] [Google Scholar]
- Jiang ZG. (2009) Study on the Biodiversity of Taohongling Sika Deer National Nature Reserve in Jiangxi. Tsinghua University Press, Beijing. [In Chinese]
- Jiang ZG, Xu XR, Liu WH, Li CL, Li CW, Lu XL, Xiao JP, Li YK, Tang SH, Ping XG, Li F, Luo ZH, Fang HX, Yu B, Zang JH, Chen QJ, Gao YM, Wu JD, Wu WG, Wang LB, Wu YF, Zu HB, Wang C, Dai J, Ying X, Wang JQ, Liu Z, Chen J, Li LL, Chen DQ, Zhang XW, Cui SP, Li J, Yuan FK, Zhang BB, Zhu JH, Gao HZ, Li HB, Chen YL, Chen YJ, Lin ZH, Wang Y, Zhang C, Zhou QH. (2012) Population status of South China sika deer in Taohongling National Nature Reserve. Chinese Journal of Wildlife 33(6): 305–308. [+332] 10.19711/j.cnki.issn2310-1490.2012.06.001 [DOI] [Google Scholar]
- Kasim M, Mukai H. (2009) Food sources of the oyster (Crassostreagigas) and the clam (Ruditapesphilippinarum) in the Akkeshi-ko estuary. Plankton & Benthos Research 4(3): 104–114. 10.3800/pbr.4.104 [DOI] [Google Scholar]
- Li ZP, Liu HL, Li GY, Bao K, Wang KY, Xu C, Feng Y, Yang YF, Wright ADG. (2013) Molecular diversity of rumen bacterial communities from tannin-rich and fiber-rich forage fed domestic sika deer (Cervusnippon) in China. BMC Microbiology 13(1): e151. 10.1186/1471-2180-13-151 [DOI] [PMC free article] [PubMed]
- Li J, Li YK, Liao LJ, Xie GY, Yuan FK. (2014) Habitat assessment of sika deer (Cevusnippon) in the Taohongling National Nature Reserve, Jiangxi Province, China. Acta Ecologica Sinica 34(5): 1274–1283. 10.5846/stxb201305171102 [DOI] [Google Scholar]
- Li FX, Du MR, Gao YP, Wang JW, Zhang YT, Zhang ZX, Jiang ZJ. (2021) Analysis of food sources of Crassostreagigas using high-throughput sequencing techniques. Yuye Kexue Jinzhan 42(5): 86–96. 10.19663/j.issn2095-9869.20200404001 [DOI] [Google Scholar]
- Lin XZ, Hu SM, Liu S, Huang H. (2018) Comparison between traditional sequencing and high throughput sequencing on the dietary analysis of juvenile fish. Chinese Journal of Applied Ecology 29(9): 3093–3101. 10.13287/j.1001-9332.201809.005 [DOI] [PubMed] [Google Scholar]
- Liu XP. (2009) Feeding habits and Food niche overlap of three Canids in Eastern Inner Mongolia. Master’s thesis, Qufu Normal University, Shandong, China. [In Chinese]
- Liu WS. (2012) Winter Diet Composition and Non-invasive Health Assessment of Feral Sika Deer. PhD thesis, Northeast Forestry University, Heilongjiang, China. [In Chinese]
- Liu Z. (2020) Sika Deer’s Feeding Habits Biochemical Analysis and Community Composition of Large and Medium-sized Mammals and Birds in Tianmu Mountains. Master’s thesis, China Jiliang University, Zhejiang, China. [In Chinese]
- Lu ZH, Zhang WQ, Liu H, Zhang HM, Li YR. (2020) Comparison on feeding habits of Cervuswallichii and sympatric ungulates and domestic animals in green grass period. Chinese Journal of Applied Ecology 31(2): 651–658. 10.13287/j.1001-9332.202002.002 [DOI] [PubMed] [Google Scholar]
- Ma J, Li KZ, Qiu DJ, Tan YH, Huang LM, Zhang JB. (2021) Dietary analysis of Flaccisagittaenflata based on high-throughput sequencing technology in Daya Bay. Ecologic Science 40(2): 9–17. 10.14108/j.cnki.1008-8873.2021.02.002 [DOI] [Google Scholar]
- Monro RH. (1982) An appraisal of some techniques used to investigate the feeding ecology of large herbivores with reference to a study on impala in the northern Transvaal. African Journal of Ecology 20(2): 71–80. 10.1111/j.1365-2028.1982.tb00277.x [DOI] [Google Scholar]
- Mulungu LS, Mahlaba TA, Massawe AW, Kennis J, Crauwels D, Eiseb S, Monadjem A, Makundi RH, Katakweba AAS, Leirs H, Belmain SR. (2011) Dietary differences of the multimammate mouse, Mastomysnatalensis (Smith, 1834), across different habitats and seasons in Tanzania and Swaziland. Wildlife Research 3(8): 640–646. 10.1071/WR11028 [DOI] [Google Scholar]
- Nakahama N, Furuta T, Ando H, Setsuko S, Takayanagi A, Isagi Y. (2020) DNA meta-barcoding revealed that sika deer foraging strategies vary with season in a forest with degraded understory vegetation. Forest Ecology and Management 484: e118637. 10.1016/j.foreco.2020.118637 [DOI]
- Noor A, Habib B, Kumar S. (2013) Habitat selection and niche segregation between chital and nilgai in Keoladeo National Park, India. European Journal Zoological Research 2: 1–9. [Google Scholar]
- Palmer SCF, Truscott AM. (2003) Seasonal habitat use and browsing by deer in Caledonian pinewoods. Forest Ecology and Management 174(1–3): 149–166. 10.1016/S0378-1127(02)00032-4 [DOI] [Google Scholar]
- Pascual-Rico R, Sánchez-Zapata JA, Navarro J, Eguía S, Anadón JD, Botella F. (2020) Ecological niche overlap between co-occurring native and exotic ungulates: Insights for a conservation conflict. Biological Invasions 22(8): 2497–2508. 10.1007/s10530-020-02265-x [DOI] [Google Scholar]
- Pianka ER. (1973) The structure of lizard communities. Annual Review of Ecology and Systematics 4(1): 53–74. 10.1146/annurev.es.04.110173.000413 [DOI] [Google Scholar]
- Pielou EC. (1969) An Introduction to Mathematical Ecology. Wiley-Interscience, New York. 10.1111/j.1558-5646.1970.tb01782.x [DOI]
- Pompanon F, Deagle BE, Symondson WOC, Brown DS, Jarman SN, Taberlet P. (2012) Who is eating what: Diet assessment using next generation sequencing. Molecular Ecology 21(8): 1931–1950. 10.1111/j.1365-294X.2011.05403.x [DOI] [PubMed] [Google Scholar]
- Reitz SR, Trumble JT. (2002) Competitive displacement among insects and arachnids. Annual Review of Entomology 47(1): 435–465. 10.1146/annurev.ento.47.091201.145227 [DOI] [PubMed] [Google Scholar]
- Schaller GB. (2000) Wildlife of the Tibetan Steppe. University of Chicago Press, Chicago. [DOI]
- Schoener TW. (1971) Theory of feeding strategies. Annual Review of Ecology and Systematics 2(1): 369–404. 10.1146/annurev.es.02.110171.002101 [DOI] [Google Scholar]
- Shannon CE. (1948) A mathematical theory of communication. The Bell Technical Journal 27(3): 379–423. 10.1002/j.1538-7305.1948.tb01338.x [DOI] [Google Scholar]
- Smith EP. (1982) Niche breadth, resource availability, and inference. Ecology 63(6): e1675. 10.2307/1940109 [DOI]
- Soininen EM, Valentini A, Coissac E, Miquel C, Gielly L, Brochmann C, Brysting AK, Sonstebo JH, Lms RA, Yoccoz NG, Taberlet P. (2009) Analysing diet of small herbivores: The efficiency of DNA barcoding coupled with high-throughput pyrosequencing for deciphering the composition of complex plant mixtures. Frontiers in Zoology 6(1): 1–16. 10.1186/1742-9994-6-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tabassum N, Lee JH, Lee SR, Kim JU, Park H, Kim HW, Kim JH. (2022) Molecular diet analysis of Adélie penguins (Pygoscelisadeliae) in the Ross Sea using fecal DNA. Biology 11(2): e182. 10.3390/biology11020182 [DOI] [PMC free article] [PubMed]
- Thomas SM, Soka GE, Mulungu LS, Makonda FBS. (2022) Spatial-temporal variations in dietary consumption of two dominant rodent species (Rhabdomysdilectus and Lophuromysacquilus) on Mount Kilimanjaro, Tanzania. Diversity 14(8): 659. 10.3390/d14080659 [DOI] [Google Scholar]
- Vázquez-Baeza Y, Pirrung M, Gonzalez A, Knight R. (2013) Emperor: A tool for visualizing highthroughput microbial community data. GigaScience 2(1): 1–16. 10.1186/2047-217X-2-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang L. (2018) The countermeasures and suggestions of improving the Cervusnippon habitat in Jiangxi Taohongling National Nature Reserve. The Journal of Hebei Forestry Science and Technology 1: 68–70. 10.16449/j.cnki.issn1002-3356.2018.01.020 [DOI] [Google Scholar]
- Wang LC, Song XZ. (2013) The Species Eco-Development Status of Chinese Sika Deer. China Animal Agriculture Association. China Deer Industry Development, Zhanjiang, 39–46. [In Chinese]
- Wang ZY, Meng DH, Luo Y, Teng LW, Gao H, Wang JF, Liu ZS. (2019) Spring food-habits of red deer (Cervuselaphusalashanicus) in Helan Mountains, China. Chinese Journal of Wildlife 40(4): 825–831. 10.19711/j.cnki.issn2310-1490.2019.04.002 [DOI] [Google Scholar]
- Wang XF, Zhong YF, Zhan JW, Liu WH, Wu WG, Gao YM, Zhang C, Shao RQ, Cao KQ, Li YK. (2021) Analysis on activity rhythm of Lophuranycthemera population in Taohongling Sika Deer National Nature Reserve based on infrared camera technology. Modern Agricultural Science and Technology 19: 182–184. [+194] 10.5846/stxb201805251148 [DOI] [Google Scholar]
- Xing XM, Ai C, Wang TJ, Li Y, Liu HT, Hu PF, Wang GW, Liu HM, Wang HL, Zhang RR, Zheng JJ, Wang XB, Wang L, Chang YX, Qian Q, Yu JH, Tang LX, Wu SG, Shao XJ, Li A, Cui P, Zhan W, Zhao S, Wu ZC, Shao XQ, Dong YM, Rong M, Tan YH, Cui XZ, Chang SZ, Song XC, Yang TG, Sun LM, Ju Y, Zhao P, Fan HH, Liu Y, Wang XH, Yang WY, Yang M, Wei T, Song SS, Xu JP, Yue ZG, Liang QQ, Li CY, Ruan J, Yang F. (2022) The First High-Quality Reference Genome of Sika Deer Provides Insights for High-Tannin Adaptation. Genomics, Proteomics & Bioinformatics. 10.1016/j.gpb.2022.05.008 [DOI] [PMC free article] [PubMed]
- Yao ZS, Xu XR, Xu P. (2010) The Analysis of Food Plants of Sika Deer in Taohongling Nature Reserve, Jiangxi Province. Proceedings and Abstracts of the 9th National Symposium on Natural Medicinal Material Resources, Guangzhou (China), July 2010, 348–353. [In Chinese]
- Ye ZW. (2015) Pharmacology of Chinese Medicine. Chongqing University Press, Chongqing, 10 pp. [In Chinese] [Google Scholar]
- Yin BF, Huai HY, Zhang YL, Zhou L, Wei WH. (2007) Trophic niches of Pantholopshodgsoni, Procaprapicticaudata and Equuskiang in Kekexili region. Chinese Journal of Ecology 18: 766–770. [In Chinese] [PubMed] [Google Scholar]
- Zhang LH, Piao HX, Jin Y. (2011a) Research progress in evolution of Chinese sika deer population. Agricultural Science Journal of Yanbian University 33(1): 73–76. 10.13478/j.cnki.jasyu.2011.01.016 [DOI] [Google Scholar]
- Zhang CY, Wu JP, Huang XY. (2011b) Evaluation of scatological analysis of sika deer food habits. Chinese Journal of Wildlife 32(4): 199–202. 10.19711/j.cnki.issn2310-1490.2011.04.005 [DOI] [Google Scholar]
- Zheng RQ, Bao YX. (2004) Study methods and procedures for ungulate food habits. Acta Ecologica Sinica 7: 1532–1539. 10.13287/j.1001-9332.202002.002 [DOI] [Google Scholar]
- Zhong LQ, Zhi XL, Sun Y, Liu XX, Sun B, Zhou SC, Zhang WQ, Zhang MH. (2020) Winter foraging of sympatric red deer and sika deer in northeast China: Diet composition, forage selection, bite diameter and browse intensity. Journal of Forest Research 25(4): 276–284. 10.1080/13416979.2020.1762025 [DOI] [Google Scholar]
- Zhou YX. (2019) Diversity of mammals and birds, ungulates activity rhythm and habitat selection by Camera Trapping in the Taohongling. Master’s thesis, Jiangxi Normal University, Jiangxi, China. [In Chinese]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Dietary of Sika deer, Reeves՚ muntjac and Chinese hare
This dataset is made available under the Open Database License (http://opendatacommons.org/licenses/odbl/1.0/). The Open Database License (ODbL) is a license agreement intended to allow users to freely share, modify, and use this Dataset while maintaining this same freedom for others, provided that the original source and author(s) are credited.
Dandan Wang, Xiaolong Hu, Minling Li, Jie Liu, Ming Tang, Wuhua Liu, Jianwen Zhan, Yongtao Xu,Weiwei Zhang
Data type
data (excel document)