

Abstract
Pregnancy is energetically demanding and, therefore, by necessity reproduction and energy balance are inextricably linked. With insufficient or excessive energy stores a female is liable to suffer complications during pregnancy or produce unhealthy offspring. Gonadotropin releasing hormone neurons are responsible for initiating both the pulsatile and subsequent surge release of luteinizing hormone to control ovulation. Meticulous work has identified two hypothalamic populations of kisspeptin (Kiss1) neurons that are critical for this pattern of release. The involvement of the hypothalamus is unsurprising as its quintessential function is to couple the endocrine and nervous systems, coordinating energy balance and reproduction. Estrogens, more specifically 17β-estradiol (E2), orchestrate the activity of a triumvirate of hypothalamic neurons within the arcuate nucleus (ARH) that govern the physiological underpinnings of these behavioral dynamics. Arising from a common progenitor pool, these cells differentiate into ARH kisspeptin, proopiomelanocortin (POMC), and agouti related peptide/neuropeptide Y (AgRP) neurons. Although the excitability of all these subpopulations is subject to genomic and rapid estrogenic regulation, Kiss1 neurons are the most sensitive, reflecting their integral function in female fertility. Based on the premise that E2 coordinates autonomic functions around reproduction, we will review recent findings on how Kiss1 neurons interact with GnRH, AgRP, and POMC neurons as well as how the rapid membrane-initiated and intracellular signaling cascades activated by E2 in these neurons are critical for control of homeostatic functions supporting reproduction. In particular, we will highlight how Kiss1 and POMC neurons conspire to inhibit AgRP neurons and diminish food motivation in service of reproductive success.
Keywords: Hypothalamus, Kisspeptin neurons, Neuropeptide Y, agouti-related peptide, proopiomelanocortin
1. Estrogenic regulation of reproduction through kisspeptin neurons
Successful reproduction is the core measure of evolutionary fitness and the quintessential function of the hypothalamus is to link the nervous and endocrine systems to support this physiological process. Within the brain, gonadotropin-releasing hormone (GnRH) neurons drive pubertal development and regulate the reproductive cycle. Postnatal GnRH neurons predominantly reside in the medial septum, diagonal band, and preoptic area (POA) of rodents (1–3), but are also found in the basal hypothalamus of sheep (2), guinea pigs (4), and primates including humans (5, 6). From these regions projections are sent to the median eminence to secrete GnRH in a pulsatile fashion, stimulating anterior pituitary gonadotrophs to release luteinizing hormone (LH) and follicle stimulating hormone (FSH). In females, LH and FSH in turn cause the synthesis and release of estrogen and progesterone from the ovaries. Following puberty, these ovarian hormones provide the requisite negative and positive feedback to maintain a normal cycle (7–10). This process is not linear, but rather relies on precisely timed GnRH pulses preceding a final surge to elicit LH release which causes ovulation and stimulates ovarian steroidogenesis. Significant effort has been expended elucidating the neural circuits involved and how estrogens regulate their activity to orchestrate the female reproductive cycle, which will be summarized in this section.
Classically, estrogenic signaling is viewed through estradiol (E2) activation of primarily ERα (11, 12), but also ERβ (13) receptors. Found in the cytosol, ERα or ERβ bind with estrogens and dimerize prior to translocation to the nucleus. Through interactions with estrogen response elements (EREs) present in certain gene promotors, estrogens can regulate transcription (14–16). In addition to cytosolic ERα and ERβ, E2 can activate membranebound estrogen receptors (mERs) to mediate rapid non-genomic actions (17, 18), though certain second messenger cascades such as phosphorylation of cAMP response element-binding protein can ultimately lead to alterations in gene expression as well (19–23). While ERα and ERβ can act as mERs, there are also the G-protein coupled estrogen receptor (GPER/GPR30/GPER1) (24, 25) and an as-yet unidentified Gq-coupled receptor (Gq-mER) (26, 27). These estrogen signaling pathways frequently co-exist and can produce divergent outcomes between cell types. For a more detailed overview of estrogen signaling cascades in hypothalamic neurons, please refer to Stincic et al., 2018 (28).
Despite the clear synchronization of GnRH neuronal activity with circulating estrogen levels (29), GnRH neurons are devoid of ERα (30–32). This revelation prompted a search for either estrogen signaling via alternative pathways or extrinsic synaptic inputs. Though ERβ is found in GnRH neurons (33–38), deletion of ERβ in either the whole animal or conditionally in GnRH neurons typically only impairs rather than eliminates fertility (39–41) by reducing the amplitude without affecting the timing of LH release (42, 43). The role of rapid estrogenic signaling is less clear but may assist in maintaining the GnRH surge for the requisite duration (44). So, while non-ERα signaling pathways contribute to GnRH release, mounting evidence has supported the existence of an extrinsic pulse generator.
Kisspeptin (Kiss1), transcribed from the Kiss1 gene, is a neuropeptide primarily produced by two populations of hypothalamic neurons found in the arcuate nucleus (ARH) and anteroventral periventricular/periventricular nuclei (AVPV/PeN) (45–49). Kiss1, also referred to as Kisspeptin-54, is the endogenous ligand of G protein-coupled receptor 54 (GPR54, aka Kiss1R) (50). Kiss1 neurons and the Kiss1Rs are indispensable for normal pubertal development (51–54) and fertility (55–57). Centrally administered kisspeptin robustly stimulates GnRH and gonadotropin secretion in both pre-pubertal and adult animals (45 , 58). Mutations in GPR54 cause autosomal recessive idiopathic hypogonadism in humans and deletion of GPR54 or Kiss1 in mice results in defective sexual development and reproductive failure (51, 53). GnRH neurons robustly express Gpr54 mRNA (59), and conditional deletion of the gene produces offspring that do not progress through puberty, have reduced gonadal size and are infertile (60). Underscoring the importance of Kiss1 signaling on GnRH function, the infertility phenotype in global GPR54 knockouts can be rescued by reintroducing the receptor solely in GnRH neurons (60). Kiss1 neurons express high levels of ERα and ERβ in both the AVPV/PeN (31, 55, 61) and the ARH (62). Therefore, estrogenic regulation of Kiss1 signaling could communicate the reproductive state of an animal to GnRH neurons.
GnRH neurons are strongly depolarized by kisspeptins. In vitro brief kisspeptin bath application produces a long-lasting depolarization in most adult GnRH neurons, regardless of sex (63–67). However, the responsiveness of GnRH neurons is much smaller in juvenile and prepubertal male mice despite a near constant level of Kiss1R mRNA expression across development (63). Based on cell signaling studies, kisspeptin excites GnRH neurons primarily through activation of canonical transient receptor potential 4 (TRPC4) channels (66, 68–70) and to a lesser extent through inhibition of inwardly rectifying K+ channels (66–68, 71, 72). TRPC channels can be activated by G-protein coupled receptors (73, 74) or participate in store-operated calcium entry (75). Trpc4, Hcn1, Cav1.3, Cav 2.2, and Cav 2.3 mRNA expression are all increased (59), which would presumably enhance responsiveness to kisspeptin. These findings demonstrate that E2 and kisspeptin signaling can drive GnRH/LH release but do not reveal how the pulses or surge are generated.
One of the more striking features of estrogenic regulation of Kiss1 neurons is the divergent effects on the ARH and AVPV/PeN populations. Kiss1ARH neurons co-express and release neurokinin B (NKB) and dynorphin (Dyn) (76–78), leading to their nickname of “KNDy neurons” (79). When circulating levels of E2 are high, Kiss1ARH neurons display enhanced expression of Vglut2 while Kiss1 mRNA expression is attenuated, biasing neurotransmission from peptidergic to glutamatergic signaling (80). Kiss1ARH neurons form a reciprocal network with other Kiss1ARH neurons (80) as well as sending projections to Kiss1AVPV/PeN neurons and GnRH “dendrons” and terminals in the median eminence (3, 81–84). Because Kiss1AVPV/PeN neurons do not express TACR3s (NKB receptor) or Kiss1Rs (85, 86), Kiss1ARH inputs to the AVPV/PeN population are essentially silent when E2 levels are low (80). However, as E2 levels peak in the lead up to proestrus and the LH surge, synchronization of Kiss1ARH neurons will exert an excitatory glutamatergic input to both Kiss1AVPV/PeN neurons and distal processes of GnRH neurons (69, 80, 87, 88). In this manner, the Kiss1ARH neurons can act as the pulse generator while driving up Kiss1AVPV/PeN activity that later initiates the LH surge.
The AVPV/PeN expresses high levels of ERα and ERβ, and the actions of the gonadal steroid hormones on Kiss1 neurons are mediated, at least in part, via nuclear-initiated signaling (transcriptional) mechanisms (31, 55, 61). For example, following E2 treatment Kiss1 mRNA expression is upregulated in the AVPV/PeN (47). Furthermore, E2 treatment enhances the expression of Vgat and tyrosine hydroxylase to, respectively, support GABAergic and dopaminergic signaling in Kiss1AVPV/PeN neurons (89, 90). Proestrus levels of E2 also elevate Kiss1AVPV/PeN excitability through positive regulation of currents such as the h-, T-type calcium and a persistent sodium current (91–93) (95). Together with previous observations that in rodents lesions of the AVPV/PeN or ER antagonist implants in the region abrogate the positive feedback effects of E2 (96–99) many have hypothesized that E2 acts on Kiss1AVPV/PeN neurons to induce the positive feedback on GnRH and LH secretion. More recently, experiments have shown that high frequency optogenetic stimulation of Kiss1AVPV/PeN neurons evokes kisspeptin release that activates TRPC4 channels in order to depolarize and excite GnRH neurons (87). However, despite the necessity of Kiss1AVPV/PeN neurons for the GnRH/LH surge (55, 87, 100, 101), these neurons are not involved in the pulsatile release of LH. Rather multi-unit recordings point to a different origin of pulse generator activity (83, 102–105).
Though the ARH was long speculated to be the origin of patterned activity, only relatively recently were Kiss1ARH neurons identified as the responsible neuronal subpopulation (81, 87, 106, 107). In vivo optogenetic stimulation of Kiss1ARH neurons proved capable of eliciting pulsatile GnRH and, subsequently, LH release in the mouse (106, 108). In vitro recordings, however, were necessary to reveal the neurocircuitry required to synchronize ARH Kiss1 activity (Figure 1) (87). To summarize, E2 levels are low prior to onset of LH pulses at which time Kiss1ARH neurons produce and co-release the neuropeptides NKB and dynorphin. Kiss1ARH neurons form a network of excitatory reciprocal connections through NKB release. Dynorphin then presynaptically inhibits further release, reducing the activity of all networked Kiss1ARH neurons (87). This sequence repeats, giving rise to oscillations in neural activity. Next, as E2 levels rise in advance of ovulation, expression of NKB, dynorphin, and kisspeptin genes falls (47, 79, 81) as Vglut2 mRNA expression and glutamate release probability dramatically rise (80). This causes a progression from peptidergic to amino acid transmission and affects not only inputs from Kiss1ARH to Kiss1AVPV/PeN neurons, but also to other ARH neurons (see below). The Kiss1ARH glutamatergic excitation of Kiss1AVPV/PeN neurons also contributes to kisspeptin-mediated excitation of GnRH neurons in the preovulatory state (87, 109). Furthermore, the precise timing of these events is synchronized to circadian rhythms through vasopressin (AVP) projections from the suprachiasmiatic nucleus to the Kiss1AVPV/PeN neurons such that ovulation occurs immediately prior to activity onset (110). Kiss1 AVPV/PeN and ARH neurons make reciprocal connections (3), which would explain why silencing of Kiss1ARH neurons causes a diurnal shift in eating patterns and obesity (111). Finally, although Kiss1AVPV/PeN neurons are responsible for the LH surge, Kiss1ARH neurons can amplify the surge when E2 is low or paired with progesterone (109).
Figure 1. Estradiol governs signaling between Kiss1 and GnRH neurons to drive LH release.
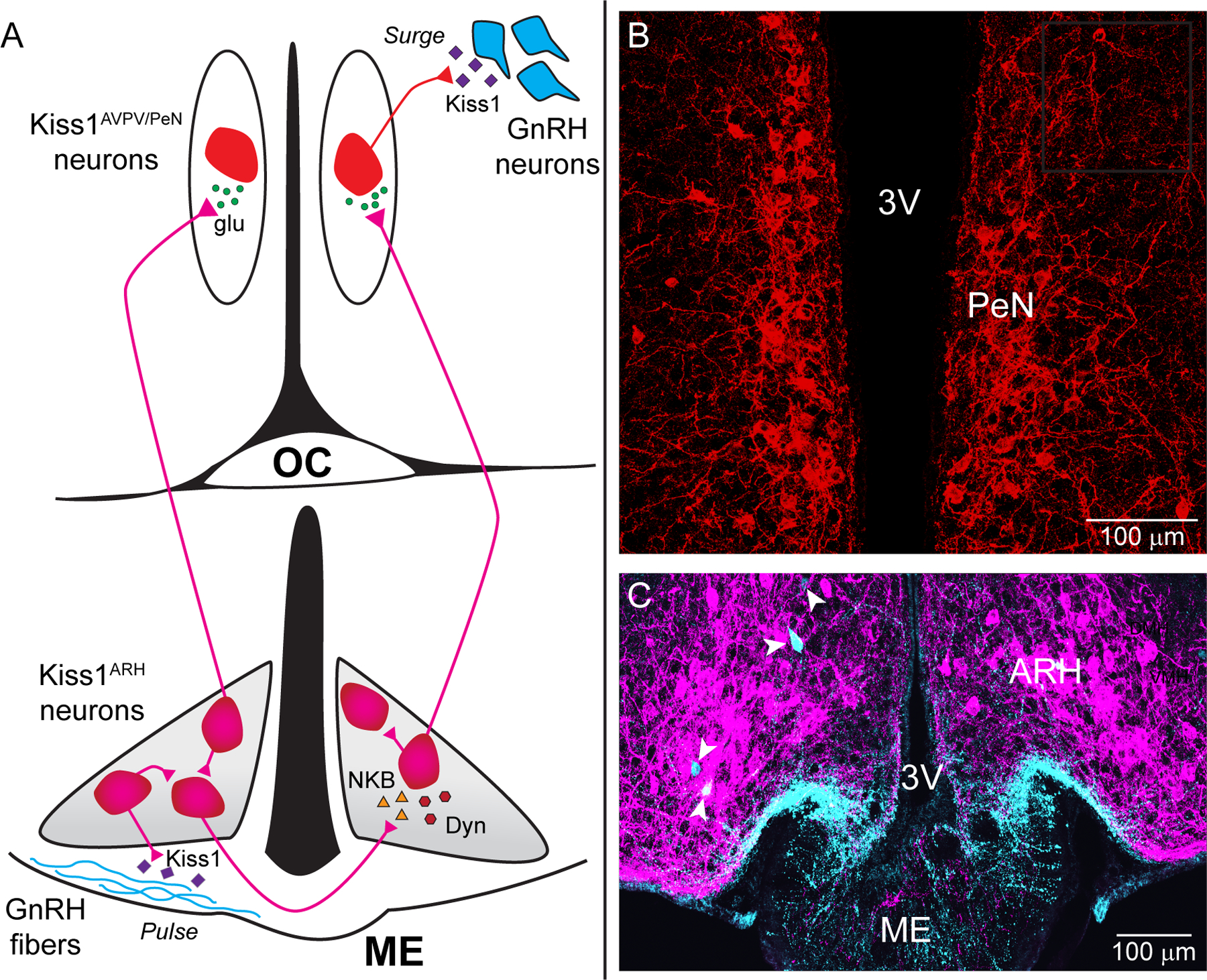
A) Prior to proestrus, estradiol (E2) levels are low and kisspeptin (Kiss1) neurons (bottom, magenta) in the arcuate nucleus of the hypothalamus (ARH, grey region) release neurokinin B (NKB, triangles) that depolarizes and recruits other Kiss1ARH neurons. Dynorphin (Dyn, hexagons) is co-released and acts presynaptically to modulate (inhibit) the release of NKB. Together the two peptides govern the synchronous activity of Kiss1ARH neurons and promote kisspeptin release (diamonds) that stimulates pulsatile gonadotrophin-releasing hormone (GnRH) release from fibers (cyan) in the median eminence (ME). As estradiol levels rise Kiss1ARH neurons transition from peptidergic to primarily fast glutamatergic (circles) neurotransmission to communicate with the Kiss1AVPV/PeN neurons, which stimulates burst-firing of Kiss1AVPV/PeN neurons. E2 also enhances the excitability and kisspeptin release of these rostral Kiss1 neurons (top, red) to robustly excite GnRH neurons via activation of the GPR54 signaling cascade, thereby stimulating the release of GnRH at the time of the preovulatory surge. Kisspeptin, GPR54, NKB, Tacr3 and GnRH are all required for normal fertility. B) Confocal micrograph of the PeN containing Kiss1 neurons found along the third ventricle. C) Confocal micrograph showing labeled Kiss1 cell bodies (magenta) and a few GnRH cell bodies (cyan with white arrowheads) in the ARH as noted in Herde et al. (J. Neuroscience 2013; 33:12689-97). GnRH fibers from the preoptic run along the ventral ARH into the ME. (3V: third ventricle; AVPV: anteroventral periventricular; ARH: arcuate nucleus of the hypothalamus; Kiss1: kisspeptin; ME: median eminence; MnPO: median preoptic nucleus; OC: Optic Chiasm; PeN: periventricular nucleus)
2. Hypothalamic neurons link reproduction and energy homeostasis
Kiss1ARH, proopiomelanocortin (POMC), and neuropeptide Y/agouti-related peptide (AgRP) neurons arise from a common precursor pool (112, 113) and together these neurons govern both reproduction and energy homeostasis. The close juxtaposition of these neuronal subpopulations to the median eminence, a circumventricular organ, enables circulating indicators of energy state (e.g. blood glucose, leptin, and insulin) to reach these neuronal populations which then convey the information downstream targets (114, 115). ARH neurons are also able to sense E2 levels by virtue of robust estrogen receptor expression (116–118), establishing a point of crosstalk between endocrine and homeostatic signaling in the brain. Historically ARH regulation of energy balance was viewed as a “tug-of-war” between two ARH subpopulations, pro-opiomelanocortin (POMC) and neuropeptide Y/agouti-related peptide (AgRP) neurons. However, Kiss1 neurons and kisspeptin/GPR54 signaling has been revealed to also influence metabolism (119), which has led to the idea that these three populations form a “triumvirate” of neurons to regulate energy homeostasis.
AgRP neurons are considered orexigenic and, regardless of energy state, can drive rapid food consumption (120–123). However, recent findings have challenged this “classical” homeostatic model of ARH function. A long-held assumption was that AgRP activity persisted during food consumption until post-ingestional effects emerged (e.g., elevated blood glucose and insulin); however in vivo photometry revealed that detection of food, depending on energy state and palatability, is sufficient to rapidly inhibit AgRP while exciting POMC neurons (124). Therefore, anticipation and extrinsic factors also influence the connectivity between and output of ARH neurons. Regardless, AgRP neurons do not send direct projections to GnRH neurons, but instead act indirectly through GABAergic inhibition of Kiss1 neurons (125). Therefore, low energy stores would lead to increased AgRP excitability and signaling to decrease Kiss1 and, consequently, GnRH activity. Indeed, chemogenetic activation of AgRP neurons prolongs the estrous cycle, whereas targeted ablation of AgRP neurons attenuates the inhibitory tone on Kiss1 neurons (125) and restores fertility in obese or leptin deficient mice (126). Fasting activates AgRP neurons (127), which reduces fertility and expression of Kiss1 in the ARH (128). Therefore, persistent AgRP activity acts as an indicator of undernutrition and inhibits reproduction.
Conversely, POMC neurons are active when energy stores are replete and decrease food intake following stimulation (120, 129, 130). In addition to the amino acid neurotransmitters GABA (131, 132) and glutamate (131), POMC neurons release a diverse complement of neuropeptides. The POMC precursor peptide is processed into several neuropeptides including, but not limited to, α-melanocyte stimulating hormone (α-MSH, excitatory) and β-endorphin (inhibitory) (133). In contrast to AgRP neurons, POMC neurons send direct projections to GnRH neurons, and the selective μ-opioid agonist DAMGO ([D-Ala2, N-Me-Phe4, Gly5-ol]-enkephalin), which mimicks β-endorphin’s action (134), hyperpolarizes (inhibits) GnRH neurons through activation of a K+ conductance (135–138). Naloxone, an opioid antagonist, stimulates GnRH release (139–143) and increases LH production (144, 145), suggesting POMC β-endorphin signaling inhibits GnRH activity and reproductive function (146), whereas selective activation of the α-MSH pathway is stimulatory (146).
More straightforward is Kiss1 to POMC neuronal signaling. Kisspeptin administered icv reduces food intake (147), optogenetic stimulation of Kiss1ARH neurons elicits glutamatergic (80, 148) and kisspeptin mediated excitation of POMC neurons (149). Reciprocal connections are made between POMC and Kiss1 neurons (150). POMC neuronal signaling may also use Kiss1 neurons as an intermediary with GnRH neurons since a subpopulation of Kiss1AVPV/PeN neurons expresses melanocortin 4 receptor (MC4R) (49), and in sheep the majority of Kiss1ARH neurons express the mRNA for MC3R (151). Blockade of melanocortin signaling in peripubertal females decreases Kiss1 mRNA expression in the Kiss1ARH neurons (146), and MC4R knockout impairs fecundity (152). Moreover, overexpression of AgRP, an inverse agonist for MCRs, causes infertility (126, 146, 153). While the specifics of POMC inputs to Kiss1 neurons remains unclear, one would assume these projections act to limit food intake in favor of reproductive behaviors.
Due to the metabolically demanding nature of pregnancy, it is no surprise that neurons involved in metabolism also affect fertility. Women suffering with anorexia often exhibit amenorrhea (154–157) and miscarriage risk is heightened in underweight females (158). Unsurprisingly, when energy reserves are low, AgRP neurons limit reproductive functions (125). Metabolism and kisspeptin neurons also affect one another. For example, hypogonadism due to undernutrition is associated with decreased Kiss1 mRNA expression across a range of ages and species (128, 159–162), Conversely, a high-fat diet is associated with elevated levels of NKB expression and precocious puberty in female rats (163), demonstrating that disruptions of energy homeostasis in either direction can affect kisspeptin signaling. In service of their antagonistic roles to maintain energy balance, AgRP and POMC neurons are inversely regulated by glucose and metabolic hormones including leptin and insulin (164–166). Leptin, a hormone produced by white adipocytes (167), signals the total body energy stores. Mice deficient in leptin or lacking the requisite receptor present an infertile, obese phenotype (168, 169). Since GnRH neurons lack leptin receptors (170, 171), leptin regulation of GnRH neurons is indirect. ARH neurons are one potential line of communication and ablation of AgRP neurons is sufficient to reverse the obese phenotype (126), but as previously stated AgRP neurons do not directly contact GnRH neurons. However, Kiss1ARH neurons express leptin receptors (172, 173), and similar to POMC neurons, leptin depolarizes and increase their firing (173). In contrast, leptin hyperpolarizes AgRP neurons via opening K+-ATP (174) and Kv2.1 channels (175). Without the inhibitory effects of leptin AgRP neurons become highly active (176) and inhibit Kiss1ARH and Kiss1AVPV/PeN neurons (Figure 2) (125) as well as POMC neurons. Therefore, AgRP activity will be relayed to GnRH neurons, at least in part, through Kiss1 and POMC neurons. Injection or overexpression of leptin into lean mice accelerates the onset of puberty (177, 178), and obesity is associated with precocious puberty in females (179, 180). Therefore, undernutrition and adiposity will affect leptin levels, which in turn influence not just fertility but pubertal timing.
Figure 2. Tri-synaptic circuit in the arcuate nucleus of the hypothalamus.
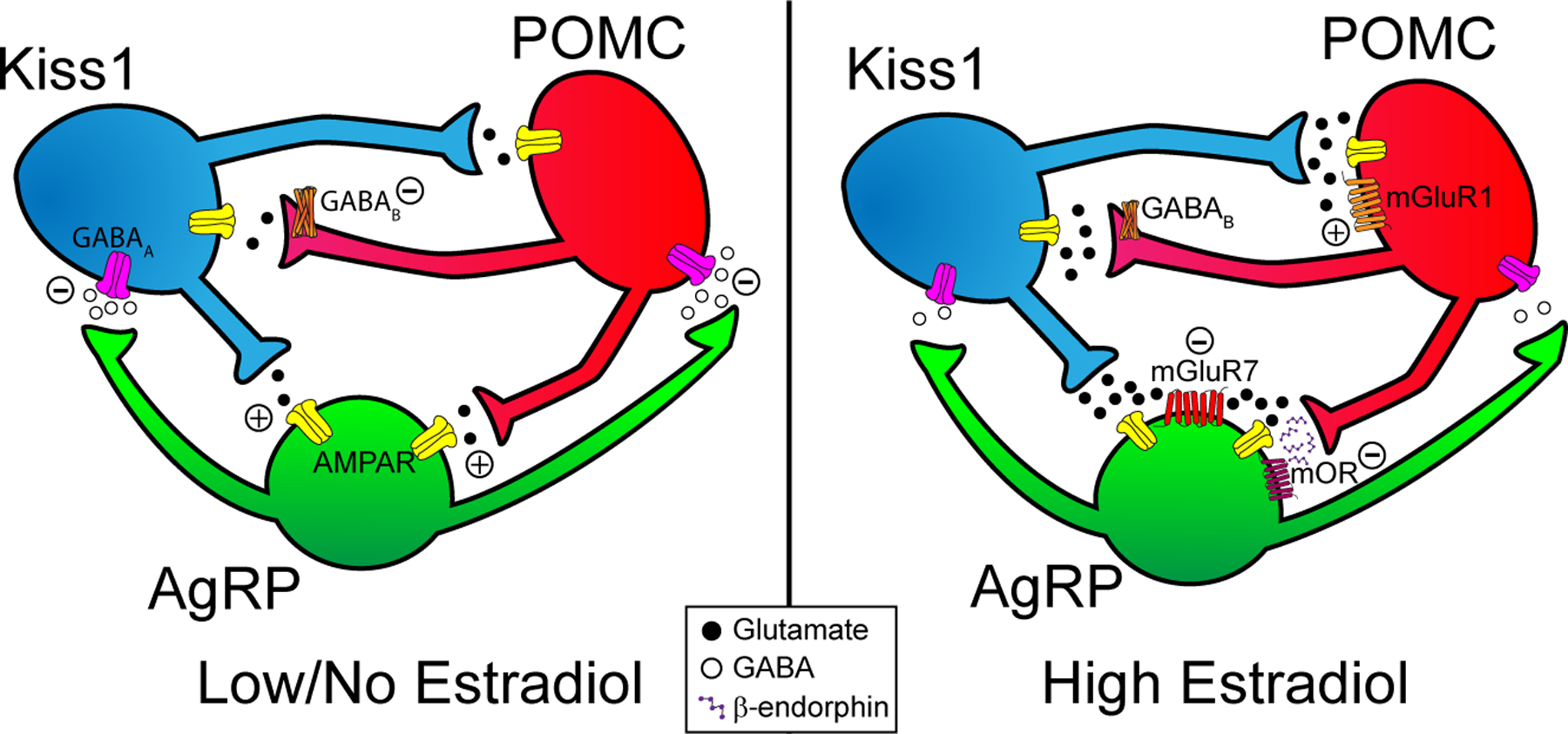
(Left panel) When circulating estradiol levels are low, AgRP neurons will release the inhibitory neurotransmitter GABA onto Kiss1 and POMC neurons. POMC and Kiss1 neurons will release a trickle of glutamate onto AgRP neurons, exciting them through AMPA receptors. (Right panel) When circulating levels of E2 are high in proestrus or with E2-replacement in ovariectomized females, AgRP will display reduced neuronal excitability and GABA release. In POMC neurons the coupling of metabotropic GABAB receptors to GIRK channels is attenuated, further diminishing GABAergic inhibition. Simultaneously in Kiss1ARH neurons Vglut2, CaV3.1, and Hcn1,2 mRNA expression is upregulated to enhance glutamate release as well as the excitatory T-type calcium and h-currents (80). Therefore, POMC and Kiss1 neurons will be disinhibited/excited at the same time their glutamate release probability is enhanced through increased Vglut2 mRNA transcription. However, E2 will also increase expression of the inhibitory mGluR7 receptor in AgRP neurons such that the greater glutamate release will recruit these extrasynaptic receptors, causing an overall inhibitory input. Additionally, POMC neurons will produce more β-endorphin to further inhibit AgRP neurons. Taken together, reciprocal connections between POMC and Kiss1 neurons will act synergistically to increase their activity and excitability while both inhibiting AgRP neurons to decrease food motivation in favor of reproductive behavior. (AgRP: neuropeptide Y/agouti-related peptide; Kiss1: kisspeptin; mOR: μ-opioid receptor; mGluR1: Group I metabotropic glutamate receptor 1; mGluR7: Group III metabotropic glutamate receptor 7; POMC: proopiomelanocortin).
Short-term indices of energy balance also affect ARH function through both genomic and rapid signaling mechanisms (Figure 2). Furthermore, the ARH is a critical control center for peripheral insulin sensitivity and glucose metabolism (181–185). In lean animals, rising blood glucose levels after meal consumption cause insulin release. Circulating insulin easily reaches the ARH neurons adjacent to the median eminence, and in vitro perfusion of insulin into the ARH rapidly depolarize POMC neuron by activating inositol triphosphate 3-kinase signaling pathways (186) and ultimately TRPC5 channels (185). TRPC channels can function as both receptor- or store-operated channels opened, respectively, by membrane delimited receptors or depletion of Ca2+ stores (75, 187). At the same time AgRP neurons are hyperpolarized by insulin through activation of ATP-sensitive potassium channels (185). However, neurons and peripheral tissues can become resistant to insulin and develop glucose intolerance. Acute activation of AgRP neurons rapidly but transiently impairs glucose and insulin tolerance (188) through NPY signaling that limits glucose uptake in brown adipose tissue (BAT) (189). This phenomenon could be a temporary physiological response to simultaneously limit energy expenditure while promoting foraging (121, 190). However, loss of insulin and glucose sensitivity can decouple ARH neurons from homeostatic feedback, promoting or perpetuating obesity (191–193). High-fat diet induced obesity also leads to leptin and ghrelin resistance as well as increased NPY/AgRP neuronal excitability (175, 194). Interestingly, while knockout of leptin receptors in AgRP neurons delays puberty (195), childhood obesity is highly associated with precocious puberty (196–198). In an obese state, TRPC channels associate with the endoplasmic reticulum protein stromal-interaction molecule (STIM1) to function as store-operated and, hence, are no longer opened by insulin (75). E2 protects females from developing insulin resistance in the brain by downregulating STIM1 in POMC neurons, increasing their excitability, and preventing conversion of TRPC to store-operated channels (199). Knockdown of Stim1 mRNA in Kiss1ARH cells enhances TRPC5 currents in response to senktide and protects against HFD-induced obesity (200). More importantly, E2 also protects against insulin resistance by preventing high-fat diet related upregulation of SOCS-3 (suppressor of cytokine signaling 3), thereby preserving insulin signaling in female rodents (199). In addition, insulin significantly increases Pomc mRNA expression within 72 hours following icv administration (201). Therefore, circulating estrogens are vital for maintaining insulin sensitivity throughout the female reproductive cycle and are neuroprotective against insulin resistance in obese states.
The ventromedial nucleus of the hypothalamus (VMH) neurons has long been recognized as an integral site for energy balance (202) that also provides positive estrogen feedback (203). Dorsomedial VMH neurons express leptin and insulin receptors (204–208), but ERα-labeled neurons appear restricted to the ventrolateral region (209). The majority of VMH neurons also play a role in glucose sensing (210, 211), displaying excitation with increasing glucose concentrations (212). Only sparse projections are sent outside the VMH; however, a few axons reach the ARH, dorsomedial hypothalamus (DMH), and paraventricular nucleus of the hypothalamus (PVH) (213, 214). On a gross functional scale, the VMH acts anorexigenically with electrical stimulation decreasing food intake (215), and lesions or injections of ERα interference RNA causing hyperphagic and obese phenotypes (216–219). Steroidogenic factor-1 (SF-1) is expressed exclusively within a subpopulation of VMH neurons (220) that innervate and excite POMC neurons (221–226). SF-1 neurons also send excitatory projections to GnRH neurons (227, 228). ERα signaling appears to stimulate energy expenditure through SF-1 neurons (114), and SF1 ablation diminishes LH secretion (229). NPY inhibits VMH neurons (230) through Y1 receptors (231), attenuating VMH-mediated excitation of POMC neurons (232). Therefore, in addition to direct reciprocal connections NPY and POMC neurons utilize VMH SF-1 neurons as an intermediary regulator. Brain-derived neurotrophic factor (BDNF) is an important protein in the differentiation and survival of neurons through development (233) that also plays a role in adult energy balance, particularly in a number of VMH neurons (234, 235). BDNF is co-expressed by most, but not all (~60%) SF-1 neurons (236) and appears to participate in energy balance (237), specifically by decreasing meal size (238–241). BDNF and ERα are co-localized in the VMH (242), and the anorexigenic efficacy of BDNF is largely dependent on E2 levels (243). So while there are clear links between the VMH and ARH in regards to metabolism and reproduction, whether or not Kiss1 neurons participate is unknown and deserves further study. If synapses are made in either direction with Kiss1 neurons, one would assume these to be excitatory and anorexigenic.
Kiss1 neurons also regulate energy balance through their projections to nuclei outside the mediobasal hypothalamus (i.e., ARH and VMH). The PVH is important for neuroendocrine and autonomic regulation of numerous functions such as food intake (244). Kiss1ARH and Kiss1AVPV/PeN neurons make close contact with PVH AVP and oxytocin neurons, and optogenetic stimulation of each Kiss1 population can, respectively, elicit postsynaptic glutamatergic or GABAergic responses, demonstrating a direct functional input to the PVH (3). As the PVH is heterogeneous in cell type and is composed of subregions with distinct projection patterns (245), further work remains to be done elucidating the functional significance of these Kiss1 inputs. Concerning the other aspect of the control of energy homeostasis, the DMH is important in thermoregulation through energy expenditure(246) (247–249). Leptin, in addition to inhibiting ARH AgRP neurons (176), also activates neurons in the DMH to drive BAT thermogenesis (250–253). Global knockout of Kiss1R/GPR54 (brain and peripheral tissues) selectively induces an obese phenotype in female mice (254–256). However, Kiss1R knockout constrained to BAT produces a lean phenotype with increased metabolism and body temperature (257), suggesting both direct and indirect signaling pathways. Kiss1ARH fibers do make close contact with leptin receptor expressing neurons in the DMH, and optogenetic stimulation elicits direct glutamatergic postsynaptic responses (3). Activation of leptin receptor-expressing DMH neurons robustly induces energy expenditure (258). Conversely, silencing these neurons decreases expenditure and increases food intake (259) with the latter effect likely the consequence of a lost inhibitory input to AgRP neurons (260). Estrogenic signaling may also play a role in adapting to changes in ambient temperature as E2 increases cFos immunocytochemical labeling in the DMH when animals are exposed to cold (261). However, the DMH exhibits low expression of ERα and ERβ (262), which would suggest either involvement of GPER, Gq-mER or an E2-sensitive input. Indeed, projections from leptin-sensitive neurons in the ARH (263), which are estrogen-sensitive, and the DMH (264) are crucial for communicating hormonal signals of energy balance to PVH neurons, which lack leptin receptors (204) and sparsely express ERα mRNA (265). AVP and OT neurons in the PVH as well as corticotrophin releasing hormone neurons do express ERβ (266) (267). Therefore, the steroid (E2)-sensitive Kiss1ARH neurons are likely not only vital for controlling pulsatile release of GnRH/LH but also coordinating energy homeostasis with reproductive activities through projections to regions such as the PVH and DMH.
3. E2 regulation of ARH circuitry and energy balance
More subtle changes are present in signaling and behavior during healthy reproductive cycles as E2 levels naturally rise and fall. Food intake, specifically sweet foods, decreases during the follicular phase (high E2) of the menstrual cycle (268–270). Female rodents also eat up to 25% less food during the evening following the LH surge (271, 272), which could represent the delay between E2-initiated transcription changes and protein synthesis. Decreases in food consumption are due to smaller meal sizes that are not fully compensated for by a slight increase in meal frequency (270, 273–275) , which may speak to how satiety is affected. Intact female rodents exhibit a “scalloping” pattern of activity, in which running activity increases on the night of proestrus (276, 277). Ovariectomy eliminates this effect and leads to an overall decrease in motor activity (278–280) as well as increased food intake (27, 275, 281–285); however, E2 replacement alone is sufficient to restore normal energy balance (23, 27, 275, 278–280). With respect to estrogenic signaling, ERα knockout mice develop an obese phenotype reflective of that observed following ovariectomy (286). Moreover, metabolic deficits are reversed by restoration of ERα, despite lacking the ERE targeting domain, which emphasizes the importance of non-classical signaling (287). Different models of ERβ knockout mice present a range of phenotypes from an impaired ovulatory cycle (288) to complete infertility (289). However, selective deletion of the ERE domain produces only a mild phenotype, once again underscoring the importance of non-classical ERβ signaling (290). GPER knockout mice display sexual dimorphism; although both sexes exhibit reduced energy expenditure and increased body weight, this phenotype emerges six weeks later in female rodents (291). While each subpopulation of ARH neurons exhibits different effects of estrogenic regulation, in general the signaling of anorexigenic neurons is enhanced while orexigenic signaling is attenuated. These are just a few examples of how estrogens are pleiotropic regulators, influencing communication between POMC, AgRP, and Kiss1 neurons in the ARH to coordinate reproduction and energy balance (Figure 2).
POMC neurons are one means by which the anorexigenic effects of E2 are communicated. Disruption of POMC signaling typically results in a positive energy balance. For example, E2 promotes physical activity by upregulating Mc4r gene expression in neurons in the VMH, sensitizing them to POMC inputs (292). POMC-specific deletion of ERα is sufficient to induce hyperphagia and increased heat production (114). The contradictory thermogenesis phenotype could be due to higher circulating levels of E2, which is suggested by blunted negative feedback of E2 on LH release that produces abnormal estrous cycles. Regardless, estrogenic signaling is of tantamount importance to the anorexigenic function of POMC neurons in females. There is an upregulation of Pomc mRNA expression with high circulating levels of E2 (i.e., proestrus/surge levels) (293, 294) when compared to ovariectomized females rodents (295, 296). Processing of the POMC precursor peptide into β-endorphin is also enhanced by E2 (297, 298). In addition, glutamatergic signaling is supported by E2 through increased expression of Vglut2 mRNA which manifests functionally as a higher release probability (150, 299). Furthermore, acutely applied E2 or Gq-coupled mER ligand STX also increases the probability of glutamate release (150), possibly by decoupling GABAB receptors from G protein-coupled inwardly rectifying K+ (GIRK) channels in POMC nerve terminals (27, 300). E2 also enhances the overall excitability of POMC neurons through upregulation of calcium channel subunits and receptor-operated TRPC channels (199, 301). Therefore, POMC neuronal excitability and neurotransmission is potentiated during proestrus to reduce food intake.
Since E2 is an anorexigenic hormone, it is logical that estrogenic signaling inhibits AgRP neuronal activity (302). Initially it was thought that AgRP neurons unilaterally sent inhibitory GABAergic projections to POMC neurons (123, 164); however, channelrhodopsin assisted circuit mapping has shown POMC neurons send reciprocal projections to AgRP neurons, primarily releasing β-endorphin and glutamate (Figure 2) (150). Initial reports noted optogenetic stimulation of POMC neurons most commonly results postsynaptic GABA-mediated currents in unidentified ARH neurons (131). However, other studies focused on responses in AgRP neurons found that optogenetic activation of POMC neurons rarely elicits a fast GABAergic or a slow excitatory (e.g. α-MSH) current (120, 123, 150). Few AgRP neurons express Mc4r but surprisingly nearly half express Mc3r (303). Therefore, the infrequency of GABA and melanocortin mediated responses suggests segregated neurotransmission from POMC neurons (304–307). The predominance of POMC glutamatergic and the infrequency of GABAergic inputs seems contradictory since one would not expect a satiety neuron to excite a hunger neuron. However, enhanced glutamate release from POMC neurons would exert an overall inhibitory tone since E2 upregulates mGluR7 (Group II/III mGluR) mRNA expression in NPY/AgRP neurons (Figure 2) (80). Together these changes in transcription will enhance POMC inhibition of AgRP neurons when circulating E2 is high.
Perhaps, Kiss1 and POMC neurons set the tone of homeostatic circuits based on the reproductive state of the female. As E2 levels peak preceding ovulation, the neuronal activity of AgRP neurons will be suppressed, unless energy reserves are critically low, to reduce food motivation. For example, fasting enhances AgRP activity and signaling by rapidly rewiring circuits (308). In those circumstances AgRP neurons will strongly inhibit Kiss1 neurons (125), disrupting the estrous cycle. Despite initial reports (309), AgRP neurons do express ERα (302); however, under normal physiological conditions rapid E2 signaling bidirectionally adjusts the activity AgRP neurons. For example, E2 alters the ability of GABAB receptors to activate G protein inwardly rectifying K+ (GIRK) channels, either strengthening or weakening the coupling based on the relative expression of ERα versus Gq-mER at the time (302, 310). That is not to say that some genomic estrogenic regulation is also present. For example, E2 also decreases AgRP excitability through increased transcription of Kcnq5 mRNA, enhancing the inhibitory M-current (311). These findings suggest that estrogenic signaling uses both genomic and rapid mechanisms to regulate POMC and Kiss1 function but relies primarily on membrane-initiated signaling for adjusting the activity of AgRP neurons.
4. Conclusions and Future Directions
In summary, estrogenic signaling governs a tripartite collaboration between ARH neurons that regulates GnRH neurons to synchronize energy balance and fertility to maximize reproductive success. When energy reserves are low, POMC activity will be minimal accompanied by a nominal amount of glutamate release onto AgRP neurons (Figure 2). With sufficient energy stores AgRP neurons become less active, leading to disinhibition of POMC neurons (312). Next, POMC activity firing rates rise to the higher frequencies (~20 Hz) (313) necessary to elicit synaptic release of β-endorphin that inhibits AgRP neurons via activation of μ-opioid receptors (150). When circulating E2 levels are high, such as during proestrus, POMC and Kiss1ARH neurons have enhanced glutamate release probability onto AgRP neurons (80). However, simultaneously increased expression of the Group III metabotropic glutamate receptor 7 postsynaptically in AgRP neurons causes this to be a net inhibitory input (80, 150). With the same enhanced glutamate release, Kiss1ARH neurons will excite POMC neurons through Group I mGluRs (80). As genomic changes lag while genes are expressed and transcribed rapid, estrogenic signaling likely eases transitions between states (low vs high E2). The positive and negative feedback between ARH neuronal subpopulations may serve to prevent small and/or transient fluctuations in the energy state of the animal from triggering dramatic shifts in the balance of ARH function (312) and enable E2 to bias, but not dictate, interactions between ARH neurons. The functional output of these neural circuit dynamics would manifest behaviorally as changes in reward salience and motivation. Briefly, when a female has sufficient energy stores to support a pregnancy, ovulation is induced, and priorities switch from ingestive behavior to mating. Indeed, ovariectomized female rodents find sucrose more rewarding than E2-treated females (314). In addition, when Vglut2 mRNA is deleted from Kiss1ARH neurons, this E2 protective effect is lost (80), suggesting that glutamatergic inhibition of AgRP neurons combined with excitation of POMC neurons may be an underlying mechanism.
Although there is little doubt that estrogenic signaling is necessary for Kiss1 neurons to control GnRH and LH release, estrogens clearly orchestrate communication between Kiss1, AgRP, and POMC neurons to regulate energy balance in the service of optimizing reproductive success. Still much work remains to be done to elucidate POMC inputs to ARH and AVPV/PeN Kiss1 neurons (146). Finally, GnRH neurons have variable responses to NPY, α-MSH, and β-endorphin (315) with at least the potency of the μ-opioid response regulated by E2 (316). Therefore, metabolic cues may be communicated directly by POMC and AgRP neurons or through Kiss1 neurons to GnRH neurons. Future studies should be aimed at further unraveling the complexity of the neuronal signaling by this triumvirate of arcuate neurons (Figure 2).
Acknowledgements
The authors thank current and former colleagues who contributed to the work described herein. Research reported in this publication was supported by National Institute of Health R01 grants NS 38809 (MJK), NS 43330 (Oline K. Rønnekleiv) and DK 68098 (MJK & OKR).
References
- 1.King JC, Tobet SA, Snavely FL, Arimura AA. The LHRH system in normal and neonatally androgenized female rats. Peptides. 1980; 185–100. [Google Scholar]
- 2.Lehman MN, Robinson JE, Karsch FJ, Silverman AJ. Immunocytochemical localization of luteinizing hormone-releasing hormone(LHRH) pathways in the sheep brain during anestrous and the mid-luteal phase of the estrous cycle. The Journal of Comparative Neurology. 1986; 24419–35. [DOI] [PubMed] [Google Scholar]
- 3.Stincic TL, Qiu J, Connors AM, Kelly MJ, Rønnekleiv OK. Arcuate and Preoptic Kisspeptin neurons exhibit differential projections to hypothalamic nuclei and exert opposite postsynaptic effects on hypothalamic paraventricular and dorsomedial nuclei in the female mouse. eneuro. 2021ENEURO.0093–21.2021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Silverman AJ. Distribution of luteinizing hormone-releasing hormone (LH-RH) in the guinea pig brain. Endocrinology. 1976; 9930–41. [DOI] [PubMed] [Google Scholar]
- 5.Silverman AJ, Antunes JL, Abrams GM, Nilaver G, Thau R, Robinson JA, Ferin M, Krey LC. The luteinizing hormone-releasing hormone pathway in rhesus (Macaca mulatta) and pigtailed (Macaca nemestrina) monkeys: New observations on thick, unembedded sections. The Journal of Comparative Neurology. 1982; 211309–17. [DOI] [PubMed] [Google Scholar]
- 6.King JC, Anthony ELP, Fitzgerald DM, Stopa EG. Luteinizing hormone-releasing hormone neurons in human preoptic/hypothalamus. The Journal of Clinical Endocrinology & Metabolism. 1985; 6088–97. [DOI] [PubMed] [Google Scholar]
- 7.Levine JE, Ramirez VD. Luteinizing hormone-releasing hormone release during the rat estrous cycle and after ovariectomy, as estimated with push-pull cannulae. Endocrinology. 1982; 1111439–48. [DOI] [PubMed] [Google Scholar]
- 8.Pau KY, Orstead KM, Hess DL, Spies HG. Feedback effects of ovarian steroids on the hypothalamichypophyseal axis in the rabbit. Biology of Reproduction. 1986; 35(4): 1009–23. [DOI] [PubMed] [Google Scholar]
- 9.Moenter SM, Brand RC, Karsch FJ. Dynamics of gonadotropin-releasing hormone (GnRH) secretion during the GnRH surge: Insights into the mechanism of GnRH surge induction. Endocrinology. 1992; 1302978–84. [DOI] [PubMed] [Google Scholar]
- 10.Wagner EJ, Rønnekleiv OK, Bosch MA, Kelly MJ. Estrogen biphasically modifies hypothalamic GABAergic function concomitantly with negative and positive control of luteinizing hormone release. The Journal of Neuroscience. 2001; 21(6): 2085–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Jensen EV, DeSombre ER. Estrogen-receptor interaction. Science. 1973; 182(4108): 126–34. [DOI] [PubMed] [Google Scholar]
- 12.Greene GL, Gilna P, Waterfield M, Baker A, Hort Y, Shine J. Sequence and expression of human estrogen receptor complementary DNA. Science. 1986; 231(4742): 1150–4. [DOI] [PubMed] [Google Scholar]
- 13.Kuiper GG, Enmark E, Pelto-Huikko M, Nilsson S, Gustafsson JÅ. Cloning of a novel estrogen receptor expressed in rat prostate and ovary. Proceedings of the National Academy of Sciences of the United States of America. 1996; 935925–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.O’Malley BW, Tsai MJ. Molecular pathways of steroid receptor action. Biology of Reproduction. 1992; 46(2): 163–7. [DOI] [PubMed] [Google Scholar]
- 15.Muramatsu M, Inoue S. Estrogen receptors: how do they control reproductive and nonreproductive functions? Biochemical and Biophysical Research Communications. 2000; 270(1): 1–10. [DOI] [PubMed] [Google Scholar]
- 16.Gruber CJ, Gruber DM, Gruber IM, Wieser F, Huber JC. Anatomy of the estrogen response element. Trends in Endocrinology & Metabolism. 2004; 15(2): 73–8. [DOI] [PubMed] [Google Scholar]
- 17.Kelly MJ, Levin ER. Rapid actions of plasma membrane estrogen receptors. Trends in Endocrinology & Metabolism. 2001; 12(4): 152–6. [DOI] [PubMed] [Google Scholar]
- 18.Kelly MJ, Rønnekleiv OK. Mini-review: neural signaling of estradiol in the hypothalamus. Molecular Endocrinology. 2015; 29(5): 645–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hammes SR, Levin ER. Extra-nuclear steroid receptors: nature and actions. Endocrine Reviews. 2007; 28726–41. [DOI] [PubMed] [Google Scholar]
- 20.Kelly MJ, Qiu J, Rønnekleiv OK. Estrogen signaling in the hypothalamus. Vitam Horm. 2005; 71123–45. [DOI] [PubMed] [Google Scholar]
- 21.Rønnekleiv OK, Malyala A, Kelly MJ. Membrane-initiated signaling of estrogen in the brain. Semin Reprod Med. 2007; 25(3): 165–77. [DOI] [PubMed] [Google Scholar]
- 22.Mhyre AJ, Dorsa DM. Estrogen activates rapid signaling in the brain: role of estrogen receptor alpha and estrogen receptor beta in neurons and glia. Neuroscience. 2006; 138(3): 851–8. [DOI] [PubMed] [Google Scholar]
- 23.Roepke TA, Xue C, Bosch MA, Scanlan TS, Kelly MJ, Rønnekleiv OK. Genes associated with membraneinitiated signaling of estrogen and energy homeostasis. Endocrinology. 2008; 149(12): 6113–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Filardo EJ, Quinn JA, Bland KI, Frackelton AR, Jr. Estrogen-induced activation of Erk-1 and Erk-2 requires the G protein-coupled receptor homolog, GPR30, and occurs via trans-activation of the epidermal growth factor receptor through release of HB-EGF. Molecular Endocrinology. 2000; 141649–60. [DOI] [PubMed] [Google Scholar]
- 25.Revankar CM, Cimino DF, Sklar LA, Arterburn JB, Prossnitz ER. A transmembrane intracellular estrogen receptor mediates rapid cell signaling. Science. 2005; 307(5715): 1625–30. [DOI] [PubMed] [Google Scholar]
- 26.Qiu J, Bosch MA, Tobias SC, Grandy DK, Scanlan TS, Rønnekleiv OK, Kelly MJ. Rapid signaling of estrogen in hypothalamic neurons involves a novel G-protein-coupled estrogen receptor that activates protein kinase C. The Journal of Neuroscience. 2003; 23(29): 9529–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Qiu J, Bosch MA, Tobias SC, Krust A, Graham S, Murphy S, Korach KS, Chambon P, Scanlan TS, Rønnekleiv OK, Kelly MJ. A G-protein-coupled estrogen receptor is involved in hypothalamic control of energy homeostasis. The Journal of Neuroscience. 2006; 26(21): 5649–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Stincic TL, Rønnekleiv OK, Kelly MJ. Diverse actions of estradiol on anorexigenic and orexigenic hypothalamic arcuate neurons. Hormones and Behavior. 2018; 104146–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Radovick S, Levine JE, Wolfe A. Estrogenic regulation of the GnRH neuron. Front Endocrinol (Lausanne). 2012; 352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Shivers BD, Harlan RE, Morrell JI, Pfaff DW. Absence of oestradiol concentration in cell nuclei of LHRH-immunoreactive neurones. Nature. 1983; 304(5924): 345–7. [DOI] [PubMed] [Google Scholar]
- 31.Wintermantel TM, Campbell RE, Porteous R, Bock D, Grone HJ, Todman MG, Korach KS, Greiner E, Perez CA, Schutz G, Herbison AE. Definition of estrogen receptor pathway critical for estrogen positive feedback to gonadotropin-releasing hormone neurons and fertility. Neuron. 2006; 52(2): 271–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Chu Z, Andrade J, Shupnik MA, Moenter SM. Differential regulation of gonadotropin-releasing hormone neuron activity and membrane properties by acutely applied estradiol: dependence on dose and estrogen receptor subtype. The Journal of Neuroscience. 2009; 29(17): 5616–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Hrabovszky E, Shughrue PJ, Merchenthaler I, Hajszan T, Carpenter CD, Liposits Z, Petersen SL. Detection of estrogen receptor-b messenger ribonucleic acid and 125I-estrogen binding sites in luteinizing hormone-releasing hormone neurons of the rat brain. Endocrinology. 2000; 1413506–9. [DOI] [PubMed] [Google Scholar]
- 34.Hrabovszky E, Steinhauser A, Barabas K, Shughrue PJ, Petersen SL, Merchenthaler I, Liposits Z. Estrogen receptor-beta immunoreactivity in luteinizing hormone-releasing hormone neurons of the rat brain. Endocrinology. 2001; 142(7): 3261–4. [DOI] [PubMed] [Google Scholar]
- 35.Herbison AE, Pape JR. New evidence for estrogen receptors in gonadotropin-releasing hormone neurons. Frontiers in Neuroendocrinology. 2001; 22(4): 292–308. [DOI] [PubMed] [Google Scholar]
- 36.Hrabovszky E, Kalló I, Szlávik N, Keller E, Merchenthaler I, Liposits Z. Gonadotropin-releasing hormone neurons express estrogen receptor-beta. J Clin Endocrinol Metab. 2007; 92(7): 2827–30. [DOI] [PubMed] [Google Scholar]
- 37.Sharifi N, Reuss AE, Wray S. Prenatal LHRH neurons in nasal explant cultures express estrogen receptor beta transcript. Endocrinology. 2002; 143(7): 2503–7. [DOI] [PubMed] [Google Scholar]
- 38.Skinner DC, Dufourny L. Oestrogen receptor beta-immunoreactive neurones in the ovine hypothalamus: distribution and colocalisation with gonadotropin-releasing hormone. J Neuroendocrinol. 2005; 17(1): 29–39. [DOI] [PubMed] [Google Scholar]
- 39.Dupont S, Krust A, Gansmuller A, Dierich A, Chambon P, Mark M. Effect of single and compound knockouts of estrogen receptors α (ERα) and β (ERβ) on mouse reproductive phenotypes. Development. 2000; 127(19): 4277–91. [DOI] [PubMed] [Google Scholar]
- 40.Shughrue PJ, Askew GR, Dellovade TL, Merchenthaler I. Estrogen-binding sites and their functional capacity in estrogen receptor double knockout mouse brain. Endocrinology. 2002; 1431643–50. [DOI] [PubMed] [Google Scholar]
- 41.Dubal DB, Zhu H, Yu J, Rau SW, Shughrue PJ, Merchenthaler I, Kindy MS, Wise PM. Estrogen receptor alpha, not beta, is a critical link in estradiol-mediated protection against brain injury. Proceedings of the National Academy of Sciences of the United States of America. 2001; 981952–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Jayes FL, Burns KA, Rodriguez KF, Kissling GE, Korach KS. The naturally occurring luteinizing hormone surge is diminished in mice lacking estrogen receptor Beta in the ovary. Biol Reprod. 2014; 90(2): 24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Novaira HJ, Negron AL, Graceli JB, Capellino S, Schoeffield A, Hoffman GE, Levine JE, Wolfe A, Wondisford FE, Radovick S. Impairments in the reproductive axis of female mice lacking estrogen receptor β in GnRH neurons. Am J Physiol Endocrinol Metab. 2018; 315(5): E1019–e33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Terasawa E, Kenealy BP. Neuroestrogen, rapid action of estradiol, and GnRH neurons. Frontiers in Neuroendocrinology. 2012; 33(4): 364–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Gottsch ML, Cunningham MJ, Smith JT, Popa SM, Acohido BV, Crowley WF, Seminara S, Clifton DK, Steiner RA. A role for kisspeptins in the regulation of gonadotropin secretion in the mouse. Endocrinology. 2004; 145(9): 4073–7. [DOI] [PubMed] [Google Scholar]
- 46.Irwig MS, Fraley GS, Smith JT, Acohido BV, Popa SM, Cunningham MJ, Gottsch ML, Clifton DK, Steiner RA. Kisspeptin activation of gonadotropin releasing hormone neurons and regulation of KiSS-1 mRNA in the male rat. Neuroendocrinology. 2004; 80(4): 264–72. [DOI] [PubMed] [Google Scholar]
- 47.Smith JT, Cunningham MJ, Rissman EF, Clifton DK, Steiner RA. Regulation of Kiss1 gene expression in the brain of the female mouse. Endocrinology. 2005; 146(9): 3686–92. [DOI] [PubMed] [Google Scholar]
- 48.Estrada KM, Clay CM, Pompolo S, Smith JT, Clarke IJ. Elevated KiSS-1 expression in the arcuate nucleus prior to the cyclic preovulatory gonadotrophin-releasing hormone/lutenising hormone surge in the ewe suggests a stimulatory role for kisspeptin in oestrogen-positive feedback. Journal of Neuroendocrinology. 2006; 18(10): 806–9. [DOI] [PubMed] [Google Scholar]
- 49.Cravo RM, Margatho LO, Osborne-Lawrence S, Donato JJ, Atkin S, Bookout AL, Rovinsky S, Frazão R, Lee CE, Gautron L, Zigman JM, Elias CF. Characterization of Kiss1 neurons using transgenic mouse models. Neuroscience. 2011; 17337–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kotani M, Detheux M, Vandenbogaerde A, Communi D, Vanderwinden JM, Le Poul E, Brezillon S, Tyldesley R, Suarez-Huerta N, Vandeput F, Blanpain C, Schiffmann SN, Vassart G, Parmentier M. The metastasis suppressor gene KiSS-1 encodes kisspeptins, the natural ligands of the orphan G protein-coupled receptor GPR54. Journal of Biological Chemistry. 2001; 276(37): 34631–6. [DOI] [PubMed] [Google Scholar]
- 51.Seminara SB, Messager S, Chatzidaki EE, Thresher RR, Acierno JS, Shagoury JK, Bo-Abbas Y, Kuohung W, Schwinof KM, Hendrick AG, Zahn D, Dixon J, Kaiser UB, Slaugenhaupt SA, Gusella JF, O’Rahilly S, Carlton MBL, Crowley WF, Aparicio SAJR, Colledge WH. The GPR54 gene as a regulator of puberty. The New England Journal of Medicine. 2003; 349(17): 1614–27. [DOI] [PubMed] [Google Scholar]
- 52.Smith JT, Clarke IJ. Kisspeptin expression in the brain: catalyst for the initiation of puberty. Reviews in Endocrine and Metabolic Disorders. 2007; 8(1): 1–9. [DOI] [PubMed] [Google Scholar]
- 53.d’Anglemont de Tassigny X, Fagg LA, Dixon JPC, Day K, Leitch HG, Hendrick AG, Zahn D, Franceschini I, Caraty A, Carlton MBL, Aparicio SAJR, Colledge WH. Hypogonadotropic hypogonadism in mice lacking a functional KiSS 1 gene. Proceedings of the National Academy of Sciences of the United States of America. 2007; 104(25): 10714–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Topaloglu AK, Reimann F, Guclu M, Yalin AS, Kotan LD, Porter KM, Serin A, Mungan NO, Cook JR, Imamoglu S, Akalin NS, Yuksel B, O’Rahilly S, Semple RK. TAC3 and TACR3 mutations in familial hypogonadotropic hypogonadism reveal a key role for Neurokinin B in the central control of reproduction. Nature Genetics. 2009; 41(3): 354–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Clarkson J, d’Anglemont de Tassigny X, Moreno AS, Colledge WH, Herbison AE. Kisspeptin-GPR54 signaling is essential for preovulatory gonadotropin-releasing hormone neuron activation and the luteinizing hormone surge. The Journal of Neuroscience. 2008; 28(35): 8691–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Yeo SH, Clarkson J, Herbison AE. Kisspeptin-gpr54 signaling at the GnRH neuron is necessary for negative feedback regulation of luteinizing hormone secretion in female mice. Neuroendocrinology. 2014; 100(2–3): 191–7. [DOI] [PubMed] [Google Scholar]
- 57.León S, Barroso A, Vázquez MJ, García-Galiano D, Manfredi-Lozano M, Ruiz-Pino F, Heras V, Romero-Ruiz A, Roa J, Schutz G, Kirilov M, Gaytan F, Pinilla L, Tena-Sempere M. Direct Actions of Kisspeptins on GnRH Neurons Permit Attainment of Fertility but are Insufficient to Fully Preserve Gonadotropic Axis Activity. Sci Rep. 2016; 619206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Kinoshita M, Tsukamura H, Adachi S, Matsui H, Uenoyama Y, Iwata K, Yamada S, Inoue K, Ohtaki T, Matsumoto H, Maeda K. Involvement of central metastin in the regulation of preovulatory luteinizing hormone surge and estrous cyclicity in female rats. Endocrinology. 2005; 146(10): 4431–6. [DOI] [PubMed] [Google Scholar]
- 59.Bosch MA, Tonsfeldt KJ, Rønnekleiv OK. mRNA expression of ion channels in GnRH neurons: subtype-specific regulation by 17β-Estradiol. Molecular and Cellular Endocrinology. 2013; 367(1–2): 85–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Kirilov M, Clarkson J, Liu X, Roa J, Campos P, Porteous R, Schütz G, Herbison AE. Dependence of fertility on kisspeptin-Gpr54 signaling at the GnRH neuron. Nature Communications. 2013; 42492-. [DOI] [PubMed] [Google Scholar]
- 61.Shughrue PJ, Lane MV, Merchenthaler I. Comparative distribution of estrogen receptor-α and -β mRNA in the rat central nervous system. The Journal of Comparative Neurology. 1997; 388507–25. [DOI] [PubMed] [Google Scholar]
- 62.Franceschini I, Lomet D, Cateau M, Delsol G, Tillet Y, Caraty A. Kisspeptin immunoreactive cells of the ovine preoptic area and arcuate nucleus co-express estrogen receptor alpha. Neuroscience Letters. 2006; 401(3): 225–30. [DOI] [PubMed] [Google Scholar]
- 63.Han S-K, Gottsch ML, Lee KJ, Popa SM, Smith JT, Jakawich SK, Clifton DK, Steiner RA, Herbison AE. Activation of gonadotropin-releasing hormone neurons by kisspeptin as a neuroendocrine switch for the onset of puberty. The Journal of Neuroscience. 2005; 25(49): 11349–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Liu X, Lee K, Herbison AE. Kisspeptin excites gonadotropin-releasing hormone (GnRH) neurons through a phospholipase C/calcium-dependent pathway regulating multiple ion channels. Endocrinology. 2008; 149(9): 4605–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Constantin S, Iremonger KJ, Herbison AE. In vivo recordings of GnRH neuron firing reveal heterogeneity and dependence upon GABAA receptor signaling. The Journal of Neuroscience. 2013; 339394–401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Zhang C, Roepke TA, Kelly MJ, Rønnekleiv OK. Kisspeptin depolarizes gonadotropin-releasing hormone neurons through activation of TRPC-like cationic channels. The Journal of Neuroscience. 2008; 28(17): 442334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Pielecka-Fortuna J, Chu Z, Moenter SM. Kisspeptin acts directly and indirectly to increase gonadotropinreleasing hormone neuron activity and its effects are modulated by estradiol. Endocrinology. 2008; 149(4): 1979–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Constantin S, Caligioni CS, Stojilkovic S, Wray S. Kisspeptin-10 facilitates a plasma membrane-driven calcium oscillator in gonadotropin-releasing hormone-1 neurons. Endocrinology. 2009; 150(3): 1400–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Iremonger KJ, Porteous R, Herbison AE. Spike and Neuropeptide-Dependent Mechanisms Control GnRH Neuron Nerve Terminal Ca(2+) over Diverse Time Scales. The Journal of Neuroscience. 2017; 37(12): 3342–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Zhang XB, Spergel DJ. Kisspeptin inhibits high-voltage activated Ca2+ channels in GnRH neurons via multiple Ca2+ influx and release pathways. Neuroendocrinology. 2012; 96(1): 68–80. [DOI] [PubMed] [Google Scholar]
- 71.Liu X, Herbison AE. Small-conductance calcium-activated potassium channels control excitability and firing dynamics in gonadotropin-releasing hormone (GnRH) neurons. Endocrinology. 2008; 149(7): 3598–604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Kroll H, Bolsover S, Hsu J, Kim SH, Bouloux PM. Kisspeptin-evoked calcium signals in isolated primary rat gonadotropin-releasing hormone neurones. Neuroendocrinology. 2011; 93(2): 114–20. [DOI] [PubMed] [Google Scholar]
- 73.Clapham DE. TRP channels as cellular sensors. Nature. 2003; 426517–24. [DOI] [PubMed] [Google Scholar]
- 74.Ambudkar IS, Ong HL. Organization and function of TRPC channelsomes. Pflügers Archiv:European Journal of Physiology. 2007; 455(2): 187–200. [DOI] [PubMed] [Google Scholar]
- 75.Salido GM, Jardin I, Rosado JA. The TRPC ion channels: association with Orai1 and STIM1 proteins and participation in capacitative and non-capacitative calcium entry. Advances in Experimental Medicine and Biology. 2011; 704413–33. [DOI] [PubMed] [Google Scholar]
- 76.Goodman RL, Lehman MN, Smith JT, Coolen LM, de Oliveira CVR, Jafarzadehshirazi MR, Pereira A, Iqbal J, Caraty A, Ciofi P, Clarke IJ. Kisspeptin neurons in the arcuate nucleus of the ewe express both dynorphin A and neurokinin B. Endocrinology. 2007; 148(12): 5752–60. [DOI] [PubMed] [Google Scholar]
- 77.Navarro VM, Gottsch ML, Wu M, García-Galiano D, Hobbs SJ, Bosch MA, Pinilla L, Clifton DK, Dearth A, Rønnekleiv OK, Braun RE, Plamiter RD, Tena-Sempere M, Alreja M, Steiner RA. Regulation of NKB pathways and their roles in the control of Kiss1 neurons in the arcuate nucleus of the male mouse. Endocrinology. 2011; 152(11): 4265–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Bartzen-Sprauer J, Klosen P, Ciofi P, Mikkelsen JD, Simonneaux V. Photoperiodic co-regulation of kisseptin, neurokinin B and dynorphin in the hypothalamus of a seasonal rodent. Journal of Neuroendocrinology. 2014; 26(8): 510–20. [DOI] [PubMed] [Google Scholar]
- 79.Lehman MN, Coolen LM, Goodman RL. Minireview: kisspeptin/neurokinin B/dynorphin (KNDy) cells of the arcuate nucleus: a central node in the control of gonadotropin-releasing hormone secretion. Endocrinology. 2010; 151(8): 3479–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Qiu J, Rivera HM, Bosch MA, Padilla SL, Stincic TL, Palmiter RD, Kelly MJ, Rønnekleiv OK. Estrogenicdependent glutamatergic neurotransmission from kisspeptin neurons governs feeding circuits in females. eLife. 2018; 7e35656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Navarro VM, Gottsch ML, Chavkin C, Okamura H, Clifton DK, Steiner RA. Regulation of gonadotropinreleasing hormone secretion by kisspeptin/dynorphin/neurokinin B neurons in the arcuate nucleus of the mouse. The Journal of Neuroscience. 2009; 29(38): 11859–66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Ohkura S, Takase K, Matsuyama S, Mogi K, Ichimaru T, Wakabayashi Y, Uenoyama Y, Mori Y, Steiner RA, Tsukamura H, Maeda KI, Okamura H. Gonadotrophin-releasing hormone pulse generator activity in the hypothalamus of the goat. Journal of Neuroendocrinology. 2009; 21(10): 813–21. [DOI] [PubMed] [Google Scholar]
- 83.Wakabayashi Y, Nakada T, Murata K, Ohkura S, Mogi K, Navarro VM, Clifton DK, Mori Y, Tsukamura H, Maeda K-I, Steiner RA, Okamura H. Neurokinin B and dynorphin A in kisspeptin neurons of the arcuate nucleus participate in generation of periodic oscillation of neural activity driving pulsatile gonadotropin-releasing hormone secretion in the goat. The Journal of Neuroscience. 2010; 30(8): 3124–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Moore AM, Desroziers E, Czieselsky K, Prescott M, Yip SH, Herbison AE, Campbell RE. Synaptic innervation of the GnRH neuron distal dendron in female mice. Endocrinology. 2018; 159(9): 3200–8. [DOI] [PubMed] [Google Scholar]
- 85.Ducret E, Gaidamaka G, Herbison AE. Electrical and morphological characteristics of anteroventral periventricular nucleus kisspeptin and other neurons in the female mouse. Endocrinology. 2010; 151(5): 2223–32. [DOI] [PubMed] [Google Scholar]
- 86.Navarro VM, Bosch MA, León S, Simavli S, True C, Pinilla L, Carroll RS, Seminara SB, Tena-Sempere M, Rønnekleiv OK, Kaiser UB. The integrated hypothalamic tachykinin-kisspeptin system as a central coordinator for reproduction. Endocrinology. 2015; 156(5): 627–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Qiu J, Nestor CC, Zhang C, Padilla SL, Palmiter RD, Kelly MJ, Rønnekleiv OK. High-frequency stimulation-induced peptide release synchronizes arcuate kisspeptin neurons and excited GnRH neurons. eLife. 2016; 5e16246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Wang L, Burger LL, Greenwald-Yarnell M, Myers MG, Moenter SM. Glutamatergic transmission to hypothalamic kisspeptin neurons is differentially regulated by estradiol through estrogen receptor α in adult female mice. The Journal of Neuroscience. 2018; 38(5): 1061–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Clarkson J, Herbison AE. Dual phenotype kisspeptin-dopamine neurones of the rostral periventricular area of the third ventricle project to GnRH hormones. Journal of Neuroendocrinology. 2011; 23293–301. [DOI] [PubMed] [Google Scholar]
- 90.Rønnekleiv OK, Qiu J, Kelly MJ. Hypothalamic Kisspeptin Neurons and the Control of Homeostasis. Endocrinology. 2022; 1631–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Piet R, Boehm U, Herbison AE. Estrous cycle plasticity in the hyperpolarization-activated current Ih is mediated by circulating 17β-estradiol in preoptic area kisspeptin neurons. The Journal of Neuroscience. 2013; 33(26): 10828–39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Zhang C, Tonsfeldt KJ, Qiu J, Bosch MA, Kobayashi K, Steiner RA, Kelly MJ, Rønnekleiv OK. Molecular mechanisms that drive estradiol-dependent burst firing of Kiss1 neurons in the rostral periventricular preoptic area. American Journal of Physiology: Endocrinology and Metabolism. 2013; 305(11): E1384–E97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Zhang C, Bosch MA, Qiu J, Ronnekleiv OK, Kelly MJ. 17beta-Estradiol increases persistent Na(+) current and excitability of AVPV/PeN Kiss1 neurons in female mice. Mol Endocrinol. 2015; 29(4): 518–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Lau OC, Shen B, Wong CO, Tjong YW, Lo CY, Wang HC, Huang Y, Yung WH, Chen YC, Fung ML, Rudd JA, Yao X. TRPC5 channels participate in pressure-sensing in aortic baroreceptors. Nature Communications. 2016; 711947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Wang L, DeFazio RA, Moenter SM. Excitability and burst generation of AVPV kisspeptin neurons are regulated by the estrous cycle via multiple conductances modulated by estradiol action. eNeuro. 2016; 3(3): e0094–16. 2016 1–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Wiegand SJ, Terasawa E, Bridson WE. Persistent estrus and blockade of progesterone-induced LH release follows lesions which do not damage the suprachiasmatic nucleus. Endocrinology. 1978; 1021645–8. [DOI] [PubMed] [Google Scholar]
- 97.Rønnekleiv OK, Kelly MJ. Plasma prolactin and luteinizing hormone profiles during the estrous cycle of the female rat: effects of surgically induced persistent estrus. Neuroendocrinology. 1988; 47(2): 133–41. [DOI] [PubMed] [Google Scholar]
- 98.Petersen SL, Barraclough CA. Suppression of spontaneous LH surges in estrogen-treated ovariectomized rats by microimplants of antiestrogens into the preoptic brain. Brain Research. 1989; 484279–89. [DOI] [PubMed] [Google Scholar]
- 99.Ma YJ, Kelly MJ, Rønnekleiv OK. Pro-gonadotropin-releasing hormone (ProGnRH) and GnRH content in the preoptic area and the basal hypothalamus of anterior medial preoptic nucleus/suprachiasmatic nucleus-lesioned persistent estrous rats. Endocrinology. 1990; 127:2654–2664. [DOI] [PubMed] [Google Scholar]
- 100.Yip SH, Boehm U, Herbison AE, Campbell RE. Conditional viral tract tracing delineates the projections of the distinct kisspeptin neuron populations to gonadotropin-releasing hormone (GnRH) neurons in the mouse. Endocrinology. 2015; 156(7): 2582–94. [DOI] [PubMed] [Google Scholar]
- 101.Piet R, Kalil B, McLennan T, Porteous R, Czieselsky K, Herbison AE. Dominant neuropeptide cotransmission in kisspeptin-GABA regulation of GnRH neuron firing driving ovulation. The Journal of Neuroscience. 2018; 38(28): 6310–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Knobil E The electrophysiology of the GnRH pulse generator in the rhesus monkey. Journal of Steroid Biochemistry. 1989; 33(4B): 669–71. [DOI] [PubMed] [Google Scholar]
- 103.Okamura H, Tsukamura H, Ohkura S, Uenoyama Y, Wakabayashi Y, Maeda K. Kisspeptin and GnRH pulse generation. Advances in Experimental Medicine and Biology. 2013; 784297–323. [DOI] [PubMed] [Google Scholar]
- 104.Kimura F, Nishihara M, Hiruma H, Funabashi T. Naloxone increases the frequency of the electrical activity of luteinizing hormone-releasing hormone pulse generator in long-term ovariectomized rats. Neuroendocrinology. 1991; 53(1): 97–102. [DOI] [PubMed] [Google Scholar]
- 105.Kinsey-Jones JS, Li XF, Luckman SM, O’Byrne KT. Effects of kisspeptin-10 on the electrophysiological manifestation of gonadotropin-releasing hormone pulse generator activity in the female rat. Endocrinology. 2008; 149(3): 1004–8. [DOI] [PubMed] [Google Scholar]
- 106.Clarkson J, Han SY, Piet R, McLennan T, Kane GM, Ng J, Porteous RW, Kim JS, Colledge WH, Iremonger KJ, Herbison AE. Definition of the hypothalamic GnRH pulse generator in mice. Proceedings of the National Academy of Sciences of the United States of America. 2017; 114(47): E10216–E23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Moore AM, Coolen LM, Lehman MN. In vivo imaging of the GnRH pulse generator reveals a temporal order of neuronal activation and synchronization during each pulse. Proceedings of the National Academy of Sciences. 2022; 119(6): e2117767119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Han SY, McLennan T, Czieselsky K, Herbison AE. Selective optogenetic activation of arcuate kisspeptin neurons generates pulsatile luteinizing hormone secretion. Proceedings of the National Academy of Sciences of the United States of America. 2015; 112(42): 13109–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Lin XH, Lass G, Kong LS, Wang H, Li XF, Huang HF, O’Byrne KT. Optogenetic Activation of Arcuate Kisspeptin Neurons Generates a Luteinizing Hormone Surge-Like Secretion in an Estradiol-Dependent Manner. Front Endocrinol (Lausanne). 2021; 12775233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Jamieson BB, Bouwer GT, Campbell RE, Piet R. Estrous Cycle Plasticity in the Central Clock Output to Kisspeptin Neurons: Implications for the Preovulatory Surge. Endocrinology. 2021; 162(6). [DOI] [PubMed] [Google Scholar]
- 111.Padilla SL, Perez JG, Ben-Hamo M, Johnson CW, Sanchez REA, Bussi IL, Palmiter RD, de la Iglesia HO. Kisspeptin neurons in the arcuate nucleus of the hypothalamus orchestrate circadian rhythms and metabolism. Current Biology. 2019; 29(4): 592–604 e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Padilla SL, Carmody JS, Zeltser LM. Pomc-expressing progenitors give rise to antagonistic neuronal populations in hypothalamic feeding circuits. Nature Medicine. 2010; 16(4): 403–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Sanz E, Quintana A, Deem JD, Steiner RA, Palmiter RD, McKnight GS. Fertility-regulating Kiss1 neurons arise from hypothalamic POMC-expressing progenitors. The Journal of Neuroscience. 2015; 35(14): 5549–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Xu X, Coats JK, Yang CF, Wang A, Ahmed OM, Alvarado M, Izumi T, Shah NM. Modular genetic control of sexually dimorphic behaviors. Cell. 2012; 148(3): 596–607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Belgardt BF, Okamura T, Brüning JC. Hormone and glucose signalling in POMC and AgRP neurons. The Journal of Physiology. 2009; 587(22): 5305–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Lauber AH, Mobbs CV, Muramatsu M, Pfaff DW. Estrogen receptor messenger RNA expression in rat hypothalamus as a function of genetic sex and estrogen dose. Endocrinology. 1991; 1293180–6. [DOI] [PubMed] [Google Scholar]
- 117.Romano GJ, Krust A, Pfaff DW. Expression and estrogen regulation of progesterone receptor mRNA in neurons of the mediobasal hypothalamus: an in situ hybridization study. Molecular Endocrinology. 1989; 3(8): 1295–300. [DOI] [PubMed] [Google Scholar]
- 118.Shughrue PJ, Komm B, Merchenthaler I. The distribution of estrogen receptor-β mRNA in the rat hypothalamus. Steroids. 1996; 61678–81. [DOI] [PubMed] [Google Scholar]
- 119.Izzi-Engbeaya C, Dhillo WS. Emerging roles for kisspeptin in metabolism. J Physiol. 2022; 600(5): 1079–88. [DOI] [PubMed] [Google Scholar]
- 120.Aponte Y, Atasoy D, Sternson SM. AGRP neurons are sufficient to orchestrate feeding behavior rapidly and without training. Nature Neuroscience. 2011; 14(3): 351–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Krashes MJ, Koda S, Ye C, Rogan SC, Adams AC, Cusher DS, Maratos-Flier E, Roth BL, Lowell BB. Rapid, reversible activation of AgRP neurons drives feeding behavior in mice. Journal of Clinical Investigation. 2011; 121(4): 1424–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Krashes MJ, Shah BP, Koda S, Lowell BB. Rapid versus delayed stimulation of feeding by the endogenously released AgRP neuron mediators GABA, NPY, and AgRP. Cell Metabolism. 2013; 18(4): 588–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Atasoy D, Betley JN, Su HH, Sternson SM. Deconstruction of a neural circuit for hunger. Nature. 2012; 488(7410): 172–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Chen Y, Lin Y-C, Kuo T-W, Knight ZA. Sensory detection of food rapidly modulates arcuate feeding circuits. Cell. 2015; 160(5): 829–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Padilla SL, Qiu J, Nestor CC, Zhang C, Smith AW, Whiddon BB, Rønnekleiv OK, Kelly MJ, Palmiter RD. AgRP to Kiss1 neuron signaling links nutritional state and fertility. Proceedings of the National Academy of Sciences. 2017; 114(9): 2413–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Wu Q, Whiddon BB, Palmiter RD. Ablation of neurons expressing agouti-related protein, but not melanin concentrating hormone, in leptin-deficient mice restores metabolic function and fertility. Proceedings of the National Academy of Sciences of the United States of America. 2012; 1093155–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Liu T, Kong D, Shah BP, Ye C, Koda S, Saunders A, Ding JB, Yang Z, Sabatini BL, Lowell BB. Fasting activation of AgRP neurons requires NMDA receptors and involves spinogenesis and increased excitatory tone. Neuron. 2012; 73(3): 511–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Matsuzaki T, Iwasa T, Kinouchi R, Yoshida S, Murakami M, Gereltsetseg G, Yamamoto S, Kuwahara A, Yasui T, Irahara M. Fasting reduces the kiss1 mRNA levels in the caudal hypothalamus of gonadally intact adult female rats. Endocrinologia Japonica. 2011; 58(11): 1003–12. [DOI] [PubMed] [Google Scholar]
- 129.Wei Q, Krolewski DM, Moore S, Kumar V, Li F, Martin B, Tomer R, Murphy GG, Deisseroth K, Watson SJ, Akil H. Uneven balance of power between hypothalamic peptidergic neurons in the control of feeding. Proceedings of the National Academy of Sciences. 2018; 115(40): E9489–E98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Zhan C, Zhou J, Feng Q, Zhang JE, Lin S, Bao J, Wu P, Luo M. Acute and long-term suppression of feeding behavior by POMC neurons in the brainstem and hypothalamus, respectively. The Journal of Neuroscience. 2013; 33(8): 3624–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Dicken MS, Tooker RE, Hentges ST. Regulation of GABA and glutamate release from proopiomelanocortin neuron terminals in intact hypothalamic networks. The Journal of Neuroscience. 2012; 32(12): 4042–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Hentges ST, Nishiyama M, Overstreet LS, Stenzel-Poore M, Williams JT, Low MJ. GABA release from proopiomelanocortin neurons. The Journal of Neuroscience. 2004; 24(7): 1578–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Allen RG, Hatfield JM, Stack J. Post-translational processing of pro-opiomelanocortin (POMC)-derived peptides during fetal monkey pituitary development. I. Adrenocorticotropin (ACTH) and alpha-melanotropins (alpha-MSHs). Dev Biol. 1988; 126(1): 156–63. [DOI] [PubMed] [Google Scholar]
- 134.Goldstein A, Naidu A. Multiple opioid receptors: ligand selectivity profiles and binding site signatures. Molecular Pharmacology. 1989; 36(2): 265–72. [PubMed] [Google Scholar]
- 135.Thind KK, Goldsmith PC. Infundibular gonadotropin-releasing hormone neurons are inhibited by direct opioid and autoregulatory synapses in juvenile monkeys. Neuroendocrinology. 1988; 47(3): 203–16. [DOI] [PubMed] [Google Scholar]
- 136.Chen W-P, Witkin JW, Silverman AJ. β-endorphin and gonadotropin-releasing hormone synaptic input to gonadotropin-releasing hormone neurosecretory cells in the male rat. The Journal of Comparative Neurology. 1989; 28685–95. [DOI] [PubMed] [Google Scholar]
- 137.Lagrange AH, Kelly MJ. Activation of PKA: a novel transduction mechanism for estrogen’s modulation of m-opioid/K+ channel coupling. Society for Neuroscience Abstracts. 1995; 211896. [Google Scholar]
- 138.Lagrange AH, Rønnekleiv OK, Kelly MJ. Estradiol-17β and μ-opioid peptides rapidly hyperpolarize GnRH neurons: A cellular mechanism of negative feedback? Endocrinology. 1995; 136(5): 2341–4. [DOI] [PubMed] [Google Scholar]
- 139.Masotto C, Sahu A, Dube MG, Kalra SP. A decrease in opioid tone amplifies the luteinizing hormone surge in estrogen-treated ovariectomized rats: comparisons with progesterone effects. Endocrinology. 1990; 126(1): 18–25. [DOI] [PubMed] [Google Scholar]
- 140.Zhang Q, Gallo RV. Effect of prodynorphin-derived opioid peptides on the ovulatory luteinizing hormone surge in the proestrous rat. Endocrine. 2002; 18(1): 27–32. [DOI] [PubMed] [Google Scholar]
- 141.Lustig R, Pfaff D, Fishman J. Induction of LH hypersecretion in cyclic rats during the afternoon of oestrus by oestrogen in conjunction with progesterone antagonism or opioidergic blockade. Journal of Endocrinology. 1988; 117(2): 229–35. [DOI] [PubMed] [Google Scholar]
- 142.Lustig RH, Pfaff DW, Fishman J. Opioidergic modulation of the oestradiol-induced LH surge in the rat: roles of ovarian steroids. Journal of Endocrinology. 1988; 116(1): 55–69. [DOI] [PubMed] [Google Scholar]
- 143.Van Vugt DA, Bakst G, Dyrenfurth I, Ferin M. Naloxone stimulation of luteinizing hormone secretion in the female monkey: influence of endocrine and experimental conditions. Endocrinology. 1983; 113(5): 1858–64. [DOI] [PubMed] [Google Scholar]
- 144.Babu GN, Marco J, Bona-Gallo A, Gallo RV. Steroid-independent endogenous opioid peptide suppression of pulsatile luteinizing hormone release between estrus and diestrus 1 in the rat estrous cycle. Brain Research. 1987; 416(2): 235–42. [DOI] [PubMed] [Google Scholar]
- 145.Rossmanith WG, Mortola JF, Yen SS. Role of endogenous opioid peptides in the initiation of the midcycle luteinizing hormone surge in normal cycling women. The Journal of Clinical Endocrinology & Metabolism. 1988; 67(4): 695–700. [DOI] [PubMed] [Google Scholar]
- 146.Manfredi-Lozano M, Roa J, Ruiz-Pino F, Piet R, Garcia-Galiano D, Pineda R, Zamora A, Leon S, Sanchez-Garrido MA, Romero-Ruiz A, Dieguez C, Vazquez MJ, Herbison AE, Pinilla L, Tena-Sempere M. Defining a novel leptin–melanocortin–kisspeptin pathway involved in the metabolic control of puberty. Molecular Metabolism. 2016; 5(10): 844–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Stengel A, Wang L, Goebel-Stengel M, Tache Y. Centrally injected kisspeptin reduces food intake by increasing meal intervals in mice. Neuroreport. 2011; 22(5): 253–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Nestor CC, Qiu J, Padilla SL, Zhang C, Bosch MA, Fan W, Aicher SA, Palmiter RD, Rønnekleiv OK, Kelly MJ. Optogenetic stimulation of arcuate nucleus Kiss1 neurons reveals a steroid-dependent glutamatergic input to POMC and AgRP neurons in male mice. Molecular Endocrinology. 2016; 30(6): 630–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Fu LY, van den Pol AN. Kisspeptin directly excites anorexigenic proopiomelanocortin neurons but inhibits orexigenic neuropeptide Y cells by an indirect synaptic mechanism. The Journal of Neuroscience. 2010; 30(30): 10205–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Stincic TL, Grachev P, Bosch MA, Rønnekleiv OK, Kelly MJ. Estradiol drives the anorexigenic activity of proopiomelanocortin neurons in female mice. eNeuro. 2018; 5(4): e0103–18. 2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Merkley CM, Shuping SL, Sommer JR, Nestor CC. Evidence That Agouti-Related Peptide May Directly Regulate Kisspeptin Neurons in Male Sheep. Metabolites. 2021; 11(3). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Sandrock M, Schulz A, Merkwitz C, Schöneberg T, Spanel-Borowski K, Ricken A. Reduction in corpora lutea number in obese melanocortin-4-receptor-deficient mice. Reproductive Biology and Endocrinology. 2009; 7(1): 24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Granholm NH, Jeppesen KW, Japs RA. Progressive infertility in female lethal yellow mice (Ay/a; strain C57BL/6J). Journal of Reproduction and Fertility. 1986; 76(1): 279–87. [DOI] [PubMed] [Google Scholar]
- 154.Chan JL, Mantzoros CS. Role of leptin in energy-deprivation states: normal human physiology and clinical implications for hypothalamic amenorrhoea and anorrexia nervosa. Lancet. 2005; 36674–85. [DOI] [PubMed] [Google Scholar]
- 155.Allaway HCM, Southmayd EA, De Souza MJ. The physiology of functional hypothalamic amenorrhea associated with energy deficiency in exercising women and in women with anorexia nervosa. Hormone Molecular Biology and Clinical Investigation. 2016; 25(2): 91–119. [DOI] [PubMed] [Google Scholar]
- 156.Golden NH, Shenker IR. Amenorrhea in anorexia nervosa neuroendocrine control of hypothalamic dysfunction. International Journal of Eating Disorders. 1994; 16(1): 53–60. [DOI] [PubMed] [Google Scholar]
- 157.Gordon CM, Ackerman KE, Berga SL, Kaplan JR, Mastorakos G, Misra M, Murad MH, Santoro NF, Warren MP. Functional Hypothalamic Amenorrhea: An Endocrine Society Clinical Practice Guideline. J Clin Endocrinol Metab. 2017; 102(5): 1413–39. [DOI] [PubMed] [Google Scholar]
- 158.Balsells M, García-Patterson A, Corcoy R. Systematic review and meta-analysis on the association of prepregnancy underweight and miscarriage. European Journal of Obstetrics & Gynecology and Reproductive Biology. 2016; 20773–9. [DOI] [PubMed] [Google Scholar]
- 159.Castellano JM, Navarro VM, Fernández-Fernández R, Nogueiras R, Tovar S, Roa J, Vazquez MJ, Vigo E, Casanueva FF, Aguilar E, Pinilla L, Dieguez C, Tena-Sempere M. Changes in hypothalamic KiSS-1 system and restoration of pubertal activation of the repoductive axis by kisspeptin in undernutrition. Endocrinology. 2005; 146(9): 3917–25. [DOI] [PubMed] [Google Scholar]
- 160.True C, Kirigiti MA, Kievit P, Grove KL, Smith MS. Leptin is not the critical signal for kisspeptin or luteinising hormone restoration during exit from negative energy balance. Journal of Neuroendocrinology. 2011; 231099–112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Yang JA, Yasrebi A, Snyder M, Roepke TA. The interaction of fasting, caloric restriction, and diet-induced obesity with 17ß-estradiol on the expression of KNDy neuropeptides and their receptors in the female mouse. Molecular and Cellular Endocrinology. 2016; 43735–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Backholer K, Smith JT, Rao A, Pereira A, Iqbal J, Ogawa S, Li Q, Clarke IJ. Kisspeptin cells in the ewe brain respond to leptin and communicate with neuropeptide Y and proopiomelanocortin cells. Endocrinology. 2010; 151(5): 2233–43. [DOI] [PubMed] [Google Scholar]
- 163.Li XF, Lin YS, Kinsey-Jones JS, O’Byrne KT. High-fat diet increases LH pulse frequency and kisspeptinneurokinin B expression in puberty-advanced female rats. Endocrinology. 2012; 153(9): 4422–31. [DOI] [PubMed] [Google Scholar]
- 164.Cowley MA, Smart JL, Rubinstein M, Cerdán MG, Diano S, Horvath TL, Cone RD, Low MJ. Leptin activates anorexigenic POMC neurons through a neural network in arcuate nucleus. Nature. 2001; 411(6836): 480–4. [DOI] [PubMed] [Google Scholar]
- 165.Faouzi M, Leshan R, Bjornholm M, Hennessey T, Jones J, Munzberg H. Differential accessibility of circulating leptin to individual hypothalamic sites. Endocrinology. 2007; 148(11): 5414–23. [DOI] [PubMed] [Google Scholar]
- 166.Ibrahim N, Bosch MA, Smart JL, Qiu J, Rubinstein M, Rønnekleiv OK, Low MJ, Kelly MJ. Hypothalamic proopiomelanocortin neurons are glucose responsive and express KATP channels. Endocrinology. 2003; 144(4): 1331–40. [DOI] [PubMed] [Google Scholar]
- 167.Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM. Positional cloning of the mouse obese gene and its human homologue. Nature. 1994; 372(6505): 425–32. [DOI] [PubMed] [Google Scholar]
- 168.Ingalls AM, Dickie MM, Snell GD. Obese, a new mutation in the house mouse. Journal of Heredity. 1950; 41(12): 317–8. [DOI] [PubMed] [Google Scholar]
- 169.Halaas JL, Gajiwala KS, Maffei M, Cohen SL, Chait BT, Rabinowitz D, Lallone RL, Burley SK, Friedman JM. Weight-reducing effects of the plasma protein encoded by the obese gene. Science. 1995; 269(5223): 543–6. [DOI] [PubMed] [Google Scholar]
- 170.Finn PD, Cunningham MJ, Pau KY, Spies HG, Clifton DK, Steiner RA. The stimulatory effect of leptin on the neuroendocrine reproductive axis of the monkey. Endocrinology. 1998; 139(11): 4652–62. [DOI] [PubMed] [Google Scholar]
- 171.Quennell JH, Mulligan AC, Tups A, Liu X, Phipps SJ, Kemp CJ, Herbison AE, Grattan DR, Anderson GM. Leptin indirectly reulates gonadotropin-releasing hormone neuronal function. Endocrinology. 2009; 1502805–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Donato JJ, Cravo RM, Frazão R, Gautron L, Scott MM, Lachey J, Castro IA, Margatho LO, Lee S, Lee C, Richardson JA, Friedman J, Chua S Jr., Coppari R, Zigman JM, Elmquist JK, Elias CF. Leptin’s effect on puberty in mice is relayed by the ventral premammillary nucleus and does not require signaling in Kiss1 neurons. The Journal of Clinical Investigation. 2011; 121355–68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Qiu J, Fang Y, Bosch MA, Rønnekleiv OK, Kelly MJ. Guinea pig kisspeptin neurons are depolarized by leptin via activation of TRPC channels. Endocrinology. 2011; 152(4): 1503–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.van den Top M, Lee K, Whyment AD, Blanks AM, Spanswick D. Orexigen-sensitive NPY/AgRP pacemaker neurons in the hypothalamic arcuate nucleus. Nature Neuroscience. 2004; 7(5): 493–4. [DOI] [PubMed] [Google Scholar]
- 175.Baver SB, Hope K, Guyot S, Bjorbaek C, Kaczorowski C, O’Connell KM. Leptin modulates the intrinsic excitability of AgRP/NPY neurons in the arcuate nucleus of the hypothalamus. The Journal of Neuroscience. 2014; 34(16): 5486–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Takahashi KA, Cone RD. Fasting induces a large, leptin-dependent increase in the intrinsic action potential frequency of orexigenic arcuate nucleus neuropeptide Y/Agouti-related protein neurons. Endocrinology. 2005; 146(3): 1043–7. [DOI] [PubMed] [Google Scholar]
- 177.Yura S, Ogawa Y, Sagawa N, Masuzaki H, Itoh H, Ebihara K, Aizawa-Abe M, Fujii S, Nakao K. Accelerated puberty and late-onset hypothalamic hypogonadism in female transgenic skinny mice overexpressing leptin. The Journal of Clinical Investigation. 2000; 105(6): 749–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Ahima RS, Dushay J, Flier SN, Prabakaran D, Flier JS. Leptin accelerates the onset of puberty in normal female mice. The Journal of Clinical Investigation. 1997; 99(3): 391–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Aksglaede L, Juul A, Olsen LW, Sørensen TIA. Age at puberty and the emerging obesity epidemic. PLOS ONE. 2009; 4(12): e8450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.De Leonibus C, Marcovecchio ML, Chiavaroli V, de Giorgis T, Chiarelli F, Mohn A. Timing of puberty and physical growth in obese children: a longitudinal study in boys and girls. Pediatric Obesity. 2014; 9(4): 292–9. [DOI] [PubMed] [Google Scholar]
- 181.Ruud J, Steculorum SM, Brüning JC. Neuronal control of peripheral insulin sensitivity and glucose metabolism. Nature Communications. 2017; 815259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Schwartz MW, Woods SC, Porte D, Jr., Seeley RJ, Baskin DG. Central nervous system control of food intake. Nature. 2000; 404(6778): 661–71. [DOI] [PubMed] [Google Scholar]
- 183.Morton GJ, Cummings DE, Baskin DG, Barsh GS, Schwartz MW. Central nervous system control of food intake and body weight. Nature. 2006; 443(7109): 289–95. [DOI] [PubMed] [Google Scholar]
- 184.Belgardt BF, Bruning JC. CNS leptin and insulin action in the control of energy homeostasis. Annals of the New York Academy of Sciences. 2010; 121297–113. [DOI] [PubMed] [Google Scholar]
- 185.Qiu J, Zhang C, Borgquist A, Nestor CC, Smith AW, Bosch MA, Ku S, Wagner EJ, Rønnekleiv OK, Kelly MJ. Insulin excites anorexigenic proopiomelanocortin neurons via activation of canonical transient receptor potential channels. Cell Metabolism. 2014; 19(4): 682–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Xu AW, Kaelin CB, Takeda K, Akira S, Schwartz MW, Barsh GS. PI3K integrates the action of insulin and leptin on hypothalamic neurons. Journal of Clinical Investigation. 2005; 115(4): 951–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Birnbaumer L The TRPC class of ion channels: a critical review of their roles in slow, sustained increases in intracellular Ca2+ concentrations. Annual Review of Pharmacology and Toxicology. 2009; 49395–426. [DOI] [PubMed] [Google Scholar]
- 188.Steculorum SM, Ruud J, Karakasilioti I, Backes H, Engstrom Ruud L, Timper K, Hess ME, Tsaousidou E, Mauer J, Vogt MC, Paeger L, Bremser S, Klein AC, Morgan DA, Frommolt P, Brinkkotter PT, Hammerschmidt P, Benzing T, Rahmouni K, Wunderlich FT, Kloppenburg P, Bruning JC. AgRP Neurons Control Systemic Insulin Sensitivity via Myostatin Expression in Brown Adipose Tissue. Cell. 2016; 165(1): 125–38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Engström Ruud L, Pereira MMA, de Solis AJ, Fenselau H, Brüning JC. NPY mediates the rapid feeding and glucose metabolism regulatory functions of AgRP neurons. Nat Commun. 2020; 11(1): 442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Dietrich MO, Zimmer MR, Bober J, Horvath TL. Hypothalamic Agrp neurons drive stereotypic behaviors beyond feeding. Cell. 2015; 160(6): 1222–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.White JD, Olchovsky D, Kershaw M, Berelowitz M. Increased hypothalamic content of preproneuropeptide-Y messenger ribonucleic acid in streptozotocin-diabetic rats. Endocrinology. 1990; 126(2): 765–72. [DOI] [PubMed] [Google Scholar]
- 192.Williams G, Gill JS, Lee YC, Cardoso HM, Okpere BE, Bloom SR. Increased neuropeptide Y concentrations in specific hypothalamic regions of streptozocin-induced diabetic rats. Diabetes. 1989; 38(3): 321–7. [DOI] [PubMed] [Google Scholar]
- 193.Loh K, Zhang L, Brandon A, Wang Q, Begg D, Qi Y, Fu M, Kulkani R, Teo J, Baldock P, Brüning JC, Cooney G, Neely G, Herzog H. Insulin controls food intake and energy balance via NPY neurons. Molecular Metabolism. 2017; 6574–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Briggs DI, Enriori PJ, Lemus MB, Cowley MA, Andrews ZB. Diet-Induced Obesity Causes Ghrelin Resistance in Arcuate NPY/AgRP Neurons. Endocrinology. 2010; 151(10): 4745–55. [DOI] [PubMed] [Google Scholar]
- 195.Egan OK, Inglis MA, Anderson GM. Leptin signalling in AgRP neurons modulates puberty onset and adult fertility in mice. The Journal of Neuroscience. 2017; doi: 10.1523/JNEUROSCI.3138-16.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Ahmed ML, Ong KK, Dunger DB. Childhood obesity and the timing of puberty. Trends Endocrinol Metab. 2009; 20(5): 237–42. [DOI] [PubMed] [Google Scholar]
- 197.Li W, Liu Q, Deng X, Chen Y, Liu S, Story M. Association between Obesity and Puberty Timing: A Systematic Review and Meta-Analysis. International Journal of Environmental Research and Public Health. 2017; 14(10): 1266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Sandhu J, Ben-Shlomo Y, Cole TJ, Holly J, Davey Smith G. The impact of childhood body mass index on timing of puberty, adult stature and obesity: a follow-up study based on adolescent anthropometry recorded at Christ’s Hospital (1936–1964). International Journal of Obesity. 2006; 30(1): 14–22. [DOI] [PubMed] [Google Scholar]
- 199.Qiu J, Bosch MA, Meza C, Navarro UV, Nestor CC, Wagner EJ, Rønnekleiv OK, Kelly MJ. Estradiol protects proopiomelanocortin neurons against insulin resistance. Endocrinology. 2018; 159(2): 647–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Qiu J, Stincic TL, Bosch MA, Connors AM, Petrie SK, Rønnekleiv OK, Kelly MJ. Deletion of Stim1 in hypothalamic arcuate nucleus Kiss1 neurons potentiates synchronous GCaMP activity and protects against dietinduced obesity. J Neurosc. 2021; 419688–701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Benoit SC, Air EL, Coolen LM, Strauss R, Jackman A, Clegg DJ, Seeley RJ, Woods SC. The catabolic action of insulin in the brain is mediated by melanocortins. The Journal of Neuroscience. 2002; 22(20): 9048–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Hetherington AW. The relationship of various hypothalamic lesions to adiposity and other phenomena in the rat. American Journal of Physiology. 1941; 133326–7. [Google Scholar]
- 203.Caraty A, Fabre-Nys C, Delaleu B, Locatelli A, Bruneau G, Karsch FJ, Herbison A. Evidence that the mediobasal hypothalamus is the primary site of action of estradiol in inducing the preovulatory gonadotropin releasing hormone surge in the ewe. Endocrinology. 1998; 1391752–60. [DOI] [PubMed] [Google Scholar]
- 204.Elmquist JK, Bjorbaek C, Ahima RS, Flier JS, Saper CB. Distributions of leptin receptor mRNA isoforms in the rat brain. The Journal of Comparative Neurology. 1998; 395(4): 535–47. [PubMed] [Google Scholar]
- 205.Scott MM, Lachey JL, Sternson SM, Lee CE, Elias CF, Friedman JM, Elmquist JK. Leptin targets in the mouse brain. The Journal of Comparative Neurology. 2009; 514(5): 518–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Cotero VE, Zhang BB, Routh VH. The response of glucose-excited neurones in the ventromedial hypothalamus to decreased glucose is enhanced in a murine model of type 2 diabetes mellitus. J Neuroendocrinol. 2010; 22(2): 65–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Levin BE, Routh VH, Kang L, Sanders NM, Dunn-Meynell AA. Neuronal glucosensing what do we know after 50 years? Diabetes. 2004; 532521–8. [DOI] [PubMed] [Google Scholar]
- 208.Klöckener T, Hess S, Belgardt BF, Paeger L, Verhagen LA, Husch A, Sohn JW, Hampel B, Dhillon H, Zigman JM, Lowell BB, Williams KW, Elmquist JK, Horvath TL, Kloppenburg P, Brüning JC. High-fat feeding promotes obesity via insulin receptor/PI3K-dependent inhibition of SF-1 VMH neurons. Nature Neuroscience. 2011; 14(7): 911–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Yokosuka M, Okamura H, Hayashi S. Postnatal development and sex difference in neurons containing estrogen receptor-alpha immunoreactivity in the preoptic brain, the diencephalon, and the amygdala in the rat. The Journal of Comparative Neurology. 1997; 389(1): 81–93. [DOI] [PubMed] [Google Scholar]
- 210.Barnes MB, Lawson MA, Beverly JL. Rate of fall in blood glucose and recurrent hypoglycemia affect glucose dynamics and noradrenergic activation in the ventromedial hypothalamus. Am J Physiol Regul Integr Comp Physiol. 2011; 301(6): R1815–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Chan O, Lawson M, Zhu W, Beverly JL, Sherwin RS. ATP-Sensitive K+ Channels Regulate the Release of GABA in the Ventromedial Hypothalamus During Hypoglycemia. Diabetes. 2007; 56(4): 1120–6. [DOI] [PubMed] [Google Scholar]
- 212.Kang L, Routh VH, Kuzhikandathil EV, Gaspers LD, Levin BE. Physiological and molecular characteristics of rat hypothalamic ventromedial nucleus glucosensing neurons. Diabetes. 2004; 53(3): 549–59. [DOI] [PubMed] [Google Scholar]
- 213.Millhouse OE. Certain ventromedial hypothalamic afferents. Brain Research. 1973; 55(1): 89–105. [DOI] [PubMed] [Google Scholar]
- 214.Canteras NS, Simerly RB, Swanson LW. Organization of projections from the ventromedial nucleus of the hypothalamus: a Phaseolus vulgaris-leucoagglutinin study in the rat. The Journal of Comparative Neurology. 1994; 348(1): 41–79. [DOI] [PubMed] [Google Scholar]
- 215.Ruffin M, Nicolaidis S. Electrical stimulation of the ventromedial hypothalamus enhances both fat utilization and metabolic rate that precede and parallel the inhibition of feeding behavior. Brain Res. 1999; 846(1): 23–9. [DOI] [PubMed] [Google Scholar]
- 216.Inoue S, Campfield LA, Bray GA. Comparison of metabolic alterations in hypothalamic and high fat dietinduced obesity. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology. 1977; 233(3): R162–R8. [DOI] [PubMed] [Google Scholar]
- 217.Sakaguchi T, Arase K, Bray GA. Sympathetic activity and food intake of rats with ventromedial hypothalamic lesions. International journal of obesity. 1988; 12(4): 285–91. [PubMed] [Google Scholar]
- 218.King BM. The rise, fall, and resurrection of the ventromedial hypothalamus in the regulation of feeding behavior and body weight. Physiology & Behavior. 2006; 87(2): 221–44. [DOI] [PubMed] [Google Scholar]
- 219.Musatov S, Chen W, Pfaff DW, Mobbs CV, Yang XJ, Clegg DJ, Kaplitt MG, Ogawa S. Silencing of estrogen receptor alpha in the ventromedial nucleus of hypothalamus leads to metabolic syndrome. Proceedings of the National Academy of Sciences of the United States of America. 2007; 104(7): 2501–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Ikeda Y, Luo X, Abbud R, Nilson JH, Parker KL. The nuclear receptor steroidogenic factor 1 is essential for the formation of the ventromedial hypothalamic nucleus. Mol Endocrinol. 1995; 9(4): 478–86. [DOI] [PubMed] [Google Scholar]
- 221.Sternson SM, Shepherd GMG, Friedman JM. Topographic mapping of VMH-› arcuate nucleus microcircuits and their reorganization by fasting. Nature Neuroscience. 2005; 81356–63. [DOI] [PubMed] [Google Scholar]
- 222.Cardinal P, André C, Quarta C, Bellocchio L, Clark S, Elie M, Leste-Lasserre T, Maitre M, Gonzales D, Cannich A, Pagotto U, Marsicano G, Cota D. CB1 cannabinoid receptor in SF1-expressing neurons of the ventromedial hypothalamus determines metabolic responses to diet and leptin. Mol Metab. 2014; 3(7): 705–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223.Kinyua AW, Yang DJ, Chang I, Kim KW. Steroidogenic Factor 1 in the Ventromedial Nucleus of the Hypothalamus Regulates Age-Dependent Obesity. PLoS One. 2016; 11(9): e0162352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Conde K, Fabelo C, Krause WC, Propst R, Goethel J, Fischer D, Hur J, Meza C, Ingraham HA, Wagner EJ. Testosterone Rapidly Augments Retrograde Endocannabinoid Signaling in Proopiomelanocortin Neurons to Suppress Glutamatergic Input from Steroidogenic Factor 1 Neurons via Upregulation of Diacylglycerol Lipase-α. Neuroendocrinology. 2017; 105(4): 341–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Fabelo C, Hernandez J, Chang R, Seng S, Alicea N, Tian S, Conde K, Wagner EJ. Endocannabinoid Signaling at Hypothalamic Steroidogenic Factor-1/Proopiomelanocortin Synapses Is Sex- and Diet-Sensitive. Front Mol Neurosci. 2018; 11214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Hernandez J, Fabelo C, Perez L, Moore C, Chang R, Wagner EJ. Nociceptin/orphanin FQ modulates energy homeostasis through inhibition of neurotransmission at VMN SF-1/ARC POMC synapses in a sex- and diet-dependent manner. Biology of Sex Differences. 2019; 10(1): 9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Boehm U, Zou Z, Buck LB. Feedback loops link odor and phermone signaling with reproduction. Cell. 2005; 123683–95. [DOI] [PubMed] [Google Scholar]
- 228.Yoon H, Enquist LW, Dulac C. Olfactory inputs to hypothalamic neurons controlling reproduction and fertility. Cell. 2005; 123(4): 669–82. [DOI] [PubMed] [Google Scholar]
- 229.Kim KW, Li S, Zhao H, Peng B, Tobet SA, Elmquist JK, Parker KL, Zhao L. CNS-specific ablation of steroidogenic factor 1 results in impaired female reproductive function. Mol Endocrinol. 2010; 24(6): 1240–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Stanley BG, Chin AS, Leibowitz SF. Feeding and drinking elicited by central injection of neuropeptide Y: Evidence for a hypothalamic site(s) of action. Brain Research Bulletin. 1985; 14(6): 521–4. [DOI] [PubMed] [Google Scholar]
- 231.Zammaretti F, Panzica G, Eva C. Sex-dependent regulation of hypothalamic neuropeptide Y-Y1 receptor gene expression in moderate/high fat, high-energy diet-fed mice. The Journal of Physiology. 2007; 583(Pt 2): 445–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Chee MJ, Myers MG Jr., Price CJ, Colmers WF. Neuropeptide Y suppresses anorexigenic output from the ventromedial nucleus of the hypothalamus. The Journal of Neuroscience. 2010; 30(9): 3380–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Huang EJ, Reichardt LF. Neurotrophins: Roles in Neuronal Development and Function. Annual Review of Neuroscience. 2001; 24(1): 677–736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Tran PV, Akana SF, Malkovska I, Dallman MF, Parada LF, Ingraham HA. Diminished hypothalamic bdnf expression and impaired VMH function are associated with reduced SF-1 gene dosage. J Comp Neurol. 2006; 498(5): 637–48. [DOI] [PubMed] [Google Scholar]
- 235.Unger TJ, Calderon GA, Bradley LC, Sena-Esteves M, Rios M. Selective deletion of Bdnf in the ventromedial and dorsomedial hypothalamus of adult mice results in hyperphagic behavior and obesity. The Journal of Neuroscience. 2007; 27(52): 14265–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Kamitakahara A, Xu B, Simerly R. Ventromedial hypothalamic expression of Bdnf is required to establish normal patterns of afferent GABAergic connectivity and responses to hypoglycemia. Molecular metabolism. 2016; 5(2): 91–101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Rios M BDNF and the central control of feeding: accidental bystander or essential player? Trends in Neurosciences. 2013; 36(2): 83–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Wang C, Bomberg E, Levine A, Billington C, Kotz CM. Brain-derived neurotrophic factor in the ventromedial nucleus of the hypothalamus reduces energy intake. American Journal of Physiology: Regulatory, Integrative and Comparative Physiology. 2007; 293(3): R1037–45. [DOI] [PubMed] [Google Scholar]
- 239.Wang C, Bomberg E, Billington CJ, Levine AS, Kotz CM. Brain-derived neurotrophic factor (BDNF) in the hypothalamic ventromedial nucleus increases energy expenditure. Brain Research. 2010; 133666–77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.Spaeth AM, Kanoski SE, Hayes MR, Grill HJ. TrkB receptor signaling in the nucleus tractus solitarius mediates the food intake-suppressive effects of hindbrain BDNF and leptin. American Journal of Physiology-Endocrinology and Metabolism. 2012; 302(10): E1252–E60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Fox EA, Byerly MS. A mechanism underlying mature-onset obesity: evidence from the hyperphagic phenotype of brain-derived neurotrophic factor mutants. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology. 2004; 286(6): R994–R1004. [DOI] [PubMed] [Google Scholar]
- 242.Blurton-Jones M, Kuan PN, Tuszynski MH. Anatomical evidence for transsynaptic influences of estrogen on brain-derived neurotrophic factor expression. The Journal of Comparative Neurology. 2004; 468(3): 347–60. [DOI] [PubMed] [Google Scholar]
- 243.Zhu Z, Liu X, Senthil Kumar SPD, Zhang J, Shi H. Central expression and anorectic effect of brain-derived neurotrophic factor are regulated by circulating estradiol levels. Hormones and Behavior. 2013; 63(3): 533–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244.Shah BP, Vong L, Olson DP, Koda S, Krashes MJ, Ye C, Yang Z, Fuller PM, Elmquist JK, Lowell BB. MC4R-expressing glutamatergic neurons in the paraventricular hypothalamus regulate feeding and are synaptically connected to the parabrachial nucleus. Proceedings of the National Academy of Sciences. 2014; 111(36): 13193–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Simmons DM, Swanson LW. Comparison of the spatial distribution of seven types of neuroendocrine neurons in the rat paraventricular nucleus: toward a global 3D model. J Comp Neurol. 2009; 516(5): 423–41. [DOI] [PubMed] [Google Scholar]
- 246.Morrison SF, Madden CJ, Tupone D. Central neural regulation of brown adipose tissue thermogenesis and energy expenditure. Cell Metabolism. 2014; 19(5): 741–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Dimicco JA, Zaretsky DV. The dorsomedial hypothalamus: a new player in thermoregulation. American Journal of Physiology: Regulatory, Integrative and Comparative Physiology. 2007; 292(1): R47–63. [DOI] [PubMed] [Google Scholar]
- 248.Sarkar S, Zaretskaia MV, Zaretsky DV, Moreno M, DiMicco JA. Stress- and lipopolysaccharide-induced c-fos expression and nNOS in hypothalamic neurons projecting to medullary raphe in rats: a triple immunofluorescent labeling study. Eur J Neurosci. 2007; 26(8): 2228–38. [DOI] [PubMed] [Google Scholar]
- 249.Yoshida K, Li X, Cano G, Lazarus M, Saper CB. Parallel preoptic pathways for thermoregulation. The Journal of Neuroscience. 2009; 29(38): 11954–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250.Haynes WG, Morgan DA, Walsh SA, Mark AL, Sivitz WI. Receptor-mediated regional sympathetic nerve activation by leptin. J Clin Invest. 1997; 100(2): 270–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251.Gavrilova O, Leon LR, Marcus-Samuels B, Mason MM, Castle AL, Refetoff S, Vinson C, Reitman ML. Torpor in mice is induced by both leptin-dependent and -independent mechanisms. Proc Natl Acad Sci U S A. 1999; 96(25): 14623–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 252.Richards RJ, Blalock A, Liao J, Reisin E. Leptin: sympathetic and cardiovascular effects. Curr Cardiol Rep. 2003; 5(6): 453–8. [DOI] [PubMed] [Google Scholar]
- 253.Zhang Y, Kerman IA, Laque A, Nguyen P, Faouzi M, Louis GW, Jones JC, Rhodes C, Münzberg H. Leptin-Receptor-Expressing Neurons in the Dorsomedial Hypothalamus and Median Preoptic Area Regulate Sympathetic Brown Adipose Tissue Circuits. The Journal of Neuroscience. 2011; 31(5): 1873–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254.Tolson KP, Garcia C, Yen S, Simonds S, Stefanidis A, Lawrence A, Smith JT, Kauffman AS. Impaired kisspeptin signaling decreases metabolism and promotes glucose intolerance and obesity. Journal of Clinical Investigation. 2014; 124(7): 3075–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255.Tolson KP, Garcia C, Delgado I, Marooki N, Kauffman AS. Metabolism and energy expenditure, but not feeding or glucose tolerance, are impaired in young Kiss1r KO female mice. Endocrinology. 2016; doi: 10.1210/en.2016-1501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 256.Tolson KP, Marooki N, Wolfe A, Smith JT, Kauffman AS. Cre/lox generation of a novel whole-body Kiss1r KO mouse line recapitulates a hypogonadal, obese, and metabolically-impaired phenotype. Molecular and Cellular Endocrinology. 2019; 498110559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 257.Tolson KP, Marooki N, De Bond JP, Walenta E, Stephens SBZ, Liaw RB, Savur R, Wolfe A, Oh DY, Smith JT, Kauffman AS. Conditional knockout of kisspeptin signaling in brown adipose tissue increases metabolic rate and body temperature and lowers body weight. Faseb j. 2020; 34(1): 107–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258.Rezai-Zadeh K, Yu S, Jiang Y, Laque A, Schwartzenburg C, Morrison CD, Derbenev AV, Zsombok A, Münzberg H. Leptin receptor neurons in the dorsomedial hypothalamus are key regulators of energy expenditure and body weight, but not food intake. Molecular Metabolism. 2014; 3(7): 681–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 259.Faber CL, Deem JD, Phan BA, Doan TP, Ogimoto K, Mirzadeh Z, Schwartz MW, Morton GJ. Leptinreceptor neurons in the dorsomedial hypothalamus regulate diurnal patterns of feeding, locomotion, and metabolism. eLife. 2021; 10e63671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 260.Garfield AS, Shah BP, Burgess CR, Li MM, Li C, Steger JS, Madara JC, Campbell JN, Kroeger D, Scammell TE, Tannous BA, Myers MG Jr., Andermann ML, Krashes MJ, Lowell BB . Dynamic GABAergic afferent modulation of AgRP neurons. Nature Neuroscience. 2016; 19(12): 1628–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 261.Uchida Y, Kano M, Yasuhara S, Kobayashi A, Tokizawa K, Nagashima K. Estrogen modulates central and peripheral responses to cold in female rats. J Physiol Sci. 2010; 60(2): 151–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262.Laflamme N, Nappi RE, Drolet G, Labrie C, Rivest S. Expression and neuropeptidergic characterization of estrogen receptors (ERa and ERb) throughout the rat brain: Anatomical evidence of distinct roles of each subtype. Journal of Neurobiology. 1998; 36357–78. [DOI] [PubMed] [Google Scholar]
- 263.Sutton AK, Myers MG Jr., Olson DP. The Role of PVH Circuits in Leptin Action and Energy Balance. Annu Rev Physiol. 2016; 78207–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 264.Otgon-Uul Z, Suyama S, Onodera H, Yada T. Optogenetic activation of leptin- and glucose-regulated GABAergic neurons in dorsomedial hypothalamus promotes food intake via inhibitory synaptic transmission to paraventricular nucleus of hypothalamus. Molecular Metabolism. 2016; 5(8): 709–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 265.Furigo IC, Kim KW, Nagaishi VS, Ramos-Lobo AM, de Alencar A, Pedroso JA, Metzger M, Donato J Jr., Prolactin-sensitive neurons express estrogen receptor-α and depend on sex hormones for normal responsiveness to prolactin. Brain Res. 2014; 156647–59. [DOI] [PubMed] [Google Scholar]
- 266.Hrabovszky E, Kallo I, Steinhauser A, Merchenthaler I, Coen CW, Petersen SL, Liposits Z. Estrogen receptor-beta in oxytocin and vasopressin neurons of the rat and human hypothalamus: Immunocytochemical and in situ hybridization studies. The Journal of Comparative Neurology. 2004; 473(3): 315–33. [DOI] [PubMed] [Google Scholar]
- 267.Miller WJS, Suzuki S, Miller LK, Handa R, Uht RM. Estrogen receptor (ER) beta isoforms rather than ER alpha regulate corticotropin-releasing hormone promoter activity through an alternate pathway. The Journal of Neuroscience. 2004; 2410628–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 268.Asarian L, Geary N. Sex differences in the physiology of eating. American Journal of Physiology: Regulatory, Integrative and Comparative Physiology. 2013; 305(11): R1215–67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 269.Bowen DJ, Grunberg NE. Variations in food preference and consumption across the menstrual cycle. Physiology & Behavior. 1990; 47(2): 287–91. [DOI] [PubMed] [Google Scholar]
- 270.Fong AK, Kretsch MJ. Changes in dietary intake, urinary nitrogen, and urinary volume across the menstrual cycle. The American Journal of Clinical Nutrition. 1993; 57(1): 43–6. [DOI] [PubMed] [Google Scholar]
- 271.Geary N Handbook of behavioral neurobiology, Vol 14, Neurobiology of food and fluid intake. 2004. [Google Scholar]
- 272.Geary N Endocrine controls of eating: CCK, leptin, and ghrelin. Physiol Behav. 2004; 81(5): 719–33. [DOI] [PubMed] [Google Scholar]
- 273.Drewett RF. The meal patterns of the oestrous cycle and their motivational significance. Q J Exp Psychol. 1974; 26(Pt3): 489–94. [DOI] [PubMed] [Google Scholar]
- 274.Blaustein JD, Wade GN. Ovarian influences on the meal patterns of female rats. Physiol Behav. 1976; 17(2): 201–8. [DOI] [PubMed] [Google Scholar]
- 275.Asarian L, Geary N. Cyclic estradiol treatment normalizes body weight and restores physiological patterns of spontaneous feeding and sexual receptivity in ovariectomized rats. Hormones and Behavior. 2002; 42(4): 461–71. [DOI] [PubMed] [Google Scholar]
- 276.Morin LP, Fitzgerald KM, Zucker I. Estradiol shortens the period of hamster circadian rhythms. Science. 1977; 196(4287): 305–7. [DOI] [PubMed] [Google Scholar]
- 277.Takasu NN, Nakamura TJ, Tokuda IT, Todo T, Block GD, Nakamura W. Recovery from Age-Related Infertility under Environmental Light-Dark Cycles Adjusted to the Intrinsic Circadian Period. Cell Rep. 2015; 12(9): 1407–13. [DOI] [PubMed] [Google Scholar]
- 278.Colvin GB, Sawyer CH. Induction of running activity by intracerebral implants of estrogen in overiectomized rats. Neuroendocrinology. 1969; 4(4): 309–20. [DOI] [PubMed] [Google Scholar]
- 279.Ahdieh HB, Wade GN. Effects of hysterectomy on sexual receptivity, food intake, running wheel activity, and hypothalamic estrogen and progestin receptors in rats. Journal of Comparative and Physiological Psychology. 1982; 96(6): 886–92. [PubMed] [Google Scholar]
- 280.Shimomura Y, Shimizu H, Takahashi M, Sato N, Uehara Y, Fukatsu A, Negishi M, Kobayashi I, Kobayashi S. The significance of decreased ambulatory activity during the generation by long-term observation of obesity in ovariectomized rats. Physiology & Behavior. 1990; 47(1): 155–9. [DOI] [PubMed] [Google Scholar]
- 281.Geary N The estrogenic inhibition of eating. In: Stricker EM, Woods SC, eds. Handbook of behavioral neurobiology. 14, Neurobiology of food and fluid intake ed. New York, NY: Kluwer Academic/Plenum; 2007: 307–45. [Google Scholar]
- 282.Czaja JA, Goy RW. Ovarian hormones and food intake in female guinea pigs and rhesus monkeys. Hormones and Behavior. 1975; 6(4): 329–49. [DOI] [PubMed] [Google Scholar]
- 283.Butera PC, Czaja JA. Intracranial estradiol in ovariectomized guinea pigs: effects on ingestive behaviors and body weight. Brain Research. 1984; 322(1): 41–8. [DOI] [PubMed] [Google Scholar]
- 284.Czaja JA. Sex differences in the activational effects of gonadal hormones on food intake and body weight. Physiology & Behavior. 1984; 33(4): 553–8. [DOI] [PubMed] [Google Scholar]
- 285.McCaffrey TA, Czaja JA. Diverse effects of estradiol-17β: concurrent suppression of appetite, blood pressure and vascular reactivity in conscious, unrestrained animals. Physiology & Behavior. 1989; 45(3): 649–57. [DOI] [PubMed] [Google Scholar]
- 286.Heine PA, Taylor JA, Iwamoto GA, Lubahn DB, Cooke PS. Increased adipose tissue in male and female estrogen receptor-α knockout mice. Proceedings of the National Academy of Sciences of the United States of America. 2000; 97(23): 12729–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 287.Park CJ, Zhao Z, Glidewell-Kenney C, Lazic M, Chambon P, Krust A, Weiss J, Clegg DJ, Dunaif A, Jameson JL, Levine JE. Genetic rescue of nonclassical ERα signaling normalizes energy balance in obese Erα-null mutant mice. Journal of Clinical Investigation. 2011; 121604–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 288.Krege JH, Hodgin JB, Couse JF, Enmark E, Warner M, Mahler JF, Sar M, Korach KS, Gustafsson JA, Smithies O. Generation and reproductive phenotypes of mice lacking estrogen receptor beta. Proceedings of the National Academy of Sciences of the United States of America. 1998; 95(26): 15677–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 289.Antal MC, Krust A, Chambon P, Mark M. Sterility and absence of histopathological defects in nonreproductive organs of a mouse ERb-null mutant. Proceedings of the National Academy of Sciences of the United States of America. 2008; 1052433–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 290.Maneix L, Antonson P, Humire P, Rochel-Maia S, Castañeda J, Omoto Y, Kim HJ, Warner M, Gustafsson J. Estrogen receptor β exon 3-deleted mouse: The importance of non-ERE pathways in ERβ signaling. Proc Natl Acad Sci U S A. 2015; 112(16): 5135–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 291.Davis KE, Carstens EJ, Irani BG, Gent LM, Hahner LM, Clegg DJ. Sexually dimorphic role of G proteincoupled estrogen receptor (GPER) in modulating energy homeostasis. Hormones and Behavior. 2014; 66(1): 196–207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 292.Krause WC, Rodriguez R, Gegenhuber B, Matharu N, Rodriguez AN, Padilla-Roger AM, Toma K, Herber CB, Correa SM, Duan X, Ahituv N, Tollkuhn J, Ingraham HA. Oestrogen engages brain MC4R signalling to drive physical activity in female mice. Nature. 2021; 599(7883): 131–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 293.Wise PM, Scarbrough K, Weiland NG, Larson GH. Diurnal pattern of proopiomelanocortin gene expression in the arcuate nucleus of proestrous, ovariectomized, and steroid-treated rats: a possible role in cyclic luteinizing hormone secretion. Molecular Endocrinology. 1990; 4(6): 886–92. [DOI] [PubMed] [Google Scholar]
- 294.Bohler HC Jr., Tracer H, Merriam GR, Petersen SL. Changes in proopiomelanocortin messenger ribonucleic acid levels in the rostral periarcuate region of the female rat during the estrous cycle. Endocrinology. 1991; 128(3): 1265–9. [DOI] [PubMed] [Google Scholar]
- 295.Martínez de Morentin PB, González-García I, Martins L, Lage R, Fernández-Mallo D, Martínez-Sánchez N, Ruíz-Pino F, Liu J, Morgan DA, Pinilla L, Gallego R, Saha AK, Kalsbeek A, Fliers E, Bisschop PH, Diéguez C, Nogueiras R, Rahmouni K, Tena-Sempere M, López M. Estradiol regulates brown adipose tissue thermogenesis via hypothalamic AMPK. Cell Metab. 2014; 20(1): 41–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 296.Pelletier G, Li S, Luu-The V, Labrie F. Oestrogenic regulation of pro-opiomelanocortin, neuropeptide Y and corticotrophin-releasing hormone mRNAs in mouse hypothalamus. Journal of Neuroendocrinology. 2007; 19(6): 426–31. [DOI] [PubMed] [Google Scholar]
- 297.Bethea CL, Hess DL, Widmann AA, Henningfeld JM. Effects of progesterone on prolactin, hypothalamic β-endorphin, hypothalamic substance P, and midbrain serotonin in guinea pigs. Neuroendocrinology. 1995; 61(6): 695–703. [DOI] [PubMed] [Google Scholar]
- 298.Thornton JE, Loose MD, Kelly MJ, Rønnekleiv OK. Effects of estrogen on the number of neurons expressing β-endorphin in the medial basal hypothalamus of the female guinea pig. The Journal of Comparative Neurology. 1994; 34168–77. [DOI] [PubMed] [Google Scholar]
- 299.Herman MA, Ackermann F, Trimbuch T, Rosenmund C. Vesicular glutamate transporter expression level affects synaptic vesicle release probability at hippocampal synapses in culture. The Journal of Neuroscience. 2014; 34(35): 11781–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 300.Qiu J, Rønnekleiv OK, Kelly MJ. Modulation of hypothalamic neuronal activity through a novel G-protein coupled estrogen membrane receptor. Steroids. 2008; 73(9–10): 985–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 301.Qiu J, Bosch MA, Jamali K, Xue C, Kelly MJ, Rønnekleiv OK. Estrogen upregulates T-type calcium channels in the hypothalamus and pituitary. The Journal of Neuroscience. 2006; 26(43): 11072–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 302.Smith AW, Bosch MA, Wagner EJ, Rønnekleiv OK, Kelly MJ. The membrane estrogen receptor ligand STX rapidly enhances GABAergic signaling in NPY/AgRP neurons: Role in mediating the anorexigenic effects of 17β-estradiol. American Journal of Physiology: Endocrinology and Metabolism. 2013; 305(5): E632–E40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 303.Mounien L, Bizet P, Boutelet I, Vaudry H, Jegou S. Expression of melanocortin MC3 and MC4 receptor mRNAs by neuropeptide Y neurons in the rat arcuate nucleus. Neuroendocrinology. 2005; 82(3–4): 164–70. [DOI] [PubMed] [Google Scholar]
- 304.Kavalali ET. The mechanisms and functions of spontaneous neurotransmitter release. Nature Reviews Neuroscience. 2015; 16(1): 5–16. [DOI] [PubMed] [Google Scholar]
- 305.Lee S, Zhang Y, Chen M, Zhou ZJ. Segregated glycine-glutamate co-transmission from vGluT3 amacrine cells to contrast-suppressed and contrast-enhanced retinal circuits. Neuron. 2016; 90(1): 27–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 306.Nishimaru H, Restrepo CE, Ryge J, Yanagawa Y, Kiehn O. Mammalian motor neurons corelease glutamate and acetylcholine at central synapses. Proceedings of the National Academy of Sciences of the United States of America. 2005; 102(14): 5245–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 307.Fortin GM, Ducrot C, Giguère N, Kouwenhoven WM, Bourque M-J, Pacelli C, Varaschin RK, Brill M, Singh S, Wiseman PW, Trudeau L-É. Segregation of dopamine and glutamate release sites in dopamine neuron axons: regulation by striatal target cells. The FASEB Journal. 2018; 33(1): 400–17. [DOI] [PubMed] [Google Scholar]
- 308.Pinto S, Roseberry AG, Liu H, Diano S, Shanabrough M, Cai X, Friedman JM, Horvath TL. Rapid rewiring of arcuate nucleus feeding circuits by leptin. Science. 2004; 304(5667): 110–5. [DOI] [PubMed] [Google Scholar]
- 309.Olofsson LE, Pierce AA, Xu AW. Functional requirement of AgRP and NPY neurons in ovarian cycledependent regulation of food intake. Proceedings of the National Academy of Sciences of the United States of America. 2009; 106(37): 15932–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 310.Smith AW, Rønnekleiv OK, Kelly MJ. Gq-mER signaling has opposite effects on hypothalamic orexigenic and anorexigenic neurons. Steroids. 2014; 8131–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 311.Roepke TA, Qiu J, Smith AW, Rønnekleiv OK, Kelly MJ. Fasting and 17β-estradiol differentially modulate the M-current in neuropeptide Y neurons. The Journal of Neuroscience. 2011; 17(33): 11825–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 312.Yang Y, Atasoy D, Su HH, Sternson SM. Hunger states switch a flip-flop memory circuit via a synaptic AMPK-dependent positive feedback loop. Cell. 2011; 146(6): 992–1003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 313.Moss RL, Kelly M, Riskind P. Tuberoinfundibular neurons: dopaminergic and norepinephrinergic sensitivity. Brain Research. 1975; 89(2): 265–77. [DOI] [PubMed] [Google Scholar]
- 314.Galea LA, Wide JK, Paine TA, Holmes MM, Ormerod BK, Floresco SB. High levels of estradiol disrupt conditioned place preference learning, stimulus response learning and reference memory but have limited effects on working memory. Behavioural Brain Research. 2001; 126(1–2): 115–26. [DOI] [PubMed] [Google Scholar]
- 315.Roa J, Herbison AE. Direct regulation of GnRH neuron excitability by arcuate nucleus POMC and NPY neuron neuropeptides in female mice. Endocrinology. 2012; 153(11): 5587–99. [DOI] [PubMed] [Google Scholar]
- 316.Lagrange AH, Rønnekleiv OK, Kelly MJ. Modulation of G protein-coupled receptors by an estrogen receptor that activates protein kinase A. Molecular Pharmacology. 1997; 51605–12. [DOI] [PubMed] [Google Scholar]