

Abstract
Background
Hippoboscid flies (Diptera: Hippoboscidae), also known as louse flies or keds, are obligate blood-sucking ectoparasites of animals, and accidentally of humans. The potential role of hippoboscids as vectors of human and veterinary pathogens is being increasingly investigated, but the presence and distribution of infectious agents in louse flies is still unknown in parts of Europe. Here, we report the use of molecular genetics to detect and characterize vector-borne pathogens in hippoboscid flies infesting domestic and wild animals in Austria.
Methods
Louse flies were collected from naturally infested cattle (n = 25), sheep (n = 3), and red deer (n = 12) across Austria between 2015 and 2019. Individual insects were morphologically identified to species level and subjected to DNA extraction for molecular pathogen screening and barcoding. Genomic DNA from each louse fly was screened for Borrelia spp., Bartonella spp., Trypanosomatida, Anaplasmataceae, Filarioidea and Piroplasmida. Obtained sequences of Trypanosomatida and Bartonella spp. were further characterized by phylogenetic and haplotype networking analyses.
Results
A total of 282 hippoboscid flies corresponding to three species were identified: Hippobosca equina (n = 62) collected from cattle, Melophagus ovinus (n = 100) from sheep and Lipoptena cervi (n = 120) from red deer (Cervus elaphus). Molecular screening revealed pathogen DNA in 54.3% of hippoboscids, including infections with single (63.39%), two (30.71%) and up to three (5.90%) distinct pathogens in the same individual. Bartonella DNA was detected in 36.9% of the louse flies. Lipoptena cervi were infected with 10 distinct and previously unreported Bartonella sp. haplotypes, some closely associated with strains of zoonotic potential. DNA of trypanosomatids was identified in 34% of hippoboscids, including the first description of Trypanosoma sp. in H. equina. Anaplasmataceae DNA (Wolbachia spp.) was detected only in M. ovinus (16%), while < 1% of the louse flies were positive for Borrelia spp. and Filarioidea. All hippoboscids were negative for Piroplasmida.
Conclusions
Molecular genetic screening confirmed the presence of several pathogens in hippoboscids infesting domestic and wild ruminants in Austria, including novel pathogen haplotypes of zoonotic potential (e.g. Bartonella spp.) and the first report of Trypanosoma sp. in H. equina, suggesting a potential role of this louse fly as vector of animal trypanosomatids. Experimental transmission studies and expanded monitoring of hippoboscid flies and hippoboscid-associated pathogens are warranted to clarify the competence of these ectoparasites as vectors of infectious agents in a One-Health context.
Graphical Abstract

Supplementary Information
The online version contains supplementary material available at 10.1186/s13071-023-05810-4.
Keywords: Bartonella, Hippobosca equina, Hippoboscidae, Lipoptena cervi, Melophagus ovinus, Vector-borne pathogens, Barcoding, Louse flies, Keds, Ruminants
Background
Hippoboscid flies (Diptera: Hippoboscidae), also known as louse flies or keds, are obligatory blood-sucking ectoparasites infesting mammals and birds worldwide [1]. To date, most research on hippoboscids has focused on understanding their biology, evolution, host specificity and impact of their hematophagous and biting behavior on animals and humans [2–8]. Various louse fly species of the genera Melophagus spp., Lipoptena spp. and Hippobosca spp. have been described to commonly infest domestic and wild ungulates in Europe [9–11], and occasionally also attack humans and pets [12–15]. Indeed, it appears that hippoboscid flies may have been attacking humans for millennia, as suggested by the identification of the common deer ked Lipoptena cervi on the late neolithic human mummy “Ötzi” in the Ötztal Alps [16]. Considering their blood-feeding nature, widespread distribution and the broad host spectrum of some species, hippoboscid flies may also act as potential vectors of infectious diseases within animal populations, and between animals and humans [17].
Hippoboscid flies have been investigated for their role as vectors of animal pathogens for over a century [18, 19], with molecular studies in the last 2 decades confirming several hippoboscid-associated pathogens of medical and veterinary importance in different louse fly species [17]. A wide range of vector-borne bacteria and protozoa have been identified in hippoboscid flies collected from domestic and wild ruminants in some European countries, including Anaplasma spp., Babesia spp., Bartonella spp., Borrelia spp., Mycoplasma spp., Rickettsia spp., Theileria spp. and Trypanosoma spp. [20–31]. Despite these research efforts, there are still major knowledge gaps regarding the presence and monitoring of emerging vector-borne diseases in hippoboscid flies in Europe, including Austria. Moreover, in view of the widespread distribution of free-ranging wild ruminants that can act as reservoirs of infectious agents in Austria [32, 33] and the increasing human presence in areas populated by wild animals due to working or leisure activities, the vector role of hippoboscids warrants further elucidation.
The aim of the present study was to detect the presence of vector-borne pathogens in hippoboscid flies infesting domestic and wild ruminants in Austria using molecular techniques. In addition, DNA barcoding of the hippoboscid flies was performed to confirm and characterize their identity.
Materials and methods
Study areas and collection of hippoboscid flies
Hippoboscid flies were collected from red deer (Cervus elaphus; n = 12), sheep (Ovis aries; n = 3) and cattle (Bos taurus; n = 25) in various locations in Austria, between 2015 and 2019 (Fig. 1). Hippoboscids infesting red deer were sampled in November/December of 2016 and 2017 from hunted animals at three sites: Schwaz (Fig. 1A) and Kufstein (Fig. 1B) in the Federal State of Tyrol and Bludenz (Fig. 1C) in the Federal State of Vorarlberg. Hunted red deer from these areas are routinely examined as part of the tuberculosis surveillance in wildlife by the Austrian Agency for Health and Food Safety (AGES). Hippoboscids were collected from the head skin of 12 recently hunted red deer submitted to the AGES Laboratory, with an estimated presence of deer keds in 20–30% of all hunted red deer investigated in the surveillance program (W. Glawischnig, personal communication). A total of 120 louse flies were sampled from the examined animals. Louse flies from sheep were obtained in March 2018 directly at a farm in Leobersdorf, Federal State of Lower Austria (Fig. 1D). At sampling, the farm had a herd of 30 adult sheep, with an observed presence of keds in 100% of the animals. A total of 100 sheep keds were collected directly from 3 adult sheep during shearing. Hippoboscids from cattle were collected from grazing animals in July/August of 2016 and 2017 in the Saalfelden area, Federal State of Salzburg (Fig. 1E), in the course of a 2-year epidemiological study involving inspection of the animals at regular intervals during the grazing season [34]. Per occasion, 31 to 57 cattle were visually examined, and louse flies were observed in up to 33% of the cattle (peak of infestation in August). A total of 61 hippoboscid individuals were collected from 24 cattle from Saalfelden between 2016 and 2017 (with some louse flies being collected from the same animal) after visual identification of the insects on the cattle’s haircoat. An additional louse fly from cattle was collected in Eisenstadt in July 2019 (Federal Sate of Burgenland, Fig. 1F). At all sampling sites, hippoboscids were separated from hair/wool manually or using fine forceps and stored immediately either dry or in ethanol in individual Eppendorf tubes. All hippoboscids were identified to species level using a stereomicroscope (Nikon SMZ1270, Tokyo, Japan) and morphological keys [35, 36], followed by DNA extraction.
Fig. 1.

Origin of hippoboscid flies from Austria. Collection sites of hippoboscid flies from domestic and wild ruminants in Schwaz (A), Kufstein (B), Bludenz (C), Leobersdorf (D), Saalfelden (E) and Eisenstadt (F), in Austria. See the main text for further information
DNA extraction
The individual hippoboscids were subjected to total DNA extraction for molecular pathogen screening and insects’ barcoding. Individual hippoboscids were mixed with 180 μl buffer ATL (DNeasy Blood & Tissue Kits, Qiagen) in 1.5-ml Eppendorf tubes, and two 1.4-mm ceramic beads (Qiagen, Hilden, Germany) were added per tube, followed by mechanical homogenization in a TissueLyser II (Qiagen, Hilden, Germany) at room temperature for 6 min. Then, 20 μl proteinase K was added, and the tubes were vortexed and incubated at 56 °C overnight. After incubation, total DNA was extracted from insect material using the QIAGEN DNeasy Blood & Tissue Kit (QIAGEN, Hilden, Germany), following the manufacturer’s instructions.
Barcoding of hippoboscid flies
To confirm the species identity and to explore the genetic diversity of the collected hippoboscids, selected specimens were subjected to DNA barcoding analysis. Total DNA extracted from 21 hippoboscids was used to amplify a region within the insects’ mitochondrial COI by conventional PCR [37] as described in Table 1. The PCR products were sequenced at LGC Genomics GmbH (Berlin, Germany). The resulting COI sequences were used for the taxonomical characterization of the hippoboscid species by comparison with available sequences on the GenBank nucleotide database for organism identification using BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi) and BOLD (www.boldsystems.org, accessed on 01 June 2022).
Table 1.
Primers and PCR cycle conditions used for the molecular characterization of pathogens and hippoboscid flies (Diptera: Hippoboscidae) collected from domestic and wild ruminants in Austria
| Organism | Target gene | Primers | Sequence (5'–3') | Length (bp) | PCR cycle conditions | References |
|---|---|---|---|---|---|---|
| Anaplasmataceae | 16S rRNA | EHR16SD_for | GGT ACC YAC AGA AGA AGT CC | 345 | 95 °C/2 min; 35 cycles: 94 °C/1 min, 54 °C/30 s, 72 °C/30 s; 72 °C/5 min | [40] |
| EHR16SR_rev | TAG CAC TCA TCG TTT ACA GC | |||||
| Borrelia | 16S rRNA | Borr_allg_for | ACG CTG GCA GTG CGT CTT AA | 674 | 94 °C/2 min; 6 cycles: 94 °C/1 min, 45 °C/1.5 min, 72 °C/75 s; 36 cycles: 94 °C/1 min, 51 °C/1.5 min, 72 °C/75 s); 72 °C/5 min | [39] |
| Borr_allg_rev | CTG ATA TCA ACA GAT TCC ACC C | |||||
| Bartonella | gltA | BhCs.781p | GGG GAC CAG CTC ATG GTG G | 379 | 94 °C/5 min; 40 cycles: 94 °C/1 min, 54 °C/1 min, 72 °C/1 min; 72 °C/10 min | [38] |
| BhCs.1137n | AAT GCA AAA AGA ACA GTA AAC A | |||||
| Filarioidea | COI | COlint-F | TGA TTG GTG GTT TTG GTA A | 668 | 94 °C/2 min; 8 cycles: 94 °C/45 s, 51 °C/45 s, 72 °C/1.5 min; 25 cycles: 94 °C/45 s, 45 °C/45 s, 72 °C/1.5 min; 72 °C/7 min | [42] |
| COlint-R | ATA AGT ACG AGT ATC AAT ATC | |||||
| Trypanosomatida | 18S rRNA | Tryp_18S_F1 (Nest 1) | GTGGACTGCCATGGCGTTGA | 960 | 94 °C/5 min; 35 cycles: 94 °C/1 min, 56 °C/1 min, 72 °C/1 min; 72 °C/5 min (Nest 1) | This study |
| Tryp_18S_R1 (Nest 1) | CAGCTTGGATCTCGTCCGTTGA | |||||
| Tryp_18S_F2 (Nest 2) | CGATGAGGCAGCGAAAAGAAATAGAG | 94 °C/5 min; 25 cycles: 94 °C/1 min, 56 °C/1 min, 72 °C/1 min; 72 °C/5 min (Nest 2) | ||||
| Tryp_18S_R2 (Nest 2) | GACTGTAACCTCAAAGCTTTCGCG | |||||
| Piroplasmida | 18S rRNA | BTH-1F (Nest 1) | CCT GAG AAA CGG CTA CCA CAT CT | 561 | 94 °C/2 min; 40 cycles: 95 °C/30 s, 68 °C/1 min, 72 °C/1 min; 72 °C/10 min (Nest 1) | [41] |
| BTH-1R (Nest 1) | TTG CGA CCA TAC TCC CCC CA | |||||
| G-2_for (Nest 2) | GTC TTG TAA TTG GAA TGA TGG | 94 °C/2 min; 40 cycles: 95 °C/30 s, 60 °C/1 min, 72 °C/1 min; 72 °C/10 min (Nest 2) | ||||
| G-2_rev (Nest 2) | CCA AAG ACT TTG ATT TCT CTC | |||||
| Lepidoptera | COI | LepF1 | ATT CAA CCA ATC ATA AA | 648 | 94 °C/2 min; 6 cycles: 94 °C/1 min, 45 °C/1.5 min, 72 °C/75 s; 36 cycles: 94 °C/1 min, 51 °C/1.5 min, 72 °C/75 s); 72 °C/5 min | [37] |
| LepR1 | TAA ACT TCT GGA TGT CAA AAA |
Molecular pathogen screening by conventional and nested PCR
The obtained DNA from each hippoboscid fly was screened for the presence of several vector-borne pathogens by targeting selected genes using primers and PCR protocols summarized in Table 1. The hippoboscids were screened by conventional PCR for bacteria of the family Anaplasmataceae (16S ribosomal RNA), the genus Borrelia (16S ribosomal RNA) and the genus Bartonella (citrate synthase gene—gltA), as well as for nematodes of the superfamily Filarioidea (mitochondrial cytochrome c oxidase subunit I gene—COI). In addition, the DNA was screened for parasites of the orders Trypanosomatida (18S ribosomal RNA) and Piroplasmida (18S ribosomal RNA) using nested PCRs. The PCR methodologies were based on previously published protocols [38–42], except for the nested PCR protocol for Trypanosomatida, which was designed for the present study. The latter primers were designed based on all 18S sequences of Trypanosomatida available on GenBank and allow the amplification of all strains. All PCR reactions were performed in an Eppendorf Mastercycler Pro (Eppendorf AG, Hamburg, Germany). The PCR products were stored at 15 °C until confirmation of the amplified regions of interest by electrophoresis in 2% agarose gels stained with Midori Green Advanced dye (Biozym Scientific, Germany). PCR products positive for the investigated pathogens were sequenced at LGC Genomics GmbH (Berlin, Germany) using amplification primers. The sequences were assembled with BioEdit [43] and compared to sequences available on NCBI GenBank (National Center for Biotechnology Information; https://blast.ncbi.nlm.nih.gov/Blast.cgi) using multiple BLAST searches.
Phylogenetic and haplotype networking analyses of Trypanosomatida and Bartonella spp.
Selected sequences of Trypanosomatida and Bartonella spp. isolated from the investigated hippoboscids were subjected to phylogenetic analyses as previously described [44], with modifications. The sequences were aligned and cut to primer binding regions, and the electropherograms were manually checked for double peaks. Double peaks were identified in 19/27 Bartonella spp. sequences, which suggested a co-infection with two distinct strains of Bartonella sp. in the same insects. In those cases, the two strains were unphased to obtain single sequences and uploaded as individual sequences to GenBank. Each strain was separately uploaded to GenBank (acc. no. ON637624—ON637640 for Trypanosomatida; OP198738—OP198806 for Bartonella spp.) and used for phylogenetic analysis.
To provide an overview of the genetic diversity of detected (and related) Trypanosomatida and Bartonella spp. strains, maximum likelihood (ML) and Bayesian inference (BI) trees were calculated for each of the two groups based on alignments including 409 sequences (991 nucleotide positions) for Trypanosoma spp. and 582 sequences (338 nucleotide positions) for Bartonella spp. Gaps in the alignments were removed using TrimAl v.1.3 [45], and the sequences were collapsed to haplotypes using DAMBE v.7.0.5.1 [46], leaving 167 haplotypes (701 nucleotide positions) for Trypanosoma spp. and 261 haplotypes for Bartonella spp. As outgroup of the Trypanosoma spp. tree, a sequence of Belchomonas wendygibsoni (KF054126) was used. No suitable sequence was available as outgroup for Bartonella spp., and this tree was instead mid-point rooted. The ML bootstrap consensus trees (1000 replicates) were calculated using the W-IQ-TREE web server [47] and applying the models TIM3e + I + G4 for Trypanosoma spp. and JC for Bartonella spp., which were suggested as best fit for the data set in the model test according to the corrected Akaike information criterion. The BI trees were calculated using MrBayes v.3.2.7 [48] applying the next complex model GTR + G + I for Trypanosoma spp. and JC for Bartonella spp. The analyses were run for 10,000,000 generations (number of chains: 4), sampling every thousandth tree. The first 25% of trees were discarded as burn-in, and a 50% majority-rule consensus tree was calculated based on the remaining 7500 trees. The sequences for the DNA haplotype network analyses were selected based on well-supported clades in the phylogenetic trees (see Additional file 1: Figs. S1 and S2). Median-joining haplotype networks were calculated with Network 10.2.0.0 (Fluxus Technology Ltd., Suffolk, UK), applying the default settings. The networks were graphically prepared and provided with information on the countries and hosts in Network Publisher v.2.1.2.5 (Fluxus Technology Ltd., Suffolk, UK) and finalized with CorelDRAW 2021 (Corel, Ottawa, Canada).
Data analysis
Data processing and descriptive statistics were performed in Microsoft Excel and GraphPad Prism 7. Statistical analyses were implemented in R version 4.0.3 [49]. Differences in total infection rate (all pathogen groups combined) and in infection prevalence of each pathogen group among the three hippoboscid species were evaluated by the test of equal or given proportions (prop.test) and pairwise comparison for proportions with Holm-Bonferroni method (pairwise.prop.test). The risks of each hippoboscid species to be infected with only one or with two concurrent pathogens in the same individual (positive/negative) were evaluated with logistic regression models (glm, family: “binomial”) using louse fly species as explanatory variable to calculate odds ratios (OR) and 95% confidence intervals (95% CI). A level of P < 0.05 was considered significant.
Results
Species and DNA barcoding of collected hippoboscid flies
A total of 282 louse flies were collected from naturally infested cattle (n = 25), sheep (n = 3) and red deer (n = 12) in different regions of Austria. The hippoboscids were identified as Hippobosca equina (n = 62; Fig. 2A) collected from cattle, Melophagus ovinus (n = 100; Fig. 2B) from sheep and L. cervi (n = 120; Fig. 2C) from red deer. Barcode analyses in BOLD of 21 individual hippoboscids (H. equina, n = 5; M. ovinus, n = 5; L. cervi, n = 11) revealed that the COI sequences of each louse fly species clustered within a respective Barcode Index Number with sequences previously reported from Europe, Northern Africa and Asia for H. equina (BOLD:AAX0882), M. ovinus (BOLD:AAX4771) and L. cervi (BOLD:ABX1452). Obtained COI sequences from the barcoded hippoboscids were submitted to GenBank under the following accession numbers: ON129173, ON129175, ON129176, ON129178, ON129181 (H. equina); ON129174, ON129177, ON129179, ON129180, ON129182 (M. ovinus); ON341137 – ON341147 (L. cervi).
Fig. 2.

Hippoboscid flies for molecular pathogen screening. Representative individuals of Hippobosca equina (A), Melophagus ovinus (B) and Lipoptena cervi (C) collected from domestic and wild ruminants in Austria
Molecular pathogen screening
Molecular screening revealed pathogen DNA in 153/282 (54.3%) of the collected hippoboscids, with substantial differences between louse fly species (Table 2). The sheep ked M. ovinus were significantly more frequently infected with pathogens in comparison with L. cervi (χ2 = 73.944, df = 1, P < 0.001) and H. equina (χ2 = 82.315, df = 1, P < 0.001), whereas no difference in total infection rate was observed between L. cervi and H. equina (χ2 = 2.7502, df = 1, P = 0.09; Table 2). Of all 153 positive individuals among the three hippoboscid species, 97 carried only one pathogen (63.4%), 47 were infected with two different pathogens (30.7%), and three distinct pathogens were confirmed in nine M. ovinus specimens (5.9%). The percentages of hippoboscids from each species infected with single or multiple pathogens in the same individuals are illustrated in Fig. 3. From the three hippoboscid species, M. ovinus had significantly higher odds to be infected with at least one pathogen compared with H. equina (OR M. ovinus: 2.8 [1.4–5.9], P < 0.01), whereas L. cervi had slightly higher odds to be infected with a single pathogen compared with H. equina (OR L. cervi [95% CI] 1.9 [0.95–4.05], P = 0.07). Melophagus ovinus was significantly most likely to be infected with two concurrent pathogens compared with H. equina (OR = 47.1 [9.8–849.2], P < 0.001). No differences were observed in the risk of carrying two pathogens between L. cervi and H. equina (OR = 1.02 [0.1–22.1, P > 0.5].
Table 2.
Pathogens detected by molecular screening in hippoboscid flies (Diptera: Hippoboscidae) collected from domestic and wild ruminants in Austria
| Host (n) | Hippoboscid flies (n) | Hippoboscids positive to pathogens/total hippoboscids screened per species (% positive) | ||||||
|---|---|---|---|---|---|---|---|---|
| Total infection rate (Positive to ≥ 1 pathogen[s]) | Bartonella | Trypanosomatida | Anaplasmataceae | Borrelia | Filarioidea | Piroplasmida | ||
| Cattle (n = 25) | Hippobosca equina (n = 62) | 14/62 (22.5%)a | 12/62 (19.3%)a | 3/62 (4.8%)a | 0/62 | 0/62 | 0/62 | 0/62 |
| Sheep (n = 3) | Melophagus ovinus (n = 100) | 96/100 (96%)b | 54/100 (54%)b | 87/100 (87%)b | 16/100 (16%) | 1/100 (100%) | 0/100 | 0/100 |
| Red deer (n = 12) | Lipoptena cervi (n = 120) | 43/120 (35.8%)a | 38/120 (31.6%)a | 6/120 (5%)a | 0/120 | 0/120 | 1/120 (0.8%) | 0/120 |
a,bDifferent letters within the same column (pathogen) represent statistically significant differences in the proportion of pathogen-positive individuals between hippoboscid species (P < 0.05)
Fig. 3.
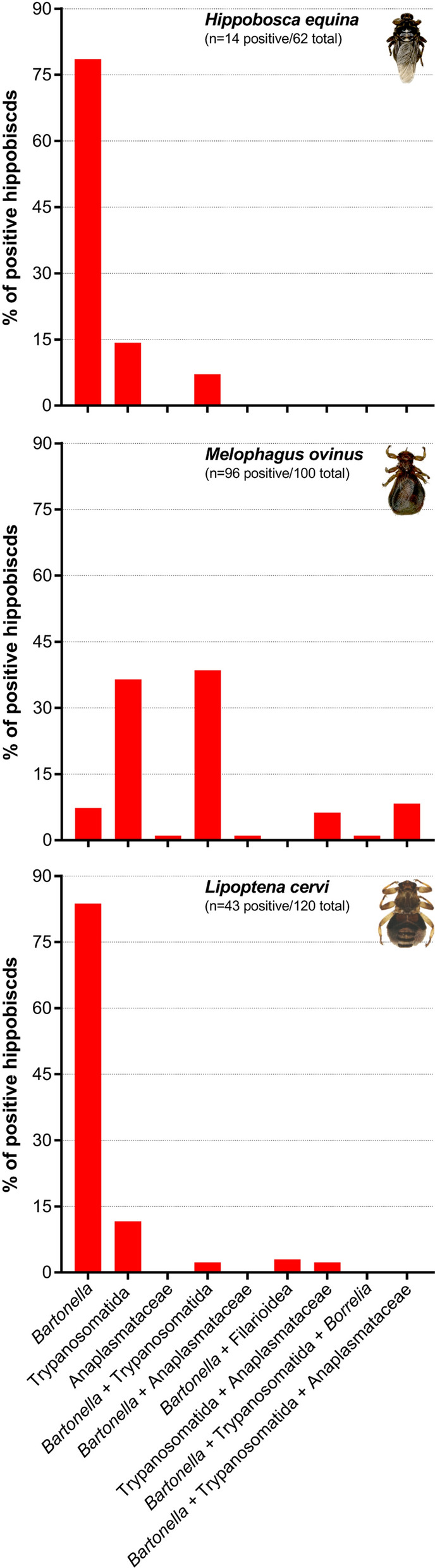
Single and co-infections in pathogen-positive hippoboscid flies. Percentage of hippoboscid flies positive to one, two or three pathogens in the same individual of Hippobosca equina, Melophagus ovinus and Lipoptena cervi collected from domestic and wild ruminants in Austria
Bartonella spp. was detected in 36.9% (104/282) of the investigated hippoboscids, followed by Trypanosomatida in 34.0% of all louse flies (96/282). Individuals of M. ovinus were found more frequently infected with Bartonella spp. in comparison with H. equina (χ2 = 17.619, df = 1, P < 0.001) and with L. cervi (χ2 = 10.283, df = 1, P < 0.001). The sheep ked M. ovinus were also more often infected with Trypanosomatida compared to H. equina (χ2 = 101.33, df = 1, P < 0.001) and to L. cervi (χ2 = 146.95, df = 1, P < 0.001). Only M. ovinus individuals were positive to Anaplasmataceae in 5.7% (16/282; 5.7% of all investigated hippoboscids) and less than 1% of the louse flies were positive for Borrelia spp. (one M. ovinus specimen) and Filarioidea (one L. cervi specimen). All investigated hippoboscids were negative for Piroplasmida. The sequences of the pathogens screened in the present study were deposited in GenBank under the following acc. no.: ON668330 (Borrelia spp.), ON678056 (Filarioidea), ON637624 – ON637640 (Trypanosomatida) and OP198738 – OP198806 (Bartonella spp.).
Bartonella spp. were the most common infectious agents detected in H. equina and L. cervi, and the second most frequently identified pathogen in M. ovinus after trypanosomatids (Table 2, Fig. 3). In L. cervi, several isolated Bartonella spp. gltA (citrate synthase gene) sequences showed 100% identity with a Bartonella sp. strain isolated from the bat Miniopterus schreibersii in Hungary (MK140014; see Additional file 2: Table S1). Furthermore, various Bartonella spp. gltA sequences from L. cervi showed > 99% identity with reported sequences of B. schoenbuchensis isolated from roe deer (Capreolus capreolus; GenBank acc. no.: AJ278184; AJ278185) and from L. cervi (AJ564634; AJ564635; Additional file 2) in Germany. Haplotype network analyses of the isolated Bartonella spp. gltA sequences from L. cervi revealed ten novel strains not previously reported (Fig. 4 and Additional file 1), including one strain (OP198738) identical to a Bartonella sp. sequence isolated from the bat M. schreibersii in Hungary (Fig. 4). In contrast to the broad diversity of Bartonella spp. strains detected in L. cervi, only one haplotype was identified in sequences isolated from H. equina (OP198794), which was 100% identical to sequences of Bartonella chomelii reported from Spain, France and New Caledonia (KM215691; KM215690; JN646657; Fig. 4 and Additional file 1). The Bartonella spp. sequences identified in M. ovinus (OP198802) showed 100% identity to sequences of Candidatus Bartonella melophagi from M. ovinus in Peru, the USA and China and from a European hedgehog (Erinaceus europaeus) in Czechia (MZ089835; MT154632; Fig. 4; Additional file 2). In the BI tree (Additional file 1), the sequences of B. chomelii, Candidatus B. melophagi and Bartonella sp. clustered in one clade with other Bartonella spp. previously reported from ruminants (BI posterior probability [BI pp] = 1.0, ML bootstrap value [ML bs] = 99). Most sequences of B. chomelii, Candidatus B. melophagi and Bartonella spp. detected in the present study clustered in one subclade with B. chomelii, B. schoenbuchensis, B. capreoli and Bartonella spp. sequences (BI = 1, ML = 100; Additional file 1). Only one Bartonella sp. sequence from L. cervi (OP198746) was placed in a separate sister clade together with B. bovis and Bartonella spp. (BI = 0.98, ML = 85).
Fig. 4.

Genetic diversity of Bartonella detected in hippoboscid flies. Median-joining haplotype network of the gltA sequences (338 bp) of selected Bartonella spp. from the present and previous studies showing their geographical distribution (A) and the reported hosts (B). Circles represent haplotypes, and numbers within the circles represent the number of individuals. If no number is shown, then only one individual is represented. Labels next to circles specify representative GenBank accession numbers of the haplotypes; white circles represent intermediate nodes; bars on branches interconnecting haplotypes represent the number of substitutions. Asterisks mark haplotypes detected in the present study
Trypanosomatid sequences (18S rRNA) were detected in all hippoboscid species and represented the most common pathogens in M. ovinus with 87% positive individuals (Table 2, Fig. 3). Trypanosoma spp. sequences isolated from M. ovinus were 100% identical to 18S sequences of Trypanosoma melophagium from Czechia, Croatia and the UK (OM256700; HQ664912; FN666409). The DNA haplotype network analysis revealed two distinct strains of T. melophagium: a new strain (ON637626) and a second one (ON637624) identical to T. melophagium sequences isolated from M. ovinus in Croatia, UK and Czechia (Fig. 5 and Additional file 1). Only three H. equina individuals (two from Saalfelden and one from Eisenstadt) were positive for trypanosomatids, featuring a Trypanosoma sp. (ON637634) that clustered together with other trypanosomatids of ruminants, including Trypanosoma theileri, T. trinaperronei, T. melophagium, T. cervi and Trypanosoma spp. (Fig. 5 and Additional file 1). The Trypanosoma sequences obtained from H. equina shared > 98% similarity with those of Trypanosoma cf. cervi isolated from white-tailed deer in the USA (JX178196), Trypanosoma sp. from horse flies in Russia (MK156792-MK15794) and T. theileri obtained from tsetse flies in the Central African Republic (KR024688). While the trypanosomatid sequences isolated from L. cervi showed > 99% identity to sequences of non-parasitic kinetoplastids of the genus Bodo from the UK and USA (AY425015; AY028450).
Fig. 5.

Genetic diversity of Trypanosoma detected in hippoboscid flies. Median-joining haplotype network of the 18S rRNA sequences (779 bp) of selected Trypanosoma spp. from the present and previous studies showing their geographical distribution (A) and the reported hosts (B). Circles represent haplotypes and numbers within the circles represent the number of individuals. If no number is shown, then only one individual is represented. Labels next to circles specify representative GenBank accession numbers of the haplotypes; white circles represent intermediate nodes; bars on branches interconnecting haplotypes represent the number of substitutions. Asterisks mark haplotypes detected in the present study
Anaplasmataceae sequences (16S rRNA) were only detected in 16 M. ovinus individuals, featuring sequences identical to those of several Wolbachia strains, including a strain previously isolated from M. ovinus (MF461472; KY224164; KY224163). Borrelia spp. (16S rRNA) was detected in a single M. ovinus and had a 93.7% similarity with a reported Borrelia sp. (CP043682) isolated from ticks associated with passeriform birds. Finally, one L. cervi individual featured the COI sequence of an unknown onchocercid nematode (Filarioidea), most similar (95.1%) to Mansonella perforata isolated from Sika deer (Cervus nippon) in Japan (AM749265).
Discussion
Here, we confirmed the molecular presence of various pathogens in blood-sucking hippoboscid flies infesting domestic and wild ruminants in Austria. The three louse fly species collected and investigated, L. cervi, M. ovinus and H. equina, have a widespread distribution in Europe [1, 50]. In the present study, L. cervi and M. ovinus were collected from their primary hosts, deer and sheep, respectively, whereas all H. equina were obtained from cattle, one of their facultative hosts [17]. The three investigated hippoboscid species differed in their total infection rates and infection prevalences to the different pathogen-groups, with M. ovinus specimens being significantly more infected than L. cervi and H. equina individuals to at least one pathogen (regardless of pathogen group), to Bartonella spp. and to trypanosomatids. Sheep keds also had a higher risk of being infected with two concurrent pathogen groups compared with L. cervi and H. equina. However, detailed molecular analyses revealed different pathogens (within each pathogen group) infecting each louse fly species; therefore, a comparison of the prevalences of the same pathogen between hippoboscids and their animal hosts is not possible. The different pathogens identified in the three hippoboscid species, and the probable role of these hippoboscids as vectors of the identified pathogens, are discussed below.
Bartonella spp. were the most frequently detected pathogens in H. equina and L. cervi, and the second most in M. ovinus. All Bartonella spp. in our study corresponded phylogenetically to species of the Bartonella linage II associated with strains that infect domestic and wild ruminants [51]. Bartonella spp. were first described in H. equina, L. cervi and M. ovinus almost 20 years ago [20, 21], with growing evidence pointing at the role of these hippoboscids as Bartonella vectors [28, 30, 52–57]. Importantly, we found ten distinct and previously unreported Bartonella spp. strains in L. cervi collected from red deer. Seven of these Bartonella spp. strains were highly similar (> 99%) to B. schoenbuchensis, a widespread pathogen infecting the midgut of deer keds [20, 28, 54]. Bartonella schoenbuchensis has been molecularly detected in blood and tissues samples from various wild ungulates, including red deer, roe deer and moose (Alces alces), all natural hosts for L. cervi [1, 28, 58–61]. Our results suggest that B. schoenbuchensis and related Bartonella spp. strains are common in L. cervi in Austria and may also be circulating in the local wild red deer populations. This is noteworthy in a One-Health context, considering that B. schoenbuchensis can be transmitted to humans, as described by a report of bacteremia in a patient suffering from fatigue, muscle pain and fever following a tick bite [62]. Moreover, B. schoenbuchensis has been suggested as the etiological agent of deer ked dermatitis in humans bitten by L. cervi [20], with similar clinical signs to cat scratch disease caused by the zoonotic Bartonella henselae [54, 63]. Therefore, the presence and distribution of B. schoenbuchensis in wild deer, deer keds and potentially other arthropod vectors in Austria warrant confirmation. Additionally, one Bartonella sp. strain isolated from deer keds in our study matched with a previously reported Bartonella sp. sequence detected in the common bent-wing bat M. schreibersii [64]. This Bartonella sp. and the B. schoenbuchensis-like strains identified in our study clustered together with B. schoenbuchensis and B. chomelii in the DNA haplotype network analysis. The other two Bartonella spp. strains detected in L. cervi clustered in a separate subclade and were highly similar to sequences of Candidatus B. melophagi reported from M. ovinus [65] and to Bartonella sp. isolated from Sika deer [66]. The diversity of Bartonella spp. lineages detected in deer keds in the present study and the presence of co-infections with two different Bartonella spp. lineages in several individuals indicate that L. cervi are reservoirs for a wide range of Bartonella spp. strains in Austria. Recent studies have also reported the recovery of several Bartonella spp. strains with zoonotic potential in deer keds (Lipoptena cervi and L. fortisetosa) and in cervids across Europe [27, 31, 33, 67], implying that these wild ungulates may act as reservoir hosts for these pathogens. Consequently, considering the common occurrence of wild cervids in Austria and the increasing reports of deer keds attacking humans in Europe [3, 6, 13, 68], it is imperative to further expand the monitoring and identification of zoonotic Bartonella spp. in deer keds and cervid populations.
Bartonella chomelii was the sole Bartonella species detected in H. equina collected from cattle in the present study. Bartonella chomelii was first described as a distinct Bartonella species from blood samples of cows in France [69], and subsequent reports in different countries confirmed its presence in both cattle [57, 70, 71] and H. equina [21, 56, 57]. Recently, molecular screenings also identified B. chomelii in ticks collected from rodents and dogs [72, 73]. In contrast, B. chomelii has not been detected in horses or other equids (the primary hosts of H. equina) or in H. equina parasitizing horses [21, 71]. It has been suggested that cattle could be accidental hosts for B. chomelii, which may be more closely related to Bartonella spp. from wild ruminants than strains isolated from domestic cattle such as B. bovis [69]. Considering that this is the first report of B. chomelii in Austria, further studies are needed to understand the occurrence and potential impact of this pathogen in cattle and wild ruminant populations. Previous work has suggested a higher prevalence of B. chomelii in older cattle (> 2 years old) and livestock managed in mountain pastures (> 600 m above sea level) [57, 71]. In the present study, B. chomelii-positive H. equina were collected from cattle grazing on mountain grasslands (~ 1000 to 1450 m above sea level; data not shown), located in the Hohe Tauern Alps of Salzburg [34], which suggests that animals during alpine grazing may be at risk of infections with B. chomelii, although this remains to be confirmed. To date, B. chomelii has not been demonstrated to induce disease in cattle, but infections with the related species B. bovis have been associated with bovine endocarditis [74].
Our results confirmed that the Bartonella spp. sequences detected in M. ovinus belonged to Candidatus B. melophagi, one of the most common pathogens in sheep ked populations [21, 53, 56]. Once thought to be only an endosymbiont of sheep keds not transmissible to ruminants [21], new evidence has confirmed the presence of Candidatus B. melophagi in sheep, including its successful culture from ovine blood, thus suggesting that sheep can serve as a host reservoir for this pathogen, with M. ovinus as its likely vector [55, 75]. However, it is still unclear whether Candidatus B. melophagi can cause clinical disease in sheep and whether M. ovinus is a competent vector transmitting these bacteria. Importantly, Candidatus B. melophagi was isolated from blood samples of two human patients presenting non-specific symptoms such as cardiovascular problems, pain and fatigue [76]. Despite these two patients having declared frequent contact with domestic and wild animals, there was no evidence of a possible route of infection or an actual causality between Candidatus B. melophagi infections and the clinical symptoms [76]. Therefore, the zoonotic potential of Candidatus B. melophagi and the role of M. ovinus as its vector require further elucidation.
In the present study, T. melophagium was the sole trypanosomatid species detected in M. ovinus, with high infection rate among the investigated sheep keds. Trypanosoma melophagium has been known to infect sheep keds and sheep for over a century [18, 77], and the present study adds to the few previous molecular genetic surveys confirming its high prevalence in M. ovinus collected from sheep in Scotland [22], Croatia [25] and recently Czechia [78]. Phylogenetic studies have concluded that T. melophagium is a single species restricted to M. ovinus and sheep; it is a member of the subgenus Megatrypanum, which includes other host-restricted pathogens infecting domestic and wild ruminants such as Trypanosoma theileri in cattle [25, 79]. Early works reported absent or very low parasitemia of T. melophagium in sheep infested by M. ovinus carrying this pathogen, and it has been suggested that sheep may become infected with T. melophagium merely by oral ingestion of sheep keds [18, 25]. To date, there is no evidence that T. melophagium can cause disease in sheep or be transmitted to other ruminant or mammalian species, and it has been proposed that it is non-pathogenic for infected M. ovinus [80]. Nevertheless, our findings suggest that sheep infested with M. ovinus in Austria could be infected with T. melophagium, and this should be confirmed, particularly in sheep farms with low or no use of ectoparasiticides for the control of sheep keds (e.g. organic farms), as previously described [25].
Our work identified a Trypanosoma sp. strain in three H. equina individuals collected from cattle in two different states of Austria (Salzburg and Burgenland). To the best of the authors’ knowledge, this is the first report of Trypanosoma sp. in H. equina, thus pointing at a potential role of this hippoboscid as a novel vector of animal trypanosomatids. In our phylogenetic analysis, this Trypanosoma sp. strain clustered together with sequences of the T. theileri group and was highly similar to Trypanosoma cf. cervi sequences isolated from white-tailed deer (Odocoileus virginianus) in the USA [81], with T. theileri-like strains from the horse flies Hybomitra tarandina, Chrysops divaricatus and Hybomitra muehlfeldi in Russia [82] and with T. theileri obtained from the tsetse fly Glossina fuscipes in the Central African Republic [83]. A previous study in Austria revealed a high prevalence of species belonging to the T. theileri/cervi complex in mosquitoes, suggesting a widespread distribution of these pathogens in animal hosts, potentially wild ruminants [84]. Therefore, the Trypanosoma sp. strain detected in H. equina in the present work might belong to the T. theileri group/complex, which may be elucidated by future molecular studies monitoring trypanosomatids in H. equina and cattle using various target genes. Furthermore, considering that T. theileri has not yet been reported to infect cattle in Austria but is present in neighboring countries [85, 86], the confirmation of T. theileri in vectors such as H. equina and tabanid flies, as well as in Austrian cattle, is warranted. Molecular genetic studies on trypanosomatids such as T. theileri-like strains have also been reported in the deer keds Lipoptena fortisetosa and L. cervi in Poland and Czechia [29, 78], but they were not detected in the current study. Regarding trypanosomatids detected in the deer ked L. cervi, we identified sequences highly similar to Bodo sp., which are non-parasitic kinetoplastids (suborder Bodonina) present in soil and water. Although Bodo spp. were isolated from bat ectoparasites and from the woylie Bettongia penicillate, an Australian marsupial, these findings were associated with environmental contamination of the mammalian hosts rather than infection [87, 88]. Therefore, we cannot exclude environmental contamination in our samples and further studies are needed to evaluate the role of Bodo sp. in hippoboscids.
In the present study, Anaplasmataceae were only detected in sheep keds and identified as Wolbachia spp. strains, which are known endosymbionts of nematodes and arthropods, including hippoboscids such as H. equina and M. ovinus [56, 65, 89, 90]. The 16S sequences of Wolbachia were identical to those of strains previously isolated from M. ovinus [91]. To date, the specific role or effects of Wolbachia on M. ovinus and other hippoboscid flies are unknown [90]. No pathogenic Anaplasmataceae were detected in the investigated hippoboscids, although M. ovinus and L. cervi can be infected with Anaplasma ovis [23, 92] and A. phagocytophilum [24], respectively. Considering that the zoonotic A. phagocytophilum has been confirmed to infect wild cervids and bovids in Austria [33, 93, 94], further work is needed to expand the monitoring of pathogenic Anaplasmataceae in hippoboscid flies and their ruminant hosts, whereas the single Borrelia-positive hippoboscid detected was a M. ovinus individual infected with a previously unreported strain 93.7% similar to a Borrelia isolate denominated A-FGy-1 and to Candidatus Borrelia mahuryensis isolated from neotropical passerine-associated ticks [95]. An early work confirmed the presence of Borrelia burgdorferi sensu lato, one of the causative agents of Lyme disease in humans, in M. ovinus [96]. The Borrelia sp. 16S sequence obtained in our study was 92.9% similar to two B. burgdorferi sequences uploaded to GenBank (AJ224138 and AJ224134). Considering the potential zoonotic risk of B. burgdorferi, new studies should characterize the identity and distribution of Borrelia spp. in sheep keds in Austria. Finally, only one L. cervi individual was positive for Filarioidea, featuring the COI sequence of an unknown onchocercid with a genetic similarity of 95% to the dermal filaroid Mansonella perforata, previously isolated from Sika deer [97, 98]. Whether this onchocercid sequence belongs to M. perforata or to another, not previously sequenced Mansonella species needs to be confirmed, as well as their potential distribution in red deer and deer ked populations in Austria.
Since the hippoboscid flies collected in the present study were isolated following a convenience sampling in selected farms with expected high infestation prevalence (sheep/cattle) and hunted animals as part of a tuberculosis surveillance program (deer), our results do not allow an accurate estimation of the country-wide prevalence of hippoboscid-associated pathogens in Austria. However, our data describe the widespread presence of pathogens in louse flies infesting ruminants in various geographic regions. Therefore, our findings support the need for the monitoring of hippoboscids and hippoboscid-borne diseases infecting domestic and wild ruminants in Austria, and potentially in other European regions, on a wider scale. Other pathogens previously reported in hippoboscid flies but not surveyed in the present study could also be included in future molecular surveys such as Rickettsia spp., Theileria ovis, Acinetobacter spp., Bacillus spp., Staphylococcus spp., Blue-tongue virus, Border Disease virus and Corynebacterium pseudotuberculosis [17]. Furthermore, the impact of climate change on the distribution and seasonal dynamics of louse flies should be investigated. Clearly, monitoring of potential zoonotic pathogens in (wild) animal reservoirs needs to be expanded in Austria, as indicated by the recent confirmation of Anaplasma phagocytophilum and Babesia spp. in wild ungulates [33].
This work contributes to the ongoing research efforts towards clarifying the role of blood-sucking hippoboscid flies as vectors of infectious agents of veterinary and medical importance. However, the presence of hippoboscid-associated pathogens confirmed by PCR and sequencing does not prove the vector competence of the investigated louse fly species for these pathogens, and only suggests their vector potential. Vector competence is the ability of arthropods to acquire and transmit the infective stage of a pathogen to a vertebrate host, including the pathogen’s replication within the vector. It requires conclusive experimental, epidemiological and clinical evidence of the pathogen’s transmission from the vector to the host, and the subsequent infection [99, 100]. For hippoboscids, there is evidence that Bartonella spp. can be vertically transmitted in L. cervi, with the pathogen being detected in adult deer keds blood feeding on ruminants, in pupae and in unfed adult flies that had not yet started to feed on blood [54]. However, whether L. cervi can transmit Bartonella spp. to a vertebrate host remains to be confirmed. Moreover, it should still be clarified whether the pathogens detected in L. cervi (e.g. Bartonella spp.) have zoonotic potential, how widespread these infectious agents are in wild ruminant populations in Austria and whether these animals may act as reservoirs for pathogens that could be vectored by deer keds. This is critically important considering the potential exposure of humans to bites by Bartonella-infected L. cervi during working or leisure activities (e.g. hunters, forestry workers, hikers).
Conclusions
In conclusion, molecular genetic screening confirmed the presence of several pathogens in three hippoboscid species infesting domestic and wild animals in Austria, with some potentially representing emerging zoonotic risks such as Bartonella spp. We report several novel pathogen sequences in hippoboscids that can contribute to the ongoing research efforts to understand the vector role of louse flies, including the first detection of Trypanosoma spp. in H. equina. Expanded monitoring of hippoboscids and hippoboscid-borne pathogens is warranted to clarify the distribution and impact of these ectoparasites as vectors of emerging infectious agents of public and animal health importance in a One-Health context.
Supplementary Information
Additional file 1: Figure S1. Bayesian inference tree featuring gltA sequencesof selected Bartonella spp. Nodes are marked with Bayesian inference posterior probabilities and maximum likelihood bootstrap values. Clades which are marked with a red bar were used for calculation of the median-joining haplotype networks containing the sequences obtained in this study. Scale bar indicates the expected mean number of substitutions per site according to the model of sequence evolution applied. Figure S2. Bayesian inference tree featuring 18S rRNA sequencesof selected Trypanosoma spp. Nodes are marked with Bayesian inference posterior probabilities and maximum likelihood bootstrap values. Clades which are marked with a red bar were used for calculation of the median-joining haplotype networks containing the sequences obtained in this study. Scale bar indicates the expected mean number of substitutions per site according to the model of sequence evolution applied.
Additional file 2: Table S1. Blast analysis of Bartonella spp. sequencesobtained from kedscollected from domestic and wild ruminants in Austria.
Acknowledgements
Financial support was provided by the Austrian Federal Ministry of Education, Science and Research (Austrian Barcode of Life—Hochschulraum-Strukturmittel).
Author contributions
HPF designed the research and raised the funding. DE, BSB, LvdV and MPE performed molecular genetic lab work. JH designed the Trypanosomatida primers. DB, GD, WG and SR were responsible for the field sampling of hippoboscid flies. JH and MSU carried out the phylogenetic analyses. MPE wrote the first draft. All authors have read and approved the final manuscript.
Funding
Open Access funding for this article was provided by the University of Veterinary Medicine Vienna (Vetmeduni Vienna). We acknowledge the financial support from the Austrian Federal Ministry of Education, Science and Research (Austrian Barcode of Life—Hochschulraum-Strukturmittel).
Availability of data and materials
All data generated or analysed during this study are included in this published article [and its supplementary information files].
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Reeves WK, Lloyd JE. Chapter20: louse flies, keds, and bat flies (Hippoboscoidea) In: Mullen GR, Durden LA, editors. Medical and veterinary entomology. Amsterdam: Elsevier Inc; 2019. pp. 421–38. [Google Scholar]
- 2.Smart J. Ked-flies. Nature. 1945;155:123. doi: 10.1038/155123a0. [DOI] [Google Scholar]
- 3.Rantanen T, Reunala T, Vuojolahti P, Hackman W. Persistent pruritic papules from deer ked bites. Acta Derm Venereol. 1982;62:307–311. [PubMed] [Google Scholar]
- 4.Small RW. A review of Melophagus ovinus (L.), the sheep ked. Vet Parasitol. 2005;130:141–55. doi: 10.1016/j.vetpar.2005.03.005. [DOI] [PubMed] [Google Scholar]
- 5.Petersen FT, Meier R, Kutty SN, Wiegmann BM. The phylogeny and evolution of host choice in the Hippoboscoidea (Diptera) as reconstructed using four molecular markers. Mol Phylogenet Evol. 2007;45:111–122. doi: 10.1016/j.ympev.2007.04.023. [DOI] [PubMed] [Google Scholar]
- 6.Mysterud A, Madslien K, Herland A, Viljugrein H, Ytrehus B. Phenology of deer ked (Lipoptena cervi) host-seeking flight activity and its relationship with prevailing autumn weather. Parasit Vectors. 2016;9:95. doi: 10.1186/s13071-016-1387-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lehikoinen A, Pohjola P, Valkama J, Mutanen M, Jaakko L. Promiscuous specialists: Host specificity patterns among generalist louse flies. PLoS ONE. 2021;16:e0247698. doi: 10.1371/journal.pone.0247698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Soliman SM, Attia MM, Al-Harbi MS, Saad AM, El-Saadony MT, Salem HM. Low host specificity of Hippobosca equina infestation in different domestic animals and pigeon. Saudi J Biol Sci. 2022;29:2112–2120. doi: 10.1016/j.sjbs.2021.11.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Taylor MA, Coop RL, Wall RL. Chapter 3—Veterinary entomology. In: Taylor MA, Coop RL, Wall RL, editors. Veterinary parasitology. 4. Wiley-Blackwell: Hoboken; 2016. [Google Scholar]
- 10.Oboňa J, Sychra O, Greš S, Heřman P, Manko P, Roháček J, et al. A revised annotated checklist of louse flies (Diptera, Hippoboscidae) from Slovakia. Zookeys. 2019;862:129–152. doi: 10.3897/zookeys.862.25992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Galecki R, Jaroszewski J, Bakula T, Xuan X. Molecular characterization of Lipoptena cervi from environmental samples collected in Poland. Int J Parasitol Parasites Wildl. 2021;14:41–47. doi: 10.1016/j.ijppaw.2020.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hermosilla C, Pantchev N, Bachmann R, Bauer C. Lipoptena cervi (deer ked) in two naturally infected dogs. Vet Rec. 2006;159:286–288. doi: 10.1136/vr.159.9.286. [DOI] [PubMed] [Google Scholar]
- 13.Härkönen S, Laine M, Vornanen M, Reunala T. Deer ked (Lipoptena cervi) dermatitis in humans—an increasing nuisance in Finland. Alces. 2009;45:73–79. [Google Scholar]
- 14.Decastello A, Farkas R. Anaphylactic reaction following forest fly (Hippobosca equina) bite: a human case. Clin Exp Med J. 2010;4:193–198. doi: 10.1556/CEMED.4.2010.1.19. [DOI] [Google Scholar]
- 15.Maslanko W, Bartosik K, Raszewska-Famielec M, Szwaj E, Asman M. Exposure of humans to attacks by deer keds and consequences of their bites—a case report with environmental background. Insects. 2020;11:859. doi: 10.3390/insects11120859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Gothe R, Schöl H. Deer keds (Lipoptena cervi) in the accompanying equipment of the Late Neolithic human mummy from the Similaun, South Tyrol. Parasitol Res. 1994;80:81–83. doi: 10.1007/BF00932630. [DOI] [PubMed] [Google Scholar]
- 17.Bezerra-Santos MA, Otranto D. Keds, the enigmatic flies and their role as vectors of pathogens. Acta Trop. 2020;209:105521. doi: 10.1016/j.actatropica.2020.105521. [DOI] [PubMed] [Google Scholar]
- 18.Hoare CA. An experimental study of the sheep-trypanosome (T. melophagium Flu, 1908), and its transmission by the sheep-ked (Melophagus ovinus L.) Parasitology. 1923;15:365–424. doi: 10.1017/S0031182000014888. [DOI] [Google Scholar]
- 19.Baker JR. A review of the role played by the Hippoboscidae (Diptera) as vectors of endoparasites. J Parasitol. 1967;53:412–418. doi: 10.2307/3276603. [DOI] [PubMed] [Google Scholar]
- 20.Dehio C, Sauder U, Hiestand R. Isolation of Bartonella schoenbuchensis from Lipoptena cervi, a blood-sucking arthropod causing deer ked dermatitis. J Clin Microbiol. 2004;42:5320–5323. doi: 10.1128/JCM.42.11.5320-5323.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Halos L, Jamal T, Maillard R, Girard B, Guillot J, Chomel B, et al. Role of hippoboscidae flies as potential vectors of Bartonella spp. infecting wild and domestic ruminants. Appl Environ Microbiol. 2004;70:6302–5. doi: 10.1128/AEM.70.10.6302-6305.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Gibson W, Pilkington JG, Pemberton JM. Trypanosoma melophagium from the sheep ked Melophagus ovinus on the island of St Kilda. Parasitology. 2010;137:1799–1804. doi: 10.1017/S0031182010000752. [DOI] [PubMed] [Google Scholar]
- 23.Hornok S, de la Fuente J, Biró N, de Fernández Mera I, Meli ML, Elek V, et al. First molecular evidence of Anaplasma ovis and Rickettsia spp. in keds (Diptera: Hippoboscidae) of sheep and wild ruminants. Vector-Borne Zoonotic Dis. 2011;11:1319–21. doi: 10.1089/vbz.2011.0649. [DOI] [PubMed] [Google Scholar]
- 24.Víchová B, Majláthová V, Nováková M, Majláth I, Čurlík J, Bona M, et al. PCR detection of re-emerging tick-borne pathogen, Anaplasma phagocytophilum, in deer ked (Lipoptena cervi) a blood-sucking ectoparasite of cervids. Biologia (Bratisl) 2011;66:1082–1086. doi: 10.2478/s11756-011-0123-1. [DOI] [Google Scholar]
- 25.Martinković F, Matanović K, Rodrigues AC, Garcia HA, Teixeira MMG. Trypanosoma (Megatrypanum) melophagium in the sheep ked Melophagus ovinus from organic farms in Croatia: phylogenetic inferences support restriction to sheep and sheep keds and close relationship with trypanosomes from other ruminant species. J Eukaryot Microbiol. 2012;59:134–144. doi: 10.1111/j.1550-7408.2011.00599.x. [DOI] [PubMed] [Google Scholar]
- 26.Duodu S, Madslien K, Hjelm E, Molin Y, Paziewska-Harris A, Harris PD, et al. Bartonella infections in deer keds (Lipoptena cervi) and moose (Alces alces) in Norway. Appl Environ Microbiol. 2013;79:322–327. doi: 10.1128/AEM.02632-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Razanske I, Rosef O, Radzijevskaja J, Klepeckiene K, Lipatova I, Paulauskas A. Infections with Bartonella spp. in free-ranging cervids and deer keds (Lipoptena cervi) in Norway. Comp Immunol Microbiol Infect Dis. 2018;58:26–30. doi: 10.1016/j.cimid.2018.06.003. [DOI] [PubMed] [Google Scholar]
- 28.Regier Y, Komma K, Weigel M, Pulliainen AT. Microbiome analysis reveals the presence of Bartonella spp. and Acinetobacter spp. in deer keds (Lipoptena cervi) Front Microbiol. 2018;9:3100. doi: 10.3389/fmicb.2018.03100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Werszko J, Steiner-Bogdaszewska Ż, Jezewski W, Szewczyk T, Kurylo G, Wolkowycki M, et al. Molecular detection of Trypanosoma spp. in Lipoptena cervi and Lipoptena fortisetosa (Diptera: Hippoboscidae) and their potential role in the transmission of pathogens. Parasitology. 2020;147:1629–35. doi: 10.1017/S0031182020001584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Werszko J, Asman M, Witecka J, Bogdaszewska ŻS. The role of sheep ked (Melophagus ovinus) as potential vector of protozoa and bacterial pathogens. Sci Rep. 2021;11:15468. doi: 10.1038/s41598-021-94895-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Galecki R, Jaroszewski J, Bakula T, Galon EM, Xuan X. Molecular detection of selected pathogens with zoonotic potential in deer keds (Lipoptena fortisetosa) Pathogens. 2021;10:324. doi: 10.3390/pathogens10030324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Cézanne R, Mrowietz N, Eigner B, Duscher GG, Glawischnig W, Fuehrer HP. Molecular analysis of Anaplasma phagocytophilum and Babesia divergens in red deer (Cervus elaphus) in Western Austria. Mol Cell Probes. 2017;31:55–58. doi: 10.1016/j.mcp.2016.07.003. [DOI] [PubMed] [Google Scholar]
- 33.Kogler S, Gotthalmseder E, Shahi-Barogh B, Harl J, Fuehrer HP. Babesia spp. and Anaplasma phagocytophilum in free-ranging wild ungulates in central Austria. Ticks Tick Borne Dis. 2021;12:101719. doi: 10.1016/j.ttbdis.2021.101719. [DOI] [PubMed] [Google Scholar]
- 34.Rehbein S, Kühnert E, Mayr S, Visser M. Louse fly (Hippobosca equina) infestation of cattle grazing at Alpine pastures in Salzburg, Austria. In: Parasitic diseases - a challenge for science and pracitce. DVG-Fachgruppe Parasitologie und parasitäre Krankheiten. Leipzig. Germany. 2019;129–30.
- 35.Hutson AM. Keds, flat-flies and bat-flies. Diptera, Hippoboscidae and Nycteribiidae. In: Handbooks for the Identification of British Insects, Part 7, 40, The Royal Entomological Society of London. 1984;10:1-40
- 36.Büttiker W. Die Lausfliegen der Schweiz (Diptera, Hippoboscidae). Documenta Faunistica Helvetiae 15. Schweizerisches Zentrum für die kartographische Erfassung der Fauna. Neuchâtel, Switzerland: Schweizerisches Zentrum für die karographische Erfassung der Fauna (SZKF); 1994.
- 37.Hebert PDN, Penton EH, Burns JM, Janzen DH, Hallwachs W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. PNAS. 2004;101:14812–14817. doi: 10.1073/pnas.0406166101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Norman AF, Regnery R, Jameson P, Greene C, Krause DC. Differentiation of Bartonella-like isolates at the species level by PCR- restriction fragment length polymorphism in the citrate synthase gene. J Clin Microbiol. 1995;33:1797–1803. doi: 10.1128/jcm.33.7.1797-1803.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Liebisch G, Sohns B, Bautsch W. Detection and typing of Borrelia burgdorferi sensu lato in Ixodes ricinus ticks attached to human skin by PCR. J Clin Microbiol. 1998;36:3355–3358. doi: 10.1128/JCM.36.11.3355-3358.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Parola P, Roux V, Camicas J-L, Baradji I, Brouqui P, Raoult D. Detection of ehrlichiae in African by polymerase chain reaction. Trans R Soc Trop Med Hyg. 2000;94:707–708. doi: 10.1016/S0035-9203(00)90243-8. [DOI] [PubMed] [Google Scholar]
- 41.Zintl A, Finnerty EJ, Murphy TM, de Waal T, Gray JS. Babesias of red deer (Cervus elaphus) in Ireland. Vet Res. 2011;42:7. doi: 10.1186/1297-9716-42-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hamer GL, Anderson TK, Berry GE, Makohon-moore AP, Crafton JC, Brawn JD, et al. Prevalence of filarioid nematodes and trypanosomes in American robins and house sparrows. Int J Parasitol Parasit Wildl. 2013;2:42–49. doi: 10.1016/j.ijppaw.2012.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Hall TA. BIOEDIT: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/ NT. Nucleic Acids Symp Ser. 1999;41:95–98. [Google Scholar]
- 44.Unterköfer MS, Harl J, Shahi B, Spergser J, Hrazdilová K, Müller F, et al. Molecular analysis of blood-associated pathogens in European wildcats (Felis silvestris silvestris) from Germany. Int J Parasitol Parasit Wildl. 2022;19:128–137. doi: 10.1016/j.ijppaw.2022.08.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Sánchez R, Serra F, Tárraga J, Medina I, Carbonell J, Pulido L, et al. Phylemon 2.0: a suite of web-tools for molecular evolution, phylogenetics, phylogenomics and hypotheses testing. Nucleic Acids Res. 2011;39:W470–4. doi: 10.1093/nar/gkr408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Xia X, Xie Z. DAMBE: software package for data analysis in molecular biology and evolution. J Hered. 2001;92:371–373. doi: 10.1093/jhered/92.4.371. [DOI] [PubMed] [Google Scholar]
- 47.Trifinopoulos J, Nguyen L-T, von Haeseler A, Minh BQ. W-IQ-TREE: a fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016;44:W232–W235. doi: 10.1093/nar/gkw256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ronquist F, Teslenko M, Van Der Mark P, Ayres DL, Darling A, Höhna S, et al. MrBayes 3.2: efficient bayesian phylogenetic inference and model choice across a large model space. Syst Biol. 2012;61:539–42. doi: 10.1093/sysbio/sys029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.R Core Team . R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing; 2022. [Google Scholar]
- 50.Aspöck H, Auer H, Walochnik J. Parasiten und Parasitäre Erkrankungen des Menschen in Mitteleuropa. Denisia. 2002;6:33–74. [Google Scholar]
- 51.Engel P, Salzburger W, Liesch M, Chang CC, Maruyama S, Lanz C, et al. Parallel evolution of a type IV secretion system in radiating lineages of the host-restricted bacterial pathogen Bartonella. PLoS Genet. 2011;7:e1001296. doi: 10.1371/journal.pgen.1001296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Reeves WK, Nelder MP, Cobb KD, Dasch GA. Bartonella spp. in deer keds, Lipoptena mazamae (Diptera: Hippoboscidae), from Georgia and South Carolina, USA. J Wildl Dis. 2006;42:391–6. doi: 10.7589/0090-3558-42.2.391. [DOI] [PubMed] [Google Scholar]
- 53.Kumsa B, Parola P, Raoult D, Socolovschi C. Bartonella melophagi in Melophagus ovinus (sheep ked) collected from sheep in northern Oromia, Ethiopia. Comp Immunol Microbiol Infect Dis. 2014;37:69–76. doi: 10.1016/j.cimid.2013.11.001. [DOI] [PubMed] [Google Scholar]
- 54.Korhonen EM, Perez Vera C, Pulliainen AT, Sironen T, Aaltonen K, Kortet R, et al. Molecular detection of Bartonella spp. in deer ked pupae, adult keds and moose blood in Finland. Epidemiol Infect. 2015;143:578–85. doi: 10.1017/S0950268814001411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Kosoy M, Bai Y, Enscore R, Rizzo MR, Bender S, Popov V, et al. Bartonella melophagi in blood of domestic sheep (Ovis aries) and sheep keds (Melophagus ovinus) from the southwestern US: cultures, genetic characterization, and ecological connections. Vet Microbiol. 2016;190:43–49. doi: 10.1016/j.vetmic.2016.05.009. [DOI] [PubMed] [Google Scholar]
- 56.Boucheikhchoukh M, Mechouk N, Benakhla A, Raoult D. Molecular evidence of bacteria in Melophagus ovinus sheep keds and Hippobosca equina forest flies collected from sheep and horses in northeastern Algeria. Comp Immunol Microbiol Infect Dis. 2019;65:103–109. doi: 10.1016/j.cimid.2019.05.010. [DOI] [PubMed] [Google Scholar]
- 57.Boularias G, Azzag N, Gandoin C, Bouillin C, Chomel B, Haddad N, et al. Bartonella bovis and Bartonella chomelii infection in dairy cattle and their ectoparasites in Algeria. Comp Immunol Microbiol Infect Dis. 2020;70:101450. doi: 10.1016/j.cimid.2020.101450. [DOI] [PubMed] [Google Scholar]
- 58.Dehio C, Lanz C, Pohl R, Behrens P, Bermond D, Piémont Y, et al. Bartonella schoenbuchii sp. nov., isolated from the blood of wild roe deer. Int J Syst Evol Microbiol. 2001;51:1557–65. doi: 10.1099/00207713-51-4-1557. [DOI] [PubMed] [Google Scholar]
- 59.Adamska M. Wild ruminants in the area of the North-Western Poland as potential reservoir hosts of Bartonella schoenbuchensis and B. bovis. Acta Parasitol. 2008;53:407–10. doi: 10.2478/s11686-008-0058-z. [DOI] [Google Scholar]
- 60.Guy L, Nystedt B, Toft C, Zaremba-Niedzwiedzka K, Berglund EC, Granberg F, et al. A gene transfer agent and a dynamic repertoire of secretion systems hold the keys to the explosive radiation of the emerging pathogen Bartonella. PLoS Genet. 2013;9:e1003393. doi: 10.1371/journal.pgen.1003393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Welc-Falȩciak R, Werszko J, Cydzik K, Bajer A, Michalik J, Behnke JM. Co-infection and genetic diversity of tick-borne pathogens in roe deer from Poland. Vector-Borne Zoonotic Dis. 2013;13:277–288. doi: 10.1089/vbz.2012.1136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Vayssier-Taussat M, Moutailler S, Féménia F, Raymond P, Croce O, La Scola B, et al. Identification of novel zoonotic activity of Bartonella spp., France. Emerg Infect Dis. 2016;22:457–62. doi: 10.3201/eid2203.150269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Guptill L. Bartonellosis. Vet Microbiol. 2010;140:347–359. doi: 10.1016/j.vetmic.2009.11.011. [DOI] [PubMed] [Google Scholar]
- 64.McKee CD, Krawczyk AI, Sándor AD, Görföl T, Földvári M, Földvári G, et al. Host phylogeny, geographic overlap, and roost sharing shape parasite communities in European bats. Front Ecol Evol. 2019;7:1–21. doi: 10.3389/fevo.2019.00069. [DOI] [Google Scholar]
- 65.Liu Y, He B, Li F, Li K, Zhang L, Li X, et al. Molecular identification of Bartonella melophagi and Wolbachia supergroup F from sheep keds in Xinjiang, China. Korean J Parasitol. 2018;56:365–370. doi: 10.3347/kjp.2018.56.4.365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Harms A, Segers FHID, Quebatte M, Mistl C, Manfredi P, Körner J, et al. Evolutionary dynamics of pathoadaptation revealed by three independent acquisitions of the VirB/D4 type IV secretion system in Bartonella. Genome Biol Evol. 2017;9:761–776. doi: 10.1093/gbe/evx042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Sacristán C, Carlos G, Faisal N, Irene S, Tengs T, Hamnes IS, et al. Bartonella spp detection in ticks, Culicoides biting midges and wild cervids from Norway. Transbound Emerg Dis. 2021;68:941–51. doi: 10.1111/tbed.13762. [DOI] [PubMed] [Google Scholar]
- 68.Buczek W, Buczek AM, Bartosik K, Buczek A. Comparison of skin lesions caused by Ixodes ricinus ticks and Lipoptena cervi deer keds infesting humans in the natural environment. Int J Environ Res Public Health. 2020;17:1–8. doi: 10.3390/ijerph17093316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Maillard R, Riegel P, Barrat F, Boullin C, Thibault D, Gandoin C, et al. Bartonella chomelii sp. nov., isolated from French domestic cattle (Bos taurus) Int J Syst Evol Microbiol. 2004;54:215–20. doi: 10.1099/ijs.0.02770-0. [DOI] [PubMed] [Google Scholar]
- 70.Mediannikov O, Davoust B, Cabre O, Rolain JM, Raoult D. Bartonellae in animals and vectors in New Caledonia. Comp Immunol Microbiol Infect Dis. 2011;34:497–501. doi: 10.1016/j.cimid.2011.09.002. [DOI] [PubMed] [Google Scholar]
- 71.Antequera-Gómez ML, Lozano-Almendral L, Barandika JF, González-Martín-Niño RM, Rodríguez-Moreno I, García-Pérez AL, et al. Bartonella chomelii is the most frequent species infecting cattle grazing in communal mountain pastures in Spain. Appl Environ Microbiol. 2015;81:623–629. doi: 10.1128/AEM.03159-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Silaghi C, Pfeffer M, Kiefer D, Kiefer M, Obiegala A. Bartonella, rodents, fleas and ticks: a molecular field study on host-vector-pathogen associations in Saxony, Eastern Germany. Microb Ecol. 2016;72:965–974. doi: 10.1007/s00248-016-0787-8. [DOI] [PubMed] [Google Scholar]
- 73.Ereqat S, Nasereddin A, Vayssier-Taussat M, Abdelkader A, Al-Jawabreh A, Zaid T, et al. Molecular evidence of Bartonella species in ixodid ticks and domestic animals in Palestine. Front Microbiol. 2016;7:1217. doi: 10.3389/fmicb.2016.01217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Maillard R, Petit E, Chomel B, Lacroux C, Schelcher F, Vayssier-Taussat M, et al. Endocarditis in cattle caused by Bartonella bovis. Emerg Infect Dis. 2007;13:1383–1385. doi: 10.3201/eid1309.070236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Bemis DA, Kania SA. Isolation of Bartonella sp. from sheep blood. Emerg. Infect. Dis. 2007;13:1565–7. [DOI] [PMC free article] [PubMed]
- 76.Maggi RG, Kosoy M, Mintzer M, Breitschwerdt EB. Isolation of candidatus Bartonella melophagi from human blood. Emerg Infect Dis. 2009;15:66–68. doi: 10.3201/eid1501.081080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Magri A, Galuppi R, Fioravanti M. Autochthonous Trypanosoma spp. in European mammals: a brief journey amongst the neglected trypanosomes. Pathogens. 2021;10:334. doi: 10.3390/pathogens10030334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Brotánková A, Fialová M, Čepička I, Brzoňová J, Svobodová M. Trypanosomes of the Trypanosoma theileri group: phylogeny and new potential vectors. Microorganisms. 2022;10:294. doi: 10.3390/microorganisms10020294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Votýpka J, D’Avila-Levy CM, Grellier P, Maslov DA, Lukeš J, Yurchenko V. New approaches to systematics of Trypanosomatidae: criteria for taxonomic (re)description. Trends Parasitol. 2015;31:460–469. doi: 10.1016/j.pt.2015.06.015. [DOI] [PubMed] [Google Scholar]
- 80.Nelson WA. Melophagus ovinus (Pupipara: Hippoboscidae): confirmation of the nonpathogenicity of Trypanosoma melophagium for sheep keds. J Invertebr Pathol. 1981;37:284–289. doi: 10.1016/0022-2011(81)90088-4. [DOI] [PubMed] [Google Scholar]
- 81.Fisher AC, Schuster G, Cobb WJ, James AM, Cooper SM, Peréz de León AA, et al. Molecular characterization of Trypanosoma (Megatrypanum) spp. infecting cattle (Bos taurus), white-tailed deer (Odocoileus virginianus), and elk (Cervus elaphus canadensis) in the United States. Vet Parasitol. 2013;197:29–42. doi: 10.1016/j.vetpar.2013.04.037. [DOI] [PubMed] [Google Scholar]
- 82.Ganyukova AI, Zolotarev AV, Malysheva MN, Frolov AO. First record of Trypanosoma theileri-like flagellates in horseflies from Northwest Russia. Protistology. 2018;12:223–230. doi: 10.21685/1680-0826-2018-12-4-6. [DOI] [Google Scholar]
- 83.Votýpka J, Rádrová J, Skalický T, Jirků M, Jirsová D, Mihalca AD, et al. A tsetse and tabanid fly survey of African great apes habitats reveals the presence of a novel trypanosome lineage but the absence of Trypanosoma brucei. Int J Parasitol. 2015;45:741–748. doi: 10.1016/j.ijpara.2015.06.005. [DOI] [PubMed] [Google Scholar]
- 84.Schoener E, Uebleis SS, Cuk C, Nawratil M, Obwaller AG, Zechmeister T, et al. Trypanosomatid parasites in Austrian mosquitoes. PLoS ONE. 2018;13:e0196052. doi: 10.1371/journal.pone.0196052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Amato B, Mira F, Di Presti Marco Lo V, Guercio A, Russotto L, Gucciardi F, et al. A case of bovine trypanosomiasis caused by Trypanosoma theileri in Sicily, Italy. Parasitol Res. 2019;118:2723–7. doi: 10.1007/s00436-019-06390-y. [DOI] [PubMed] [Google Scholar]
- 86.Bittner L, Krämer K, Wöckel A, Snedec T, Delling C, Böttcher D, et al. Malnutrition as the cause of recumbency in suckler cows associated with Trypanosoma theileri infection. Acta Vet Scand. 2021;63:2. doi: 10.1186/s13028-020-00567-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Szöke K, Sándor AD, Boldogh SA, Görföl T, Votýpka J, Takács N, et al. DNA of free-living bodonids (Euglenozoa: Kinetoplastea) in bat ectoparasites: Potential relevance to the evolution of parasitic trypanosomatids. Acta Vet Hung. 2017;65:531–540. doi: 10.1556/004.2017.051. [DOI] [PubMed] [Google Scholar]
- 88.Northover AS, Godfrey SS, Keatley S, Lymbery AJ, Wayne AF, Cooper C, et al. Increased Trypanosoma spp. richness and prevalence of haemoparasite co-infection following translocation. Parasit Vectors. 2019;12:126. doi: 10.1186/s13071-019-3370-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Nováková E, Husník F, Šochová E, Hypša V. Arsenophonus and Sodalis symbionts in louse flies: an analogy to the Wigglesworthia and Sodalis system in tsetse flies. Appl Environ Microbiol. 2015;81:6189–6199. doi: 10.1128/AEM.01487-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Madhav M, Baker D, Morgan JAT, Asgari S, James P. Wolbachia: a tool for livestock ectoparasite control. Vet Parasitol. 2020;288:109297. doi: 10.1016/j.vetpar.2020.109297. [DOI] [PubMed] [Google Scholar]
- 91.Šochová E, Husník F, Nováková E, Halajian A, Hypša V. Arsenophonus and Sodalis replacements shape evolution of symbiosis in louse flies. PeerJ. 2017;5:e4099. doi: 10.7717/peerj.4099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Zhao L, He B, Li KR, Li F, Zhang LY, Li XQ, et al. First report of Anaplasma ovis in pupal and adult Melophagus ovinus (sheep ked) collected in South Xinjiang. China Parasites & Vectors. 2018;11:258. doi: 10.1186/s13071-018-2788-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Silaghi C, Hamel D, Thiel C, Pfister K, Passos LMF, Rehbein S. Genetic variants of Anaplasma phagocytophilum in wild caprine and cervid ungulates from the Alps in Tyrol, Austria. Vector-Borne Zoonotic Dis. 2011;11:355–362. doi: 10.1089/vbz.2010.0051. [DOI] [PubMed] [Google Scholar]
- 94.Silaghi C, Hamel D, Pfister K, Rehbein S. Babesia species and co-infection with Anaplasma phagocytophilum in free-ranging ungulates from Tyrol (Austria) Wien Tierarztl Monatsschr. 2011;98:268–274. [Google Scholar]
- 95.Binetruy F, Garnier S, Boulanger N, Talagrand-Reboul É, Loire E, Faivre B, et al. A novel Borrelia species, intermediate between Lyme disease and relapsing fever groups, in neotropical passerine-associated ticks. Sci Rep. 2020;10:10596. doi: 10.1038/s41598-020-66828-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Chu CY, Jiang BG, Qiu EC, Zhang F, Zuo SQ, Yang H, et al. Borrelia burgdorferi sensu lato in sheep keds (Melophagus ovinus), Tibet, China. Vet Microbiol. 2011;149:526–529. doi: 10.1016/j.vetmic.2010.11.031. [DOI] [PubMed] [Google Scholar]
- 97.Uni S, Bain O, Takaoka H. Affinities between Cutifilaria (Nematoda: Filarioidea), parasites of deer, and Mansonella as seen in a new onchocercid, M. (C.) perforata n. sp, from Japan. Parasite. 2004;11:131–40. doi: 10.1051/parasite/2004112131. [DOI] [PubMed] [Google Scholar]
- 98.Ferri E, Barbuto M, Bain O, Galimberti A, Uni S, Guerrero R, et al. Integrated taxonomy: traditional approach and DNA barcoding for the identification of filarioid worms and related parasites (Nematoda) Front Zool. 2009;6:1. doi: 10.1186/1742-9994-6-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Pultorak EL, Maggi RG, Breitschwerdt EB. Bartonellosis: a one health perspective. In: Yamada A, Kahn LH, Kaplan B, Monath TP, Woodall J, Conti L, editors. Confronting emerging zoonoses: the one health paradigm. Springer Japan: Tokyo; 2014. pp. 113–49. [Google Scholar]
- 100.Wilson AJ, Morgan ER, Booth M, Norman R, Perkins SE, Hauffe HC, et al. What is a vector? Philos Trans R Soc B Biol Sci. 2017;372:20160085. doi: 10.1098/rstb.2016.0085. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1: Figure S1. Bayesian inference tree featuring gltA sequencesof selected Bartonella spp. Nodes are marked with Bayesian inference posterior probabilities and maximum likelihood bootstrap values. Clades which are marked with a red bar were used for calculation of the median-joining haplotype networks containing the sequences obtained in this study. Scale bar indicates the expected mean number of substitutions per site according to the model of sequence evolution applied. Figure S2. Bayesian inference tree featuring 18S rRNA sequencesof selected Trypanosoma spp. Nodes are marked with Bayesian inference posterior probabilities and maximum likelihood bootstrap values. Clades which are marked with a red bar were used for calculation of the median-joining haplotype networks containing the sequences obtained in this study. Scale bar indicates the expected mean number of substitutions per site according to the model of sequence evolution applied.
Additional file 2: Table S1. Blast analysis of Bartonella spp. sequencesobtained from kedscollected from domestic and wild ruminants in Austria.
Data Availability Statement
All data generated or analysed during this study are included in this published article [and its supplementary information files].