

Abstract
Estrogen has a well-known effect of reducing salt intake in rats. This mini review focuses on recent findings regarding the interaction of estradiol with brain angiotensin II to control increased sodium palatability that occurs as a result of sodium appetite in spontaneously hypertensive rats.
Keywords: estradiol, palatability, hypertension, sodium, angiotensin II
1 Introduction
The ingestion of water and sodium is a necessary behavioral response for terrestrial animals to compensate for body fluid deficits. Sodium appetite occurs in response to prolonged extracellular dehydration or sodium depletion and is an innate and behaviorally motivated state influenced by learning, environmental conditions, and emotions (Denton et al., 1982; Schulkin, 1991).
Hormones play a significant role in inducing or constraining sodium intake in humans and rats (Santollo et al., 2023). This role involves the organization of future behavior from gestation and birth onward, in addition to the immediate effect of hormones on the ingestion of salt (Epstein, 1991; Mecawi et al., 2015). Angiotensin II (ANG II), for example, acts in the lamina terminalis of the third brain ventricle to produce sodium appetite, in collaboration with steroids that control fluid balance, such as aldosterone (Epstein, 1991; De Luca et al., 2010). In contrast, other hormones, some peptides and sex steroids among them, counteract the effect of ANG II (Mecawi et al., 2015). In addition to the effect on sodium consumption, estrogens also have a profound influence on hydromineral balance and cardiovascular parameters, and exert a beneficial effect in terms of control of arterial blood pressure in rats and humans (Hay et al., 2014; Vivas et al., 2014).
The objective of this mini-review is to highlight the role of taste in the inhibitory effect that estrogens exert on hypertonic NaCl intake as an expression of sodium appetite in rats. Thus, the neurobiology of sodium appetite is briefly reviewed, followed by description of the inhibitory effect of estrogens on sodium appetite, sodium preference, and sodium palatability in normotensive and hypertensive rats. Subsequently, we discuss the inhibitory effect of estrogens on sodium palatability in a model of sodium appetite produced by water deprivation and partial rehydration (WD-PR) in spontaneously hypertensive rats (SHR).
2 Neurobiology of sodium appetite
The control of sodium and water intake depends on forebrain and hindbrain areas involved in a circuitry that integrates facilitatory and inhibitory mechanisms (Johnson and Thunhorst, 2007; Formenti et al., 2013; Menani et al., 2014). Facilitatory mechanisms that produce water and sodium intake include ANG II and aldosterone. The action of the facilitatory mechanisms is counterbalanced by inhibitory mechanisms derived from visceral or brain sensory information (Johnson and Thunhorst, 2007; Menani et al., 2014).
The renin–angiotensin–aldosterone system (RAAS) is a key hormonal system activated by hypovolemia-related conditions (Fitzsimons, 1998). Central ANG II receptors are expressed in neuronal cells in several brain areas, such as the subfornical organ (SFO), organum vasculosum of the lamina terminalis (OVLT), paraventricular nucleus (PVN), median preoptic nucleus (MnPO), area postrema (AP), and nucleus of the solitary tract (NTS) (Lewis et al., 1986; McKinley et al., 1987; Allen et al., 1988; Allen et al., 1999). Among ANG II receptors, the most important for its dipsogenic effect are the AT1 receptors present in forebrain circumventricular organs, such as the SFO and OVLT (Fitzsimons, 1998).
Changes in blood pressure and volume are detected by receptors located in the aortic arch and carotid sinus (baroreceptors) and in the walls of large veins and the atria (volume receptors). Afferent signals from baroreceptors and cardiopulmonary receptors reach the brainstem via cranial nerves IX and X, with most of these nerve endings making their first synapse in the NTS (Johnson and Thunhorst, 2007). The NTS sends projections to the lateral parabrachial nucleus (LPBN), a pontine structure located dorsal to the superior cerebellar peduncle.
The LPBN acts as an important inhibitory mechanism for sodium intake control (for reviews, see Andrade et al., 2014; Menani et al., 2014). The LPBN also has important reciprocal connections with several other hindbrain and forebrain areas involved in the control of fluid and electrolyte balance (Ciriello et al., 1984; Fulwiler and Saper, 1984; Jhamandas et al., 1996; Johnson and Thunhorst, 2007). Signals from peripheral baroreceptors, cardiopulmonary volume receptors, taste receptors, or osmoreceptors ascend to the LPBN, and from this nucleus, they may influence the activity of forebrain areas important for the control of sodium intake.
Sodium appetite can be experimentally induced by various protocols. A slow-onset protocol involves subcutaneous injection of the diuretic furosemide (10 mg/mL/rat), followed by access to only water and sodium-deficient food for 24 h. This method promotes a loss of between 1.5 and 2.0 mEq of sodium and induces a consistent sodium appetite, which expresses or “matures” several hours after the injection of furosemide (Sakai et al., 1986; Rowland and Morian, 1999). An acute, rapid-onset sodium appetite can be induced by furosemide combined with a low dose of the angiotensin-converting enzyme blocker captopril (FURO + CAP). The FURO + CAP protocol advances the expression of sodium appetite to within 2 h (Thunhorst and Johnson, 1994; Thunhorst et al., 1998). The rapid induction of water and 0.3 M NaCl intake by FURO + CAP treatment results from mild hypotension - which unloads low-pressure volume receptors, combined with the action of ANG II in the brain (Fitts and Masson, 1989; Thunhorst et al., 1994; Thunhorst and Johnson, 1994).
The water deprivation–partial rehydration (WD-PR) protocol provides a method for production of clearly distinct thirst and sodium appetite, in which the rat feeds on regular laboratory chow, a sodium-excess diet relative to the animal’s need (De Luca et al., 2010). The WD-PR protocol involves a sequence of water deprivation with free access to regular laboratory chow (WD), followed by partial rehydration (PR), during which period the rat is allowed to drink only water to satiation. Drinking water to satiation during PR leads to the dilution of body fluids and partial recovery of the extracellular volume with sustained high systemic activation of the RAS (De Luca et al., 2002; De Luca et al., 2010). Thereafter, a sodium appetite two-bottle test (0.3 M NaCl and water) is conducted. Increased NaCl intake relative to water intake results in recovery of blood volume (De Luca et al., 2010). It also induces differential neuronal activity in the lamina terminalis and other brain areas relative to the preceding hydrated, WD, and WD-PR conditions (Pereira-Derderian et al., 2010; Dalmasso et al., 2015).
3 Estrogens and sodium appetite
Estrogens have a dual effect on sodium appetite in rats. On the one hand, their perinatal action confers a “female” character on offspring, thus being responsible for female sodium preference in adulthood (Epstein, 1991). On the other hand, estrogens limit the hydromineral fluid intake of females during estrus (Antunes Rodrigues and Covian, 1963; Danielsen and Buggy, 1980). This effect is associated with modulation of the hypothalamic secretion of inhibitory peptides, e.g., oxytocin, and downregulation of the brain renin–angiotensin system (RAS) (Mecawi et al., 2015; Almeida-Pereira et al., 2019).
Estrogens also modulate the local RAS in different organs. For example, in the kidney, estradiol (E2) decreases the activity of angiotensin-converting enzyme (ACE) and renin, as well as the mRNA expression of angiotensin II type 1 receptor (AT1R), but increases the mRNA expression of AT2R and the production of ANG-(1-7) (Shenoy et al., 2009; Brown et al., 2012; Bhatia et al., 2013). In the brain, E2 inhibits the mRNA expression of AT1R and ACE in the lamina terminalis (Kisley et al., 1999; Dean et al., 2006; Krause et al., 2006; Santollo and Daniels, 2015).
4 Sodium palatability
Humans, other, primates, and rats exhibit stereotyped orofacial reactions to different tastes applied directly to the oral cavity, and the evaluation of these reactions can be used as a behavioral measure of the hedonic value of taste: that is, taste palatability (Grill and Norgren, 1978; Berridge, 2000; Berridge and Kringelbach, 2015). Using this approach, previous studies have shown that the palatability of salty taste increases after sodium depletion (Berridge et al., 1984; Grill and Bernstein, 1988; Berridge and Schulkin, 1989). In other words, sodium depletion promotes an increase in the frequency of hedonic reactions and a decrease in aversive reactions in response to intraoral (IO) administration of NaCl solution, thus characterizing a hedonic shift in sodium palatability induced by sodium depletion (Berridge et al., 1984; Grill and Bernstein, 1988; Berridge and Schulkin, 1989).
A recent study from our laboratory with normotensive rats showed that antagonism of brain AT1R reduces hedonic responses and increases aversive responses to intraoral infusion of 0.3 M NaCl (Zenatti et al., 2021). Moreover, in the same strain of rats, increased endogenous production of brain ANG II sensitizes hedonic responses to intraoral infusion of the same solution (Fernandes et al., 2022).
5 Salt preference
Several studies have demonstrated an increase in preference for sodium in humans undergoing alterations in fluid and electrolyte balance, such as administration of diuretics accompanied by a low-sodium diet and water deprivation, as well as sweating, hemodialysis, and gestation (Beauchamp et al., 1990; Takamata et al., 1994; Leshem and Rudoy, 1997; Duffy et al., 1998). However, excessive NaCl intake is a risk factor for chronic diseases, including hypertension. Limited sodium intake (less than 2 g/day) is recommended to control blood pressure and reduce the risk of cardiovascular diseases (He and Macgregor, 2012; World Health Organization, 2016); this recommendation is difficult to follow because dietary NaCl increases food palatability (Keast and Breslin, 2003; Liem et al., 2011). Moreover, increased salt preference may be associated with pathological conditions. This can be observed in medicated, compensated patients undergoing chronic treatment for heart failure (de Souza et al., 2012). The same is observed in patients with primary aldosteronism, who show reduced taste perception of NaCl (Adolf et al., 2021). This favors a higher dietary intake, which in turn accelerates the progress of arterial hypertension (Adolf et al., 2021).
Investigations of preference for salt in hypertension can benefit from the use of animal models of palatability.
6 Sodium intake and sodium palatability in spontaneously hypertensive rats
The spontaneously hypertensive rat (SHR) is a traditional model for the study of essential hypertension (Elmarakby and Sullivan, 2021). In addition to high blood pressure, the adult SHR exhibits enhanced sodium intake, as measured by ingestion of NaCl solution (for early reviews, see Ely et al., 2000; Di Nicolantonio et al., 2004). This intense NaCl intake arises from a hyperactive brain RAS rather than from altered fluid–electrolyte balance (DiNicolantonio et al., 1982; Di Nicolantonio et al., 2004). Brain injection of captopril, an ACE inhibitor, reduces daily NaCl intake in the SHR (DiNicolantonio et al., 1982). Accordingly, the SHR is suitable for modeling of excess sodium intake in the human context under different hydration conditions and difficulty in controlling arterial pressure (Takamata et al., 1994; Millard-Stafford et al., 2012; Lucia and Curtis, 2018; Mill et al., 2019; Minegishi et al., 2020; Orysiak et al., 2022; Zhang et al., 2022).
As predicted, the SHR exhibits higher hypertonic NaCl intake than normotensive strains in response to WD-PR (Pereira-Derderian et al., 2010). The SHR also exhibits a different pattern of encephalic c-Fos expression, particularly in the lamina terminalis, in response to WD-PR (Pereira-Derderian et al., 2010). In addition, the lamina terminalis of this strain expresses more Fos-IR in response to either brain or systemic infusions of ANG II (Rowland et al., 1995; Blume et al., 1997). Finally, inhibition of brain AT1R-mediated ERK1/2 activation reduces 0.3 M NaCl intake and the ANG II pressor response in the SHR (Andrade-Franze et al., 2020). Taken together, these findings suggest that the high intake of hypertonic NaCl exhibited by the SHR in response to WD-PR results from hyperactive brain RAS activity, particularly in the lamina terminalis.
By investigating sodium palatability in male SHRs and normotensive rats under euhydrated and dehydrated conditions, we have previously shown that SHRs also have enhanced palatability for NaCl taste, as measured by an increased number of hedonic versus aversive responses to IO infusion (1 mL/1 min) of 0.3 M NaCl in a taste reactivity test, irrespective of their body fluid status (Pereira et al., 2021).
7 Discussion
Considering that estrogens exert an inhibitory influence on sodium intake and that SHRs constitute a model for the study of increased sodium intake and palatability in the context of essential hypertension, we aimed to investigate sodium palatability in female SHRs in comparison to normotensive rats. In this section, we compare the findings of recent studies with female SHRs (Pereira et al., 2019; Pereira et al., 2022) to those of a study with male SHRs (Pereira et al., 2021), discussing the possible mechanisms involved in the effects of estrogen on sodium palatability in SHRs.
Using hormone replacement therapy (Sampson et al., 2012; Tazumi et al., 2016), we have investigated the role of E2 in salt taste in ovariectomized female SHRs “clamped” in simulated physiological estrus (Pereira et al., 2019). Under the WD-PR protocol, E2 exerted a strong effect (>50% alteration) on the occurrence of hedonic (reduced) and aversive (increased) orofacial reactions to intraoral infusion of 0.3 M NaCl in SHRs (Figure 1, left). There was also a reduction of approximately 75% in 0.3 M NaCl intake in a subsequent sodium appetite test (not shown). In contrast, ovariectomized (OVX) control SHRs that received no estogens exhibited orofacial reactions comparable to those of male SHRs (Figure 1, left). OVX SHRs also ingested the same amount of 0.3 M NaCl as male SHRs (not shown). Recall that male SHRs exhibit not only notably high 0.3 M NaCl intake but also strong orofacial hedonic reactions and a low rate of aversive reactions to IO of this solution, even in a euhydrated condition, relative to normotensive strains (Pereira-Derderian et al., 2010; Pereira et al., 2021). As shown in Figure 1 (left, bottom), E2 significantly increased the occurrence of aversive reactions in SHRs in a euhydrated condition.
FIGURE 1.

Adapted from our previous studies (Pereira et al., 2021; Pereira et al., 2022) with permission from Appetite and Hormones and Behavior. Left: the number of hedonic (top) and aversive (bottom) responses to IO infusion of 0.3 M NaCl in a euhydrated condition, after WD-PR and following the sodium appetite test in female and male SHRs submitted to this protocol. Right: the number of hedonic (top) and aversive (bottom) responses to IO infusion of 0.3 M NaCl after WD-PR in ovariectomized (OVX) SHRs treated with vehicle or estradiol (E2, 10 μg/kg/rat) s.c. combined with saline or losartan (100 μg/μL) ICV. Results are presented as means ± SEM. Differences were analyzed by two-way ANOVA and the Student–Newman–Keuls test.
In a follow-up investigation of responses to WD-PR (Pereira et al., 2022), intracerebroventricular (ICV) injection of losartan (an AT1R antagonist) was found to have no effect on orofacial reactions to IO of 0.3 M NaCl in E2-treated SHRs (Figure 1, right). In contrast, losartan strongly reduced hedonic reactions and increased aversive reactions in OVX SHRs. ICV losartan also abolished 0.3 M NaCl intake in a sodium appetite test among OVX and E2-treated SHRs (not shown). Finally, and similarly to the outcome in the brains of normotensive rats (Krause et al., 2006), E2-treated SHRs exhibited reduced hypothalamic mRNA expression of AT1R (Pereira Jr. et al., 2022).
The SHR/WD-PR model, along with data from a normotensive strain, provides fresh evidence for the roles of brain ANG II and estrogen in emotion-based control of behavior in relation to salty taste (Pereira et al., 2019; Pereira et al., 2021; Zenatti et al., 2021; Fernandes et al., 2022; Pereira Jr. et al., 2022). We conclude that alteration of salt palatability is a crucial key in understanding why estrogen inhibits NaCl intake in SHRs. The underlying mechanism resides in the brain, involving the attenuation of ANG II-induced increases in salt palatability. The extent to which this attenuation depends on the modulation of salt taste afferents (Shigemura et al., 2013; Martin and Sollars, 2017) is open to further research.
Orofacial reactions to taste correspond directly to the consummation of the motivation to ingest salt, a behavior modulated by the mesolimbic system (Berridge, 2018; Hsu et al., 2018). Estrogens modulate neuronal activity in the preoptic/hypothalamic area in connection with mesolimbic dopaminergic transmission (Champagne and Curley, 2016). As a final note, Figure 2 illustrates how estrogens interact with brain ANG II to inhibit salty taste, thereby inhibiting sodium appetite.
FIGURE 2.
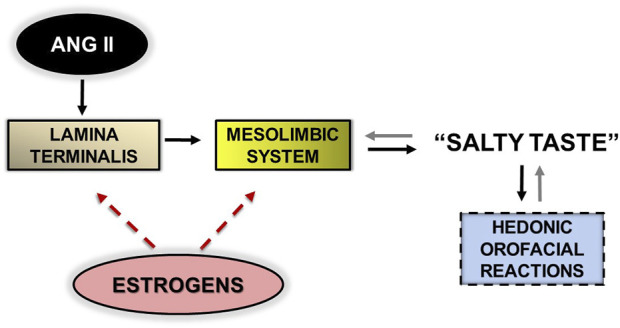
Flow diagram representing the sequence of brain events triggered by the action of angiotensin II (ANG II) on the lamina terminalis, leading to “pleasure” in experience of salty taste, expressed by hedonic, orofacial motor somatic reactions (solid black arrows). The presence of salt in the oral cavity feeds back sensory information (via the brainstem; not shown) to the mesolimbic system to control hedonic responses (solid gray arrows). Estrogen activates brain mechanisms that attenuate this flow, thereby opposing the production of hedonic reactions to salty taste; it acts on the lamina terminalis (or adjacent areas; see text) and on the mesolimbic system (red dotted arrows).
Funding Statement
This research was funded by CNPq (306833/2021-2), FAPESP, UNESP.
Author contributions
Conceptualization: CA, JM, and LL; methodology: EP and AZ; writing—original draft preparation: EP and CA; writing—review and editing: JM, LL, EP, AZ, and CA; supervision: CA. All authors contributed to the article and approved the submitted version.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
- Adolf C., Gorge V., Heinrich D. A., Hoster E., Schneider H., Handgriff L., et al. (2021). Altered taste perception for sodium chloride in patients with primary aldosteronism: A prospective cohort study. Hypertension 77, 1332–1340. 10.1161/HYPERTENSIONAHA.120.16440 [DOI] [PubMed] [Google Scholar]
- Allen A. M., Macgregor D. P., Mckinley M. J., Mendelsohn F. A. (1999). Angiotensin II receptors in the human brain. Regul. Pept. 79, 1–7. 10.1016/s0167-0115(98)00138-4 [DOI] [PubMed] [Google Scholar]
- Allen A. M., Mckinley M. J., Mendelsohn F. A. O. (1988). Comparative neuroanatomy of angiotensin II receptor localization in the mammalian hypothalamus. Clin. Exp. Pharmacol. Physiol. 15, 137–145. 10.1111/j.1440-1681.1988.tb01055.x [DOI] [PubMed] [Google Scholar]
- Almeida-Pereira G., Vilhena-Franco T., Coletti R., Cognuck S. Q., Silva H. V. P., Elias L. L. K., et al. (2019). 17β-Estradiol attenuates p38MAPK activity but not PKCα induced by angiotensin II in the brain. J. Endocrinol. 240, 345–360. 10.1530/JOE-18-0095 [DOI] [PubMed] [Google Scholar]
- Andrade C. A., Andrade-Franze G. M., De Paula P. M., De Luca L. A., Jr., Menani J. V. (2014). Role of α2-adrenoceptors in the lateral parabrachial nucleus in the control of body fluid homeostasis. Braz J. Med. Biol. Res. 47, 11–18. 10.1590/1414-431X20133308 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andrade-Franze G. M. F., Pereira E. D., Jr., Yosten G. L. C., Samson W. K., Menani J. V., De Luca L. A., Jr., et al. (2020). Blockade of erk1/2 activation with U0126 or Pep7 reduces sodium appetite and angiotensin ii-induced pressor responses in spontaneously hypertensive rats. Peptides 136, 170439. 10.1016/j.peptides.2020.170439 [DOI] [PubMed] [Google Scholar]
- Antunes Rodrigues J., Covian M. R. (1963). Hypothalamic control of sodium chloride and water intake. Acta Physiol. Lat. Am. 13, 94–100. [PubMed] [Google Scholar]
- Beauchamp G. K., Bertino M., Burke D., Engelman K. (1990). Experimental sodium depletion and salt taste in normal human volunteers. Am. J. Clin. Nutr. 51, 881–889. 10.1093/ajcn/51.5.881 [DOI] [PubMed] [Google Scholar]
- Berridge K. C. (2018). Evolving concepts of emotion and motivation. Front. Psychol. 9, 1647. 10.3389/fpsyg.2018.01647 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berridge K. C., Flynn F. W., Schulkin J., Grill H. J. (1984). Sodium depletion enhances salt palatability in rats. Behav. Neurosci. 98, 652–660. 10.1037//0735-7044.98.4.652 [DOI] [PubMed] [Google Scholar]
- Berridge K. C., Kringelbach M. L. (2015). Pleasure systems in the brain. Neuron 86, 646–664. 10.1016/j.neuron.2015.02.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berridge K. C. (2000). Measuring hedonic impact in animals and infants: Microstructure of affective taste reactivity patterns. Neurosci. Biobehav Rev. 24, 173–198. 10.1016/s0149-7634(99)00072-x [DOI] [PubMed] [Google Scholar]
- Berridge K. C., Schulkin J. (1989). Palatability shift of a salt-associated incentive during sodium depletion. Q. J. Exp. Psychol. B 41, 121–138. [PubMed] [Google Scholar]
- Bhatia K., Zimmerman M. A., Sullivan J. C. (2013). Sex differences in angiotensin-converting enzyme modulation of Ang (1-7) levels in normotensive WKY rats. Am. J. Hypertens. 26, 591–598. 10.1093/ajh/hps088 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blume A., Lebrun C. J., Herdegen T., Bravo R., Linz W., Mollenhoff E., et al. (1997). Increased brain transcription factor expression by angiotensin in genetic hypertension. Hypertension 29, 592–598. 10.1161/01.hyp.29.2.592 [DOI] [PubMed] [Google Scholar]
- Brown R. D., Hilliard L. M., Head G. A., Jones E. S., Widdop R. E., Denton K. M. (2012). Sex differences in the pressor and tubuloglomerular feedback response to angiotensin II. Hypertension 59, 129–135. 10.1161/HYPERTENSIONAHA.111.178715 [DOI] [PubMed] [Google Scholar]
- Champagne F. A., Curley J. P. (2016). Plasticity of the maternal brain across the lifespan. New Dir. Child. Adolesc. Dev. 2016, 9–21. 10.1002/cad.20164 [DOI] [PubMed] [Google Scholar]
- Ciriello J., Lawrence D., Pittman Q. J. (1984). Electrophysiological identification of neurons in the parabrachial nucleus projecting directly to the hypothalamus in the rat. Br Res. 322, 388–392. 10.1016/0006-8993(84)90140-9 [DOI] [PubMed] [Google Scholar]
- Dalmasso C., Antunes-Rodrigues J., Vivas L., De Luca L. A., Jr. (2015). Mapping brain Fos immunoreactivity in response to water deprivation and partial rehydration: Influence of sodium intake. Physiol. Behav. 151, 494–501. 10.1016/j.physbeh.2015.08.020 [DOI] [PubMed] [Google Scholar]
- Danielsen J., Buggy J. (1980). Depression of ad lib and angiotensin-induced sodium intake at oestrus. Br Res. Bull. 5, 501–504. 10.1016/0361-9230(80)90253-1 [DOI] [PubMed] [Google Scholar]
- De Luca L. A., Jr., Pereira-Derderian D. T., Vendramini R. C., David R. B., Menani J. V. (2010). Water deprivation-induced sodium appetite. Physiol. Behav. 100, 535–544. 10.1016/j.physbeh.2010.02.028 [DOI] [PubMed] [Google Scholar]
- De Luca L. A., Jr., Xu Z., Schoorlemmer G. H., Thunhorst R. L., Beltz T. G., Menani J. V., et al. (2002). Water deprivation-induced sodium appetite: Humoral and cardiovascular mediators and immediate early genes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 282, R552–R559. 10.1152/ajpregu.00295.2000 [DOI] [PubMed] [Google Scholar]
- De Souza J. T., Matsubara L. S., Menani J. V., Matsubara B. B., Johnson A. K., De Gobbi J. I. (2012). Higher salt preference in heart failure patients. Appetite 58, 418–423. 10.1016/j.appet.2011.09.021 [DOI] [PubMed] [Google Scholar]
- Dean S. A., Tan J., White R., O'brien E. R., Leenen F. H. (2006). Regulation of components of the brain and cardiac renin-angiotensin systems by 17beta-estradiol after myocardial infarction in female rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 291, R155–R162. 10.1152/ajpregu.00497.2005 [DOI] [PubMed] [Google Scholar]
- Denton D. A., Cox P., Mckinley M., Nelson J., Weisinger R. (1982). Salt and water homeostasis. Clin. Exp. Pharmacol. Physiology 9, 518. [Google Scholar]
- Di Nicolantonio R., Kren V., Zidek V., Pravenec M. (2004). Salt preference of congenic strains derived from the spontaneously hypertensive rat. Physiol. Behav. 80, 617–622. 10.1016/j.physbeh.2003.11.001 [DOI] [PubMed] [Google Scholar]
- Dinicolantonio R., Hutchinson J. S., Mendelsohn F. A. (1982). Exaggerated salt appetite of spontaneously hypertensive rats is decreased by central angiotensin-converting enzyme blockade. Nature 298, 846–848. 10.1038/298846a0 [DOI] [PubMed] [Google Scholar]
- Duffy V. B., Bartoshuk L. M., Moore R. S., Rodin J. (1998). Taste changes across pregnancy. Ann. N. Y. Acad. Sci. 855, 805–809. 10.1111/j.1749-6632.1998.tb10663.x [DOI] [PubMed] [Google Scholar]
- Elmarakby A. A., Sullivan J. C. (2021). Sex differences in hypertension: Lessons from spontaneously hypertensive rats (SHR). Clin. Sci. (Lond) 135, 1791–1804. 10.1042/CS20201017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ely D., Turner M., Milsted A. (2000). Review of the Y chromosome and hypertension. Braz J. Med. Biol. Res. 33, 679–691. 10.1590/s0100-879x2000000600009 [DOI] [PubMed] [Google Scholar]
- Epstein A. N. (1991). Neurohormonal control of salt intake in the rat. Br Res. Bull. 27, 315–320. 10.1016/0361-9230(91)90118-4 [DOI] [PubMed] [Google Scholar]
- Fernandes N. A. R., Santos B. M., Andrade-Franze G. M. F., De Paula P. M., Andrade C. A. F., Vanderlei Menani J., et al. (2022). Rapid onset sodium appetite and orofacial responses to intraoral capsaicin and hypertonic NaCl in the rat. Appetite 174, 106014. 10.1016/j.appet.2022.106014 [DOI] [PubMed] [Google Scholar]
- Fitts D. A., Masson D. B. (1989). Forebrain sites of action for drinking and salt appetite to angiotensin or captopril. Behav. Neurosci. 103, 865–872. 10.1037/h0092457 [DOI] [PubMed] [Google Scholar]
- Fitzsimons J. T. (1998). Angiotensin, thirst, and sodium appetite. Physiol. Rev. 78, 583–686. 10.1152/physrev.1998.78.3.583 [DOI] [PubMed] [Google Scholar]
- Formenti S., Bassi M., Nakamura N. B., Schoorlemmer G. H., Menani J. V., Colombari E. (2013). Hindbrain mineralocorticoid mechanisms on sodium appetite. Am. J. Physiol. Regul. Integr. Comp. Physiol. 304, R252–R259. 10.1152/ajpregu.00385.2011 [DOI] [PubMed] [Google Scholar]
- Fulwiler C. E., Saper C. B. (1984). Subnuclear organization of the efferent connections of the parabrachial nucleus in the rat. Br Res. Rev. 7, 229–259. 10.1016/0165-0173(84)90012-2 [DOI] [PubMed] [Google Scholar]
- Grill H. J., Bernstein I. L. (1988). Strain differences in taste reactivity to NaCl. Am. J. Physiol. 255, R424–R430. 10.1152/ajpregu.1988.255.3.R424 [DOI] [PubMed] [Google Scholar]
- Grill H. J., Norgren R. (1978). The taste reactivity test.I. Mimetic responses to gustatory stimuli in neurologically normal rats. Br Res. 143, 263–279. 10.1016/0006-8993(78)90568-1 [DOI] [PubMed] [Google Scholar]
- Hay M., Xue B., Johnson A. K. (2014). Yes! Sex matters: Sex, the brain and blood pressure. Curr. Hypertens. Rep. 16, 458. 10.1007/s11906-014-0458-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- He F. J., Macgregor G. A. (2012). Salt intake, plasma sodium, and worldwide salt reduction. Ann. Med. 44 (1), S127–S137. 10.3109/07853890.2012.660495 [DOI] [PubMed] [Google Scholar]
- Hsu T. M., Mccutcheon J. E., Roitman M. F. (2018). Parallels and overlap: The integration of homeostatic signals by mesolimbic dopamine neurons. Front. Psychiatry 9, 410. 10.3389/fpsyt.2018.00410 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jhamandas J. H., Petrov T., Harris K. H., Vu T., Krukoff T. L. (1996). Parabrachial nucleus projection to the amygdala in the rat: Electrophysiological and anatomical observations. Br. Res. Bull. 39, 115–126. 10.1016/0361-9230(95)02084-5 [DOI] [PubMed] [Google Scholar]
- Johnson A., Thunhorst R. (2007). “The neuroendocrinology, neurochemistry and molecular biology of thirst and salt appetite,” in Handbook of neurochemistry and molecular neurobiology: Behavioral neurochemistry, neuroendocrinology and molecular neurobiology Lajtha A., Laustein J. 3rd (New York, NY: Springer; ), 641–687. [Google Scholar]
- Keast R. S. J., Breslin P. A. (2003). An overview of binary taste–taste interactions. Food Qual. Prefer. 14, 111–124. 10.1016/s0950-3293(02)00110-6 [DOI] [Google Scholar]
- Kisley L. R., Sakai R. R., Fluharty S. J. (1999). Estrogen decreases hypothalamic angiotensin II AT1 receptor binding and mRNA in the female rat. Br. Res. 844, 34–42. 10.1016/s0006-8993(99)01815-6 [DOI] [PubMed] [Google Scholar]
- Krause E. G., Curtis K. S., Stincic T. L., Markle J. P., Contreras R. J. (2006). Oestrogen and weight loss decrease isoproterenol-induced Fos immunoreactivity and angiotensin type 1 mRNA in the subfornical organ of female rats. J. Physiol. 573, 251–262. 10.1113/jphysiol.2006.106740 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leshem M., Rudoy J. (1997). Hemodialysis increases the preference for salt in soup. Physiol. Behav. 61, 65–69. 10.1016/s0031-9384(96)00319-8 [DOI] [PubMed] [Google Scholar]
- Lewis S. J., Allen A. M., Verberne A. J. M., Figdor R., Jarrot B., Mendelsohn F. a. O. (1986). Angiotensin II receptor binding in the rat nucleus tractus solitarii is reduced after unilateral nodose ganglionectomy or vagotomy. Eur. J. Pharmacol. 125, 305–307. 10.1016/0014-2999(86)90043-9 [DOI] [PubMed] [Google Scholar]
- Liem D. G., Miremadi F., Keast R. S. (2011). Reducing sodium in foods: The effect on flavor. Nutrients 3, 694–711. 10.3390/nu3060694 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lucia K. J., Curtis K. S. (2018). Behavioral responses and fluid regulation in male rats after combined dietary sodium deficiency and water deprivation. Physiol. Behav. 184, 162–171. 10.1016/j.physbeh.2017.11.025 [DOI] [PubMed] [Google Scholar]
- Martin L. J., Sollars S. I. (2017). Contributory role of sex differences in the variations of gustatory function. J. Neurosci. Res. 95, 594–603. 10.1002/jnr.23819 [DOI] [PubMed] [Google Scholar]
- Mckinley M. J., Allen A. M., Clevers J., Paxinos G., Mendelsohn F. A. O. (1987). Angiotensin receptor binding in human hypothalamus: Autoradiographic localization. Br. Res. 420, 375–379. 10.1016/0006-8993(87)91260-1 [DOI] [PubMed] [Google Scholar]
- Mecawi A. S., Macchione A. F., Nunez P., Perillan C., Reis L. C., Vivas L., et al. (2015). Developmental programing of thirst and sodium appetite. Neurosci. Biobehav Rev. 51, 1–14. 10.1016/j.neubiorev.2014.12.012 [DOI] [PubMed] [Google Scholar]
- Menani J. V., De Luca L. A., Jr., Johnson A. K. (2014). Role of the lateral parabrachial nucleus in the control of sodium appetite. Am. J. Physiol. Regul. Integr. Comp. Physiol. 306, R201–R210. 10.1152/ajpregu.00251.2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mill J. G., Baldo M. P., Molina M., Schmidt M. I., Barreto S. M., Chor D., et al. (2019). Sex-specific patterns in the association between salt intake and blood pressure: The ELSA-Brasil study. J. Clin. Hypertens. (Greenwich) 21, 502–509. 10.1111/jch.13509 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Millard-Stafford M., Wendland D. M., O'dea N. K., Norman T. L. (2012). Thirst and hydration status in everyday life. Nutr. Rev. 70 (2), S147–S151. 10.1111/j.1753-4887.2012.00527.x [DOI] [PubMed] [Google Scholar]
- Minegishi S., Luft F. C., Titze J., Kitada K. (2020). Sodium handling and interaction in numerous organs. Am. J. Hypertens. 33, 687–694. 10.1093/ajh/hpaa049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orysiak J., Mlynarczyk M., Tomaszewski P. (2022). Hydration status in men working in different thermal environments: A pilot study. Int. J. Environ. Res. Public Health 19, 5627. 10.3390/ijerph19095627 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pereira E. D., Jr., Dantas R. M., Andrade-Franze G. M. F., De Luca L. A., Jr., Menani J. V., Andrade C. A. F. (2019). Estradiol modulates the palatability of 0.3M NaCl in female rats during sodium appetite. Appetite 133, 252–261. 10.1016/j.appet.2018.11.012 [DOI] [PubMed] [Google Scholar]
- Pereira E. D., Jr., Faria A. M., Andrade-Franze G. M. F., Menani J. V., De Luca L. A., Jr., Andrade C. A. F. (2021). Sodium palatability in male spontaneously hypertensive rats. Horm. Behav. 130, 104952. 10.1016/j.yhbeh.2021.104952 [DOI] [PubMed] [Google Scholar]
- Pereira E. D., Jr., Oliveira L. M., Coletto-Nunes G., Souza P. P. C., Menani J. V., De Luca L. A., Jr., et al. (2022). Central angiotensinergic mechanisms in female spontaneously hypertensive rats treated with estradiol. Appetite 174, 106012. 10.1016/j.appet.2022.106012 [DOI] [PubMed] [Google Scholar]
- Pereira-Derderian D. T., Vendramini R. C., Menani J. V., De Luca L. A., Jr. (2010). Water deprivation-induced sodium appetite and differential expression of encephalic c-Fos immunoreactivity in the spontaneously hypertensive rat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 298, R1298–R1309. 10.1152/ajpregu.00359.2009 [DOI] [PubMed] [Google Scholar]
- Rowland N. E., Li B. H., Fregly M. J., Smith G. C. (1995). Fos induced in brain of spontaneously hypertensive rats by angiotensin II and co-localization with AT-1 receptors. Br. Res. 675, 127–134. 10.1016/0006-8993(95)00047-t [DOI] [PubMed] [Google Scholar]
- Rowland N. E., Morian K. R. (1999). Roles of aldosterone and angiotensin in maturation of sodium appetite in furosemide-treated rats. Am. J. Physiol. 276, R1453–R1460. 10.1152/ajpregu.1999.276.5.R1453 [DOI] [PubMed] [Google Scholar]
- Sakai R. R., Nicolaidis S., Epstein A. N. (1986). Salt appetite is suppressed by interference with angiotensin II and aldosterone. Am. J. Physiol. 251, R762–R768. 10.1152/ajpregu.1986.251.4.R762 [DOI] [PubMed] [Google Scholar]
- Sampson A. K., Hilliard L. M., Moritz K. M., Thomas M. C., Tikellis C., Widdop R. E., et al. (2012). The arterial depressor response to chronic low-dose angiotensin II infusion in female rats is estrogen dependent. Am. J. Physiol. Regul. Integr. Comp. Physiol. 302, R159–R165. 10.1152/ajpregu.00256.2011 [DOI] [PubMed] [Google Scholar]
- Santollo J., Daniels D. (2015). Control of fluid intake by estrogens in the female rat: Role of the hypothalamus. Front. Syst. Neurosci. 9, 25. 10.3389/fnsys.2015.00025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santollo J., Daniels D., Leshem M., Schulkin J. (2023). Sex differences in salt appetite: Perspectives from animal models and human studies. Nutrients 15, 208. 10.3390/nu15010208 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schulkin J. (1991). The allure of salt. Psychobiology 19, 116–121. 10.3758/bf03327180 [DOI] [Google Scholar]
- Shenoy V., Grobe J. L., Qi Y., Ferreira A. J., Fraga-Silva R. A., Collamat G., et al. (2009). 17beta-Estradiol modulates local cardiac renin-angiotensin system to prevent cardiac remodeling in the DOCA-salt model of hypertension in rats. Peptides 30, 2309–2315. 10.1016/j.peptides.2009.09.005 [DOI] [PubMed] [Google Scholar]
- Shigemura N., Iwata S., Yasumatsu K., Ohkuri T., Horio N., Sanematsu K., et al. (2013). Angiotensin II modulates salty and sweet taste sensitivities. J. Neurosci. 33, 6267–6277. 10.1523/JNEUROSCI.5599-12.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takamata A., Mack G. W., Gillen C. M., Nadel E. R. (1994). Sodium appetite, thirst, and body fluid regulation in humans during rehydration without sodium replacement. Am. J. Physiol. 266, R1493–R1502. 10.1152/ajpregu.1994.266.5.R1493 [DOI] [PubMed] [Google Scholar]
- Tazumi S., Yokota N., Kawakami M., Omoto S., Takamata A., Morimoto K. (2016). Effects of estrogen replacement on stress-induced cardiovascular responses via renin-angiotensin system in ovariectomized rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 311, R898–R905. 10.1152/ajpregu.00415.2015 [DOI] [PubMed] [Google Scholar]
- Thunhorst R. L., Johnson A. K. (1994). Renin-angiotensin, arterial blood pressure, and salt appetite in rats. Am. J. Physiol. 266, R458–R465. 10.1152/ajpregu.1994.266.2.R458 [DOI] [PubMed] [Google Scholar]
- Thunhorst R. L., Morris M., Johnson A. K. (1994). Endocrine changes associated with a rapidly developing sodium appetite in rats. Am. J. Physiol. 267, R1168–R1173. 10.1152/ajpregu.1994.267.5.R1168 [DOI] [PubMed] [Google Scholar]
- Thunhorst R. L., Xu Z., Cicha M. Z., Zardetto-Smith A. M., Johnson A. K. (1998). Fos expression in rat brain during depletion-induced thirst and salt appetite. Am. J. Physiol. 274, R1807–R1814. 10.1152/ajpregu.1998.274.6.r1807 [DOI] [PubMed] [Google Scholar]
- Vivas L., Godino A., Dalmasso C., Caeiro X. E., Macchione A. F., Cambiasso M. J. (2014). “Neurochemical circuits subserving fluid balance and baroreflex: A role for serotonin, oxytocin, and gonadal steroids,” in Neurobiology of body fluid homeostasis: Transduction and integration. Editors De LucaJr. L. A., Menani J. V., Johnson A. K. (Boca Raton, FL: ). [PubMed] [Google Scholar]
- World Health Organization (2016). Global NCD target: Reduce high blood pressure. Available at: https://apps.who.int/iris/handle/10665/312279 (Accessed September 1, 2016).
- Zenatti A. A., Pereira E. D., Jr., Possari J., Andrade C. a. F., Menani J. V., De Luca L. A., Jr. (2021). Interference with the renin-angiotensin system reduces the palatability of 0.3 M NaCl in sodium-deplete rats. Appetite 158, 105037. 10.1016/j.appet.2020.105037 [DOI] [PubMed] [Google Scholar]
- Zhang J., Zhang N., Liu S., Du S., Ma G. (2022). Young adults with higher salt intake have inferior hydration status: A cross-sectional study. Nutrients 14, 287. 10.3390/nu14020287 [DOI] [PMC free article] [PubMed] [Google Scholar]