

Abstract
At the top of the midbrain is the inferior colliculus (IC), which functions as the major hub for processing auditory information. Despite the functional significance of neurons in the IC, our understanding of their formation is limited. In this study, we identify the embryonic patterning gene Dbx1 as a key molecular player that governs genetic programs for IC survival. We find that Dbx1 plays a critical role in preventing apoptotic cell death in postnatal IC by transcriptionally repressing c-Jun and pro-apoptotic BH3 only factors. Furthermore, by employing combined approaches, we uncover that Tcf7l2 functions downstream of Dbx1. Loss of Tcf7l2 function causes IC phenotypes with striking similarity to those of Dbx1 mutant mice, which include defective embryonic maturation and postnatal deletion of the IC. Finally, we demonstrate that the Dbx1-Tcf7l2 cascade functions upstream of Ap-2δ, which is essential for IC development and survival. Together, these results unravel a novel molecular mechanism for IC maintenance, which is indispensable for normal brain development.
Subject terms: Development, Gene regulation
Introduction
The auditory midbrain or inferior colliculus (IC) is the major hub serving as a critical integration center of the central auditory pathway [1]. To process the perception of sound stimuli, neurons in the auditory system form precise circuits, which emerge from cochlea ganglion neurons. The IC receives virtually all ascending brainstem inputs and descending projection from the auditory cortex [2, 3]. In turn, the IC projects to the medial geniculate body, the thalamic relay nucleus [4, 5]. Neurons in the IC also receive many non-auditory inputs from the superior colliculus (SC), which is known as the optic midbrain. The IC is functionally subdivided into core (central nucleus) and shell (dorsal and lateral cortex) subregions according to dendritic morphology and axonal trajectories [1]. Despite the functional significance of IC neurons, our understanding of their formation is limited.
During early embryogenesis, the territories of midbrain progenitors are specified and assigned into seven subdivisions (m1-m7) with specific combinations of gene expression codes [6, 7]. The m1-m2 regions are organized into the dorsal midbrain or tectum, which further subdivide into the IC and SC. Early growth and patterning of the tectum are tightly linked to the key signaling molecules of the isthmic organizer [8–11]. FGF signaling functions in a dose- and time- dependent manner to control tectal growth, specification, and survival [10, 12, 13]. Distinct levels of FGF activity play crucial roles in regulating SC and IC fates, as reduced FGF signaling results in disruption of the IC [10, 12, 14–16]. In addition, inactivating Fgf8 at different stages leads to variable truncation of the developing tectum [17]. Similarly, loss of the tectum has been also observed in embryos that lack Wnt1 or En1 function although the phenotypes of Wnt1- or En1-deficient embryos with altered midbrain and cerebellum can vary [18–20]. Impaired cellular processes including defective proliferation, cell fate transformation, and loss of tectal stem zone are likely to be responsible for the loss of the tectum [12, 13, 17]. Furthermore, abnormal apoptosis also appears to be one of the major causes for embryonic tectum deletion [10, 21]. Pax and Meis2 play differential roles in forming the tectum [14, 22–24], and Otx2 contributes to the control of multiple aspects of midbrain development [25–28]. During neurogenesis between E11.5 and E14.5 [29–32], the tectum produces glutamatergic and GABAergic neurons in the SC and IC [6, 7]. The transcription factors Ascl1, Helt, Gata2 and Tal2 play selective functions in determining GABAergic neuronal identity along the DV axis [6, 7, 33–37]. Although previous works have greatly broadened our understanding of the early phase of tectal development, little is known about the molecular mechanisms underlying the differentiation and survival of the IC at the late embryonic and adult stages.
Herein we present evidence that the transcriptional cascade of the Dbx1, Tcf7l2 and Ap-2δ (Tfap2d) transcription factors regulates the survival of the IC. The homeodomain transcription factor Dbx1 is expressed in various regions of the developing nervous system [38], and is required for regulating neuronal cell fate determination. In the spinal cord, Dbx1 is necessary for specifying distinct identities of V0 and V1 interneurons that coordinate the alternation of left and right limbs [39–41]. In the hindbrain, Dbx1 is required for regulating the identity and function of the preBötzinger respiratory neurons [42, 43]. In the forebrain, Dbx1 is critical for regulating the specification of hypothalamic neurons that control innate behaviors [44]. A recent study by gain- and loss-of-function analyses revealed crucial roles of Dbx1 in regulating midline crossing by midbrain commissural axons [45]. During early embryogenesis, Dbx1 expression in the midbrain is restricted to the m1-m2 progenitor domains, which generate IC and SC neurons [28, 38]. At late embryonic stages, Dbx1 expression is excluded from the SC, and persists in the IC (Allen Brain Atlas). These observations raise the possibility that Dbx1 has a later role in regulating IC formation. Tcf7l2, a high mobility group (HMG) box-containing transcription factor, has been known as both the major transducer of Wnt/β-catenin activity and high diabetic risk factor [46–48]. Tcf7l2 expression initiates at E10.5, and is abundantly detected in the diencephalon and midbrain [49]. Loss of Tcf7l2 function leads to pituitary hyperplasia [50], defective oligodendrogenesis [51–53], and disruption of thalamic and habenula development [54–56]. Despite strong and persistent expression in the dorsal midbrain during embryonic and postnatal stages, the possible role of Tcf7l2 in tectal differentiation remains unexplored.
By employing combined approaches, we demonstrate that Dbx1-Tcf7l2 transcriptional cascade orchestrates a genetic program for the development of the IC. Notably, this regulatory cascade is required for postnatal survival of the IC by playing an essential anti-apoptotic role through transcriptional suppression of c-Jun and BH3-only factors such as Hrk and Bim. We further show that the Dbx1-Tcf7l2 cascade acts upstream of Ap-2δ, which regulates neuronal survival in the IC [57]. Together, our data begin to define a novel molecular pathway for IC maintenance, which is indispensable for normal brain development.
Results
Dbx1 orchestrates a genetic program for the development of the inferior colliculus
To investigate the role of Dbx1 in the regulation of IC differentiation, we first generated Dbx1del mice in which Dbx1 is inactivated at the pre-implantation stages, by crossing Dbx1loxp/loxp mice [44] to EIIa-Cre [58] transgenic mice. We then analyzed global gene expression in the IC of control and Dbx1del mutant embryos on E18.5 by RNA-seq. In this comparison, we found 1,524 genes differentially expressed (≥0.4 or ≤ −0.4 log2 fold-change; Padj < 0.05), 1,213 of them upregulated and 311 downregulated in Dbx1del mutants (Fig. 1A; Table S1; Table S2). The differentially expressed genes (DEGs) were significantly enriched in gene ontology (GO) terms associated with transcription factor activity, neuron projection, cell adhesion, synaptic function, apoptotic process, Wnt signaling, ion channel activity, extracellular matrix, and neurotransmitter signaling (Fig. 1B; Table S3). To assess whether the DEGs from these groups are specific to the IC, we surveyed corresponding gene expression data from the Allen Brain Atlas (https://mouse.brain-map.org). We found that 100% of the DEGs highly enriched in the IC belonged to the downregulated target genes (Fig. 1C, F, H). To validate this loss of Dbx1-altered gene expression in the IC, we selected a subset of the identified DEGs for further analysis by in situ hybridization (ISH). IC-enriched genes included transcription factors Tcf7l2, Foxp2, Irx1/2, Shox2, Pou4f3, Mef2c, Meis2, Maf, Rora, and Bcl11a/b; neuronal adhesion Reln [59]; axon projection Gas7 and Robo2 [45, 60]; synaptic transmission Grm5, Grik2, Nrgn, Kcnk9 and Ndst4 [61] (Figs. 1D, G, I, S1A, B). By contrast, the DEGs that were upregulated in the Dbx1del mutant were dominated by genes that were either depleted from the IC, broadly expressed, or undetectable (Fig. 1C, E, F, G, H, I). Among them were transcription factor Otx2, Pou4f1, Barhl1, Ebf1/3, Irx5/6, and Lhx2; chromatin remodeling factor Chd7; axon guidance Robo1, Nrp2, Ntn1, Efna5, and Islr2 [62]; basement membrane Col4a5 [63]; cell adhesion Pcdh19 [64] (Fig. 1C, F, H). Taken together, these data identify a set of Dbx1-regulated genes with potentially novel roles in late phases of IC development.
Fig. 1. Differential expression profiling identifies Dbx1 target genes in the IC.

A Volcano plot representing transcriptional changes in Dbx1del IC versus wildtype IC at E18.5. Orange dots correspond to upregulated genes whereas blue dots correspond to downregulated genes. B Gene ontology (GO) analysis of differentially expressed genes in the IC of Dbx1del embryos at E18.5 compared with wildtype embryos. Bar charts showing 10 GO terms for biological process, molecular function and cellular components, ranked by fold enrichment. C, F, H Clustering heatmaps represent expression patterns of the genes with overrepresented GO terms: transcription factors (C), neuron projection and adhesion (F), and synaptic transmission and excitability (H). Each column represents an independent biological replicate. The log2 values of expression in relation to the median of the row are shown as a blue-red color scale. Localization of each gene expression in the brain was estimated according to the Allen Brain Atlas, and is indicated by boxes. IC-enriched, expression enriched in the IC; IC-depleted, expressed broadly in the brain except for the IC; Ubiquitous, expressed throughout the brain; Low, low overall expression in the brain; N/A, not available in the Allen Brain Atlas. D, E, G, I In situ hybridization on E18.5 sagittal brain sections. Scale bars, 200 μm.
The expression of Tcf7l2 and Meis2 identified the whole IC except for the ventrolateral region, whereas other IC-enriched genes such as Nrgn, Foxp2, Maf, Ndst4, Pou4f3, and Barhl1 could serve as subregion-specific IC markers (Fig. S1A, B). Foxp2, Maf, and Ndst4 are expressed along the DV axis, and partially overlapped with the expression of Nrgn, which is restricted to the dorsolateral region. The ventromedial region in the IC was subdivided into two different cell populations, Ndst4+ Pou4f3- cells and Ndst4- Pou4f3+ cells (asterisk in Fig. S1A, B). Therefore, our datasets provided a list of molecular markers that distinguish the IC from the SC and other brain regions, and could selectively identify different neuronal types in the IC.
Loss of Dbx1 leads to upregulation of BH3 only pro-apoptotic genes and robust neuronal cell death in the IC
Because the RNA profile analysis revealed alterations in the expression of genes involved in apoptotic cell death, we next investigated the potential cellular mechanism of the IC defects in the Dbx1del mutant by determining whether Dbx1 disruption affects neuronal survival in the IC. Among the BH3 only factors implicated in triggering neuronal apoptosis [65–67], the RNA profile showed upregulation of Bim and Hrk (Tables S2, S3; Fig. 2A). Also, c-Jun, the transcription and phosphorylation of which play critical roles in the regulation of BH3 only factors [67, 68], was also upregulated in Dbx1-deficient IC. This effect was further validated by RT-PCR and ISH (Fig. 2A, B). Ectopic expression of Bim, Hrk, and c-Jun was specifically detected in the IC of Dbx1del mutants. To determine whether Dbx1- deficient IC neurons undergo cell death, we next performed immunostaining for cleaved caspase 3 (Cas3) in the developing midbrain of controls and Dbx1del mutants. At E18.5, loss of Dbx1 resulted in ~700% increase in the number of IC neurons undergoing apoptosis, compared with control IC (Fig. 2C). By contrast, in the SC, there was no major difference in apoptotic cell death between controls and Dbx1del mutants (Fig. 2C). To determine the timing of the onset of abnormal cell death, we measured immunoreactivity for active Cas3 at earlier stages. At E12.5 and E15.5, there was no significant difference in the number of Cas3+ cells in the developing midbrain between controls and Dbx1del mutants (Fig. 2D, E). Therefore, most of the cell death in the Dbx1del -IC occurred at late embryonic stages. These data indicate that Dbx1 inactivation leads to specific loss of IC cells.
Fig. 2. Loss of Dbx1 leads to increased levels of c-Jun and BH3 only factors, and to cell death in the IC.

A Differential expression of BH3 only (Bid, Bad, Bim, Puma, Hrk) factors and c-Jun in E18.5 Dbx1del compared with control based on RNA-seq (pink bars; ****FDR q ≤ 0.0001) and qRT-PCR (light blue bars; Student’s t-test; ***p ≤ 0.001); error bars indicate standard error for log2 fold change). B In situ hybridization with Bim, Hrk, and c-Jun riboprobes in E18.5 sagittal brain sections. C–E Immunohistochemistry using anti-cleaved caspase 3 (Cas3) was performed on E18.5 coronal sections (C) and E12.5 (D) and E15.5 (E) sagittal sections to detect apoptosis. E18.5, but not E12.5 or E15.5 Dbx1del mutants exhibited increased numbers of apoptotic cells in the IC. For E12.5, Cas3+ cells were counted from 5 sections from each embryo (n = 5). For counting Cas3+ cells at E15.5 and E18.5, 10–20 counting sites (counting grid for SC, 200 μm × 200 μm; for IC, 100 μm × 100 μm) were evaluated in one section. The values were calculated from 5 sections from each embryo (n = 5). *P < 0.01 (Student’s t-test). Scale bars, 200 μm.
Postnatal deletion of the IC in the absence of Dbx1
We next explored the consequence of Dbx1-deficiency on the postnatal development of IC neurons. Null mutation of Dbx1 causes perinatal lethality due to asphyxia resulting from loss of the core respiratory rhythm generator in the hindbrain preBötC complex [42, 43], which precludes analysis of the long- term consequences on dorsal midbrain development. To overcome this limitation, we generated a mouse line driving Cre- mediated recombination by using a Sox2 regulatory element (Sre), which was shown to direct the transcription of a reporter gene to distinct brain regions except for the hindbrain [69] (Fig. S2A, B). In Sre-Cre; Dbx1loxp/del mice (Dbx1cko), Dbx1 expression was abrogated in the midbrain at E12.5 (Fig. S2C). At P3, when the fissure between the SC and IC is apparent, external anatomical examination of postnatal brains enabled us to identify Dbx1cko mice among their littermates. Dbx1cko mutant IC was slightly reduced in size, compared with controls, whereas no overt morphological malformations were found in other brain regions (Fig. 3A). Consistent with the Dbx1del mutant, the IC in Dbx1cko mutants lost expression of Tcf7l2, Nrgn, and Mef2c, but acquired expression of the SC-specific markers Otx2, Pou4f1 and Ap-2α (Fig. 3B). Furthermore, the Dbx1cko mutants displayed robust apoptotic cell death in the IC (Fig. 3C). By P20, the IC continued to grow and form the two rounded posterior eminences of the quadrigeminal plate in controls, whereas in the absence of Dbx1, the IC was almost completely missing, resulting in abutment of the SC on the anterior cerebellum (Fig. 3D). This was corroborated by examination of Tcf7l2, Nrgn and Otx2 expressions on sagittal sections (Fig. 3D). These data demonstrate that the embryonic changes mentioned above result in alterations of the postnatal IC, and that Dbx1 is selectively required for IC neuronal survival by playing an essential anti-apoptotic role.
Fig. 3. Absence of the inferior colliculus in Dbx1cko mutant mice.

A A close-up dorsal view of P3 control and Dbx1cko mutant brains. Dashed lines indicate the position of the IC. Compared with controls, Dbx1cko mutant brains showed a reduction in IC size. B In situ hybridization on P3 sagittal sections of control and Dbx1cko brains. C Immunohistochemistry with anti-Cas3 on P3 sagittal sections of control and Dbx1cko brains. Cas3+ cells were counted from five sections from three embryos using 20 counting sites (counting grid 100 μm × 100 μm) in one section. *P < 0.05; **P < 0.001 (Student’s t-test). D The P20 brain phenotype of Dbx1cko mutants. The entire brain (dorsal view) and histological sections of the midbrain. Section ISH on sagittal sections indicates an almost complete lack of the IC with the SC fused to the cerebellum in Dbx1cko mutants. Scale bars, 200 μm.
Transcriptome analyses reveal similar regulatory functions for Dbx1 and Tcf7l2
Among the Dbx1-regulated genes that our transcriptome analyses identified, Tcf7l2 is a good candidate for mediating the function of Dbx1, given the temporal and spatial pattern of Tcf7l2 expression in the developing midbrain. Dbx1 and Tcf7l2 transcription in the midbrain was limited to m1-m2 domains, which are organized into the IC and SC. Dbx1 was expressed in the progenitor region, and Tcf7l2 in the post-mitotic region (Fig. S3). Early loss of Dbx1 severely downregulated Tcf7l2 expression in the IC and SC, whereas later depletion of Dbx1 abrogated Tcf7l2 expression in the IC despite normal expression in the SC (Fig. S3), indicating distinct temporal requirements for Dbx1 in the regulation of Tcf7l2 transcription.
To explore the potential role of Tcf7l2 in the regulation of tectal development, we generated Tcf7l2del mice by crossing Tcf7l2loxp/loxp mice to the EIIa-Cre line, and analyzed transcriptome changes in the IC of control and Tcf7l2del mutant embryos on E18.5 by RNA-Seq. This comparison revealed 633 DEGs (≥0.4 or ≤ −0.4 log2 fold-change; Padj < 0.05), 371 of them upregulated and 262 downregulated in Tcf7l2del mutant (Tables S1, S4, S5; Fig. 4A, B). To identify genes that are co-regulated by Dbx1 and Tcf7l2, we intersected these datasets from Tcf7l2del mutants with our list of Dbx1-regulated genes. Out of 633 DEGs that were regulated by Tcf7l2, 342 targets (54%; 198 upregulated and 144 downregulated), were commonly affected to a similar extent in Dbx1del- and Tcf7l2del-mutants (Table S6; Fig. 4C, D). We selected a subset of the common targets to validate their expression in Tcf7l2del mutants by ISH (Fig. 4E–J). Among them were transcription factors Mef2c, Foxp2, Irx1/2, Shox2, Maf, Rora, Bcl11a, Barhl1, Ebf3, and Irx5; neuron projection/adhesion Gas7, Nrp2, and Islr2; synaptic activity Grm5, Grik2, Nrgn, Ndst4, and Kcnk9. These observations indicate that Tcf7l2-deficient IC closely resembles Dbx1-depleted IC in its gene expression profile. Taken together, these data suggest that Tcf7l2 regulates an IC-specific genetic program, and that Dbx1 and Tcf7l2 have a significant degree of overlapping regulatory function in the development of the IC.
Fig. 4. Tcf7l2 acts downstream of Dbx1 to orchestrate a genetic program of morphological maturation in the developing IC.

A Volcano plot of differential gene expression measured by RNA-seq of wildtype and Tcf7l2del IC. Upregulated or downregulated genes are indicated by orange and blue dots, respectively. B GO term enrichment for gene sets that are differentially regulated in Tcf7l2del embryos. Bar charts showing 10 GO terms for biological process, molecular function, and cellular components, ranked by fold enrichment. C Intersections of differentially expressed genes in Dbx1del and Tcf7l2del IC identify commonly upregulated and downregulated gene sets. D Correlation between the significantly differentially expressed genes (n = 342) observed in Dbx1del and Tcf7l2del mutants. Thresholds used to determine differential expression are indicated by dashed lines (log2FC ≥ 0.4 and ≤ −0.4; Padj ≤ 0.05). The Pearson correlation coefficient is indicated in the lower right corner. E, G, I Clustering heatmap indicating relative expression of the genes with the overrepresented GO terms: transcription factors (E), neuron projection and adhesion (G), and synaptic transmission and excitability (I). Each column represents an independent biological replicate. The log2 values of expression in relation to the median of the row are shown as a blue-red color scale. F, H, J In situ hybridization on E18.5 sagittal brain sections. Scale bars, 200 μm.
Deletion of Tcf7l2 leads to specific loss of the IC due to increased apoptosis
Considering that Dbx1 and Tcf7l2 share IC phenotypes through the regulation of common target genes, we asked whether loss of Tcf7l2 also results in abnormal cell death in the developing midbrain. The RNA profile showed upregulation of Bim, Hrk, and Jun in Tcf7l2-deficient IC with a pattern similar to that in Dbx1del mutants (Table S4; Fig. 5A). This effect was further confirmed by RT-PCR and ISH (Fig. 5A, B). To determine whether Tcf7l2 inactivation leads to apoptotic cell death in the developing midbrain, we performed immunostaining for Cas3 in control and Tcf7l2del mutant embryos. Loss of Tcf7l2 resulted in a ~500% increase in the number of Cas3+ cells in the IC at E18.5, while control and Tcf7l2- deficient SC both had low levels of apoptotic cells (Fig. 5C). We observed no significant differences in the numbers of phospho-histone H3+ mitotic cells in the developing midbrain between normal and mutant ICs (Fig. 5D). To further corroborate that prevention of apoptotic cell death by Tcf7l2 is a critical event during IC development, we examined the effects of Tcf7l2 on the postnatal development of IC neurons. Because Tcf7l2del mutants die shortly after birth due to abnormal intestine development [47, 70], we crossed Tcf7l2loxp/loxp mice with a Sre-Cre line to bypass perinatal lethality (Fig. 5E). Superficial examination of the brains of Tcf7l2cko mutants revealed large deletion of IC with fusion of the SC to the vermis of the cerebellum (Fig. 5F). Loss of Tcf7l2 resulted in remarkable reduction in the size of Nrgn+ domain, while the Otx2+ SC region was normally detected. Thus, the phenotype of the Tcf7l2-depleted IC was essentially the same as that of the Dbx1-deficient IC. Taken together, our data indicate that Tcf7l2 functions downstream of Dbx1 to regulate survival of the IC.
Fig. 5. Tcf7l2 inactivation leads to apoptosis and lack of the inferior colliculus.

A Differential expression of BH3-only apoptotic factors and Jun in E18.5 Tcf7l2del mutants compared with controls based on RNA-seq (pink bars, **FDR q ≤ 0.005; ****FDR q ≤ 0.0001) and qRT-PCR (light blue bars, Student’s t-test; ***p ≤ 0.001; error bars indicate standard error for log2 fold change). B ISH with Bim, Hrk and c-Jun probe in E18.5 sagittal brain sections. C Immunohistochemistry using anti-Cas3 was performed on E15.5 and E18.5 sagittal sections to detect apoptosis. For counting of Cas3+ cells at E15.5 and E18.5, 10–20 counting sites (counting grid 100 μm x 100 μm) were evaluated in one section. The values were calculated for five sections from six embryos for each genotype. *P < 0.001 (Student’s t-test). D Immunohistochemistry for phospho-histone H3 (PH) on sagittal sections of E15.5 midbrain. The number of PH3-positive cells was counted for three sections from three embryos for each genotype. E Section ISH on coronal sections of E12.5 control and Tcf7l2cko embryos with a riboprobe derived from exon 10 of Tcf7l2. F The dorsal view of control and Tcf7l2cko mutant P20 brains. Section ISH on sagittal sections indicates large deletion of the IC, whereas the SC abutted on the cerebellum in Tcf7l2cko mutants. Scale bars, 200 μm.
Ap-2δ is an essential regulator downstream of the Dbx1-Tcf7l2 cascade
The AP-2 family of transcription factors plays important roles in regulating cell differentiation and apoptosis [57, 71, 72]. Among the five AP-2 genes that are expressed in the developing midbrain, only Ap-2δ has expression that persists in the IC during embryonic and adult stages. Interestingly, loss of Ap-2δ causes massive apoptotic cell death, specifically in the posterior midbrain, and postnatal deletion of the IC [57], resembling the phenotypes of Dbx1 and Tcf7l2 mutants. We therefore characterized the transcript levels of Ap-2δ in the midbrain in the absence of Dbx1 or Tcf7l2. RNA-seq and qRT-PCR revealed significant reduction in the mRNA expression of Ap-2δ in Dbx1-deficient or Tcf7l2-deficient IC (Fig. 6A; Tables S2, S4, S6). In Dbx1del embryos, Ap-2δ staining was not detectable in the entire midbrain despite normal expression in other regions during embryogenesis (Fig. 6B). In Dbx1cko embryos, Ap-2δ expression was largely unaffected in the SC, but abolished in the IC (Fig. 6B). Similarly, loss of Tcf7l2 did not alter Ap-2δ expression in the SC, whereas it severely downregulated expression in the IC (Fig. 6B). Therefore, our results and those of others suggest that Ap-2δ functions downstream of the Dbx1-Tcf7l2 regulatory axis to control IC morphogenesis by preventing cell death.
Fig. 6. Ap-2δ is co-regulated by Dbx1 and Tcf7l2 in the IC.
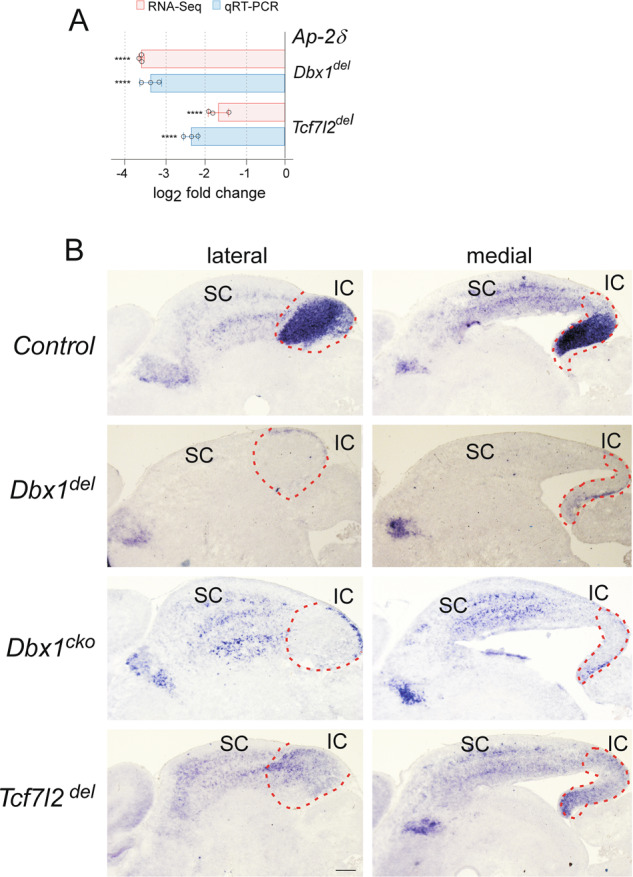
A Differential expression of Ap-2δ in Dbx1del and Tcf7l2del mouse embryos compared with controls based on RNA-seq (****FDR q ≤ 0.0001) and qRT-PCR (Student’s t-test; ****p ≤ 0.0001). Error bars indicate standard error for log2 fold change). B Representative midbrain sections of wild-type, Dbx1del, Dbx1cko, and Tcf7l2del mouse embryos hybridized with Ap-2δ at E18.5. Scale bars, 200 μm.
Tcf7l2 and Ap-2δ are necessary for Dbx1-mediated IC survival
The above-described observations led us to ask whether Tcf7l2 and Ap-2δ are essential components of Dbx1-mediated genetic programs that support the survival of the IC. To address this question, we established IC cell cultures from E15.5 embryonic brains of control and Dbx1del mice. Primary cultures from control IC tissues were found to contain many cells that co-expressed Tcf7l2 and Nrgn immunoreactivity, indicating normal IC cell identity (Fig. 7A). By contrast, consistent with our embryo experiments, the majority of primary cultured Dbx1-deficient IC cells were Tcf7l2−, Nrgn− (Fig. 7A). Compared to controls, Dbx1del IC cells that had been cultured for 5~7 days in vitro (DIV) showed apoptosis in significantly increased amounts (Fig. 7B). Notably, almost none of Cas3+ apoptotic cells expressed Tcf7l2 immunoreactivity (Fig. 7B). When IC cells were allowed to develop for up to 21 DIV, control IC cells were healthy, with round cell bodies and a complex network of neurites (Fig. 7A, C). However, In Dbx1-deficient cell cultures, their cell bodies were smaller and shrunken with fragmented neurites (Fig. 7A, C). Some of Dbx1del IC cells had begun detaching from the substrate and neurites were not visible. We estimated cell viability during a 28 DIV time course by using Calcein AM, a cell permeable dye, which is converted to fluorescent Calcein in live cells. The number of viable Dbx1del IC cells continued to decrease drastically and was <5% by 28 DIV, whereas >50% of control IC cells were viable (Fig. 7D). To address whether restoration of Tcf7l2 and Ap-2δ expression is able to prevent apoptosis and promote survival in Dbx1- deficient IC cells, Tcf7l2 and/or Ap-2δ expression constructs were transfected into Dbx1del mutant IC cells that had been cultured for 2 DIV together with a GFP expression vector to identify transfected cells. After 7 additional days of culture, GFP-positive cells were evaluated for apoptosis by assessing Cas3 immunoreactivity. The level of apoptosis was significantly decreased in primary cultures of Dbx1del IC cells transfected with Tcf7l2 and/or Ap-2δ expression constructs, compared with mock-transfected Dbx1del IC cells (Fig. 7E). To determine whether Dbx1-deficient IC cells remained viable following transfection, we assessed cell survival using Calcein-AM during a 21 DIV time course. The viability of mock-transfected Dbx1del IC cells continued to decrease drastically and was <5% by 21 DIV, whereas >30% of Dbx1del IC cells transfected with Tcf7l2 and/or Ap-2δ expression constructs were viable (Fig. 7F), indicating a significant rescue of IC apoptosis by Dbx1 deficiency. Taken together, these results suggest that Tcf7l2 and Ap-2δ transcription factors are important mediators of Dbx1 function for IC survival.
Fig. 7. Restored expression of Tcf7l2 and/or Ap-2δ attenuates apoptosis in Dbx1-deficient IC cells.

A Isolation and primary cultures of IC cells from E15.5 control and Dbx1del mutant embryonic brains. IC cells were cultured on coverslips coated with L-ornithine and laminin in 24-well plates. IC cell identity was verified by immunocytochemistry of 5 days in vitro (DIV5) cells using antibodies against Nrgn and Tcf7l2. Bright-field (BF) images display cellular processes including neurite outgrowth. Consistent with gene expression profiles in embryos, the majority of primary cultured Dbx1-deficient IC cells are Tcf7l2−, Nrgn−. B Caspase 3 (Cas3) immunostaining were performed at DIV7 cells to identify quantity of apoptosis. Co-immunostaining revealed that most of Cas3+ cells are Tcf7l2− (n = 5 biological replicates; Student’s t-test; *p ≤ 0.001). C Primary IC cells were established from E15.5 embryos and kept in culture for 21 days. Compared to control IC cells with round cell bodies and a complex network of neurites, most of Dbx1del mutant cell bodies were smaller, shrunken, and the neurites were fragmented and lost. D Calcein AM staining of viable cells. The number of viable cells was determined by Calcein fluorescence at DIV14, DIV21, and DIV28. Six counting sites were evaluated in one coverslip (n = 5 biological replicates). *P < 0.01; **P < 0.001 (Student’s t-test). E, F IC cells isolated from Dbx1del mutant E15.5 brains were cultured for 2 DIV and transfected with Tcf7l2 (p043mTcf-4B), Ap-2δ (pCMV-Tfap2d) expression constructs or both DNAs. Nuclear GFP expression construct (CAG-NLS-GFP) was co-transfected to identify the transfected cells. E Quantification of apoptosis was achieved from IC cells cultured for 7 additional DIV by counting the proportion of Cas3+ cells among GFP+ cells. Approximately 3000 GFP+ cells were considered for each experiments. Histograms represent the means normalized to 1 in the proportion of mock control (empty vector). Experiments were performed in 4 biological replicates. Error bars correspond to standard deviation. Means were compared using Student’s t-test; *p ≤ 0.01). F Quantification of survival was achieved from IC cells cultured for 7, 14 and 21 additional DIV following transfection. The number of viable cells was determined by Calcein fluorescence for 6 counting sites in each coverslip (n = 4 biological replicates). (Student’s t-test; *p ≤ 0.05; **p ≤ 0.01).
Discussion
Previous studies defined core (central nucleus) and shell (dorsal and lateral nucleus) regions of the IC in adult brain mainly by cell size, dendrite morphology, and axonal trajectories according to anterograde or retrograde tracing [2, 73–76]. IC neuronal populations and distributions during embryogenesis remains poorly understood, as assigning subdivisions is not straightforward because of a lack of gene expression profiling. To the best of our knowledge, this is the first study to systematically capture the gene expression profile of the developing IC at the genomic level, and identify an exhaustive list of molecular markers that distinguish the IC from the SC and other brain regions. Some of the IC-enriched genes revealed by our datasets served as molecular markers that could selectively identify different neuronal types in the IC. Most IC cells are Tcf7l2+ and Meis2+, and these neuron populations can be further subdivided into several distinct clusters according to differential distributions of other IC-specific genes including Nrgn, Foxp2, Maf, and Ndst4 (Fig. S1), indicating diverse populations of embryonic IC neurons. Further mapping of the co-expression of more genes in those neuronal clusters at postnatal stages will be required to determine the precise contribution of individual embryonic neuronal subtypes to adult IC organization and connectivity.
Our study shows that Dbx1 is required for the specification of the progenitor pool in the posterior tectum that generates the Tcf7l2+ Meis+ neuronal population in the IC via positive and negative regulation of target genes. It is not surprising that the majority (~80%) of the DEGs are upregulated in Dbx1 mutants as Dbx1 functions as a transcriptional repressor that contains the eh1 domain to interact with Gro/TLE corepressor. The downregulated DEGs including Tcf7l2 and Meis2, are likely to be indirectly regulated by Dbx1, and other transcriptional repressors released by loss of Dbx1 may mediate the negative regulation. A question that arises is how this single transcription factor can regulate distinct gene programs along the AP axis of the developing tectum. Our study uncovered striking similarities in IC phenotypes between Dbx1 and Tcf7l2 mutants. Interestingly, Dbx1 is also required for Tcf7l2 expression in the SC, but unlike the loss of Dbx1, loss of Tcf7l2 did not affect SC differentiation and morphogenesis, indicating that Dbx1 depends on Tcf7l2 to promote IC development, and requires other factor(s) for SC differentiation.
Our study uncovered 342 genes that are co-regulated by Dbx1 and Tcf7l2, many of which belong to the list of genes with previously uncharacterized expression and role in the IC, suggesting them as strong candidates for follow-up studies to investigate their functions in IC morphogenesis. Hesse et al. reported the identification of 12 genes, whose expression in the IC is regulated by Ap-2δ [57]. Interestingly, five of these Ap-2δ targets; Mef2c, Rgs4, Myh8, Ndst4, and Hjurp; are also co-regulated by Dbx1 and Tcf7l2. Mef2c is expressed in multiple brain regions including the cerebral cortex, midbrain, and cerebellum, and is crucial for neuronal differentiation, synaptic activity, learning, and memory [77]. However, the functional roles of these target genes in the midbrain remain unexplored. It should be noted that this Dbx1-regulated transcriptional program is not likely to be mediated entirely by the action of the Tcf7l2-Ap-2δ cascade. For example, Meis2, which marks most IC cells along with Tcf7l2, is regulated by Dbx1, but not by Tcf7l2. Previous gain-of-function experiments performed in chicken embryos demonstrated the ability of Meis2 to cause a diencephalic-to-tectal fate change without inducing the isthmic organizer [24]. The question of whether a parallel action by other transcription factors participates in the regulation of IC development by Dbx1 needs further investigation.
A remarkable feature common to Dbx1, Tcf7l2 and Ap-2δ mutants is abnormal cell death in the posterior tectum that commences at a late developmental phase and leads postnatally to specific loss of the IC. Considering the known roles and temporal expression window of Dbx1, we anticipated that Dbx1 function in the midbrain may be restricted to early embryonic patterning events. However, we discovered a novel function of Dbx1 as a survival factor that can transcriptionally suppress the pro-apoptotic factors c-Jun, Bim and Hrk. The apoptotic functions of c-Jun and BH3-only proteins are limited by multiple mechanisms (transcriptional, post-transcriptional, and post-translational), and stimuli by death signals release these controls [78]. Apoptotic stimuli induce transcriptional upregulation of c-Jun, Bim and Hrk, causing neuronal cell death [65, 68, 79]. We show that like the loss of Dbx1, the loss of Tcf7l2 also results in ectopic induction of c-Jun, Bim and Hrk. During embryonic development, Wnt signaling facilitates or inhibits apoptosis depending on the cellular context [48, 80]. Loss of Wnt1 leads to the absence of midbrain and cerebellum due to increased apoptotic cell death at E9.5 [10, 18], and β-catenin-deficient mice exhibit pronounced apoptosis at E7.5 [81]. Unlike loss of Wnt1 or β-catenin, loss of Tcf7l2 resulted in abnormal cell death in the IC at ~E18.5. At early stages, all four Tcf/Lef factors are expressed in the midbrain, but at late embryonic and postnatal stages, only Tcf7l2 expression is strongly present in the IC (Allen Brain Atlas). Therefore, it is likely that other Tcf/Lef factors may compensate for the loss of Tcf7l2 in the early stages, but not at later stages, when Tcf7l2 is the only gene expressed in the IC. Like in Dbx1 and Tcf7l2 mutants, at the early stages, apoptosis does not occur in Ap-2δ mutants, although Ap-2δ is already expressed [57]. Interestingly, Ap-2 genes are expressed in the developing midbrain in a pattern similar to that of Tcf/Lef factors. Five AP-2 genes are expressed in the developing midbrain, but only Ap-2δ expression continues in the IC during the late embryonic and adult stages [57] (Allen Brain Atlas), suggesting that functional redundancy may exist or that early midbrain development may not be dependent on Ap-2δ [57, 74, 75]. As Tcf7l2 and Ap-2δ are positively regulated by Dbx1, we assessed the ability of Tcf7l2 and Ap-2δ to rescue apoptosis induced by Dbx1 deficiency. We found that these two factors were able to decrease apoptosis and increase survival of Dbx1-deficient IC cells. These observations suggest that Tcf7l2 and Ap-2δ are required for Dbx1-mediated IC survival. However, individual or combined expression of these factors showed incomplete rescue, suggesting that the ability of Dbx1 to promote IC survival is not likely to be mediated entirely by the anti-apoptotic function of Tcf7l2 and Ap-2δ . Among the IC-enriched genes regulated by Dbx1, but not by Tcf7l2, Meis2 has been shown to prevent neuronal apoptosis [82]. Yang et al. generated Meis2 conditional mutant mice by using Dlx5/6-Cre to inactivate Meis2 in the ventral telencephalon. Loss of Meis2 resulted in significantly increased apoptotic cell death in the developing striatum, and altered transcription factor networks involved in regulating neuronal apoptosis [82]. Whether Meis2 participates in the regulation of IC survival requires further investigation.
Materials and methods
Mouse lines
All animal procedures were carried out in accordance with the guidelines and protocols approved by the Kyung Hee University Institutional Animal Care and Use Committee. The generation of Dbx1loxp/loxp, Tcf7l2loxp/loxp, ROSA-Tomato and EIIa-Cre mice obtained from The Jackson Laboratory, Bar Harbor, ME. was described previously [44, 58, 70, 83]. All animals used for these experiments were maintained on mixed backgrounds.
Production of transgenic mice
The Sre sequence was amplified by PCR with the primers (5′-ATTAGCGGCCGCAT TGCCATGGCAACGACTGCTC-3′; 5′-ATTAGCGGCCGCGGTCCATTTTGAGGTA AATTACAGG), and cloned into the NotI site of an expression vector containing a β-globin minimal promoter, the Cre gene, and an SV40 poly(A) signal. The resulting plasmid transgene was prepared and linearized with SalI for microinjection as previously described [69]. Transient transgenic embryos or mouse lines were generated by pronuclear injection into fertilized eggs derived from the FVBN mouse strain.
Immunohistochemistry
For immunohistochemistry, embryos fixed in 4% formaldehyde for 2–3 h at 4 °C were immersed in 30% sucrose, and cryosectioned at 25 µm. The following primary antibodies were used: anti-Caspase3 (Cell Signaling, 9661; 1:1000), anti-phospho-Histone H3 (Upstate Biotechnology, 06–570; 1:1000), anti-GFP (Abcam ab6673, 1:500), anti-Tcf7l2 (Millipore, 05–511, 1:200), anti-Tdtomato (SICGEN, AB8181–200, 1:500), and anti-Nrgn (Millipore, AB5620, 1:500). Following incubation with primary antibody, sections were incubated with species-specific secondary antibodies conjugated to Alexa Fluor (Molecular Probes, A11034, A11003, A11010, A11029, A11055, A11056; 1:200).
Whole-mount and section in situ hybridization
Whole-mount and section in situ hybridization were performed using digoxigenin-UTP-labeled riboprobes essentially as described previously [54]. The pairs of PCR primer sequences to generate template DNA for in vitro transcription by T7 RNA polymerase were obtained from the Allen Brain Atlas or designed against unique regions of transcripts to avoid cross-reactivity. At least three independent experiments were performed for any given riboprobes, which showed highly reproducible gene expression data.
Real-time RT-PCR
After IC tissues were harvested in TRIzol Reagent (Invitrogen), total RNA was isolated using an RNeasy Mini kit (Qiagen) and used to prepare cDNA using a SuperScriptIII first-strand synthesis system (Invitrogen). Real-time PCR was performed in duplicates for each RNA sample using QuantiTect SYBR Green Supermix (Qiagen) following the manufacturers’ instructions. Reactions were monitored using the Mx4000 multiplex quantitative PCR system (Stratagene, La Jolla, CA). All results were normalized to the housekeeping gene β-actin and relative quantification was calculated using comparative threshold cycle values for three biological replicates.
RNA-seq analysis
E18.5 IC regions were dissected, and RNA was extracted using the TRIzol Plus RNA Purification Kit (Invitrogen). Triplicates of nine IC tissues for each genotypes were run for RNA-seq (Table S1). After adaptor sequences and low-quality reads were filtered out, the clean reads from each sample were aligned to the mouse genome mm10 assembly from the University of California, Santa Cruz (UCSC), and tag counts were performed with Bowtie2 using HISAT2 (Table S1). After normalization of the read counts, transcript levels were calculated and presented by FPKM and TPM. Genes with log2(FC) ≥ 0.4 and log2(FC) ≤ −0.4 and FDR adjusted P value (q-value) ≤ 0.05 were considered to be differentially expressed. After statistical tests, we performed functional annotation and GO term assignment with the g:Profiler tool (https://biit.cs.ut.ee/gprofiler/), Panther (http://www.pantherdb.org) and KEGG database (http://www.genome.jp/kegg/). GO term enrichments were tested with Fisher’s exact test, and FDR-adjusted P values (q-value) ≤ 0.01 were considered significant.
Primary cell culture and transfection
Brain tissues were isolated from individual embryos at E15.5 and dissected in 3 cm sterile Petridish containing ice cold dissection medium, consisting of Hanks’ balanced salt solution (HBSS) (Gibco), 20 mM D-glucose (Sigma), 100 units/ml Penicillin/Streptomycin (Gibco), and 200 μM ascorbic acid (Sigma). IC tissues were collected in a 1.5 ml sterile tube and digested with StemPro Accutase Cell Dissociation reagent (Thermo Fisher Scientific) for 10–20 min at 37 °C. The digestion was stopped by removing the supernatant and washing the tissue with dissociation medium, consisting of Neurobasal medium (Invitrogen), 200 μM L-glutamine (Thermo Fisher Scientific), 2% B-27 supplement (Thermo Fisher Scientific), 2% B-27 supplement Plus (Thermo Fisher Scientific), 1% FBS (Gibco), and 200 μM ascorbic acid. The tissue was resuspended in 1 ml dissociation medium and triturated to achieve single cell suspension. After cells were spun at 200 × g, the cell pellet was resuspended in the dissociation medium. For immunofluorescence microscopy, the cell suspension was transferred on top of 12 mm coverslips coated with 20 μg/ml poly-L-ornithine (Sigma) and 10 μg/ml laminin (Thermo Fisher Scientific) (100,000 cells per coverslip). After 1 h incubation in a humidified tissue culture incubator at 37 °C at 5% CO2, the coverslips were carefully transferred into 24-well plates containing the dissociation medium. Transient transfection was performed at 3 days in vitro. Transfected plasmids (0.3 μg) consisted of a mixture of 0.1 μg CAG-NLS-GFP [84] (a gift from Viviana Gradinaru; Addgene plasmid # 104061) + 0.2 μg p043-mTcf4B [85] (a gift from Ramesh Shivdasani; Addgene plasmid # 11031); a mixture of 0.1 μg CAG-NLS-GFP + 0.2 μg pCMV6-Tfap2d (Origene); a mixture of 0.1 μg CAG-NLS-GFP + 0.1 μg p043-mTcf4 + 0.1 μg pCMV6-Tfap2d. Mock transfection consisted of 0.1 μg CAG-NLS-GFP + 0.2 μg empty vector. For individual wells in 24-well plates, a total of 0.3 μg DNA was diluted in 25 μl Opti-MEM Reduced Serum Medium (Gibco), and 2.5 μl Lipofectamine 2000 (Invitrogen) was diluted in 25 μl Opti-MEM in a separate tube. The diluted DNA-lipofectamine mixture was incubated at RT for 20 min and applied to cells.
Cell viability
The viability of cells in culture was assessed by measuring the activity of intracellular esterase using a Live Cell Imaging kit (Invitrogen). The non-fluorescent acetomethoxy derivative of calcein (calcein AM) is a cell permeable dye that can be transported into live cells. In living cells, calcein AM is converted to fluorescent calcein after hydrolysis by esterase. As dead cells lack active esterase, only live cells are labeled. IC-derived cells were plated on poly-L-ornithine and laminin coated coverslips in 24-well plates and allowed to culture for 1–4 weeks. After Calcein AM was added to the cells and incubated for 15 min, six randomly chosen fields from each coverslip were photographed, and the number of surviving cells was computed.
Statistical analysis
All mutant phenotypes we report in Dbx1del, Dbx1cko, Tcf7l2del, and Tcf7l2cko mice were completely penetrant in both embryos and postnatal animals. The number of independent values in each experiment was as follows: Fig. 1, three independent biological replicates for RNA-seq. Each replicate was prepared from nine IC tissues of E18.5 control and Dbx1del. n = 3 for E18.5 ISH; Fig. 2, n = 3 for real-time RT-PCR, n = 3 for E18.5 ISH, n = 5 for E12.5-E18.5 Caspase3 staining; Fig. 3, n = 3 for P3 ISH, n = 5 for P3 Caspase3 staining, n = 5 for P20 ISH; Fig. 4, three independent biological replicates for RNA-seq. Each replicate was prepared from nine IC tissues of E18.5 control and Tcf7l2del. n = 3 for E18.5 ISH; Fig. 5, n = 3 for real-time RT-PCR, n = 3 for E18.5 ISH, n = 5 for Caspase3 and PH3 immunohistochemistry; n = 5 for P20 ISH; Fig. 6, n = 3 for real-time RT-PCR, n = 3 for ISH; Fig. 7, n = 5 for Caspase3 and calcein AM staining in control and Dbx1del IC cells, n = 4 for Caspase3 and calcein AM staining in transfected Dbx1del IC cells. Relevant information for each experiment including statistical tests and p values are included in the legend corresponding to each figure. In all cases, p ≤ 0.05 is considered statistically significant and error bars represent s.e.m.
Supplementary information
{kind=link}
{kind=link}
{kind=link}
Acknowledgements
This work was supported by the National Research Foundation of Korea (NRF) grants funded by the Korea Ministry of Science and ICT (No. 2019R1F1A1042941 and No. 2022R1F1A1064555).
Author contributions
Conceived project: YJ. Designed experiments: HT, YJ. Performed experiments: HT, QN, JJ, DL. Analyzed data: HT, QN, JJ, DL, YJ. Provided reagents: YN, TK, JY, KB, Wrote paper: YJ. Reviewed and revised the paper: HT, QN, JJ, DL, YJ.
Data availability
RNA-seq raw FASTQ files of wildtype and Dbx1 mutants have been deposited into the NCBI Sequence Read Archive under bioproject ID PRJNA793387. RNA-seq raw FASTQ files of wildtype and Tcf7l2 mutants have been deposited into the NCBI Sequence Read Archive under bioproject ID PRJNA801436.
Competing interests
The authors declare no competing interests.
Ethics approval
All animal experiments were conducted in accordance with the guidelines of the Kyung Hee University Institutional Animal Care and Use Committee (KHGASP-20–015).
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
The online version contains supplementary material available at 10.1038/s41418-023-01165-6.
References
- 1.Winer JA, Schreiner CE. The Inferior Colliculus. New York, NY: Springer New York; 2005. 10.1007/b138578.
- 2.Oliver DL, Morest DK. The central nucleus of the inferior colliculus in the cat. J Comp Neurol. 1984;222:237–64. doi: 10.1002/cne.902220207. [DOI] [PubMed] [Google Scholar]
- 3.Winer JA, Larue DT, Diehl JJ, Hefti BJ. Auditory cortical projections to the cat inferior colliculus. J Comp Neurol. 1998;400:147–74. doi: 10.1002/(SICI)1096-9861(19981019)400:2<147::AID-CNE1>3.0.CO;2-9. [DOI] [PubMed] [Google Scholar]
- 4.Peruzzi D, Bartlett E, Smith PH, Oliver DL. A Monosynaptic GABAergic Input from the Inferior Colliculus to the Medial Geniculate Body in Rat. J Neurosci. 1997;17:3766–77. doi: 10.1523/JNEUROSCI.17-10-03766.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Winer JA, Saint Marie RL, Larue DT, Oliver DL. GABAergic feedforward projections from the inferior colliculus to the medial geniculate body. Proc Natl Acad Sci. 1996;93:8005–10. doi: 10.1073/pnas.93.15.8005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Nakatani T, Minaki Y, Kumai M, Ono Y. Helt determines GABAergic over glutamatergic neuronal fate by repressing Ngn genes in the developing mesencephalon. Development. 2007;134:2783–93. doi: 10.1242/dev.02870. [DOI] [PubMed] [Google Scholar]
- 7.Kala K, Haugas M, Lilleväli K, Guimera J, Wurst W, Salminen M, et al. Gata2 is a tissue-specific post-mitotic selector gene for midbrain GABAergic neurons. Development. 2009;136:253–62. doi: 10.1242/dev.029900. [DOI] [PubMed] [Google Scholar]
- 8.Joyner AL, Liu A, Millet S. Otx2, Gbx2 and Fgf8 interact to position and maintain a mid-hindbrain organizer. Curr Opin Cell Biol. 2000;12:736–41. doi: 10.1016/S0955-0674(00)00161-7. [DOI] [PubMed] [Google Scholar]
- 9.Wurst W, Bally-Cuif L. Neural plate patterning: upstream and downstream of the isthmic organizer. Nat Rev Neurosci. 2001;2:99–108. doi: 10.1038/35053516. [DOI] [PubMed] [Google Scholar]
- 10.Chi CL, Martinez S, Wurst W, Martin GR. The isthmic organizer signal FGF8 is required for cell survival in the prospective midbrain and cerebellum. Development. 2003;130:2633–44. doi: 10.1242/dev.00487. [DOI] [PubMed] [Google Scholar]
- 11.Shamim H, Mahmood R, Logan C, Doherty P, Lumsden A, Mason I. Sequential roles for Fgf4, En1 and Fgf8 in specification and regionalisation of the midbrain. Development. 1999;126:945–59. doi: 10.1242/dev.126.5.945. [DOI] [PubMed] [Google Scholar]
- 12.Basson MA, Echevarria D, Ahn CP, Sudarov A, Joyner AL, Mason IJ, et al. Specific regions within the embryonic midbrain and cerebellum require different levels of FGF signaling during development. Development. 2008;135:889–98. doi: 10.1242/dev.011569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Dee A, Li K, Heng X, Guo Q, Li JYH. Regulation of self-renewing neural progenitors by FGF/ERK signaling controls formation of the inferior colliculus. Development. 2016;143:3661–73. doi: 10.1242/dev.138537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sgaier SK, Lao Z, Villanueva MP, Berenshteyn F, Stephen D, Turnbull RK, et al. Genetic subdivision of the tectum and cerebellum into functionally related regions based on differential sensitivity to engrailed proteins. Development. 2007;134:2325–35. doi: 10.1242/dev.000620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Trokovic R. FGFR1 is independently required in both developing mid- and hindbrain for sustained response to isthmic signals. EMBO J. 2003;22:1811–23. doi: 10.1093/emboj/cdg169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Xu J, Liu Z, Ornitz DM. Temporal and spatial gradients of Fgf8 and Fgf17 regulate proliferation and differentiation of midline cerebellar structures. Development. 2000;127:1833–43. doi: 10.1242/dev.127.9.1833. [DOI] [PubMed] [Google Scholar]
- 17.Sato T, Joyner AL. The duration of Fgf8 isthmic organizer expression is key to patterning different tectal-isthmo-cerebellum structures. Development. 2009;136:3617–26. doi: 10.1242/dev.041210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.McMahon AP, Bradley A. The Wnt-1 (int-1) proto-oncogene is required for development of a large region of the mouse brain. Cell. 1990;62:1073–85. doi: 10.1016/0092-8674(90)90385-R. [DOI] [PubMed] [Google Scholar]
- 19.Thomas KR, Capecchi MR. Targeted disruption of the murine int-1 proto-oncogene resulting in severe abnormalities in midbrain and cerebellar development. Nature. 1990;346:847–50. doi: 10.1038/346847a0. [DOI] [PubMed] [Google Scholar]
- 20.Wurst W, Auerbach AB, Joyner AL. Multiple developmental defects in Engrailed-1 mutant mice: an early mid-hindbrain deletion and patterning defects in forelimbs and sternum. Development. 1994;120:2065–75. doi: 10.1242/dev.120.7.2065. [DOI] [PubMed] [Google Scholar]
- 21.Serbedzija GN, Dickinson M, McMahon AP. Cell death in the CNS of the Wnt-1 mutant mouse. J Neurobiol. 1996;31:275–82. doi: 10.1002/(SICI)1097-4695(199611)31:3<275::AID-NEU1>3.0.CO;2-7. [DOI] [PubMed] [Google Scholar]
- 22.Urbánek P, Wang ZQ, Fetka I, Wagner EF, Busslinger M. Complete block of early B cell differentiation and altered patterning of the posterior midbrain in mice lacking Pax5/BSAP. Cell. 1994;79:901–12. doi: 10.1016/0092-8674(94)90079-5. [DOI] [PubMed] [Google Scholar]
- 23.Matsunaga E, Araki I, Nakamura H. Role of Pax3/7 in the tectum regionalization. Development. 2001;128:4069–77. doi: 10.1242/dev.128.20.4069. [DOI] [PubMed] [Google Scholar]
- 24.Agoston Z, Schulte D. Meis2 competes with the Groucho co-repressor Tle4 for binding to Otx2 and specifies tectal fate without induction of a secondary midbrain-hindbrain boundary organizer. Development. 2009;136:3311–22. doi: 10.1242/dev.037770. [DOI] [PubMed] [Google Scholar]
- 25.Broccoli V, Boncinelli E, Wurst W. The caudal limit of Otx2 expression positions the isthmic organizer. Nature. 1999;401:164–8. doi: 10.1038/43670. [DOI] [PubMed] [Google Scholar]
- 26.Millet S, Campbell K, Epstein DJ, Losos K, Harris E, Joyner AL. A role for Gbx2 in repression of Otx2 and positioning the mid/hindbrain organizer. Nature. 1999;401:161–4. doi: 10.1038/43664. [DOI] [PubMed] [Google Scholar]
- 27.Puelles E, Acampora D, Lacroix E, Signore M, Annino A, Tuorto F, et al. Otx dose-dependent integrated control of antero-posterior and dorso-ventral patterning of midbrain. Nat Neurosci. 2003;6:453–60. doi: 10.1038/nn1037. [DOI] [PubMed] [Google Scholar]
- 28.Di Giovannantonio LG, Di Salvio M, Omodei D, Prakash N, Wurst W, Pierani A, et al. Otx2 cell-autonomously determines dorsal mesencephalon versus cerebellum fate independently of isthmic organizing activity. Development. 2014;141:377–88. doi: 10.1242/dev.102954. [DOI] [PubMed] [Google Scholar]
- 29.Cowan WM, Martin AH, Wenger E. Mitotic patterns in the optic tectum of the chick during normal development and after early removal of the optic vesicle. J Exp Zool. 1968;169:71–92. doi: 10.1002/jez.1401690110. [DOI] [PubMed] [Google Scholar]
- 30.Altman J, Bayer SA. Time of origin of neurons of the rat inferior colliculus and the relations between cytogenesis and tonotopic order in the auditory pathway. Exp Brain Res. 1981;42–42:411–23. doi: 10.1007/BF00237506. [DOI] [PubMed] [Google Scholar]
- 31.Altman J, Bayer SA. Time of origin of neurons of the rat superior colliculus in relation to other components of the visual and visuomotor pathways. Exp Brain Res. 1981;42:424–34. doi: 10.1007/BF00237507. [DOI] [PubMed] [Google Scholar]
- 32.Edwards MA, Caviness VS, Schneider GE. Development of cell and fiber lamination in the mouse superior colliculus. J Comp Neurol. 1986;248:395–409. doi: 10.1002/cne.902480308. [DOI] [PubMed] [Google Scholar]
- 33.Miyoshi G, Bessho Y, Yamada S, Kageyama R. Identification of a novel basic helix-loop-helix gene, Heslike, and its role in GABAergic neurogenesis. J Neurosci. 2004;24:3672–82. doi: 10.1523/JNEUROSCI.5327-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Guimera J, Weisenhorn DV, Wurst W. Megane/Heslike is required for normal GABAergic differentiation in the mouse superior colliculus. Development. 2006;133:3847–57. doi: 10.1242/dev.02557. [DOI] [PubMed] [Google Scholar]
- 35.Peltopuro P, Kala K, Partanen J. Distinct requirements for Ascl1 in subpopulations of midbrain GABAergic neurons. Dev Biol. 2010;343:63–70. doi: 10.1016/j.ydbio.2010.04.015. [DOI] [PubMed] [Google Scholar]
- 36.Achim K, Peltopuro P, Lahti L, Tsai HH, Zachariah A, Astrand M, et al. The role of Tal2 and Tal1 in the differentiation of midbrain GABAergic neuron precursors. Biol Open. 2013;2:990–7. doi: 10.1242/bio.20135041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wende C-Z, Zoubaa S, Blak A, Echevarria D, Martinez S, Guillemot F, et al. Hairy/Enhancer-of-Split MEGANE and Proneural MASH1 Factors Cooperate Synergistically in Midbrain GABAergic Neurogenesis. PLoS ONE. 2015;10:e0127681. doi: 10.1371/journal.pone.0127681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Shoji H, Ito T, Wakamatsu Y, Hayasaka N, Ohsaki K, Oyanagi M, et al. Regionalized expression of the Dbx family homeobox genes in the embryonic CNS of the mouse. Mech Dev. 1996;56:25–39. doi: 10.1016/0925-4773(96)00509-6. [DOI] [PubMed] [Google Scholar]
- 39.Pierani A, Moran-Rivard L, Sunshine MJ, Littman DR, Goulding M, Jessell TM. Control of interneuron fate in the developing spinal cord by the progenitor homeodomain protein Dbx1. Neuron. 2001;29:367–84. doi: 10.1016/S0896-6273(01)00212-4. [DOI] [PubMed] [Google Scholar]
- 40.Lanuza GM, Gosgnach S, Pierani A, Jessell TM, Goulding M. Genetic identification of spinal interneurons that coordinate left-right locomotor activity necessary for walking movements. Neuron. 2004;42:375–86. doi: 10.1016/S0896-6273(04)00249-1. [DOI] [PubMed] [Google Scholar]
- 41.Talpalar AE, Bouvier J, Borgius L, Fortin G, Pierani A, Kiehn O. Dual-mode operation of neuronal networks involved in left-right alternation. Nature. 2013;500:85–8. doi: 10.1038/nature12286. [DOI] [PubMed] [Google Scholar]
- 42.Bouvier J, Thoby-Brisson M, Renier N, Dubreuil V, Ericson J, Champagnat J, et al. Hindbrain interneurons and axon guidance signaling critical for breathing. Nat Neurosci. 2010;13:1066–74. doi: 10.1038/nn.2622. [DOI] [PubMed] [Google Scholar]
- 43.Gray PA, Hayes JA, Ling GY, Llona I, Tupal S, Picardo MC, et al. Developmental origin of preBötzinger complex respiratory neurons. J Neurosci. 2010;30:14883–95. doi: 10.1523/JNEUROSCI.4031-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Sokolowski K, Esumi S, Hirata T, Kamal Y, Tran T, Lam A, et al. Specification of select hypothalamic circuits and innate behaviors by the embryonic patterning gene dbx1. Neuron. 2015;86:403–16. doi: 10.1016/j.neuron.2015.03.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Inamata Y, Shirasaki R. Dbx1 triggers crucial molecular programs required for midline crossing by midbrain commissural axons. Development. 2014;141:1260–71. doi: 10.1242/dev.102327. [DOI] [PubMed] [Google Scholar]
- 46.Grant SF, Thorleifsson G, Reynisdottir I, Benediktsson R, Manolescu A, Sainz J, et al. Variant of transcription factor 7-like 2 (TCF7L2) gene confers risk of type 2 diabetes. Nat Genet. 2006;38:320–3. doi: 10.1038/ng1732. [DOI] [PubMed] [Google Scholar]
- 47.Korinek V, Barker N, Moerer P, van Donselaar E, Huls G, Peters PJ, et al. Depletion of epithelial stem-cell compartments in the small intestine of mice lacking Tcf-4. Nat Genet. 1998;19:379–83. doi: 10.1038/1270. [DOI] [PubMed] [Google Scholar]
- 48.Li F, Chong ZZ, Maiese K. Winding through the WNT pathway during cellular development and demise. Histol Histopathol. 2006;21:103–24. doi: 10.14670/hh-21.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Ah Cho E, Dressler GR. TCF-4 binds β-catenin and is expressed in distinct regions of the embryonic brain and limbs. Mech Dev. 1998;77:9–18. doi: 10.1016/S0925-4773(98)00131-2. [DOI] [PubMed] [Google Scholar]
- 50.Brinkmeier ML, Potok MA, Davis SW, Camper SA. TCF4 deficiency expands ventral diencephalon signaling and increases induction of pituitary progenitors. Dev Biol. 2007;311:396–407. doi: 10.1016/j.ydbio.2007.08.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Ye F, Chen Y, Hoang T, Montgomery RL, Zhao XH, Bu H, et al. HDAC1 and HDAC2 regulate oligodendrocyte differentiation by disrupting the beta-catenin-TCF interaction. Nat Neurosci. 2009;12:829–38. doi: 10.1038/nn.2333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Hammond E, Lang J, Maeda Y, Pleasure D, Angus-Hill M, Xu J, et al. The Wnt effector transcription factor 7-like 2 positively regulates oligodendrocyte differentiation in a manner independent of Wnt/β-catenin signaling. J Neurosci. 2015;35:5007–22. doi: 10.1523/JNEUROSCI.4787-14.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Zhao C, Deng Y, Liu L, Yu K, Zhang L, Wang H, et al. Dual regulatory switch through interactions of Tcf7l2/Tcf4 with stage-specific partners propels oligodendroglial maturation. Nat Commun. 2016;7:10883. doi: 10.1038/ncomms10883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Lee M, Yoon J, Song H, Lee B, Lam DT, Yoon J, et al. Tcf7l2 plays crucial roles in forebrain development through regulation of thalamic and habenular neuron identity and connectivity. Dev Biol. 2017;424:62–76. doi: 10.1016/j.ydbio.2017.02.010. [DOI] [PubMed] [Google Scholar]
- 55.Tran H-N, Park W, Seong S, Jeong JE, Nguyen QH, Yoon J, et al. Tcf7l2 transcription factor is required for the maintenance, but not the initial specification, of the neurotransmitter identity in the caudal thalamus. Dev Dyn. 2020;249:646–55. doi: 10.1002/dvdy.146. [DOI] [PubMed] [Google Scholar]
- 56.Lipiec MA, Bem J, Koziński K, Chakraborty C, Urban-Ciećko J, Zajkowski T, et al. TCF7L2 regulates postmitotic differentiation programmes and excitability patterns in the thalamus. Development. 2020;147:dev190181. doi: 10.1242/dev.190181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Hesse K, Vaupel K, Kurt S, Buettner R, Kirfel J, Moser M. AP-2δ is a crucial transcriptional regulator of the posterior midbrain. PLoS ONE. 2011;6:e23483. doi: 10.1371/journal.pone.0023483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Lakso M, Pichel JG, Gorman JR, Sauer B, Okamoto Y, Lee E, et al. Efficient in vivo manipulation of mouse genomic sequences at the zygote stage. Proc Natl Acad Sci USA. 1996;93:5860–5. doi: 10.1073/pnas.93.12.5860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Matsunaga Y, Noda M, Murakawa H, Hayashi K, Nagasaka A, Inoue S, et al. Reelin transiently promotes N-cadherin-dependent neuronal adhesion during mouse cortical development. Proc Natl Acad Sci USA. 2017;114:2048–53. doi: 10.1073/pnas.1615215114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Chao CC-K, Chang P-Y, Lu HH-P. Human Gas7 isoforms homologous to mouse transcripts differentially induce neurite outgrowth. J Neurosci Res. 2005;81:153–62. doi: 10.1002/jnr.20552. [DOI] [PubMed] [Google Scholar]
- 61.Garrido-García A, de Andrés R, Jiménez-Pompa A, Soriano P, Sanz-Fuentes D, Martínez-Blanco E, et al. Neurogranin Expression Is Regulated by Synaptic Activity and Promotes Synaptogenesis in Cultured Hippocampal Neurons. Mol Neurobiol. 2019;56:7321–37. doi: 10.1007/s12035-019-1593-3. [DOI] [PubMed] [Google Scholar]
- 62.Panza P, Sitko AA, Maischein H-M, Koch I, Flötenmeyer M, Wright GJ, et al. The LRR receptor Islr2 is required for retinal axon routing at the vertebrate optic chiasm. Neural Dev. 2015;10:23. doi: 10.1186/s13064-015-0050-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Xiao T, Baier H. Lamina-specific axonal projections in the zebrafish tectum require the type IV collagen Dragnet. Nat Neurosci. 2007;10:1529–37. doi: 10.1038/nn2002. [DOI] [PubMed] [Google Scholar]
- 64.Cooper SR, Emond MR, Duy PQ, Liebau BG, Wolman MA, Jontes JD. Protocadherins control the modular assembly of neuronal columns in the zebrafish optic tectum. J Cell Biol. 2015;211:807–14. doi: 10.1083/jcb.201507108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Putcha GV, Moulder KL, Golden JP, Bouillet P, Adams JA, Strasser A, et al. Induction of BIM, a proapoptotic BH3-only BCL-2 family member, is critical for neuronal apoptosis. Neuron. 2001;29:615–28. doi: 10.1016/S0896-6273(01)00238-0. [DOI] [PubMed] [Google Scholar]
- 66.Coultas L, Terzano S, Thomas T, Voss A, Reid K, Stanley EG, et al. Hrk/DP5 contributes to the apoptosis of select neuronal populations but is dispensable for haematopoietic cell apoptosis. J Cell Sci. 2007;120:2044–52. doi: 10.1242/jcs.002063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Ma C, Ying C, Yuan Z, Song B, Li D, Liu Y, et al. dp5/HRK is a c-Jun target gene and required for apoptosis induced by potassium deprivation in cerebellar granule neurons. J Biol Chem. 2007;282:30901–9. doi: 10.1074/jbc.M608694200. [DOI] [PubMed] [Google Scholar]
- 68.Whitfield J, Neame SJ, Paquet L, Bernard O, Ham J. Dominant-Negative c-Jun Promotes Neuronal Survival by Reducing BIM Expression and Inhibiting Mitochondrial Cytochrome c Release. Neuron. 2001;29:629–43. doi: 10.1016/S0896-6273(01)00239-2. [DOI] [PubMed] [Google Scholar]
- 69.Lee B, Song H, Rizzoti K, Son Y, Yoon J, Baek K, et al. Genomic code for Sox2 binding uncovers its regulatory role in Six3 activation in the forebrain. Dev Biol. 2013;381:491–501. doi: 10.1016/j.ydbio.2013.06.016. [DOI] [PubMed] [Google Scholar]
- 70.Angus-Hill ML, Elbert KM, Hidalgo J, Capecchi MR. T-cell factor 4 functions as a tumor suppressor whose disruption modulates colon cell proliferation and tumorigenesis. Proc Natl Acad Sci. 2011;108:4914–9. doi: 10.1073/pnas.1102300108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Schorle H, Meier P, Buchert M, Jaenisch R, Mitchell PJ. Transcription factor AP-2 essential for cranial closure and craniofacial development. Nature. 1996;381:235–8. doi: 10.1038/381235a0. [DOI] [PubMed] [Google Scholar]
- 72.Moser M, Pscherer A, Roth C, Becker J, Mücher G, Zerres K, et al. Enhanced apoptotic cell death of renal epithelial cells in mice lacking transcription factor AP-2beta. Genes Dev. 1997;11:1938–48. doi: 10.1101/gad.11.15.1938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Malmierca MS, Blackstad TW, Osen KK, Karagülle T, Molowny RL. The central nucleus of the inferior colliculus in rat: A Golgi and computer reconstruction study of neuronal and laminar structure. J Comp Neurol. 1993;333:1–27. doi: 10.1002/cne.903330102. [DOI] [PubMed] [Google Scholar]
- 74.Meininger V, Pol D, Derer P. The inferior colliculus of the mouse. A Nissl and Golgi study. Neuroscience. 1986;17:1159–79. doi: 10.1016/0306-4522(86)90085-0. [DOI] [PubMed] [Google Scholar]
- 75.Chen C, Cheng M, Ito T, Song S. Neuronal Organization in the Inferior Colliculus Revisited with Cell-Type-Dependent Monosynaptic Tracing. J Neurosci. 2018;38:3318–32. doi: 10.1523/JNEUROSCI.2173-17.2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Goyer D, Silveira MA, George AP, Beebe NL, Edelbrock RM, Malinski PT, et al. A novel class of inferior colliculus principal neurons labeled in vasoactive intestinal peptide-Cre mice. Elife. 2019;8:e43770. doi: 10.7554/eLife.43770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Li H, Radford JC, Ragusa MJ, Shea KL, McKercher SR, Zaremba JD, et al. Transcription factor MEF2C influences neural stem/progenitor cell differentiation and maturation in vivo. Proc Natl Acad Sci USA. 2008;105:9397–402. doi: 10.1073/pnas.0802876105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Fricker M, Tolkovsky AM, Borutaite V, Coleman M, Brown GC. Neuronal Cell Death. Physiol Rev. 2018;98:813–80. doi: 10.1152/physrev.00011.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Imaizumi K. Critical Role for DP5/Harakiri, a Bcl-2 Homology Domain 3-Only Bcl-2 Family Member, in Axotomy-Induced Neuronal Cell Death. J Neurosci. 2004;24:3721–5. doi: 10.1523/JNEUROSCI.5101-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Ellies DL, Church V, Francis-West P, Lumsden A. The WNT antagonist cSFRP2 modulates programmed cell death in the developing hindbrain. Development. 2000;127:5285–95. doi: 10.1242/dev.127.24.5285. [DOI] [PubMed] [Google Scholar]
- 81.Huelsken J, Vogel R, Brinkmann V, Erdmann B, Birchmeier C, Birchmeier W. Requirement for beta-catenin in anterior-posterior axis formation in mice. J Cell Biol. 2000;148:567–78. doi: 10.1083/jcb.148.3.567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Yang L, Su Z, Wang Z, Li Z, Shang Z, Du H, et al. Transcriptional profiling reveals the transcription factor networks regulating the survival of striatal neurons. Cell Death Dis. 2021;12:262. doi: 10.1038/s41419-021-03552-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Madisen L, Zwingman TA, Sunkin SM, Oh SW, Zariwala HA, Gu H, et al. A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat Neurosci. 2010;13:133–40. doi: 10.1038/nn.2467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Challis RC, Ravindra Kumar S, Chan KY, Challis C, Beadle K, Jang MJ, et al. Systemic AAV vectors for widespread and targeted gene delivery in rodents. Nat Protoc. 2019;14:379–414. doi: 10.1038/s41596-018-0097-3. [DOI] [PubMed] [Google Scholar]
- 85.Lee YJ, Swencki B, Shoichet S, Shivdasani RA. A possible role for the high mobility group box transcription factor Tcf-4 in vertebrate gut epithelial cell differentiation. J Biol Chem. 1999;274:1566–72. doi: 10.1074/jbc.274.3.1566. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
RNA-seq raw FASTQ files of wildtype and Dbx1 mutants have been deposited into the NCBI Sequence Read Archive under bioproject ID PRJNA793387. RNA-seq raw FASTQ files of wildtype and Tcf7l2 mutants have been deposited into the NCBI Sequence Read Archive under bioproject ID PRJNA801436.