

Abstract
Saliva is well-described in oral food processing, but its role in taste responsiveness remains understudied. Taste stimuli must dissolve in saliva to reach their receptor targets. This allows the constituents of saliva the opportunity to interact with taste stimuli and their receptors at the most fundamental level. Yet, despite years of correlational data suggesting a role for salivary proteins in food preference, there were few experimental models to test the role of salivary proteins in taste-driven behaviors. Here we review our experimental contributions to the hypothesis that salivary proteins can alter taste function. We have developed a rodent model to test how diet alters salivary protein expression, and how salivary proteins alter diet acceptance and taste. We have found that salivary protein expression is modified by diet, and these diet-induced proteins can, in turn, increase the acceptance of a bitter diet. The change in acceptance is in part mediated by a change in taste signaling. Critically, we have documented increased detection threshold, decreased taste nerve signaling, and decreased oromotor responding to quinine when animals have increases in a subset of salivary proteins compared to control conditions.
1. Introduction
Choosing a food item is an extremely complex decision, with learned responses, conditioned and unconditioned preferences, metabolic state, food availability, and a host of social and biopsychological factors contributing to the decision. Humans currently live in a complex food environment with high availability of palatable, energy-dense, nutrient-poor foods. This modern shift in food availability is often highlighted as a contributor to the rise in a host of diet-related disease states. Dietary interventions to address the health crisis are plentiful [1,2], often focused on increasing vegetable intake [3–5] and very often unsuccessful [1,4,6]. The most commonly reported barrier to increasing vegetable intake is flavor [7,8]. There is, however, a very large, and well-documented, variation in vegetable preference, suggesting that consuming vegetables in the modern food environment is not a lost cause; and underlying variation in one of the decision drivers can be targeted to increase acceptance. One of the most effective interventions to increase vegetable intake is repeated exposure [9,10] which results in increased liking for the target vegetable [9].
Although the literature is equivocal about whether the variation in vegetable intake is linked to genetic variation in bitter perception [11–14], it is clear that vegetable consumers rate vegetables as more pleasant to consume than vegetable avoiders do [9]. Bitterness is the taste quality that most people associate with non-preferred vegetables [8]. The casual evolutionary explanation for bitter rejection is that this taste quality represents the potential presence of a toxin and therefore should be avoided. However, while this rule of thumb is often repeated, it is not well supported. First, bitterness is a poor predictor of toxicity [15] and animals do not avoid bitter stimuli but instead appear to use bitterness as a signal to exercise caution (i.e., “go-slow”) when sampling bitter stimuli [16].
Most bitter compounds in our evolutionary diet are plant secondary compounds (PSCs). Plants defend their photosynthesizing tissues from herbivory through chemical or physical defense (e.g., PSCs, and thorns or needles, respectively). Omnivores and herbivores rely on plant tissue for nutrients and therefore entered a necessary evolutionary arms-race with plants, defending themselves against potential toxins. However, PSCs are a diverse group of compounds, including alkaloids, cyanogenic glycosides, glucosinolates, terpenoids, saponins, and phenolics, among others. Since herbivores and omnivores cannot avoid bitter foods, they instead evolved elaborate and extensive biotransformation capacity to consume some of them. The consumer must neutralize the threat of the PSC by complexing the compound or breaking it down in the gut. If the gut cannot defend the animal against the compound or if the product is still reactive, then the liver acts as the major site of biotransformation [17]. This biotransformation capacity leads to the compound no longer being “toxic” to the animal, although the bitter signal may remain.
Salivary proteins (SPs) were identified in the 1980s as part of this defense system, and were often referred to as the “first line of defense” against tannins, a class of phenolic PSCs. Tannins make up an extremely common class of PSCs; they are considered bitter and astringent. Tannins are anti-nutritional, meaning they reduce the efficiency of nutrient metabolism, particularly for proteins [18], in species without the biotransformation capacity to consume them [19]. Tannins are the unpleasant flavor in unripe bananas, are noticeable in red wines and black tea, and are used by hosts of plants as a defense. A group of salivary proteins complex to tannins and reduce absorption of the compound [20]. Variation in tannin acceptance is one of the most noticeable divisions in mammalian foraging [21] and drives food selection patterns [21–23]. Thus, much of the work exploring the role of SPs and bitterness was first described as an adaptation to tannins in diet.
2. Salivary proteins are associated with diet
Mehansho et al. [24] experimentally demonstrated that rats could upregulate a class of SPs, the proline-rich proteins (PRPs), in response to a tannin-containing diet, and could safely consume the diet while maintaining body mass. In contrast, hamsters could not upregulate PRPs in response to the tannin-containing diet, and lost dangerous amounts of body mass. This work contributed to the idea that SPs were acting as a defense against PSCs. Glendinning [22] later demonstrated that mice with upregulated PRPs increased preference for solutions containing tannins, compared to mice without PRPs. This was an incredibly important finding, because it suggested that, not only did the PRPs protect the animals against the negative effects of tannins, but they also may have made the tannin more acceptable.
PRPs were also implicated in the reduction of astringency as a mechanism for increased tannin acceptance. In addition to being bitter, tannins are also known to be astringent, i.e., elicit a dry, puckering mouthfeel [25,26]. The mechanism of this sensation, which is considered tactile rather than taste and is carried via trigeminal mechanoreceptors, is relatively well-studied [27]. One hypothesis poses that the sensation of astringency arises from the interaction between compounds found in food (notably, tannins and alums), and certain PRPs which would otherwise contribute to the lubrication of the oral cavity. When PRPs bind to tannins to minimize their anti-nutritional effects, they form complexes that remain bound [28] and attract other tannin-PRP complexes, as well as other salivary proteins (including mucins), and eventually this complex reaches a size at which it is no longer soluble and precipitates from saliva [28,29]. This precipitate removes PRPs from saliva, resulting in a solution with decreased lubrication and increased friction. In support of this, mixing saliva with tannins results in a decreased solution viscosity and increased friction coefficient [30,31]. Alternatively, a second hypothesis poses that tannins interact directly with the oral mucosa, decreasing its lubricating ability [32]. In this hypothesis, PRPs actually prevent the perception of astringency, rather than participating in it, by scavenging tannins so they are unable to interact with the oral mucosa. In support of this, the sensation of astringency is reported even when humans have rinsed the saliva from their mouths, and addition of saliva decreases the perception of astringency. Additionally, tannin aggregating to the oral mucosa increases friction force, while the presence of PRPs prevent aggregation of tannins [32].
While much of the research to date has targeted tannins and the sensation of astringency, a few groups have focused their efforts on implicating SPs in variations in taste perception and food acceptance in humans. The primary methods for identification of proteins or protein classes of interest include protein separation by 1- or 2-D gel electrophoresis followed by excision of important bands (1-D) or spots (2-D) and identification of the sample by mass spectrometry [33–37]. Proteins of interest can be targeted by ELISA or Western blot [34–36,38,39]. Total sample proteomic analysis is often used to give a high-resolution, large-scale overview of a given protein profile via mass spectrometry (HPLC-MS [40–42]). Each of these methods has its own limitations and none of the methods alone is able to identify or quantify the full complement of SPs, Nonetheless, several studies have identified a handful of proteins that appear to relate to diet choices.
Salivary α-amylase has long been known to catalyze the digestion of starch in the oral cavity, and there is a wealth of research on the relationship between salivary α-amylase and starch perception [43]. Amylase activity has been correlated to changes in the perception of starch-thickened food [44] as well as hedonic ratings of fat and sweetness [38]. Adults with low sweet taste sensitivity have higher expression of α-amylase, as well as other proteins such as carbonic anhydrase VI (formerly known as gustin), and cystatins (cysteine protease inhibitors), than those who are more sensitive to sucrose [34]. SP profiles are also correlated with fat and salt taste sensitivity [35,45] and liking for salt [38] and, in children, a limited diet is marked by a different SP profile than an expanded (or “normal”) diet [33]. Subjects who are particularly sensitive to oleic acid (thought to be representative of fat taste) have high expression of cystatins (cystatin-SN, cystatin-D), zinc-α–2-glyco-protein, which is highly expressed glycoprotein, and carbonic anhydrase VI [35]. The relationship between bitter taste and SP expression has also had considerable focus. There are links between SPs (in particular carbonic anhydrase VI) and 6-n-propylthiouracil (PROP) taster status [40], and oral administration of certain PRPs (Il-2 and Ps-1, which are constitutively higher in PROP-identified supertasters) can facilitate increases in PROP intensity [46]. PROP-identified supertasters have higher constitutive expression of carbonic anhydrase VI and S-type cystatins [47]. In infants, SP profiles (particularly cystatins) are predictive of the acceptance of a urea-containing solution [39], and finally, those considered sensitive to the taste of caffeine have differences in SP profile from those considered insensitive [35,36]. In particular, increased caffeine taste sensitivity was related to more amylase and higher immunoglobulins (specifically, IgA) and lower expression of cystatin-SN [36].
3. Barriers and solutions to using animal models
Despite the years of data suggesting a role for SPs in taste and diet selection, experimental data were limited. The ability to carefully manipulate diet is difficult with human subjects but the inability to regularly collect saliva from animal models meant that there were very few intervention studies examining how salivary proteins are altered by diet or experimental demonstrations of how the proteins alter diet acceptance.
Traditionally, if large volumes of saliva were needed from rodents, experimenters would use a mixture of pilocarpine and isoproterenol to induce salivary flow and protein expression, respectively. Isoproterenol (IPR) is a beta-adrenergic agonist and pilocarpine is a muscarinic cholinergic agonist. Salivary secretion is controlled by the sympathetic and parasympathetic nervous systems, and these drugs are nonspecific activators; their use comes with a large number of serious side effects, including signs of illness, modifications of heart function, and changes in water balance. These side effects severely limit the ability to reliably collect behavioral data on food preference and intake, and animals cannot withstand consistent use of the drugs. To address this limitation, our lab adapted a technique originally developed by Pavlov; we developed a protocol to collect saliva from awake animals using classical conditioning. We pipette a sucrose and citric acid solution (2 mLs/day of 30 mM citric acid in 1 M sucrose) into the oral cavity daily for 2 weeks; at the conclusion of this training, the animals are conditioned to salivate in the presence of a pipette. When collecting saliva, we are able to use an empty pipette to aspirate saliva from around the teeth and under the tongue. We chose to use a “sweet” and “sour”2 mixture for two reasons. First, this mixture is predicted to release saliva from multiple salivary glands [48]. Second, the sucrose makes the solution palatable to the animals. This method gave us the tools to begin to explore how SPs are affected by diet, and if those changes correlate with diet acceptance behaviors.
4. Diet changes salivary protein expression
Because the most extensive data on SPs altering diet choice was the relationship between PRPs and tannic acid tolerance, the first study we conducted using our saliva collection method was to track the upregulation of SPs after rats were exposed to a tannin-containing diet. It had previously been demonstrated that rats given a tannic acid diet would upregulate PRPs, but the ingestion microstructure of the increased acceptance and the time course of the protein changes were unknown. We gave male rats a control diet that was designed to have little or no PSCs [49]. We collected saliva from the animals on the control diet for baseline comparison, and then transitioned animals to a diet containing 3% tannic acid. Whole saliva was analyzed by 1-D gel electrophoresis. In agreement with previous research, animals on the tannic acid diet upregulated the expression of a subset of their SPs in response to the diet exposure. This upregulation took approximately 4 days, and the upregulation of specific SP bands were correlated with changes in diet acceptance, specifically with rate of ingestion and meal size. Rate of ingestion can be considered a behavioral measure of orosensory information. When an animal, including a human, is fed an increasingly hedonically positive stimulus, they eat faster [50,51]. As the SP content increased in saliva, the animals ate faster, suggesting that the orosensory information from the food was less negative with the SPs present. This finding was consistent with the literature describing the relationship between PRPs and tannic acid. Surprisingly, we also identified a subset of non-PRPs altered by our treatment. Notably, we saw changes in cystatin-S, which has been implicated in several studies looking at saliva and acceptance of normally avoided stimuli [36,52].
Next, we repeated the aforementioned study but used a diet adulterated with quinine, an alkaloid PSC that is described as bitter, but not astringent. We wanted to determine if salivary modifications were a specific evolutionary response to tannic acid, or if other diets were capable of modulating SP expression. Animals who consumed the quinine diet upregulated a subset of SPs, while there was no change in the control animals. We also pair-fed the control diet to a separate group of rats in which the total amount of food offered was based on the consumption of the quinine group; these rats exhibited no differences in SP expression. This suggested that the changes were due to the presence of quinine, and not repeated saliva collections or a reduction in food intake. This was the first report we are aware of that non-tannin diet exposure could cause long-term alterations of SPs, as opposed to within-meal or meal anticipatory effects [53,54]. The changes in the quinine-fed animals mirrored the tannin study in several ways. Proteins were upregulated across a similar period of time (4 days) and were correlated with the same microstructural measures of consumption: meal size and rate of ingestion. We also saw a similar protein expression pattern to tannin consumption (Fig. 1). Injection with IPR, and diets containing tannin or quinine indudce very similar patterns of protein expression, which implicate parotid-derived SPs as contributing to the oral modification of tannin and quinine given that the parotid gland has been identified as the main site of IPRs effect [55].
Fig. 1.
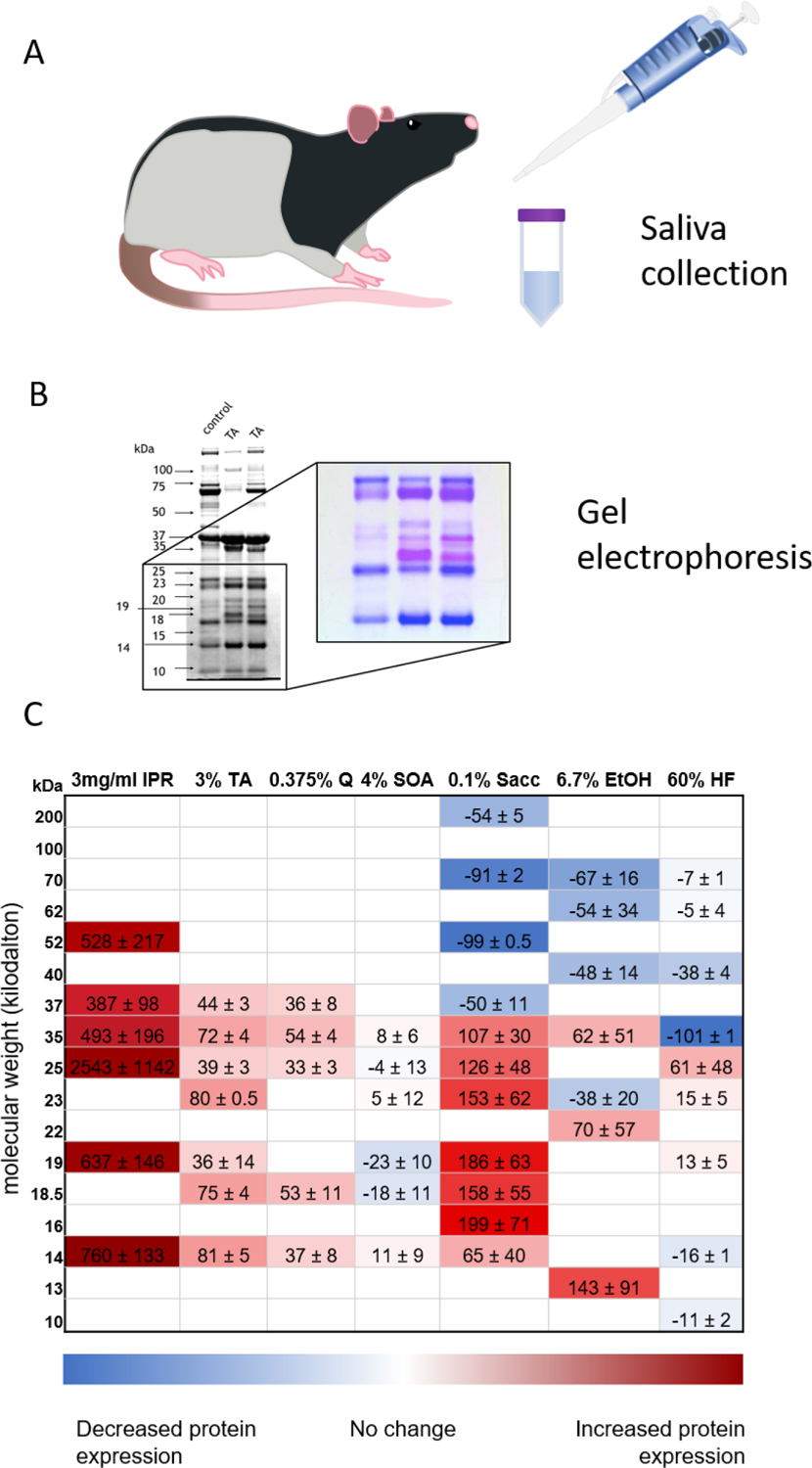
Panels A & B are methodological illustrations. Panel A) is an illistrutive reminder that saliva is collected by pipetting from the oral cavity of rats conditioned to salivate in the presence of a pipette. B) is an example of a gel highlighting the comparison between saliva collected while the animals are on a control diet (control) and saliva collected after tannic acid exposure (TA). A portion of the gel is enlarged and is presented in coomassie stain. Due to the binding characteristics of proline-rich proteins the bands containing this class of proteins stain pink. Several bands containing these proteins are visable in the upregulated lanes of the insert. C) is a heat map showing SP profiles for animals after injection of isoproterenol or dietary exposure to various diets: isoproterenol (IPR; n = 4), tannin acid (TA; n = 12), quinine (Q; n = 8), 4% sucrose octaacetate (SOA; n = 5), 0.1% saccharin solution (Sacc; n = 8), 6.7% ethanol (EtOH; n = 5), and 60% high fat (HF; n = 4). Data are relative protein expression densities in kilodaltons (kDa) presented as a difference from baseline, corrected by control group, and multiplied by 100 to enhance colors (average (change in protein density of experimental animal from its own baseline to the last day of diet treatment) – (average change in control group across the same time period) *100). The intensity of the red represents relative increase in protein expression, and the intensity of the blue represents relative decrease in protein expression. IPR injection methods: 3 mg/ml isoproterenol and 30 mg/ml pilocarpine are injected subcutaneously in anesthetized animals as described in [66]. Diet treatment methods: Animals were placed on a control diet for ~2 weeks and control saliva samples were collected as previously described on the last four days of control diet. Half of the animals in each group were then switched onto their experimental diet (or saccharin solution) for a minimum of 8 days while the other half remained on the control diet. Saliva was collected from all animals throughout the experimental diet. Sample analysis methods: As described in [58,61,66], saliva samples were diluted 1:1 with trifluoroacetic acid (TFA) and spun to remove debris from samples. Sample protein concentrations were determined using the Pierce BCA Protein Assay Kit (Thermo Fisher Scientific). Samples were combined with 1/3 vol of 4x Invitrogen sample buffer, heated at 82°C for 10 mins and resolved on a 12% SDS-PAGE with MOPS running buffer. Molecular standards were run simultaneously along with the samples to determine molecular mass of each band. Gels images were captured using the Azure c400 imager and densitometric analyses were performed using AzureSpot software. It is important to note that this figure represents changes in protein bands, which implies different patterns of expression. Each band contains multiple proteins, therefore a protein specific comparison cannot be inferred from this data presentation.
To ask if all bitter compounds produce the same pattern of protein expression, we offered rats the synthetic compound, sucrose octaacteate (SOA), which is described as “bitter” and activates a T2R “bitter” receptor [56]. Surprisingly, SOA did not alter SP expression in any measurable way ([58, 59], Fig. 1), even at concentrations that produced a similar level of rejection to the quinine diet. Although SOA is considered “bitter,” several studies have documented behavioral differences between SOA and quinine acceptance patterns [59,60]. The lack of response to SOA suggests it is not just “bitter” stimulation that alters protein expression, but there is an unknown characteristic that is absent in SOA. We later exposed rats to a wide assortment of diets for at least 2 weeks each: 0.1% saccharin, 6.7% EtOH, and a 60% high-fat diet (Fig. 1). Visual comparison of the SP expression on these diverse diets shows that each diet seems to drive salivary protein patterns unique to that diet. We have not yet systematically considered the down-regulation of SPs, but we know that for tannin- and quinine-induced changes, SP expression persists at least 6 days after the animals returned to a control diet [61,62]. We also have not systematically explored the relationship between the rate or scale of SP change with total intake or intensity of the experimental diet. For example, some of the same protein bands we see upregulated by quinine exposure are upregulated by exposure to saccharin, which is more readily consumed than the “bitter” diets although it is thought to interact with “bitter” taste receptors.
In addition to abundant data describing variation in human SP expression [33,34,38,39], the ability to alter SP expression with diet does not appear to be limited to rodent models. There are distinct changes in the saliva of habitual alcohol users (with nicotine [63], nicotine not reported [64]) and in children with habitually limited diets [33]. Running and colleagues recently used an intervention model and demonstrated that a dietary introduction of daily polyphenolics was able to alter SP expression in human participants [65].
Though the unique SP expression patterns driven by different diets are both important and interesting, our focus has remained on bitter stimuli, and using behavioral and electrophysiological analyses to elucidate the mechanisms of increased bitter diet acceptance. The correlation between SP expression and rate of ingestion of quinine-adulterated food suggested that the SPs were contributing to a change in taste. Unlike tannins, quinine is not considered to be an astringent compound. The major limitation of the early data sets was that the implication of the SPs was entirely correlational. To try to establish a causal relationship between SP expression and diet acceptance, we decided to take advantage of the similarity in protein expression after IPR injection, tannin and quinine diets (e.g. Fig. 1).
5. Salivary proteins change diet acceptance
We reasoned that if the proteins were driving a change in the taste or acceptance of the diet, then we should be able to increase measures of acceptance by pre-treating the animals with treatments that increased those proteins. Nature gave us an easy way to test this hypothesis since IPR, quinine and tannic acid had such similar SP expression patterns after exposure, therefore we measured how animals with different pre-treatments consumed a quinine diet. One group of rats was injected with IPR daily (for 3 days) to upregulate SP expression, while another was fed tannic acid diet to increase SPs. There were also several control conditions: some rats were fed a control diet ad-lib, some were pair-fed the control diet to match the average intake of IPR-treated rats, and a third control group was given 4% sucrose octaacetate (SOA). As was discussed above, IPR injections cause a dramatic reduction in food intake, and the pair-fed group was a control for that reduction. SOA-treated animals were a control for “bitter” diet exposure but without altered SP expression. After pre-treatment, all animals were offered a 0.375% quinine diet. The two groups that had SPs upregulated by their pre-treatment (IPR and tannic acid) showed a significant increase in the rate of ingestion the first time they were offered a quinine diet (Fig. 2) compared to their controls [58]. We did not see this effect in any of the control manipulations (SOA or pair-feeding) on rate of ingestion. These data suggested to us that salivary protein profiles could alter the acceptance of a novel diet via a taste-driven mechanism.
Fig. 2.
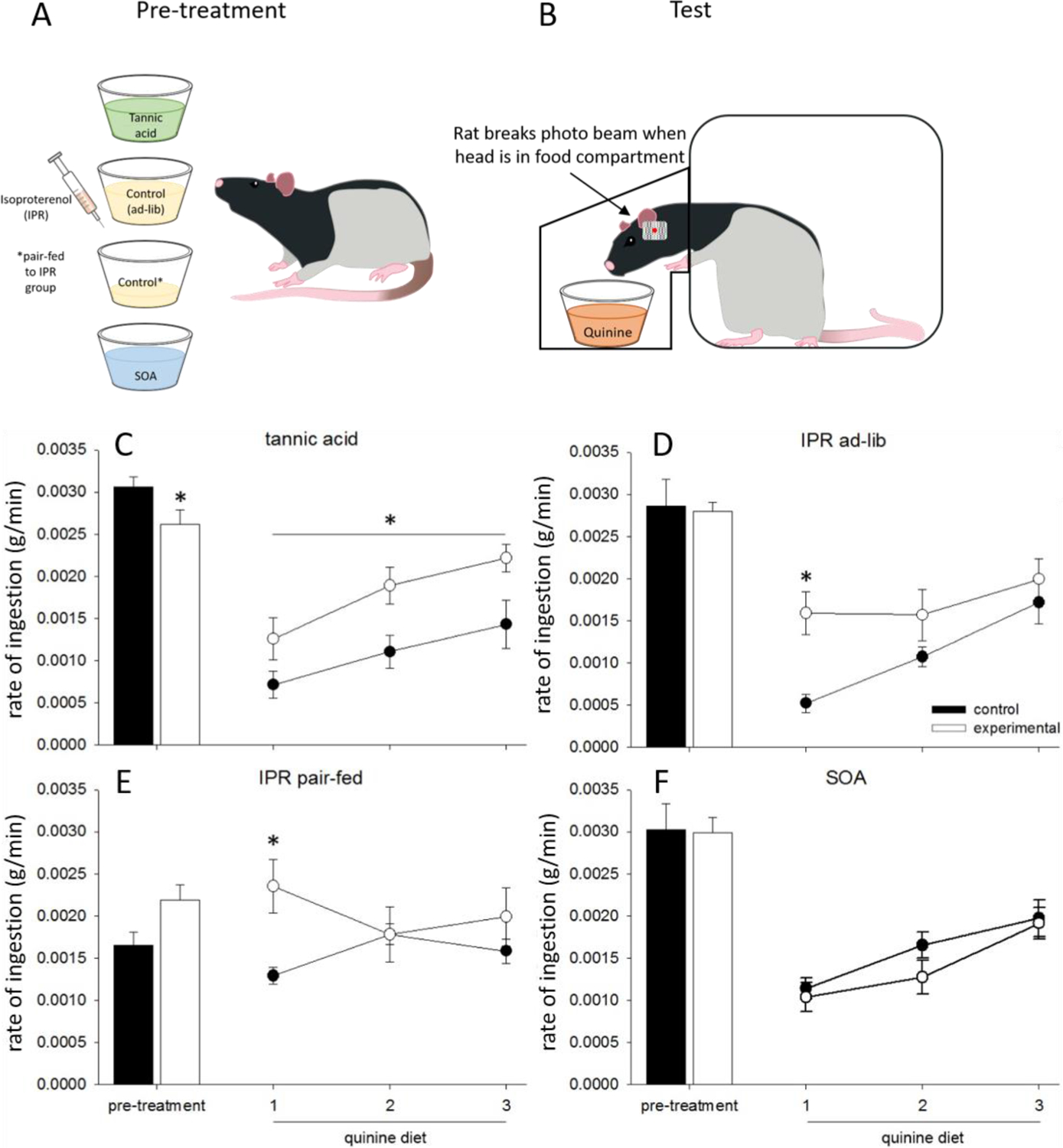
Panels A and B are methodological illustrations. A) Groups of rats were pre-treated with treatments designed to upregulate salivary proteins (tannic acid diet or injections of isoproterenol (IPR)) or one of two control conditions (sucrose octaacetate or a control diet pair fed to mirror intake of the IPR treated group). B) Rats were then given access to a diet containing quinine and the custom caging was able to measure the rate of ingestion as well as other measures. C) This figure is modified from one originally published in Martin et al. 2019 [58] and is reprinted with permission of the publisher. Data are average rates of ingestion (g/min) ±SEM in rats fed tannic acid (TA), sucrose octaacetate (SOA), or control diet. (C–F). Closed bars represent the 3-day average of rate of ingestion rate of ingestion for control animals during the pre-treatment phase, during which they consumed control diet, while open bars represent experimental animals who were exposed to either TA (A), IPR injections (B, C), or SOA (D). Closed circles represent quinine intake of control animals, while open circles represent quinine intake of pretreated animals. TA (A) and IPR pre-treated (C, D) animals ate quinine faster than their controls. * indicates significant differences (p < 0.05).
6. Salivary proteins modify measures of taste
Rate of ingestion was correlated with SP expression, and altered bySP upregulating pre-treatment which suggested there was a change in orosensory information. However post-oral feedback, especially in long-term tests, can contribute to rate of ingestion. To examine the role of SPs in more narrow measures of taste-guided behavior, we first used a relatively simple model: the brief-access taste test. Animals were given various concentrations of tannin solution [62], quinine solution [61] or sucrose solution [66] for 30-s trials. Animals were tested in a custom designed cage known as the Davis Rig (Med Associates, DiLog Instruments). The Davis Rig has a plexi-glass cage with a slot at the front that gives the animal access to a sipper tube, which is mounted on a sliding table. A mechanical shutter controls access to the sipper tubes. Once the shutter opens, the animals can choose to lick the solution for up to 30 s, then the shudder will close for 10 s while the mechanized table changes the available tube. In this way, animals are given very brief exposure to each stimulus. It is thought that this brief access will limit integration of oral and post-oral signals, and that the licking seen within the 30-s trial is driven by orosensory stimulation alone. Each of the taste qualities were tested in three 30-min tests, made up of as many trials an animal could initiate, across 3 non-consecutive days. Animals were tested in this paradigm before and after they were fed a diet to upregulate SP expression. In this paradigm, we are able to record concentration-dependent behavior and fit psychometric curves.
We found that rats with SPs upregulated by tannin increased licking to intermediate concentrations of tannin [62], and rats with SPs upregulated by quinine increased licking to intermediate concentrations of quinine and did not change licking to sucrose [61]. When testing ligands reported as “bitter” in the paradigm, we water-deprive the animals and thus at low stimulus concentrations, they drink at near-maximal rates. As the concentration of a bitter stimulus increases, licking decreases until the animals are only sampling the tube. The effects of the SPs were specifically seen at the intermediate solution concentrations, where control animals began decreasing licking, presumably due to the increasing aversive properties of the stimulus; SP-upregulated animals acted as if these concentrations were not yet aversive. These data suggested once again that taste responsiveness was being altered by salivary proteins.
Brief-access licking is a convenient and straightforward test, but the data rely heavily on the hedonic evaluation of the stimulus and motivational state. We saw that the animals were more accepting of intermediate concentrations of tannin or quinine, but this task is unable to disentangle if this is a change in the perceived intensity of the stimulus (i.e., it is less strong), a change in the hedonic evaluation of the stimulus (i.e., it is less negative), or some other change in the motivational state of the animal. We hypothesized that the SPs were actually changing the taste detection of the stimulus, i.e., SPs were masking the “bitter” taste of quinine, making it seem less intense. To minimize the confounds inherent in the brief access test, we employed a psychophysical procedure to test animals for their ability to detect perithreshold concentrations of quinine in a 2-response operant task before and after they were treated with tannin, quinine, SOA or control diet. In this task, we used a stimulus delivery system/operant chamber referred to as the gustometer (Fig. 3). This gustometer has been extensively used to asses taste sensitivity in rats [57,67–69]. Briefly, it is a modified Skinner box with three open slots in the front of the cage. The animal has access to a very small amount of a stimulus through a center slot. The animal is trained to respond to the taste of the stimulus by licking at a manipulanda behind one of the side slots to receive a water reward. For example, if the stimulus is bitter, the animal may be trained to lick to a manipulanda to the right, and if the stimulus is not bitter (i.e., water), then the animal would lick at the left manipulanda. The taste-associated manipulandum is usually counterbalanced across the groups of animals. With training, rats are very adept at this task; at high concentrations, rats perform with an accuracy rate over 90% (Fig. 3). The task is also robust; when rats are tested and re-tested two weeks later while maintained on a non-bitter control diet, there is no change in the psychometric curve (Fig. 3E).
Fig. 3.
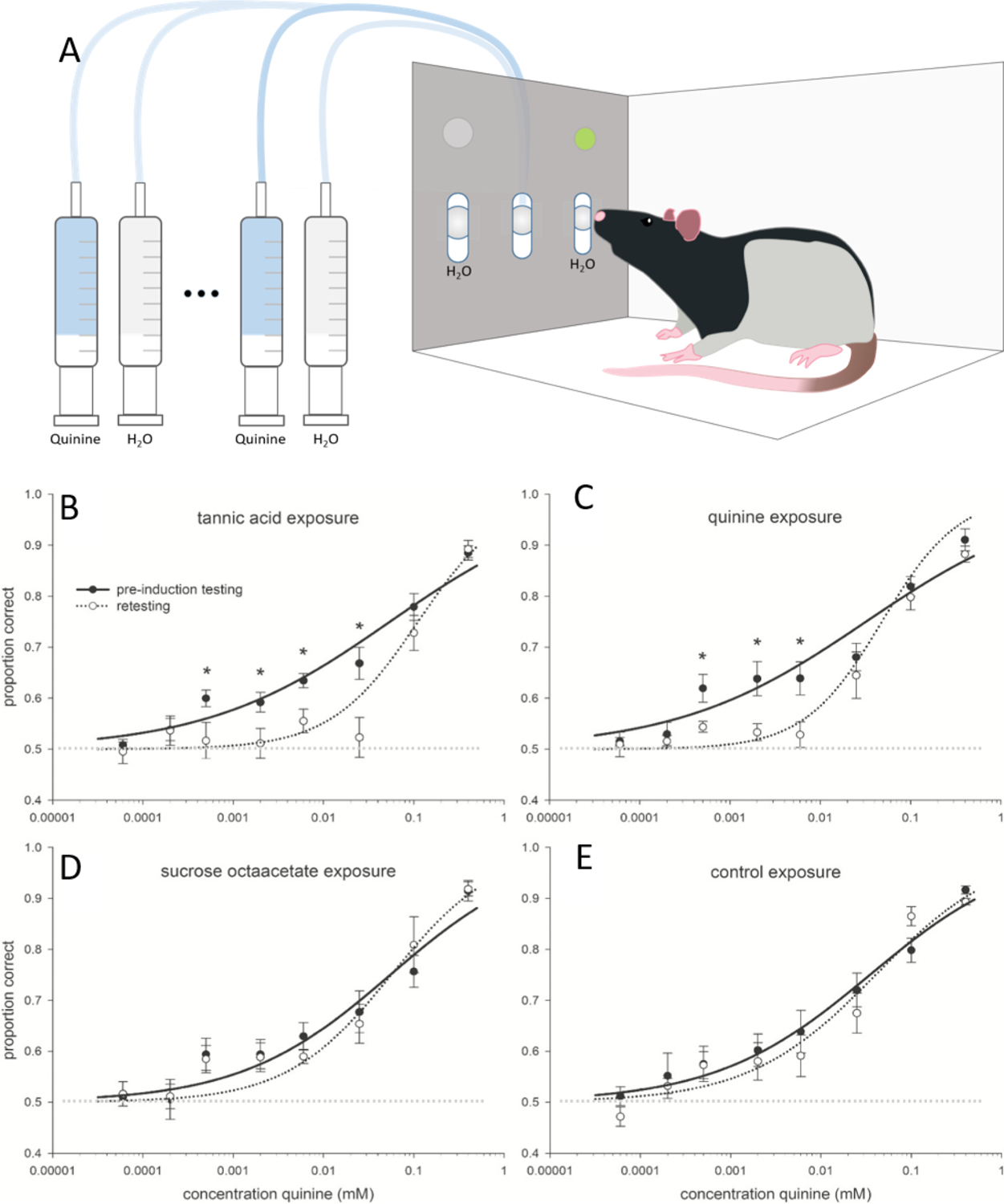
A) is a simplified methodological illustration of the gustometer. Small amounts of either water or various concentrations of quinine were delivered by a series of syringe pumps to the center slot and rats were trained to respond to the left or right manipulandum to report the presence or absence of quinine in the stimulus. B) This figure is modified from one originally published in Martin et al. 2019 [57] and is reprinted with permission of the publisher. Data are mean (±SE) proportion correct as a function of stimulus concentration for quinine. The solid lines represent performance accuracy during the preinduction testing phase, whereas dotted lines represent performance during the retesting phase. A faint dotted line also marks the 50% point of the graph. At 50% where animals are choosing a response at random i.e. are no longer able to discriminate between the water and quinine samples. Asterisks represent significant within-group differences between preinduction testing and retesting. Animals were tested, then exposed to 3% tannic acid (A), 0.375% quinine (B), 4% sucrose octaacetate (C), or a nonbitter control (D), and retested. The curves were fit to the data based on a 3-parameter logistic function#. SE, standard error. * indicates significant differences (p < 0.05).
# 3-parameter logistic function to calculate the curves : [(a − 0.5)/(1 + 10(x − c)*b)]+0.5[(a − 0.5)/(1 + 10(x − c)*b)]+0.5, where the curve parameters, a, b, and c, were defined as the asymptotic maximum performance as determined by the mean of the stimulus control sessions, the slope of the curve, and the midpoint concentration between asymptotic maximum and minimum (EC50), respectively.
To test if the upregulation of SPs would alter detection thresholds, we tested all of the animals while they were maintained on a control diet (pre-induction testing). Some rats were then given diets that altered SP expression (quinine, tannin) for 2 weeks or were maintained on the control diet or given SOA. We then returned rats to the control diet and re-tested them. We found that both of the treatments that altered SP expression (tannic acid; Fig. 3B. and quinine; Fig. 3C) dramatically altered performance. Rats in these conditions began responding at 50% correct (i.e., chance) at much higher concentrations than they had in their pre-induction test. We do not believe this was due to experience with “bitter” taste or a hedonically negative diet, since we do not see the same change in the group that had been offered SOA (Fig. 3D). It is also unlikely to be due to having prior experience with the test, because neither the control group nor the SOA group demonstrated any changes between their tests. To determine if having SPs upregulated altered the way an animal could perform the task in general, we repeated the experiment with tannic acid-exposed animals performing the threshold detection task for sucrose. SPs upregulated by tannin did not alter detection thresholds to sucrose. Together, these data suggest that the presence of SPs upregulated by exposure to quinine and tannic acid increased bitter detection threshold (i.e. decreased sensitivity). As with the brief-access test, they responded at intermediate concentrations as if the solution was less concentrated.
While the work we had done up to this point supported the hypothesis that SPs altered taste, the designs we used could not exclude learning from the paradigm. We controlled for “bitter” exposure by feeding animals SOA, but to appropriately test the hypothesis that SPs modify taste responsiveness, we needed naïve rats with upregulated SPs, and none of the negative effects of IPR injections. To do this, we developed another paradigm: the donor saliva system. We had previously demonstrated that pre-treatment with IPR altered SPs in a way very similar to quinine and increased quinine acceptance (Fig. 1,Fig. 2 and [58]), so we reasoned that if we delivered saliva from IPR-treated animals to bitter naïve animals, we should see the same increased quinine tolerance. We chose to use IPR in our donor animals instead of diet treatments because with dietary treatment we can collect only around 50 μl of saliva per day at a protein concentration of approximately 3 mg/mL. IPR and pilocarpine treatment allow us to collect around 2 mLs per collection at approximately 12 mg/mL total protein content. This means that a single donor animal is able to donate approximately 8 mLs of saliva when the saliva has been diluted to physiologically relevant levels. Being able to collect large volumes of saliva is essential for studies where it is continuously infused.
To ask if the presence of SPs alone was able to alter taste responsiveness, we took advantage of the taste reactivity paradigm. Taste reactivity tests quantify the reflexive and taste-guided [70] oral motor responses to taste stimuli (Fig. 4A). For example, rats given a “sweet” stimulus display ingestive responses including rhythmic mouth movements and tongue protrusions, while rats given a “bitter” stimulus display rejection responses including gapes, forelimb flails, and chin rubs. Taste reactivity was the ideal paradigm for this test because stimuli can be delivered while the animals are need-free. Additionally, the stimuli can be delivered in predetermined volumes. For solution delivery, animals were implanted with oral catheters. Once they were healed and trained, male rats were tested in a specialized infusion chamber equipped with a camera under a clear plexiglass floor (Fig. 4B), which allowed visual access to the oromotor and somatic behaviors of the animal during the infusions. Animals were infused with donor saliva, donor saliva which had the proteins removed by centrifugal filters, artificial saliva, or water; each of these four conditions was delivered both with and without quinine. Animals that were given quinine in the presence of SPs (i.e. whole saliva) showed no increase of aversive or rejection oromotor movements over an infusion of saliva alone (Fig. 4C and [66]). In contrast, addition of quinine to protein-free saliva, artificial saliva, and water resulted in large increases in rejection behaviors. These data further support the hypothesis that the presence of SPs decreases the salience of quinine in such a way that the animals treat it as less concentrated. SP infusions did not generate increased positive orosensory movements or alter responding to non-bitter stimuli, suggesting that the effect is specific to reducing “bitterness,” [66]. The effect is also concentration-dependent; as we decreased the concentration of SPs, we saw an increase in aversive behavior to quinine in the solution, even though the quinine concentration remained constant [66].
Fig. 4.
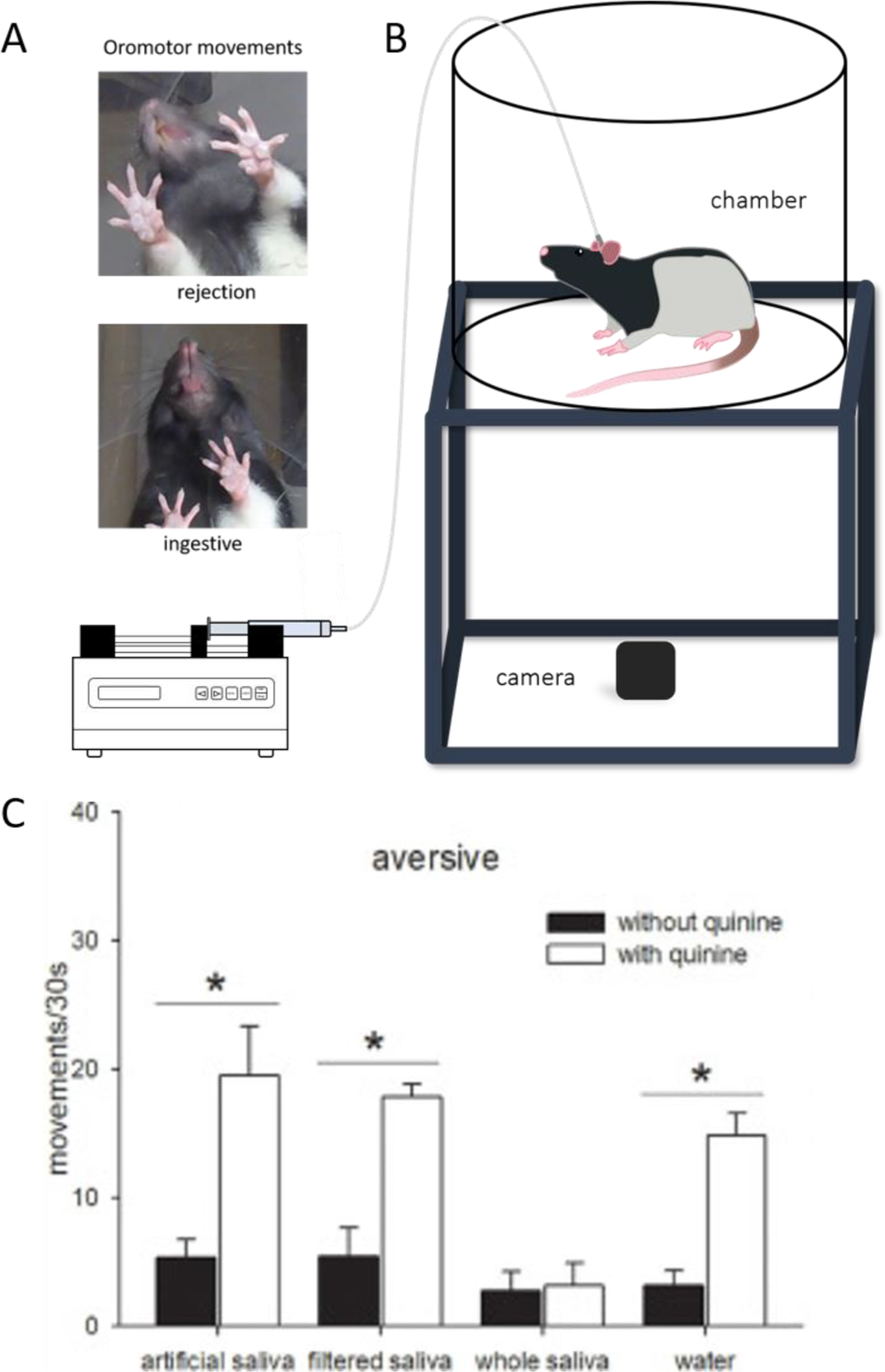
Panel A) depicts examples of rejection and ingestive responses in the taste reactivity paradigm. B) is an illustration of the taste reactivity chamber. An animal is filmed by a camera placed beneath a clear floor while an infusion pump delivers a predetermined stimulus directly into the oral cavity. C) This figure was originally published in Martin et al. 2020 [66] and is reprinted with permission of the publisher. Data are average (n = 6, ±SEM) number of movements in response to 30 s infusions of artificial saliva, filtered saliva, whole saliva, and water, with quinine (white bars) or without quinine (black bars). Movements are categorized as aversive (gapes, head shakes, chin rubs, forelimb flails, and face washes). * indicates significant differences (p < 0.05).
This work offered strong evidence that SPs were altering taste-driven behavior. We have also collaborated with Dr. Robert Contreras to test if these changes were happening peripherally. We used the donor saliva model to deliver quinine, with and without SPs, to the fungiform papillae of anesthetized rats while the Contreras lab recorded multi-unit activity from the chorda tympani (CT) nerve. The CT is a branch of the facial nerve which innervates the front of the tongue. Consistent with the behavioral data, we found that when SPs were present there was suppressed CT activity compared to when quinine was delivered alone [62]. Again, this effect was specific to quinine because we saw no effect of SPs on salt-evoked CT responses.
7. Summary
There has been strong evidence in the human literature that SPs are correlated with food preferences [33,35–39]. Our work in rodent models suggests that diet exposure changes the pattern of SP expression. In the case of exposure to a quinine-laced diet, the particular pattern of proteins that are upregulated can then alter the acceptance of a quinine diet, at least in part, by altering the taste of the diet. Rats behave as if the quinine is less intense in the presence of the SPs than in the absence of the proteins. While the evidence that SPs (broadly speaking) alter taste is becoming more convincing, there is still a considerable number of unknowns.
8. Unknowns and future work
8.1. The protein identification mystery
The most pressing and obvious question is which proteins are responsible for driving taste signal modification. However, this question has so far proved to be difficult to answer. The salivary environment is incredibly complex and can contain over 1000 proteins [71]. These proteins are produced from major and minor salivary glands and mix in the oral cavity where they are cleaved or modified by each other and the oral environment. Protein identification is also complex as many of the proteins of interest are either proline- or glycan-rich, making them hard to identify by mass spectrometry. To date, some proteins have been identified in correlational analyses as playing a role in taste or flavor responding including proline-rich proteins [19,21,40,41,49,72,73], mucins [74,32], prolactin inducible protein [37], cystatins [36,37,39], carbonic anhydrase VI [37,75], gliadoralin A [76], SMR1 [61], Von Ebner’s gland protein [77] and α-amylase [36,44,74,78]. More recently work has begun to suggest a role for biotransformation enzymes such as glutathione-S-transferase in altering taste compounds using in vitro systems [79]. To our knowledge, none of these proteins have been directly tested in controlled taste tests, although some work has been done with carbonic anhydrase VI and proline-rich proteins, demonstrating that manipulations of the protein alter diet acceptance [75]. It is to be elucidated if these proteins work independently, act on other proteins in the saliva, or work in combination to form complexes. Although rodent and human saliva are distinct and have many species specific proteins [80] the major classes of proteins discussed here (e.g. proline rich proteins, mucins, cystatins) and many of the specific proteins are expressed in both species (e.g. amylase, carbonic anhydrase, Von Ebner’s gland proteins) making rodents a good model for taking these studies forward into experimental manipulations. If the protein or protein complexes could be experimentally identified, perhaps they could be used to modify bitter compounds as part of pediatric medical formulations or as part of dietary treatments.
8.2. The mechanism mystery
While it is becoming very clear that SPs do play a role in taste signaling, the mechanism is yet unknown. Three potential mechanisms have been proposed. The first is that SPs form a mucosal pellicle to reduce access of the stimulus to the receptor (pellicle hypothesis). In support of this hypothesis, a number of the proteins that correlate with taste alterations are involved in pellicle formation [36,76]. Inconsistent with this hypothesis is that the pattern of SP expression we have worked with specifically alters “bitter” but not sucrose or NaCl taste [76] suggesting that the pellicle would have to be permeable to some but not all stimuli. Furthermore, saliva from donor animals washed across the tongue decreases taste nerve signaling [76] which suggests that pellicle formation is not necessary for SPs to exert their action. The second hypothesis proposes that SPs bind to the stimulus and keep it from interacting with the receptor (stimulus-binding hypothesis). In support of this hypothesis, there is evidence that SPs form complexes with a variety of stimuli [49,81,82] and as discussed earlier, precipitation of tannins by SPs is one of the proposed mechanisms for changes in tannin astringency. However, the ability to bind to the stimulus is not sufficient evidence that it is mediating the change in taste response. The role of most of the proteins produced in saliva is to bind [83]. SPs bind to each other to form pellicles that protect the tissue and teeth, and can bind to bacteria, the tooth surface, tissue surfaces, and so on. Due to the “sticky” nature of these proteins, binding assays alone have historically revealed little about the true nature of the protein interactions. The third hypothesis states that SPs bind to the target taste receptor and alter its responsiveness to stimuli (receptor-binding hypothesis), but with few SP targets to explore, there is no work to our knowledge that has systematically tested this hypothesis.
9. Other potential actions of saliva
Though taste is an important part of its perception, food never generates a taste sensation alone. Notably, much of what humans consider “taste” is actually “flavor”, or the confluence of taste and food odor [84]. Though food odors can be sensed orthonasally (through the nose), retronasal olfaction plays the chief role in food and drink flavor perception. Retronasal odors are generated during chewing when food particles are broken down both manually and chemically; volatile compounds in food or drink are released and mixed with air, which travels back up the nasal cavity to the olfactory epithelium [85]. It is just as likely that SPs alter the activity of odor volatiles as they do taste stimuli, as odorants have the same opportunity for interaction with saliva [86]. In fact, there is some work already demonstrating the interaction of SPs and odorants. Mucins and α-amylase have been shown to prevent the release of volatile esters and ketones (i.e., volatiles that are perceived as fruity and pungent, respectively) [74]. True to their binding nature, mucins appear to bind and prevent the release of some hydrophobic compounds, but can also decrease the retention of volatile compounds [87]. Volatiles are also released through metabolism; changes in salivary lipase activity have been shown to alter the perception of cheese aroma [88]. Finally, perception of wine aroma correlates with SP profile [89,90].
This work is critical to understanding the acceptance of foods that are called “bitter,” because perceived bitterness is not the only reason people reject green vegetables. The quality of vegetable odor differs between vegetable preferers and avoiders [91], and is inversely related to vegetable consumption [8,92]. Notably, some PSCs found in vegetables are described as pungent and sulfurous. During vegetable preparation or chewing, glucosinolates, found in vegetables, are hydrolyzed into free sulfur volatiles as well as isothiocyanates, indoles, and nitriles [93]. These compounds have been implicated in the acceptance of Brassica vegetables. Increased sulfur volatile production by in-mouth salivary activity is related to changes in odor quality perception [94], and is inversely related to liking of raw vegetables [95]. More work is necessary to understand if SPs are able to alter flavor perception of vegetables in the same way they can modify bitter taste.
10. Role of saliva in post-oral feedback
During eating, saliva is mixed in with food in the oral cavity and swallowed; this means that the ability of saliva to interact with our foods does not necessarily end in the mouth. There are a few lines of evidence suggesting that some SPs can survive the gut. First, proline-rich SPs can survive through the gut of ruminants [96], which is a substantially higher pH than rat or human [97]. Second, the proline is excreted in the feces of rats consuming tannin diets [49] as the tannin and proteins are apparently still complexed together. There are two ways that SPs, if they are surviving the stomach environment, could continue to act in the gut. It has been demonstrated that SPs can reduce the transport of tannins across cultured intestinal cells [81], suggesting that if the SP is bound to a bitter stimulus it may alter absorption. The second way SPs could work in the gut mirrors their action in the mouth: by changing the way the stimulus interacts with “bitter” (i.e., T2R) receptors. These are not mutually exclusive possibilities.
In our prior work, we have demonstrated that altering SP profiles leads to changes in diet acceptance by altering the rate of ingestion and the size of meals consumed [61,62]. We have discussed rate of ingestion at length, but meal size is also an interesting microstructural measure because it is often associated with post-oral feedback. As an individual consumes a bitter, and potentially toxic, food source, the associated risk is not simply that the food contains a toxin, but rather that the toxin is consumed in quantities that surpass the animal’s capacity for biotransformation. The risk is in the dose, which is determined by meal size [16, 98,99]. Activation of several mechanisms in the gut reduce meal size; however, in the case of bitter stimuli, one mechanism that is notable is that activation of the intestinal T2R “bitter” receptors causes the release of satiety signals like cholecystokinin [100]. This suggests that if SPs interact with the stimulus or the receptor, they could show a similar action in the gut as they do in the oral cavity, which would explain the increased meal size exhibited by rats with upregulated SPs. Our lab is currently working to test hypotheses about the role of SPs in modified post-ingestive feedback.
11. Conclusions
The circular relationship between repeated diet exposure and increased acceptance is well documented [9,10,101–103]. We postulate that SPs may be in part mediating increased acceptance of bitter foods after repeated exposure, and that they serve this function as a physiological facilitator for learned food safety. If a foraging animal (or a toddler) consumes a diet with no ill effects, the animal should be able to incorporate that food source into their niche. Perhaps SPs are upregulated by repeated sampling of a food, so when an animal encounters a given plant again, it does not have to “remember” that this particular plant is safe, and instead it just does not taste as bad as it used to. We have only examined the relationship between the pattern of proteins that we can duplicate with IPR and quinine diet because of the experimental convenience of the model, but it is possible that the pattern of proteins that is present after saccharin or alcohol exposure could alter taste responsiveness to other taste qualities. We have also focused on males and are now examining the phenomenon in cycling female rats. Females may be particularly interesting with the added complexities that the estrous cycle alters food intake [104], and estrogen receptors are differentially expressed in the salivary glands [105]. This is a new field of inquiry and we expect that as more labs begin investigating the phenomena described here we will learn the extent to which saliva contributes to food choice.
Acknowledgements
From A-M Torregrossa: This manuscript is based on the work that was to be presented during the 2022 Annual Meeting of the Society for the Study for Ingestive Behavior. Laura Martin and Verenice Ascencio Gutierrez collected the data in Fig. 1 and wrote sections of the manuscript. I regret that due to a family emergency my trip to Porto was canceled at the last minute. Missing this meeting is a deep regret. I cannot state strongly enough how honored I am to have received the Alan N. Epstein Research Award this year. I attended my first SSIB meeting in Philadelphia in 2001 as technician in the Bourne lab. For the last 20+ years SSIB has been a great source of friends, collaborations, support and one of my scientific homes. I am so grateful to the Society for the Study of Ingestive Behavior awards committee and those who supported my nomination. I want to thank my current and former (official and unofficial) mentors Drs. Gerard P. Smith, Denise M. Dearing, James C. Smith, Lisa Eckel, and Alan Spector for your unwavering guidance and support throughout my training and career. I would also like to thank the three anonymous reviewers for their thoughtful comments and to note that Dr. Laura E. Martin created all figure illustrations. This work was supported by NIH R01DC016869 and NSF Career Award IOS 1942291.
Footnotes
2 Note that references to “bitter,” “sweet” “sour” etc. will be presented in quotation marks when discussing animal work. One can never be sure that the perceptual experience of a rodent is identical to that of the human experience described by these adjectives.
References
- [1].Chao AM, Quigley KM, Wadden TA, Dietary interventions for obesity: clinical and mechanistic findings, J. Clin. Invest (2021) 131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [2].Hoare JK, Jebeile H, Garnett SP, Lister NB, Novel dietary interventions for adolescents with obesity: a narrative review, Pediatr. Obes 16 (2021) e12798. [DOI] [PubMed] [Google Scholar]
- [3].Dreher ML, Ford NA, A comprehensive critical assessment of increased fruit and vegetable intake on weight loss in women, Nutrients 12 (2020) 1919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Kaiser KA, Brown AW, Bohan Brown MM, Shikany JM, Mattes RD, Allison DB, Increased fruit and vegetable intake has no discernible effect on weight loss: a systematic review and meta-analysis, Am. J. Clin. Nutr 100 (2014) 567–576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Appleton KM, Hemingway A, Saulais L, Dinnella C, Monteleone E, Depezay L, et al. , Increasing vegetable intakes: rationale and systematic review of published interventions, Eur. J. Nutr 55 (2016) 869–896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Al-Khudairy L, Loveman E, Colquitt JL, Mead E, Johnson RE, Fraser H, et al. , Diet, physical activity and behavioural interventions for the treatment of overweight or obese adolescents aged 12 to 17 years, Cochrane Database System. Rev (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Schatzer M, Rust P, Elmadfa I, Fruit and vegetable intake in Austrian adults: intake frequency, serving sizes, reasons for and barriers to consumption, and potential for increasing consumption, Public Health Nutr 13 (2010) 480–487. [DOI] [PubMed] [Google Scholar]
- [8].Dinnella C, Morizet D, Masi C, Cliceri D, Depezay L, Appleton KM, et al. , Sensory determinants of stated liking for vegetable names and actual liking for canned vegetables: a cross-country study among European adolescents, Appetite 107 (2016) 339–347. [DOI] [PubMed] [Google Scholar]
- [9].Appleton KM, Hemingway A, Rajska J, Hartwell H, Repeated exposure and conditioning strategies for increasing vegetable liking and intake: systematic review and meta-analyses of the published literature, Am. J. Clin. Nutr 108 (2018) 842–856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Anzman-Frasca S, Savage JS, Marini ME, Fisher JO, Birch LL, Repeated exposure and associative conditioning promote preschool children’s liking of vegetables, Appetite 58 (2012) 543–553. [DOI] [PubMed] [Google Scholar]
- [11].Keller KL, Steinmann L, Nurse RJ, Tepper BJ, Genetic taste sensitivity to 6-n-propylthiouracil influences food preference and reported intake in preschool children, Appetite 38 (2002) 3–12. [DOI] [PubMed] [Google Scholar]
- [12].Bell KI, Tepper BJ, Short-term vegetable intake by young children classified by 6- n-propylthoiuracil bitter-taste phenotype, Am. J. Clin. Nutr 84 (2006) 245–251. [DOI] [PubMed] [Google Scholar]
- [13].Yackinous CA, Guinard J−X, Relation between PROP (6-n-propylthiouracil) taster status, taste anatomy and dietary intake measures for young men and women, Appetite 38 (2002) 201–209. [DOI] [PubMed] [Google Scholar]
- [14].Feeney EL, O’Brien SA, Scannell AGM, Markey A, Gibney ER, Genetic and environmental influences on liking and reported intakes of vegetables in Irish children, Food Qual. Prefer 32 (2014) 253–263. [Google Scholar]
- [15].Glendinning JI, Is the bitter rejection response always adaptive? Physiol. Behav 56 (1994) 1217–1227. [DOI] [PubMed] [Google Scholar]
- [16].Torregrossa A−M, Dearing MD, Nutritional toxicology of mammals: regulated intake of plant secondary compounds, Funct. Ecol 23 (2009) 48–56. [Google Scholar]
- [17].McArthur C, Haggerman AE, Robbins CT, Physiological strategies of mammalian herbivores against plant defenses, in: Paolo RT, Robbins CT (Eds.), Plant Defenses Against Mammalian Herbivory, CRC Press Inc, Ann Arbor MI, 1991, pp. 103–114. [Google Scholar]
- [18].Butler LG, Antinutritional effects of condensed and hydrolyzable tannins, Basic Life. Sci 59 (1992) 693–698. [DOI] [PubMed] [Google Scholar]
- [19].Mehansho H, Butler LG, Carlson DM, Dietary Tannins and Salivary Proline-Rich Proteins: interactions, Induction, and Defense Mechanisms, Annu. Rev. Nutr 7 (1987) 423–440. [DOI] [PubMed] [Google Scholar]
- [20].Cai K, Bennick A, Effect of salivary proteins on the transport of tannin and quercetin across intestinal epithelial cells in culture, Biochem. Pharmacol 72 (2006) 974–980. [DOI] [PubMed] [Google Scholar]
- [21].Shimada T, Salivary Proteins as a Defense Against Dietary Tannins, J. Chem. Ecol 32 (2006) 1149–1163. [DOI] [PubMed] [Google Scholar]
- [22].Glendinning JI, Effect of salivary proline-rich proteins on ingestive responses to tannic acid in mice, Chem. Senses 17 (1992) 1–12. [Google Scholar]
- [23].Juntheikki M−R, Comparison of tannin-binding proteins in saliva of Scandinavian and North American moose (Alces alces), Biochem. Syst. Ecol 24 (1996) 595–601. [Google Scholar]
- [24].Mehansho H, Ann DK, Butler LG, Rogler J, Carlson DM, Induction of proline-rich proteins in hamster salivary glands by isoproterenol treatment and an unusual growth inhibition by tannins, J. Biol. Chem 262 (1987) 12344–12350. [PubMed] [Google Scholar]
- [25].Bajec M, Pickering G, Astringency: mechanisms and perception, Crit. Rev. Food Sci. Nutr 48 (2008) 858–875. [DOI] [PubMed] [Google Scholar]
- [26].Ma W, Guo A, Zhang Y, Wang H, Liu Y, Li H, A review on astringency and bitterness perception of tannins in wine, Trends Food Sci. Technol 40 (2014) 6–19. [Google Scholar]
- [27].Dinnella C, Recchia A, Fia G, Bertuccioli M, Monteleone E, Saliva characteristics and individual sensitivity to phenolic astringent stimuli, Chemical Senses. 34 (2009) 295–304. [DOI] [PubMed] [Google Scholar]
- [28].Lee CA, Ismail B, Vickers ZM, The role of salivary proteins in the mechanism of astringency, J. Food Sci 77 (2012) C381–C3C7. [DOI] [PubMed] [Google Scholar]
- [29].Baxter NJ, Lilley TH, Haslam E, Williamson MP, Multiple interactions between polyphenols and a salivary proline-rich protein repeat result in complexation and precipitation†, Biochemistry 36 (1997) 5566–5577. [DOI] [PubMed] [Google Scholar]
- [30].Prinz J, Lucas P, Saliva tannin interactions, J. Oral. Rehabil 27 (2000) 991–994. [DOI] [PubMed] [Google Scholar]
- [31].Brossard N, Cai H, Osorio F, Bordeu E, Chen J, Oral” tribological study on the astringency sensation of red wines, J. Texture Stud 47 (2016) 392–402. [Google Scholar]
- [32].Ployon S, Morzel M, Belloir C, Bonnotte A, Bourillot E, Briand L, et al. , Mechanisms of astringency: structural alteration of the oral mucosal pellicle by dietary tannins and protective effect of bPRPs, Food Chem 253 (2018) 79–87. [DOI] [PubMed] [Google Scholar]
- [33].Morzel M, Truntzer C, Neyraud E, Brignot H, Ducoroy P, Lucchi G, et al. , Associations between food consumption patterns and saliva composition: specificities of eating difficulties children, Physiol. Behav 173 (2017) 116–123. [DOI] [PubMed] [Google Scholar]
- [34].Rodrigues L, Costa G, Cordeiro C, Pinheiro C, Amado F, Lamy E, Salivary proteome and glucose levels are related with sweet taste sensitivity in young adults, Food Nutr. Res 61 (2017), 1389208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Mounayar R, Morzel M, Brignot H, Tremblay-Franco M, Canlet C, Lucchi G, et al. , Salivary markers of taste sensitivity to oleic acid: a combined proteomics and metabolomics approach, Metabolomics 10 (2014) 688–696. [DOI] [PubMed] [Google Scholar]
- [36].Dsamou M, Palicki O, Septier C, Chabanet C, Lucchi G, Ducoroy P, et al. , Salivary Protein Profiles and Sensitivity to the Bitter Taste of Caffeine, Chem. Senses 37 (2012) 87–95. [DOI] [PubMed] [Google Scholar]
- [37].Morzel M, Chabanet C, Schwartz C, Lucchi G, Ducoroy P, Nicklaus S, Salivary protein profiles are linked to bitter taste acceptance in infants, Eur. J. Pediatr 173 (2014) 575–582. [DOI] [PubMed] [Google Scholar]
- [38].Méjean C, Morzel M, Neyraud E, Issanchou S, Martin C, Bozonnet S, et al. , Salivary composition is associated with liking and usual nutrient intake, PLoS ONE 10 (2015), e0137473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Morzel C. Chabanet, Schwartz C, Lucchi G, Ducoroy P, Nicklaus S, Salivary protein profiles are linked to bitter taste acceptance in infants, Eur. J. Pediatr 173 (2014) 575–582. [DOI] [PubMed] [Google Scholar]
- [40].Cabras T, Melis M, Castagnola M, Padiglia A, Tepper BJ, Messana I, et al. , Responsiveness to 6-n-Propylthiouracil (PROP) is associated with salivary levels of two specific basic proline-rich proteins in humans, PLoS ONE 7 (2012) e30962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Melis M, Aragoni MC, Arca M, Cabras T, Caltagirone C, Castagnola M, et al. , Marked increase in PROP taste responsiveness following oral supplementation with selected salivary proteins or their related free amino acids, PLoS ONE 8 (2013) 1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Stolle T, Grondinger F, Dunkel A, Meng C, Médard G, Kuster B, et al. , Salivary proteome patterns affecting human salt taste sensitivity, J. Agric. Food Chem 65 (2017) 9275–9286. [DOI] [PubMed] [Google Scholar]
- [43].Butterworth PJ, Warren FJ, Ellis PR, Human α-amylase and starch digestion: an interesting marriage, Starch-Stärke 63 (2011) 395–405. [Google Scholar]
- [44].Ferry A-LS, Mitchell JR, Hort J, Hill SE, Taylor AJ, Lagarrigue S, et al. , In-mouth amylase activity can reduce perception of saltiness in starch-thickened foods, J. Agric. Food Chem 54 (2006) 8869–8873. [DOI] [PubMed] [Google Scholar]
- [45].Stolle T, Grondinger F, Dunkel A, Meng C, Médard G, Kuster B, et al. , Salivary proteome patterns affecting human salt taste sensitivity, J. Agric. Food Chem 65 (2017) 9275–9286. [DOI] [PubMed] [Google Scholar]
- [46].Melis M, Aragoni MC, Arca M, Cabras T, Caltagirone C, Castagnola M, et al. , Marked increase in PROP taste responsiveness following oral supplementation with selected salivary proteins or their related free amino acids, PLoS ONE 8 (2013) e59810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Rodrigues L, Costa G, Cordeiro C, Pinheiro C, Amado F, Lamy E, Salivary proteome and glucose levels are related with sweet taste sensitivity in young adults, Food Nutr Res (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Stuchell RN, Mandel ID, A comparative study of salivary lysozyme in caries-resistant and caries-susceptible adults, J. Dent. Res 62 (1983) 552–554. [DOI] [PubMed] [Google Scholar]
- [49].Skopec MM, Hagerman AE, Karasov WH, Do salivary proline-rich proteins counteract dietary Hydrolyzable tannin in laboratory rats? J. Chem. Ecol 30 (2004) 1679–1692. [DOI] [PubMed] [Google Scholar]
- [50].Davis JD, The effectiveness of some sugars in stimulating licking behavior in the rat, Physiol. Behav 11 (1973) 39–45. [DOI] [PubMed] [Google Scholar]
- [51].Smith JC, Microstructure of the rat’s intake of food, sucrose and saccharin in 24-hour tests, Neurosci. Biobehav. Rev 24 (2000) 199–212. [DOI] [PubMed] [Google Scholar]
- [52].Katsukawa H, Ninomiya Y, Capsaicin induces cystatin s-like substances in submandibular saliva of the rat, J. Dent. Res 78 (1999) 1609–1616. [DOI] [PubMed] [Google Scholar]
- [53].Neyraud E, Sayd T, Morzel M, Dransfield E, Proteomic analysis of human whole and parotid Salivas following stimulation by different tastes, J. Proteome Res 5 (2006) 2474–2480. [DOI] [PubMed] [Google Scholar]
- [54].Quintana M, Palicki O, Lucchi G, Ducoroy P, Chambon C, Salles C, et al. , Short-term modification of human salivary proteome induced by two bitter tastants, urea and quinine, Chemosens Percept 2 (2009) 133–142. [Google Scholar]
- [55].Hagen KGT, Balys MM, Tabak LA, Melvin JE, Analysis of isoproterenol-induced changes in parotid gland gene expression, Physiol. Genomics 8 (2002) 107–114. [DOI] [PubMed] [Google Scholar]
- [56].Brockhoff A, Behrens M, Massarotti A, Appendino G, Meyerhof W, Broad tuning of the human bitter taste receptor hTAS2R46 to various sesquiterpene lactones, clerodane and labdane diterpenoids, strychnine, and denatonium, J. Agric. Food Chem 55 (2007) 6236–6243. [DOI] [PubMed] [Google Scholar]
- [57].Martin LE, Kay KE, Torregrossa AM, Bitter-induced salivary proteins increase detection threshold of quinine, but not sucrose, Chem Senses. (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [58].Martin LE, Nikonova LV, Kay KE, Torregrossa AM, Altering salivary protein profile can increase acceptance of a novel bitter diet, Appetite 136 (2019) 8–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Aravich PF, Sclafani A, Dietary preference behavior in rats fed bitter tasting quinine and sucrose octa acetate adulterated diets, Physiol. Behav 25 (1980) 157–160. [DOI] [PubMed] [Google Scholar]
- [60].Sclafani A, Aravich PF, Schwartz J, Hypothalamic hyperphagic rats overeat bitter sucrose octa acetate diets but not quinine diets, Physiol. Behav 22 (1979) 759–766. [DOI] [PubMed] [Google Scholar]
- [61].Martin LE, Nikonova LV, Kay K, Paedae AB, Contreras RJ, Torregrossa A−M, Salivary proteins alter taste-guided behaviors and taste nerve signaling in rat, Physiol. Behav 184 (2018) 150–161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [62].Torregrossa A−M, Nikonova L, Bales MB, Leal M. Villalobos, Smith JC, Contreras RJ, et al. , Induction of salivary proteins modifies measures of both orosensory and postingestive feedback during exposure to a tannic acid diet, PLoS ONE 9 (2014), e105232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [63].Batista TBD, Chaiben CL, Penteado CAS, Nascimento JMC, Ventura TMO, Dionizio A, et al. , Salivary proteome characterization of alcohol and tobacco dependents, Drug Alcohol Depend. 204 (2019), 107510. [DOI] [PubMed] [Google Scholar]
- [64].Dutta SK, Orestes M, Vengulekur S, Kwo P, Ethanol and human saliva: effect of chronic alcoholism on flow rate, composition, and epidermal growth factor, Am. J. Gastroenterol 87 (1992) 350–354. [PubMed] [Google Scholar]
- [65].Crawford CR, Running CA, Addition of chocolate milk to diet corresponds to protein concentration changes in human saliva, Physiol. Behav 225 (2020), 113080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [66].Martin LE, Kay KE, James KF, Torregrossa A−M, Altering salivary protein profile can decrease aversive oromotor responding to quinine in rats, Physiol. Behav 223 (2020), 113005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [67].St John S, Spector A, Behavioral discrimination between quinine and KCl is dependent on input from the seventh cranial nerve: implications for the functional roles of the gustatory nerves in rats, J. Neurosci 18 (1998) 4353–4362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [68].Spector AC, Kopka SL, Rats fail to discriminate quinine from denatonium: implications for the neural coding of bitter-tasting compounds, J. Neurosci 22 (2002) 1937–1941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [69].Martin LE, Kay KE, Torregrossa A−M, Rats are unable to discriminate quinine from diverse bitter stimuli, Am. J. Physiol.-Regul. Integrat. Comparat. Physiol 317 (2019) R793–R802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [70].Grill HJ, Norgren R, The taste reactivity test. I. Mimetic responses to gustatory stimuli in neurologically normal rats, Brain Res. 143 (1978) 263–279. [DOI] [PubMed] [Google Scholar]
- [71].Denny P, Hagen FK, Hardt M, Liao L, Yan W, Arellanno M, et al. , The proteomes of human parotid and submandibular/sublingual gland salivas collected as the ductal secretions, J. Proteome Res 7 (2008) 1994–2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [72].Luck G, Liao H, Murray NJ, Grimmer HR, Warminski EE, Williamson MP, et al. , Polyphenols, astringency and proline-rich proteins, Phytochemistry 37 (1994) 357–371. [DOI] [PubMed] [Google Scholar]
- [73].Lu Y, Bennick A, Interaction of tannin with human salivary proline-rich proteins, Arch. Oral Biol 43 (1998) 717–728. [DOI] [PubMed] [Google Scholar]
- [74].Pagès-Hélary S, Andriot I, Guichard E, Canon F, Retention effect of human saliva on aroma release and respective contribution of salivary mucin and α-amylase, Food Research Int. 64 (2014) 424–431. [DOI] [PubMed] [Google Scholar]
- [75].Patrikainen M, Pan P, Kulesskaya N, Voikar V, Parkkila S, The role of carbonic anhydrase VI in bitter taste perception: evidence from the Car6 –/– mouse model, J. Biomed. Sci 21 (2014) 82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [76].Martin LE, Nikonova LV, Kay K, Paedae AB, Contreras RJ, Torregrossa AM, Salivary proteins alter taste-guided behaviors and taste nerve signaling in rat, Physiol. Behav 184 (2018) 150–161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [77].Gurkan S, Secretions of von Ebner’s glands influence responses from taste buds in rat circumvallate papilla, Chem. Senses 13 (1988) 655–661. [Google Scholar]
- [78].da Costa G, Lamy E, Capela e Silva F, Andersen J, Sales Baptista E, Coelho A, Salivary amylase induction by tannin-enriched diets as a possible countermeasure against tannins, J. Chem. Ecol 34 (2008) 376–387. [DOI] [PubMed] [Google Scholar]
- [79].Schwartz M, Brignot H, Feron G, Hummel T, Zhu Y, von Koskull D, et al. , Role of human salivary enzymes in bitter taste perception, Food Chem 386 (2022), 132798. [DOI] [PubMed] [Google Scholar]
- [80].Karn RC, Chung AG, Laukaitis CM, Shared and unique proteins in human, mouse and rat saliva proteomes: footprints of functional adaptation, Proteomes 1 (2013) 275–289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [81].Cai K, Hagerman AE, Minto RE, Bennick A, Decreased polyphenol transport across cultured intestinal cells by a salivary proline-rich protein, Biochem. Pharmacol 71 (2006) 1570–1580. [DOI] [PubMed] [Google Scholar]
- [82].Frenkel ES, Ribbeck K, Salivary mucins in host defense and disease prevention, J. Oral. Microbiol 7 (2015) 29759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [83].Fábian TK, Hermann P, Beck A, Fejérdy P, Fábián G, Salivary defense proteins: their network and role in innate and acquired oral immunity, Int J Mol Sci 13 (2012) 4295–4320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [84].Canon F, Neiers F, Guichard E, Saliva and flavor perception: perspectives, J. Agric. Food Chem 66 (2018) 7873–7879. [DOI] [PubMed] [Google Scholar]
- [85].Salles C, Chagnon M−C, Feron G, Guichard E, Laboure H, Morzel M, et al. , In-mouth mechanisms leading to flavor release and perception, Crit. Rev. Food Sci. Nutr 51 (2010) 67–90. [DOI] [PubMed] [Google Scholar]
- [86].Ployon S, Morzel M, Canon F, The role of saliva in aroma release and perception, Food Chem. 226 (2017) 212–220. [DOI] [PubMed] [Google Scholar]
- [87].Van Ruth S, Roozen J, Influence of mastication and saliva on aroma release in a model mouth system, Food Chem 71 (2000) 339–345. [Google Scholar]
- [88].Guichard E, Repoux M, Qannari E, Labouŕe H, Feron G, Model cheese aroma perception is explained not only by in vivo aroma release but also by salivary composition and oral processing parameters, Food Funct. 8 (2017) 615–628. [DOI] [PubMed] [Google Scholar]
- [89].Piombino P, Genovese A, Esposito S, Moio L, Cutolo PP, Chambery A, et al. , Saliva from obese individuals suppresses the release of aroma compounds from wine, PLoS ONE 9 (2014) e85611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [90].Piombino P, Moio L, Genovese A, Orthonasal vs. retronasal: studying how volatiles’ hydrophobicity and matrix composition modulate the release of wine odorants in simulated conditions, Food Res. Int 116 (2019) 548–558. [DOI] [PubMed] [Google Scholar]
- [91].Lim J, Padmanabhan A, Retronasal olfaction in vegetable liking and disliking, Chem. Senses 38 (2012) 45–55. [DOI] [PubMed] [Google Scholar]
- [92].Duffy VB, Hayes JE, Sharafi M, Interactions between retronasal olfaction and taste influence vegetable liking and consumption: a psychophysical investigation, J. Agric. Food Res 2 (2020), 100044. [Google Scholar]
- [93].Barba FJ, Nikmaram N, Roohinejad S, Khelfa A, Zhu Z, Koubaa M, Bioavailability of glucosinolates and their breakdown products: impact of processing, Front Nutr 3 (2016) 24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [94].Frank D, Piyasiri U, Archer N, Jenifer J, Appelqvist I Influence of saliva on individual in-mouth aroma release from raw cabbage (Brassica oleracea var. capitata f. rubra L.) and links to perception, Heliyon 4 (2018) e01045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [95].Frank D, Piyasiri U, Archer N, Heffernan J, Poelman AA, In-mouth volatile production from brassica vegetables (Cauliflower) and associations with liking in an adult/child cohort, J. Agric. Food Chem 69 (2021) 11646–11655. [DOI] [PubMed] [Google Scholar]
- [96].Austin P, Suchar L, Robbins C, Hagerman A, Tannin-binding proteins in saliva of deer and their absence in saliva of sheep and cattle, J. Chem. Ecol 15 (1989) 1335–1347. [DOI] [PubMed] [Google Scholar]
- [97].Beasley DE, Koltz AM, Lambert JE, Fierer N, Dunn RR, The evolution of stomach acidity and its relevance to the human microbiome, PLoS ONE 10 (2015), e0134116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [98].Torregrossa A−M, Azzara AV, Dearing MD, Differential regulation of plant secondary compounds by herbivorous rodents, Funct. Ecol (2011), 10.1111/j.365-2435.011.01896.x. [DOI] [PubMed] [Google Scholar]
- [99].Torregrossa A−M, Azzara AV, Dearing MD, Testing the diet-breadth trade-off hypothesis: differential regulation of novel plant secondary compounds by a specialist and a generalist herbivore, Oecologia 168 (2012) 711–718. [DOI] [PubMed] [Google Scholar]
- [100].Hao S, Sternini C, Raybould HE, Role of CCK1 and Y2 receptors in activation of hindbrain neurons induced by intragastric administration of bitter taste receptor ligands. American journal of physiology, Regulat. Integrat. Comparat. Physiol 294 (2008) R33–RR8. [DOI] [PubMed] [Google Scholar]
- [101].Distel RA, Provenza FD, Experience early in life affects voluntary intake of blackbrush by goats, J. Chem. Ecol 17 (1991) 431–450. [DOI] [PubMed] [Google Scholar]
- [102].Sullivan SA, Birch LL, Infant dietary experience and acceptance of solid foods, Pediatrics 93 (1994) 271–277. [PubMed] [Google Scholar]
- [103].Ventura Alison K., Worobey J, Early influences on the development of food preferences, Current Biology 23 (2013) R401–R4R8. [DOI] [PubMed] [Google Scholar]
- [104].Asarian L, Geary N, Sex differences in the physiology of eating, Am. J. Physiol. Regul. Integr. Comp. Physiol 305 (2013) R1215–R1267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [105].Campbell PS, Ben-Aryeh H, Swanson KA, Differential distribution of an estrogen receptor in the submandibular and parotid salivary glands of female rats, Endocr. Res 16 (1990) 333–345. [DOI] [PubMed] [Google Scholar]