

Abstract
The use of bacteriophages, viruses that specifically infect bacteria, as antibiotics has become an area of great interest in recent years as the effectiveness of conventional antibiotics recedes. The detection of phage interactions with specific bacteria in a rapid and quantitative way is key for identifying phages of interest for novel antimicrobials. Outer membrane vesicles (OMVs) derived from Gram-negative bacteria can be used to make supported lipid bilayers (SLBs) and therefore in vitro membrane models that contain naturally occurring components of the bacterial outer membrane. In this study, we employed Escherichia coli OMV derived SLBs and use both fluorescent imaging and mechanical sensing techniques to show their interactions with T4 phage. We also integrate these bilayers with microelectrode arrays (MEAs) functionalized with the conducting polymer PEDOT:PSS and show that the pore forming interactions of the phages with the SLBs can be monitored using electrical impedance spectroscopy. To highlight our ability to detect specific phage interactions, we also generate SLBs using OMVs derived from Citrobacter rodentium, which is resistant to T4 phage infection, and identify their lack of interaction with the phage. The work presented here shows how interactions occurring between the phages and these complex SLB systems can be monitored using a range of experimental techniques. We believe this approach can be used to identify phages that work against bacterial strains of interest, as well as more generally to monitor any pore forming structure (such as defensins) interacting with bacterial outer membranes, and thus aid in the development of next generation antimicrobials.
Keywords: outer membrane vesicles, supported lipid bilayer, fluorescent microscopy, QCM-D, PEDOT:PSS, bacteriophage-membrane interactions
It is estimated that 25 000 people in Europe die every year from antibiotic resistant bacteria.1 This number is rising and it is projected that antibiotic resistance could be the cause of 10 million global deaths by 2050.2 The problem is compounded by the slowdown in effective development of antibiotic treatments. In the last 30 years, there has been a 90% reduction in novel antibiotics approved by the U.S. Food and Drug Administration (FDA).3 Out of this concerning landscape, dominated by the dual issues of increasing antibiotic resistance and decreasing novel antibiotic development, bacteriophage therapy has arisen as an alternative strategy. Bacteriophages, or phages for short, are viruses that specifically infect bacteria. They are the most abundant biological entities on earth, with an estimated 4.8 × 1031 phage particles in the whole biosphere.4 Phages are capable of infecting a specific range of bacterial strains depending on the components existing in the cell membrane of these bacteria. This specificity, coupled with the fact phages cannot infect eukaryotic cells, means that phage therapy can be a highly precise form of antibiotic treatment with minimal side effects.5
A hurdle to the usage of phage therapy becoming more prevalent is the screening of interactions between phages and bacteria. Currently, the main method used to identify and enumerate phage interactions with specific bacteria is the double agar overlay assay.6 This assay relies on mixing phage and bacterial cultures together with soft agar; if the phage can infect the bacteria, clear spots (plaques) appear on the bacterial lawn. Although this technique has been used for many years, it is time-consuming and not suited to rapid phage screening. Therefore, new methods are required that seek to provide a simpler way to precisely evaluate any given phage-host interaction.
The ability of a phage particle to infect a bacterial cell relies first and foremost on the interaction made with specific components in the outermost layer of the cell.7 In the case of Gram-negative bacteria, whose members make up the bulk of the antimicrobial resistant strains, this outermost layer is the outer membrane (OM).8 The OM consists of an asymmetric lipid bilayer with phospholipids in the inner leaflet and glycolipids, predominantly lipopolysaccharides (LPS), in the outer leaflet.9 The OM also contains outer membrane proteins (OMPs) that play a variety of roles in cellular functioning, notably in the permeation of specific molecules.10 Phages have tail structures which recognize components of the LPS, OMPs, or both,11 attaching themselves to the cell and initiating membrane penetration and genome injection. For instance, the well-characterized T4 phage binds to both OmpC and LPS on the surface of Escherichia coli prior to infection, while the T7 phage binds to only LPS molecules.7,12
Supported lipid bilayers (SLBs) have been used extensively as tools to investigate membrane interactions in an in vitro setting. The strength of the SLB platform lies in its ability to be integrated with a range of measurement techniques to probe the events occurring in these membrane mimic systems. However, SLBs generated from synthetic lipids do not faithfully represent the true structure of a cell membrane and this has been a major weakness. By using outer membrane vesicles (OMVs), extracellular vesicles of diameter 20–250 nm that are naturally produced by Gram-negative bacteria, it has been possible to generate SLBs that contain components (such as LPS and OMPs) that are found in the OM of a cell.13,14 These OM SLBs have been characterized using high resolution microscopy methods with their components elucidated by such techniques as structured illumination microscopy (SIM) and atomic force microscopy (AFM) where height profiles allow for both synthetic and native lipids components to be spatially mapped.15 Indeed, the presence of outer membrane components in addition to the retention of their native orientation has been confirmed with these super resolution microscopy techniques but also by engineering cleavable fluorescent tags. By tagging extracellular and intracellular facing proteins and forming SLBs using OMVs that express such proteins, Daniel et al. have determined the “parachute”-like mechanism of SLB formation and demonstrated that only the extracellular-facing tag is cleavable confirming the native orientation of membrane proteins in the SLB.13,16,17
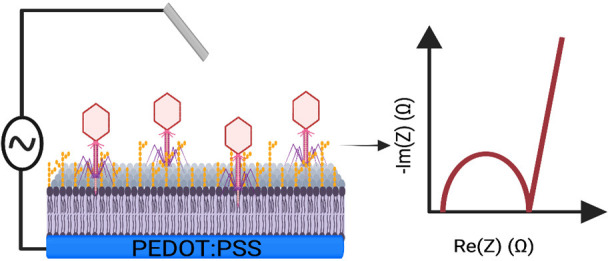
A key part of the attraction of the SLB platform is its ability to be integrated with a range of measurement techniques. One such measurement technique is electrochemical impedance spectroscopy (EIS), used to probe the resistance and capacitance characteristics of the SLB.18 SLBs can be generated upon microelectrode arrays (MEAs), and moreover these MEAs can be functionalized with biocompatible conducting polymers, such as the polymer poly(3,4-ethylenedioxythiophene) polystyrenesulfonate (PEDOT:PSS) to increase the device sensitivity. PEDOT:PSS, owing to its ion-to-electron mixed conductivity and cushioned mechanical properties, is an ideal material for interfacing with biological materials (such as SLBs), leading to the emergence of organic bioelectronic devices.19,20 OM SLBs on PEDOT:PSS coated MEAs have been probed by EIS previously; the actions of the antibiotics polymyxin B, bacitracin and meropenem with Escherichia coli OM SLBs were monitored using this technique.21 However, the OM SLB has not yet been investigated in the context of bacteriophage interactions.
In this study we use T4 phage, a phage that specifically infects E. coli cells, measuring its interactions with SLBs using optical and electrical techniques. The basis for the study relies on the interaction occurring between T4 phage particles and E. coli OM components, as shown schematically in Figure 1a. Briefly, the phage first attaches to the OM via interactions with LPS and OmpC components in the OM, and after this initial attachment the phage forms a pore in the OM through which it injects its DNA contents into the host cell. We start by showing the specificity of the infection with whole cells using Citrobacter rodentium, bacteria that are a closely related species to E. coli but crucially not susceptible to T4 infection, as a negative control. We generated OM SLBs from both E. coli and C. rodentium derived OMVs and assessed their interactions with T4 phage using Structured Illumination Microscopy (SIM). We further investigated their interaction using Quartz Crystal Microbalance with Dissipation monitoring (QCM-D), a technique that measures surface mass changes with high sensitivity. Finally, we integrated SLBs with PEDOT:PSS coated MEAs and performed EIS, showing that the specific interaction between the T4 phage and E. coli OM SLBs can be detected electrically. By combining the impedance signature with the optical data, we show here the ability of the OMV derived SLB as a quantitative phage screening platform that can play a role in furthering the prominence of phage therapy as a viable alternative to conventional antibiotic treatments.
Figure 1.

Characterization of T4 phage whole cell infection. (a) Schematic of the stages of T4 phage interaction with the OM of an E. coli cell. (bi) AFM image of a single T4 phage particle. Scale bar = 100 nm; height bar = 0–36 nm. (bii) TEM image of a single T4 phage particle. Scale bar = 100 nm. (ci) SIM imaging of E. coli cells after incubation with SYBR green stained T4 phage, imaged in the (left) 488 nm wavelength and (right) brightfield channels. (cii) SIM imaging of C. rodentium cells after incubation with SYBR green stained T4 phage, imaged in the (left) 488 nm wavelength and (right) brightfield channels. Scale bar = 10 μm. (d) Plaque assay in which the same concentration of T4 phage is mixed with E. coli and C. rodentium cells. The mixture is combined with soft agar and left to set overnight. The plaques seen in the E. coli are evidence of T4 infection. (e) T4 phage adsorption assay conducted with E. coli cells (multiplicity of infection (MOI) = 0.01). Samples were taken at regular time intervals; the number of free phages over time was enumerated by spot assay (n = 3) from which the concentration of free phage in PFU mL–1 was calculated.
Experimental Section
Bacterial Strains and Culture Conditions
The bacterial strains are shown in Table 1. E. coli and C. rodentium strains were grown at 37 °C in Lysogeny Broth (LB).
Table 1. Table of bacterial strains, description and reference/source in the study.
| Strains | Description | Reference/Source |
|---|---|---|
| Escherichia coli BL21 (DE3) | Lab strain | Invitrogen |
| Citrobacter rodentium DBS100 | Wild type (WT) strain | GPCS Lab Strain Collection; Popov et al.22 |
Phage Propagation
Ten microliters of serial dilutions of phages was mixed with 200 μL of overnight E. coli culture and 4 mL of molten top agar (0.35% agar) and then poured onto a LB agar plate (1.5% agar) and incubated overnight at 30 °C. Phage concentration was determined by counting plaques on the plate to give the concentration in plaque forming units per mL (PFU mL–1). To harvest the phages, the top agar was scraped off the plate, and the surface washed with 3 mL of LB. The wash was added to the harvested top agar and vortexed vigorously with 500 μL of chloroform for 2 min. Chloroform is used to extract T4 phages as per well established protocol.23 The agar mix was then centrifuged at 2220 g for 20 min at 4 °C. The supernatant was removed, and 10 μL of NaHCO3 chloroform was added and vortexed briefly. The phage lysate was stored at 4 °C for further use.
AFM Imaging of T4 Phage
AFM images were acquired in Scanasyst mode using ScanasystFluid+ probes (Bruker), with a nominal spring constant of 0.7 N m–1 and a resonant frequency of 150 kHz. Images were recorded at scan speeds of 1.5 Hz and tip–sample interaction forces between 200 and 300 pN. T4 phage sample was incubated on mica for 20 min before washing and imaging. The resulting images were analyzed using Nanoscope to evaluate the T4 phage dimensions.
SIM Imaging of T4 Phage
SYBR green 10 000×(Invitrogen) stock was diluted 1:10 in PBS. Phage were stained by incubating 1 μL of the 1:10 dilution with 200 μL of phage lysate for 15 min. Excess dye was removed through a Zeba spin desalting column (Thermo Scientific). To acquire structured illumination microscopy images, a × 60/1.2 NA water immersion lens (UPLSAPO 60XW, Olympus) focused the structured illumination pattern onto the sample, and the same lens was also used to capture the fluorescence emission light before imaging onto an sCMOS camera (C11440, Hamamatsu). The wavelengths used for excitation was 488 nm (iBEAM-SMART-488, Toptica) for imaging the SYBR green stained phage. Images were acquired using customized SIM software described previously.24
Optical Infection Assay
SYBR green stained T4 phages were added to overnight cultures of E. coli and C. rodentium in a 1:100 dilution ratio. The mix of phages and bacteria were incubated for 30 min at 37 °C, before being passed through a Zeba spin desalting column (Thermo Scientific). The mixture was centrifuged at 3000 g for 5 min and then resuspended in phosphate buffered saline (PBS, pH 7.4) for imaging. Two microliters of sample was deposited on a glass coverslip and an agarose pad was placed on top of the sample to prevent the bacteria from moving, as described previously. A glass coverslip was then placed on top of the agarose to prevent it from drying out. Images were acquired using SIM; the wavelength used for excitation was 488 nm (iBEAM-SMART-488, Toptica).
T4 Phage Adsorption Assay
The T4 phage adsorption assay was performed as described previously.25 In this case, 10 mL of overnight E. coli culture was used as the host, infected with 1 μL of ∼1010 PFU/mL T4 phage. As a no cell control, the same volume of phage was mixed with 10 mL of LB. The culture and the control were placed in an incubator at 37 °C, shaking at 150 rpm. Samples (100 μL) were taken at t = 0, 5, 15, 30, and 60 min and added to 900 μL of LB and 20 μL of NaHCO3 chloroform. The number of plaque forming units for each sample time point was determined by plaque assay as described in the phage propagation section above.
OMV Isolation from E. coli and C. rodentium
Five milliliters of LB broth was inoculated with E. coli cells and grown for 16–20 h. Two milliliters of the overnight culture was added to 200 mL of LB and allowed to incubate at 37 °C for ∼3 h until the OD600 of the culture was ∼1.5. The cells were then centrifuged (4000 g, 4 °C, 15 min) to remove cell debris, and the supernatant was collected. The supernatant was further passed through a 0.22 μm filter. The outer membrane vesicles (OMVs) were then isolated by ultracentrifugation (140 000 g, 4 °C) for 3 h (Beckman Coulter, Type 50.2 Ti Fixed-Angle Rotor) and the OMV pellets were resuspended in 250 μL of PBS supplemented with 2 mM MgCl2 solution. Finally, the OMV dispersion was centrifuged (16 000 g, 4 °C) for 30 min to remove any final contaminants such as flagella. The supernatant was collected and resuspended in 500 μL of PBS supplemented with 2 mM MgCl2 solution. The final OMV stocks were then aliquoted and stored at −80 °C for further experiments. OMVs were stored for up one month and any thawed OMVs were either used immediately or discarded.
For C. rodentium OMVs, the same protocol was followed except the culture was grown to an OD of ∼2.0 prior to the same centrifugation and filtration steps.
Preparation of POPC–PEG5kPE Synthetic Liposomes
POPC and PEG5kPE (Avanti) were mixed in a 99.5:0.5 molar ratio, with a nitrogen stream used to evaporate the chloroform, and the sample was further desiccated for 1 h in a vacuum. The lipids were then hydrated in PBS supplemented with 2 mM MgCl2 to give a final lipid concentration of 4 mg mL–1. Single unilamellar vesicles were made by lipid extrusion through a 50 nm pore sized polycarbonate membrane, and samples were stored for up to 2 weeks at 4 °C.
Nanoparticle Tracking Analysis (NTA)
Nanoparticle Tracking Analysis (NTA) was carried out using a Nanosight NS500 (Malvern Panalytical) fitted with an Electron Multiplying Charged Couple Device (EMCCD) camera configured with a 522 nm laser. Prior to analysis, samples were diluted (1:500) in PBS. Five ×60 s videos were recorded for each sample analyzed, with a temperature range of 20.8–21.5 °C and a camera level of 15. NTA 3.2 software was used to analyze the data with a detection threshold of 5.
Transmission Electron Microscopy (TEM)
For imaging OMVs, 10 μL of sample (concentration 1010 particles/mL) was negatively stained with 1% (w/v) uranyl acetate solution for 2 min at room temperature before being visualized with a Tecnai G2 80–200 keV transmission electron microscope, operating at 200 keV with images recorded with a bottom-mounted AMT CCD camera. For T4 phage imaging, 10 μL of sample (concentration 109 PFU mL–1) was used while for T4 and OMV combined imaging, phage (∼109 PFU mL–1) was mixed with E. coli OMVs (∼1010 PFU mL–1) in a 1:1 ratio, and left to incubate at room temperature for 20 min before the negative staining procedure was conducted.
Formation of Supported Lipid Bilayers on Glass Coverslips
Glass coverslips (Academy, 22 × 40 mm, 0.16–0.19 mm thick) were first cleaned with acetone and isopropanol in a 1:1 ratio. 100 μL of ∼1010 OMV particles mL–1 was added to the glass slide and allowed to incubate for 20 min before washing twice with PBS solution to remove excess unadhered OMVs. 100 μL of POPC–PEG5kPE liposomes were then added for 1 h to induce rupturing of the OMVs. The well was then washed again twice with PBS; the SLBs were then kept in PBS solution for imaging.
Characterization of SLBs Using Fluorescence Recovery after Photobleaching (FRAP)
Prior to analysis by fluorescence recovery after photobleaching (FRAP), OMVs must be fluorescently labeled. This was achieved by adding 1 μL of 0.36 mM R18 dye (Invitrogen) to 200 μL of OMV stock and sonicating for 15 min. A G25 spin column (GE healthcare) was used to remove unbound/excess R18 by centrifugation at 1500 g for 3 min at room temperature. Lipid bilayers were formed using the protocol outlined above. FRAP measurements were conducted using an inverted Zeiss LSM800 confocal microscope with a 10× objective lens. A 30 μm diameter bleaching spot was made, and recovery of the fluorescence intensity of this spot was measured over time relative to a 50 μm diameter reference spot. The data were analyzed using MATLAB, and the fluorescence recovery was modeled using a modified Bessel function as previously described.26 The model fit was used to extract the diffusion coefficient (D) according to the equation D = r2/4τ where r is the radius of the photobleached spot and τ is the characteristic diffusion time. The fit was also used to extract the mobile fraction (MF) according to the equation (IE – I0)/(II – I0), where IE is the final postbleach intensity value, I0 is the first postbleach intensity value, and II is the initial prebleach intensity value.
SIM Imaging of Phage SLB Interactions
SLBs were formed in the manner described above, where the lipids were stained with R18. SYBR green stained phages (concentration ∼1010 PFU mL–1) were added to the SLBs (E. coli/ C. rodentium/ POPC–PEG5kPE) and incubated for 20 min, washed with PBS before imaging using SIM. To acquire SIM images, a ×60/1.2 NA water immersion lens (UPLSAPO 60XW, Olympus) focused the structured illumination pattern onto the sample, and the same lens was also used to capture the fluorescence emission light before imaging onto an sCMOS camera (C11440, Hamamatsu). The wavelengths used for excitation were 561 nm (OBIS 561, Coherent) for the lipid bilayers and 488 nm (iBEAM-SMART-488, Toptica) for the SYBR green stained phage.
QCM-D Measurements
QCM measurements were performed using a Q-sense analyzer (QE401, Biolin Scientific). Piezoelectric silicon sensors were used for all the experiments. First the frequency and dissipation of energy signals were stabilized in PBS and the different lipid vesicles stocks in PBS were pumped into the chamber with a constant flow rate of 70 μL min–1 controlled by a peristaltic pump. All vesicles were given enough time to adsorb on the surface of the crystal. After that PBS 1X solution was used to wash unbound vesicles. When a stable frequency signal was established, phages were pumped in the chamber at a constant flow rate of 70 μL min–1 and incubated for about an hour on both synthetic lipid-functionalized sensors and outer-membrane lipids functionalized QCM sensors. Finally, the sensors were rinsed with PBS. The mass adsorbed per unit area was extracted from raw frequency and dissipation of energy data by modeling the third, fifth, seventh, and ninth overtones with the Kelvin–Voigt viscoelastic model.27−29 Q-tools, D-find, and Q-soft software were used for the modeling the raw data.30
Preparation of PEDOT:PSS Mixture
PEDOT:PSS dispersion (Heraeus) was mixed with 5% (v/v) ethylene glycol (EG) and 0.5% (v/v) dodecylbenzenesulfonic acid (DBSA) in order to enhance film formation and conductivity. 1% (v/v) 3-glycidoxypropyltrimethoxysilane (GOPS), which is a polymer cross-linking agent, was then added and the final mixture was sonicated for 10 min and sequentially filtered through a 0.8 μm and a 0.45 μm syringe filter prior to use.
Microelectrode Array Device Fabrication
PEDOT:PSS microelectrode arrays were fabricated using two different device architectures with both parylene C and silicon oxide used as electrode insulation layers. Parylene C devices are designed into arrays with circular electrodes with 450 μm in diameter or 200 μm by 200 μm square electrodes. To fabricate the devices, 4 in. glass wafers first were cleaned by sonication in acetone and then isopropanol for 15 min. The wafers are rinsed with DI water and baked 15 min at 150 °C. To pattern for contact tracks, a negative photoresist, AZ nLOF2035 (Microchemicals GmbH) was spun on the glass wafer with 3000 rpm for 45 s and exposed with UV light using mask aligner (Karl Suss MA/BA6). The photoresist was developed in AZ 726 MIF developer (MicroChemicals) developer for 28 s. Ti (5 nm)/Au (100 nm) layer as conductive tracks was deposited by e-beam evaporation on top of the wafer and the Ti–Au metal layer was lifted-off by soaking in Ni555 (Microchemicals GmbH) overnight. Prior to the deposition of 2 μm layer (sacrificial layer) of parylene C ((SCS), the wafer was soaked with 3% A174 (3-(trimethoxysilyl)propyl methacrylate) in ethanol solution (0.1% acetic acid in ethanol) for 60 s to promoteparylene C adhesion onto the wafer. An antiadhesive layer of Micro-90 in DI water (2% v/v solution) was spun (1000 rpm for 45 s), and then the second layer of 2 μm parylene C (SCS) was deposited. A layer of positive photoresist AZ 10XT (Microchemicals GmbH) was spun at 3000 rpm for 45 s, exposed to UV, and developed in AZ 726 MIF developer (MicroChemicals) for 6 min to pattern electrode areas. Reactive ion etching (Oxford 80 Plasmalab plus) opened the window for deposition of Clevios PH500 PEDOT:PSS (Heraeus). The PEDOT:PSS mixture (prepared as described above) was spin coated at 3000 rpm for 45 s. The device was baked at 90 °C for 1 min, and the sacrificial parylene C layer was peeled off. Finally, the sample was baked at 130 °C for 1 h. For devices fabricated using the silicon oxide method, glass wafers were first cleaned by heating in 9:1 sulfuric acid/hydrogen peroxide for 20 min at 120 °C. Metal contacts (50 nm Au between two 5 nm Ti adhesion layers) were e-beam evaporated and photolithographically patterned using a lift-off process. 230 nm SiO2 insulation layer was deposited with chemical vapor deposition and Au contacts were photolithographically patterned and then exposed using inductively coupled CHF3 plasma reactive ion etching. Wafers were treated with O2 plasma and then PEDOT:PSS layer was deposited via spin-coating the PH 1000 (Ossila) dispersion containing 5% v/v ethylene glycol and 1% v/v (3-glycidyloxypropyl)trimethyoxysilane at 2000 rpm for 2 min and baked at 140 °C for 30 min. 100 nm Ge hard mask was deposited using e-beam evaporation. Ge layer and underlying PEDOT:PSS were patterned using photolithography and inductively coupled plasma reactive ion etching with CF4 and O2, respectively. Devices were soaked in deionized water for 48 h to oxidize and remove the Ge layer. Electrodes were 500 × 500 μm square electrodes.
EIS Measurements
EIS was performed using a potentiostat (Autolab PG-STAT204) in a three-electrode configuration with Ag/AgCl and Pt electrodes being used as the reference and counter electrodes, respectively. Each PEDOT:PSS coated gold electrode in a single array was sequentially used as the working electrode. The AC current was recorded within the frequency range 50–100 000 Hz, with 10 data points per decade (equally spaced on a logarithmic scale). An AC voltage of 0.01 V and a DC voltage of 0 mV versus OCP were applied. For all experiments, LB was used as the electrolyte. Measurements were recorded for the baseline (ie. no bilayer), after bilayer formation (as described above), and after phage (concentration ∼1010 PFU mL–1) incubation with the bilayer for 20 min. Data were collected and analyzed using NOVA 2.1.3 software (Metrohm Autolab).
Results and Discussion
We first characterized the T4 phage particles using both AFM and TEM imaging (Figure 1bi, bii). The total length of the phage was 239 ± 13 nm, with the hexagonal head having length and width of 119 ± 9 nm and 105 ± 17 nm, respectively. These dimensions were in agreement with previously reported values for T4 phage.31E. coli and C. rodentium are both members of the attaching and effacing (A/E) family of bacteria and are genetically very similar, sharing large parts of their genomic sequences.32,33 Even though these two types of bacteria are similar, T4 phage was only able to infect E. coli and not C. rodentium as shown by SIM imaging of infectivity assays, since T4 phage particles could be stained with SYBR green (Figure S1). After incubation with stained T4 phage (Figure 1ci, 1cii), a significant difference was seen in the fluorescence of the two types of bacteria (558 ± 80 and 360 ± 5 AU for E. coli and C. rodentium respectively, P = 0.0006). Further evidence that T4 phage was only able to infect E. coli and not C. rodentium was shown by plaque assays, thus providing a means to exploit the difference in specificity to validate the eventual screening platform (Figure 1d). The area of clearance, or plaques, in the E. coli plate evidenced the phage infection. We investigated the kinetics of the T4 interaction with E. coli cells by conducting a phage adsorption assay (Figure 1e, Figure S2). By counting the number of plaques, and therefore measuring the PFU mL–1 over time (indicative of free, unbound phage), we observed the concentration significantly decreased from 2.4 ± 0.4 (× 104 PFU mL–1) to 0.9 ± 0.1 (× 104 PFU mL–1) after 15 min (P = 0.018). This was in contrast to the control case, where the concentration did not significantly change between 0 and 15 min (concentrations were 2.8 ± 0.1 and 2.7 ± 0.1 (x 104 PFU mL–1) respectively. Combining all this information, we concluded that the choice of bacteria and phage were appropriate for moving forward with developing the SLB screening platform.
The process of generating E. coli OM SLBs is depicted in Figure 2a. Briefly, OMVs isolated from an E. coli culture were induced to rupture and fuse, producing a complete SLB with the addition of POPC–PEG fusogenic liposomes. The presence of OMVs was confirmed using nanoparticle tracking analysis (NTA), a technique that measures the size distribution of particles based on their diffusivity.34 The average size of the particles was 186.6 ± 18.3 nm, in line with the expected size of E. coli OMVs.35 TEM was used to image the OMVs in the sample (Figure S3a), verifying their morphology in line with previous images of OMVs.36 The quality of the resulting SLB after vesicle fusion was evaluated using fluorescence recovery after photobleaching (FRAP). This technique relies on fluorescently staining OMVs with rhodamine-18 (R18) in the SLBs and then observing how, when a spot in the bilayer is bleached, fluorescence intensity recovers over time due to the lipid lateral mobility. As shown by Figure 2b, the E. coli SLB fluorescence recovered over time, indicating that the bilayer was complete and mobile. The diffusion coefficient (D) and mobile fraction (MF) values were 0.59 ± 0.04 μm2/s and 0.84 ± 0.04, respectively, similar to the values obtained by Hsia et al. for OM SLBs on glass.13 Since C. rodentium also produce OMVs,37 a crucial aspect of the screening procedure was the ability to form SLBs using the same process as for E. coli SLBs (Figure 2a). We confirmed the isolation of C. rodentium OMVs using NTA (Figure 2ci) and TEM (Figure S3b) with the OMVs having an average size of 134.6 ± 4.9 nm. SLBs were also generated, with the FRAP data showing the bilayers to have D and MF values of 0.39 ± 0.03 μm2/s and 1.02 ± 0.02 respectively.
Figure 2.

Formation of SLBs derived from bacterial OMVs. (a) Schematic showing the process of forming OM SLBs using E. coli OMVs. (bi) NTA characterization of the E. coli OMVs, with the main peak at 143 nm and the average size being 186.6 ± 18.3 nm. The blue shading represents the standard error of the mean. (bii) FRAP data for the E. coli OM SLB, showing the recovery of the bleached spot over time due to the mobility of the lipids. The corresponding diffusion coefficient and mobile fraction values are 0.59 ± 0.04 μm2/s and 0.84 ± 0.04, respectively. The diameter of the bleached spot is 30 μm. (ci) NTA characterization of the C. rodentium OMVs, showing a mean peak at 142 nm and an average size at 134.6 ± 4.9 nm. The blue shading represents the standard error of the mean. (cii) FRAP data for the C. rodentium SLB, where the diffusion coefficient and mobile fraction values are 0.39 ± 0.03 μm2/s and 1.02 ± 0.02, respectively.
Having established that SLBs could be generated using the two strains of bacterial OMVs, T4 phage binding to the E. coli and C. rodentium SLB systems was evaluated. SYBR green stained T4 phage incubated with R18 stained SLBs were imaged by SIM. With E. coli SLBs, spots of green fluorescence can be observed, indicating that the phage particles could interact with the SLB due to the presence of LPS and OmpC – the two components required for initial T4 phage binding at the start of the infection process in whole cells38 (Figure 3ai). Conversely, no binding was observed with the C. rodentium SLB (Figure S4); this shows the high specificity of the binding interaction since even though the bacterial strains are closely related, the outer membrane did not allow for phage binding. The fluorescence quantification verified this–the fluorescence in the 488 nm wavelength region was ∼3 times higher for the E. coli SLBs compared to the C. rodentium SLBs (Figure 3aii). POPC only SLBs were used as an additional negative control showing a similar lack of T4 phage binding (Figure S5).
Figure 3.

Optical and QCM-D screening of T4 phage interactions with SLBs. (ai) Panel of SIM images taken from E. coli and C. rodentium OM SLBs (stained with R18) incubated with SYBR green stained T4 phage. Scale bar = 2 μm. (aii) Measurement of the fluorescence intensity in the 488 nm wavelength region (measured in arbitrary units) for the 3 types of bilayers, including POPC SLB, after phage incubation. Error bars represent the standard error (two-way analysis of variance; *P ≤ 0.05; **P ≤ 0.01; ***P ≤ 0.001; n = 6). (b) QCM-D measurements for phage interaction with SLBs. Schematic of the sequence of events in the formation of the OM SLB and subsequent phage addition is shown. (1) Addition of OMVs, (2) addition of POPC–PEG and SLB formation, (3) addition of phage. Graph below shows mass changes over time due to phage addition to the OM SLB. Phage is added at ∼7800 s and washing commences at ∼10 600 s. (c) Corresponding QCM-D measurements for the control POPC SLB and subsequent phage addition. Zoom in of the portion of the graph related to the phage addition. Phage is added to the system at ∼1000 s and washing commences at ∼4400 s.
QCM-D is a technique that has been used extensively to monitor SLB formation and subsequent interactions and so we used it to verify the phage SLB interaction.39 This technique relies on measuring changes in frequency and dissipation at various overtones (these correspond to various penetration depths of the signal) on a SiO2 or gold sensor. The changes in frequency (Δf) relate to changes of mass absorbed on the sensor; a decrease in Δf denotes an increase in mass. Alternatively, changes in dissipation (ΔD) are related to a change in the rigidity where an increase in ΔD denotes a decrease in rigidity. Preliminary QCM-D measurements were conducted on gold sensors, where it is well established that a supported vesicle layer (SVL) is formed as opposed to an SLB, as the OMVs do not rupture well on unmodified gold surfaces.40,41 In this context, the specificity of phage to OMVs was demonstrated (Figure S5a,b). We then went on to monitor the formation of the E. coli OM SLB using QCM-D on a SiO2 sensor. Focusing first on the Δf over time (Figure S6ai), we observed that after OMVs were added, there was a decrease of ∼20 Hz due to the mass increase of the absorbed OMVs. This Δf is in line with previously reported values for Gram-negative bacteria OMVs adsorbed to the SiO2 sensor. After addition of the POPC–PEG liposomes, there appeared a sharp drop in Δf and concurrent increase in ΔD (Figure S6aii) as the liposomes adsorbed to the surface. These were followed by an increase in Δf and drop in ΔD, a signature that is associated with a release of coupled-mass water seen in these systems as the vesicles rupture and fuse to form the SLB.13,14,17Figure 3b shows how the mass of the adsorbed material on the sensor was extracted from the Δf and ΔD data presented in the Supporting Information. At ∼8000 s, the T4 phage was added and an increase in mass was observed, suggesting that the phage bound to the surface of the SLB. At ∼ 10 600 s, when buffer was washed over the sensor, there was a mass difference of ∼100 ng cm–2 after washing. This change in mass was indicative of permanent and specific phage binding–since the mass of a single phage virion is 194 MDa (when the capsid head is filled with DNA), this mass increase corresponded to the binding of ∼3.11 × 108 phage cm–2. To further confirm the specificity of T4 phage binding, a control QCM-D measurement was conducted with a POPC bilayer. The rapid decrease in Δf followed by an increase (occurring simultaneously with the opposite trends in ΔD) in the signal indicated that the POPC–PEG liposomes were adsorbed to the sensor surface, and then rapidly ruptured and fused to form an SLB with the associated release of coupled-mass water (Figure S6bi, S5bii). The Δf signal stabilized at -28 Hz which is in line with synthetic bilayer formation monitored by QCM-D on SiO2 sensors.42Figure 3c illustrates the mass changes extracted from the Δf and ΔD data. At ∼1000 s, phage was added to the system and an increase in mass was observed from ∼820 ng/cm2 to ∼1030 ng/cm2. However, when the sensor was washed with buffer in the same manner as for the E. coli SLB system, the mass dropped back to the pre phage incubation level of ∼820 ng/cm2. This indicated that the phage could not bind to the SLB specifically, in contrast to what was seen in the E. coli SLB case. The QCM-D experiments thus complemented the findings of the SIM experiments and expanded upon them to an extent since not only the phage-SLB interactions but also the bilayer formation itself were monitored to a high degree of precision.
A fundamental part of this study was the creation of a platform that provided a quick and quantitative evaluation of phage interaction with a given bilayer system. EIS can be used to measure the properties of an SLB integrated on a MEA device. EIS measures the compound effect of resistance and capacitance properties of an SLB by applying an alternating voltage over a frequency range and measuring the corresponding impedance (Z), which is a measure of resistance to current flow in the system. By decoupling the real part and imaginary part of SLB impedance, the membrane resistance and capacitance can be extracted. In our setup, when integrating the SLB with PEDOT:PSS coated MEA devices, we used a well-established equivalent circuit in which the bilayer is modeled as a resistor and capacitor in parallel16,43 (Figure 4a). The complex Z consists of an imaginary and real part; when these are plotted against each other for each frequency, a Nyquist plot is generated. The Nyquist plot is a useful way of expressing EIS data since the width of the semicircle portion of the graph denotes the membrane resistance.
Figure 4.

Electrical screening of phage interaction with SLBs. (a) Image of a MEA used for the EIS measurements. Scale bar = 5 mm. Each array consists of eight PEDOT:PSS coated electrodes upon which the SLB is formed. The impedance of the SLB is modeled using the equivalent circuit in the inset shown. (bi) Nyquist plot from a representative EIS recording for T4 phage incubated with E. coli SLBs. Measurements were taken before the SLB was formed, after the SLB formation, and after phage incubation. The decrease in the impedance is hypothesized to be the result of the phage particles binding and initiating pore formation in the OMV component of the SLB due to phage tail contraction and tube penetration. (bii) TEM images of T4 phage, comparing the morphology with and without interaction with E. coli OMVs. Scale bar = 100 nm. (biii) Nyquist plot from a representative EIS recording for T4 phage incubated with C. rodentium SLBs. Measurements were taken before the SLB was formed, after the SLB formation and after phage incubation. (c) Membrane resistance% change (i.e., change in extracted membrane resistance before and after phage incubation) on each electrode for three types of SLBs (E. coli SLB, C. rodentium SLB, POPC SLB). Error bars represent the standard error (two-way analysis of variance; *P ≤ 0.05; **P ≤ 0.01; ***P ≤ 0.001; n = 3).
When EIS was used to monitor the E. coli SLB formation on the MEA, the characteristic semicircle shape on the Nyquist plot appeared, denoting the presence of an additional RC element, indicating the presence of an SLB (Figure 3bi). The Bode plot also showed the presence of this insulating barrier (Figure S7a). The extracted resistance for the SLB was 251.6 ± 34.1 Ω cm2, in agreement with previously reported values for SLBs containing mammalian cell components.44 When T4 phage was added to the bilayer, a 14% reduction in the membrane resistance was observed (217.5 ± 30.1 Ω cm2)—we hypothesized that this was due to the interaction process occurring between the phage particles and the outer membrane components on the SLB surface. When T4 interacts with a cell, the initial process involves the adhesion of the phage to the outer membrane surface, followed by sheath contraction and subsequent tail tube penetration through the outer membrane.45 This penetration by the tail tube and effective pore formation would explain the reduction in resistance observed here—as the phage particles interact with the E. coli SLB, the attachment and penetration processes are initiated, leading to increased ion flow through the SLB and subsequent drop in membrane resistance. To investigate whether intact OMV components could trigger this membrane penetration process, we used TEM to image the phage bound to E. coli OMVs since these are the components of the SLB that would interact with the phage (Figure 4bii). The difference between phage particles bound and unbound to OMVs was stark—the bound phages exhibited contracted tails (about half of the uncontracted length), with this contraction driving the penetration of the tail tube through the OMV membrane.46 Moreover, the morphology of the capsid head was altered, which we postulate to be associated with the release of DNA through the tail tube. This supported the notion that the drop in impedance seen in the EIS readings was due to the phage tail tube penetration of the bilayer taking place.
C. rodentium SLBs were also generated on MEAs and probed by EIS. The Nyquist (Figure 4biii) and Bode (Figure S7b) plots confirmed the formation of the SLB, with the extracted membrane resistance being 230.2 ± 46.7 Ω cm2 which was very similar to the resistance observed for the E. coli SLB. Crucially, when phage was added here, a 1% decrease in membrane resistance was observed (227.9 ± 44.9 Ω cm2), indicating that T4 phage was unable to attach and subsequently penetrate these bilayers due to the absence of the necessary outer membrane components. Finally, POPC SLBs were used as an additional control (Figure S7ci, 7cii). EIS measurements showed the bilayer resistance to be 481.4 ± 168.8 Ω cm2 initially and 447.0 ± 149.7 Ω cm2 after phage addition. Due to electrode-to-electrode variation in bilayer quality, it was important to measure relative changes on each electrode individually to meaningfully compare between data sets. When relative changes in membrane resistance were measured, it became clear that the decrease in membrane resistance upon phage incubation was found only in the E. coli SLB case (Figure 4biv). Here, the membrane resistance decreased by 13.5 ± 1.2% while it decreased by only 0.1 ± 0.8% and 2.8 ± 2.5% in the C. rodentium and POPC SLBs, respectively. Therefore, the difference in membrane resistance changes between the three SLB cases provides a quantitative screening mechanism for detecting phage – SLB interactions.
The results here describe a multiparametric, quantitative readout of T4 phages interacting with OM SLBs. T4 phages belong to the myoviridae family of phages characterized by their long contractile tails. Podoviridae, exemplified by T7 phage, have short noncontractile tails and therefore exhibit a different mechanism of interaction with outer membranes and subsequent pore formation.46,47 On the other hand, the siphoviridae family of phages (e.g. λ phage) have long noncontractile tails and contain a central tape measure protein used to dictate tail length as well as to facilitate DNA injection through the outer membrane.48 We therefore envisage this platform to measure the interaction not only with myoviridae and OM SLBs but also with these other phage families to analyze whether the results we see depend on the mechanistic variation of phage infection. Perhaps there is a certain electrical signature that distinguishes these various mechanisms, in a similar manner to how EIS can be used to distinguish between different mechanisms of antibiotic interaction with membranes.21
In terms of advancing phage therapy, there is considerable interest in how variations on both the tail structures of phage, as well as the receptors present in the outer membrane, help or hinder an infection event. Recently, Zeng and Salmond were able to increase the host range specificity of lambda phage by expressing its receptor LamB in three different bacterial genera.49 The OM SLB platform could act as a complementary technique to those used in this study, due to the ease of generating OMVs derived from these various mutant bacteria. In a similar manner, rational engineering of the proteins in the phage tails responsible for receptor binding, the so-called phage receptor binding proteins (RBPs), has been used to program the host specificity of the phage. For instance, Yehl et al. used site directed mutagenesis to create a T3 phage library with different RBP sequences and thus different host range specificities.50 The SLB platform could help with evaluating whether a given phage tail mutation increases or decreases the propensity of the phage to interact with bacterial membranes via interpretation of the optical, mechanical sensing and electrochemical interaction signatures described in this paper.
Bacteria produce structures as part of their innate immune defense systems that are morphologically very similar to phage tail structures. These so-called tailosins are produced by bacteria to destroy closely related strains, and their mechanism of action relies on compromising the bacterial cell membrane.51 For instance, S5 pyocins destroy Pseudomonas aeruginosa strains by delivering a pore forming complex across the OM.52 By measuring interactions between tailocins and OM SLBs, this would provide information on the potential of the structures as antimicrobials against a given pathogenic bacteria. Similarly, defensins are phage tail like structures produced by mammalian immune system cells to destroy pathogenic bacteria. Studies on alpha defensins have shown their pore forming mechanism of action in synthetic lipid bilayers,53 and we hope to demonstrate the ability of our OMV derived SLB system to monitor the disruption effects of novel defensins and therefore extend the field of defensin research as an alternative to current antimicrobial strategies.
Conclusion
In conclusion, we have shown that SLBs containing the naturally occurring components of bacterial outer membranes provide a platform for specifically detecting bacteriophage interactions. After first confirming two types of bacteria, namely E. coli and C. rodentium, that can be used as the basis for a T4 phage screening platform, we showed that OM SLBs using OMVs isolated from these two bacteria can be generated. Optical imaging with SIM revealed that T4 was only able to interact with the E. coli SLB, and this interaction was further quantified using the highly sensitive QCM-D monitoring. The culmination of the study was the electrochemical monitoring of the differential phage interaction with the three types of SLBs, thus providing a quick and quantitative readout of the interaction (or lack thereof). In addition to this, the MEA platform has the potential to be integrated with a multiplexed, microfluidic setup which could prove pivotal in developing high throughput phage screening methods. Although in this study we sequentially use optical, mechanical sensing, and electrical techniques, future work will be based on simultaneous monitoring on a single multipurpose device. Overall, the study outlined here is the first time SLBs have been used to interact with phages, and the setup could prove important in easily and quantitatively determining hereto unknown phage-bacteria interactions. This will help accelerate the advent of phage therapy–a crucial avenue that needs to be explored further if our fight against antibiotic resistance is to prove successful.
Acknowledgments
All figures were created using BioRender.com. K.B. was funded by an Engineering and Physical Sciences Research Council (EPSRC)-DTP PhD studentship (project 2266415). R.M. acknowledges funding from the EPSRC Cambridge NanoDTC under award EP/S022953/1. Part of this work was performed at the Stanford Nano Shared Facilities (SNSF), supported by the National Science Foundation under award ECCS-2026822. Part of this work was performed in part in the nano@Stanford laboratories, which are supported by the National Science Foundation as part of the National Nanotechnology Coordinated Infrastructure under award ECCS-2026822. We acknowledge the Cambridge Royce facilities grant EP/P024947/1 and Sir Henry Royce Institute - recurrent grant EP/R00661X/1 for EQCM-D facility at Maxwell Centre, University of Cambridge. A.S. acknowledges funding from the European Union’s Horizon 2020 research and innovation program under the Marie Skłodowska-Curie grant, MultiStem (No. 895801). C.F.K. acknowledges funding from the EPSRC (EP/H018301/1 and EP/L015889/1), the Wellcome Trust (089703/Z/09/Z and 3-3249/Z/16/Z), the Medical Research Council (MR/K015850/1 and MR/K02292X/1), MedImmune, and Infinitus (China). I.M. acknowledges funding from the Royal Society (URF/R1/221795). R.M.O. acknowledges funding for this project sponsored by the Defense Advanced Research Projects Agency (DARPA) Army Research Office and accomplished under Cooperative Agreement W911NF-18-2-0152. The views and conclusions contained in this document are those of the authors and should not be interpreted as representing the official policies, either expressed or implied, of DARPA or the Army Research Office or the U.S. Government. The U.S. Government is authorized to reproduce and distribute reprints for Government purposes notwithstanding any copyright notation herein. Please note: for the purpose of open access, the author has applied a Creative Commons Attribution (CC BY) licence to any Author Accepted Manuscript version arising from this submission.
Supporting Information Available
The Supporting Information is available free of charge at https://pubs.acs.org/doi/10.1021/acsbiomaterials.3c00021.
Plates used for the adsorption assay and TEM images of OMVs used to make SLBs, a panel of SIM images for the fluorescent screening of phage interactions, and extra QCM-D and EIS data (PDF)
Author Contributions
K.B. grew and isolated OMVs, propagated phage samples, and performed FRAP, EIS, and plaque and adsorption assay measurements. G.S. and R. Monson provided bacterial glycerol stocks and T4 phage. Z.L., J.T., and A. Salleo designed and fabricated PEDOT:PSS microelectrode arrays. A. Savva performed QCM-D measurements and analyzed these data. K.B., C.F.K., and I.M. performed SIM measurements. R. McCoy performed NTA measurements. K.B. and R. McCoy analyzed the data, wrote and revised the manuscript with R. Monson and R.M.O. R. Monson and R.M.O. supervised the study and provided funding support. All authors approved the manuscript prior to submission.
The authors declare no competing financial interest.
Supplementary Material
References
- Laxminarayan R.; Duse A.; Wattal C.; Zaidi A. K. M.; Wertheim H. F. L.; Sumpradit N.; Vlieghe E.; Hara G. L.; Gould I. M.; Goossens H.; Greko C.; So A. D.; Bigdeli M.; Tomson G.; Woodhouse W.; Ombaka E.; Peralta A. Q.; Qamar F. N.; Mir F.; Kariuki S.; Bhutta Z. A.; Coates A.; Bergstrom R.; Wright G. D.; Brown E. D.; Cars O. Antibiotic Resistance—the Need for Global Solutions. Lancet Infectious Diseases 2013, 13 (12), 1057–1098. 10.1016/S1473-3099(13)70318-9. [DOI] [PubMed] [Google Scholar]
- Luepke K. H.; Suda K. J.; Boucher H.; Russo R. L.; Bonney M. W.; Hunt T. D.; Mohr J. F. Past, Present, and Future of Antibacterial Economics: Increasing Bacterial Resistance, Limited Antibiotic Pipeline, and Societal Implications. Pharmacotherapy 2017, 37 (1), 71–84. 10.1002/phar.1868. [DOI] [PubMed] [Google Scholar]
- Spellberg B. Future of Antibiotics 2014, 18, 228. 10.1186/cc13948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cobián Güemes A. G.; Youle M.; Cantú V. A.; Felts B.; Nulton J.; Rohwer F. Viruses as Winners in the Game of Life. Annu. Rev. Virol. 2016, 3 (1), 197–214. 10.1146/annurev-virology-100114-054952. [DOI] [PubMed] [Google Scholar]
- Kutter E. M.; Kuhl S. J.; Abedon S. T. Re-Establishing a Place for Phage Therapy in Western Medicine. Future Microbiology 2015, 10 (5), 685–688. 10.2217/fmb.15.28. [DOI] [PubMed] [Google Scholar]
- Ács N.; Gambino M.; Brøndsted L. Bacteriophage Enumeration and Detection Methods. Front. Microbiol. 2020, 11, 594868. 10.3389/fmicb.2020.594868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bertozzi Silva J.; Storms Z.; Sauvageau D. Host Receptors for Bacteriophage Adsorption. FEMS Microbiology Letters 2016, 363, fnw002. 10.1093/femsle/fnw002. [DOI] [PubMed] [Google Scholar]
- Breijyeh Z.; Jubeh B.; Karaman R. Resistance of Gram-Negative Bacteria to Current Antibacterial Agents and Approaches to Resolve It. Molecules 2020, 25 (6), 1340. 10.3390/molecules25061340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kamio Y.; Nikaido H. Outer Membrane of Salmonella Typhimurium: Accessibility of Phospholipid Head Groups to Phospholipase C and Cyanogen Bromide Activated Dextran in the External Medium. Biochemistry 1976, 15 (12), 2561–2570. 10.1021/bi00657a012. [DOI] [PubMed] [Google Scholar]
- Vergalli J.; Bodrenko I. V.; Masi M.; Moynié L.; Acosta-Gutiérrez S.; Naismith J. H.; Davin-Regli A.; Ceccarelli M.; van den Berg B.; Winterhalter M.; Pagès J.-M. Porins and Small-Molecule Translocation across the Outer Membrane of Gram-Negative Bacteria. Nat. Rev. Microbiol 2020, 18 (3), 164–176. 10.1038/s41579-019-0294-2. [DOI] [PubMed] [Google Scholar]
- Chen P.; Sun H.; Ren H.; Liu W.; Li G.; Zhang C. LamB, OmpC, and the Core Lipopolysaccharide of Escherichia Coli K-12 Function as Receptors of Bacteriophage Bp7. J. Virol 2020, 94 (12), e00325-20 10.1128/JVI.00325-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- González-García V. A.; Bocanegra R.; Pulido-Cid M.; Martín-Benito J.; Cuervo A.; Carrascosa J. L. Characterization of the Initial Steps in the T7 DNA Ejection Process. Bacteriophage 2015, 5 (3), e1056904 10.1080/21597081.2015.1056904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsia C.-Y.; Chen L.; Singh R. R.; DeLisa M. P.; Daniel S. A Molecularly Complete Planar Bacterial Outer Membrane Platform. Sci. Rep. 2016, 6 (1), 32715. 10.1038/srep32715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mohamed Z.; Shin J.-H.; Ghosh S.; Sharma A. K.; Pinnock F.; Bint E Naser Farnush S.; Dorr T.; Daniel S. Clinically Relevant Bacterial Outer Membrane Models for Antibiotic Screening Applications. ACS Infectious Diseases 2021, 7, 2707. 10.1021/acsinfecdis.1c00217. [DOI] [PubMed] [Google Scholar]
- Bali K.; Mohamed Z.; Scheeder A.; Pappa A.-M.; Daniel S.; Kaminski C. F.; Owens R. M.; Mela I. Nanoscale Features of Tunable Bacterial Outer Membrane Models Revealed by Correlative Microscopy. Langmuir 2022, 38 (29), 8773–8782. 10.1021/acs.langmuir.2c00628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu H.-Y.; Pappa A.-M.; Pavia A.; Pitsalidis C.; Thiburce Q.; Salleo A.; Owens R. M.; Daniel S. Self-Assembly of Mammalian Cell Membranes on Bioelectronic Devices with Functional Transmembrane Proteins. Langmuir 2020, 36, 7325. 10.1021/acs.langmuir.0c00804. [DOI] [PubMed] [Google Scholar]
- Richards M. J.; Hsia C.-Y.; Singh R. R.; Haider H.; Kumpf J.; Kawate T.; Daniel S. Membrane Protein Mobility and Orientation Preserved in Supported Bilayers Created Directly from Cell Plasma Membrane Blebs. Langmuir 2016, 32 (12), 2963–2974. 10.1021/acs.langmuir.5b03415. [DOI] [PubMed] [Google Scholar]
- Bali K.; Guffick C.; McCoy R.; Lu Z.; Kaminski C. F.; Mela I.; Owens R. M.; van Veen H. W. Biosensor for Multimodal Characterization of an Essential ABC Transporter for Next-Generation Antibiotic Research. ACS Appl. Mater. Interfaces 2023, 15 (10), 12766–12776. 10.1021/acsami.2c21556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu Z.; van Niekerk D.; Savva A.; Kallitsis K.; Thiburce Q.; Salleo A.; Pappa A.-M.; Owens R. M. Understanding Electrochemical Properties of Supported Lipid Bilayers Interfaced with Organic Electronic Devices. J. Mater. Chem. C 2022, 10 (20), 8050–8060. 10.1039/D2TC00826B. [DOI] [Google Scholar]
- Zhang Y.; Inal S.; Hsia C.-Y.; Ferro M.; Ferro M.; Daniel S.; Owens R. M. Supported Lipid Bilayer Assembly on PEDOT:PSS Films and Transistors. Adv. Funct. Mater. 2016, 26 (40), 7304–7313. 10.1002/adfm.201602123. [DOI] [Google Scholar]
- Ghosh S.; Mohamed Z.; Shin J.-H.; Bint E Naser S. F.; Bali K.; Dörr T.; Owens R. M.; Salleo A.; Daniel S. Impedance Sensing of Antibiotic Interactions with a Pathogenic E. Coli Outer Membrane Supported Bilayer. Biosens. Bioelectron. 2022, 204, 114045. 10.1016/j.bios.2022.114045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Popov G.; Fiebig-Comyn A.; Shideler S.; Coombes B. K.; Savchenko A. Complete Genome Sequence of Citrobacter Rodentium Strain DBS100. Microbiol Resour Announc 2019, 8 (24), e00421-19 10.1128/MRA.00421-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bourdin G.; Schmitt B.; Marvin Guy L.; Germond J.-E.; Zuber S.; Michot L.; Reuteler G.; Brüssow H. Amplification and Purification of T4-Like Escherichia Coli Phages for Phage Therapy: From Laboratory to Pilot Scale. Appl. Environ. Microbiol. 2014, 80 (4), 1469–1476. 10.1128/AEM.03357-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Young L. J.; Ströhl F.; Kaminski C. F. A Guide to Structured Illumination TIRF Microscopy at High Speed with Multiple Colors. J. Vis Exp 2016, (111), 53988. 10.3791/53988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petty N. K.; Foulds I. J.; Pradel E.; Ewbank J. J.; Salmond G. P. C. A Generalized Transducing Phage (ΦIF3) for the Genomically Sequenced Serratia Marcescens Strain Db11: A Tool for Functional Genomics of an Opportunistic Human Pathogen. Microbiology 2006, 152 (6), 1701–1708. 10.1099/mic.0.28712-0. [DOI] [PubMed] [Google Scholar]
- Soumpasis D. M. Theoretical Analysis of Fluorescence Photobleaching Recovery Experiments. Biophys. J. 1983, 41 (1), 95–97. 10.1016/S0006-3495(83)84410-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mohamed Z.; Shin J. H.; Ghosh S.; Sharma A. K.; Pinnock F.; Bint E Naser Farnush S.; Dörr T.; Daniel S. Clinically Relevant Bacterial Outer Membrane Models for Antibiotic Screening Applications. ACS Infectious Diseases 2021, 7 (9), 2707–2722. 10.1021/acsinfecdis.1c00217. [DOI] [PubMed] [Google Scholar]
- Kaufmann S.; Ilg K.; Mashaghi A.; Textor M.; Priem B.; Aebi M.; Reimhult E. Supported Lipopolysaccharide Bilayers. Langmuir 2012, 28 (33), 12199–12208. 10.1021/la3020223. [DOI] [PubMed] [Google Scholar]
- Cho N.-J.; Frank C. W.; Kasemo B.; Höök F. Quartz Crystal Microbalance with Dissipation Monitoring of Supported Lipid Bilayers on Various Substrates. Nat. Protoc. 2010, 5 (6), 1096–1106. 10.1038/nprot.2010.65. [DOI] [PubMed] [Google Scholar]
- Savva A.; Hallani R.; Cendra C.; Surgailis J.; Hidalgo T. C.; Wustoni S.; Sheelamanthula R.; Chen X.; Kirkus M.; Giovannitti A.; Salleo A.; McCulloch I.; Inal S. Balancing Ionic and Electronic Conduction for High-Performance Organic Electrochemical Transistors. Adv. Funct. Mater. 2020, 30 (11), 1907657. 10.1002/adfm.201907657. [DOI] [Google Scholar]
- Leiman P. G.; Kanamaru S.; Mesyanzhinov V. V.; Arisaka F.; Rossmann M. G. Structure and Morphogenesis of Bacteriophage T4. Cellular and Molecular Life Sciences (CMLS) 2003, 60 (11), 2356–2370. 10.1007/s00018-003-3072-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magaziner S. J.; Zeng Z.; Chen B.; Salmond G. P. C.. The Prophages of Citrobacter Rodentium Represent a Conserved Family of Horizontally Acquired Mobile Genetic Elements Associated with Enteric Evolution towards Pathogenicity. J. Bacteriol. 2019, 201 ( (9), ), 10.1128/JB.00638-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deng W.; de Hoog C. L.; Yu H. B.; Li Y.; Croxen M. A.; Thomas N. A.; Puente J. L.; Foster L. J.; Finlay B. B. A Comprehensive Proteomic Analysis of the Type III Secretome of Citrobacter Rodentium. J. Biol. Chem. 2010, 285 (9), 6790–6800. 10.1074/jbc.M109.086603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Filipe V.; Hawe A.; Jiskoot W. Critical Evaluation of Nanoparticle Tracking Analysis (NTA) by NanoSight for the Measurement of Nanoparticles and Protein Aggregates. Pharm. Res. 2010, 27 (5), 796–810. 10.1007/s11095-010-0073-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jan A. T. Outer Membrane Vesicles (OMVs) of Gram-Negative Bacteria: A Perspective Update. Front. Microbiol. 2017, 8, 1053. 10.3389/fmicb.2017.01053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kulkarni H. M.; Nagaraj R.; Jagannadham M. V. Protective Role of E. Coli Outer Membrane Vesicles against Antibiotics. Microbiological Research 2015, 181, 1–7. 10.1016/j.micres.2015.07.008. [DOI] [PubMed] [Google Scholar]
- Sinha A.; Nyongesa S.; Viau C.; Gruenheid S.; Veyrier F. J.; Le Moual H.. PmrC (EptA) and CptA Negatively Affect Outer Membrane Vesicle Production in Citrobacter Rodentium. J. Bacteriol. 2019, 201 ( (7), ), 10.1128/JB.00454-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Montag D.; Hashemolhosseini S.; Henning U. Receptor-Recognizing Proteins of T-Even Type Bacteriophages: The Receptor-Recognizing Area of Proteins 37 of Phages T4 TuIa and TuIb. J. Mol. Biol. 1990, 216 (2), 327–334. 10.1016/S0022-2836(05)80324-9. [DOI] [PubMed] [Google Scholar]
- Cho N.-J.; Frank C. W.; Kasemo B.; Höök F. Quartz Crystal Microbalance with Dissipation Monitoring of Supported Lipid Bilayers on Various Substrates. Nat. Protoc 2010, 5 (6), 1096–1106. 10.1038/nprot.2010.65. [DOI] [PubMed] [Google Scholar]
- Choi S.-E.; Greben K.; Wördenweber R.; Offenhäusser A. Positively Charged Supported Lipid Bilayer Formation on Gold Surfaces for Neuronal Cell Culture. Biointerphases 2016, 11 (2), 021003. 10.1116/1.4945306. [DOI] [PubMed] [Google Scholar]
- Reimhult E.; Höök F.; Kasemo B. Intact Vesicle Adsorption and Supported Biomembrane Formation from Vesicles in Solution: Influence of Surface Chemistry, Vesicle Size, Temperature, and Osmotic Pressure. Langmuir 2003, 19 (5), 1681–1691. 10.1021/la0263920. [DOI] [Google Scholar]
- Renner L. D.; Weibel D. B. MinD and MinE Interact with Anionic Phospholipids and Regulate Division Plane Formation in Escherichia Coli. J. Biol. Chem. 2012, 287 (46), 38835–38844. 10.1074/jbc.M112.407817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pappa A.-M.; Liu H.-Y.; Traberg-Christensen W.; Thiburce Q.; Savva A.; Pavia A.; Salleo A.; Daniel S.; Owens R. M. Optical and Electronic Ion Channel Monitoring from Native Human Membranes. ACS Nano 2020, 14, 12538. 10.1021/acsnano.0c01330. [DOI] [PubMed] [Google Scholar]
- Tang T.; Savva A.; Traberg W. C.; Xu C.; Thiburce Q.; Liu H.-Y.; Pappa A.-M.; Martinelli E.; Withers A.; Cornelius M.; Salleo A.; Owens R. M.; Daniel S. Functional Infectious Nanoparticle Detector: Finding Viruses by Detecting Their Host Entry Functions Using Organic Bioelectronic Devices. ACS Nano 2021, 15 (11), 18142–18152. 10.1021/acsnano.1c06813. [DOI] [PubMed] [Google Scholar]
- Hu B.; Margolin W.; Molineux I. J.; Liu J. Structural Remodeling of Bacteriophage T4 and Host Membranes during Infection Initiation. Proc. Natl. Acad. Sci. U. S. A. 2015, 112 (35), E4919-E4928 10.1073/pnas.1501064112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aksyuk A. A.; Leiman P. G.; Kurochkina L. P.; Shneider M. M.; Kostyuchenko V. A.; Mesyanzhinov V. V.; Rossmann M. G. The Tail Sheath Structure of Bacteriophage T4: A Molecular Machine for Infecting Bacteria. EMBO J. 2009, 28 (7), 821–829. 10.1038/emboj.2009.36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cuervo A.; Fàbrega-Ferrer M.; Machón C.; Conesa J. J.; Fernández F. J.; Pérez-Luque R.; Pérez-Ruiz M.; Pous J.; Vega M. C.; Carrascosa J. L.; Coll M. Structures of T7 Bacteriophage Portal and Tail Suggest a Viral DNA Retention and Ejection Mechanism. Nat. Commun. 2019, 10 (1), 3746. 10.1038/s41467-019-11705-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arnaud C.-A.; Effantin G.; Vivès C.; Engilberge S.; Bacia M.; Boulanger P.; Girard E.; Schoehn G.; Breyton C. Bacteriophage T5 Tail Tube Structure Suggests a Trigger Mechanism for Siphoviridae DNA Ejection. Nat. Commun. 2017, 8 (1), 1953. 10.1038/s41467-017-02049-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeng Z.; Salmond G. P. C. Bacteriophage Host Range Evolution through Engineered Enrichment Bias, Exploiting Heterologous Surface Receptor Expression. Environmental Microbiology 2020, 22 (12), 5207–5221. 10.1111/1462-2920.15188. [DOI] [PubMed] [Google Scholar]
- Yehl K.; Lemire S.; Yang A. C.; Ando H.; Mimee M.; Torres M. D. T.; de la Fuente-Nunez C.; Lu T. K. Engineering Phage Host-Range and Suppressing Bacterial Resistance through Phage Tail Fiber Mutagenesis. Cell 2019, 179 (2), 459–469. 10.1016/j.cell.2019.09.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carim S.; Azadeh A. L.; Kazakov A. E.; Price M. N.; Walian P. J.; Lui L. M.; Nielsen T. N.; Chakraborty R.; Deutschbauer A. M.; Mutalik V. K.; Arkin A. P. Systematic Discovery of Pseudomonad Genetic Factors Involved in Sensitivity to Tailocins. ISME J. 2021, 15 (8), 2289–2305. 10.1038/s41396-021-00921-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Behrens H. M.; Lowe E. D.; Gault J.; Housden N. G.; Kaminska R.; Weber T. M.; Thompson C. M. A.; Mislin G. L. A.; Schalk I. J.; Walker D.; et al. Pyocin S5 Import into Pseudomonas Aeruginosa Reveals a Generic Mode of Bacteriocin Transport. mBio 2020, 11 (2), e03230-19 10.1128/mBio.03230-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y.; Lu W.; Hong M. Membrane-Bound Structure and Topology of a Human Alpha Defensin Indicates A Dimer Pore Mechanism for Membrane Disruption. Biochemistry 2010, 49 (45), 9770–9782. 10.1021/bi101512j. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.