

Abstract
One of the primary methods by which the gut microbiome interacts with its host is through the interactions that occur through the production of the metabolites produced, either directly, or indirectly, through microbial metabolism. Decades of research has demonstrated that these metabolic products play a vital role in human health, either for its benefit or detriment. This review article highlights the main metabolites produced by the interactions between diet and the gut microbiome, bile acids and the gut microbiome, and products produced by the gut microbiome alone. Additionally, this article reviews the literature on the effects that these metabolites play on human health.
Keywords: Gut metabolites, microbiome, short chain fatty acids, trimethylamine-N-oxide, bile acids
Introduction
The gastrointestinal tract is home to trillions of microbes which collectively contain more genes than the human genome and produce various metabolites that affect the human host [1]. Decades of research have come to demonstrate that these metabolic end-products play a vital role in humans, affecting components in cardiovascular, neurologic, and metabolic health [2]. Collectively, these various metabolic end-products produced by the microbiome are both synthesized and utilized in various ways [3]. The objective of this review article is to highlight three main mechanisms by which the core metabolites are synthesized and discuss their function and impact on human health.
Metabolites produced by the interaction between the diet and gut microbiome of the host
Short chain fatty acids
Gut microbes feed on the diet of the host and through this interaction metabolic end-products including short chain fatty acids (acetate, propionate, and butyrate), trimethylamine-N-oxide (TMAO), and a litany of tryptophan catabolites are produced.
Fermentable dietary fiber that pass through the small intestine without being digested or absorbed, are utilized by the bacteria in the colon [4]. They undergo fermentation that results in the production of acetate, propionate, butyrate, and gases (H2 and CO2). The production occurs in a 60:20:20 ratio for acetate, propionate, and butyrate respectively with anywhere from 90 to 99% being used in the gut [5,6]. Collectively, all of the SCFA can bind to the G-protein coupled receptor 41 and 43 (GPR41/43), which are expressed on the enteroendocrine L-cells to exert their metabolic effects [7,8]. Binding to GPR41/43 induces secretion of gut hormones glucagon-like peptide-1 (GLP-1) and peptide YY which can increase energy expenditure, increases fat oxidation, reduces pro-inflammatory cytokines, and decreases appetite [9,10]. These receptors are known to be found in the most metabolically active tissues including muscle, liver, and adipose tissue, indicating the direct role SCFA play in systemic energy metabolism [2,10]. Table 1 highlights the effects of the three short chain fatty acids.
Table 1.
Effects of the various short chain fatty acids
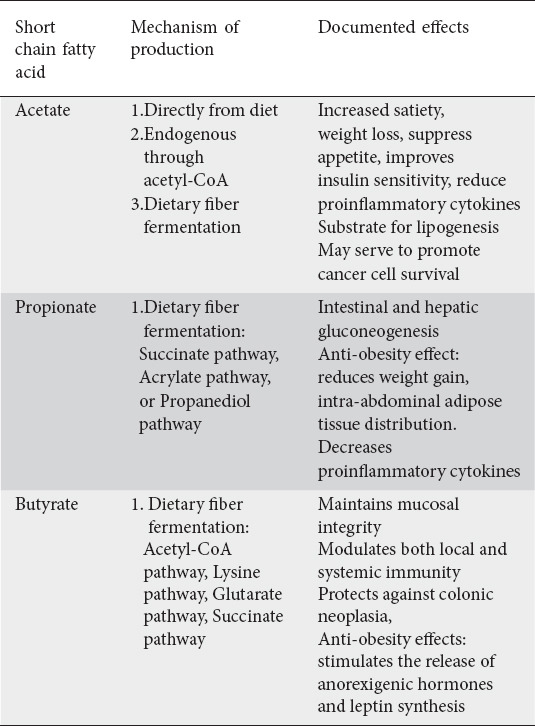
Acetate may be generated by three pathways either by direct consumption of acetate-containing foods, through endogenous production in tissues through acetyl-CoA, or by the fermentation of dietary fiber, particularly acetogenic fibers which include inulin and galacto-oligosaccharides [11,12]. Of the various SCFAs, acetate is the most abundant, being produced in a 60:20:20 ratio in the colon for acetate, propionate, and butyrate respectively. Peripherally, this ratio becomes 180:5:1, indicating that a majority of propionate and butyrate are utilized at the site of production [4]. Interestingly, this ratio is altered for hosts that consume western-diets rich on fat and low in fiber with lower amounts of peripheral acetate [13]. This is a notable finding given the role acetate plays in metabolic diseases and in particular with type 2 diabetes mellitus (T2DM). Controversy, however, still surrounds the health benefits of acetate. While some studies associate acetate with increased satiety and weight loss through its interaction with GPR41/43, others report its obesogenic properties as it functions as a substrate for both hepatic and adipocyte lipogenesis [11,14]. Gao et al found that acetate functions as an epigenetic metabolite to promote cancer cell survival under hypoxic states by serving as a carbon source for lipid synthesis [15]. Still, few studies suggest acetate suppresses weight gain, improves insulin sensitivity, stimulates the gut-brain axis to suppress appetite, and can downregulate inflammation in obesity by reducing proinflammatory cytokines and increasing regulatory T-cells [16-19]. This discrepancy in data surrounding acetate is likely related to its dynamic signaling which may vary based on the physiologic state [20].
Propionate is generated through bacterial fermentation of indigestible fibers through predominantly the succinate pathway and to a lesser extent the acrylate and propanediol pathway. The ability to ferment the various SCFAs are dependent on genes encoded in the microbiome such as the mmdA gene that encodes for the methylmalonyl-CoA decarboxylase in Bacteroidetes and in many Negativicutes families of bacteria [21]. Once formed, propionate is utilized at the level of the colonocytes as a substrate for intestinal gluconeogenesis through the FFAR3 signaling pathway, or it is absorbed into the portal system and taken to the liver where it is utilized as a substrate in hepatic gluconeogenesis [22]. Human studies demonstrate propionate to have an overall anti-obesity effect. In a randomized controlled study, daily ingestion of 10g of propionate resulted in significantly increased post-prandial GLP-1 and peptide PYY plasma levels, reduced weight gain, intra-abdominal adipose tissue distribution, intrahepatocellular lipid content and prevented the development of insulin sensitivity seen in the control group [23]. Propionate also has been shown to have anti-inflammatory properties by decreasing levels of interleukin-8 and TNF-α release from neutrophils [24,25].
Butyrate is perhaps the best studied and the most beneficial to human health. Four pathways for butyrate synthesis have been described, and occur predominantly through the acetyl-CoA pathway, followed by the lysine, glutarate, and succinate pathways [26]. Butyrate has wide-ranging clinical benefits not only for the luminal colonocytes where it is the preferred energy molecule, but also at the systemic level. These benefits include maintaining mucosal integrity, modulating both local and systemic immunity, and inhibiting neoplastic changes at the cellular level [27,28].
Butyrate supports mucosal integrity through activation of the peroxisome proliferator-activated receptor-γ (PPARγ). and stimulates β-oxidation and oxygen consumption in the gut, resulting to a rich luminal anaerobic environment [29]. In addition, it increases mucin production by goblet cells, increases immunoglobulin synthesis, and enhances secretion of antimicrobial peptides [30,31]. It further supports antimicrobial function by enabling the conversion of the proinflammatory M1 macrophage to the resolution-phase M2 macrophage [32]. Apart from the significant immunity role butyrate is shown to support a strong anti-neoplastic effect. Termed the “butyrate paradox”, they describe the contradictory effects of proliferation caused by butyrate in undifferentiated neoplastic cells where glucose is the preferred energy molecule, and differentiated colonocytes where butyrate is used. In these neoplastic cells, at the intra-cellular level, butyrate accumulates leading to histone modification which alters transcription and halts cell cycle progression, therefore protecting against colonic neoplasia [33]. Finally, similar to propionate, butyrate also serves to have an anti-obesity effect through its ability to stimulate the release of anorexigenic hormones and stimulates leptin synthesis [34,35].
Trimethylamine-N-oxide (TMAO)
There has been increasing evidence regarding the role trimethylamine-N-oxide (TMAO), a common gut microbiome derived dietary metabolite, plays in ischemic atherosclerotic disease risk [36,37]. Importantly, the essential role of gut flora in the ultimate production of plasma TMAO has been independently established. Suppression of intestinal flora with broad spectrum antibiotics results to decreased plasma TMAO; reversal of this affects bacterial recolonization [37]. The gut microbiota plays an obligatory role in converting various dietary nutrients, such as choline, betaine, L-carnitine, ergothioneine, trimethyllysine, y-butyrobetaine, phosphatidylcholine, glycerophosphocholine, and TMAO, into trimethylamine (TMA) gas which is rapidly absorbed into portal circulation where it is subsequently oxidized to TMAO by hepatic flavin-containing mono-oxygenase (FMO) [38]. These TMA precursor nutrients are primarily derived from animal products such as red meat, poultry, fish, and eggs [2]. Romano et al identified nine human intestinal strains capable of producing TMA derived from choline within the Firmicutes and Proteobacteria phyla, and colonization of gnotobiotic mice with these microbes resulted in TMAO accumulation in serum [39]. A number of specific bacterial enzymes have been implicated in the generation of TMA, including choline-TMA lyase, carnitine monooxygenase, betaine reductase, and TMAO reductase [38,40]. Table 2 highlight the main effects of TMAO.
Table 2.
Effects of TMAO
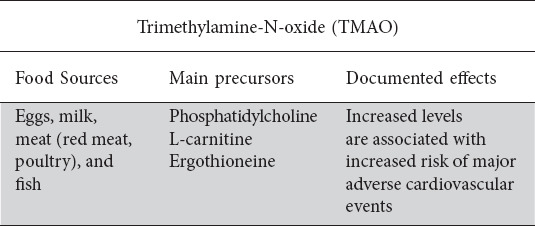
TMAO has emerged having an important role in cardiovascular disease. Wang et al first demonstrated a strong correlation between systemic TMAO levels and coronary atherosclerotic burden and cardiac risk [41]. This was followed by Tang et al who demonstrated that increased plasma TMAO levels were associated with significantly increased risk of major adverse cardiovascular events, even when adjusted for traditional risk factors [36]. The exact mechanism by which TMAO plays a role in this space is less clear; in rodent models, dietary TMAO or its precursors lead to accelerated arteriosclerosis and platelet aggregation, while inhibition of TMA production by selective TMA-lyase inhibition attenuates these effects [41,42]. On the contrary, mice specifically fed L-carnitine diets to increase plasma TMAO levels were interestingly shown to have reduced aortic atherosclerosis- perhaps suggesting differential downstream effects from different nutrient precursors [43]. Separately, the role TMAO plays in patients in heart failure, has also been investigated. Those with heart failure are known to have functional intestinal dysbiosis secondary associated visceral mucosal ischemia, leading to chronic inflammation, increased intestinal permeability, and importantly a relative shift in the composition of gut microbiota to favor TMA-forming species such as Firmicutes and Proteobacteria [44]. TMAO, in turn, has been shown to have a number of direct and indirect effects that exacerbate heart failure, including promoting myocardial hypertrophy and fibrosis, activation of inflammatory pathways to induce endothelial dysfunction, pathologic ventricular remodeling, and renal interstitial fibrosis [45,46].
A number of interventions have been studied to alter the gut microbiome-TMAO-cardiovascular disease axis, including dietary changes to reduce plasma TMAO levels, probiotic supplementation, and potential enzymatic drug targets to reduce TMAO formation [47-50]. As work continues to better characterize this pathway, opportunities to identify novel therapeutic interventions will continue to emerge which are likely to have a profound influence on patient related outcomes in cardiovascular disease. Although levels of TMAO are dependent on the linkage of diet and gut microbiome it is important to note that, its regulation is associated with other host-environmental factors such as host comorbidities and genetics.
Tryptophan metabolites
While microbiota-generated SCFA and TMAO metabolites have been studied for over a century, the role metabolites generated by proteolysis has received little attention. Historically, products from proteolysis have been associated with negative effects, however newer data suggest that tryptophan metabolites play a beneficial role in intestinal homeostasis. Table 3 highlights the effects of the various tryptophan metabolites.
Table 3.
Effects of tryptophan metabolites

Tryptophan is one of the nine essential amino acids that humans are unable to produce, thus it must be consumed from protein sources including meats, fish, eggs, and nuts [2]. A majority is absorbed in the small intestine and a smaller amount does reach the colon where bacteria convert tryptophan into indole and various indole derivates. These derivatives include indoleacetic acid, indolepropionic acid, indolelactic acid, indoleacrylic acid, indolealdehyde, indoleethanol, tryptamine, and skatole [51,52]. The function of these tryptophan metabolites are multiple including antimicrobial effects and modulating the immune system, as well as maintaining mucosal homeostasis by affecting systemic hormone secretion and possessing anti-oxidant properties [53,54].
Studies on indole demonstrate its antimicrobial effects. It exerts anti-bacterial activity against Staphylococcus aureus, Salmonella, Lactobacillus, E. coli, and B. cereus. In addition, indoleethanol inhibits bacteriophage replication in certain bacterial strains and also prevents proliferation of parasitic protozoa [55,56]. Metabolites of tryptophan are also capable of modulating the innate and adaptive immune system, through its ability to bind to the aryl hydrocarbon receptor (AHR), present on cells of the immune system including dendritic cells and T-cells [53]. These metabolites function as AHR ligands and in several murine models have been shown to limit intestinal inflammation. For example, low levels of AHR ligands are implicated in the pathogenesis of inflammatory bowel disease [57,58]. Additionally, indole and IA maintain the intestinal epithelial barrier by promoting goblet cell differentiation and mucus production, which further aids in mitigating potential intestinal inflammation [54,59].
Systemically, tryptophan and its metabolites also play a role in hormone secretion with anti-inflammatory properties. Indole functions as a signaling molecule at the colonic enteroendocrine L cells to stimulate GLP-1 secretion, thereby affecting insulin secretion from pancreatic B-cells, suppressing appetite, and slowing gastric emptying [60,61]. Further, IPA serves as scavenger for free oxygen radicals to prevent oxidative damage [62]
Metabolites produced by interaction between bile acids and gut microbiome
Bile acids
Bile acids (BA) are amphipathic molecules, which contain both hydrophilic and hydrophobic regions. They serve to solubilize dietary lipids by forming micelles in the small intestine to help facilitate lipid absorption or excretion. In addition to their role in the absorption of dietary fat and homeostasis of cholesterol, bile acids also serve as signal molecules through interactions with several nuclear hormone receptors, including farnesoid X receptor (FXR), peroxisome proliferator-activated receptor (PPAR), G-protein coupled receptor (TGR5), vitamin D receptor (VDR), and thyroid hormone receptor, allowing bile acids to act as hormones with far reaching effects throughout the body [63].
Approximately 1 liter of bile is produced daily by hepatocytes as the end product of cholesterol metabolism. BA synthesis is initiated in one of two pathways in humans: through cholesterol 7α-hydroxylase termed the classic pathway, or through sterol 27-hydroxylase termed the alternative pathway. The end products of these reactions form the primary BAs cholic acid and chenodeoxycholic acid. These are subsequently conjugated with glycine or taurine, forming bile salts which are then stored in the gallbladder. Table 4 highlights the effects of bile acids.
Table 4.
Bile acids and their effects
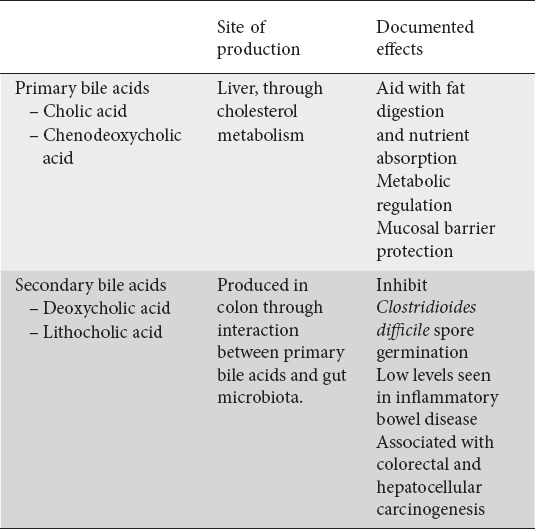
Postprandial secretion of cholecystokinin leads to primary BA release into the proximal duodenum to aid with fat digestion, nutrient absorption, metabolic regulation, and mucosal barrier protection [64]. While ~95% of BA are reabsorbed in the distal ileum through the apical sodium-dependent bile acid transporter (ASBT) and recycled into enterohepatic circulation, bacterial deconjugation prevents reuptake into enterocytes allowing ~5% of the BA to continue to the colon. In the colon, this population of BA interact with the gut microbiota to produce secondary BA deoxycholic acid (DCA) and lithocholic acid (LCA). Over 50 different secondary bile acids have been characterized as the products of the interaction between primary bile acids and the gut microbiota through various deconjugation, dehydrogenation, and dihydroxylation reactions with DCA and LCA being the two most abundant [65].
The balance between primary and secondary bile acids are crucial to host health, as an imbalance is associated with detrimental effects on the host. This is likely due to such intricacies as host microenvironment, antibiotic exposure, diet, and microbiota composition. For example, Clostridioides difficile infection (CDI) can occur when the normal gut microbiota is depleted by antibiotics. The native microbiota is required to convert primary to secondary bile acids which help prevent CDI. Theriot et al were able to demonstrate that specific bile acids are able to initiate C. difficile spore germination, while other secondary bile acids are able to inhibit its growth [66]. An imbalance is also seen in inflammatory bowel disease where BA metabolism is distinctly dysregulated. Stool studies demonstrate elevated levels of primary BAs and reduced levels of secondary BAs. Duboc et al demonstrated that this change induces a greater inflammatory response and may participate in the chronic inflammation loop of IBD [67].
The association of BA with inflammation, also implicates the secondary BAs, namely DCA and LCA, to gastrointestinal cancer, particularly colorectal cancer and hepatocellular cancer. Because DCA and LCA are more hydrophobic than the primary BAs, they more readily disrupt cellular membranes and induce cell damage. DCA can also directly degrade p53. An additional mechanism leading to carcinogenesis is through the creation of reactive oxygen species which formed through cell damage caused by DCA and LCA. Finally, taurine-conjugated BA are deconjugated and metabolized into H2S, a known potent carcinogen.
While such findings implicate bile acids in the mechanisms of antibiotic resistance and carcinogenesis, it also may position bile acids as novel therapeutic targets. The central role of bile acids in regulation of metabolic and immune homeostasis is beginning to be explored for therapeutic use in other settings as well. Oral delivery of a probiotic in a multi-site randomized control trial was able to increase bile acids with a subsequent decrease in markers of inflammation including CRP and TNF-a and modulation of bile acids through FXR signaling is now being investigated as a therapy for NAFLD and non-alcoholic steatohepatitis [68,69]. Additionally, LCA activates vitamin D receptors in a highly selective manner, leading to expression of CYP3A, a cytochrome P450 enzyme which helps metabolize LCA to less harmful metabolites, and may be a mechanism through which vitamin D exerts a protective effect against colon cancer [70,71].
Metabolites produced by the gut microbiome alone
Polyamines
Polyamines (PA) are defined as small polycationic molecules, present in millimolar concentrations that spontaneously interact with various macromolecules such as DNA, RNA, phospholipids and proteoglycans. The three main PAs important to human health are putrescine, spermidine and spermine. PAs are the non-protein amino acids that are essential across all living organisms are associated with a wide range of biological functions that include gene and stress regulation, cell proliferation and differentiation, and in the regulation of enzymatic activity. PAs are found in both natural and processed foods, and the intestinal lumen concentration is dependent on both diet and synthesis by gut microbiota [72]. It is believed that the exogenous PAs derived from food are nearly almost absorbed by the upper gastrointestinal (GI) tract while the PA in the lower part of the GI tract are synthesized by the gut microbiota [73]. Table 5 highlights the effects of PAs.
Table 5.
Polyamines and their effects

Gut microbiota are able to synthesize polyamines using constitutive or inducible forms of amino acid decarboxylase enzymes located in the cytosol. Synthesized by ornithine decarboxylase (ODC), putrescine can then be converted into spermidine with the addition of an aminopropyl moiety donated from decarboxylated S-adenosylmethionine (dcAdoMet) by spermidine synthase [74,75]. Additionally, Matsumoto et al, recently demonstrated that the intestinal luminal levels of putrescine and spermidine are mainly dependent on colonic microbiota [76]. Spermine can then be created through the donation of an additional aminopropyl group to the amino butyl end of spermidine, through the use of spermine synthase. After synthesis, PAs are transported to the proximal gut via the portal circulation and biliary tree [75].
Putrescine, spermidine and spermine are some of the most important metabolites produced by the gut microbiota, as they affect the overall health of the host. PAs have been shown to have antioxidant effects and inhibit production of inflammatory cytokines, as well as influence the intestinal mucosal barrier. These benefits, in addition to resistance to oxidative stress, can increase the longevity of the host through administration of probiotics that lead to suppression of chronic low-grade inflammation as a result of higher PA levels [76]. Recent epidemiological studies have demonstrated a decrease in cardiovascular events and mortality with an increased PA intake, specifically spermidine [76-78]. In addition, AdoMet is a major contributor to DNA methylation, which is a molecular marker used to both monitor aging and predict life expectancy [79]. Recent studies have demonstrated that low levels of spermidine and spermine levels can increase the accumulation of dcAdoMet, and therefore reduce DNA methylation levels [75].
However, dysregulation of PA metabolism can lead to pro-carcinogenic effects, as high concentrations of these PAs have been suggested to be involved in the tumorigenesis of colorectal cancer and other tumors [80]. Tumors have been shown to induce the PA biosynthetic pathway, and a high level of PAs can therefore create a beneficial environment for tumor growth. Recent work has been investigating this by looking at inhibiting the ODC enzyme; however, this has not been successful as the tumors supplement their PA environments through exogenous sources. For patients with advanced adenomas, a recent proposal suggests that a PA deficient diet could help avoid recurrence after use of PA-inhibitory drugs [75]. Although systemic PA concentrations are tightly controlled through complex networks, additional work is needed to understand how the gut microbiota and regulation of PA biosynthesis can serve as an effective prevention or treatment of human diseases.
Branched-chain amino acids (BCAAs)
Of the twenty amino acids, nine are unable to be produced within the human body and are therefore considered to be essential. Although diet is the largest source of leucine, isoleucine and valine, these branched-chain amino acids (BCAAs) are also both degraded and synthesized by the metabolic pathways of the gut microbiota [81,82]. Given that BCAAs are important synthesis substrates, there is an increased demand for them during bacterial invasion [81]. In addition to providing energy via catabolism, activating the mTOR pathway and serving as signaling molecules that regulate glucose, lipid and protein synthesis, BCAAs are required for the upkeep of the high metabolic status of activated T-cells [81,83]. Disruption in the levels of BCAAs and their derivatives have been identified as potential biomarkers for diseases such as insulin resistance, T2DM, cancer and cardiovascular diseases [83]. Given that gut microbiota are able to produce BCAAs via their own biosynthetic pathways, it is hypothesized that intestinal microbiota can also contribute to and effect BCAA host availability. When accessing the microbial source of amino acids (AAs), the ileal microbiota is of most importance because the small intestine is most responsible for AA uptake [82]. In vitro experiments have demonstrated Clostridium, Bacillus-Lactobacillus-Streptococcus, and Proteobacteria groups as the most abundant AA-fermenting bacteria in the small intestine, while Clostridia and Peptostreptococci groups are the most abundant in the large intestine [83].
Three mechanistic studies involving mice have demonstrated that manipulation of the gut microbiota, including the low-abundant bacteria, can have significant effects on the systemic BCAA pool and host metabolism [82,84]. Elevated BCAA concentrations are also associated with obesity, diabetes and cancer in humans [82]. Altered BCAA concentrations are significantly associated prior to and after development of T2DM. Pedersen et al demonstrated this in a study looking at 277 insulin-resistant, non-diabetic subjects, of which insulin resistance positively correlated with BCAA synthesis [83,85]. This is further correlated with a gut microbiome that has an enhanced biosynthetic potential for BCAAs. Furthermore, a metagenome wide study demonstrated that obese individuals have a depleted BCAA degradation pathway and thus have a higher capacity to produce aromatic amino acids and BCAAs [86]. Additionally, BCAAs are essential for cancer growth, and the tumors are able to use BCAAs as an energy source [83]. Increased plasma levels of BCAAs are also found in both pancreatic and breast cancers [83,87].
Recent studies are looking at modulating host systemic BCAA’s by manipulating the gut microbiota and have shown promising results [82]. Although these studies are in their early stages and data is limited, these studies warrant further investigation in order to fully understand the effect of the BCAAs produced from gut microbiota.
Bacterial vitamins
Essential vitamins are either obtained from the diet or synthesized by gut microbiota [81]. Vitamins can either be fat-soluble or water-soluble, and each serve a vast array of functions within the body. While fat-soluble vitamins are important components of the cell membrane, water-soluble vitamins function as coenzymes in metabolic reactions. It has been demonstrated that the gut microbiota are able to synthesize vitamin K2 and most water-soluble B vitamins, such as biotin, cobalamin, folates, nicotinic acid, pantothenic acid, pyridoxine, riboflavin and thiamine [88]. While dietary vitamins are absorbed in the small intestine, the majority of the uptake of vitamins produced from gut microbiota occur in the colon [81,88]. Table 6 outlines the symptoms caused by these vitamin deficiencies.
Table 6.
Symptoms caused by vitamin deficiencies
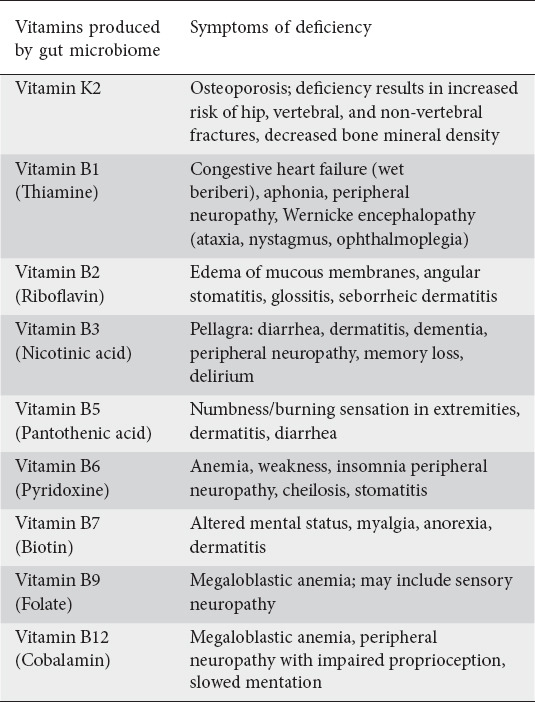
The main vitamins produced by the gut microbiota include vitamin K2 and various members of the vitamin B family, including vitamin B1 (thiamine), B2 (riboflavin), B3 (nicotinic acid), B5 (pantothenic acid), B6 (pyridoxine), B7 (biotin), B9 (folate), and B12 (cobalamin) [81]. Recent studies have also shown the production of vitamin C (ascorbate) from gut microbiota as well, although less abundant. While the metabolic pathways utilized by microbes to produce ascorbic acid via gut microbiota are unknown, low concentrations of ascorbate have been reported in the inflamed mucosa of patients with IBD [88]. In addition, ascorbate was reported to suppress T-effector cells and inhibit T-cell activation [89]. A recent study by Pham et al demonstrated that colon-delivered vitamin C results in significantly increased microbial alpha diversity and fecal SCFAs [90]. Further investigation is needed to determine both the metabolic pathways and the effect of ascorbate levels on preventing microbiota-related human diseases.
Synthesis of these B-vitamins occur through a variety of bacterial strains. For example, B12 (cobalamin) synthesis occurs in the phylum Fusobacteria and B2 (riboflavin) is synthesized by phyla Bacteroidetes, Fusobacteria, Proteo-bacteria, and Firmicutes [81]. Recent studies have shown that the intermediate 5-(2-oxopropylideneamino)-6-D-ribitylami-nouracil (5-OP-RU) during riboflavin synthesis regulates the mucosal-associated invariant T (MAIT) cells and respond to microbiota in an MHC-related molecule 1 (MR1) in a dose dependent manner [81,91]. Although typically an unstable intermediate, 5-OP-RU becomes trapped in MR1 and is thus used as an antigen to activate MAIT cells. This activation leads to cytotoxic effector functions, migration, and proliferative expansion [92].
In contrast, vitamin B9 metabolite 6-formylpterin (6-FP) competes with 5-OP-RU and inhibits this MAIT activation by binding to MR1 [81,92]. Although MAIT cells were discovered more than 25 years ago, it has only recently captured the attention of researchers. While some studies have reported that MAIT cells have a role in various diseases, there is considerable disagreement in the amplitude of their affect. However, it is known that MAIT cell abundance varies among people, with those at high risk, such as the very young, elderly and immunocompromised, having low abundance. It is important to continue investigation in this area to determine how 5-OP-RU could be used to enhance adaptive immunity, and how 6-FP can block MAIT functions when MAIT cells become lymphomas [92].
Limitations
A limitation inherent to this review article is that the majority of the published data focus on the blood and serum metabolome. An increasing emphasis in recent literature has highlighted the need to also address the impact of fecal, urine, and saliva metabolome on health as the two do not always correlate [93]. It is possible that an incomplete picture and conclusions are being drawn when studying and evaluating only one biofluid. Additionally, many confounders affect the gut microbiome and in effect, the metabolites produced. Therefore, more studies stratifying those confounders are warranted
Concluding remarks
Decades of research have impacted our understanding of the gut microbiome and the role they play in maintaining homeostasis. Through their litany of metabolic end-products, produced in a variety of pathways, these products alter human physiology, pathology, immunity, and metabolism. There ultimately still remain a plethora of unknown chemical metabolites that have yet to be discovered and much research is still needed to fully elucidate the effects of the known gut microbiome-derived metabolites. In addition, this review covers the metabolites produced by the bacterial inhabitants of the gut, yet there still remains the need to evaluate the metabolic end-products produced by viruses, fungi, and bacteriophages. While much of this research is still in its discovery phase, future studies in this area will undoubtedly reveal novel strategies, therapies, and treatments that will integrate the gut microbiome to inform clinical practice.
Biography
Oregon Health and Science University, Portland, OR, USA
Footnotes
Conflict of Interest: None
References
- 1.Gilbert JA, Blaser MJ, Caporaso JG, Jansson JK, Lynch SV, Knight R. Current understanding of the human microbiome. Nat Med. 2018;24:392–400. doi: 10.1038/nm.4517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Fan Y, Pedersen O. Gut microbiota in human metabolic health and disease. Nat Rev Microbiol. 2021;19:55–71. doi: 10.1038/s41579-020-0433-9. [DOI] [PubMed] [Google Scholar]
- 3.Han S, Van Treuren W, Fischer CR, et al. A metabolomics pipeline for the mechanistic interrogation of the gut microbiome. Nature. 2021;595:415–420. doi: 10.1038/s41586-021-03707-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Rahman S, Trone K, Kelly C, Stroud A, Martindale R. All Fiber is Not Fiber. Curr Gastroenterol Rep. 2023;25:1–12. doi: 10.1007/s11894-022-00858-1. [DOI] [PubMed] [Google Scholar]
- 5.den Besten G, van Eunen K, Groen AK, Venema K, Reijngoud DJ, Bakker BM. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J Lipid Res. 2013;54:2325–2340. doi: 10.1194/jlr.R036012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Parada Venegas D, De la Fuente MK, Landskron G, et al. Short chain fatty acids (SCFAS)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front Immunol. 2019;10:277. doi: 10.3389/fimmu.2019.00277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kimura I, Ozawa K, Inoue D, et al. The gut microbiota suppresses insulin-mediated fat accumulation via the short-chain fatty acid receptor GPR43. Nat Commun. 2013;4:1829. doi: 10.1038/ncomms2852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ang Z, Ding JL. GPR41 and GPR43 in obesity and inflammation - protective or causative? Front Immunol. 2016;7:28. doi: 10.3389/fimmu.2016.00028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Tolhurst G, Heffron H, Lam YS, et al. Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein-coupled receptor FFAR2. Diabetes. 2012;61:364–371. doi: 10.2337/db11-1019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Larraufie P, Martin-Gallausiaux C, Lapaque N, et al. SCFAs strongly stimulate PYY production in human enteroendocrine cells. Sci Rep. 2018;8:74. doi: 10.1038/s41598-017-18259-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hernández MAG, Canfora EE, Jocken JWE, Blaak EE. The short-chain fatty acid acetate in body weight control and insulin sensitivity. Nutrients. 2019;11:1943. doi: 10.3390/nu11081943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Canfora EE, Blaak EE. Acetate:a diet-derived key metabolite in energy metabolism:good or bad in context of obesity and glucose homeostasis? Curr Opin Clin Nutr Metab Care. 2017;20:477–483. doi: 10.1097/MCO.0000000000000408. [DOI] [PubMed] [Google Scholar]
- 13.Moffett JR, Puthillathu N, Vengilote R, Jaworski DM, Namboodiri AM. Acetate revisited:a key biomolecule at the nexus of metabolism, epigenetics and oncogenesis-part 1:acetyl-CoA, acetogenesis and acyl-CoA short-chain synthetases. Front Physiol. 2020;11:580167. doi: 10.3389/fphys.2020.580167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Duwaerts CC, Maher JJ. Macronutrients and the adipose-liver axis in obesity and fatty liver. Cell Mol Gastroenterol Hepatol. 2019;7:749–761. doi: 10.1016/j.jcmgh.2019.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gao X, Lin SH, Ren F, et al. Acetate functions as an epigenetic metabolite to promote lipid synthesis under hypoxia. Nat Commun. 2016;7:11960. doi: 10.1038/ncomms11960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Balmer ML, Ma EH, Thompson AJ, et al. Memory CD8+ T cells balance pro- and anti-inflammatory activity by reprogramming cellular acetate handling at sites of infection. Cell Metab. 2020;32:457–467. doi: 10.1016/j.cmet.2020.07.004. [DOI] [PubMed] [Google Scholar]
- 17.Xu M, Jiang Z, Wang C, et al. Acetate attenuates inflammasome activation through GPR43-mediated Ca2+-dependent NLRP3 ubiquitination. Exp Mol Med. 2019;51:1–13. doi: 10.1038/s12276-019-0276-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Priyadarshini M, Villa SR, Fuller M, et al. An acetate-specific GPCR, FFAR2, regulates insulin secretion. Mol Endocrinol. 2015;29:1055–1066. doi: 10.1210/me.2015-1007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Frost G, Sleeth ML, Sahuri-Arisoylu M, et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat Commun. 2014;5:3611. doi: 10.1038/ncomms4611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Moffett JR, Puthillathu N, Vengilote R, Jaworski DM, Namboodiri AM. Acetate revisited:a key biomolecule at the nexus of metabolism, epigenetics, and oncogenesis - part 2:acetate and ACSS2 in health and disease. Front Physiol. 2020;11:580171. doi: 10.3389/fphys.2020.580171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Reichardt N, Duncan SH, Young P, et al. Phylogenetic distribution of three pathways for propionate production within the human gut microbiota. ISME J. 2014;8:1323–1335. doi: 10.1038/ismej.2014.14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Tedelind S, Westberg F, Kjerrulf M, Vidal A. Anti-inflammatory properties of the short-chain fatty acids acetate and propionate:a study with relevance to inflammatory bowel disease. World J Gastroenterol. 2007;13:2826–2832. doi: 10.3748/wjg.v13.i20.2826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Chambers ES, Byrne CS, Morrison DJ, et al. Dietary supplementation with inulin-propionate ester or inulin improves insulin sensitivity in adults with overweight and obesity with distinct effects on the gut microbiota, plasma metabolome and systemic inflammatory responses:a randomised cross-over trial. Gut. 2019;68:1430–1438. doi: 10.1136/gutjnl-2019-318424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.De Vadder F, Kovatcheva-Datchary P, Goncalves D, et al. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell. 2014;156:84–96. doi: 10.1016/j.cell.2013.12.016. [DOI] [PubMed] [Google Scholar]
- 25.Chambers ES, Viardot A, Psichas A, et al. Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adults. Gut. 2015;64:1744–1754. doi: 10.1136/gutjnl-2014-307913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Vital M, Howe AC, Tiedje JM. Revealing the bacterial butyrate synthesis pathways by analyzing (meta)genomic data. mBio. 2014;5:e00889. doi: 10.1128/mBio.00889-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Yip W, Hughes MR, Li Y, et al. Butyrate shapes immune cell fate and function in allergic asthma. Front Immunol. 2021;12:628453. doi: 10.3389/fimmu.2021.628453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Li Q, Cao L, Tian Y, et al. Butyrate suppresses the proliferation of colorectal cancer cells via targeting pyruvate kinase M2 and metabolic reprogramming. Mol Cell Proteomics. 2018;17:1531–1545. doi: 10.1074/mcp.RA118.000752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Byndloss MX, Olsan EE, Rivera-Chávez F, et al. Microbiota-activated PPAR-g signaling inhibits dysbiotic Enterobacteriaceae expansion. Science. 2017;357:570–575. doi: 10.1126/science.aam9949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Toni T, Alverdy J, Gershuni V. Re-examining chemically defined liquid diets through the lens of the microbiome. Nat Rev Gastroenterol Hepatol. 2021;18:903–911. doi: 10.1038/s41575-021-00519-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Chen J, Vitetta L. The role of butyrate in attenuating pathobiont-induced hyperinflammation. Immune Netw. 2020;20:e15. doi: 10.4110/in.2020.20.e15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ji J, Shu D, Zheng M, et al. Microbial metabolite butyrate facilitates M2 macrophage polarization and function. Sci Rep. 2016;6:24838. doi: 10.1038/srep24838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Salvi PS, Cowles RA. Butyrate and the intestinal epithelium:modulation of proliferation and inflammation in homeostasis and disease. Cells. 2021;10:1775. doi: 10.3390/cells10071775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Li Z, Yi CX, Katiraei S, et al. Butyrate reduces appetite and activates brown adipose tissue via the gut-brain neural circuit. Gut. 2018;67:1269–1279. doi: 10.1136/gutjnl-2017-314050. [DOI] [PubMed] [Google Scholar]
- 35.Xiong Y, Miyamoto N, Shibata K, et al. Short-chain fatty acids stimulate leptin production in adipocytes through the G protein-coupled receptor GPR41. Proc Natl Acad Sci U S A. 2004;101:1045–1050. doi: 10.1073/pnas.2637002100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Tang WH, Wang Z, Levison BS, et al. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N Engl J Med. 2013;368:1575–1584. doi: 10.1056/NEJMoa1109400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wang Z, Klipfell E, Bennett BJ, et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature. 2011;472:57–63. doi: 10.1038/nature09922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Zhang Y, Wang Y, Ke B, Du J. TMAO:how gut microbiota contributes to heart failure. Transl Res. 2021;228:109–125. doi: 10.1016/j.trsl.2020.08.007. [DOI] [PubMed] [Google Scholar]
- 39.Romano KA, Vivas EI, Amador-Noguez D, Rey FE. Intestinal microbiota composition modulates choline bioavailability from diet and accumulation of the proatherogenic metabolite trimethylamine-N-oxide. mBio. 2015;6:e02481. doi: 10.1128/mBio.02481-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Rath S, Rud T, Pieper DH, Vital M. Potential TMA-producing bacteria are ubiquitously found in mammalia. Front Microbiol. 2019;10:2966. doi: 10.3389/fmicb.2019.02966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wang Z, Roberts AB, Buffa JA, et al. Non-lethal inhibition of gut microbial trimethylamine production for the treatment of atherosclerosis. Cell. 2015;163:1585–1595. doi: 10.1016/j.cell.2015.11.055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Zhu W, Gregory JC, Org E, et al. Gut microbial metabolite TMAO enhances platelet hyperreactivity and thrombosis risk. Cell. 2016;165:111–124. doi: 10.1016/j.cell.2016.02.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Collins HL, Drazul-Schrader D, Sulpizio AC, et al. L-Carnitine intake and high trimethylamine N-oxide plasma levels correlate with low aortic lesions in ApoE(-/-) transgenic mice expressing CETP. Atherosclerosis. 2016;244:29–37. doi: 10.1016/j.atherosclerosis.2015.10.108. [DOI] [PubMed] [Google Scholar]
- 44.Chen K, Zheng X, Feng M, Li D, Zhang H. Gut microbiota-dependent metabolite trimethylamine n-oxide contributes to cardiac dysfunction in western diet-induced obese mice. Front Physiol. 2017;8:139. doi: 10.3389/fphys.2017.00139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Savi M, Bocchi L, Bresciani L, et al. Trimethylamine-N-oxide (TMAO)-induced impairment of cardiomyocyte function and the protective role of urolithin B-glucuronide. Molecules. 2018;23:549. doi: 10.3390/molecules23030549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Tang WH, Wang Z, Kennedy DJ, et al. Gut microbiota-dependent trimethylamine N-oxide (TMAO) pathway contributes to both development of renal insufficiency and mortality risk in chronic kidney disease. Circ Res. 2015;116:448–455. doi: 10.1161/CIRCRESAHA.116.305360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.De Filippis F, Pellegrini N, Vannini L, et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut. 2016;65:1812–1821. doi: 10.1136/gutjnl-2015-309957. [DOI] [PubMed] [Google Scholar]
- 48.Gan XT, Ettinger G, Huang CX, et al. Probiotic administration attenuates myocardial hypertrophy and heart failure after myocardial infarction in the rat. Circ Heart Fail. 2014;7:491–499. doi: 10.1161/CIRCHEARTFAILURE.113.000978. [DOI] [PubMed] [Google Scholar]
- 49.Qiu L, Yang D, Tao X, Yu J, Xiong H, Wei H. Enterobacter aerogenes ZDY01 attenuates choline-induced trimethylamine N-oxide levels by remodeling gut microbiota in mice. J Microbiol Biotechnol. 2017;27:1491–1499. doi: 10.4014/jmb.1703.03039. [DOI] [PubMed] [Google Scholar]
- 50.Roberts AB, Gu X, Buffa JA, et al. Development of a gut microbe-targeted nonlethal therapeutic to inhibit thrombosis potential. Nat Med. 2018;24:1407–1417. doi: 10.1038/s41591-018-0128-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Roager HM, Licht TR. Microbial tryptophan catabolites in health and disease. Nat Commun. 2018;9:3294. doi: 10.1038/s41467-018-05470-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Platten M, Nollen EAA, Röhrig UF, Fallarino F, Opitz CA. Tryptophan metabolism as a common therapeutic target in cancer, neurodegeneration and beyond. Nat Rev Drug Discov. 2019;18:379–401. doi: 10.1038/s41573-019-0016-5. [DOI] [PubMed] [Google Scholar]
- 53.Rothhammer V, Quintana FJ. The aryl hydrocarbon receptor:an environmental sensor integrating immune responses in health and disease. Nat Rev Immunol. 2019;19:184–197. doi: 10.1038/s41577-019-0125-8. [DOI] [PubMed] [Google Scholar]
- 54.Powell DN, Swimm A, Sonowal R, et al. Indoles from the commensal microbiota act via the AHR and IL-10 to tune the cellular composition of the colonic epithelium during aging. Proc Natl Acad Sci U S A. 2020;117:21519–21526. doi: 10.1073/pnas.2003004117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Qin HL, Liu J, Fang WY, Ravindar L, Rakesh KP. Indole-based derivatives as potential antibacterial activity against methicillin-resistance Staphylococcus aureus (MRSA) Eur J Med Chem. 2020;194:112245. doi: 10.1016/j.ejmech.2020.112245. [DOI] [PubMed] [Google Scholar]
- 56.Jin M, Xu C, Zhang X. The effect of tryptophol on the bacteriophage infection in high-temperature environment. Appl Microbiol Biotechnol. 2015;99:8101–8111. doi: 10.1007/s00253-015-6674-2. [DOI] [PubMed] [Google Scholar]
- 57.Natividad JM, Agus A, Planchais J, et al. Impaired aryl hydrocarbon receptor ligand production by the gut microbiota is a key factor in metabolic syndrome. Cell Metab. 2018;28:737–749. doi: 10.1016/j.cmet.2018.07.001. [DOI] [PubMed] [Google Scholar]
- 58.Ghiboub M, Verburgt CM, Sovran B, Benninga MA, de Jonge WJ, Van Limbergen JE. Nutritional therapy to modulate tryptophan metabolism and aryl hydrocarbon-receptor signaling activation in human diseases. Nutrients. 2020;12:2846. doi: 10.3390/nu12092846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Yang S, Yu M. Role of goblet cells in intestinal barrier and mucosal immunity. J Inflamm Res. 2021;14:3171–3183. doi: 10.2147/JIR.S318327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Acar I, Cetinkaya A, Lay I, Ileri-Gurel E. The role of calcium sensing receptors in GLP-1 and PYY secretion after acute intraduodenal administration of L-Tryptophan in rats. Nutr Neurosci. 2020;23:481–489. doi: 10.1080/1028415X.2018.1521906. [DOI] [PubMed] [Google Scholar]
- 61.Buckley MM, O'Brien R, Brosnan E, et al. Glucagon-like peptide-1 secreting L-cells coupled to sensory nerves translate microbial signals to the host rat nervous system. Front Cell Neurosci. 2020;14:95. doi: 10.3389/fncel.2020.00095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Herraiz T, Galisteo J. Endogenous and dietary indoles:a class of antioxidants and radical scavengers in the ABTS assay. Free Radic Res. 2004;38:323–331. doi: 10.1080/10611860310001648167. [DOI] [PubMed] [Google Scholar]
- 63.Wahlström A, Sayin SI, Marschall HU, Bäckhed F. Intestinal crosstalk between bile acids and microbiota and its impact on host metabolism. Cell Metab. 2016;24:41–50. doi: 10.1016/j.cmet.2016.05.005. [DOI] [PubMed] [Google Scholar]
- 64.de Aguiar Vallim TQ, Tarling EJ, Edwards PA. Pleiotropic roles of bile acids in metabolism. Cell Metab. 2013;17:657–669. doi: 10.1016/j.cmet.2013.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Hamilton JP, Xie G, Raufman JP, et al. Human cecal bile acids:concentration and spectrum. Am J Physiol Gastrointest Liver Physiol. 2007;293:G256–G263. doi: 10.1152/ajpgi.00027.2007. [DOI] [PubMed] [Google Scholar]
- 66.Theriot CM, Bowman AA, Young VB. Antibiotic-induced alterations of the gut microbiota alter secondary bile acid production and allow for Clostridium difficile spore germination and outgrowth in the large intestine. mSphere. 2016;1:e00045–e00015. doi: 10.1128/mSphere.00045-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Duboc H, Rajca S, Rainteau D, et al. Connecting dysbiosis, bile-acid dysmetabolism and gut inflammation in inflammatory bowel diseases. Gut. 2013;62:531–539. doi: 10.1136/gutjnl-2012-302578. [DOI] [PubMed] [Google Scholar]
- 68.Schaap FG, Trauner M, Jansen PL. Bile acid receptors as targets for drug development. Nat Rev Gastroenterol Hepatol. 2014;11:55–67. doi: 10.1038/nrgastro.2013.151. [DOI] [PubMed] [Google Scholar]
- 69.Neuschwander-Tetri BA, Loomba R, Sanyal AJ, et al. NASH Clinical Research Network. Farnesoid X nuclear receptor ligand obeticholic acid for non-cirrhotic, non-alcoholic steatohepatitis (FLINT):a multicentre, randomised, placebo-controlled trial. Lancet. 2015;385:956–965. doi: 10.1016/S0140-6736(14)61933-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Adachi R, Honma Y, Masuno H, et al. Selective activation of vitamin D receptor by lithocholic acid acetate, a bile acid derivative. J Lipid Res. 2005;46:46–57. doi: 10.1194/jlr.M400294-JLR200. [DOI] [PubMed] [Google Scholar]
- 71.Zhu Y, Wang PP, Zhai G, et al. Vitamin D receptor and calcium-sensing receptor polymorphisms and colorectal cancer survival in the Newfoundland population. Br J Cancer. 2017;117:898–906. doi: 10.1038/bjc.2017.242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Sánchez-Jiménez F, Medina MÁ, Villalobos-Rueda L, Urdiales JL. Polyamines in mammalian pathophysiology. Cell Mol Life Sci. 2019;76:3987–4008. doi: 10.1007/s00018-019-03196-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Matsumoto M, Benno Y. The relationship between microbiota and polyamine concentration in the human intestine:a pilot study. Microbiol Immunol. 2007;51:25–35. doi: 10.1111/j.1348-0421.2007.tb03887.x. [DOI] [PubMed] [Google Scholar]
- 74.Hanfrey CC, Pearson BM, Hazeldine S, et al. Alternative spermidine biosynthetic route is critical for growth of Campylobacter jejuni and is the dominant polyamine pathway in human gut microbiota. J Biol Chem. 2011;286:43301–43312. doi: 10.1074/jbc.M111.307835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Ramos-Molina B, Queipo-Ortuño MI, Lambertos A, Tinahones FJ, Peñafiel R. Dietary and gut microbiota polyamines in obesity- and age-related diseases. Front Nutr. 2019;6:24. doi: 10.3389/fnut.2019.00024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Matsumoto M, Kurihara S, Kibe R, Ashida H, Benno Y. Longevity in mice is promoted by probiotic-induced suppression of colonic senescence dependent on upregulation of gut bacterial polyamine production. PLoS One. 2011;6:e23652. doi: 10.1371/journal.pone.0023652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Eisenberg T, Abdellatif M, Schroeder S, et al. Cardioprotection and lifespan extension by the natural polyamine spermidine. Nat Med. 2016;22:1428–1438. doi: 10.1038/nm.4222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Kiechl S, Pechlaner R, Willeit P, et al. Higher spermidine intake is linked to lower mortality:a prospective population-based study. Am J Clin Nutr. 2018;108:371–380. doi: 10.1093/ajcn/nqy102. [DOI] [PubMed] [Google Scholar]
- 79.Ciccarone F, Tagliatesta S, Caiafa P, et al. DNA methylation dynamics in aging:how far are we from understanding the mechanisms? Mech Ageing Dev. 2018;174:3–17. doi: 10.1016/j.mad.2017.12.002. [DOI] [PubMed] [Google Scholar]
- 80.Sugiyama Y, Nara M, Sakanaka M, et al. Comprehensive analysis of polyamine transport and biosynthesis in the dominant human gut bacteria:potential presence of novel polyamine metabolism and transport genes. Int J Biochem Cell Biol. 2017;93:52–61. doi: 10.1016/j.biocel.2017.10.015. [DOI] [PubMed] [Google Scholar]
- 81.Yang W, Cong Y. Gut microbiota-derived metabolites in the regulation of host immune responses and immune-related inflammatory diseases. Cell Mol Immunol. 2021;18:866–877. doi: 10.1038/s41423-021-00661-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Gojda J, Cahova M. Gut Microbiota as the link between elevated BCAA serum levels and insulin resistance. Biomolecules. 2021;11:1414. doi: 10.3390/biom11101414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Nie C, He T, Zhang W, Zhang G, Ma X. Branched chain amino acids:beyond nutrition metabolism. Int J Mol Sci. 2018;19:954. doi: 10.3390/ijms19040954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Herman MA, She P, Peroni OD, Lynch CJ, Kahn BB. Adipose tissue branched chain amino acid (BCAA) metabolism modulates circulating BCAA levels. J Biol Chem. 2010;285:11348–11356. doi: 10.1074/jbc.M109.075184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Pedersen HK, Gudmundsdottir V, Nielsen HB, et al. MetaHIT Consortium. Human gut microbes impact host serum metabolome and insulin sensitivity. Nature. 2016;535:376–381. doi: 10.1038/nature18646. [DOI] [PubMed] [Google Scholar]
- 86.Liu R, Hong J, Xu X, et al. Gut microbiome and serum metabolome alterations in obesity and after weight-loss intervention. Nat Med. 2017;23:859–868. doi: 10.1038/nm.4358. [DOI] [PubMed] [Google Scholar]
- 87.DeBerardinis RJ, Chandel NS. Fundamentals of cancer metabolism. Sci Adv. 2016;2:e1600200. doi: 10.1126/sciadv.1600200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Pham VT, Dold S, Rehman A, Bird JK, Steinert RE. Vitamins, the gut microbiome and gastrointestinal health in humans. Nutr Res. 2021;95:35–53. doi: 10.1016/j.nutres.2021.09.001. [DOI] [PubMed] [Google Scholar]
- 89.Chang YL, Rossetti M, Vlamakis H, et al. A screen of Crohn's disease-associated microbial metabolites identifies ascorbate as a novel metabolic inhibitor of activated human T cells. Mucosal Immunol. 2019;12:457–467. doi: 10.1038/s41385-018-0022-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Pham VT, Fehlbaum S, Seifert N, et al. Effects of colon-targeted vitamins on the composition and metabolic activity of the human gut microbiome- a pilot study. Gut Microbes. 2021;13:1–20. doi: 10.1080/19490976.2021.1875774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Corbett AJ, Eckle SB, Birkinshaw RW, et al. T-cell activation by transitory neo-antigens derived from distinct microbial pathways. Nature. 2014;509:361–365. doi: 10.1038/nature13160. [DOI] [PubMed] [Google Scholar]
- 92.Godfrey DI, Koay HF, McCluskey J, Gherardin NA. The biology and functional importance of MAIT cells. Nat Immunol. 2019;20:1110–1128. doi: 10.1038/s41590-019-0444-8. [DOI] [PubMed] [Google Scholar]
- 93.Deng K, Xu JJ, Shen L, et al. Comparison of fecal and blood metabolome reveals inconsistent associations of the gut microbiota with cardiometabolic diseases. Nat Commun. 2023;14:571. doi: 10.1038/s41467-023-36256-y. [DOI] [PMC free article] [PubMed] [Google Scholar]