

Abstract
Folate is a crucial nutrient that supports physiological functions. Low folate levels is a risk factor for several diseases, including cardiovascular diseases and neural tube defects. The most used folate supplement is folic acid, a synthetic oxidative form, and folic acid grain fortification is a success story of public health. However, the metabolic conversion of folic acid to bioactive tetrahydrofolate requires several enzymes and cofactors. Therefore, these factors influence its bioavailability and efficacy. In contrast, 5-methyltetrahydrofolate is used directly and participates in one-carbon metabolism, and the use of 5-methyltetrahydrofolate as an alternative folate supplement has increased. The metabolism of 5-methyltetrahydrofolate is primarily dependent on the transmembrane transporter, reduced folate carrier (RFC), and the RFC gene SLC19A1 variant is a functional polymorphism that affects folate status indexes. Recent studies demonstrated that the expression of RFC and cystathionine β-synthase, another enzyme required for homocysteine clearance, increases significantly by supplementation with calcitriol (vitamin D3), suggesting that calcitriol intake promotes the bioavailability of folate and has synergistic effects in homocysteine clearance. The advancements in biomedical and cohort studies and clinical trials have enhanced our understanding of the critical roles of folate and the regulation of one-carbon metabolism. We anticipate that the field of folate supplementation is poised to evolve from one size for all to personalized, precision, poly-paths (3Ps), which is a critical measure to meet individual needs, maximize health benefits, and minimize side effects.
Key words: folate, homocysteine, 5-methyltetrahydrofolate, vitamin D, precision nutrition
Introduction
Folate is the generic term for the water-soluble B-complex vitamin B9. The term folate stems from the Latin word folium, which means leaf; thus, folates are present in substantial amounts in green leafy vegetables.[1] Many vegetables and legumes are rich sources of folate, with the concentrations being around 200 pg/ 100 g in leafy vegetables and up to 600 pg/100 g in some beans and chickpeas. In contrast, meat and meat products, except for the liver, contain negligible amounts of folate.[2] The most oxidized and stable form is folic acid, a synthetic form of folate, which is composed of three covalently linked components: a pteridine ring, p-aminobenzoic acid, and a glutamate residue.[3] Mammals can synthesize the pteridine ring, but cannot couple this ring to other compounds, and are thus dependent on folate uptake from exogenous sources.[4]
Adequate folate intake is essential, as folate is a crucial cofactor in one-carbon metabolism and plays a vital role in methylation reactions. Folate deficiency is associated with elevated plasma concentrations of homocysteine (Hcy), which is demonstrated to be a risk factor or marker of many diseases and disorders.
The purpose of this review is to summarize the advancements in folate supplementation and fortification over the past two decades and the latest viewpoints from experimental and clinical studies, which provide an evolutionary history of precision nutrition.
Overview: The Functions and Requirements of Folate
Folate is a water-soluble B vitamin that plays a critical role in nucleic acid biosynthesis, DNA repair, and methylation.[5] Adequate folate intake is vital for cell division and homeostasis because folate coenzymes play essential roles in nucleic acid synthesis, methionine regeneration, and the shuttling, oxidation, and reduction of one-carbon units required for normal metabolism and regulation.[6] Low folate status is associated with elevated plasma Hcy, which are both risk factors for cardiovascular disease,[7,8] stroke,[9,10] megaloblastic anemia, neural tube defects (NTDs; congenital malformation of the fetus),[11] depression,[12] Alzheimer’s disease,[13,14] and tumors.[15]
Food folates exist as pteroyl polyglutamates and must be hydrolyzed in the gut to monoglutamates before absorption.[16] 5-Methyltetrahydrofolate (5-MTHF) is the predominant form of dietary folate and the only form of folate normally found in circulation. Thus, this is the form that is typically transported to the peripheral tissues for use in cellular metabolism. 5-MTHF accounts for approximately 98% of folate in human plasma.[17]
The daily Dietary Reference Intake of folate for adults is 400 μg of dietary folate equivalents.[18] However, many people do not have sufficient natural folate in their diet, especially populations with high folate demands, such as pregnant and lactating women. Historically, folic acid deficiency has been a global problem, and even today, it remains a severe problem in countries where dietary supplementation is not mandatory. Maintaining adequate folic acid levels requires sufficient intake, efficient intestinal absorption, and a specific transport system to achieve optimal tissue distribution.[19] Folic acid supplementation and fortification are the first choices to target folate deficiency as the most effective direct approach.
Metabolic Pathway of Folate
One-carbon metabolism encompasses a complex metabolic network based on the biochemical reactions of folate compounds. Folic acid is a hub of one-carbon metabolism that activates and transfers methyl groups for biosynthesis, including purine and thymidine syntheses.[20] However, folic acid has no coenzyme activity and must be reduced to the metabolically active form, 5-MTHF, through a series of steps. First, folic acid is reduced to dihydrofolate (DHF) and then tetrahydrofolate (THF) by dihydrofolate reductase (DHFR), which is converted to the bioactive form 5-MTHF by methylenetetrahydrofolate reductase (MTHFR).[21]
Hcy is a sulfur-containing, non-protein, vascular, toxic amino acid located at a branch point within the one-carbon metabolic pathway. Hcy is metabolized via two different pathways: (i) irreversibly degradation via the transsulfuration pathway to cysteine and (ii) remethylated back to methionine via the remethylation pathway (Figure 1). The latter process is disrupted by insufficient or metabolic abnormalities of folic acid levels and a deficiency in vitamin B12, which can eventually lead to hyperhomocysteinemia. As confirmed by epidemiological studies, high Hcy levels increase the risk of hypertension significantly and cardiovascular and cerebrovascular diseases.[8,22]
Figure 1.
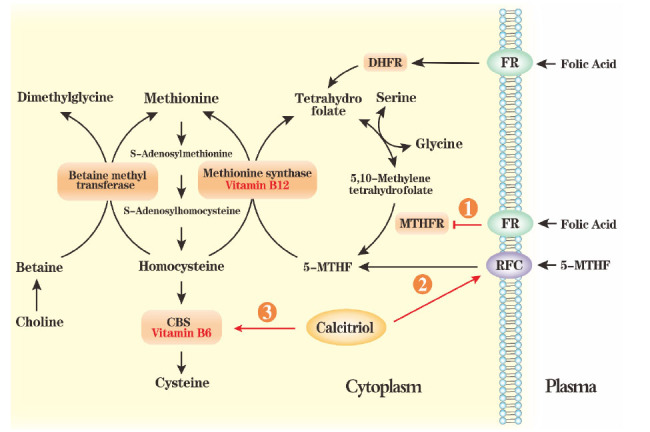
An overview of one-carbon metabolism and the folate cycle. One-carbon metabolism mainly involves the folate and homocysteine cycles. Plasma folic acid enters the tissue cells via FR, and then DHFR converts it into tetrahydrofolate. Next, tetrahydrofolate is transformed into 5,10-methylenetetrahydrofolate. Then, MTHFR converts 5,10-methylenetetrahydrofolate into 5-MTHF, providing a methyl group for converting homocysteine into methionine in a reaction catalyzed by MTR, which needs vitamin B12 as a cofactor. 5-MTHF enters into cells via RFC. Betaine, the end-product of oxidative metabolism of choline, is an alternative methyl donor for methylating homocysteine, which is catalyzed via BHMT. In the transfer-sulfuration pathway, CBS produces cysteine, which requires vitamin B6 as a coenzyme mediator. ①: Folic acid as an inhibitor of MTHFR; ②: Calcitriol as an enhancer of RFC; ③: Calcitriol as an activator of CBS. 5-MTHF: 5-methyl-tetrahydrofolate; BMT: betaine–homocysteine methyltransferase; CBS: cystathionine β-synthase; DHFR: dihydrofolate reductase; FR: folate receptor; MTHFR: methylenetetrahydrofolate reductase; MTR: methionine synthase; RFC: reduced folate carrier.
In the transfer-sulfuration pathway, cysteine is produced by the action of cystathionine β-synthase (CBS), a process that requires vitamin B6 as a coenzyme mediator.[23,24] 5-MTHF serves as a methyl donor in the remethylation pathway. Under the action of methionine synthetase and with vitamin B12 as a coenzyme, Hcy is methylated to methionine[25] and further transformed into S-adenosyl methionine (SAM). SAM is an essential active methyl donor and the second most common enzyme cofactor after adenosine triphosphate (ATP). SAM plays a significant role in epigenetics and biosynthetic processes, including phosphatidylcholine, creatine and polyamine syntheses, and sulfur metabolism.[26,27] This cross-linking of folic acid and methionine cycles results in an inverse relationship between Hcy and folic acid levels. Thus, plasma Hcy is regarded as a functional indicator of folate status in the body.[28]
The Northern Manhattan Study affirmed that elevated total Hcy (tHcy) is a significant and robust predictor for ischemic stroke and vascular death.[29] Clearance of tHcy conferred health benefits at all levels, but the lowest tHcy levels enjoyed the most significant risk reduction.
Folic Acid Supplements
Folic acid (pteroylmonoglutamate) is absorbed, whereas food folates (polyglutamate derivatives) must be hydrolyzed to monoglutamates in the gut by a brush border hydrolase before absorption.
Folate supplements include their oxidized form, folic acid, and reduced form, mainly 5-MTHF or 5-MTHF calcium salt, which is commercially available as a crystalline form. Food folates exist primarily as 5-MTHF and formyltetrahydrofolate; however, native folates are generally polyglutamates, where each glutamate residue is linked to another by a γ-glutamyl bond. Polyglutamyl folates are hydrolyzed to folylmonoglutamates by pteroyl-γ-glutamyl hydrolase and then metabolized within enterocytes to 5-MTHF.[30]
Folic acid is used widely as a supplement and in food fortification because of its low price, high thermostability, and high bioavailability.[31] Nonetheless, folic acid lacks coenzyme activity and must be reduced to the metabolically active form, tetrahydrofolate, within cells. 5-MTHF is the predominant form of dietary folate and the only species normally detected in the circulation and used for cellular metabolism.
5-MTHF and folic acid have similar physiological activity, bioavailability, and absorption at equimolar doses. Bioavailability studies have measured blood concentrations of folate and functional indicators of folate status and plasma Hcy and concluded that 5-MTHF is at least as effective as folic acid in improving the folate status.
Some countries have mandated the fortification of flour with folic acid or other vitamins and minerals (USA 1.4 mg/kg from 1998; Canada 1.5 mg/kg from 1998; Chile 2.2 mg/kg from 2000).[32] Currently, 92 countries have legislation to mandate fortification of at least one industrially milled cereal grain (Table 1); 91 of these countries requested wheat flour fortification, either alone or with other grains.[33] The benefits of folic acid supplementation are apparent: in the USA, Canada, and Chile, mandatory fortification of flour substantially improved folate and Hcy status and the NTDs’ rates fell by between 31% and 78%.[34] A recent study estimated that 65,380 congenital disabilities of the brain and spine were prevented in 2019 because the flour was fortified with folic acid.[35] This causality was further demonstrated in an observational study that identified significant reduction in the risk of congenital heart disease and cleft lip and palate in children whose mothers received folic acid supplements or multivitamins.[36]
Table 1.
Mandatory fortified cereal grain and the number of countries with corresponding legislation – July 2022[30]
| Mandatory fortified cereal grain | Number |
|---|---|
| Wheat flour | 67 |
| Wheat flour, maize flour | 17 |
| Wheat flour, rice | 5 |
| Wheat flour, maize flour, rice | 2 |
| Rice | 1 |
Over the last two decades, folic acid supplementation doses have been defined using general population characteristics. During this period, official agencies such as the World Health Organization (WHO) and the European Food Safety Agency (EFSA) specifically distinguished between the fortification requirements of different groups, including infants, children, adults, pregnant women, and others. Another consideration for folic acid dosage recommendation is the presence or lack of supplementation or food fortification.[37] Alternatively, some studies have used Hcy as an evaluation index for supplemental folic acid dosage.[28,38]
As previously mentioned, vitamin B6 and B12 participate as coenzymes in the metabolism of folic acid and Hcy. For this reason, folic acid supplementation alone may not be able to maximize the reduction of Hcy levels. Related studies have explored the optimization of folic acid status indicators with folic acid supplementation, as well as vitamin B6 and B12. The results showed that at 0.8 mg/d folic acid, the Hcy levels were reduced by 23%. Additional supplementation with vitamin B12 was associated with a further reduction in Hcy by 7%.[35] Another study reported that folic acid supplementation lowered fasting tHcy by 19.6% (P < 0.001), and that following folic acid supplementation, low-dose vitamin B6 lowered fasting plasma tHcy by 7.5%.[39] A systematic meta-analysis of prospective studies to examine the relationship between dietary (from diet and supplements) intake of these B vitamins and the risk of stroke (with 12 prospective studies comprising 389,938 participants and 10,749 cases included in the final analysis) indicated that both vitamin B6 and folate intake were inversely correlated with the risk of stroke. Each 0.5 mg/d increase in vitamin B6 intake reduced the risk of stroke by 6%.[40] This observation shows that when used in combination with vitamin B12, B6, and other B vitamins, folic acid can achieve a greater level of Hcy reduction than when used alone.
Reduced Folate Supplement
Previous studies have indicated that single-nucleotide polymorphisms (SNPs) in genes encoding enzymes of one-carbon metabolism are crucial determinants of Hcy concentration.[41] A recent study confirmed that one-carbon metabolism gene variants affect the response to folate treatment.[42]
In 1995, Frosst et al.[43] identified a common mutation in MTHFR : a C to T substitution at bp 677. This mutation alters a highly conserved amino acid, and in both heterozygous and homozygous individuals, it causes a reduction in enzyme activity and increased thermolability. Individuals homozygous for the mutation (MTHFR 677TT) have significantly elevated Hcy[43,44] and altered folate distribution within red blood cells.[44] An increased circulating concentration of Hcy is associated with a higher risk of stroke.[9] Further investigations have shown that the mutation frequency of C677T is higher among the Chinese population when compared to the European and American populations.[45] Research has suggested that TT individuals require higher folate intake than those with the CT or CC genotype to counterbalance the effect of the MTHFR 677 mutation on tHcy levels and achieve similar tHcy concentrations.[45,46] The consensus is that the precise folic acid supplementation dose should be based on individual genotypes. Although accurate precision supplementation has not yet been realized, progress has been made through the subdivision of populations by age and genotype combined with other factors.
The average dose of folic acid expected from food fortification in the USA and Canada is approximately 100 μg/d, which is mainly converted to 5-MTHF.[3] However, an important issue has emerged from research on folic acid supplementation: above 200 μg/d, small amounts of folic acid appear unchanged in the circulation.[47] Thus, people taking supplements or consuming fortified foods may face a buildup of unmetabolized folic acid (UMFA).[48] Before the body uses folic acid, it must first be reduced to DHF and subsequently to THF. Production of DHF and THF is catalyzed by DHFR in the liver.[49] DHFR activity in the human liver has been shown to be low and highly variable.[50] This observation points to a potentially limited ability to activate the synthetic form of folate and raises concerns about the clinical trials using high levels of folic acid (ranging from 0.5 to 15.0 mg/d).[51] The extremely low rate of conversion of folic acid suggests that the potential benefits of its use in high doses are outweighed by saturation of DHFR, especially in individuals with lower-than-average DHFR enzyme activity.[50] DHFR is saturated when folic acid intake exceeds a physiological threshold and conversion to the reduced form is impaired, resulting in the appearance of UMFA in circulation.[25] The most publicized safety risk concerns the correcting and masking neurologic sequelae of cobalamin deficiency in patients exposed to UMFA, as UMFA may mask the underlying disease.[52, 53, 54]
Additionally, the presence of UMFA may cause accumulation of DHF. In vitro, DHF can inhibit the activities of MTHFR and thymidylate synthase, two enzymes required for one-carbon metabolism, theoretically leading to paradox functional folate deficiency.[15] Given these risks, there has been growing interest in determining the amount of dietary folic acid required for UMFA to accumulate within the body. In one study, participants on a 5-day regimen of fortified, ready-to-eat-cereal and bread added to their regular diet had a threshold intake of 266 μg folic acid per meal for UMFA to appear in serum.[47] Another study found that 1 mg of folic acid consumed in 10 equal doses of 100 μg also caused folic acid to appear in serum.[55] Thus, UMFA was detected in people consuming either folic acid-fortified foods or supplements.[48,56]
Given the potential downsides of folic acid supplementation, there is a clear need for an alternative solution. Rather than using synthetic folate, that is, folic acid, a more direct option is to supplement with an already bioactive reduced form, such as 5-MTHF, which is the most available folate form in human plasma[57] and constitutes 95%–98% of folate in serum.[17,58]
5-MTHF has many advantages over folic acid as a supplement. 5-MTHF participates directly in one-carbon metabolism without activation. Thus, 5-MTHF supplements should bypass the limitation of low DHFR activity and the decreased efficiency caused by several common polymorphisms of MTHFR and other enzymes. Unlike folic acid, data suggest that 5-MTHF will not mask vitamin B12 deficiency. 5-MTHF is also more effective than folic acid supplementation in improving folate status.[59] Studies on the safety, tolerability, and retention rates of 5-MTHF/5-MTHF-Ca have suggested that 5-MTHF-Ca is a safe alternative to folic acid as a source of folate and may be particularly advantageous for individuals with MTHFR defects, who could have difficulty processing folic acid from supplements or fortified foods.[60,61] Importantly, a study demonstrated that after administering 5-MTHF, UMFA was not detected in the plasma of women who were of MTHFR 677CT genotype.[62] Products supplementing the active form of folic acid are also becoming increasingly abundant.
Folate Transporters
As a coenzyme, folate is distributed primarily in the cytosol and mitochondria of liver cells, where one-carbon metabolism occurs.[21] This requires folate molecules to cross membranes into these compartments. Because of the hydrophilic nature of the charged folate molecule, there is minimal passive diffusion across cell membranes. Instead, specific transporters are required to facilitate intestinal folate absorption and the transport of folates into systemic tissues.[19]
Three folate transporters account for folate influx activities observed in mammalian cells.
(1) Folate receptors (FRs) are membrane proteins with an endocytic transport mechanism that have a high affinity for folic acid (approximately 1 nmol/L) and a lower affinity for reduced folates.[63] The genes encoding FR-α, -β, and -γ are located on chromosome 11 (q11.3–q13.5).[64] FR-α is expressed in epithelial tissues. FR-β is expressed in cells of hematopoietic origin.[65] FR-γ expression has been detected by polymerase chain reaction (PCR) analysis in normal and malignant hematopoietic cells present in the spleen, bone marrow and thymus, ovarian, cervical, and uterine carcinoma.[66] The affinity of FRs for reduced folate cofactors and methotrexate (MTX) is approximately threefold and 100-fold lower, respectively, when compared to the affinity of FRs for folic acid.
(2) Proton-coupled folate transporter (PCFT) is a member of the superfamily of facilitative carriers (SLC46A1). From a physiological perspective, PCFT is required for intestinal folate absorption and transport of folates across the choroid plexus–cerebrospinal fluid barrier. PCFT has a very high affinity for folic acid at pH 5.5 and a low folic acid transport activity at pH 7.4.[67]
(3) Reduced folate carrier (RFC; SLC19A1) is an anion exchanger with a high affinity for reduced folates (Km of 1–5 μmol/L) and a low affinity for folic acid (Km of 200–400 μmol/L), with optimum activity at physiological pH.[63]
The human RFC gene is located on chromosome 21q22.3. Five exons encode RFC, which consists of 591 amino acids.[68] The gene for this transporter is ubiquitously expressed in human tissues.[69] RFC is a bidirectional folate transporter, functions as an anion exchanger, and is a major uptake route for transporting folates into mammalian cells at ambient neutral pH.[70] RFC is a member of the solute carrier family (SLC) of facilitative carriers, which currently comprises 300 genes that have been classified into 43 subfamilies.[1] The SLC family belongs to an even larger superfamily known as the major facilitator superfamily of transporters.[71]
5-MTHF, like all folates, is hydrophilic and incapable of permeating plasma membranes by diffusion alone. RFC is a high-capacity, bidirectional transporter of 5-MTHF, which mediates the transport of 5-MTHF from blood into the cells of peripheral tissues.[72,73]
Regulation of Folate Transporters
Although 5-MTHF has apparent advantages over folic acid as a supplement, 5-MTHF is not a perfect solution. The RFC gene SLC19A1 variant has been reported to have functional SNPs; the most extensively studied variant is G80A. This base change results in the exchange of arginine with histidine at residue 27 in the amino acid sequence. A study has shown that the RFC G80A polymorphism can impact folate status.[73] In this study, women with the 80GA and 80AA genotypes had higher red blood cell folate concentrations when compared to women with the 80GG genotype,[73] which is consistent with another study demonstrating that after stratification by the C677T genotype, individuals who were 80AA/677CT had higher plasma folate levels than those who were 80GG/677CT (P = 0.02).[74] The RFC G80A polymorphism is ubiquitous among several ethnic populations. The G and A alleles at base 80 are distributed almost equally among Ashkenazi Jews, Hispanics, and Caucasians, with the A frequency ranging from 47.2% to 56.4%.[75]
Unfortunately, there is a paucity of data to illustrate the exact relationship between the G80A polymorphism of RFC and the folate status after 5-MTHF supplementation. We hypothesize that as variable activities of DHFR and polymorphisms of MTHFR and other enzymes are bypassed by supplementation with reduced folate, RFC, as an essential functional carrier of reduced folate, may become an important factor in defining dosage selection with 5-MTHF supplementation.
Indeed, the expression and regulation of RFC may interact with folate levels. One study showed that folate deficiency increased mRNA synthesis for folate transporters, including RFC and PCFT. This seems to be the underlying transcriptional regulatory mechanism controlling the expression of folate transporters,[76] which may be a maintenance mechanism for folate metabolism homeostasis. Another study demonstrated that activation of vitamin D receptor (VDR) through calcitriol (1,25-dihydroxyvitamin D3) exposure upregulates RFC mRNA and protein expression and function in immortalized cultures of human cerebral microvascular endothelial cells (hCMEC/D3). Moreover, in isolated mouse brain capillaries, the role of VDR in the direct regulation of RFC was further confirmed. Subsequently, an in vivo study using Folr1-knockout (Folr1 KO) mice showed that a loss of FR-α led to a substantial decrease of folate delivery to the brain, and that pretreatment of Folr1 KO mice with the VDR-activating ligand, calcitriol, resulted in an over sixfold increase in the concentration of [13C5]-5-formyltetrahydrofolate ([13C5]-5-formyl THF) in brain tissues, with levels comparable to wild-type animals.[77,78]
The transsulfuration pathway is necessary for Hcy clearance and culminates in cysteine synthesis. The first step of this metabolic pathway is the condensation of Hcy and serine to form cystathionine, which is catalyzed by the vitamin B6-dependent enzyme CBS.[79] In one study, a microarray experiment on MC3T3-E1 murine preosteoblasts treated with calcitriol revealed that the transcription and expression and the enzymatic function of CBS were rapidly and strongly induced by calcitriol.[80]
Calcitriol acts as a ligand for the nuclear transcription factor, VDR, regulating gene transcription and cell function. Calcitriol was found to regulate the transcription of about 3% of mouse and human genomes. Calcitriol metabolites may also function as hormones or cytokines for many cellular functions like cell cycle control and differentiation.[81]
These results suggest that calcitriol supplementation may enhance the bioavailability and bioefficacy of folate by improving its transport and activating one of the metabolic pathways for Hcy clearance. Therefore, folate combined with vitamin D intervention may be a therapeutic strategy to increase the absorption and distribution of folate in tissues of individuals with poor RFC activity or folate metabolism defects and promote the reduction of Hcy. This novel intervention aims to optimize folate supplementation by increasing its absorption and capacity in the body.
Folate Supplement in Special Gro
Folate supplementation for MTX-treated patients To establish individualized medication for patients receiving specific drug treatments, some issues need to be illustrated based on research data in this field.
MTX is a folate antagonist and a well-established therapy for autoimmune and cancer patients.[82,83] RFC is the major route of folate and MTX uptake from extracellular fluids. A study showed that RFC mRNA expression levels are more important as a factor linked to MTX efficacy, and individual differences in RFC expression levels among subjects were observed. Therefore, individual differences in MTX treatment efficacy may be related to individual differences in RFC mRNA expression levels. Moreover, responsiveness to MTX may be predictable based on these individual differences in RFC mRNA expression.[84] Additionally, the most studied SNP of RFC, G80A, was associated with altered MTX concentrations in leukemia patients treated with MTX,[85] suggesting that the RFC 80A allele may potentiate the effect of low-dose MTX in patients with rheumatoid arthritis.[86]
In high doses, as used in cancer chemotherapy, MTX depletes the intracellular folate stores and thus disrupts nucleotide synthesis. MTX treatment is associated with significant side effects and toxicity in some patients. Folate supplementation is often used to ameliorate MTX-associated side effects and toxicities. According to studies reviewed, the use of folate supplements in patients treated with MTX reduces the incidence of hepatotoxicity and gastrointestinal intolerance without impairing the efficacy of MTX.
Furthermore, studies reported that MTX treatment may lead to increased plasma Hcy concentrations, and concomitant folate supplementation decreases plasma Hcy levels.[87] Moreover, a study showed that low-dose MTX of 7.5–10.0 mg per week induces a significant rise in plasma Hcy during the first 4 weeks of treatment in arthritis patients without folic acid deficiency.[88] However, relatively few studies have addressed folate supplementation with the use of MTX for treating psoriasis. Thus, folate supplementation has been recommended for every patient who receives MTX.[89]
In summary, folate reduces the adverse effects and toxicities without affecting the efficacy in studies of rheumatoid arthritis. The use of folate may increase the likelihood of efficacious long-term, tolerable, and toxicity-free therapy for patients receiving MTX.
Folate supplementation for epileptic patients
The interaction between cytochrome P450 enzyme-inducing antiepileptic drugs (P450-inducing antiepileptic drugs [AEDs]) and folate metabolism may be clinically relevant and balanced by supplementation with small doses of folic acid. The results of a survey support the recommendation that women of childbearing age treated with AEDs receive folic acid supplementation, particularly those taking P450-inducing AEDs.[90]
Patients with epilepsy exhibit elevated plasma tHcy levels more frequently than the general population, which is caused by chronic treatment with older AEDs and polymorphisms in the MTHFR gene. Epileptic patients exhibit, in a percentage range of 20%–40%, supraphysiological plasma levels of tHcy; however, this tHcy level is only observed in approximately 5% of the general population.[91] In another study, epileptic children with hyper-tHcy had significantly lower folate and cobalamin concentrations.[92]
Among the various variables analyzed, hyper-tHcy has been indicated as a risk factor for the progression of atherosclerosis in epileptic patients.[93, 94, 95, 96, 97, 98] Noteworthy, literature data suggest that patients with epilepsy also exhibit an increased risk for stroke.[99] In this sense, the definition of a standardized clinical assessment is essential for developing new therapeutic strategies, including folate and other B vitamin supplements, and reducing morbidity and mortality in epilepsy patients under long-term therapy.
Results have shown that the condition of hyper-tHcy can be easily corrected by the intake of folate and other B vitamins. This raises the question of how folic acid and/or B vitamins are provided to patients with epilepsy. Belcastro et al.[100] summarized previous studies and claimed that supplementation of 0.4 mg/ d folic acid plus a low B vitamin dose (i.e., B6, B12) is an option to treat or avoid consequences of folate deficiency in patients on chronic treatment with older AEDs.
The above two cases reveal that people treated with drugs for nervous system disorders need carefully regulated and/or measured folate supplements to ensure benefits from them, when compared to the folate supplements used by the general population. Although the functional indicator of folate status, Hcy, is influenced by various factors such as genetic or environmental, as an independent risk of many disorders, monitoring Hcy levels deserves more attention by clinicians. Diversified and personalized solutions for this issue are promising, and significant progress and advances in this research field continue to be made.
Conclusion
Over the past 20 years, universal folic acid grain fortification in many countries, including the USA, has been a success story in medicine and public health. Rapid advances in biomedical sciences (e.g., human genetics, epigenetics, proteomics, metabolomics) and high-throughput biotechnology, along with large-scale cohort studies and clinical trials, have greatly enhanced our understanding of the critical role and regulation of folate in the context of one-carbon metabolism. We anticipate that the field of folate supplementation is poised to evolve from one size for all to personalized, precision, poly-paths (3Ps). This evolution is critically needed to meet individual needs, maximize health benefits, and minimize adverse effects. Well-designed clinical trials and prospective cohort studies will be required to deepen our knowledge and pave the road toward 3Ps. Findings from this line of research will be transformative and will shape the future of 3P folate nutrition.
Funding Statement
This work was supported by the following aspects: National Key Research and Development Program of China (2021YFC2500600/2021YFC2500601, to Li JP).
Footnotes
Author Contribution
He QQ, Li JP: Conceptualization; He QQ: Writing—Original draft preparation; He QQ, Li JP: Writing—Reviewing and Editing; Li JP: Supervision, Project administration.
Conflict of Interest
The authors declare no competing interest.
References
- 1.Ifergan I, Assaraf YG. Molecular mechanisms of adaptation to folate deficiency. Vitam Horm. 2008;79:99. doi: 10.1016/S0083-6729(08)00404-4. - [DOI] [PubMed] [Google Scholar]
- 2.Witthöft CM, Forssén K, Johannesson L, Jägerstad M.. Folates - food sources, analyses, retention and bioavailability. Näringsforskning. 2016;43:138. - [Google Scholar]
- 3.Pietrzik K, Bailey L, Shane B. Folic acid and l-5-methyltetrahydrofolate: Comparison of clinical pharmacokinetics and pharmacodynamics. Clin Pharmacokinet. 2010;49:535. doi: 10.2165/11532990-000000000-00000. - [DOI] [PubMed] [Google Scholar]
- 4.Birn H. The kidney in vitamin b12 and folate homeostasis: Characterization of receptors for tubular uptake of vitamins and carrier proteins. Am J Physiol Renal Physiol. 2006;291:22. doi: 10.1152/ajprenal.00385.2005. - [DOI] [PubMed] [Google Scholar]
- 5.Bailey LB, Gregory JF.. Folate metabolism and requirements. J Nutr. 1999;129:779. doi: 10.1093/jn/129.4.779. - [DOI] [PubMed] [Google Scholar]
- 6.Oaks BM, Dodd KW, Meinhold CL, Jiao L, Church TR, Stolzenberg-Solomon RZ. Folate intake, post–folic acid grain fortification, and pancreatic cancer risk in the prostate, lung, colorectal, and ovarian cancer screening trial. Am J Clin Nutr. 2010;91:449. doi: 10.3945/ajcn.2009.28433. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Moat SJ, Lang D, McDowell IF, Clarke ZL, Madhavan AK, Lewis MJ. Folate, homocysteine, endothelial function and cardiovascular disease. J Nutr Biochem. 2004;15:64. doi: 10.1016/j.jnutbio.2003.08.010. et al. - [DOI] [PubMed] [Google Scholar]
- 8.Wald DS, Law M, Morris JK. Homocysteine and cardiovascular disease: Evidence on causality from a meta-analysis. BMJ. 2002;325:1202. doi: 10.1136/bmj.325.7374.1202. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Casas JP, Bautista LE, Smeeth L, Sharma P, Hingorani AD. Homocysteine and stroke: Evidence on a causal link from mendelian randomisation. Lancet. 2005;365:224. doi: 10.1016/S0140-6736(05)17742-3. - [DOI] [PubMed] [Google Scholar]
- 10.Li R, Weng H, Pan Y, Meng X, Liao X, Wang M. Relationship between homocysteine levels and post-stroke cognitive impairment in female and male population: from a prospective multicenter study. J Transl Intern Med. 2021;9:264. doi: 10.2478/jtim-2021-0035. et al . - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Stanger O. Physiology of folic acid in health and disease. Curr Drug Metab. 2002;3:211. doi: 10.2174/1389200024605163. - [DOI] [PubMed] [Google Scholar]
- 12.Alpert JE, Fava M. Nutrition and depression: The role of folate. Nutr Rev. 1997;5:145. doi: 10.1111/j.1753-4887.1997.tb06468.x. - [DOI] [PubMed] [Google Scholar]
- 13.Flaubert T, Shea TB. Folate deprivation, the methionine cycle, and alzheimer’s disease. Vitam Horm. 2008;79:83. doi: 10.1016/S0083-6729(08)00403-2. - [DOI] [PubMed] [Google Scholar]
- 14.Kim T. Can gamma entrainment of the brain rhythms prevent or alleviate Alzheimer’s disease? J Transl Intern Med. 2021;9:231. doi: 10.2478/jtim-2021-0048. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kawakita D, Lee YA, Gren LH, Buys SS, La Vecchia C, Hashibe M. The impact of folate intake on the risk of head and neck cancer in the prostate, lung, colorectal, and ovarian cancer screening trial (plco) cohort. Br J Cancer. 2018;118:299. doi: 10.1038/bjc.2017.383. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Scaglione F, Panzavolta G. Folate, folic acid and 5-methyltetrahydrofolate are not the same thing. Xenobiotica. 2014;44:480. doi: 10.3109/00498254.2013.845705. - [DOI] [PubMed] [Google Scholar]
- 17.Page R, Robichaud A, Arbuckle TE, Fraser WD, MacFarlane AJ. Total folate and unmetabolized folic acid in the breast milk of a cross-section of canadian women. Am J Clin Nutr. 2017;105:1101. doi: 10.3945/ajcn.116.137968. - [DOI] [PubMed] [Google Scholar]
- 18.Suitor CW, Bailey, Lynn B. Dietary folate equivalents: Interpretation and application. J Am Diet Assoc. 2000;100:88. doi: 10.1016/S0002-8223(00)00027-4. - [DOI] [PubMed] [Google Scholar]
- 19.Zhao R, Matherly LH, Goldman ID. Membrane transporters and folate homeostasis: Intestinal absorption and transport into systemic compartments and tissues. Expert Rev Mol Med. 2009;11:e4. doi: 10.1017/S1462399409000969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Locasale JW. Serine, glycine and one-carbon units: Cancer metabolism in full circle. Nat Rev Cancer. 2013;13:572. doi: 10.1038/nrc3557. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ducker GS, Rabinowitz JD. One-carbon metabolism in health and disease. Cell Metab. 2017;25:27. doi: 10.1016/j.cmet.2016.08.009. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Blom HJ, Smulders Y. Overview of homocysteine and folate metabolism. With special references to cardiovascular disease and neural tube defects. J Inherit Metab Dis. 2011;34:75. doi: 10.1007/s10545-010-9177-4. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ubbink JB, Merwe Avd, Delport R, Allen RH, Stabler SP, Riezler R. The effect of a subnormal vitamin b-6 status on homocysteine metabolism. J Clin Invest. 1996;98:177. doi: 10.1172/JCI118763. et al. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mattson MP, Shea TB. Folate and homocysteine metabolism in neural plasticity and neurodegenerative disorders. Trends Neurosci. 2003;26:13746. doi: 10.1016/S0166-2236(03)00032-8. [DOI] [PubMed] [Google Scholar]
- 25.Obeid R, Herrmann W. The emerging role of unmetabolized folic acid in human diseases: Myth or reality? Curr Drug Metab. 2012;13:1184. doi: 10.2174/138920012802850137. - [DOI] [PubMed] [Google Scholar]
- 26.Su X, Wellen KE, Rabinowitz JD. Metabolic control of methylation and acetylation. Curr Opin Chem Biol. 2016;30:52. doi: 10.1016/j.cbpa.2015.10.030. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zhu M, Ding Q, Lin Z, Chen X, Chen S, Zhu Y. New insights of epigenetics in vascular and cellular senescence. J Transl Intern Med. 2021;9:239. doi: 10.2478/jtim-2021-0049. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Wald DS, Bishop L, Wald NJ, Law M, Hennessy E, Weir D. Randomized trial of folic acid supplementation and serum homocysteine levels. Arch Intern Med. 2001;161:695. doi: 10.1001/archinte.161.5.695. et al. - [DOI] [PubMed] [Google Scholar]
- 29.Sacco RL, Anand K, Lee H-S, Boden-Albala B, Stabler S, Allen R. Homocysteine and the risk of ischemic stroke in a triethnic cohort: The northern manhattan study. Stroke. 2004;35:2263. doi: 10.1161/01.STR.0000142374.33919.92. et al. - [DOI] [PubMed] [Google Scholar]
- 30.Lucock M. Folic acid: Nutritional biochemistry, molecular biology, and role in disease processes. Mol Genet Metab. 2000;71:121. doi: 10.1006/mgme.2000.3027. - [DOI] [PubMed] [Google Scholar]
- 31.Molloy AM. Folate bioavailability and health. Int J Vitam Nutr Res. 2002;72:46. doi: 10.1024/0300-9831.72.1.46. - [DOI] [PubMed] [Google Scholar]
- 32.Wright AJA, Dainty JR, Finglas PM. Folic acid metabolism in human subjects revisited: Potential implications for proposed mandatory folic acid fortification in the uk. Br J Nutr. 2007;98:667. doi: 10.1017/S0007114507777140. - [DOI] [PubMed] [Google Scholar]
- 33.Food fortification initiative. 2022. https://www.ffinetwork.org/globalprogress. Available from. Accessed August 30, 2022.
- 34.Eichholzer M, Tönz O, Zimmermann R. Folic acid: A public-health challenge. Lancet. 2006;367:1352. doi: 10.1016/S0140-6736(06)68582-6. - [DOI] [PubMed] [Google Scholar]
- 35.Kancherla V, Wagh K, Pachón H, Oakley GP. A 2019 global update on folic acid-preventable spina bifida and anencephaly. Birth Defects Res. 2020;113:77. doi: 10.1002/bdr2.1835. - [DOI] [PubMed] [Google Scholar]
- 36.Bailey LB, Berry RJ. Folic acid supplementation and the occurrence of congenital heart defects, orofacial clefts, multiple births, and miscarriage. Am J Clin Nutr. 2005;81:1213S. doi: 10.1093/ajcn/81.5.1213. - [DOI] [PubMed] [Google Scholar]
- 37.Crider KS, Bailey LB, Berry RJ. Folic acid food fortification-its history, effect, concerns, and future directions. Nutrients. 2011;3:370. doi: 10.3390/nu3030370. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Collaboration HLT. Dose-dependent effects of folic acid on blood concentrations of homocysteine: A meta-analysis of the randomized trials. Am J Clin Nutr. 2005;82:806. doi: 10.1093/ajcn/82.4.806. - [DOI] [PubMed] [Google Scholar]
- 39.McKinley MC, McNulty H, McPartlin J, Strain J, Pentieva K, Ward M. Low-dose vitamin b-6 effectively lowers fasting plasma homocysteine in healthy elderly persons who are folate and riboflavin replete. Am J Clin Nutr. 2001;73:759. doi: 10.1093/ajcn/73.4.759. et al. - [DOI] [PubMed] [Google Scholar]
- 40.Chen L, Li Q, Fang X, Wang X, Min J, Wang F. Dietary intake of homocysteine metabolism-related b-vitamins and the risk of stroke: A dose-response meta-analysis of prospective studies. Adv Nutr. 2020;00:1. doi: 10.1093/advances/nmaa061. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Meurs JBv, Pare G, Schwartz SM, Hazra A, Tanaka T, Vermeulen SH. Common genetic loci influencing plasma homocysteine concentrations and their effect on risk of coronary artery disease. Am J Clin Nutr. 2013;98:668. doi: 10.3945/ajcn.112.044545. et al. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Li D, Zhao Q, Huang X, Zhang C, Godfrey O, Zhang W. Association of genetic and epigenetic variants in one-carbon metabolism gene with folate treatment response in hyperhomocysteinaemia. Eur J Clin Nutr. 2020;74:1073. doi: 10.1038/s41430-020-0611-x. - [DOI] [PubMed] [Google Scholar]
- 43.Frosst P, Blom HJ, Milos R, Goyette P, Sheppard CA, Matthews RG. A candidate genetic risk factor for vascular disease a common mutation in methylenetetrahydrofolate reductase. Nat Genet. 1995;10:111. doi: 10.1038/ng0595-111. et al. - [DOI] [PubMed] [Google Scholar]
- 44.Herrmann W, Obeid R, Schorr H, Zarzour W, Geisel J. Homocysteine, methylenetetrahydrofolate reductase c677t polymorphism and the b-vitamins: A facet of nature-nurture interplay. Clin Chem Lab Med. 2003;41:547. doi: 10.1515/CCLM.2003.083. - [DOI] [PubMed] [Google Scholar]
- 45.Huang X, Qin X, Yang W, Liu L, Jiang C, Zhang X. Mthfr gene and serum folate interaction on serum homocysteine lowering: Prospect for precision folic acid treatment. Arterioscler Thromb Vasc Biol. 2018;38:679. doi: 10.1161/ATVBAHA.117.310211. et al. - [DOI] [PubMed] [Google Scholar]
- 46.Ashfield-Watt PA, Pullin CH, Whiting JM, Clark ZE, Moat SJ, Newcombe RG. Methylenetetrahydrofolate reductase 677c→t genotype modulates homocysteine responses to a folate-rich diet or a low-dose folic acid supplement: A randomized controlled trial. Am J Clin Nutr. 2002;76:180. doi: 10.1093/ajcn/76.1.180. et al. - [DOI] [PubMed] [Google Scholar]
- 47.Kelly P, McPartlin J, Goggins M, Weir DG, Scott JM. Unmetabolized folic acid in serum: Acute studies in subjects consuming fortified food and supplements. Am J Clin Nutr. 1997;65:1790. doi: 10.1093/ajcn/65.6.1790. - [DOI] [PubMed] [Google Scholar]
- 48.Pfeiffer CM, Sternberg MR, Fazili Z, Yetley EA, Lacher DA, Bailey RL. Unmetabolized folic acid is detected in nearly all serum samples from us children, adolescents, and adults. J Nutr. 2015;145:520. doi: 10.3945/jn.114.201210. et al. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Patanwala I, King MJ, Barrett DA, Rose J, Jackson R, Hudson M. Folic acid handling by the human gut: Implications for food fortification and supplementation. Am J Clin Nutr. 2014;100:593. doi: 10.3945/ajcn.113.080507. et al. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Bailey SW, Ayling JE. The extremely slow and variable activity of dihydrofolate reductase in human liver and its implications for high folic acid intake. Proc Natl Acad Sci USA. 2009;106:15424. doi: 10.1073/pnas.0902072106. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Wang X, Qin X, Demirtas H, Li J, Mao G, Huo Y. Efficacy of folic acid supplementation in stroke prevention: A meta-analysis. Lancet. 2007;369:1876. doi: 10.1016/S0140-6736(07)60854-X. et al. - [DOI] [PubMed] [Google Scholar]
- 52.Schneider JA, Tangney CC, Morris MC. Folic acid and cognition in older persons. Expert Opin. Drug Saf. 2006;5:511. doi: 10.1517/14740338.5.4.511. - [DOI] [PubMed] [Google Scholar]
- 53.Selhub J, Paul L. Folic acid fortification: Why not vitamin b12 also? BioFactors. 2011;37:269. doi: 10.1002/biof.173. - [DOI] [PubMed] [Google Scholar]
- 54.Dickinson CJ. Does folic acid harm people with vitamin b12 deficiency? Q J Med. 1995;88:357. - [PubMed] [Google Scholar]
- 55.Sweeney MR, McPartlin J, Weir DG, Daly L, Scott JM. Postprandial serum folic acid response to multiple doses of folic acid in fortified bread. Br J Nutr. 2006;95:145. doi: 10.1079/bjn20051618. - [DOI] [PubMed] [Google Scholar]
- 56.Steluti J, Reginaldo C, Selhub J, Paul L, Fisberg RM, Marchioni DM. Presence of circulating folic acid in plasma and its relation with dietary intake, vitamin b complex concentrations and genetic variants. Eur J Nutr. 2019;58:3069. doi: 10.1007/s00394-018-1852-5. - [DOI] [PubMed] [Google Scholar]
- 57.Obeid R, Kasoha M, Kirsch SH, Munz W, Herrmann W. Concentrations of unmetabolized folic acid and primary folate forms in pregnant women at delivery and in umbilical cord blood. Am J Clin Nutr. 2010;92:1416. doi: 10.3945/ajcn.2010.29361. - [DOI] [PubMed] [Google Scholar]
- 58.Obeid R, Holzgreve W, Pietrzik K. Is 5-methyltetrahydrofolate an alternative to folic acid for the prevention of neural tube defects? J Perinat Med. 2013;41:469. doi: 10.1515/jpm-2012-0256. - [DOI] [PubMed] [Google Scholar]
- 59.Lamers Y, Prinz-Langenohl R, Bramswig S, Pietrzik K. Red blood cell folate concentrations increase more after supplementation with [6s]-5-methyltetrahydrofolate than with folic acid in women of childbearing age. Am J Clin Nutr. 2006;84:156. doi: 10.1093/ajcn/84.1.156. - [DOI] [PubMed] [Google Scholar]
- 60.Niederberger KE, Dahms I, Broschard TH, Boehni R, Moser R. Safety evaluation of calcium l-methylfolate. Toxicol Rep. 2019;6:1018. doi: 10.1016/j.toxrep.2019.09.012. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Zajecka JM, Fava M, Shelton RC, Barrentine LW, Young P, Papakostas GI. Long-term efficacy, safety, and tolerability of l-methylfolate calcium 15 mg as adjunctive therapy with selective serotonin reuptake inhibitors: A 12-month, open-label study following a placebo-controlled acute study. J Clin Psychiatry. 2016;77:654. doi: 10.4088/JCP.15m10181. - [DOI] [PubMed] [Google Scholar]
- 62.Fohr IP, Prinz-Langenohl R, Brönstrup A, Bohlmann AM, Nau H, Berthold HK. 5,10-methylenetetrahydrofolate reductase genotype determines the plasma homocysteine-lowering effect of supplementation with 5-methyltetrahydrofolate or folic acid in healthy young women. Am J Clin Nutr. 2002;2:275. doi: 10.1093/ajcn/75.2.275. et al. - [DOI] [PubMed] [Google Scholar]
- 63.Zhao R, Qiu A, Tsai E, Jansen M, Akabas MH, Goldman ID. The proton-coupled folate transporter: Impact on pemetrexed transport and on antifolates activities compared with the reduced folate carrier. Mol Pharmacol. 2008;74:854. doi: 10.1124/mol.108.045443. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Spiegelstein O, Eudy JD, Finnell RH. Identification of two putative novel folate receptor genes in humans and mouse. Gene. 2000;258:117. doi: 10.1016/s0378-1119(00)00418-2. - [DOI] [PubMed] [Google Scholar]
- 65.Manohar-Ratnam MDAS.. The folate receptor: What does it promise in tissue-targeted therapeutics? Cancer Metastasis Rev. 2007;26:141. doi: 10.1007/s10555-007-9048-0. - [DOI] [PubMed] [Google Scholar]
- 66.Shen F, Wu M, Ross JF, Miller D, Ratnam M. Folate receptor type γ is primarily a secretory protein due to lack of an efficient signal for glycosylphosphatidylinositol modification: Protein characterization and cell type specificity. Biochemistry. 1995;34:5660. doi: 10.1021/bi00016a042. - [DOI] [PubMed] [Google Scholar]
- 67.Zhao R, Goldman ID. The molecular identity and characterization of a proton-coupled folate transporter—pcft; biological ramifications and impact on the activity of pemetrexed. Cancer Metastasis Rev. 2007;26:129. doi: 10.1007/s10555-007-9047-1. - [DOI] [PubMed] [Google Scholar]
- 68.Zhao R, Diop-Bove N, Visentin M, Goldman ID. Mechanisms of membrane transport of folates into cells and across epithelia. Annu Rev Nutr. 2011;31:177. doi: 10.1146/annurev-nutr-072610-145133. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Whetstine JR, Flatley RM, Matherly LH. The human reduced folate carrier gene is ubiquitously and differentially expressed in normal human tissues: Identification of seven non-coding exons and characterization of a novel promoter. Biochem J. 2002;367:629. doi: 10.1042/BJ20020512. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Freisheim JH, Price EM, Ratnam M. Folate coenzyme and antifolate transport proteins in normal and neoplastic cells. Adv Enzyme Regul. 1989;29:13. doi: 10.1016/0065-2571(89)90091-5. - [DOI] [PubMed] [Google Scholar]
- 71.Matherly LH, Wilson MR, Hou Z. The major facilitative folate transporters solute carrier 19a1 and solute carrier 46a1: Biology and role in antifolate chemotherapy of cancer. Drug Metab Disposition. 2014;42:632. doi: 10.1124/dmd.113.055723. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Matherly LH, Hou Z, Deng Y. Human reduced folate carrier: Translation of basic biology to cancer etiology and therapy. Cancer Metastasis Rev. 2007;26:111. doi: 10.1007/s10555-007-9046-2. - [DOI] [PubMed] [Google Scholar]
- 73.Stanisławska-Sachadyn A, Mitchell LE, Woodside JV, Buckley PT, Kealey C, Young IS. The reduced folate carrier (slc19a1) c.80g>a polymorphism is associated with red cell folate concentrations among women. Ann Hum Genet. 2009;73:484. doi: 10.1111/j.1469-1809.2009.00529.x. et al. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Chango A, Emery-Fillon N, Courcyt GPd, Lambert D, Pfiste M, Rosenblatt DS. A polymorphism (80g->a) in the reduced folate carrier gene and its associations with folate status and homocysteinemia. Mol Genet Metab. 2000;70:310. doi: 10.1006/mgme.2000.3034. et al. - [DOI] [PubMed] [Google Scholar]
- 75.Rady PL, Szucs S, Matalon RK. Genetic polymorphism (g80a) of reduced folate carrier gene in ethnic populations. Mol Genet Metab. 2001;73:285. doi: 10.1006/mgme.2001.3197. - [DOI] [PubMed] [Google Scholar]
- 76.Thakur S, Rahat B, Hamid A, Najar RA, Kaur J. Identification of regulatory mechanisms of intestinal folate transport in condition of folate deficiency. J Nutr Biochem. 2015;26:1084. doi: 10.1016/j.jnutbio.2015.05.002. - [DOI] [PubMed] [Google Scholar]
- 77.Alam C, Hoque MT, Finnell RH, Goldman ID, Bendayan R. Regulation of reduced folate carrier (rfc) by vitamin d receptor at the blood-brain barrier. Mol Pharm. 2017;14:3848. doi: 10.1021/acs.molpharmaceut.7b00572. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Alam C, Aufreiter S, Georgiou CJ, Hoque MT, Finnell RH, O’Connor DL. Upregulation of reduced folate carrier by vitamin d enhances brain folate uptake in mice lacking folate receptor alpha. Proc Natl Acad Sci U S A. 2019;116:17531. doi: 10.1073/pnas.1907077116. et al. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Finkelstein JD. Pathways and regulation of homocysteine metabolism in mammals. Semin Thromb Hemost. 2000;26:219. doi: 10.1055/s-2000-8466. - [DOI] [PubMed] [Google Scholar]
- 80.Kriebitzsch C, Verlinden L, Eelen G, Schoor NMv, Swart K, Lips P. 1,25-dihydroxyvitamin d3 influences cellular homocysteine levels in murine preosteoblastic mc3t3-e1 cells by direct regulation of cystathionine β-synthase. J Bone Miner Res. 2011;26:2991. doi: 10.1002/jbmr.493. et al. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Bouillon R, Bischoff-Ferrari H, Willett W. Vitamin d and health: Perspectives from mice and man. J Bone Miner Res. 2008;23:974. doi: 10.1359/jbmr.080420. - [DOI] [PubMed] [Google Scholar]
- 82.Weinblatt ME, Coblyn JS, Fox DA, Fraser PA, Holdsworth DE, Glass DN. Efficacy of low-dose mrthotrexate in rheumatoid arthritis. N Eng J Med. 1985;312:818. doi: 10.1056/NEJM198503283121303. et al. - [DOI] [PubMed] [Google Scholar]
- 83.Turner AG, Hendry WF, Williams GB, Bloom HJG. The treatment of advanced bladder cancer with methotrexate. Br J Urol. 2010;49:673. doi: 10.1111/j.1464-410x.1977.tb04551.x. - [DOI] [PubMed] [Google Scholar]
- 84.Tazoe Y, Hayashi H, Tsuboi S, Shioura T, Matsuyama T, Yamada H. Reduced folate carrier 1 gene expression levels are correlated with methotrexate efficacy in japanese patients with rheumatoid arthritis. Drug Metab Pharmacokinet. 2015;30:227. doi: 10.1016/j.dmpk.2015.02.001. et al. - [DOI] [PubMed] [Google Scholar]
- 85.Laverdiere C, Chiasson S, Costea I, Moghrabi A, Krajinovic M. Polymorphism g80a in the reduced folate carrier gene and its relationship to methotrexate plasma levels and outcome of childhood acute lymphoblastic leukemia. Blood. 2002;100:3832. doi: 10.1182/blood.V100.10.3832. - [DOI] [PubMed] [Google Scholar]
- 86.Hideki H, Yui T, Seiji T, Masato H, Mariko M, Teruhisa A. A single nucleotide polymorphism of reduced folate carrier 1 predicts methotrexate efficacy in japanese patients with rheumatoid arthritis. Drug Metab Pharmacokinet. 2013;28:164. doi: 10.2133/dmpk.dmpk-12-nt-038. et al. - [DOI] [PubMed] [Google Scholar]
- 87.Ede AEv, Laan RFJM, Blom HJ, Boers GHJ, Haagsma CJ, Thomas CMG. Homocysteine and folate status in methotrexate-treated patients with rheumatoid arthritis. Rheumatology. 2002;6:658. doi: 10.1093/rheumatology/41.6.658. et al. - [DOI] [PubMed] [Google Scholar]
- 88.Slot O. Changes in plasma homocysteine in arthritis patients starting treatment with low-dose methotrexate subsequently supplemented with folic acid. Acta Rheumatol Scand. 2001;30:305. doi: 10.1080/030097401753180408. - [DOI] [PubMed] [Google Scholar]
- 89.Strober BE, Menon K. Folate supplementation during methotrexate therapy for patients with psoriasis. J Am Acad Dermatol. 2005;53:652. doi: 10.1016/j.jaad.2005.06.036. - [DOI] [PubMed] [Google Scholar]
- 90.Kampman MT. Folate status in women of childbearing age with epilepsy. Epilepsy Res. 2007;75:52. doi: 10.1016/j.eplepsyres.2007.04.003. - [DOI] [PubMed] [Google Scholar]
- 91.Belcastro V, Gaetano G, Italiano D, Oteri G, Caccamo D, Pisani LR. Antiepileptic drugs and mthfr polymorphisms influence hyper-homocysteinemia recurrence in epileptic patients. Epilepsia. 2007;48:1990. doi: 10.1111/j.1528-1167.2007.01164.x. et al. - [DOI] [PubMed] [Google Scholar]
- 92.Huemer M, Ausserer B, Graninger G, Hubmann M, Huemer C, Schlachter K. Hyperhomocysteinemia in children treated with antiepileptic drugs is normalized by folic acid supplementation. Epilepsia. 2005;46:1677. doi: 10.1111/j.1528-1167.2005.00264.x. et al. - [DOI] [PubMed] [Google Scholar]
- 93.Elliott JO, Jacobson MP, Haneef Z. Cardiovascular risk factors and homocysteine in epilepsy. Epilepsy Res. 2007;76:113. doi: 10.1016/j.eplepsyres.2007.07.005. - [DOI] [PubMed] [Google Scholar]
- 94.Chuang Y, Chuang H, Lin T, Chang C, Lu C, Chang W. Effects of long-term antiepileptic drug monotherapy on vascular risk factors and atherosclerosis. Epilepsia. 2012;53:120. doi: 10.1111/j.1528-1167.2011.03316.x. et al. - [DOI] [PubMed] [Google Scholar]
- 95.Hamed SA, Hamed EA, Hamdy R, Nabeshima T. Vascular risk factors and oxidative stress as independent predictors of asymptomatic atherosclerosis in adult patients with epilepsy. Epilepsy Res. 2007;74:18392. doi: 10.1016/j.eplepsyres.2007.03.010. [DOI] [PubMed] [Google Scholar]
- 96.Hamed SA, Nabeshima T. The high atherosclerotic risk among epileptics: The atheroprotective role of multivitamins. Jap J Pharmacol. 2005;98:340. doi: 10.1254/jphs.crj05003x. - [DOI] [PubMed] [Google Scholar]
- 97.Li S, Gao Y, Ma K, Li Y, liu C, Yan Y. Lipid-related protein NECTIN2 is an important marker in the progression of carotid atherosclerosis: An intersection of clinical and basic studies. J Transl Intern Med. 2021;9:294. doi: 10.2478/jtim-2021-0044. et al. - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Qian H, Zhang Y, Wu B, Wu S, You S, Zhang N. Structure and function of HECT E3 ubiquitin ligases and their role in oxidative stress. J Transl Intern Med. 2020;8:71. doi: 10.2478/jtim-2020-0012. et al . - [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Cleary P, Shorvon S, Tallis R. Late-onset seizures as a predictor of subsequent stroke. The Lancet. 2004;363:1184. doi: 10.1016/S0140-6736(04)15946-1. - [DOI] [PubMed] [Google Scholar]
- 100.Belcastroa V, Striano P. Antiepileptic drugs, hyperhomocysteinemia and b-vitamins supplementation in patients with epilepsy. Epilepsy Res. 2012;102:1. doi: 10.1016/j.eplepsyres.2012.07.003. - [DOI] [PubMed] [Google Scholar]