

Abstract
Sitotroga cerealella is one of the major pests of cereals in the field and storage conditions throughout the world. The main objective was to study the life tables of S. cerealella on wheat, maize and barley and its implications on percent parasitism of Trichogramma chilonis. S. cerealella is reared under lab conditions as its eggs are utilized for rearing T. chilonis. Fresh eggs of S. cerealella were collected and after hatching the neonate larvae of S. cerealella were transferred onto each host plant species for obtaining first (F1) generation (G). Seventy eggs were used for each host and each egg was used as a replicate. Daily observations were made for recording the life-table parameters of the S. cerealella. The data showed that the developmental time of S. cerealella eggs and pupae was maximum (5.68 and 7.75 days) when reared on wheat, while the maximum larval duration (19.77 days) of S. cerealella was recorded on barley. The maximum fecundity (290.30 ± 22.47 eggs/female) was recorded on maize, while minimum fecundity per female was recorded on barley (159.30 eggs/ female). The S. cerealella reared on maize had significantly higher values of finite rate of increase (λ), intrinsic rate of increase (r), and net reproductive rate (Ro) (0.14 ± 0.04 day− 1, 1.16 ± 0.05 day− 1, and 136.85 ± 20.25 eggs/ female) respectively. The mean generation time (T) (35.18 ± 0.61 days) was higher on wheat. Likewise, the gross reproductive rate (GRR) and the age-stage specific reproductive values (vxj) of newly oviposited eggs of S. cerealella were recorded higher (136.85 ± 20.25; 1.160 offspring) on maize. The data regarding the efficacy of T. chilonis for different parameters were recorded higher on maize i.e., percent parasitism (89.00 ± 2.30%), percent adult emergence (81.60 ± 1.20%), adult longevity (3.80 ± 0.10 days) and total adult longevity (9.90 ± 0.20 days) as compared to wheat and barley. Our findings revealed that S. cerealella can be best reared on maize under laboratory conditions as it prefers this host as compared to wheat and barley. Therefore, assigning the most susceptible and favorite host (maize) would help us to improve T. chilonis mass production under laboratory conditions.
Subject terms: Biological techniques, Environmental biotechnology, Plant biotechnology
Introduction
Cereal crops contain essential nutrients necessary for both human beings and other animals. These nutrients are needed and consumed by human beings regularly in their diet. The Food and Agriculture Organization estimated the total production of cereal crops as 2777 million tons during 2022, however the production of coarse grains used as a feed for animals, except wheat and rice reached 1330.02 million tons1. Global warming and other risks that have emerged in recent years show that cereal crop production and storage have critical roles in world food supply. Increasing demand for cereal crops can be met to some extent by implementing better storage conditions to reduce postharvest crop loss. Grain pests are serious threats to the stored grains and stored products in both developing and underdeveloped countries where a large portion of the stored grains and products are lost due to these storage pests2. The average grain losses due to the storage pests are about 12% of the total grain produced and in some cases these losses could approach 50%. An estimated 10% of stored grain losses globally are attributed to insect pest infestations3. This loss is caused by 70 different species of insects belonging to orders like Coleoptera and Lepidoptera. Among these insect pests, S. cerealella (Olivier) (Lepidoptera: Gelechiidae) is one of the most devastating primary lepidopteran pests of wheat, maize, sorghum, and rice under field and in storage conditions4.
The Angoumois grain moth, S. cerealella is one of the major pests of maize, wheat, rice, and sorghum under pre- and postharvest field and storage conditions throughout the world5. Damage is caused by the larvae boring into the seed and feeding on seed contents (embryo and endosperm). The silk made by the larvae on grains under high humidity imparts an unhealthy appearance, smell and intense growth of mold on grains6. Such damage causes substantial economic loss and reduces the viability of the seeds to germinate. Under favorable conditions, S. cerealella breeding may continue throughout the years in suitable stores7,8.
Although various control measures have been used against this destructive pest, using resistance varieties for suppression of S. cerealella infestation in stored products has been proposed in recent years9. Studying the life table parameters on resistant hosts is one of the most important tools in integrated pest management programs in stored products10. However, the proper use of resistant hosts in pest management techniques requires knowledge of the life table and biological parameters of pests11. Many scientists have used the life table theory for predicting the growth and size of populations12,13. Studying the age-stage, two-sex life table is a very comprehensive approach for measuring the growth, survival and reproductive potential of an insect population. This approach can faithfully represent the actual life history of an insect species since it easily depicts stage difference and incorporates both sexes. Life table studies are helpful in studying the various ecological aspects of interest in relation to insect pests and their associated natural enemies for the development of integrated control14,15. Traditional female-based age-specific life tables, on the other hand, neglect the male component of a population and stage structure in metamorphosing species and as a result, are fundamentally unable to accurately describe a population16,17. Additionally, the food quality has also been taken into consideration because it may influence the life history parameters of insects including survival rate, longevity and fecundity18,19.
In addition to being a serious pest of stored products, S. cerealella is also used as a host for mass rearing of the biological control agent, T. chilonis. In recent years, different cereal grains varieties have been used for large-scale production of S. cerealella as an alternative host for rearing of parasitoids and predators20. S. cerealella was originally used as a factitious host for the mass rearing of Trichogramma by Flander in 1929. Mclaren and Rye, (1983) devised an efficient method for mass rearing of Trichogramma wasps on S. cerealella eggs. The ability to rear T. chilonis and other biological control agents on factitious hosts like S. cerealella has greatly improved21,22. Effectiveness of bio-agents on host eggs is significantly influenced by the quality of the cereals e.g., T. chilonis performance was highly influenced by Corcyra cephalonica eggs when reared on different cereals. More adults of T. chilonis emerged on C. cephalonica eggs reared on maize compared to wheat and barley23. Although some biological parameters (development and fecundity) of S. cerealella on various stored products have been documented by entomologists24,25, a complete study of all the lifetables is not available. Therefore, the present research was designed to study the age and stage specific lifetables of S. cerealella and bio-efficacy of the T. chilonis against S. cerealella reared on different hosts under laboratory conditions.
Results
The results showed that the food source affects the population parameters of S. cerealella (Table 1). The eggs developmental time (days) of S. cerealella was significantly lower when reared on maize and barely (5.45 and 5.43 days, F = 278.57, DFs = 2, 207), respectively as compared to the developmental time of S. cerealella eggs reared on wheat (5.68 days) (P = 0.0167). The total larval duration (days) of S. cerealella when reared on maize was recorded significantly lower (P = 0.0079, 17.97 days) as compared to S. cerealella reared on barley (19.77 days), while non-significant difference was observed when reared on maize and wheat (P = 0.4826, (F = 228.02, DFs = 2, 207). The data regarding the pupal duration (days) of S. cerealella show that significantly lower pupal duration of S. cerealella was recorded when reared on maize (6.57 ± 0.12 days) (P = 0.00, (F = 1142.59, DFs = 2, 195) as compared to wheat and barley, while non- significant difference was recorded in the pupal duration of S. cerealella reared on wheat (7.75 ± 0.14 days) and barley (7.42 ± 0.11 days) (P = 0.076). In the same way, significantly lower pre-adult survival rate of S. cerealella was recorded when reared on maize (30.69 ± 0.36 days) (P = 0.001, F = 0.002, DFs = 2, 207), while the greatest pre-adult survival rate of S. cerealella was recorded on barley (33.28 ± 0.33 days), followed by wheat (32.87 ± 0.39 days). The total developmental time of S. cerealella adults was significantly extended on maize (11.22 ± 0.30 days, F = 64.09, DFs = 2, 207) and barley (10.27 ± 0.32 days) as compared to wheat (10.90 ± 0.40 days) (P = 0.016). The total pre oviposition period (TPOP) of S. cerealella reared individually on barley and wheat was not significantly different (P = 0.5615). A significantly higher number of oviposition days of S. cerealella were observed when reared on maize (6.96 ± 0.360 days) (P = 0.00, F = 431.06, DFs = 2, 96), while no significant difference was observed in the oviposition days of S. cerealella reared on wheat and barley (P = 0.300). In the same way, significantly maximum number of eggs (fecundity/ female) of S. cerealella was recorded when reared on maize (290.30 ± 22.47 eggs) (P = 0.000, (F = 435.13, DFs = 2, 96), while no significance difference in the mean fecundity of S. cerealella was recorded when reared on wheat and barley (P = 0.082). Similarly, a significantly lower doubling time (days) of S. cerealella was recorded when reared on maize (6.96 ± 0.36 days, (F = 452.91, DFs = 2, 207) as compared to wheat and barley (5.37 ± 0.200 and 5.611 ± 0.227 days) respectively. The total female longevity (days) of S. cerealella was recorded lower on maize (42.15 ± 0.750 days, (F = 64.55, DFs = 2, 96), followed by wheat and barley with (42.84 ± 0.88, 44.72 ± 0.92 days) respectively. A non-significant difference was recorded in the female longevity of S. cerealella when reared on wheat and maize as compared to barley (P = 0.5434). Similarly, the total male longevity (days) of S. cerealella was recorded significantly lower on maize as compared to wheat (P = 0.006; F = 142.50, DFs = 2, 96) (41.69 ± 0.635 days), while no significant difference was observed in the mean total male longevity of S. cerealella reared on maize and barely (P = 0.163).
Table 1.
Stage durations, oviposition days and fecundity (Mean ± standard error) of Sitotroga cerealella reared individually on wheat, barley, and maize under laboratory conditions.
| Parameter | n* | Wheat | n | Barely | N | Maize |
|---|---|---|---|---|---|---|
| x̄ ± S. E | x̄ ± S. E | x̄ ± S. E | ||||
| Egg (days) | 70 | 5.68 ± 7.423a | 70 | 5.42 ± 0.076b | 70 | 5.45 ± 0.059b |
| Larva (days) | 70 | 18.51 ± 0.571ab | 70 | 19.77 ± 0.45a | 70 | 17.97 ± 0.50b |
| Pupa (days) | 66 | 7.757 ± 0.148a | 66 | 7.424 ± 0.113a | 66 | 6.575 ± 0.125b |
| Preadult/immature duration (days) | 66 | 32.88 ± 0.398a | 66 | 33.26 ± 0.343a | 66 | 30.70 ± 0.368b |
| Adult (days) | 66 | 10.90 ± 0.409ab | 66 | 10.27 ± 0.323b | 66 | 11.22 ± 0.304a |
| TPOP (days) | 33 | 32.87 ± 0.560a | 33 | 33.30 ± 0.484a | 33 | 30.69 ± 0.521b |
| Oviposition days | 33 | 5.30 ± 0.303b | 33 | 4.90 ± 0.22bc | 33 | 6.96 ± 0.360a |
| Fecundity (eggs per female) | 33 | 198.87 ± 18.89b | 33 | 159.30 ± 12.77bc | 33 | 290.30 ± 22.47a |
| Doubling time (days) | 70 | 5.37 ± 0.200a | 70 | 5.611 ± 0.227a | 70 | 4.65 ± 0.154b |
| Female total longevity (days) | 33 | 42.84 ± 0.88ab | 33 | 44.21 ± 0.591a | 33 | 42.15 ± 0.750b |
| Male total longevity (days) | 33 | 44.72 ± 0.923a | 33 | 42.90 ± 0.602ab | 33 | 41.69 ± 0.635bc |
Where x̄ = Mean and S. E = standard error, were estimated by using the bootstrap (100000) resampling method. Figures with same letters in a row are not statistically different from each other using the bootstrap (100000) resampling technique.
*n indicates several replicates.
Population parameters of Sitotroga cerealella reared on different hosts
The population parameters viz intrinsic rate of increase (r), finite rate of increase (λ), reproductive rate (Ro), generation time (T) and gross reproductive rate (GRR) of S. cerealella reared individually on wheat, barley and maize are presented in Table 2. The means and standard error were estimated by using the bootstrap method. S. cerealella reared on maize had high values of the intrinsic rate of increase (r) (F = 531.54, DFs = 2, 207), finite rate of increase (λ) (F = 535.43, DFs = 2, 207) and net reproductive rate (Ro) (F = 277.57, DFs = 2, 207), (0.148 ± 0.004 per day, 1.160 ± 0.005 per day and 136.85 ± 20.25 offspring) respectively as compared to S. cerealella reared on wheat and barley. A significant difference was recorded in the gross reproductive rate (GRR) of S. cerealella reared on maize and barley (P = 0.000, F = 306.09, DFs = 2, 207), while no statistical variation was recorded in the gross reproductive rate of S. cerealella reared on maize and wheat (P = 0.0865). As presented in Table 2, different hosts had a significant effect on the mean generation time (T) of S. cerealella (F = 253.29, DFs = 2, 207). The mean generation-time (T) was found to be lowest on maize (33.056 ± 0.642) (P = 0.0167) as compared to wheat (35.182 ± 0.61 days).
Table 2.
Life table parameters (Mean ± Standard error) of Sitotroga cerealella reared individually on wheat, barley and maize under laboratory conditions.
| Parameters | n* | Wheat | N | Barley | n | Maize |
|---|---|---|---|---|---|---|
| x̄ ± S. E | x̄ ± S. E | x̄ ± S. E | ||||
| The intrinsic rate of increase (r) per day | 70 | 0.129 ± 0.004b | 70 | 0.123 ± 0.004b | 70 | 0.148 ± 0.004a |
| The finite rate of increase (λ) per day | 70 | 1.137 ± 0.005b | 70 | 1.131 ± 0.005b | 70 | 1.160 ± 0.005a |
| The net reproductive rate (Ro) offsprings | 70 | 93.757 ± 14.79ab | 70 | 75.122 ± 11.25b | 70 | 136.857 ± 20.25a |
| The mean generation time (T) in days | 70 | 35.182 ± 0.610a | 70 | 34.963 ± 0.595a | 70 | 33.056 ± 0.642b |
| Gross reproductive rate (GRR) offsprings | 70 | 93.75 ± 14.79b | 70 | 75.1 ± 11.25c | 70 | 136.85 ± 20.25a |
Where x̄ = Mean and S. E = standard error, were estimated by using the bootstrap (100000) resampling method. Figures with same letters in a row are not statistically different from each other using the bootstrap (100000) resampling technique.
*n indicates number of replicates.
Survival rate of Sitotroga cerealella reared on different hosts
The age stage specific survival-rate (sxj) is the probability that an individual can survive to age x and stage j. The different curves in Fig. 1 showed that there was significant overlapping between the curves because of different times of development that happened among S. cerealella individuals. The mean (total) developmental time of S. cerealella eggs (Fig. 1) was significantly lower on maize and wheat (5 days) as compared when reared on barley (7 days) (P = 0.0167, (F = 64.09, DFs = 2, 207). Similarly, the total larval developmental duration (days) of S. cerealella (29 days) was noted significantly lower when reared on maize as compared to wheat and barley (P = 0.007, (F = 228.02, DFs = 2, 207). In the same way, less pupal developmental time of S. cerealella was recorded when reared on maize (37 days) as compared to barley and wheat (39 days). Similarly, the total developmental time of adult S. cerealella males was (48, 53 and 49 days) and for females it was (51, 54 and 51 days) when reared on maize, wheat, and barley respectively.
Figure 1.
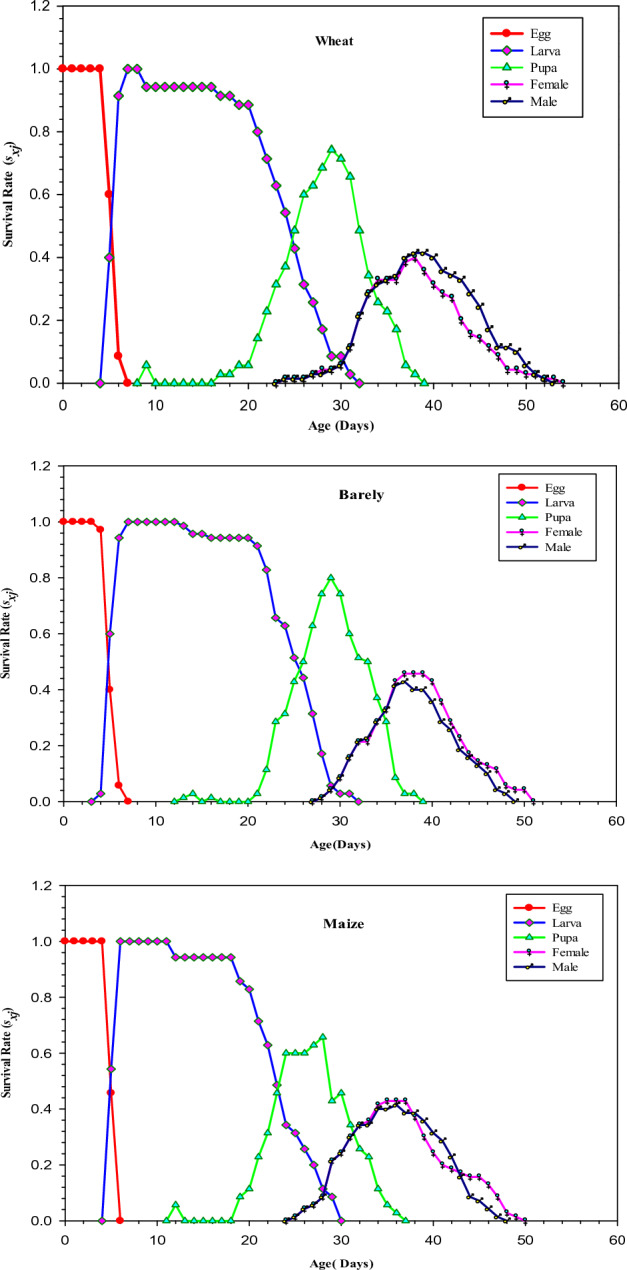
Survival rate (sxj) of Sitotroga cerealella reared individually on wheat, barley, and maize under laboratory conditions.
Age-specific survival rate of S. cerealella reared on different hosts
The age specific survival-rate (lx) of S. cerealella dropped slowly on maize and barley as compared to wheat (Fig. 2). The value for the age-specific fecundity (fx) (114.00 offspring per day) was recorded on day 25 in maize which was greater than the peak value of S. cerealella reared on barley (110.0 per day) on day 28 and wheat (4.0 per day) on the 24th day. The curve of the age specific fecundity (mx) exhibit that egg-laying of S. cerealella started on day 25 and maximum egg-laying (14.38 eggs per day) of S. cerealella was recorded on day 25 when reared on maize, while the egg-laying of S. cerealella started on day 24 and maximum egg-laying (12.71 eggs per day) was recorded on day 34 when wheat was used as food source. Similarly, egg-laying of S. cerealella started on day 28 on barley and the maximum egg-laying (10.52 eggs per day) of S. cerealella was recorded on day 36. In the same way the greatest age-specific maternity (lxmx) of S. cerealella (12.88 offspring) was observed on maize on day 31 followed by 11.75 offspring and 9.77 offspring in wheat and barley on days 33 and 36 respectively as shown in Fig. 2.
Figure 2.
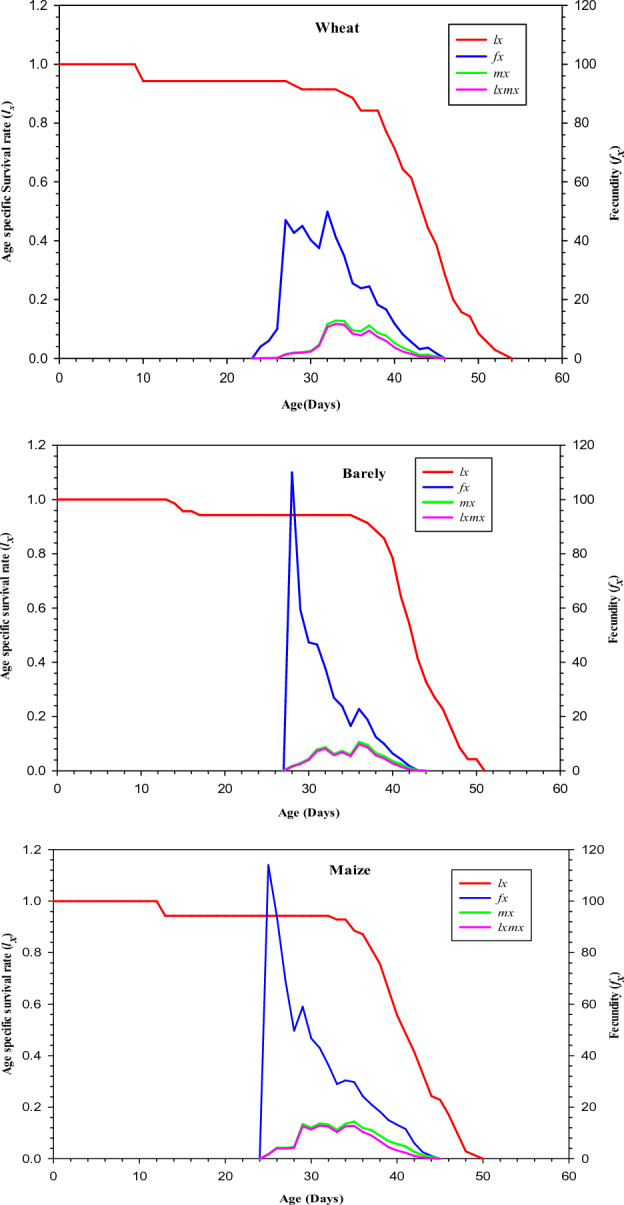
The Age-specific survival rate (lx), age-stage fecundity of the female stage (fx) age-specific fecundity (mx), and age-specific maternity (lxmx) of Sitotroga cerealella reared on wheat, barley and maize under laboratory conditions.
Age stage-specific life expectancy
The age specific life-expectancy (exj) of a newborn egg of S. cerealella was 41.851 and 41.942 days on maize and barley respectively which were longer than the life expectancy (exj) of a newly oviposited egg of S. cerealella reared on wheat (40.271 days). As seen in Fig. 3 the life expectancy (exj) of male and female S. cerealella adults was significantly high at the time of emergence (16.512 and 17.101) days respectively when reared on maize (P = 0.001). Similarly, the life expectancy (exj) of male and female adults of S. cerealella at the time of emergence was 14.77 and 16.207 days respectively on barley while in wheat, the male and female life expectancy at the time of emergence was 12.459 and 14.318 days respectively which was lower as compared to S. cerealella reared on barley and maize (Fig. 3).
Figure 3.
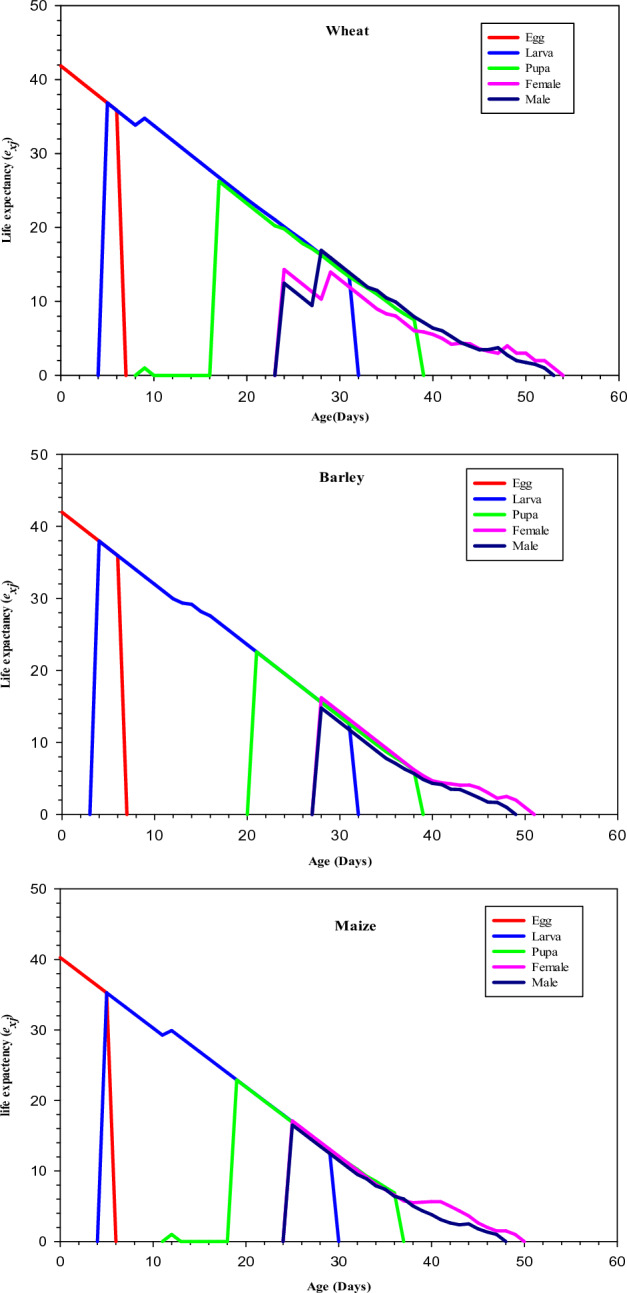
Age-stage specific life expectancy (exj) of Sitotroga cerealella reared on wheat, barley and maize under laboratory conditions.
Age stage-specific reproductive rate
The reproductive value (vxj) of individuals of age x and Stage j is shown in Fig. 4. The age stage specific reproductive values (vxj) of a newly oviposited egg of S. cerealella were recorded highest (1.160 offspring per day) when reared on maize followed by the reproductive value (vxj) of S. cerealella reared on wheat (1.137 offspring per day) and barley (1.131 offspring per day) respectively. The (vxj) curve for the female reproductive value was significantly increased at the start of reproduction (Fig. 4). A newly emerged female showed the highest reproductive value (vxj) of (398.74 offspring) on day 26 when reared on maize as compared to the reproductive value (vxj) when reared on barley and wheat (310.39 and 153.29 offspring per day) respectively.
Figure 4.
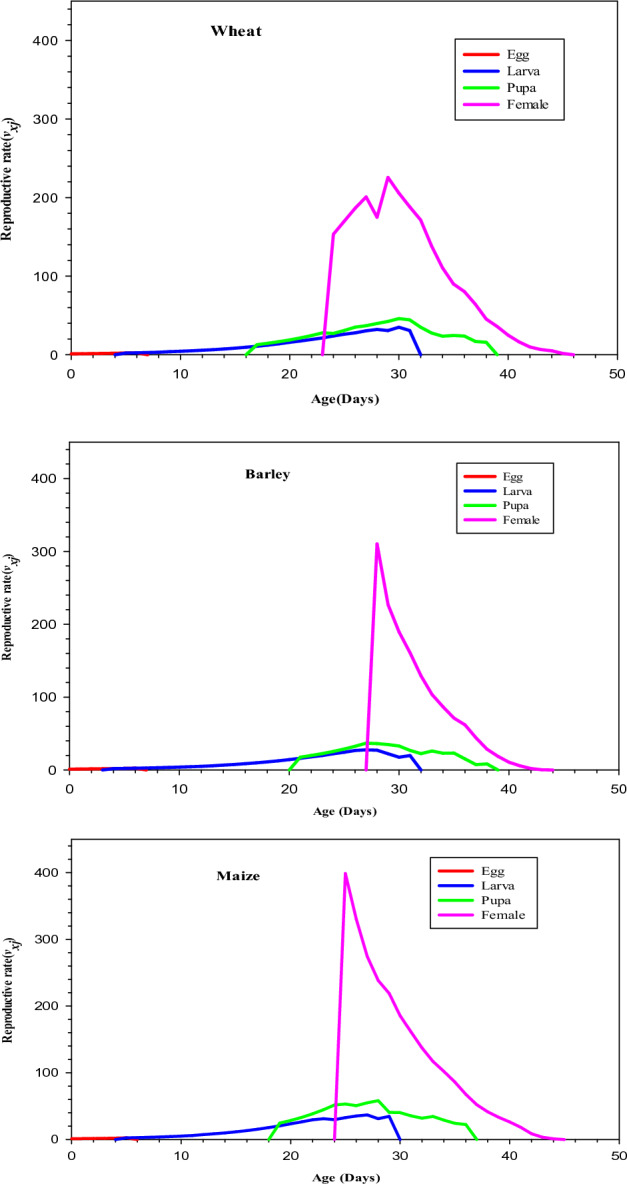
Age-stage specific reproductive rate (vxj) of Sitotroga cerealella reared on wheat, barley, and maize under laboratory conditions.
Population projection
The population size of the different stages of S. cerealella reared on maize wheat and barley simulated from an initial population of 10 eggs in each host were used to project the population growth of S. cerealella for 60 days duration using the TIMING-MSChart program is shown in Fig. 5. The different curves in Fig. 5 shows the trend and emergence time of different stages of S. cerealella reared on different hosts. According to the simulation results, the population growth of S. cerealella was the quickest in maize followed by wheat and slowest in barley. The S. cerealella population on maize reached 15,679.72 adults, on wheat reached 1,577.56 adults, and on barley reached 1,004.87 adults after 60 days (Fig. 5).
Figure 5.

Population projection of S. cerealella reared on wheat, barley and maize under laboratory conditions.
Efficacy of Trichogramma chilonis against Sitotroga cerealella eggs under laboratory conditions
The mean percent parasitism of T. chilonis was significantly higher on eggs of S. cerealella reared on maize (89.00 ± 2.31%) compared with other cereals, this was followed by values for barley (77.67 ± 2.40%) and wheat (71.33 ± 1.45%) (Table 3). The maximum mean percent adults of T. chilonis (81.66 ± 1.20%) emerged from eggs of S. cerealella reared on maize followed by barley (72.00 ± 1.53%) while the minimum mean percent adult emergence of T. chilonis was recorded (72.00 ± 1.53%) on eggs reared on wheat. Maximum adult longevity (3.80 ± 0.12 days) and total longevity (9.96 ± 0.20 days) of T. chilonis were recorded on S. cerealella eggs reared on maize while significantly lower adult longevity (3.066 ± 0.12 days) and total longevity (7.90 ± 0.32 days) of T. chilonis were recorded on S. cerealella eggs reared on wheat.
Table 3.
Percent parasitism, percent adult emergence, adult longevity and total adult longevity of Trichogramma chilonis on S. cerealella eggs developed on wheat, barley and maize under laboratory conditions.
| Hosts | Percent parasitism (%) | Percent adult emergence (%) | Longevity (days) | Total longevity (days) |
|---|---|---|---|---|
| Maize | 89.00 ± 2.31a | 81.66 ± 1.20a | 3.80 ± 0.12a | 9.96 ± 0.20a |
| Barley | 77.66 ± 2.40b | 72.00 ± 1.53b | 3.43 ± 0.23ab | 8.50 ± 0.29b |
| Wheat | 71.33 ± 1.45b | 72.00 ± 1.53b | 3.066 ± 0.12b | 7.90 ± 0.32b |
| LSD Values | 7.2648 | 8.1835 | 0.5729 | 0.9535 |
Figures with same letters in a column are not statistically different from each other using the least Significance Difference test at 5% significance level.
Discussion
Life table studies of insects reared on different hosts can reflect the suitability of the different hosts for that specific pest species26. Several researchers have reported that the life table parameters of the stored grain pests could be affected by the physiochemical properties (morphology and hardness of the grain, nutritional value and availability of the food) of the stored products27. In the current study, the lifetable and population parameters of Angoumois grain moth, S. cerealella were studied individually on wheat, barley and maize. The greatest fecundity and oviposition period was recorded when S. cerealella was reared on maize as compared to barley and wheat. The eggs and larval developmental time (days) of S. cerealella was recorded lower on maize as compared to wheat and barley. Similarly, the developmental time of pupae (days) was observed lower on maize as compared to wheat and barley. Also, minimum male and female total longevity (days) was recorded when S. cerealella was reared on maize as compared to wheat and barley respectively. In the same way the total Pre-Ovi-Position period (TPOP), doubling time and pre adult survival rate (%) was recorded lower on maize and wheat as compared to barley. In previous studies, the population parameters of S. cerealella reared on the different hosts were reported by other researchers. A study conducted by Chi and Su reported that rapid growth of insects on a given diet suggests high host suitability28. Further the loss of weight, emergence of adult and gain in adult weight was observed by stored insect pests on susceptible varieties as compared to resistant varieties24.
The highest intrinsic rate of increase (r) per day for S. cerealella was recorded on maize and the mean generation time (T) of S. cerealella was recorded on wheat. Similarly, the highest finite-rate of increase (λ) per day, net reproductive-rate (R0), age specific fecundity (fx), age specific maternity (lxmx), life expectancy of both adults, age stage specific reproductive values (vxj) and Gross reproductive rate (GRR) of S. cerealella were recorded on maize. The greatest age-stage specific life expectancy (exj) of newly oviposited eggs of S. cerealella was recorded on barley. Also, the net reproductive rate (R0) and gross reproductive rate (GRR) metrics only measure the reproductive potential of diets, not their total fitness. Therefore, care should be taken when using these two metrics (R0 and GRR) to assess if a diet is suitable for S. cerealella. Similar results were reported for S. cerealella when it was reared on maize29, wheat20 and rice30 and these studies show that the life table parameters of S. cerealella were greatly influenced by the host phenology and nutritional qualities of food sources. Another study conducted by Demissie et al., also documented that the developmental time of S. cerealella eggs, larvae and pupae on rice grain was 5.5 days, 25.2 days and 5.0 days respectively31.
Our findings on total developmental time of immature stages (33.26 days) are in complete conformity with the study of Murad and Batool, who recorded that the total developmental time of immature stages of S. cerealella on barley was 32.1 days32. Our results agree with the work of Akter et al., who reported the lifetable parameters of S. cerealella on different barley varieties and reported the incubation period (7.00 ± 0.00), larval period (36.80 ± 0.38), pupal period (9.48 ± 0.17) and adult longevity of both male and female (6.32 ± 0.13 and 6.74 ± 0.26) days respectively33.
Our results further showed that the oviposition days of S. cerealella on barley was (4.909 ± 0.22 eggs), with age-stage specific life expectancy (exj) (41.942 days) and mean number of eggs female− 1 (159.30 ± 12.77 eggs) were recorded. These findings confirm the findings of Akter et al., who observed almost similar oviposition days (4.80 ± 0.25), age-stage specific life expectancy (exj) ranges from 36.6–48.25 days and 161.37 ± 15.11 eggs female− 1 of S. cerealella on different cultivars of barley33.
The age stage specific survival rate (sxj) of S. cerealella was recorded higher on wheat as compared to maize and barely. While the survival rate of S. cerealella eggs, larvae and pupa reared on barley were 7, 32 and 39 days respectively. Also, the life expectancy (exj) of S. cerealella and the age stage specific reproductive values (vxj) of newly oviposited egg of S. cerealella was high on barley as compared to wheat and maize. Similar results were also published by Rizwan et al., who studied the survival rate and other developmental parameters of S. cerealella on barely and recorded maximum total life expectancy of S. cerealella (58.459 days) on different varieties of barley30. Similarly, the age specific life expectancy of a newly oviposited egg of S. cerealella was also recorded higher on barley as compared to wheat and maize. These three hosts were further assessed for their proximate compositions (Supplementary file “Table S1”). The proximate results showed higher moisture (12.5%), crude protein (13.00%), ash content (2.20%) and crude fat (3.60%) in maize as compared to wheat and barley, while no significant different were recorded in % carbohydrate content in all the three types of grains. These results further confirm that these grain’s constituents have a direct effect on the biology of the S. cerealella31. Grains with high moisture, crude protein and ash contents are positively correlated with the progeny emerged and susceptibility index31,32. Similarly, grains with high crude fats have positive impact on the mean fecundity data of S. cerealella10. These findings support the current research finding where maximum mean number eggs laid of S. cerealella was recorded when reared on maize grains.
The projection of S. cerealella population reared on wheat, barley and maize reflects the usefulness of age-stage two-sex life tables for forecasting the population under favorable environmental conditions. According to the simulation results the population growth was the quickest on maize and it was followed by wheat and slowest on barley. The S. cerealella population on maize reached 15,679.72 adults, while on wheat it reached 1,577.56 adults and on barley reached 1,004.87 adults after 60 days. Predicting and anticipating the population growth of insect pests are critical in formulating the correct timing schedule for a pest management programme34. A computer-based program using life table data is therefore a very important tool in pest management and decision making35.
The mean parasitism, mean adult emergence, longevity of adult and total adult longevity of T. chilonis were recorded highest on S. cerealella eggs reared on maize. A research study conducted by Akter et al., on the lifetables of T. chilonis feeding on S. cerealella reported that T. chilonis parasitism ranged from 61 to 95%, with adult emergence 51 to 96.30%. They further reported adult longevity ranged from 4.2 to 7.4 days33. Current findings are further on par with the work of Nadeem et al., who recorded the greatest adult’s emergence (98%) of T. chilonis from host eggs36. According to evolutionary models, more females will oviposit in large hosts than in small hosts37,38. A similar study conducted by Khan et al., also reported the same parameters of T. chilonis reared on S. cerealella egg under free choice method and observed percent parasitism, percent adult emergence, adult longevity and total longevity of adults as 94.0 ± 0.33, 94.0 ± 0.49, 9.6 ± 0.16 and 3.6 ± 0.27) days respectively27. Similarly, Charnov et al. also stated that sex-ratio manipulation may be an adaptation in response to the differing impact of host size on the fitness of female versus male wasps39. Our findings are further in line with these studies in that large size eggs from maize produced proportionately more female wasps than smaller size eggs from wheat and barley, which produced the lowest proportion of females but still produced more females than males.
Based on present study, S. cerealella completed their life cycle on all tested grains but maize was the best food source among the tested grains. The simulation data results showed that the growth of the S. cerealella population was faster on maize than it was on wheat and barley. Also, the results show that different food sources used for rearing of host insect significantly affected the percent parasitism, adult emergence, and adult longevity of the T. chilonis. The current study provides valuable results since understanding the life-tables study of S. cerealella on various hosts will be useful in mass production of biocontrol agents. Also, utilizing a susceptible host (maize) could help researchers and farmers in improving mass production of biocontrol agents. Further study on the demographic parameters of S. cerealella can help to understand the population dynamics and to develop more effective management programs for this pest.
Conclusions
The study results showed that S. cerealella completed its life cycle on all the grains however maize seems to be a better food source for mass production of this pest compared to wheat and barley. The simulation data results showed that the growth of the S. cerealella was faster on maize as compared to wheat and barley. Understanding the life table studies of S. cerealella on different hosts is useful in designing a comprehensive scheme for an IPM program of S. cerealella.
Additionally, T. chilonis parasitism rate, adult emergence, and adult longevity were affected by the food sources of host insects. Therefore, assigning the most susceptible and favorite host (maize) would help us to improve T. chilonis mass production. These findings will provide a concrete basis for further studies in the real environment for both S. cerealella and T. chilonis.
Materials and methods
The study was conducted at the Department of Plant Protection, The University of Agriculture Peshawar during 2020. The insecticide-free seeds of wheat (variety Saleem 2000), barley (variety AAJ) and maize (cultivar Azam) were purchased from the local market in Peshawar. The seeds were disinfected by sterilization at 121 °C for 30 min in an autoclave. After sterilization, the seeds were air dried and then were used for the experiments.
Rearing of the Sitotroga cerealella
For the rearing of Sitotroga cerealella, fresh eggs of S. cerealella were obtained from (SCRI,) Mardan during the first week of April 2020 and were maintained under laboratory conditions at 27 ± 2 °C and 65 ± 5% R.H. with 08:16 h (L: D) cycle. After hatching the neonate larvae of S. cerealella were transferred onto each host under laboratory conditions. These S. cerealella were reared on different hosts for one generation. When enough eggs (~ 1500) were produced, the life-table studies of S. cerealella were initiated on sterilized barley, maize and wheat.
Life table parameters
The life table parameters of Angoumois grain moth were studied on wheat, barley, and maize under laboratory conditions at 27 ± 2 °C and 65 ± 5% R.H. with 08:16 h (L: D) cycle. After rearing for one generation, S. cerealella eggs were collected using a soft hand brush in Petri dishes. These eggs were then transferred into Petri dishes containing different grain hosts. A replicate consisted of a single egg and a total of 70 eggs (replicates) were used for each host as mentioned above. The cultures were maintained under the laboratory conditions at 27 ± 2 °C and 65 ± 5% R.H. with 08:16 h (L: D) cycle. A total of 70 petri dishes(replicates) were used for each diet. Daily observations were made for eggs hatching and larval duration. The pupal duration of S. cerealella in different hosts was confirmed by the presence of a tiny round semitransparent hole in the grains until adult emergence. Upon emergence, adults were paired and then each pair (one male and one female) was transferred into a plastic funnel (approximately 12–13 cm diameter), then covered with a lid of 75 mesh size sieve at the top face. The plastic funnel was then placed upside down over a dish containing starch for eggs data collection. The number of eggs were sieved daily through the 85-mesh size sieve and counted under a binocular magnifying instrument. The data on number of eggs were continued until the death of all individuals in the experiments.
Population projection
The population potential and growth of S. cerealella reared on wheat, maize, and barley were utilized for population and growth projection of S. cerealella by the Chart i.e., TIMING-MS program40,41. For the initial population of S. cerealella on maize, wheat, and barley, 10 eggs in each host were used to project the population growth of S. cerealella for 60 days duration.
Rearing of Trichogramma chilonis
When enough host eggs were available, rearing of T. chilonis was initiated. Sticky gum was spread uniformly on a piece of hard paper (15 × 10 cm). Then fresh eggs (300) of the S. cerealella (F1 generation) reared on each host were sprinkled on each sticky card pieces. These egg cards were then exposed to UV radiation for one hour to kill the larvae present inside eggs. After the radiation treatment, these eggs cards were transferred to a glass jar containing adults of T. chilonis in the ratio of 6:1 (female: male) for 3 h. After 3 h, the exposed egg cards were kept in separate glass jars under laboratory conditions at 25 ± 2 °C, 65 ± 5% R.H. with 08:16 h (L: D) cycle. T. chilonis adults upon emergence were fed on 10% honey drops solution. The emergence of T. chilonis adults was programmed i.e., emergence of adults from parasitized eggs which have been parasitized by keeping them at low temperatures (5 °C) during the pupal stage.
Efficacy of Trichogramma chilonis against Sitotroga cerealella eggs under laboratory conditions
To study the efficacy of T. chilonis against S. cerealella reared on wheat, barley and maize, fresh eggs of S. cerealella (100) of F2 generation from each host were collected separately from already established cultures. Those eggs were then scattered uniformly on sticky cards (3 × 2 cm). A total of five cards each having 20 eggs/card were used for each treatment. These egg cards were then placed separately in the glass jar containing freshly mated female adult of T. chilonis for parasitization for 24 h. The exposed eggs were then kept in separate glass jar under laboratory conditions at 25 ± 2 °C, 65 ± 5% R.H. with 08:16 h (L: D) cycle until they turned black indicating development of the parasitoid inside. Upon turning black, the cards containing eggs were collected from the jars and were examined for percent parasitism. Percent Parasitism was calculated using formula42.
Proximate composition
Chemical analyses of the tested grains of wheat, maize and barely grains were performed using the standard official methods of analysis of Association of Official Analytical Chemists (AOAC), (2016).
Statistical analysis
Data on eggs hatching, larval and pupal durations, adult emergence, adult longevity, preoviposition period and oviposition period were analyzed utilizing age stage, two sex life-table theory26,28,40,43,44. The Intrinsic rate of increase (r), the finite rate of increase (λ) and net reproductive rate (R0) were calculated with the help of a TWOSEX-MS Chart program. Bootstraps methods (10000 resampling) were used for the estimation of standard errors and comparison of population parameters of S. cerealella reared on wheat, barley and maize hosts. The data regarding the efficacy studies of T. chilonis were subjected to one way analysis of variances. The data on mean parasitism, mean adult emergence, adult longevity and mean (total) adult longevity were analyzed using Statistical package (Statistix 8.1) means were compared using LSD test at 5% level of probability.
Ethical declaration
All the experiments were reviewed and approved by the Department of Plant Protection, The University of Agriculture, Peshawar ethical committee as per university set guidelines and were in accordance with the Pakistan Agricultural Pesticide Act. 1997.
Supplementary Information
Acknowledgements
This project was supported by the Researchers Supporting Project number (RSP2023R191), King Saud University, Riyadh, Saudi Arabia. We would also like to thank the Department of Plant Protection for providing the necessary equipment. The authors would also like to thank Michael D. Bryan (Plant Industry Program Specialist Pesticide and Plant Pest Management Division) State of Michigan, Department of Agriculture and Rural Development for his diligent proofreading of this manuscript.
Author contributions
M.S., A.U.R.S., A.G. and S.A. conceived and designed the study. I.U. conducted data gathering. M. S. wrote the article. M.H.A, A.A.S, L.A. M.M.A.D and M.S helped in data analysis and revision of the manuscript.
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-023-37852-0.
References
- 1.FAOSTAT. URL: http://www.fao.org/faostat/en/-data/QC. Food and agriculture organization of the United Nations (FAO) (2022).
- 2.Tripathi, A. K. Pests of stored grains. In: Pests and Their Management (pp. 311–359). Springer, Singapore. (2018).
- 3.Gabriel, A. H. and Hundie, B. Farmers' post-harvest grain management choices under liquidity constraints and impending risks: Implications for achieving food security objectives in Ethiopia (No. 1004-2016-78504). (2006).
- 4.Hodges, R. J. et al. Postharvest losses and waste in developed and less developed countries: Opportunities to improve resource. (2011).
- 5.Rajendran, S. Postharvest pest losses in Pimentel D, editor. Encyclopedia of pest management. New York (NY): Marcel Dekker; pp. 654–656. (2002).
- 6.Borzoui E, et al. Effects of food quality on biology and physiological traits of Sitotroga cerealella (Lepidoptera: Gelechiidae) J. Econ. Entomol. 2016;110(1):266–273. doi: 10.1093/jee/tow284. [DOI] [PubMed] [Google Scholar]
- 7.Hayma, J. AD31E The storage of tropical agricultural products (No. 31). Agromisa Foundation. (2003).
- 8.Weston PA, Rattlingourd PL. Progeny production by Tribolium castaneum (Coleoptera: Tenebrionidae) and Oryzaephilus surinamensis (Coleoptera: Silvanidae) in maize previously infested by Sitotroga cerealella (Lepidoptera: Gelichiidae) J. Econ. Entomol. 2000;93(2):533–536. doi: 10.1603/0022-0493-93.2.533. [DOI] [PubMed] [Google Scholar]
- 9.Hansen LS, Skovgard H, Hell K. Life table study of Sitotroga cerealella (Lepidoptera: Gelechiidae), a strain from West Africa. J. Econ. Entomol. 2004;97(4):1484–1490. doi: 10.1093/jee/97.4.1484. [DOI] [PubMed] [Google Scholar]
- 10.Rizwana S, et al. Biological parameters of Trichogramma chilonis Ishii (Trichogrammatidae: Hymenoptera) feeding on Sitotroga cerealella eggs at three constant temperatures. Pak. J. Agric. Res. 2013;26(1):65–70. [Google Scholar]
- 11.Hamed M, Nadeem S. Effect of cereals on the development of Sitotroga cerealella (Olivier) (Lepidoptera: Gelechiidae) and subsequent quality of the egg parasitoid, Trichogramma chilonis (Ishii) (Hymenoptera: Trichogrammatidae) Pak. J. Zool. 2012;4(4):923–929. [Google Scholar]
- 12.Razmjou J, Tavakkoli H, Fallahi A. Effect of soybean cultivar on life history parameters of Tetranychus urticae Koch (Acari: Tetranychidae) J. Pest Sci. 2009;82(1):89–94. doi: 10.1007/s10340-008-0227-8. [DOI] [Google Scholar]
- 13.Carey JR. Applied demography for biologists with special emphasis on insects. New York: Oxford University Press; 1993. [Google Scholar]
- 14.Legaspi JC. Life history of Podisus maculiventris (Heteroptera: Pentatomidae) adult females under different constant temperatures. Environ. Entomol. 2004;33:1200–1206. doi: 10.1603/0046-225X-33.5.1200. [DOI] [Google Scholar]
- 15.Li J, et al. Effects of tomato chlorosis virus on the performance of its key vector, Bemisia tabaci in China. J. Appl. Entomol. 2018;142(3):296–304. doi: 10.1111/jen.12477. [DOI] [Google Scholar]
- 16.Chi H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988;17:26–34. doi: 10.1093/ee/17.1.26. [DOI] [Google Scholar]
- 17.Birch LC. The intrinsic rate of natural increase of an insect population. J. Animal Ecol. 1948;17:15–26. doi: 10.2307/1605. [DOI] [Google Scholar]
- 18.Carey JR. Applied demography for biologists with special emphasis on insects. Oxford University Press; 1993. [Google Scholar]
- 19.Awmack CS, Leather SR. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002;47(1):817–844. doi: 10.1146/annurev.ento.47.091201.145300. [DOI] [PubMed] [Google Scholar]
- 20.Salim M, Gökçe A, Bakhsh A. Demographic study of imidacloprid-resistant colorado potato beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomelidae) fed on transgenic and commercial potato cultivars. Phytoparasitica. 2022;50(1):201–221. doi: 10.1007/s12600-021-00937-5. [DOI] [Google Scholar]
- 21.Saljoqi AR, et al. Screening of various irrigated wheat varieties against angoumois grain moth, Sitotroga cerealella (Olivier) (Lepidoptera: Gelechiidae) J. Entomol. Zool. Stud. 2015;3:36–41. [Google Scholar]
- 22.El-Hafez A, et al. Acceptance and preference of pink bollworm and some lepidopterous eggs for parasitism by Trichogramma. Egypt. J. Agric. Res. 2001;79(2):123–132. [Google Scholar]
- 23.El-Wakeil NE. Evaluation of efficiency of Trichogramma evanescens reared on different factitious hosts to control Helicoverpa armigera. J. Pest. Sci. 2007;80(1):29. doi: 10.1007/s10340-006-0150-9. [DOI] [Google Scholar]
- 24.Nathan SS, et al. Effects of millet, wheat, rice, and sorghum diets on development of Corcyra cephalonica (Stainton) (Lepidoptera: Galleriidae) and its suitability as a host for Trichogramma chilonis Ishii (Hymenoptera: Trichogrammatidae) J. Environ. Entomol. 2006;35(3):784–788. doi: 10.1603/0046-225X-35.3.784. [DOI] [Google Scholar]
- 25.Ahmed S, Raza A. Antibiosis of physical characteristics of maize grains to Sitotroga cerealella (Olivier.) (Gelechiidae: Lepidoptera) in the free choice test. Pak. J. Life Soc. Sci. 2010;8(2):142–147. [Google Scholar]
- 26.Chi H, Su HY. Age-stage, two-sex life tables of Aphidius gifuensis (Ashmead) (Hymenoptera: Braconidae) and its host Myzus persicae (Sulzer) (Homoptera: Aphididae) with mathematical proof of the relationship between female Fecundity and the net reproductive rate. Environ. Entomol. 2006;35:10–21. doi: 10.1603/0046-225X-35.1.10. [DOI] [Google Scholar]
- 27.Yang T, Chi H. Life table and development of Bemisia argentifolii (Homoptera: Aleyrodidae) at different temperatures. J. Econ. Entomol. 2006;99(3):691–698. doi: 10.1093/jee/99.3.691. [DOI] [PubMed] [Google Scholar]
- 28.Huang YB, Chi H. Age-stage, two-sex life tables of Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae) with a discussion on the problem of applying female age-specific life tables to insect populations. Insect Sci. 2012;19:263–273. doi: 10.1111/j.1744-7917.2011.01424.x. [DOI] [Google Scholar]
- 29.Van Lenteren, J.C. & Noldus, L.P.J. Whitefly-plant relationship: behavioral and ecological aspects. In: Whiteflies: their Bionomics, Pest Status and Management (pp. 47–89), D. Gerling (Ed.), Intercept, Andover, Hants, UK. (1990).
- 30.Shafique M, Chaudry MA. Susceptibility of maize grains to storage insects. Pak. J. Zool. 2007;39(2):77. [Google Scholar]
- 31.Rizwana S, et al. Resistance in stored rice varieties against Angoumois grain moth, Sitotroga cerealella (Oliver) (Lepidoptera: Gelechiidae) Pak. J. Zool. 2011;43(2):343–348. [Google Scholar]
- 32.Demissie G, et al. Biochemical basis of resistance in different varieties of maize for their relative susceptibility to Sitotroga cerealella (Olivier) (Lepidoptera: Gelechiidae) J. Stored Prod. Postharvest Res. 2015;6:1–12. [Google Scholar]
- 33.Murad SM, Batool Z. Relative biochemical basis of susceptibility in commercial wheat varieties against angoumois grain moth, Sitotroga cerealella (Olivier) and construction of its life table. J. Biom. Biostat. 2017;8:333. doi: 10.4172/2155-6180.1000333. [DOI] [Google Scholar]
- 34.Akter T, Jahan M, Bhuiyan MSI. Biology of the Angoumois grain moth, Sitotroga cerealella (Oliver) on stored rice grain in laboratory condition. J. Asia. Bangladesh Sci. 2013;39(1):61–67. doi: 10.3329/jasbs.v39i1.16034. [DOI] [Google Scholar]
- 35.Shazali MEH, Smit RH. Life history studies of internally feeding pests of stored sorghum: Sitotroga cerealella (Olivier) and Sitophilus oryzae (L.) J. Stored Prod. Res. 1985;21(4):171–178. doi: 10.1016/0022-474X(85)90012-8. [DOI] [Google Scholar]
- 36.Sultan, R. et al. Biological parameters of Trichogramma chilonis Ishii (Trichogrammatidae: Hymenoptera) feeding on Sitotroga cerealella eggs at three constant temperatures. Pak. J. Agric. Res.26(1). (2013).
- 37.King BH. Offspring sex ratios in parasitoid wasps. Q. Rev. Biol. 1987;62:367–396. doi: 10.1086/415618. [DOI] [Google Scholar]
- 38.King BH. Host-size-dependent sex ratios among parasitoid wasps: Does host growth matter? Oecologia. 1989;78:420–426. doi: 10.1007/BF00379119. [DOI] [PubMed] [Google Scholar]
- 39.Charnov EL, et al. Sex ratio evolution in a variable environment. Nature. 1981;289:27–33. doi: 10.1038/289027a0. [DOI] [PubMed] [Google Scholar]
- 40.Ashamo MO, Khanna SC. Relative resistance of some corn varieties to the angoumois grain moth, Sitotroga cerealella (Olivier) J. Zool. Res. 2006;17(2):212–216. [Google Scholar]
- 41.Chi H, Liu H. Two new methods for the study of insect’s population ecology. Bull. Inst. Zool. Acad. Sin. 1985;24(2):225–240. [Google Scholar]
- 42.Chi, H. TWOSEX-MSChart: a computer program for the age-stage, two-sex life table analysis. (2021).
- 43.Khan I, et al. Appraisal of different wheat genotypes against Angoumois grain moth, Sitotroga cerealella (Oliv) Pak. J. Zool. 2010;42:161–168. [Google Scholar]
- 44.Chi H. Life-table analysis incorporating both sexes and variable development rate among individuals. Environ. Entomol. 1988;17:26–34. doi: 10.1093/ee/17.1.26. [DOI] [Google Scholar]
- 45.Khan J, et al. Studies on the biological attributes and parasitic potential of Trichogramma Chilonis (Ishii) reared on Sitotroga cerealella (Olivier) and Corcyra cephalonica (Stainton) eggs under laboratory conditions. Int. J. Entomol. Res. 2019;4(5):36–41. [Google Scholar]
- 46.Zeinalzadeh L, et al. Effect of four commercial barley varieties on life table parameters of Sitotroga cerealella (Olivier) (Lepidoptera: Gelechiidae) J. Crop Prot. 2016;5(2):293–305. doi: 10.18869/modares.jcp.5.2.293. [DOI] [Google Scholar]
- 47.Nadeem S, et al. Comparative rearing of Trichogramma chilonis (Ishii) (Hymenoptera: Trichogrammatidae) at different temperature conditions. Pak. Entomol. 2009;31(1):33–36. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.