

Abstract
Ischemic stroke is one of the most common causes of mortality and disability worldwide. However, treatment efficacy and the progress of research remain unsatisfactory. As the critical support system and essential components in neurovascular units, glial cells and blood vessels (including the blood-brain barrier) together maintain an optimal microenvironment for neuronal function. They provide nutrients, regulate neuronal excitability, and prevent harmful substances from entering brain tissue. The highly dynamic networks of this support system play an essential role in ischemic stroke through processes including brain homeostasis, supporting neuronal function, and reacting to injuries. However, most studies have focused on postmortem animals, which inevitably lack critical information about the dynamic changes that occur after ischemic stroke. Therefore, a high-precision technique for research in living animals is urgently needed. Two-photon fluorescence laser-scanning microscopy is a powerful imaging technique that can facilitate live imaging at high spatiotemporal resolutions. Two-photon fluorescence laser-scanning microscopy can provide images of the whole-cortex vascular 3D structure, information on multicellular component interactions, and provide images of structure and function in the cranial window. This technique shifts the existing research paradigm from static to dynamic, from flat to stereoscopic, and from single-cell function to multicellular intercommunication, thus providing direct and reliable evidence to identify the pathophysiological mechanisms following ischemic stroke in an intact brain. In this review, we discuss exciting findings from research on the support system after ischemic stroke using two-photon fluorescence laser-scanning microscopy, highlighting the importance of dynamic observations of cellular behavior and interactions in the networks of the brain’s support systems. We show the excellent application prospects and advantages of two-photon fluorescence laser-scanning microscopy and predict future research developments and directions in the study of ischemic stroke.
Key Words: astrocytes, blood-brain barrier, calcium signaling, glymphatic system, ischemic stroke, microglia, network, remodel, two-photon fluorescence laser-scanning microscopy, vessels
Introduction
Ischemic stroke is a major cause of mortality and morbidity worldwide (GBD 2019 Stroke Collaborators, 2021). Currently, reperfusion therapy performed within the appropriate time window for rescuing neurons in the ischemic penumbra is recommended as the first-line treatment (Powers et al., 2019; Turc et al., 2019; Berge et al., 2021). However, many patients cannot undergo reperfusion therapy due to contraindications. Even with emergency reperfusion treatments, there is still a considerable portion of patients with poor prognoses (Bhaskar et al., 2018; Sun et al., 2018; Rabinstein et al., 2019; Wollenweber et al., 2019; Valkonen et al., 2022). Removing the initiating factor of ischemia alone without addressing the subsequent cascade responses is not enough to achieve ideal outcomes.
Due to a lack of understanding of the communication among neuronal networks, glial cells, and vessels, the efficiency of alternative pro-neurogenesis or neuroprotective therapy remains uncertain (Li et al., 2018; Shi et al., 2018; Bang and Kim, 2019; Hatakeyama et al., 2020; Xu et al., 2020; Paul and Candelario-Jalil, 2021). As the critical support system for the brain and essential components of the neurovascular unit, glial cells and vessels (including the blood-brain barrier [BBB]) maintain an optimal microenvironment for neuronal function (Ransohoff, 2016b). They provide nutrients, regulate neuronal excitability, and prevent harmful substances from entering brain tissue (Allen and Lyons, 2018; Verkhratsky and Nedergaard, 2018; Caporarello et al., 2019; Garcia-Caceres et al., 2019; Liu et al., 2019; Sweeney et al., 2019; Schaeffer and Iadecola, 2021). The synchrony and coordination of neurons, glia, and blood vessels allow the brain to function well. Based on the idea that “structure determines function,” the functional network concept was first proposed for connective neurons. However, both neurons and the support system form functional networks, and the support system is involved in the regulation of neuronal networks (Kabba et al., 2018; Santello et al., 2019; Kirst et al., 2020; Schaeffer and Iadecola, 2021; Sun et al., 2021). These complex, highly heterogeneous, multicomponent, dynamic, and topologically heterogeneous networks of the support system in the brain ensure neuronal function and are involved in the development of neuropsychiatric disorders (Freeman, 2010; Chai et al., 2017; Iadecola, 2017; Greenhalgh et al., 2020; Kirst et al., 2020; Schaeffer and Iadecola, 2021; Endo et al., 2022; Silvin et al., 2022). Direct and reliable information about the dynamic and subtle changes in the networks of the support system for the brain can only be acquired from living tissue (Schaeffer and Iadecola, 2021). However, commonly used paradigms in ischemic stroke research do not provide high-level spatiotemporal observations in living systems (Table 1). It is difficult to reconstruct the dynamic changes in the networks of the support system from postmortem brain sections. Emerging fluorescent micro-optical sectioning tomography techniques can be used to rebuild whole brain networks but fail to reflect dynamic network functions, such as blood flow direction and velocity. Thus, techniques that can be used to understand the dynamic changes in the support system during ischemic stroke are essential to explain the pathophysiological processes and identify potential therapeutic targets. In 1990, Denk et al. first presented two-photon fluorescence laser-scanning microscopy (2PLSM; Figure 1), which has been widely used in live imaging. In this method, two long-wavelength photons are used to simultaneously excite one fluorescent probe from the ground state to an excited electronic state. Compared to conventional laser scanning confocal microscopy, 2PLSM uses infrared light (IR) for excitation, which greatly reduces absorption and scattering in biological tissues, allowing it to illuminate deeper tissues. This imaging protocol restricts fluorescence and photobleaching to the focal plane, greatly reducing damage to specimens. In addition, a wide-field photon detector without the use of pinholes recovers the scattered emission photons and further increases imaging depth. Thus, 2PLSM can be used to observe pathophysiological processes down to 600–800 μm in living tissue, depending on the degree of absorption and scattering and the power of the femtosecond laser (Helmchen and Denk, 2005; Diaspro et al., 2006). Nowadays, modern 2PLSM systems have now achieved imaging depths of up to 1000 μm throughout almost the entire gray matter of the mouse neocortex (Theer et al., 2003).
Table 1.
Commonly used ischemic stroke study paradigms
| Model | Description | Research technology | Merits | Demerits | |
|---|---|---|---|---|---|
| In vitro | Cell culture, OGD model | Cell cultrue under oxygen and glucose deprivation environment | Biochemical and molecular biological techniques | Simplification of the pathophysiological process | Far from reality, the absence of intact brain structure |
| Ex vivo | Acute brain slices, OGD model | Alive acute slices under oxygen and glucose deprivation environment | Electrophysiological, microscopy | Close to the in vivo environment Observe the deep brain tissue Facilitate for drug delivery | Influence cellular state when performing brain tissue sectioning |
| In vivo | MCAO | Introducing a filament into the origin of the middle cerebral artery through the internal carotid artery to block the blood flow | Postmortem research: biochemical and molecular biological techniques, the omics, optical microscopy, and micro-optical sectioning tomography | Reflect on the process in an intact postmortem animal | Reflect states at specific time points, inability to remodel dynamic processes |
| Photothrombotic stroke | Systemically applying a photosensitive dye (e.g., Rose Bengal) and illuminate it with light of a specific wavelength to block the blood flow | Living observation technologies: neuroimaging, laser Doppler flowmetry, in vivo electrophysiological and optogenetics | Dynamic recording of morphological and functional changes of cells and vessels in living animals. | The spatiotemporal resolution is limited. Glial cells and vascular elements are electrically silent. | |
| Endothelin-1 model | Locally applying the endothelin-1 on the brain to reduce the blood flow using its vasoconstrictive properties |
MCAO: Middle cerebral artery occlusion; OGD: oxygen-glucose deprivation.
Figure 1.
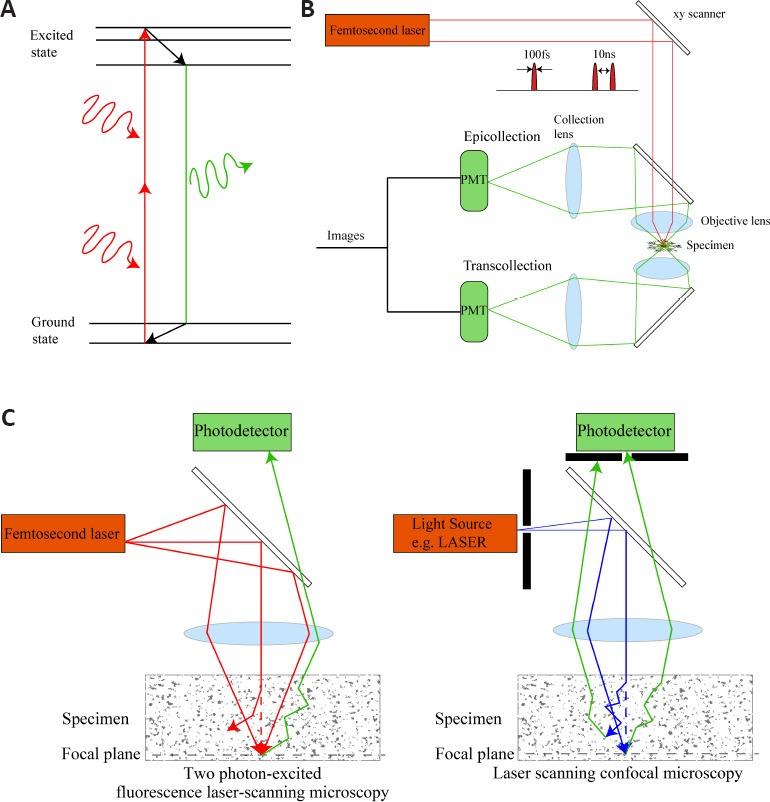
Simplified imaging strategy of 2PLSM and comparison of 2PLSM with laser scanning confocal microscopy.
(A) A simplified Jablonski diagram of the imaging process for 2PLSM. The two long-wavelength photons are used to simultaneously excite one fluorescent probe from the ground state to an excited electron state. (B) A simplified schematic of a 2PLSM with epifluorescence and transfluorescence collection. A femtosecond pulsed laser with a pulse width of 100 fs and a repetition period of 10 ns will excite light with a high density of photons, increasing the chance of excitation on the focal plane while reducing tissue damage. For intravital imaging, only epifluorescence collection is permitted. PMT refers to photomultiplier tube, used to detect weak photon signals and multiply them, converting them into electrical signals and finally forming images. (C) Different imaging strategies for scattered specimens using 2PLSM and laser scanning confocal microscopy. In 2PLSM, the paths of two ballistic photons and one scattered photon are shown (red lines). The energy of the scattered photon away from the focal plane is too low to excite fluorescent molecules. In laser scanning confocal microscopy, the paths of one ballistic photon and one scattered photon are shown (blue lines). The corresponding fluorescent photons are shown (green lines). The laser scanning confocal microscopy uses the pinhole to filter the off-focus plane fluorescence. However, fluorescent photons on the focal plane may also be scattered and filtered by the pinhole. 2PLSM: Two photon-excited fluorescence laser-scanning microscopy.
By introducing different kinds of fluorescence probes, 2PLSM can be used to observe dynamic changes in morphologies and functions in cells in living brains before and after ischemic stroke (Ricard et al., 2018). By combining 2PLSM with other techniques, researchers can obtain multimodal, in situ, time-resolved, and spatial-sequential brain imaging data and information in living animals. 2PLSM imaging turns static research into dynamic research, revealing structural and functional changes and reconstructing the functional networks in ischemic stroke, which can help identify new therapeutic targets and refine the time window for ischemic stroke. In this review, we first show the simple steps required to perform 2PLSM imaging. We mainly discuss the major findings from 2PLSM-based research on the support system during ischemic stroke, focusing on interactions between glial cells, vessels, the BBB, and neuronal networks. We hope this review will help researchers gain a better understanding of the pathophysiological processes of the support system after ischemic stroke.
Search Strategy and Selection Criteria
Studies cited in this review were obtained from searching the PubMed database (https://pubmed.ncbi.nlm.nih.gov) using the following keywords: stroke, ischemia, ischemic, astrocyte, microglia, vessel, blood-brain barrier, two-photon microscopy, multiphoton microscopy, reprogram, and glymphatic system. Studies cited in this review were published between 1989 and 2022. The literature search was completed by the author YF on July 25, 2022.
General Procedures for In Vivo Two-Photon Fluorescence Laser-Scanning Microscopy Imaging of Experimental Ischemic Stroke
The first procedure for in vivo 2PLSM imaging of experimental ischemic stroke is the preparation of the cranial window for imaging in living animals. The thick and inhomogeneous structures of intact skulls cause severe optical aberrations and scattering noise, which greatly reduces image quality (Helmchen and Denk, 2005; Yoon et al., 2020). Thus, craniotomy or thinned-skull cranial window surgery using dental drills is needed (Xu et al., 2007; Marker et al., 2010; Yang et al., 2010). Different types of cranial windows are shown in Figure 2A–C. In general, craniotomy enables researchers to perform operations on the brain parenchyma, such as direct dye coverage and stereotactic injection of viruses or substances. Acute cranial windows can be combined with electrophysiology to measure neuronal electrical signals with electrodes simultaneously with 2PLSM imaging. Compared with acute or chronic craniotomy, the thinned-skull cranial window minimizes damage to brain tissue at the expense of imaging quality. Thinned-skull cranial window surgery is widely used, especially for imaging sensitive cells, such as microglia, or for structures that do not require high precision, such as imaging amyloid plaques (Xu et al., 2007; Yang et al., 2010). The second procedure for imaging of experimental ischemic stroke is visualization the components of interest in the brain, which is the selection of the fluorescent labeling strategies (Trachtenberg et al., 2002). A schematic of 2PLSM in vivo imaging of neurons and support systems is shown in Figure 2D. The fluorescent labeling strategies can be roughly divided into cell morphology and cell function labeling, namely, labeling the soma and processes to show cell morphology or labeling important intracellular functional ions, such as intracellular calcium, to show cell function. The commonly used fluorescent labeling methods are shown in Table 2 (Hartmann et al., 2015a; Hierro-Bujalance et al., 2018; Tong et al., 2021). The third procedure is the selection of specific imaging strategies using a 2PLSM system, including different excitation wavelengths different fluorescent labels as well as frame acquisition rates and timescales for different components. This aspect requires extensive experience and practical tests. The relevant parameters used are summarized in other articles (Benninger and Piston, 2013; Fumagalli et al., 2014; Ricard et al., 2018; Adhikari et al., 2021; Leben et al., 2022). The fourth procedure is inducing the experimental ischemic stroke. There are two commonly used experimental ischemic stroke models for in vivo 2PLSM imaging, middle cerebral artery occlusion (MCAO) and photothrombotic stroke (PT). A schematic diagram and images of in vivo 2PLSM imaging for MCAO and PT are shown in Figure 2E–H. MCAO is a widely used model of experimental ischemic stroke. To simulate stroke infarction and reperfusion, a single filament is introduced into the internal carotid artery from the external carotid artery and advanced to block the origin of the middle cerebral artery, blocking blood flow, and left in place for a period of time before being removed (Longa et al., 1989; Chiang et al., 2011). MCAO can form the ischemic penumbra, but the infarct area is large and the corresponding damage to the animals is also large (Sigler and Murphy, 2010; Krafft et al., 2012). In PT, the infarct is induced by the systematic application of a photosensitive dye (usually rose bengal) and illumination with light of a specific wavelength to block focal cortical blood flow (Kim et al., 2000; Labat-gest and Tomasi, 2013). This approach results in a rapid, stable, sharp-edged infarct suitable for studies of cortical plasticity; however, the infarct is permanent and lacks the ischemic penumbra (Nishimura et al., 2006; Zhang and Murphy, 2007; Krafft et al., 2012; Labat-gest and Tomasi, 2013; Li and Zhang, 2021).
Figure 2.
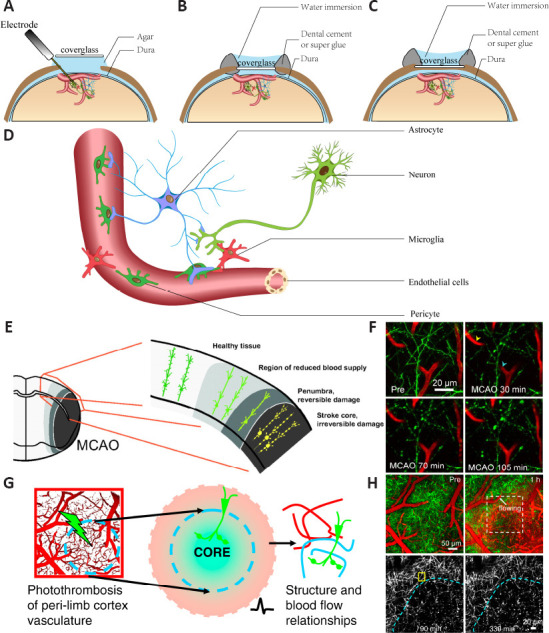
Schematic of in vivo 2PLSM imaging for experimental ischemic stroke.
(A–C) Different types of cranial windows for 2PLSM imaging. (A) Acute craniotomy skull window. This method can be combined with electrophysiology. (B) Chronic craniotomy skull window. To reduce the regeneration of the skull, the dura mater can be removed during surgery before covering the glass. (C) Thinned-skull window. The thickness of the skull is reduced to ~20 μm, reducing optical aberrations and scattering noise caused by the intact skull. (D) Schematic of 2PLSM in vivo imaging of neurons and the support system. (E) Schematic of in vivo 2PLSM imaging for middle cerebral artery occlusion (MCAO). The red box on the left side is out of the range where 2PLSM can be seen through the cranial window. Ideally, 2PLSM can continuously image the infarct core, penumbra, regions of reduced blood supply, and normal tissue after MCAO. (F) Real images obtained via 2PLSM after unilateral ligation of the common carotid artery followed by suture occlusion in MCAO. The picture shows rapid and extensive dendritic injury. (G) Schematic of in vivo 2PLSM imaging for photothrombotic stroke (PT). (H) Real images obtained via 2PLSM after PT. The picture shows the sharp-edged infarct in PT. 2PLSM: Two photon-excited fluorescence laser-scanning microscopy. E is reprinted from Sigler and Murphy (2010) with permission. F–H are reprinted from Zhang and Murphy (2007).
Table 2.
The commonly respective used fluorescent labeling methods for vessels, astrocytes and microglial cells
| Visualization strategy | Visualization methods | Comments | Reference | |
|---|---|---|---|---|
| Vessels | ||||
| Structure labeling | Intravenous injection fluorescently labeled vascular tracers, e.g., Evans blue, Fluorophore conjugated dextran (including FITC-dextran, Texas red-dextran, and Alexa 680-dextran) | Fluorescent label the serum | Dirnagl et al., 1992; Rovainen et al., 1993; Kubotera et al., 2019 | |
| Intravenous injection plant lectin conjugated with fluorophores, e.g., tomato lectin, FITC-wheat germ agglutinin lectin | Imaging the endothelial glycocalyx. | Robertson et al., 2015; Yoon et al., 2017 | ||
| Tg eGFP-Claudin5 transgenic mice | Imaging the tight junction between endothelial cells. Expressing eGFP and Claudin5 under the direction of the endothelial-specific receptor tyrosine kinase (Tek, formerly, and Tie2) promoter. |
Knowland et al., 2014 | ||
| Tie2-GFP transgenic mice or crossing Tie2-Cre with floxed fluorescent protein reporter mouse line | Expressing GFP under the direction of the endothelial-specific receptor tyrosine kinase (Tek, formerly, Tie2) promoter. Widely used in imaging the endothelial cells. The homozygous Tie2-GFP mice have retinal degeneration and are blind in adulthood, which may affect their performance in behavioral tests after ischemic stroke. |
Motoike et al., 2000; Kisanuki et al., 2001 | ||
| Crossing Tie2-CreER transgenic mice with floxed fluorescent protein reporter mouse line | Forde et al., 2002 | |||
| NG2DsRedBAC transgenic mice or crossing NG2-CreERTM with floxed fluorescent protein reporter mouse line. | Expressing fluorescent protein driven by the promoter for NG2 proteoglycan. Mice express red fluorescent protein, DsRed, driven by the control of promoter for NG2 proteoglycan. Labeling mural cells as well as polydendrocytes and oligodendrocyte progenitor cells. |
Hartmann et al., 2015b | ||
| PDGFRβ-tdTomato or PDGFRβ-CreERT2 transgenic mice | Expressing fluorescent protein driven by the promoter for PDGFRβ, labeling mural cells. Cannot separate pericytes and smooth muscle cell. |
Hartmann et al., 2015b | ||
| αSMA-mChrerry or crossing αSMA-CreERT2 with floxed fluorescent protein reporter mouse line | Expressing fluorescent protein driven by the promoter for αSMA, labeling mural cells. | Wendling et al., 2009; Armstrong et al., 2010 | ||
| Functional labeling | Intravenous injection fluorescent vascular tracers, including: Fluorophore conjugated dextran, e.g., FITC-dextran Texas red-dextran Alexa 680-dextran | Fluorescent label the serum. The velocity and direction of blood flow are calculated from the unlabeled red blood cell flow paths through time lapse 2PLSM imaging. |
Ngai and Winn, 1996; Kleinfeld et al., 1998; Saunders et al., 2015 | |
| Crossing PDGFRβ-CreERT2 with Rosa26< LSL-GCaMP6s> (Ai96) transgenic mice | Expressing GCaMP6s driven by the promoter of PDGFRβ. GCaMP cpGFP, CaM, and the Ca2+/CaM-binding M13pep. Intracellular calcium concentration fluctuations shown by the fluorescence intensity. Imaging the fluctuation of intracellular calcium in mural cells. |
Glück et al., 2021 | ||
| Acta2-RCaMP1.07 | Expressing RCaMP1.07 driven by the promoter of αSMA. RCaMP1.07 is a red fluorescent protein with a calmodulin binding domain. |
Meza-Resillas et al., 2021 | ||
| Intravenous injection of fluorescently labeled vascular tracer that does not cross the blood-brain barrier under normal conditions, e.g., Evans blue, fluorophore conjugated dextran (including FITC-dextran, Texas red-dextran, Alexa 680-dextran), Alexa-albumin | Calculation the permeability of BBB. | Saunders et al., 2015; Ahishali and Kaya, 2021 | ||
| Astrocytes | ||||
| Structure labeling | Direct application of dyes to the cortical surface or intravenous injection sulforhodamine 101 (SR101) sulforhodamine B (SRB) | Not very specific for labeling astrocytes. | Nimmerjahn et al., 2004; Vérant et al., 2008a; Appaix et al., 2012; Hill and Grutzendler, 2014 | |
| GFAP-GFP or GFAP-eGFP transgenic mice | Expressing GFP or eGFP driven by the promoter of GFAP. Only reactive astrocytes are labeled. Simultaneous labeling of neurons. |
Nolte et al., 2001 | ||
| Aldhlll-eGFP | Expressing eGFP dricen by the aldehyde dehydrogenase 1 family member L1 (Aldh1l1) promoter/enhancer regions. | Mills et al., 2022 | ||
| Functional labeling | Direct coloading Fluo-4 acetoxymethyl (AM) ester to the cerebral cortex | Imaging in vivo cytosolic calcium fluctuation in astrocytes. | Hirase et al., 2004 | |
| Injection the Ca2+-sensitive fluorescent indicator Oregon Green 488 BAPTA-1 (OGB-1) and SR101 into the brain. | Not very specific for labeling astrocytes. | Ma et al., 2021 | ||
| Crossing Aldh1l1-Cre/ERT2 transgenic mice with floxed fluorescent protein reporter mouse line or stereotactic injection Cre-dependent FLEX-GFP AAVs in Aldh1l1-Cre/ERT2 BAC transgenic mice | Srinivasan et al., 2016 | |||
| Microglial cells | ||||
| Structure labeling | Incubating brain slices in FITC-IB4 | Imaging microglia in living brain slices and slice cultures | Dailey et al., 2013 | |
| Intravenous injection plant lectin conjugated with fluorophores (tomato lectin, isolectin IB4) | No need to breed transgenic mice. Quick and easy marking. Labeled volumes are limited. Simultaneous labeling of partial endothelial cells. Longitudinal imaging is not possible since the requirement for intraparenchymal injection of the fluorophore. |
Brawek and Garaschuk, 2017 | ||
| Intravenous injection peridinin chlorophyll conjugated Iba-1 and aminomethyl coumarin acetate conjugated CD68 into wild type mice before 2PLSM | No transgenic mice required. Blocking receptors on microglia. |
Bok et al., 2015 | ||
| F4/80 or CD68 promoter in combination with the surface-mutated AAV6 capsid | Selective GFP expression in vivo after intracerebroventricular injections in wild-type mice. May induce pathological conditions. | Rosario et al., 2016 | ||
| LV.miR-9.T, microRNA-9-regulated vector | Microglia of rodent brain do not express miR-9. Exclusion of peripheral bone marrow cells. Not entirely selective, depending on cell expression level of microRNA-9. |
Åkerblom et al., 2013 | ||
| CX3CR1-GFP transgenic mice CX3CR1-EGFP transgenic mice | Expressing GFP or EGFP sequence replacing the first 390 bp of the coding exon (exon 2) of the chemokine (C-X3-C motif) receptor 1 (Cx3cr1) gene. The most widely used fluorescent labeling techniques of microglia. Simultaneous labeling microglia and peripheral monocytic cells. Co-imaging microglia and neurons requires cross with Thy1-GFP transgenic mice, thus neurons and microglia have the same fluorophore (GFP) and may not be well distinguished from neurons by morphology. Partial knockdown of CX3CL1/fractalkine pathway, which may influence the pathophysiological process. |
Jung et al., 2000; Davalos et al., 2005; Nimmerjahn et al., 2005; Wake et al., 2009; Goldmann et al., 2016; Wieghofer and Prinz, 2016; Haimon et al., 2018 Fumagalli et al., 2013; Brawek and Garaschuk, 2017; Hickman et al., 2019 | ||
| Iba1-EGFP transgenic mice | Expressing the EGFP under the direction of Iba1 gene. Widely used fluorescent labeling technology of microglia. Simultaneous labeling microglia and peripheral monocytic cells. Weak GFP fluorescence expression. Iba-1 expression is unstable in the same cells, correlating with Iba-1 protein expression level. |
Hirasawa et al., 2005 | ||
| CD11b-CreERT2 cross R26-tdTomato transgenic mice | Only labels partial microglia. Simultaneous labeling of microglia and peripheral blood mononuclear macrophages. |
Füger et al., 2017 | ||
| CD11b-CreERT2;R26-tdTomato;Iba1-eGFP | Labels all microglia. Simultaneous labeling of microglia and peripheral blood mononuclear macrophages. |
Füger et al., 2017 | ||
| Tmem119-tdTomato reporter mouse Tmem119-EGFP and Tmem119-CreERT2 transgenic mice | Expressing tdTomato or EGFP driven by the promoter of TMEM119. Specifically label microglia. Not affect cell function and physiological function. Using the CRISPR-Cas9 technology. |
Ruan et al., 2020 Kaiser and Feng, 2019 | ||
| Functional labeling | Staining calcium in microglia with OGB-1 using a single-cell electroporation technique | First fluorescent labeling of intracellular calcium signals in microglia. Failing to render calcium signaling changes in the entire microglial cell network. |
Eichhoff et al., 2011 | |
| Injection of recombinant retroviral vector to introduce GCaMP2 | Labeling microglia network calcium signaling. Retroviruses infect only dividing cells; stab wound injury is required to stimulate microglia proliferation. | Seifert et al., 2011 | ||
| Twitch-2B in microglia by a microglia-specific microRNA-9-regulated viral vector | Expression of a genetically-encoded ratiometric Ca2+ sensor Twitch-2B in microglia. Not entirely selective, depending on cell expression level of microRNA-9. |
Brawek et al., 2017 | ||
| A Cre-dependent GCaMP5G line, termed PC::G5-tdT (Polr2a, CAG, GCaMP5G, tdTomato) | No effect on physiological functions of transgenic mice. Visualizing calcium signaling of microglia. Low expression level of calcium fluorescence signal. Need to breed transgenic animals. |
Gee et al., 2014 |
2PLSM: Two-photon fluorescence laser-scanning microscopy; AAV: adeno-associated virus; BBB: blood-brain barrier; CaM: calmodulin; cpGFP: circularly permuted green fluorescent protein; eGFP: enhanced green fluorescent protein; FITC: fluorescein isothiocyantate; GFAP: glial fibrillary acidic protein; GFP: green fluorescent protein; Iba1: ionized calcium-binding adaptor molecule 1; M13pep: “M13” peptide; NG2: neural/glial antigen 2; PDGFRβ: platelet-derived growth factor receptor β; TMEM119: transmembrane protein 119; αSMA: α-smooth muscle actin.
Vessel Dysfunction and Recovery after Ischemic Stroke
Blood vessels, as pipelines for blood circulation, supply neurons and glial cells with essential nutrients, carry away metabolic waste, and form the barrier between the blood and cerebral parenchyma. In intact brains, blood vessels constitute a unique hierarchical 3D network (Kirst et al., 2020). Dysfunction and recovery in hemorheology, hemodynamics, and architecture of the vessel network significantly affect the pathological process of ischemic stroke and are associated with prognosis. In the acute phase of ischemic stroke, reperfusion therapy saves neurons in the ischemic penumbra and reduce mortality; this is regarded as the first-line therapy in ischemic stroke (Powers et al., 2019; Turc et al., 2019; Berge et al., 2021). However, timely and effective recanalization is not always accompanied by downstream tissue reperfusion, which has been described as the “no-reflow phenomenon” (Kloner, 2011; Bai and Lyden, 2015). Good collateral circulation is an independent predictor of improved outcomes and is a potential therapeutic target (Vernieri et al., 2001; Miteff et al., 2009; Shuaib et al., 2011; Vagal et al., 2018; Guglielmi et al., 2019; Broocks et al., 2020). In the recovery phase of ischemic stroke, angiogenesis in the peri-infarct region has been correlated with longer survival times in patients and coupled with neuronal remodeling (Kanazawa et al., 2019; Hatakeyama et al., 2020). Therefore, detailed intravital information on post-stroke hemodynamics, hemorheology, and vascular network architecture can help to gain a comprehensive understanding of the pathophysiological mechanisms of ischemic stroke and to develop corresponding therapies. By applying intravenous vascular tracers to label blood plasma, researchers can efficiently study the hemorheology, hemodynamics, and structure of hierarchical vessels (Figure 3A–C). Using transgenic fluorescent markers in animals, researchers can further image different cellular components of microvessel networks (Table 2). In this section, the network of vessels mainly refers to the cerebral microvessel system.
Figure 3.
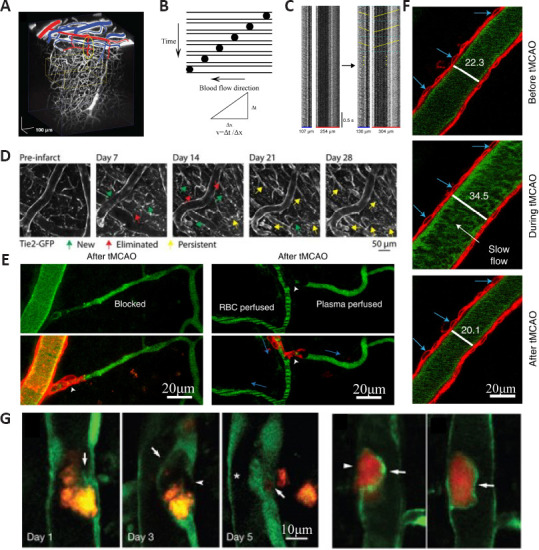
Vessels observed by 2PLSM.
(A) Reconstructing the 3D network structure of blood arteries in the cortex from in vivo 2PLSM. Surface arterioles are marked in red and venules are marked in blue. Yellow ovals indicate the target penetrating arteriole. (B) Intravenous injection of fluorescent dyes to label blood plasma and measure blood flow velocity using high-speed line-scan technique. The fluorescent-labeled blood plasma is bright, and red blood cells are dark, represented by dark dots. Red blood cells flow within the vessel following the blood flow direction and form diagonal streaks. Calculating the slopes of the streaks can obtain the blood flow velocity. (C) Kymographs of the same vessels after nonlinear transformation so that the horizontal axis represents 3D distance. For flow speed measurement, each kymograph was divided into 0.5-second blocks (cyan dashed lines). (D) The recovery process of vessels from photothrombotic stroke (PT) in Tie2-GFP transgenic mice. (E) Real images obtained from in vivo 2PLSM in transgenic mice labeling mural cells after transient middle cerebral artery occlusion (tMCAO). This image shows the flow outcome of an arteriole and its downstream capillary after tMCAO. (F) Example image showing an arteriole before, during, and after tMCAO (blue arrows indicate the same vascular smooth muscle cells; numbers indicate vessel diameter). Scale bar, 20 µm. (G) Real images for embolus extravasation obtained from in vivo 2PLSM. 2PLSM: Two photon-excited fluorescence laser-scanning microscopy; GFP: green fluorescent protein. A is reprinted from Nishimura et al. (2010) with permission. C–G are reprinted from Fan et al. (2020), Williamson et al., (2020), Hill et al. (2015), Tong et al. (2021), and Lam et al. (2010) with permission, respectively.
Hemodynamics and hemorheology in the acute phase of ischemic stroke
The strong and rapid compensatory capacity and plasticity of the vascular network against local ischemia are important for the survival of neurons in the infarct core in the acute phase of ischemic stroke. Schaffer et al. (2006) found that, after photothrombotic occlusion to individual pial arterioles, the blood flow was rapidly re-established at the first branch downstream without neuronal apoptosis or peri-photothrombosis, revealing a robust redistribution and compensatory ability of blood flow after surface arteriole occlusion. Shih et al. (2009) found that the MCAO model caused red blood cell flow reversal in surface arterioles in the penumbra to ensure blood flow in penetrating arterioles, which compensated for the incomplete recovery of blood flow during reperfusion. However, this compensatory capacity is related to the structural level of the occluded vessel, and not all levels can be compensated by endogenous mechanisms of blood flow after vascular occlusion. Using 2PLSM, researchers can obtain cortical vascular network structures, classify vascular hierarchy, and target vessels for embolization (the 3D network structure of cortical vasculature is shown in Figure 3A). They can identify the surface communication network, the subsurface microcirculation network, and penetrating arterioles that connect the two. After occlusion of penetrating arterioles, nearby penetrating and surface arterioles did not dilate, and there was no restoration of blood flow in occluded penetrating arterioles, although the capillaries directly downstream from the occluded arteriole dilated (Nishimura et al., 2007; Nishimura et al., 2010). To further assess the response of different levels of vessels and collateral blood flow to focal cortical ischemia, Luo et al. (2017) further used in vivo 2PLSM to observe interactions between the surface and subsurface microcirculatory networks after ischemia. They performed five different combinations of occlusion of the target arterioles, their collateral surface vessels, and penetrating arterioles (Luo et al., 2017). After in vivo staining of dead cells, behavioral tests, and histological analysis, they observed protective roles of collateral flow within the subsurface microcirculation network in neural circuits in the ischemic penumbra (Luo et al., 2017). They first demonstrated that collateral blood vessels at different levels of the endogenous cerebrovascular hierarchy compensated for decreased blood flow during ischemia by penetrating arterioles (Luo et al., 2017). These findings demonstrated the important compensatory function of collateral blood flow during the acute phase of ischemic stroke and highlighted the different responses and different roles of vessels at different levels of the vascular network during acute ischemic conditions. However, the specific compensatory mechanisms of the cerebral arteriole system in response to ischemic stroke is not clear and may be related to the response of precapillary sphincters, smooth muscle cells, pericytes, and even astrocytes to hypoxia and diffuse depolarization as shown in previous studies (Chuquet et al., 2007; Fernández-Klett et al., 2010; Mishra et al., 2016; Alarcon-Martinez et al., 2020; Hartmann et al., 2021; Meza-Resillas et al., 2021; Zambach et al., 2021).
To enhance the compensatory capacity of the collateral circulation and neural remodeling, remote ischemic conditioning is considered an effective treatment (Liao et al., 2019; McDonald et al., 2021). Remote ischemic preconditioning is a kind of remote ischemic conditioning which involves inducing acute transient ischemia in the distant limb after the onset of ischemic stroke but before reperfusion treatment (Ma et al., 2020a, b). Ma et al. (2020a, b) used aged rats to mimic human ischemic stroke and found that using remote ischemic preconditioning before experimental ischemic stroke could enhance collateral flow by preventing the stroke-induced narrowing of pial arterioles, which reduced the infarct area. However, the mechanisms of enhanced collateral blood flow induced by remote ischemic preconditioning was not further discussed in this study. The role of humoral factors needs to be further studied.
Recovery of hemodynamics, hemorheology, and vascular network architecture after ischemic stroke
Dynamic changes seen with embolisms in vivo are important because thromboses are the main initiator of ischemic stroke and are thought to be closely related to revascularization and prognosis (Powers et al., 2019; Turc et al., 2019; Berge et al., 2021). In 2010, using in vivo 2PLSM, Lam et al. (2010) first observed dynamic changes in microemboli in cerebral microvessels in living mouse brains. They used fluorescently labeled microemboli and found that embolus extravasation is an alternative recanalization mechanism when hemodynamic forces and the fibrinolytic system fail to clear the thrombus. The process of embolus extravasation occurs 2–7 days after thrombosis and is mediated by a novel mechanism of microvascular plasticity (Figure 3G; Lam et al., 2010). The rate of embolus extravasation is significantly reduced in aged mice or after inhibiting matrix metalloproteinase 2/9 activity (Lam et al., 2010). El Amki et al. (2020) induced a fibrin rich clot thrombin in the middle cerebral artery (MCA) followed by intravenous t-PA thrombolysis to mimic stroke and intravenous thrombolytic therapy, which differed from the classical MCAO model using filament to induce ischemia.They fluorescently labeled neutrophils, used in vivo 2PLSM to image the distal capillary flow after recanalization in mice and demonstrated that the no-reflow phenomenon after reperfusion therapy might be due to cortical microvascular occlusion caused by neutrophils (El Amki et al., 2020). These in vivo 2PLSM studies, using fibrin thrombi or cholesterol emboli, closely simulated the pathophysiological process of human ischemic stroke, which enabled researchers to dynamically observe changes in embolism in vivo with high spatiotemporal resolution. By monitoring blood flow in vivo after PT, Schrandt et al. (2015) tracked long-term vascular changes and found that a flow deficit remained even 35 days after occlusion, suggesting that more time was necessary for full perfusion recovery.
Endothelial cells are indispensable components of the vasculature, and labeling endothelial cells rather than plasma can enable imaging of the whole vascular system without missing any capillaries (Williamson et al., 2020). Williamson et al. (2020) used transgenic mice expressing fluorescent proteins in endothelial cells to observe changes in the microvessel system after ischemic stroke (Figure 3D). Using in vivo 2PLSM and multi-exposure speckle imaging, they visualized the processes of structural vascular plasticity and the reconstruction of peri-infarct blood flow. They associated the rebuilding of blood vessels with neuronal network function recovery based on 2PLSM imaging of microvessels, multi-exposure speckle imaging of blood flow, and behavioral testing. They demonstrated that the extent of vascular structural plasticity predicted local blood flow reconstruction, which predicted the recovery of neuronal function.
Breakdown of the BBB and the roles of peripherally derived cells
The BBB is a critical diffusion barrier between the brain parenchyma and cerebral capillaries that can prevent the influx of most toxic substances and provide essential nutrients (e.g., oxygen and glucose) to maintain brain homeostasis (Liebner et al., 2018; Vanlandewijck et al., 2018; Sweeney et al., 2019; Cheng et al., 2022). The BBB mainly comprises endothelial cells and tight junctions, astrocyte endfeet, and pericytes (Liebner et al., 2018; Sweeney et al., 2019; Profaci et al., 2020; Zou et al., 2021). To date, all of these components are considered to be indispensable to maintain the integrity and function of the BBB (Liebner et al., 2018; Caporarello et al., 2019; Heithoff et al., 2021). BBB dysfunction is commonly involved in the pathological processes that occur during the acute phase of ischemic stroke and is associated with ischemic stroke outcomes (Lasek-Bal et al., 2019; Li et al., 2019b; Bernardo-Castro et al., 2020). The integrity of the BBB is essential for maintaining homeostasis and supporting the function of neuronal and glial networks, which can be easily represented by permeability to fluorescent dyes during in vivo 2PLSM (see detailed information of calculation methods in Figure 4). In this section, we focus on the roles of endothelial cells and pericytes in the breakdown of the BBB and the roles of peripherally derived immune cells in ischemic stroke, while the role of astrocytes in maintaining the BBB is detailed in the section “Roles of communication between astrocytes and multiple cell types in the acute phase of ischemic stroke”.
Figure 4.
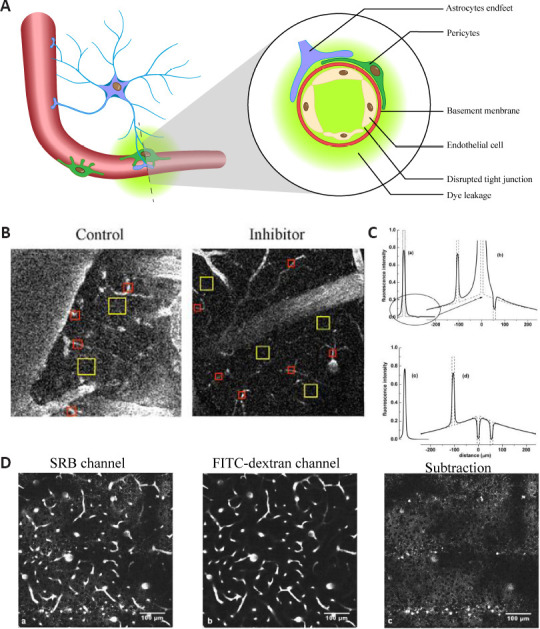
Calculation of BBB permeability.
(A) Leakage pattern of the BBB. (B) Calculating BBB permeability by quantifying the regions of interest and measuring fluorescence intensity before and after ischemic stroke. Red squares represent vascular areas and yellow squares represent interstitial spaces; they were manually defined. (C) Schematic numerical simulation of the differential imaging method for calculating the permeability of the BBB without the need for shape recognition. Co-injection of two dyes, one of high molecular weight (fluorescein-labeled dextran, FITC-dextran, 70 kDa, green fluorescence) and one of low molecular weight (sulforhodamine B [SRB], 559 Da, red fluorescence). The gains of the two channels were adjusted so that the signals coming from within the vessels were equal. Subtraction of the image of the FITC-dextran from that of the SRB gave images in which the vasculature was invisible and the SRB in the parenchyma could be imaged with high resolution. Algorithms are presented for rapidly quantifying the extravasation. (D) Actual pictures from the differential imaging method. The left image is under the SRB channel, the middle image is under the FITC-dextran channel, and the right image was obtained by subtracting the FITC-dextran image from the SRB image. BBB: Blood-brain barrier; FITC: fluorescein isothiocyantate. B is reprinted from Caballero-Garrido et al. (2015) with permission. C and D are reprinted from Vérant et al. (2008b) with permission.
Researchers using in vivo 2PLSM found that the PT model was more inclined to produce early BBB injury than MCAO, revealing that the two commonly used models of experimental ischemic stroke may differ in their mechanisms (Frederix et al., 2007; Bragin et al., 2016). This makes photochemical embolization a useful tool to study the pathological mechanisms of BBB breakdown. However, whether the pathophysiology of BBB damage caused by thromboembolism and that caused by direct photodamage are different remains to be studied (Frederix et al., 2007; Kleinschnitz et al., 2008; Labat-gest and Tomasi, 2013; Bragin et al., 2016; Cotrina et al., 2017).
Endothelial cells and tight junctions play an essential role in maintaining the integrity of the BBB. Knowland et al. (2014) used Tg eGFP-Claudin5 transgenic mice to label tight junctions between endothelial cells (Knowland et al., 2014). They found that the recruitment of transcellular and paracellular pathways of endothelial cells, rather than tight junction breakdown, mediated early-stage BBB dysfunction in the transient MCAO experimental model (Knowland et al., 2014). This discovery revealed a new mechanism of BBB damage after ischemic stroke. In addition, Ernesto Caballero-Garrido et al. (2015) found that injection of an inhibitor of miR-155, a potential regulator of endothelial morphogenesis, could reduce BBB permeability and increase red blood cell flow in a distal MCAO model.
Regarding pericytes, the soma of pericytes is immobile, while the processes exhibit dynamic extension and retraction. After selective ablation of individual pericytes, neighboring pericytes extend their processes to contact uncovered regions of the endothelium (Berthiaume et al., 2018b). Pericyte-labeled transgenic mice were imaged using in vivo 2PLSM to examine the relationship between pericytes and the integrity of cortical capillaries after PT (Underly et al., 2017; Berthiaume et al., 2018a, b; Underly and Shih, 2020). This showed that, during the acute phase of PT, matrix-metalloproteinase-9 synthesis in pericytes mediated tight junction injury, as reflected by greater leakage of fluorescent dye in regions where pericyte somata adjoined the endothelium (Underly et al., 2017; Berthiaume et al., 2018a, b; Underly and Shih, 2020). Further study of the related molecular mechanisms showed that mice with pericyte-specific deletion of SENP1, a protein with reverse SUMOylation function, promoted the formation of cerebral microvascular thrombosis faster than that observed in wild-type mice and exacerbated poststroke neuronal damage after PT (Sun et al., 2020).
Peripherally derived cells that infiltrate into the brain parenchyma are also largely involved in the post-stroke pathophysiology (Jayaraj et al., 2019; Yang et al., 2019; Cai et al., 2022; Wicks et al., 2022). Combining intravital 2PLSM with flow cytometry and quantitative real-time PCR, Neumann et al. (2018) described the dynamics of tdTomato-labeled neutrophils and GFP-labeled microglia after PT in bone marrow chimeric mice. A cross of CX3CR1-GFP and LysM-eGFP mice was used to simultaneously image neutrophils and microglia. They found that neutrophils rapidly entered the brain parenchyma from the activated site of endothelial cells in ischemic stroke and mediated the microglial immune response (Neumann et al., 2018). Systemic blockade of VLA-4 could effectively reduce the entry of neutrophils and reduce post-ischemic injury (Neumann et al., 2018). This study confirmed neutrophil infiltration into the brain parenchyma and long-term contact with microglia (Neumann et al., 2018). However, the lethal irradiation used may destroy the BBB and be a limited reflection of the real pathophysiological processes of ischemic stroke (Li et al., 2004).
For imaging specific immune cells (T cells and B cells) after stroke, the observation time must be relatively prolonged. Using sagittal brain slices from adoptive lymphocyte transfer mice and hCD2-GFP transgenic mice after ischemic stroke, Ortolano et al. (2010) performed in situ real-time T-cell imaging in ischemic stroke. In 2011, Fumagalli et al. further used hCD2-GFP transgenic mice to exhibit GFP-labeled T cells combined with in vivo multiphoton microscopy to visualize, define, and quantitatively analyze the movement and behavior of T cells. Li et al. (2017) used intravital 2PLSM imaging and showed that C–C chemokine receptor type 5 was critical for regulatory T cells to dock in the injured vessel wall and interact with blood-derived neutrophils/macrophages after MCAO. In vivo and in vitro experiments showed that C–C chemokine receptor type 5 was important for regulatory T cells to reduce BBB damage after ischemic stroke. Ortega et al. (2020) used whole-brain volumetric serial two-photon tomography and a custom-developed image analysis pipeline to visualize and quantify B cell diapedesis throughout the brain in B cell adoptive transfer mice after temporary MCAO.
Dual Roles of Astrocytes in Ischemic Stroke
Astrocytes, highly heterogeneous glial cells in the brain, tile the entire CNS, form interconnected astrocytic networks, and interact with neurons and blood vessels (Nedergaard et al., 2003; Volterra and Meldolesi, 2005; Perea et al., 2009; Santello et al., 2019). Astrocytes assist in maintaining brain tissue homeostasis and supporting neurons (Liddelow and Barres, 2017; Verkhratsky and Nedergaard, 2018; Santello et al., 2019). Single-cell sequencing and transcriptome sequencing have shown that astrocytes transform into protective reactive astrocytes after ischemic stroke and are characterized by upregulated neurotrophic factors and extracellular matrix-associated protein expression (Zamanian et al., 2012; Liddelow et al., 2017). However, the exact impact of reactive astrocytes and the astrocytic scar remains controversial (Diniz et al., 2019; Yang et al., 2020; Zhou et al., 2020). On the one hand, reactive astrocytes regulate the extracellular matrix and form the astrocytic scar after ischemic stroke, which can isolate and limit the harmful environment of the infarction core. On the other hand, astrocytic scars may impair axonal regeneration and neuronal plasticity (Pekny et al., 2014; Sun et al., 2019; Shen et al., 2021). Using 2PLSM, researchers can observe the longitudinal morphological and functional changes in astrocytes; the communication among astrocytes, neurons, and vessels; and the influence of astrocytes on neuronal survival and blood vessel rebuilding after ischemic stroke. Thus, we can learn more about the dual roles of astrocytes in ischemic stroke and their effects on outcomes.
Roles of astrocytes in the acute phase of ischemic stroke
Changes in astrocyte behaviors
2PLSM has been used to investigate the dynamic morphological and positional changes in astrocytes after ischemic stroke. Using an in vivo cardiac arrest model and ex vivo oxygen/glucose deprivation (OGD) model in living cortical slices, Risher et al. (2009, 2012) found that swollen astrocytes are the primary sources of cytotoxic cerebral edema after ischemic stroke (Risher et al., 2009, 2012). They found that the spreading depolarization (SD) mediated the volume increase in astrocytes, and astrocyte volume changes could be reversed, possibly due to the distribution of aquaporin 4 (AQP4) protein on astrocytes (Risher et al., 2012). However, Rakers et al. (2017) used AQP4-knockout transgenic mice and found that AQP4 did not affect SD-related edema of astrocytes in vivo. These findings illustrated a contribution of the astrocyte volume increase in post-stroke cell-derived edema, but the mechanisms of this volume increase were likely unrelated to AQP4. Although this water channel protein is specifically expressed in astrocytes, it is mainly densely expressed on the endfeet and is more likely to be associated with the maintenance of the BBB and has a limited relationship with astrocyte volume regulation (Manley et al., 2004; Friedman et al., 2009; Akdemir et al., 2014; Yao et al., 2015; Heithoff et al., 2021).
Roles of communication between astrocytes and multiple cell types in the acute phase of ischemic stroke
Traditionally, astrocytes are considered to be electrically silent cells. However, recent studies have found that the critical role of multiple ion channels in modulating cell membrane potential in astrocytes (McNeill et al., 2021; Armbruster et al., 2022). Using calcium imaging techniques, researchers have found that astrocytes frequently bidirectionally communicate with surrounding glia, neurons, and microvessels, and this communication process can be reflected by changes in intercellular signaling in astrocytes (Perea et al., 2009). Astrocytes integrate and respond to signals from neurons and vessels. 2PLSM is well suited for tracing dynamic intercellular signaling changes in astrocytes through calcium imaging techniques in live tissue. Interestingly, the [Ca2+]i transients in the processes of astrocytes differ from those in the soma in terms of spatiotemporal characteristics (Bazargani and Attwell, 2016). [Ca2+]i transients in astrocytic processes originate from two pathways: extracellular calcium influx through ion channels (i.e., transient receptor potential channels) and release from intracellular stores. [Ca2+]i transients in the soma, on the other hand, depend largely on release from intracellular stores, which is mainly mediated by IP3R2 signaling (Bazargani and Attwell, 2016; Bindocci et al., 2017). The [Ca2+]i transients in astrocytes may contribute to the Ca2+-dependent release of glial glutamate after ischemic stroke, which mediates excitotoxicity in neuronal networks (Takano et al., 2009; Siracusa et al., 2019; Kirdajova et al., 2020).
Bidirectional communication between astrocytes and neurons received attention as early as the 1990s, and the term “tripartite synapse” was proposed to describe this bidirectional communication (Cornell-Bell et al., 1990; Charles et al., 1991). Changes in communication between astrocytes and neurons are worthy of exploration.
Ding et al. (2009) used in vivo 2PLSM imaging and found that, during the acute phase of PT, [Ca2+]i transients increased synchronously in astrocytes and propagated as waves in the astrocytic network. Using BAPTA to selectively inhibit [Ca2+]i transients in astrocytes could significantly reduce infarct volume. Additional studies have shown that IP3R2 receptor knockout mice and TRPV4 knockout mice, which exhibit reduced internal calcium release and external calcium inflow in astrocytes, respectively, also exhibited reduced glia-dependent glutamate release and performed better after experimental ischemic stroke (Ding et al., 2009; Dong et al., 2013; Rakers and Petzold, 2017; Rakers et al., 2017). Moreover, in aged mice, spontaneous Ca2+ activity in astrocytes and the infarct area were higher than those in adult mice after ischemic stroke, while spontaneous Ca2+ activity in neurons was unchanged (Figure 5A) (Fordsmann et al., 2019; Murmu et al., 2019). These studies showed that, after experimental ischemic stroke, [Ca2+]i increased in astrocytes, which may aggravate neuronal excitotoxicity through glia-dependent glutamate release (Ding et al., 2009; Dong et al., 2013; Rakers and Petzold, 2017; Rakers et al., 2017; Fordsmann et al., 2019; Murmu et al., 2019). Calcium waves in astrocytes and peri-infarct depolarizations are shown in Figure 5B. Shinotsuka et al. (2014) used acute cortical slices under OGD and showed differing results regarding the calcium wave in astrocytes. They found that the astrocytic gap junction network acted as a buffer for intercellular calcium fluctuations in neurons during the acute phase of ischemia. After blocking gap junctions between astrocytes in mouse cortical slices under OGD, the SD occurred earlier (Shinotsuka et al., 2014), showing a protective effect of astrocyte networks on neurons. However, in ex vivo studies, the severity of ischemia and hypoxia in brain tissue may differ from that in in vivo studies, affecting the roles of astrocytes after ischemia.
Figure 5.
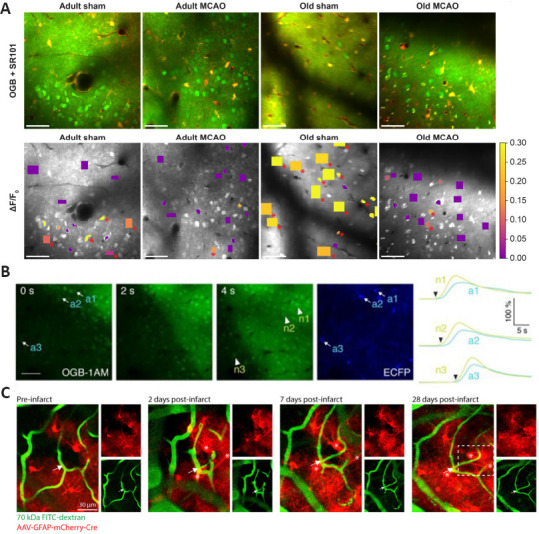
Images from 2PLSM observation of astrocytes.
(A) Representative in vivo 2PLSM images recorded in adult and aged mice after sham operation or permanent middle cerebral artery occlusion (MCAO). In the top line, green indicates Oregon Green 488 BAPTA-1/AM (OGB), red indicates sulforhodamine 101 (SR101). In the lower line, OGB is shown in gray. Mean ΔF/F0 during 2-Hz whisker stimulation is color-coded for neuronal somas (n.s.) and neuropil (n.p.). Red asterisks (*) indicate responsive regions of interest (ROIs). Scale bars: 50 μm. (B) Calcium images in astrocytes in Cx43-ECFP+/– mice. Peri-infarct depolarizations were identified as propagating calcium waves in astrocytes (a1–a3, identified by ECFP expression) and neurons (n1–n3). Scale bar: 50 μm. (C) Two-photon images showing reactive astrocytes facilitating vascular repair and remodeling after PT. Green indicates vessels and red indicates astrocytes. These images show a new capillary segment (arrow) formed after stroke contacted by astrocytes (*). 2PLSM: Two photon-excited fluorescence laser-scanning microscopy. A is reprinted from Murmu et al., 2019. B is reprinted from Rakers et al. (2017) with permission. C is reprinted from Williamson et al. (2021).
After quantifying the contacts between astrocytes and vessels in vivo using 2PLSM, almost all cortical astrocytes were in direct contact with blood vessels, which provided the structural basis for synapse-astrocyte-vessel communication (Hösli et al., 2022). Astrocytes integrate signals from synapses and vessels and are involved in the functioning of the neurovascular unit, which has been widely recognized in healthy live brains (Zonta et al., 2003; Perea et al., 2009; Bazargani and Attwell, 2016). 2PLSM can simultaneously observe the [Ca2+]i fluctuations in astrocytes and vessels diameters and selectively release Ca2+ from astrocytes using photolysis of the Ca2+ cage DMNP-EDTA. Studies in normal live brain slices showed that elevated [Ca2+]i in astrocytes could regulate the diameter of arterioles adjacent to the endfeet to modulate blood flow for adaptation to neuronal activity, mainly through the phospholipase A2-arachidonic acid pathway, which produces metabolites prostaglandin and epoxyeicosatrienoic acid for vasodilation and 20-hydroxyeicosatetraenoic acid for vasoconstriction (Zonta et al., 2003; Mulligan and MacVicar, 2004; Metea and Newman, 2006; Gordon et al., 2008; Rosenegger et al., 2015). However, other in vivo photolysis experiments using caged Ca2+ in astrocytic endfeet adjacent to cortical penetrating arterioles showed that the increase of [Ca2+]i in astrocyte endfeet was related to vasodilation via release of prostaglandin messengers to ensure basal blood flow in the brain (Takano et al., 2006). Whether the elevated [Ca2+]i in astrocytes causes vasodilation or constriction may be related to the pre-existing tone of the vessel and the astrocyte microenvironment (Gordon et al., 2008; Attwell et al., 2010; Rosenegger et al., 2015). Thus, the specific role of astrocytes in the regulation of the neurovascular unit during the acute phase of ischemic stroke needs to be better determined in vivo rather than ex vivo; intravital experiments are thought to more precisely show communication between astrocytes, neurons, and mural cells (Grubb et al., 2021). Chuquet et al. (2007) imaged the neurovascular unit in vivo during KCl and cardiac arrest-induced SD and found that the hyperacute vasoconstriction of arterioles that occurred after SD was associated with fast [Ca2+]i waves in astrocytes, mediated by a phospholipase A2 derivative. This vasoconstriction of arterioles mediated the hyperemic phase and might have contributed to endogenous compensation after ischemic stroke (Chuquet et al., 2007). However, this study partially ignored the contribution of mural cells after ischemic stroke, which construct the walls of blood vessels and are thought to be the crucial components for regulating vasomotor tone (Grubb et al., 2021). Therefore, labeling mural cells and calcium signaling in mural cells as well as clarifying the sequential order of the communication between astrocytes and mural cells and responses to ischemia are essential. Moreover, it is necessary to differentiate the vascular hierarchy, as the structures in the arterioles and capillaries are different (Grubb et al., 2020). Mishra et al. (2016), using in vivo 2PLSM imaging, showed that the role of astrocytes in the regulation of hemodynamics was related to the vascular level; astrocytes seemed to regulate capillaries but not arterioles. They applied calcium coupling agents and several receptor inhibitors and simultaneous imaging of astrocyte calcium signals and vessel diameters both in acute brain slices and in vivo (Mishra et al., 2016). The increased [Ca2+]i in astrocytes mediated capillary dilation through ATP-gated channels P2X purinoceptor 1, but the increased [Ca2+]i in astrocytes did not mediate arteriole dilation (Mishra et al., 2016). However, there is a lack of studies related to astrocyte-vessel communication and the roles of astrocytes in regulating hemodynamics in experimental ischemic stroke.
Moreover, astrocytes are the essential component of the BBB (Heithoff et al., 2021). However, Tóth et al. (2019) found no changes in astrocyte-vessel connections and no plasma extravasation over two hours of time-lapse 2PLSM imaging in PT and bilateral carotid ligation. Only photochemical embolization, a direct vascular injury, resulted in fluid extravasation (Tóth et al., 2019). This surprising result suggested that morphological changes in astrocytes in response to injury may be much slower than previously thought. However, several previous studies have shown that AQP4, a protein densely expressed on astrocytic endfeet, was associated with stroke-related vascular disruption (Manley et al., 2004; Friedman et al., 2009; Akdemir et al., 2014; Yao et al., 2015). Researchers found that AQP4–/– transgenic mice exhibited lower brain parenchymal water content and better neurological function, and animals with a normal AQP4 phenotype showed reduced AQP4 expression after ischemic stroke (Manley et al., 2004; Friedman et al., 2009; Akdemir et al., 2014; Yao et al., 2015).
The glymphatic system
An exciting discovery from 2PLSM-based research in astrocytes is the glymphatic system. Iliff et al. (2012) first described the glymphatic system as “a sewage system in the brain” that promotes cerebrospinal fluid (CSF) flow through the brain parenchyma and removes interstitial solutes. They demonstrated that the glymphatic system consists of peri-arterial CSF inflow, CSF mixed with interstitial fluid, and a mixture of CSF and interstitial fluid flowing out through the peri-venous space and leaving the brain along the cranial and spinal nerves (Iliff et al., 2012). CSF mixing with interstitial fluid is mediated by the AQP4 protein densely expressed on astrocytic endfeet (Iliff et al., 2012). This sewage system in the brain has never been represented in postmortem animal tissue sections, which inevitably have damage to the CSF flow pathway due to fixation (Benias et al., 2018; Mestre et al., 2018). In contrast, in vivo 2PLSM imaging provides high-resolution dynamic evidence of fluid flow. Iliff et al. (2012) used in vivo 2PLSM to record the movement of fluorescent tracers of different molecular weights injected into the ventricles. In Aqp4 gene knockout mice, the medium molecular-weight tracer could flow into the perivascular space while the movement from the perivascular space into the brain parenchyma was blocked (Iliff et al., 2012). The glymphatic system could perform clearance functions, including clearing soluble amyloid β, a key protein in Alzheimer’s disease (Iliff et al., 2012). Xie et al. (2013) further explored 2PLSM imaging of tetramethylammonium diffusion in awake, anesthetized, and sleeping mice and found that neither natural sleep nor anesthesia was associated with increased interstitial space, leading to a significant increase in convective exchange between CSF and interstitial fluid, suggesting that sleep drives metabolite clearance in the adult brain. As for studying ischemic stroke, they used in vivo 2PLSM and magnetic resonance imaging and demonstrated that CSF influx is an essential source of edema fluid after ischemic stroke (Mestre et al., 2020b). The influx of CSF with high sodium levels could further drive brain tissue swelling. Thus, they pioneered the term ionic edema to describe the intermediary brain edema phase between early cytotoxic and late vasogenic brain edema stages after ischemic stroke. They used genetically encoded calcium indicators to visualize the depolarization of neurons (Mestre et al., 2020b). They demonstrated that there was a temporal link between SD and CSF transport through the perivascular spaces to the brain parenchyma after ischemic stroke, indicating that SD likely mediated CSF transport to brain parenchyma through the glymphatic system (Mestre et al., 2020b). This emphasized the crucial role of the astrocyte network in post-stroke edema. Targeting AQP4 to treat brain edema after ischemic stroke may be a possible treatment option.
However, several studies have challenged the opinion that the glymphatic system mediates solute clearance from brain parenchyma through convective flow (Hladky and Barrand, 2014; Spector et al., 2015; Holter et al., 2017; Smith et al., 2017). Holter et al. (2017) used 3D electron microscopy to reconstruct interstitial solute transport in hippocampal tissue. Their results suggested that solutes were more likely to be transported through interstitial spaces by diffusion rather than by convective flow. Smith et al. (2017) used 2PLSM to photobleach dextran in the parenchymal extracellular space and to image the fluorescence recovery process. They found that fluorescent dextran transport in brain parenchyma depended on the size of the molecule, was mainly diffusive rather than convective transport, and was unaffected after cardiac arrest (Smith et al., 2017). Further imaging in AQP4-deficient animals revealed that AQP4 deletion did not impair the transport of fluorescent solutes from the subarachnoid space to the brain parenchyma (Smith et al., 2017). Mestre et al. (2020a) published a paper, summarizing and responding to the controversies around the glymphatic system. They argued that mathematical models ignored the irregularities of the perivascular space and that AQP4 knockout mice should show a high degree of age uniformity, as the activity of the glymphatic system declines rapidly with aging. However, the specific role and mechanisms of astrocytic AQP4 in mediating the influx of CSF into the brain parenchyma in ischemic stroke remains to be investigated (Spector et al., 2015; Mestre et al., 2020a).
Roles of astrocytes in recovery phase of ischemic stroke
The roles of astrocytes in the recovery phase of ischemic stroke are also complex. Based on postmortem techniques, Li et al. (2015) found that astrocyte IP3R2 knockout mice showed attenuated excessive astrogliosis and relieved brain injury, neuronal death, and behavioral deficits compared to controls 14 days after PT. This indicated that the calcium signal pathway in astrocytes may be harmful to neural recovery after ischemic stroke. However, other studies using dynamic in vivo 2PLSM showed that astrocytes played an essential role in angiogenesis, vascular network reorganization, and neural recovery. Heras-Romero et al. (2022) administered extracellular vesicles released by primary cortical astrocytes in the ventricles of mice and continuously observed the dynamic recovery process after ischemic stroke for 21 days. They found that the extracellular vesicles could mediate recovery of structure and function in neurons. Williamson et al. (2021) traced the morphology of astrocytes and the relationship between astrocytes and angiogenesis after PT for 28 days. They demonstrated that reactive astrocytes colocalized with and contacted newly formed vessels (Figure 5C). Chemogenetic ablation of peri-infarct reactive astrocytes dramatically impaired vascular remodeling and impeded the recovery of neurological function. Göbel et al. (2020) explored the specific mechanisms of astrocytes in vessel remodeling. They observed a prominent mitochondria-enriched compartment in astrocytic endfeet and mediation of vascular remodeling after stab-wound injury. However, this finding requires further study in ischemic stroke models.
The contradictory results regarding astrocytes in the recovery of ischemic stroke may be related to the differentiation of diverse subpopulations of astrocytes after injury; different subpopulations of astrocytes may perform different functions. Gene knockouts in all astrocytes without differentiating between subpopulations may obscure experimental results. In 2013, Bardehle et al. used time-lapse 2PLSM to observe dynamic responses of astrocytes to acute traumatic brain injury in live mice for up to 28 days. They found significant heterogeneity in the responses of astrocytes to injury, with only a distinct subset located at juxtavascular sites proliferating, one subset retaining their initial morphology, and another directing their processes toward the lesion (Bardehle et al., 2013). However, there is a lack of research on the dynamics of astrocyte behaviors in ischemic stroke. Thus, subtle subpopulation studies of astrocytes may be more practical for the study of ischemic stroke.
The Response of Microglia in the Acute Phase of Ischemic Stroke
Microglia, the critical resident immune cells in the brain, are dedicated to neuronal network homeostasis and function (Pena-Ortega, 2017). Microglia transform into activated microglia after ischemic stroke, which is a process marked by changes from ramified to amoeboid proliferative and morphological features, and activated microglia exhibit enhanced phagocytosis and release cytokines and growth factors (Otxoa-de-Amezaga et al., 2019; Qin et al., 2019; Song et al., 2019; Ronaldson and Davis, 2020). Complex functions and multiple phenotypes of reactive microglia have been widely demonstrated using single-cell sequencing and transcriptomics (Ransohoff, 2016a; Li et al., 2019a; Qin et al., 2019; Sierra et al., 2019; Stratoulias et al., 2019; Jurga et al., 2020). However, the specific roles of microglia in ischemic stroke have not been well studied. Microglia are highly dynamic and frequently communicate with neurons and vessels, meaning that postmortem animal studies inevitably miss a great deal of data on dynamic changes, and neuroimaging has limited spatiotemporal resolution. Furthermore, it is inevitable that different microglial activation states will occur during ex vivo preparations due to their high sensitivity. Therefore, with advanced fluorescent labeling techniques (Table 2), in vivo time-lapse 2PLSM is an excellent tool for studying microglial networks, especially in the immediate response after ischemic stroke.
Changes in microglia behaviors after ischemic stroke
The transformation from ramified into amoeboid morphological features is an essential indicator of microglial activation (Kreutzberg, 1996; Zhang, 2019). Using 2PLSM, Nimmerjahn et al. (2005) and Davalos et al. (2005) found that, even in the resting state, microglial processes were remarkably motile, likely to monitor surroundings and interact with neighboring cells (Davalos et al., 2005; Nimmerjahn et al., 2005). After external stimulation such as laser ablation or the application of ATP, microglia rapidly converged to the lesion site, even without complete activation (Hanisch and Kettenmann, 2007). This finding demonstrated that microglia are highly dynamic and play an important role in brain microenvironment homeostasis in both healthy and diseased brain tissue.
Wake et al. (2009) found that, in the normal resting state, microglial processes change dynamically and contact neuronal synapses at a specific frequency and contact duration. After transient cerebral ischemia, the contact time between microglia and synapses was prolonged, corresponding to the disappearance of the presynaptic bouton (Wake et al., 2009). Li et al. (2013) used a parabiosis model of CX3CR1-GFPand wild-type mice to demonstrate that proliferating microglia in the brain parenchyma after ischemic stroke were derived from resident microglia rather than circulating monocytes or myeloid progenitor cells. Masuda et al. (2011) demonstrated that the activation of microglia in the ischemic penumbra was associated with a decrease in blood flow, and that this activation was energy-dependent and required residual blood flow. However, further studies on the pathophysiological impacts of activated microglia after ischemic stroke have been controversial. Several in vivo 2PLSM studies further showed that blocking important receptors expressed on microglia, including chemokine receptors, colony-stimulating factor receptors, and complement receptors (CX3CR1, C3aR, CSF1R, and CXCR3), could have neuroprotective effects (Fumagalli et al., 2013; Walter et al., 2015; Surugiu et al., 2019; Hou et al., 2020). Other studies have shown in vivo that selective elimination of microglia increased infarct size; exogenously applied BV2 microglial cell lines reduced neural damage in organotypic hippocampal slice cultures, indicating that microglia are protective (Neumann et al., 2006; Szalay et al., 2016; Zhang, 2019).
In addition to the infarct core and ischemic penumbra, secondary neurodegeneration (SND) is also an important research focus. SND involves the progressive death of neurons associated with the site of infarction but not initially damaged during the ischemic stroke (Jayaraj et al., 2019; Stuckey et al., 2021). Researchers have used acute slices from the thalamus, a major site of SND, to observe morphological changes in microglia (Kluge et al., 2017, 2019). In these 2PLSM studies, they found that microglial specificity became nonresponsive in SND, as evidenced by weakened microglial process movements (Kluge et al., 2017, 2019). In contrast, phagocytosis and the levels of classic microglial molecular markers remained elevated. The appearance of nonresponsive microglia was strongly associated with neuronal damage.
Microglia communication with multiple cell types after ischemic stroke
As a kind of non-excitable cell, microglia highly depend on changes in intracellular [Ca2+]i to perform cellular functions (Heo et al., 2015). Using in vivo two-photon calcium imaging, Eichhoff et al. (2011) found that most (80%) microglia showed no spontaneous Ca2+ transients at rest or under conditions of strong neuronal activity or intercellular astrocytic Ca2+ waves. However, microglia reliably responded with large, generalized Ca2+ transients to damage individual neurons (Eichhoff et al., 2011). This suggested that changes in microglial calcium signaling are primarily involved in pathological processes in response to neuronal injury, contrary to the results of in vitro experiments. Using in vivo 2PLSM, they found frequent Ca2+ transients in microglia after triggering cortical SD, which was induced by applying exogenous KCl solution (Tvrdik et al., 2019; Kearns et al., 2020, 2022; Liu et al., 2021). This linked neuronal damage, neuronal electrical signals, and microglial functional. A calcium wave in cortical microglia during ischemic stroke is shown in Figure 6B. This study also provided a reliable paradigm for studying the relationship between microglia and neuronal networks after ischemic stroke.
Figure 6.
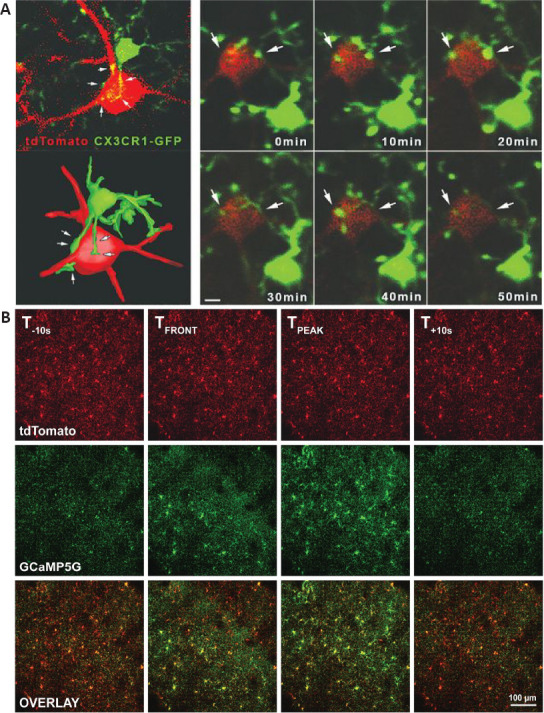
Images of in vivo 2PLSM of microglia.
(A) The left row shows a single imaging plane (upper panel) and 3D reconstruction (lower panel) from an in vivo two-photon Z-stack showing a neocortical neuron (red) being contacted by microglial processes (green). The right panel shows in vivo 2PLSM time-lapse imaging showing temporal dynamics of microglia–neuron contacts. (B) A traveling calcium wave in cortical microglia during ischemic stroke. Individual frames were extracted from a high-speed two-photon microscopy recording. tdTomato and GCaMP5G signals are shown separately and in overlay. GCaMP is composed of permuted green fluorescent protein (cpGFP), calmodulin (CaM), and the Ca2+/CaM-binding “M13” peptide (M13pep). Intracellular calcium concentration fluctuations are shown by the fluorescence intensity. Tfront and Tpeak indicate the emergence and peak intensity of the calcium wave, respectively. The flanking panels (T−10s and T+10s) show the frames recorded 10 seconds before and after these events. Scale bar: 100 μm. 2PLSM: Two photon-excited fluorescence laser-scanning microscopy. A and B are reprinted from Cserép et al. (2020) and Tvrdik et al. (2019) with permission, respectively.
Moreover, Cserép et al. (2020) used post hoc confocal laser scanning microscopy and electron microscopy to further illustrate the highly dynamic ultrastructure of microglia-neuron junctions in healthy brains, which provided valid structural evidence for the novel microglia-neuron junction concept (Figure 6A). They demonstrated that the microglia-neuron junctions consisted of closely apposed mitochondria, reticular membrane structures, intracellular tethers, and associated vesicle-like membrane structures within the neuronal cell body (Cserép et al., 2020). They used mitochondrial fluorescent labeling and in vivo 2PLSM to demonstrate that the foundation of microglia-neuron junctions relied on mitochondria-dependent neuronal exocytosis release signals (Cserép et al., 2020). This signaling process was highly correlated with ATP and ADP, which are important ligands for the regulation of microglial processes via the microglial purinoceptor P2Y12 (Cserép et al., 2020). They proposed that healthy neurons may constitutively release ATP and other signaling molecules at the microglia-neuron junction and that P2Y12 receptors on microglia receive the relevant signals, thus allowing microglia to monitor neuronal status (Cserép et al., 2020). Blocking microglial P2Y12 receptors after MCAO led to a strong increase in neuronal calcium load and to a significantly larger lesion volume than those observed in control mice, which may indicate that microglia play a neuroprotective post-stroke role through microglia-neuron junctions (Cserép et al., 2020).
Microglia also interact with blood vessels. Bisht et al. (2021) used in vivo 2PLSM to observe fluorescently labelled pericapillary cells in CX3CR1-GFPtransgenic mice. They confirmed that the pericapillary CX3CR1-GFPcells were capillary-associated microglia using molecular, morphological, and ultrastructural approaches. After elimination of perivascular microglia using CSF1R inhibitors, capillary diameters increased and cerebral blood flow measured by laser speckle imaging increased, while vascular responsiveness to CO2 decreased. This demonstrated that, in the healthy brain, microglia communicate with blood vessels and play an important role in regulating vascular structure and function (Bisht et al., 2021). After ischemic stroke, changes in microglia-vessel communication also affects prognosis. Using microglia-labeled transgenic mice (CX3CR1-GFP), Jolivel et al. (2015) co-imaged microglia and blood vessels. Soon after reperfusion in transient MCAO, the microglia in the penumbra were activated and began to expand towards adjacent blood vessels, phagocytize vascular endothelial cells, and mediate injury to the BBB.
The role of interactions between different types of glia in ischemic stroke has also received a lot of attention (Garcia-Caceres et al., 2019; Liu et al., 2020; Matejuk and Ransohoff, 2020). However, regarding interactions between glial cells, current two-photon studies lack data on communication and cooperation between different types of glia and their specific roles in the network structure after ischemic stroke.
Limitations and Innovations
Although the high spatial and temporal resolution for in vivo studies using 2PLSM are remarkable for the longitudinal study of ischemic stroke, 2PLSM is far from perfect. With its development, many limitations have been already resolved. First, before imaging, skull window surgery is required. For an open-skull glass window, damage to brain tissue and associated inflammatory reactions are inevitable (Xu et al., 2007; Marker et al., 2010; Yang et al., 2010). Researchers also need long training to complete the surgery. Thinned cranial windows limit damage to the brain, while performing relative long-term observation requires repeated thinning of the skull due to bone regrowth (Yang et al., 2010). Zhao et al. (2018) have developed a technique for safe clearing of a skull window, which resulted in in vivo 2PLSM imaging of cortical structures at synaptic resolution without removing the skull or excessive thinning of the skull, minimizing damage to brain tissue. Second, the maximum imaging depth of 2PLSM is limited, making whole-brain in vivo live imaging not possible. Changes in neural circuits after stroke are important for neuronal plasticity and functional compensation (Fox, 2018; Herbet and Duffau, 2020). Stereotactic injection of tracer viruses offers a possible solution for in vivo visualization of neural circuits (Chatterjee et al., 2018; Zhang et al., 2018; Macknik et al., 2019; Poinsatte et al., 2019; De La Crompe et al., 2020; Tang et al., 2020). Third, the achievement of multicolor 2PLSM imaging is also a crucial problem in studies targeting intercellular communication. The emission spectra of different fluorescent probes may overlap, generating spectral crosstalk and making it hard to clearly distinguish the sources of emitted light, which limits imaging of neurovascular units. To achieve multicolor 2PLSM imaging, multiple pairs of lasers and detectors or filters and detectors are used to separate different fluorescence signals, but this increases the cost (Le Grand et al., 2008). Several post-processing techniques and the combination of 2PLSM with other optical imaging principles can be used to solve this problem (Ueki et al., 2020; Olson et al., 2022; Yan et al., 2022). Moreover, novel quantum dots have been used as fluorescent labels in 2PLSM, and their optical properties are suitable for multicolor imaging (Mashinchian et al., 2014; He et al., 2018; Qi et al., 2018; Wang et al., 2019; Qin et al., 2020). Fourth, for highly specific fluorescent labeling of certain components, such as microglia, the prior methodology for cell visualization uses fluorescent reporters, requiring cross-breeding of genetic lines of interest. This requires a fairly long breeding cycle. Moreover, commonly used 2PLSM devices need heavy shock-resistant benchwork that is too bulky to miniaturize. For ischemic stroke studies, the common imaging strategy is head fixation after anesthesia, and the results may not reflect the actual cellular function in the natural state (Groothuis et al., 2007; Cao et al., 2017; Guo et al., 2021; Bharioke et al., 2022). Therefore, it is important to have a two-photon imaging platform that can assess post-stroke pathophysiological processes in awake, behaving animals. Several studies have used 2PLSM to image awake animals (Stobart et al., 2018; Yang et al., 2018; Piatkevich et al., 2019; Chow et al., 2020). Zong et al. (2021) developed a miniature two-photon microscope called FHIRM-TPM 2.0, which can reflect the activity characteristics of neurons in mice in a natural state rather than with fixation or under anesthesia. Finally, imaging data obtained from 2PLSM are extensive, and image processing is highly complex. Researchers have introduced artificial intelligence to assist with data processing (Barbastathis et al., 2019; Borhani et al., 2019; Gur et al., 2020). However, the robustness and interpretability of results obtained using artificial intelligence need to be carefully evaluated.
The development of two-photon fluorescent probes is also continuing to progress. Novel, specific organelle or subcellular component-localizable, two-photon fluorescent probes are now under development, which would allow researchers to explore biological molecular-level changes in important subcellular structures, such as mitochondria, lysosomes, nuclei, the Golgi apparatus, and the endoplasmic reticulum (Collot et al., 2018; Dana et al., 2019; Ye et al., 2019; Zhao et al., 2020; Cheng et al., 2021). A novel multiplexed dynamic intravital multiphoton imaging method allowed researchers to distinguish seven fluorophore signals corresponding to various cellular and tissue compartments using four detector channels (Rakhymzhan et al., 2021). Label-free imaging is also a good alternative. Wu et al. (2020) were able to receive autofluorescence signals with short wavelength two-photon excitation. They developed a time-resolved two-photon excitation microscopy system using a homemade 520-nm femtosecond fiber laser as the excitation source, which could noninvasively achieve intravital high-resolution 3D imaging of a microvascular network.
Conclusions
After discussing and summarizing studies of ischemic stroke using 2PLSM over the past decades, we found that the pathophysiological processes after ischemic stroke cannot be fully explored by only observing changes in neurons after stroke without considering the neural support system represented mainly by blood vessels, astrocytes, and microglia. Moreover, when studying the roles of blood vessels and glia, it needs to be fully recognized that they constitute a large and sophisticated spatiotemporal network whose structural hierarchy, behavior, and function change dynamically with the environment and over time and have important implications for the fate of neurons after ischemic stroke. The same cellular component at different temporal and spatial nodes may have different functions. However, we can easily find that the specific roles of glia and blood vessels in ischemic stroke are still controversial. We believe that this phenomenon is partly due to different responses of different subgroups of glial cells and different hierarchies in vascular networks in different environments and that knockdown or overexpression certain genes, as well as imaging only a portion of cells in the neurovascular unit for a certain period of time, are likely to represent only a few aspects of cellular functions. This problem is partially ignored in a majority of studies but is gradually being recognized (Schaeffer and Iadecola, 2021).
Despite the limitations of imaging depth, 2PLSM is invaluable for recording both structural and functional information of multiple cells and cell types simultaneously with subcellular resolution in live brains. Although 2PLSM provides solid and reliable in vivo evidence and reflects subtle in vivo processes that cannot be obtained from postmortem animals, results based on 2PLSM still vary between studies. The use of in vivo animal models or live brain slices, the age of the animal, the method of astrocyte labeling, and modeling methods can bias results away from the natural disease state. Moreover, the commonly used in vivo experimental ischemic stroke models (i.e., MCAO and PT) are far from reality, which is an obstacle in clinical translation (Sommer, 2017). Imaging evidence provided by 2PLSM is at the cellular or subcellular level. It is a challenge to connect the high-quality cellular-level in vivo evidence provided by 2PLSM with the molecular-level evidence provided by biochemical techniques. Relying on specific ablation of cells or gene knockouts or upregulation may not perfectly merge the advantages of both. The emerging spatial transcriptomics technique gives a possibility to fuse the two types of technologies, but the tedious data analysis and interpretation processes are not easily applicable.
Overall, future research will shift from postmortem animal research to live animal research, from static research to dynamic research, from flat to three-dimensional, and from single-cell function and structure to intercellular communication. 2PLSM is a robust tool to translate study discovery from bench to bedside.
Acknowledgments:
We are very grateful to Yunyun Han from Huazhong University of Science and Technology for the helpful comments on the manuscript, and we also thank her valuable suggestions on the content of the article.
Footnotes
Author contributions: Manuscript design and conception: XW and JRL; manuscript writing: XW; literature search: YF. Figure and table design and preparation: DYC. Language polishing, manuscript review and editing: HN and ZPT. All authors read and approved the final version of the manuscript.
Conflicts of interest: The authors declare no conflicts of interest.
Data availability statement: No additional data are available.
C-Editor: Zhao M; S-Editor: Li CH; S-Editors: Li CH, Song LP; T-Editor: Jia Y
References
- 1.Adhikari DP, Biener G, Stoneman MR, Badu DN, Paprocki JD, Eis A, Park PS, Popa I, Raicu V. Comparative photophysical properties of some widely used fluorescent proteins under two-photon excitation conditions. Spectrochim Acta A Mol Biomol Spectrosc. 2021;262:120133. doi: 10.1016/j.saa.2021.120133. [DOI] [PubMed] [Google Scholar]
- 2.Ahishali B, Kaya M. Evaluation of blood-brain barrier integrity using vascular permeability markers: Evans blue, sodium fluorescein, albumin-alexa fluor conjugates, and horseradish peroxidase. Methods Mol Biol. 2021;2367:87–103. doi: 10.1007/7651_2020_316. [DOI] [PubMed] [Google Scholar]
- 3.Akdemir G, Ratelade J, Asavapanumas N, Verkman AS. Neuroprotective effect of aquaporin-4 deficiency in a mouse model of severe global cerebral ischemia produced by transient 4-vessel occlusion. Neurosci Lett. 2014;574:70–75. doi: 10.1016/j.neulet.2014.03.073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Åkerblom M, Sachdeva R, Quintino L, Wettergren EE, Chapman KZ, Manfre G, Lindvall O, Lundberg C, Jakobsson J. Visualization and genetic modification of resident brain microglia using lentiviral vectors regulated by microRNA-9. Nat Commun. 2013;4:1770. doi: 10.1038/ncomms2801. [DOI] [PubMed] [Google Scholar]
- 5.Alarcon-Martinez L, Villafranca-Baughman D, Quintero H, Kacerovsky JB, Dotigny F, Murai KK, Prat A, Drapeau P, Di Polo A. Interpericyte tunnelling nanotubes regulate neurovascular coupling. Nature. 2020;585:91–95. doi: 10.1038/s41586-020-2589-x. [DOI] [PubMed] [Google Scholar]
- 6.Allen NJ, Lyons DA. Glia as architects of central nervous system formation and function. Science. 2018;362:181–185. doi: 10.1126/science.aat0473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Appaix F, Girod S, Boisseau S, Römer J, Vial JC, Albrieux M, Maurin M, Depaulis A, Guillemain I, van der Sanden B. Specific in vivo staining of astrocytes in the whole brain after intravenous injection of sulforhodamine dyes. PLoS One. 2012;7:e35169. doi: 10.1371/journal.pone.0035169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Armbruster M, Naskar S, Garcia JP, Sommer M, Kim E, Adam Y, Haydon PG, Boyden ES, Cohen AE, Dulla CG. Neuronal activity drives pathway-specific depolarization of peripheral astrocyte processes. Nat Neurosci. 2022;25:607–616. doi: 10.1038/s41593-022-01049-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Armstrong JJ, Larina IV, Dickinson ME, Zimmer WE, Hirschi KK. Characterization of bacterial artificial chromosome transgenic mice expressing mCherry fluorescent protein substituted for the murine smooth muscle alpha-actin gene. Genesis. 2010;48:457–463. doi: 10.1002/dvg.20638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Attwell D, Buchan AM, Charpak S, Lauritzen M, Macvicar BA, Newman EA. Glial and neuronal control of brain blood flow. Nature. 2010;468:232–243. doi: 10.1038/nature09613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Bai J, Lyden PD. Revisiting cerebral postischemic reperfusion injury: new insights in understanding reperfusion failure, hemorrhage, and edema. Int J Stroke. 2015;10:143–152. doi: 10.1111/ijs.12434. [DOI] [PubMed] [Google Scholar]
- 12.Bang OY, Kim EH. Mesenchymal stem cell-derived extracellular vesicle therapy for stroke: challenges and progress. Front Neurol. 2019;10:211. doi: 10.3389/fneur.2019.00211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Barbastathis G, Ozcan A, Situ G. On the use of deep learning for computational imaging. Optica. 2019;6:921–943. [Google Scholar]
- 14.Bardehle S, Krüger M, Buggenthin F, Schwausch J, Ninkovic J, Clevers H, Snippert HJ, Theis FJ, Meyer-Luehmann M, Bechmann I, Dimou L, Götz M. Live imaging of astrocyte responses to acute injury reveals selective juxtavascular proliferation. Nat Neurosci. 2013;16:580–586. doi: 10.1038/nn.3371. [DOI] [PubMed] [Google Scholar]
- 15.Bazargani N, Attwell D. Astrocyte calcium signaling: the third wave. Nat Neurosci. 2016;19:182–189. doi: 10.1038/nn.4201. [DOI] [PubMed] [Google Scholar]
- 16.Benias PC, Wells RG, Sackey-Aboagye B, Klavan H, Reidy J, Buonocore D, Miranda M, Kornacki S, Wayne M, Carr-Locke DL, Theise ND. Structure and distribution of an unrecognized interstitium in human tissues. Sci Rep. 2018;8:4947. doi: 10.1038/s41598-018-23062-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Benninger RK, Piston DW. Two-photon excitation microscopy for the study of living cells and tissues. Curr Protoc Cell Biol Chapter 4. 2013 doi: 10.1002/0471143030.cb0411s59. Unit 4.11.11-24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Berge E, Whiteley W, Audebert H, De Marchis GM, Fonseca AC, Padiglioni C, de la Ossa NP, Strbian D, Tsivgoulis G, Turc G. European Stroke Organisation (ESO) guidelines on intravenous thrombolysis for acute ischaemic stroke. Eur Stroke J. 2021;6:I–LXII. doi: 10.1177/2396987321989865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bernardo-Castro S, Sousa JA, Brás A, Cecília C, Rodrigues B, Almendra L, Machado C, Santo G, Silva F, Ferreira L, Santana I, Sargento-Freitas J. Pathophysiology of blood-brain barrier permeability throughout the different stages of ischemic stroke and its implication on hemorrhagic transformation and recovery. Front Neurol. 2020;11:594672. doi: 10.3389/fneur.2020.594672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Berthiaume AA, Hartmann DA, Majesky MW, Bhat NR, Shih AY. Pericyte structural remodeling in cerebrovascular health and homeostasis. Front Aging Neurosci. 2018a;10:210. doi: 10.3389/fnagi.2018.00210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Berthiaume AA, Grant RI, McDowell KP, Underly RG, Hartmann DA, Levy M, Bhat NR, Shih AY. Dynamic remodeling of pericytes in vivo maintains capillary coverage in the adult mouse brain. Cell Rep. 2018b;22:8–16. doi: 10.1016/j.celrep.2017.12.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Bharioke A, Munz M, Brignall A, Kosche G, Eizinger MF, Ledergerber N, Hillier D, Gross-Scherf B, Conzelmann KK, Macé E, Roska B. General anesthesia globally synchronizes activity selectively in layer 5 cortical pyramidal neurons. Neuron. 2022;110:2024–2040.e10. doi: 10.1016/j.neuron.2022.03.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Bhaskar S, Stanwell P, Cordato D, Attia J, Levi C. Reperfusion therapy in acute ischemic stroke: dawn of a new era? BMC Neurol. 2018;18:8. doi: 10.1186/s12883-017-1007-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bindocci E, Savtchouk I, Liaudet N, Becker D, Carriero G, Volterra A. Three-dimensional Ca(2+) imaging advances understanding of astrocyte biology. Science. 2017;356:eaai8185. doi: 10.1126/science.aai8185. [DOI] [PubMed] [Google Scholar]
- 25.Bisht K, Okojie KA, Sharma K, Lentferink DH, Sun YY, Chen HR, Uweru JO, Amancherla S, Calcuttawala Z, Campos-Salazar AB, Corliss B, Jabbour L, Benderoth J, Friestad B, Mills WA, 3rd, Isakson BE, Tremblay M, Kuan CY, Eyo UB. Capillary-associated microglia regulate vascular structure and function through PANX1-P2RY12 coupling in mice. Nat Commun. 2021;12:5289. doi: 10.1038/s41467-021-25590-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Bok S, Wang T, Lee CJ, Jeon SU, Kim YE, Kim J, Hong BJ, Yoon CJ, Kim S, Lee SH, Kim HJ, Kim IH, Kim KH, Ahn GO. In vivo imaging of activated microglia in a mouse model of focal cerebral ischemia by two-photon microscopy. Biomed Opt Express. 2015;6:3303–3312. doi: 10.1364/BOE.6.003303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Borhani N, Bower AJ, Boppart SA, Psaltis D. Digital staining through the application of deep neural networks to multi-modal multi-photon microscopy. Biomed Opt Express. 2019;10:1339–1350. doi: 10.1364/BOE.10.001339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bragin DE, Peng Z, Bragina OA, Statom GL, Kameneva MV, Nemoto EM. Improvement of impaired cerebral microcirculation using rheological modulation by drag-reducing polymers. Adv Exp Med Biol. 2016;923:239–244. doi: 10.1007/978-3-319-38810-6_32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Brawek B, Garaschuk O. Monitoring in vivo function of cortical microglia. Cell Calcium. 2017;64:109–117. doi: 10.1016/j.ceca.2017.02.011. [DOI] [PubMed] [Google Scholar]
- 30.Brawek B, Liang Y, Savitska D, Li K, Fomin-Thunemann N, Kovalchuk Y, Zirdum E, Jakobsson J, Garaschuk O. A new approach for ratiometric in vivo calcium imaging of microglia. Sci Rep. 2017;7:6030. doi: 10.1038/s41598-017-05952-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Broocks G, Kniep H, Schramm P, Hanning U, Flottmann F, Faizy T, Schonfeld M, Meyer L, Schon G, Aulmann L, Machner B, Royl G, Fiehler J, Kemmling A. Patients with low Alberta Stroke Program Early CT Score (ASPECTS) but good collaterals benefit from endovascular recanalization. J Neurointerv Surg. 2020;12:747–752. doi: 10.1136/neurintsurg-2019-015308. [DOI] [PubMed] [Google Scholar]
- 32.Caballero-Garrido E, Pena-Philippides JC, Lordkipanidze T, Bragin D, Yang Y, Erhardt EB, Roitbak T. In vivo inhibition of miR-155 promotes recovery after experimental mouse stroke. J Neurosci. 2015;35:12446–12464. doi: 10.1523/JNEUROSCI.1641-15.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Cai W, Hu M, Li C, Wu R, Lu D, Xie C, Zhang W, Li T, Shen S, Huang H, Qiu W, Liu Q, Lu Y, Lu Z. FOXP3+macrophage represses acute ischemic stroke-induced neural inflammation. Autophagy. 2022:1–20. doi: 10.1080/15548627.2022.2116833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Cao R, Li J, Ning B, Sun N, Wang T, Zuo Z, Hu S. Functional and oxygen-metabolic photoacoustic microscopy of the awake mouse brain. Neuroimage. 2017;150:77–87. doi: 10.1016/j.neuroimage.2017.01.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Caporarello N, D'Angeli F, Cambria MT, Candido S, Giallongo C, Salmeri M, Lombardo C, Longo A, Giurdanella G, Anfuso CD, Lupo G. Pericytes in microvessels: from “mural” function to brain and retina regeneration. Int J Mol Sci. 2019;20:6351. doi: 10.3390/ijms20246351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Chai H, Diaz-Castro B, Shigetomi E, Monte E, Octeau JC, Yu X, Cohn W, Rajendran PS, Vondriska TM, Whitelegge JP, Coppola G, Khakh BS. Neural circuit-specialized astrocytes: transcriptomic, proteomic, morphological, and functional evidence. Neuron. 2017;95:531–549.e9. doi: 10.1016/j.neuron.2017.06.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Charles AC, Merrill JE, Dirksen ER, Sanderson MJ. Intercellular signaling in glial cells: calcium waves and oscillations in response to mechanical stimulation and glutamate. Neuron. 1991;6:983–992. doi: 10.1016/0896-6273(91)90238-u. [DOI] [PubMed] [Google Scholar]
- 38.Chatterjee S, Sullivan HA, MacLennan BJ, Xu R, Hou Y, Lavin TK, Lea NE, Michalski JE, Babcock KR, Dietrich S, Matthews GA, Beyeler A, Calhoon GG, Glober G, Whitesell JD, Yao S, Cetin A, Harris JA, Zeng H, Tye KM, et al. Nontoxic, double-deletion-mutant rabies viral vectors for retrograde targeting of projection neurons. Nat Neurosci. 2018;21:638–646. doi: 10.1038/s41593-018-0091-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Cheng F, Qiang T, Ren L, Liang T, Gao X, Wang B, Hu W. Observation of inflammation-induced mitophagy during stroke by a mitochondria-targeting two-photon ratiometric probe. Analyst. 2021;146:2632–2637. doi: 10.1039/d1an00208b. [DOI] [PubMed] [Google Scholar]
- 40.Cheng YQ, Wu CR, Du MR, Zhou Q, Wu BY, Fu JY, Balawi E, Tan WL, Liao ZB. CircLphn3 protects the blood-brain barrier in traumatic brain injury. Neural Regen Res. 2022;17:812–818. doi: 10.4103/1673-5374.322467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Chiang T, Messing RO, Chou WH. Mouse model of middle cerebral artery occlusion. J Vis Exp. 20112761 doi: 10.3791/2761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Chow BW, Nuñez V, Kaplan L, Granger AJ, Bistrong K, Zucker HL, Kumar P, Sabatini BL, Gu C. Caveolae in CNS arterioles mediate neurovascular coupling. Nature. 2020;579:106–110. doi: 10.1038/s41586-020-2026-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Chuquet J, Hollender L, Nimchinsky EA. High-resolution in vivo imaging of the neurovascular unit during spreading depression. J Neurosci. 2007;27:4036–4044. doi: 10.1523/JNEUROSCI.0721-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Collot M, Fam TK, Ashokkumar P, Faklaris O, Galli T, Danglot L, Klymchenko AS. Ultrabright and fluorogenic probes for multicolor imaging and tracking of lipid droplets in cells and tissues. J Am Chem Soc. 2018;140:5401–5411. doi: 10.1021/jacs.7b12817. [DOI] [PubMed] [Google Scholar]
- 45.Cornell-Bell AH, Finkbeiner SM, Cooper MS, Smith SJ. Glutamate induces calcium waves in cultured astrocytes: long-range glial signaling. Science. 1990;247:470–473. doi: 10.1126/science.1967852. [DOI] [PubMed] [Google Scholar]
- 46.Cotrina ML, Lou N, Tome-Garcia J, Goldman J, Nedergaard M. Direct comparison of microglial dynamics and inflammatory profile in photothrombotic and arterial occlusion evoked stroke. Neuroscience. 2017;343:483–494. doi: 10.1016/j.neuroscience.2016.12.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Cserép C, Pósfai B, Lénárt N, Fekete R, László ZI, Lele Z, Orsolits B, Molnár G, Heindl S, Schwarcz AD, Ujvári K, Környei Z, Tóth K, Szabadits E, Sperlágh B, Baranyi M, Csiba L, Hortobágyi T, Maglóczky Z, Martinecz B, et al. Microglia monitor and protect neuronal function through specialized somatic purinergic junctions. Science. 2020;367:528–537. doi: 10.1126/science.aax6752. [DOI] [PubMed] [Google Scholar]
- 48.Dailey ME, Eyo U, Fuller L, Hass J, Kurpius D. Imaging microglia in brain slices and slice cultures. Cold Spring Harb Protoc. 2013;2013:1142–1148. doi: 10.1101/pdb.prot079483. [DOI] [PubMed] [Google Scholar]
- 49.Dana H, Sun Y, Mohar B, Hulse BK, Kerlin AM, Hasseman JP, Tsegaye G, Tsang A, Wong A, Patel R, Macklin JJ, Chen Y, Konnerth A, Jayaraman V, Looger LL, Schreiter ER, Svoboda K, Kim DS. High-performance calcium sensors for imaging activity in neuronal populations and microcompartments. Nat Methods. 2019;16:649–657. doi: 10.1038/s41592-019-0435-6. [DOI] [PubMed] [Google Scholar]
- 50.Davalos D, Grutzendler J, Yang G, Kim JV, Zuo Y, Jung S, Littman DR, Dustin ML, Gan WB. ATP mediates rapid microglial response to local brain injury in vivo. Nat Neurosci. 2005;8:752–758. doi: 10.1038/nn1472. [DOI] [PubMed] [Google Scholar]
- 51.De La Crompe B, Coulon P, Diester I. Functional interrogation of neural circuits with virally transmitted optogenetic tools. J Neurosci Methods. 2020;345:108905. doi: 10.1016/j.jneumeth.2020.108905. [DOI] [PubMed] [Google Scholar]
- 52.Denk W, Strickler JH, Webb WW. Two-photon laser scanning fluorescence microscopy. Science. 1990;248:73–76. doi: 10.1126/science.2321027. [DOI] [PubMed] [Google Scholar]
- 53.Diaspro A, Bianchini P, Vicidomini G, Faretta M, Ramoino P, Usai C. Multi-photon excitation microscopy. Biomed Eng Online. 2006;5:36. doi: 10.1186/1475-925X-5-36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Ding S, Wang T, Cui W, Haydon PG. Photothrombosis ischemia stimulates a sustained astrocytic Ca2+signaling in vivo. Glia. 2009;57:767–776. doi: 10.1002/glia.20804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Diniz LP, Matias I, Siqueira M, Stipursky J, Gomes FCA. Astrocytes and the TGF-beta 1 pathway in the healthy and diseased brain: a double-edged sword. Mol Neurobiol. 2019;56:4653–4679. doi: 10.1007/s12035-018-1396-y. [DOI] [PubMed] [Google Scholar]
- 56.Dirnagl U, Villringer A, Einhäupl KM. In-vivo confocal scanning laser microscopy of the cerebral microcirculation. J Microsc. 1992;165:147–157. doi: 10.1111/j.1365-2818.1992.tb04312.x. [DOI] [PubMed] [Google Scholar]
- 57.Dong QP, He JQ, Chai Z. Astrocytic Ca(2+) waves mediate activation of extrasynaptic NMDA receptors in hippocampal neurons to aggravate brain damage during ischemia. Neurobiol Dis. 2013;58:68–75. doi: 10.1016/j.nbd.2013.05.005. [DOI] [PubMed] [Google Scholar]
- 58.Eichhoff G, Brawek B, Garaschuk O. Microglial calcium signal acts as a rapid sensor of single neuron damage in vivo. Biochim Biophys Acta. 2011;1813:1014–1024. doi: 10.1016/j.bbamcr.2010.10.018. [DOI] [PubMed] [Google Scholar]
- 59.El Amki M, Glück C, Binder N, Middleham W, Wyss MT, Weiss T, Meister H, Luft A, Weller M, Weber B, Wegener S. Neutrophils obstructing brain capillaries are a major cause of no-reflow in ischemic stroke. Cell Rep. 2020;33:108260. doi: 10.1016/j.celrep.2020.108260. [DOI] [PubMed] [Google Scholar]
- 60.Endo F, Kasai A, Soto JS, Yu X, Qu Z, Hashimoto H, Gradinaru V, Kawaguchi R, Khakh BS. Molecular basis of astrocyte diversity and morphology across the CNS in health and disease. Science. 2022;378:eadc9020. doi: 10.1126/science.adc9020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Fan JL, Rivera JA, Sun W, Peterson J, Haeberle H, Rubin S, Ji N. High-speed volumetric two-photon fluorescence imaging of neurovascular dynamics. Nat Commun. 2020;11:6020. doi: 10.1038/s41467-020-19851-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Fernández-Klett F, Offenhauser N, Dirnagl U, Priller J, Lindauer U. Pericytes in capillaries are contractile in vivo, but arterioles mediate functional hyperemia in the mouse brain. Proc Natl Acad Sci U S A. 2010;107:22290–22295. doi: 10.1073/pnas.1011321108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Forde A, Constien R, Gröne HJ, Hämmerling G, Arnold B. Temporal Cre-mediated recombination exclusively in endothelial cells using Tie2 regulatory elements. Genesis. 2002;33:191–197. doi: 10.1002/gene.10117. [DOI] [PubMed] [Google Scholar]
- 64.Fordsmann JC, Murmu RP, Cai C, Brazhe A, Thomsen KJ, Zambach SA, Lønstrup M, Lind BL, Lauritzen M. Spontaneous astrocytic Ca(2+) activity abounds in electrically suppressed ischemic penumbra of aged mice. Glia. 2019;67:37–52. doi: 10.1002/glia.23506. [DOI] [PubMed] [Google Scholar]
- 65.Fox MD. Mapping symptoms to brain networks with the human connectome. N Engl J Med. 2018;379:2237–2245. doi: 10.1056/NEJMra1706158. [DOI] [PubMed] [Google Scholar]
- 66.Frederix K, Chauhan AK, Kisucka J, Zhao B-Q, Hoff EI, Spronk HMH, Ten Cate H, Wagner DD. Platelet adhesion receptors do not modulate infarct volume after a photochemically induced stroke in mice. Brain Res. 2007;1185:239–245. doi: 10.1016/j.brainres.2007.07.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Freeman MR. Specification and morphogenesis of astrocytes. Science. 2010;330:774–778. doi: 10.1126/science.1190928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Friedman B, Schachtrup C, Tsai PS, Shih AY, Akassoglou K, Kleinfeld D, Lyden PD. Acute vascular disruption and aquaporin 4 loss after stroke. Stroke. 2009;40:2182–2190. doi: 10.1161/STROKEAHA.108.523720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Füger P, Hefendehl JK, Veeraraghavalu K, Wendeln AC, Schlosser C, Obermüller U, Wegenast-Braun BM, Neher JJ, Martus P, Kohsaka S, Thunemann M, Feil R, Sisodia SS, Skodras A, Jucker M. Microglia turnover with aging and in an Alzheimer's model via long-term in vivo single-cell imaging. Nat Neurosci. 2017;20:1371–1376. doi: 10.1038/nn.4631. [DOI] [PubMed] [Google Scholar]
- 70.Fumagalli S, Coles JA, Ejlerskov P, Ortolano F, Bushell TJ, Brewer JM, De Simoni MG, Dever G, Garside P, Maffia P, Carswell HV. In vivo real-time multiphoton imaging of T lymphocytes in the mouse brain after experimental stroke. Stroke. 2011;42:1429–1436. doi: 10.1161/STROKEAHA.110.603704. [DOI] [PubMed] [Google Scholar]
- 71.Fumagalli S, Perego C, Ortolano F, De Simoni MG. CX3CR1 deficiency induces an early protective inflammatory environment in ischemic mice. Glia. 2013;61:827–842. doi: 10.1002/glia.22474. [DOI] [PubMed] [Google Scholar]
- 72.Fumagalli S, Ortolano F, De Simoni MG. A close look at brain dynamics: cells and vessels seen by in vivo two-photon microscopy. Prog Neurobiol. 2014;121:36–54. doi: 10.1016/j.pneurobio.2014.06.005. [DOI] [PubMed] [Google Scholar]
- 73.Garcia-Caceres C, Balland E, Prevot V, Luquet S, Woods SC, Koch M, Horvath TL, Yi CX, Chowen JA, Verkhratsky A, Araque A, Bechmann I, Tschop MH. Role of astrocytes, microglia, and tanycytes in brain control of systemic metabolism. Nat Neurosci. 2019;22:7–14. doi: 10.1038/s41593-018-0286-y. [DOI] [PubMed] [Google Scholar]
- 74.GBD 2019 Stroke Collaborators. Global, regional, and national burden of stroke and its risk factors, 1990-2019: a systematic analysis for the Global Burden of Disease Study 2019. Lancet Neurol. 2021;20:795–820. doi: 10.1016/S1474-4422(21)00252-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Gee JM, Smith NA, Fernandez FR, Economo MN, Brunert D, Rothermel M, Morris SC, Talbot A, Palumbos S, Ichida JM, Shepherd JD, West PJ, Wachowiak M, Capecchi MR, Wilcox KS, White JA, Tvrdik P. Imaging activity in neurons and glia with a Polr2a-based and cre-dependent GCaMP5G-IRES-tdTomato reporter mouse. Neuron. 2014;83:1058–1072. doi: 10.1016/j.neuron.2014.07.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Glück C, Ferrari KD, Binini N, Keller A, Saab AS, Stobart JL, Weber B. Distinct signatures of calcium activity in brain mural cells. Elife. 2021;10:e70591. doi: 10.7554/eLife.70591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Goldmann T, Wieghofer P, Jordão MJ, Prutek F, Hagemeyer N, Frenzel K, Amann L, Staszewski O, Kierdorf K, Krueger M, Locatelli G, Hochgerner H, Zeiser R, Epelman S, Geissmann F, Priller J, Rossi FM, Bechmann I, Kerschensteiner M, Linnarsson S, et al. Origin, fate and dynamics of macrophages at central nervous system interfaces. Nat Immunol. 2016;17:797–805. doi: 10.1038/ni.3423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Gordon GR, Choi HB, Rungta RL, Ellis-Davies GC, MacVicar BA. Brain metabolism dictates the polarity of astrocyte control over arterioles. Nature. 2008;456:745–749. doi: 10.1038/nature07525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Greenhalgh AD, David S, Bennett FC. Immune cell regulation of glia during CNS injury and disease. Nat Rev Neurosci. 2020;21:139–152. doi: 10.1038/s41583-020-0263-9. [DOI] [PubMed] [Google Scholar]
- 80.Groothuis DR, Vavra MW, Schlageter KE, Kang EW, Itskovich AC, Hertzler S, Allen CV, Lipton HL. Efflux of drugs and solutes from brain: the interactive roles of diffusional transcapillary transport, bulk flow and capillary transporters. J Cereb Blood Flow Metab. 2007;27:43–56. doi: 10.1038/sj.jcbfm.9600315. [DOI] [PubMed] [Google Scholar]
- 81.Grubb S, Cai C, Hald BO, Khennouf L, Murmu RP, Jensen AGK, Fordsmann J, Zambach S, Lauritzen M. Precapillary sphincters maintain perfusion in the cerebral cortex. Nat Commun. 2020;11:395. doi: 10.1038/s41467-020-14330-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Grubb S, Lauritzen M, Aalkjær C. Brain capillary pericytes and neurovascular coupling. Comp Biochem Physiol A Mol Integr Physiol. 2021;254:110893. doi: 10.1016/j.cbpa.2020.110893. [DOI] [PubMed] [Google Scholar]
- 83.Guglielmi V, LeCouffe NE, Zinkstok SM, Compagne KCJ, Eker R, Treurniet KM, Tolhuisen ML, van der Worp HB, Jansen IGH, van Oostenbrugge RJ, Marquering HA, Dippel DWJ, Emmer BJ, Majoie CBLM, Roos YBWEM, Coutinho JM, et al. MR-CLEAN Registry Investigators. Collateral circulation and outcome in atherosclerotic versus cardioembolic cerebral large vessel occlusion. Stroke. 2019;50:3360–3368. doi: 10.1161/STROKEAHA.119.026299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Guo J, Ran M, Gao Z, Zhang X, Wang D, Li H, Zhao S, Sun W, Dong H, Hu J. Cell-type-specific imaging of neurotransmission reveals a disrupted excitatory-inhibitory cortical network in isoflurane anaesthesia. EBioMedicine. 2021;65:103272. doi: 10.1016/j.ebiom.2021.103272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Gur S, Wolf L, Golgher L, Blinder P. Microvascular dynamics from 4D microscopy using temporal segmentation. Pac Symp Biocomput. 2020;25:331–342. [PubMed] [Google Scholar]
- 86.G?bel J, Engelhardt E, Pelzer P, Sakthivelu V, Jahn HM, Jevtic M, Folz-Donahue K, Kukat C, Schauss A, Frese CK, Giavalisco P, Ghanem A, Conzelmann KK, Motori E, Bergami M. Mitochondria-endoplasmic reticulum contacts in reactive astrocytes promote vascular remodeling. Cell Metab. 2020;31:791–808.e798. doi: 10.1016/j.cmet.2020.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Haimon Z, Volaski A, Orthgiess J, Boura-Halfon S, Varol D, Shemer A, Yona S, Zuckerman B, David E, Chappell-Maor L, Bechmann I, Gericke M, Ulitsky I, Jung S. Re-evaluating microglia expression profiles using RiboTag and cell isolation strategies. Nat Immunol. 2018;19:636–644. doi: 10.1038/s41590-018-0110-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Hanisch UK, Kettenmann H. Microglia: active sensor and versatile effector cells in the normal and pathologic brain. Nat Neurosci. 2007;10:1387–1394. doi: 10.1038/nn1997. [DOI] [PubMed] [Google Scholar]
- 89.Hartmann DA, Underly RG, Watson AN, Shih AY. A murine toolbox for imaging the neurovascular unit. Microcirculation. 2015a;22:168–182. doi: 10.1111/micc.12176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Hartmann DA, Underly RG, Grant RI, Watson AN, Lindner V, Shih AY. Pericyte structure and distribution in the cerebral cortex revealed by high-resolution imaging of transgenic mice. Neurophotonics. 2015b;2:041402. doi: 10.1117/1.NPh.2.4.041402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Hartmann DA, Berthiaume AA, Grant RI, Harrill SA, Koski T, Tieu T, McDowell KP, Faino AV, Kelly AL, Shih AY. Brain capillary pericytes exert a substantial but slow influence on blood flow. Nature Neuroscience. 2021;24:633–645. doi: 10.1038/s41593-020-00793-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Hatakeyama M, Ninomiya I, Kanazawa M. Angiogenesis and neuronal remodeling after ischemic stroke. Neural Regen Res. 2020;15:16–19. doi: 10.4103/1673-5374.264442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.He S, Song J, Qu J, Cheng Z. Crucial breakthrough of second near-infrared biological window fluorophores: design and synthesis toward multimodal imaging and theranostics. Chem Soc Rev. 2018;47:4258–4278. doi: 10.1039/c8cs00234g. [DOI] [PubMed] [Google Scholar]
- 94.Heithoff BP, George KK, Phares AN, Zuidhoek IA, Munoz-Ballester C, Robel S. Astrocytes are necessary for blood-brain barrier maintenance in the adult mouse brain. Glia. 2021;69:436–472. doi: 10.1002/glia.23908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Helmchen F, Denk W. Deep tissue two-photon microscopy. Nat Methods. 2005;2:932–940. doi: 10.1038/nmeth818. [DOI] [PubMed] [Google Scholar]
- 96.Heo DK, Lim HM, Nam JH, Lee MG, Kim JY. Regulation of phagocytosis and cytokine secretion by store-operated calcium entry in primary isolated murine microglia. Cell Signal. 2015;27:177–186. doi: 10.1016/j.cellsig.2014.11.003. [DOI] [PubMed] [Google Scholar]
- 97.Heras-Romero Y, Morales-Guadarrama A, Santana-Martínez R, Ponce I, Rincón-Heredia R, Poot-Hernández AC, Martínez-Moreno A, Urrieta E, Bernal-Vicente BN, Campero-Romero AN, Moreno-Castilla P, Greig NH, Escobar ML, Concha L, Tovar-Y-Romo LB. Improved post-stroke spontaneous recovery by astrocytic extracellular vesicles. Mol Ther. 2022;30:798–815. doi: 10.1016/j.ymthe.2021.09.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Herbet G, Duffau H. Revisiting the functional anatomy of the human brain: toward a meta-networking theory of cerebral functions. Physiol Rev. 2020;100:1181–1228. doi: 10.1152/physrev.00033.2019. [DOI] [PubMed] [Google Scholar]
- 99.Hickman SE, Allison EK, Coleman U, Kingery-Gallagher ND, El Khoury J. Heterozygous CX3CR1 deficiency in microglia restores neuronal β-amyloid clearance pathways and slows progression of Alzheimer's like-disease in PS1-APP mice. Front Immunol. 2019;10:2780. doi: 10.3389/fimmu.2019.02780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Hierro-Bujalance C, Bacskai BJ, Garcia-Alloza M. In vivo imaging of microglia with multiphoton microscopy. Front Aging Neurosci. 2018;10:218. doi: 10.3389/fnagi.2018.00218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Hill RA, Grutzendler J. In vivo imaging of oligodendrocytes with sulforhodamine 101. Nat Methods. 2014;11:1081–1082. doi: 10.1038/nmeth.3140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Hill RA, Tong L, Yuan P, Murikinati S, Gupta S, Grutzendler J. Regional blood flow in the normal and ischemic brain is controlled by arteriolar smooth muscle cell contractility and not by capillary pericytes. Neuron. 2015;87:95–110. doi: 10.1016/j.neuron.2015.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Hirasawa T, Ohsawa K, Imai Y, Ondo Y, Akazawa C, Uchino S, Kohsaka S. Visualization of microglia in living tissues using Iba1-EGFP transgenic mice. J Neurosci Res. 2005;81:357–362. doi: 10.1002/jnr.20480. [DOI] [PubMed] [Google Scholar]
- 104.Hirase H, Qian L, Barthó P, Buzsáki G. Calcium dynamics of cortical astrocytic networks in vivo. PLoS Biol. 2004;2:E96. doi: 10.1371/journal.pbio.0020096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Hladky SB, Barrand MA. Mechanisms of fluid movement into, through and out of the brain: evaluation of the evidence. Fluids Barriers CNS. 2014;11:26. doi: 10.1186/2045-8118-11-26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Holter KE, Kehlet B, Devor A, Sejnowski TJ, Dale AM, Omholt SW, Ottersen OP, Nagelhus EA, Mardal KA, Pettersen KH. Interstitial solute transport in 3D reconstructed neuropil occurs by diffusion rather than bulk flow. Proc Natl Acad Sci U S A. 2017;114:9894–9899. doi: 10.1073/pnas.1706942114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Hösli L, Zuend M, Bredell G, Zanker HS, Porto de Oliveira CE, Saab AS, Weber B. Direct vascular contact is a hallmark of cerebral astrocytes. Cell Rep. 2022;39:110599. doi: 10.1016/j.celrep.2022.110599. [DOI] [PubMed] [Google Scholar]
- 108.Hou B, Jiang C, Wang D, Wang G, Wang Z, Zhu M, Kang Y, Su J, Wei P, Ren H, Ju F. Pharmacological targeting of CSF1R inhibits microglial proliferation and aggravates the progression of cerebral ischemic pathology. Front Cell Neurosci. 2020;14:267. doi: 10.3389/fncel.2020.00267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Iadecola C. The neurovascular unit coming of age: a journey through neurovascular coupling in health and disease. Neuron. 2017;96:17–42. doi: 10.1016/j.neuron.2017.07.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Iliff JJ, Wang M, Liao Y, Plogg BA, Peng W, Gundersen GA, Benveniste H, Vates GE, Deane R, Goldman SA, Nagelhus EA, Nedergaard M. A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid β. Sci Transl Med. 2012;4:147ra111. doi: 10.1126/scitranslmed.3003748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Jayaraj RL, Azimullah S, Beiram R, Jalal FY, Rosenberg GA. Neuroinflammation: friend and foe for ischemic stroke. J Neuroinflammation. 2019;16:142. doi: 10.1186/s12974-019-1516-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Jolivel V, Bicker F, Binamé F, Ploen R, Keller S, Gollan R, Jurek B, Birkenstock J, Poisa-Beiro L, Bruttger J, Opitz V, Thal SC, Waisman A, Bäuerle T, Schäfer MK, Zipp F, Schmidt MHH. Perivascular microglia promote blood vessel disintegration in the ischemic penumbra. Acta Neuropathol. 2015;129:279–295. doi: 10.1007/s00401-014-1372-1. [DOI] [PubMed] [Google Scholar]
- 113.Jung S, Aliberti J, Graemmel P, Sunshine MJ, Kreutzberg GW, Sher A, Littman DR. Analysis of fractalkine receptor CX(3)CR1 function by targeted deletion and green fluorescent protein reporter gene insertion. Mol Cell Biol. 2000;20:4106–4114. doi: 10.1128/mcb.20.11.4106-4114.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Jurga AM, Paleczna M, Kuter KZ. Overview of general and discriminating markers of differential microglia phenotypes. Front Cell Neurosci. 2020;14:198. doi: 10.3389/fncel.2020.00198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Kabba JA, Xu Y, Christian H, Ruan W, Chenai K, Xiang Y, Zhang L, Saavedra JM, Pang T. Microglia: housekeeper of the central nervous system. Cell Mol Neurobiol. 2018;38:53–71. doi: 10.1007/s10571-017-0504-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Kaiser T, Feng G. Tmem119-EGFP and Tmem119-CreERT2 transgenic mice for labeling and manipulating microglia. eNeuro. 2019;6:ENEURO.0448–18.2019. doi: 10.1523/ENEURO.0448-18.2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Kanazawa M, Takahashi T, Ishikawa M, Onodera O, Shimohata T, Del Zoppo GJ. Angiogenesis in the ischemic core: A potential treatment target? J Cereb Blood Flow Metab. 2019;39:753–769. doi: 10.1177/0271678X19834158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Kearns KN, Liu L, Sharifi KA, Stauderman KA, Park MS, Tvrdik P. Microglial calcium activity during ischemic stroke. Circulation. 2020 doi:10.1161/circ.142.suppl_3.14278. [Google Scholar]
- 119.Kearns KN, Liu L, Soldozy S, Sharifi KA, Shaffrey ME, Park MS, Tvrdik P. Microglia modulate cortical spreading depolarizations after ischemic stroke: a narrative review. Neurocrit Care. 2022;37(Suppl 1):133–138. doi: 10.1007/s12028-022-01469-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Kim GW, Sugawara T, Chan PH. Involvement of oxidative stress and caspase-3 in cortical infarction after photothrombotic ischemia in mice. J Cereb Blood Flow Metab. 2000;20:1690–1701. doi: 10.1097/00004647-200012000-00008. [DOI] [PubMed] [Google Scholar]
- 121.Kirdajova DB, Kriska J, Tureckova J, Anderova M. Ischemia-triggered glutamate excitotoxicity from the perspective of glial cells. Front Cell Neurosci. 2020;14:51. doi: 10.3389/fncel.2020.00051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Kirst C, Skriabine S, Vieites-Prado A, Topilko T, Bertin P, Gerschenfeld G, Verny F, Topilko P, Michalski N, Tessier-Lavigne M, Renier N. Mapping the fine-scale organization and plasticity of the brain vasculature. Cell. 2020;180:780–795.e725. doi: 10.1016/j.cell.2020.01.028. [DOI] [PubMed] [Google Scholar]
- 123.Kisanuki YY, Hammer RE, Miyazaki J, Williams SC, Richardson JA, Yanagisawa M. Tie2-Cre transgenic mice: a new model for endothelial cell-lineage analysis in vivo. Dev Biol. 2001;230:230–242. doi: 10.1006/dbio.2000.0106. [DOI] [PubMed] [Google Scholar]
- 124.Kleinfeld D, Mitra PP, Helmchen F, Denk W. Fluctuations and stimulus-induced changes in blood flow observed in individual capillaries in layers 2 through 4 of rat neocortex. Proc Natl Acad Sci U S A. 1998;95:15741–15746. doi: 10.1073/pnas.95.26.15741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Kleinschnitz C, Braeuninger S, Pham M, Austinat M, Nölte I, Renné T, Nieswandt B, Bendszus M, Stoll G. Blocking of platelets or intrinsic coagulation pathway-driven thrombosis does not prevent cerebral infarctions induced by photothrombosis. Stroke. 2008;39:1262–1268. doi: 10.1161/STROKEAHA.107.496448. [DOI] [PubMed] [Google Scholar]
- 126.Kloner RA. No-reflow phenomenon: maintaining vascular integrity. J Cardiovasc Pharmacol Ther. 2011;16:244–250. doi: 10.1177/1074248411405990. [DOI] [PubMed] [Google Scholar]
- 127.Kluge MG, Kracht L, Abdolhoseini M, Ong LK, Johnson SJ, Nilsson M, Walker FR. Impaired microglia process dynamics post-stroke are specific to sites of secondary neurodegeneration. Glia. 2017;65:1885–1899. doi: 10.1002/glia.23201. [DOI] [PubMed] [Google Scholar]
- 128.Kluge MG, Abdolhoseini M, Zalewska K, Ong LK, Johnson SJ, Nilsson M, Walker FR. Spatiotemporal analysis of impaired microglia process movement at sites of secondary neurodegeneration post-stroke. J Cereb Blood Flow Metab. 2019;39:2456–2470. doi: 10.1177/0271678X18797346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Knowland D, Arac A, Sekiguchi KJ, Hsu M, Lutz SE, Perrino J, Steinberg GK, Barres BA, Nimmerjahn A, Agalliu D. Stepwise recruitment of transcellular and paracellular pathways underlies blood-brain barrier breakdown in stroke. Neuron. 2014;82:603–617. doi: 10.1016/j.neuron.2014.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Krafft PR, Bailey EL, Lekic T, Rolland WB, Altay O, Tang J, Wardlaw JM, Zhang JH, Sudlow CL. Etiology of stroke and choice of models. Int J Stroke. 2012;7:398–406. doi: 10.1111/j.1747-4949.2012.00838.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Kreutzberg GW. Microglia: a sensor for pathological events in the CNS. Trends Neurosci. 1996;19:312–318. doi: 10.1016/0166-2236(96)10049-7. [DOI] [PubMed] [Google Scholar]
- 132.Kubotera H, Ikeshima-Kataoka H, Hatashita Y, Allegra Mascaro AL, Pavone FS, Inoue T. Astrocytic endfeet re-cover blood vessels after removal by laser ablation. Sci Rep. 2019;9:1263. doi: 10.1038/s41598-018-37419-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Labat-gest V, Tomasi S. Photothrombotic ischemia: a minimally invasive and reproducible photochemical cortical lesion model for mouse stroke studies. J Vis Exp. 2013:50370. doi: 10.3791/50370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Lam CK, Yoo T, Hiner B, Liu Z, Grutzendler J. Embolus extravasation is an alternative mechanism for cerebral microvascular recanalization. Nature. 2010;465:478–482. doi: 10.1038/nature09001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Lasek-Bal A, Jedrzejowska-Szypulka H, Student S, Warsz-Wianecka A, Zareba K, Puz P, Bal W, Pawletko K, Lewin-Kowalik J. The importance of selected markers of inflammation and blood-brain barrier damage for short-term ischemic stroke prognosis. J Physiol Pharmacol. 2019;70:209–217. doi: 10.26402/jpp.2019.2.04. [DOI] [PubMed] [Google Scholar]
- 136.Le Grand Y, Leray A, Guilbert T, Odin C. Non-descanned versus descanned epifluorescence collection in two-photon microscopy: Experiments and Monte Carlo simulations. Opt Commun. 2008;281:5480–5486. [Google Scholar]
- 137.Leben R, Lindquist RL, Hauser AE, Niesner R, Rakhymzhan A. Two-photon excitation spectra of various fluorescent proteins within a broad excitation range. Int J Mol Sci. 2022;23:13407. doi: 10.3390/ijms232113407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Li H, Xie Y, Zhang N, Yu Y, Zhang Q, Ding S. Disruption of IPƒR2-mediated Ca²z signaling pathway in astrocytes ameliorates neuronal death and brain damage while reducing behavioral deficits after focal ischemic stroke. Cell Calcium. 2015;58:565–576. doi: 10.1016/j.ceca.2015.09.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Li P, Wang L, Zhou Y, Gan Y, Zhu W, Xia Y, Jiang X, Watkins S, Vazquez A, Thomson AW, Chen J, Yu W, Hu X. C-C chemokine receptor type 5 (CCR5)-mediated docking of transferred tregs protects against early blood-brain barrier disruption after stroke. J Am Heart Assoc. 2017;6:e006387. doi: 10.1161/JAHA.117.006387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Li PY, Stetler RA, Leak RK, Shi YJ, Li Y, Yu WF, Bennett MVL, Chen J. Oxidative stress and DNA damage after cerebral ischemia: Potential therapeutic targets to repair the genome and improve stroke recovery. Neuropharmacology. 2018;134:208–217. doi: 10.1016/j.neuropharm.2017.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Li Q, Cheng Z, Zhou L, Darmanis S, Neff NF, Okamoto J, Gulati G, Bennett ML, Sun LO, Clarke LE, Marschallinger J, Yu G, Quake SR, Wyss-Coray T, Barres BA. Developmental heterogeneity of microglia and brain myeloid cells revealed by deep single-cell RNA sequencing. Neuron. 2019a;101:207–223.e10. doi: 10.1016/j.neuron.2018.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Li T, Pang S, Yu Y, Wu X, Guo J, Zhang S. Proliferation of parenchymal microglia is the main source of microgliosis after ischaemic stroke. Brain. 2013;136:3578–3588. doi: 10.1093/brain/awt287. [DOI] [PubMed] [Google Scholar]
- 143.Li Y, Zhang J. Animal models of stroke. Animal Model Exp Med. 2021;4:204–219. doi: 10.1002/ame2.12179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Li YC, Zhong W, Jiang Z, Tang XQ. New progress in the approaches for blood-brain barrier protection in acute ischemic stroke. Brain Res Bull. 2019b;144:46–57. doi: 10.1016/j.brainresbull.2018.11.006. [DOI] [PubMed] [Google Scholar]
- 145.Li YQ, Chen P, Jain V, Reilly RM, Wong CS. Early radiation-induced endothelial cell loss and blood-spinal cord barrier breakdown in the rat spinal cord. Radiat Res. 2004;161:143–152. doi: 10.1667/rr3117. [DOI] [PubMed] [Google Scholar]
- 146.Liao Z, Bu Y, Li M, Han R, Zhang N, Hao J, Jiang W. Remote ischemic conditioning improves cognition in patients with subcortical ischemic vascular dementia. BMC Neurol. 2019;19:206. doi: 10.1186/s12883-019-1435-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Liddelow SA, Barres B. Reactive astrocytes: production, function, and therapeutic potential. Immunity. 2017;46:957–967. doi: 10.1016/j.immuni.2017.06.006. [DOI] [PubMed] [Google Scholar]
- 148.Liddelow SA, Guttenplan KA, Clarke LE, Bennett FC, Bohlen CJ, Schirmer L, Bennett ML, Münch AE, Chung WS, Peterson TC, Wilton DK, Frouin A, Napier BA, Panicker N, Kumar M, Buckwalter MS, Rowitch DH, Dawson VL, Dawson TM, Stevens B, Barres BA, et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature. 2017;541:481–487. doi: 10.1038/nature21029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Liebner S, Dijkhuizen RM, Reiss Y, Plate KH, Agalliu D, Constantin G. Functional morphology of the blood-brain barrier in health and disease. Acta Neuropathol. 2018;135:311–336. doi: 10.1007/s00401-018-1815-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Liu L, Kearns KN, Eli I, Sharifi KA, Soldozy S, Carlson EW, Scott KW, Sluzewski MF, Acton ST, Stauderman KA, Kalani MYS, Park M, Tvrdik P. Microglial calcium waves during the hyperacute phase of ischemic stroke. Stroke. 2021;52:274–283. doi: 10.1161/STROKEAHA.120.032766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Liu LR, Liu JC, Bao JS, Bai QQ, Wang GQ. Interaction of microglia and astrocytes in the neurovascular unit. Front Immunol. 2020;11:1024. doi: 10.3389/fimmu.2020.01024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Liu YU, Ying Y, Li Y, Eyo UB, Chen T, Zheng J, Umpierre AD, Zhu J, Bosco DB, Dong H, Wu LJ. Neuronal network activity controls microglial process surveillance in awake mice via norepinephrine signaling. Nat Neurosci. 2019;22:1771–1781. doi: 10.1038/s41593-019-0511-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Longa EZ, Weinstein PR, Carlson S, Cummins R. Reversible middle cerebral artery occlusion without craniectomy in rats. Stroke. 1989;20:84–91. doi: 10.1161/01.str.20.1.84. [DOI] [PubMed] [Google Scholar]
- 154.Luo C, Liang F, Ren H, Yao X, Liu Q, Li M, Qin D, Yuan TF, Pei Z, Su H. Collateral blood flow in different cerebrovascular hierarchy provides endogenous protection in cerebral ischemia. Brain Pathol. 2017;27:809–821. doi: 10.1111/bpa.12458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Ma J, Ma Y, Shuaib A, Winship IR. Impaired collateral flow in pial arterioles of aged rats during ischemic stroke. Transl Stroke Res. 2020a;11:243–253. doi: 10.1007/s12975-019-00710-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Ma J, Ma Y, Shuaib A, Winship IR. Improved collateral flow and reduced damage after remote ischemic perconditioning during distal middle cerebral artery occlusion in aged rats. Sci Rep. 2020b;10:12392. doi: 10.1038/s41598-020-69122-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Ma Z, Wei L, Du X, Hou S, Chen F, Jiao Q, Liu A, Liu S, Wang J, Shen H. Two-photon calcium imaging of neuronal and astrocytic responses: the influence of electrical stimulus parameters and calcium signaling mechanisms. J Neural Eng. 2021 doi: 10.1088/1741-2552/ac0b50. doi:10.1088/1741-2552/ac0b50. [DOI] [PubMed] [Google Scholar]
- 158.Macknik SL, Alexander RG, Caballero O, Chanovas J, Nielsen KJ, Nishimura N, Schaffer CB, Slovin H, Babayoff A, Barak R, Tang SM, Ju NS, Yazdan-Shahmorad A, Alonso JM, Malinskiy E, Martinez-Conde S. Advanced circuit and cellular imaging methods in nonhuman primates. J Neurosci. 2019;39:8267–8274. doi: 10.1523/JNEUROSCI.1168-19.2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Manley GT, Binder DK, Papadopoulos MC, Verkman AS. New insights into water transport and edema in the central nervous system from phenotype analysis of aquaporin-4 null mice. Neuroscience. 2004;129:983–991. doi: 10.1016/j.neuroscience.2004.06.088. [DOI] [PubMed] [Google Scholar]
- 160.Marker DF, Tremblay ME, Lu SM, Majewska AK, Gelbard HA. A thin-skull window technique for chronic two-photon in vivo imaging of murine microglia in models of neuroinflammation. J Vis Exp. 2010:2059. doi: 10.3791/2059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Mashinchian O, Johari-Ahar M, Ghaemi B, Rashidi M, Barar J, Omidi Y. Impacts of quantum dots in molecular detection and bioimaging of cancer. Bioimpacts. 2014;4:149–166. doi: 10.15171/bi.2014.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Masuda T, Croom D, Hida H, Kirov SA. Capillary blood flow around microglial somata determines dynamics of microglial processes in ischemic conditions. Glia. 2011;59:1744–1753. doi: 10.1002/glia.21220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Matejuk A, Ransohoff RM. Crosstalk between astrocytes and microglia: an overview. Front Immunol. 2020;11:1416. doi: 10.3389/fimmu.2020.01416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.McDonald MW, Dykes A, Jeffers MS, Carter A, Nevins R, Ripley A, Silasi G, Corbett D. Remote ischemic conditioning and stroke recovery. Neurorehabil Neural Repair. 2021;35:545–549. doi: 10.1177/15459683211011224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.McNeill J, Rudyk C, Hildebrand ME, Salmaso N. Ion channels and electrophysiological properties of astrocytes: implications for emergent stimulation technologies. Front Cell Neurosci. 2021;15:644126. doi: 10.3389/fncel.2021.644126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Mestre H, Tithof J, Du T, Song W, Peng W, Sweeney AM, Olveda G, Thomas JH, Nedergaard M, Kelley DH. Flow of cerebrospinal fluid is driven by arterial pulsations and is reduced in hypertension. Nat Commun. 2018;9:4878. doi: 10.1038/s41467-018-07318-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Mestre H, Mori Y, Nedergaard M. The brain's glymphatic system: current controversies. Trends Neurosci. 2020a;43:458–466. doi: 10.1016/j.tins.2020.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Mestre H, Du T, Sweeney AM, Liu G, Samson AJ, Peng W, Mortensen KN, Stæger FF, Bork PAR, Bashford L, Toro ER, Tithof J, Kelley DH, Thomas JH, Hjorth PG, Martens EA, Mehta RI, Solis O, Blinder P, Kleinfeld D, et al. Cerebrospinal fluid influx drives acute ischemic tissue swelling. Science. 2020b;367:eaax7171. doi: 10.1126/science.aax7171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Metea MR, Newman EA. Glial cells dilate and constrict blood vessels: a mechanism of neurovascular coupling. J Neurosci. 2006;26:2862–2870. doi: 10.1523/JNEUROSCI.4048-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Meza-Resillas J, Ahmadpour N, Stobart M, Stobart J. Brain pericyte calcium and hemodynamic imaging in transgenic mice in vivo. J Vis Exp. 2021 doi: 10.3791/62725. doi:10.3791/62725. [DOI] [PubMed] [Google Scholar]
- 171.Mills WA, 3rd, Woo AM, Jiang S, Martin J, Surendran D, Bergstresser M, Kimbrough IF, Eyo UB, Sofroniew MV, Sontheimer H. Astrocyte plasticity in mice ensures continued endfoot coverage of cerebral blood vessels following injury and declines with age. Nat Commun. 2022;13:1794. doi: 10.1038/s41467-022-29475-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Mishra A, Reynolds JP, Chen Y, Gourine AV, Rusakov DA, Attwell D. Astrocytes mediate neurovascular signaling to capillary pericytes but not to arterioles. Nat Neurosci. 2016;19:1619–1627. doi: 10.1038/nn.4428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Miteff F, Levi CR, Bateman GA, Spratt N, McElduff P, Parsons MW. The independent predictive utility of computed tomography angiographic collateral status in acute ischaemic stroke. Brain. 2009;132:2231–2238. doi: 10.1093/brain/awp155. [DOI] [PubMed] [Google Scholar]
- 174.Motoike T, Loughna S, Perens E, Roman BL, Liao W, Chau TC, Richardson CD, Kawate T, Kuno J, Weinstein BM, Stainier DY, Sato TN. Universal GFP reporter for the study of vascular development. Genesis. 2000;28:75–81. doi: 10.1002/1526-968x(200010)28:2<75::aid-gene50>3.0.co;2-s. [DOI] [PubMed] [Google Scholar]
- 175.Mulligan SJ, MacVicar BA. Calcium transients in astrocyte endfeet cause cerebrovascular constrictions. Nature. 2004;431:195–199. doi: 10.1038/nature02827. [DOI] [PubMed] [Google Scholar]
- 176.Murmu RP, Fordsmann JC, Cai C, Brazhe A, Thomsen KJ, Lauritzen M. Sensory stimulation-induced astrocytic calcium signaling in electrically silent ischemic penumbra. Front Aging Neurosci. 2019;11:223. doi: 10.3389/fnagi.2019.00223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Nedergaard M, Ransom B, Goldman SA. New roles for astrocytes: Redefining the functional architecture of the brain. Trends Neurosci. 2003;26:523–530. doi: 10.1016/j.tins.2003.08.008. [DOI] [PubMed] [Google Scholar]
- 178.Neumann J, Gunzer M, Gutzeit HO, Ullrich O, Reymann KG, Dinkel K. Microglia provide neuroprotection after ischemia. FASEB J. 2006;20:714–716. doi: 10.1096/fj.05-4882fje. [DOI] [PubMed] [Google Scholar]
- 179.Neumann J, Henneberg S, von Kenne S, Nolte N, Müller AJ, Schraven B, Görtler MW, Reymann KG, Gunzer M, Riek-Burchardt M. Beware the intruder: Real time observation of infiltrated neutrophils and neutrophil-Microglia interaction during stroke in vivo. PLoS One. 2018;13:e0193970. doi: 10.1371/journal.pone.0193970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Ngai AC, Winn HR. Estimation of shear and flow rates in pial arterioles during somatosensory stimulation. Am J Physiol. 1996;270:H1712–1717. doi: 10.1152/ajpheart.1996.270.5.H1712. [DOI] [PubMed] [Google Scholar]
- 181.Nimmerjahn A, Kirchhoff F, Kerr JN, Helmchen F. Sulforhodamine 101 as a specific marker of astroglia in the neocortex in vivo. Nat Methods. 2004;1:31–37. doi: 10.1038/nmeth706. [DOI] [PubMed] [Google Scholar]
- 182.Nimmerjahn A, Kirchhoff F, Helmchen F. Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science. 2005;308:1314–1318. doi: 10.1126/science.1110647. [DOI] [PubMed] [Google Scholar]
- 183.Nishimura N, Schaffer CB, Friedman B, Tsai PS, Lyden PD, Kleinfeld D. Targeted insult to subsurface cortical blood vessels using ultrashort laser pulses: three models of stroke. Nat Methods. 2006;3:99–108. doi: 10.1038/nmeth844. [DOI] [PubMed] [Google Scholar]
- 184.Nishimura N, Schaffer CB, Friedman B, Lyden PD, Kleinfeld D. Penetrating arterioles are a bottleneck in the perfusion of neocortex. Proc Natl Acad Sci U S A. 2007;104:365–370. doi: 10.1073/pnas.0609551104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Nishimura N, Rosidi NL, Iadecola C, Schaffer CB. Limitations of collateral flow after occlusion of a single cortical penetrating arteriole. J Cereb Blood Flow Metab. 2010;30:1914–1927. doi: 10.1038/jcbfm.2010.157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Nolte C, Matyash M, Pivneva T, Schipke CG, Ohlemeyer C, Hanisch UK, Kirchhoff F, Kettenmann H. GFAP promoter-controlled EGFP-expressing transgenic mice: a tool to visualize astrocytes and astrogliosis in living brain tissue. Glia. 2001;33:72–86. [PubMed] [Google Scholar]
- 187.Olson JE, Yu JH, Thimes RL, Camden JP. Vibrational two-photon microscopy for tissue imaging: Short-wave infrared surface-enhanced resonance hyper-Raman scattering. J Biophotonics. 2022;15:e202100158. doi: 10.1002/jbio.202100158. [DOI] [PubMed] [Google Scholar]
- 188.Ortega SB, Torres VO, Latchney SE, Whoolery CW, Noorbhai IZ, Poinsatte K, Selvaraj UM, Benson MA, Meeuwissen AJM, Plautz EJ, Kong X, Ramirez DM, Ajay AD, Meeks JP, Goldberg MP, Monson NL, Eisch AJ, Stowe AM. B cells migrate into remote brain areas and support neurogenesis and functional recovery after focal stroke in mice. Proc Natl Acad Sci U S A. 2020;117:4983–4993. doi: 10.1073/pnas.1913292117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Ortolano F, Maffia P, Dever G, Rodolico G, Millington OR, De Simoni MG, Brewer JM, Bushell TJ, Garside P, Carswell HV. Advances in imaging of new targets for pharmacological intervention in stroke: real-time tracking of T-cells in the ischaemic brain. Br J Pharmacol. 2010;159:808–811. doi: 10.1111/j.1476-5381.2009.00527.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Otxoa-de-Amezaga A, Miro-Mur F, Pedragosa J, Gallizioli M, Justicia C, Gaja-Capdevila N, Ruiz-Jaen F, Salas-Perdomo A, Bosch A, Calvo M, Marquez-Kisinousky L, Denes A, Gunzer M, Planas AM. Microglial cell loss after ischemic stroke favors brain neutrophil accumulation. Acta Neuropathologica. 2019;137:321–341. doi: 10.1007/s00401-018-1954-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Paul S, Candelario-Jalil E. Emerging neuroprotective strategies for the treatment of ischemic stroke: An overview of clinical and preclinical studies. Exp Neurol. 2021;335:113518. doi: 10.1016/j.expneurol.2020.113518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Pekny M, Wilhelmsson U, Pekna M. The dual role of astrocyte activation and reactive gliosis. Neurosci Lett. 2014;565:30–38. doi: 10.1016/j.neulet.2013.12.071. [DOI] [PubMed] [Google Scholar]
- 193.Pena-Ortega F. Pharmacological tools to activate microglia and their possible use to study neural network patho-physiology. Curr Neuropharmacol. 2017;15:595–619. doi: 10.2174/1570159X14666160928151546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Perea G, Navarrete M, Araque A. Tripartite synapses: astrocytes process and control synaptic information. Trends Neurosci. 2009;32:421–431. doi: 10.1016/j.tins.2009.05.001. [DOI] [PubMed] [Google Scholar]
- 195.Piatkevich KD, Bensussen S, Tseng HA, Shroff SN, Lopez-Huerta VG, Park D, Jung EE, Shemesh OA, Straub C, Gritton HJ, Romano MF, Costa E, Sabatini BL, Fu Z, Boyden ES, Han X. Population imaging of neural activity in awake behaving mice. Nature. 2019;574:413–417. doi: 10.1038/s41586-019-1641-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Poinsatte K, Betz D, Torres VO, Ajay AD, Mirza S, Selvaraj UM, Plautz EJ, Kong XM, Gokhale S, Meeks JP, Ramirez DMO, Goldberg MP, Stowe AM. Visualization and quantification of post-stroke neural connectivity and neuroinflammation using serial two-photon tomography in the whole mouse brain. Front Neurosci. 2019;13:1055. doi: 10.3389/fnins.2019.01055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Powers WJ, Rabinstein AA, Ackerson T, Adeoye OM, Bambakidis NC, Becker K, Biller J, Brown M, Demaerschalk BM, Hoh B, Jauch EC, Kidwell CS, Leslie-Mazwi TM, Ovbiagele B, Scott PA, Sheth KN, Southerland AM, Summers DV, Tirschwell DL. Guidelines for the Early Management of Patients With Acute Ischemic Stroke:2019 Update to the 2018 Guidelines for the Early Management of Acute Ischemic Stroke: A Guideline for Healthcare Professionals From the American Heart Association/American Stroke Association. Stroke. 2019;50:e344–418. [Google Scholar]
- 198.Profaci CP, Munji RN, Pulido RS, Daneman R. The blood-brain barrier in health and disease: Important unanswered questions. J Exp Med. 2020;217:e20190062. doi: 10.1084/jem.20190062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Qi J, Sun CW, Li DY, Zhang HQ, Yu WB, Zebibula A, Lam JWY, Xi W, Zhu L, Cai FH, Wei PF, Zhu CL, Kwok RTK, Streich LL, Prevedel R, Qian J, Tang BZ. Aggregation-induced emission luminogen with near-infrared-II excitation and near-infrared-I emission for ultradeep intravital two-photon microscopy. ACS Nano. 2018;12:7936–7945. doi: 10.1021/acsnano.8b02452. [DOI] [PubMed] [Google Scholar]
- 200.Qin C, Zhou LQ, Ma XT, Hu ZW, Yang S, Chen M, Bosco DB, Wu LJ, Tian DS. Dual functions of microglia in ischemic stroke. Neurosci Bull. 2019;35:921–933. doi: 10.1007/s12264-019-00388-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Qin W, Alifu N, Lam JWY, Cui YH, Su HF, Liang GD, Qian J, Tang BZ. Facile synthesis of efficient luminogens with AIE features for three-photon fluorescence imaging of the brain through the intact skull. Adv Mater. 2020;32:e2000364. doi: 10.1002/adma.202000364. [DOI] [PubMed] [Google Scholar]
- 202.Rabinstein AA, Albers GW, Brinjikji W, Koch S. Factors that may contribute to poor outcome despite good reperfusion after acute endovascular stroke therapy. Int J Stroke. 2019;14:23–31. doi: 10.1177/1747493018799979. [DOI] [PubMed] [Google Scholar]
- 203.Rakers C, Petzold GC. Astrocytic calcium release mediates peri-infarct depolarizations in a rodent stroke model. J Clin Invest. 2017;127:511–516. doi: 10.1172/JCI89354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Rakers C, Schmid M, Petzold GC. TRPV4 channels contribute to calcium transients in astrocytes and neurons during peri-infarct depolarizations in a stroke model. Glia. 2017;65:1550–1561. doi: 10.1002/glia.23183. [DOI] [PubMed] [Google Scholar]
- 205.Rakhymzhan A, Acs A, Leben R, Winkler TH, Hauser AE, Niesner RA. Method for multiplexed dynamic intravital multiphoton imaging. Methods Mol Biol. 2021;2350:145–156. doi: 10.1007/978-1-0716-1593-5_10. [DOI] [PubMed] [Google Scholar]
- 206.Ransohoff RM. A polarizing question: do M1 and M2 microglia exist? Nat Neurosci. 2016a;19:987–991. doi: 10.1038/nn.4338. [DOI] [PubMed] [Google Scholar]
- 207.Ransohoff RM. How neuroinflammation contributes to neurodegeneration. Science. 2016b;353:777–783. doi: 10.1126/science.aag2590. [DOI] [PubMed] [Google Scholar]
- 208.Ricard C, Arroyo ED, He CX, Portera-Cailliau C, Lepousez G, Canepari M, Fiole D. Two-photon probes for in vivo multicolor microscopy of the structure and signals of brain cells. Brain Struct Funct. 2018;223:3011–3043. doi: 10.1007/s00429-018-1678-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Risher WC, Andrew RD, Kirov SA. Real-time passive volume responses of astrocytes to acute osmotic and ischemic stress in cortical slices and in vivo revealed by two-photon microscopy. Glia. 2009;57:207–221. doi: 10.1002/glia.20747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Risher WC, Croom D, Kirov SA. Persistent astroglial swelling accompanies rapid reversible dendritic injury during stroke-induced spreading depolarizations. Glia. 2012;60:1709–1720. doi: 10.1002/glia.22390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Robertson RT, Levine ST, Haynes SM, Gutierrez P, Baratta JL, Tan Z, Longmuir KJ. Use of labeled tomato lectin for imaging vasculature structures. Histochem Cell Biol. 2015;143:225–234. doi: 10.1007/s00418-014-1301-3. [DOI] [PubMed] [Google Scholar]
- 212.Ronaldson PT, Davis TP. Regulation of blood-brain barrier integrity by microglia in health and disease: A therapeutic opportunity. J Cereb Blood Flow Metab. 2020;40(1_suppl):S6–24. doi: 10.1177/0271678X20951995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Rosario AM, Cruz PE, Ceballos-Diaz C, Strickland MR, Siemienski Z, Pardo M, Schob KL, Li A, Aslanidi GV, Srivastava A, Golde TE, Chakrabarty P. Microglia-specific targeting by novel capsid-modified AAV6 vectors. Mol Ther Methods Clin Dev. 2016;3:16026. doi: 10.1038/mtm.2016.26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Rosenegger DG, Tran CH, Wamsteeker Cusulin JI, Gordon GR. Tonic local brain blood flow control by astrocytes independent of phasic neurovascular coupling. J Neurosci. 2015;35:13463–13474. doi: 10.1523/JNEUROSCI.1780-15.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Rovainen CM, Woolsey TA, Blocher NC, Wang DB, Robinson OF. Blood flow in single surface arterioles and venules on the mouse somatosensory cortex measured with videomicroscopy, fluorescent dextrans, nonoccluding fluorescent beads, and computer-assisted image analysis. J Cereb Blood Flow Metab. 1993;13:359–371. doi: 10.1038/jcbfm.1993.49. [DOI] [PubMed] [Google Scholar]
- 216.Ruan C, Sun L, Kroshilina A, Beckers L, De Jager P, Bradshaw EM, Hasson SA, Yang G, Elyaman W. A novel Tmem119-tdTomato reporter mouse model for studying microglia in the central nervous system. Brain Behav Immun. 2020;83:180–191. doi: 10.1016/j.bbi.2019.10.009. [DOI] [PubMed] [Google Scholar]
- 217.Santello M, Toni N, Volterra A. Astrocyte function from information processing to cognition and cognitive impairment. Nat Neurosci. 2019;22:154–166. doi: 10.1038/s41593-018-0325-8. [DOI] [PubMed] [Google Scholar]
- 218.Saunders NR, Dziegielewska KM, Møllgård K, Habgood MD. Markers for blood-brain barrier integrity: how appropriate is Evans blue in the twenty-first century and what are the alternatives? Front Neurosci. 2015;9:385. doi: 10.3389/fnins.2015.00385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Schaeffer S, Iadecola C. Revisiting the neurovascular unit. Nat Neurosci. 2021;24:1198–1209. doi: 10.1038/s41593-021-00904-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Schaffer CB, Friedman B, Nishimura N, Schroeder LF, Tsai PS, Ebner FF, Lyden PD, Kleinfeld D. Two-photon imaging of cortical surface microvessels reveals a robust redistribution in blood flow after vascular occlusion. PLoS Biol. 2006;4:e22. doi: 10.1371/journal.pbio.0040022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Schrandt CJ, Kazmi SM, Jones TA, Dunn AK. Chronic monitoring of vascular progression after ischemic stroke using multiexposure speckle imaging and two-photon fluorescence microscopy. J Cereb Blood Flow Metab. 2015;35:933–942. doi: 10.1038/jcbfm.2015.26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Seifert S, Pannell M, Uckert W, Färber K, Kettenmann H. Transmitter- and hormone-activated Ca(2+) responses in adult microglia/brain macrophages in situ recorded after viral transduction of a recombinant Ca(2+) sensor. Cell Calcium. 2011;49:365–375. doi: 10.1016/j.ceca.2011.03.005. [DOI] [PubMed] [Google Scholar]
- 223.Shen XY, Gao ZK, Han Y, Yuan M, Guo YS, Bi X. Activation and role of astrocytes in ischemic stroke. Front Cell Neurosci. 2021;15:755955. doi: 10.3389/fncel.2021.755955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Shi LG, Rocha M, Leak RK, Zhao JY, Bhatia TN, Mu HF, Wei ZS, Yu F, Weiner SL, Ma FF, Jovin TG, Chen J. A new era for stroke therapy: Integrating neurovascular protection with optimal reperfusion. J Cereb Blood Flow Metab. 2018;38:2073–2091. doi: 10.1177/0271678X18798162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Shih AY, Friedman B, Drew PJ, Tsai PS, Lyden PD, Kleinfeld D. Active dilation of penetrating arterioles restores red blood cell flux to penumbral neocortex after focal stroke. J Cereb Blood Flow Metab. 2009;29:738–751. doi: 10.1038/jcbfm.2008.166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Shinotsuka T, Yasui M, Nuriya M. Astrocytic gap junctional networks suppress cellular damage in an in vitro model of ischemia. Biochem Biophys Res Commun. 2014;444:171–176. doi: 10.1016/j.bbrc.2014.01.035. [DOI] [PubMed] [Google Scholar]
- 227.Shuaib A, Butcher K, Mohammad AA, Saqqur M, Liebeskind DS. Collateral blood vessels in acute ischaemic stroke: a potential therapeutic target. Lancet Neurol. 2011;10:909–921. doi: 10.1016/S1474-4422(11)70195-8. [DOI] [PubMed] [Google Scholar]
- 228.Sierra A, Paolicelli RC, Kettenmann H. Cien Años de microglía: milestones in a century of microglial research. Trends Neurosci. 2019;42:778–792. doi: 10.1016/j.tins.2019.09.004. [DOI] [PubMed] [Google Scholar]
- 229.Sigler A, Murphy TH. In vivo 2-photon imaging of fine structure in the rodent brain: before, during, and after stroke. Stroke. 2010;41:S117–123. doi: 10.1161/STROKEAHA.110.594648. [DOI] [PubMed] [Google Scholar]
- 230.Silvin A, Uderhardt S, Piot C, Da Mesquita S, Yang K, Geirsdottir L, Mulder K, Eyal D, Liu Z, Bridlance C, Thion MS, Zhang XM, Kong WT, Deloger M, Fontes V, Weiner A, Ee R, Dress R, Hang JW, Balachander A, et al. Dual ontogeny of disease-associated microglia and disease inflammatory macrophages in aging and neurodegeneration. Immunity. 2022;55:1448–1465.e6. doi: 10.1016/j.immuni.2022.07.004. [DOI] [PubMed] [Google Scholar]
- 231.Siracusa R, Fusco R, Cuzzocrea S. Astrocytes: role and functions in brain pathologies. Front Pharmacol. 2019;10:1114. doi: 10.3389/fphar.2019.01114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Smith AJ, Yao X, Dix JA, Jin BJ, Verkman AS. Test of the 'glymphatic'hypothesis demonstrates diffusive and aquaporin-4-independent solute transport in rodent brain parenchyma. Elife. 2017;6:e27679. doi: 10.7554/eLife.27679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Sommer CJ. Ischemic stroke: experimental models and reality. Acta Neuropathol. 2017;133:245–261. doi: 10.1007/s00401-017-1667-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Song YY, Li ZW, He TT, Qu MJ, Jiang L, Li WL, Shi XJ, Pan JJ, Zhang LY, Wang YT, Zhang ZJ, Tang YH, Yang GY. M2 microglia-derived exosomes protect the mouse brain from ischemia-reperfusion injury via exosomal miR-124. Theranostics. 2019;9:2910–2923. doi: 10.7150/thno.30879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.Spector R, Robert Snodgrass S, Johanson CE. A balanced view of the cerebrospinal fluid composition and functions: Focus on adult humans. Exp Neurol. 2015;273:57–68. doi: 10.1016/j.expneurol.2015.07.027. [DOI] [PubMed] [Google Scholar]
- 236.Srinivasan R, Lu TY, Chai H, Xu J, Huang BS, Golshani P, Coppola G, Khakh BS. New transgenic mouse lines for selectively targeting astrocytes and studying calcium signals in astrocyte processes in situ and in vivo. Neuron. 2016;92:1181–1195. doi: 10.1016/j.neuron.2016.11.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Stobart JL, Ferrari KD, Barrett MJP, Gluck C, Stobart MJ, Zuend M, Weber B. Cortical circuit activity evokes rapid astrocyte calcium signals on a similar timescale to neurons. Neuron. 2018;98:726–735.e4. doi: 10.1016/j.neuron.2018.03.050. [DOI] [PubMed] [Google Scholar]
- 238.Stratoulias V, Venero JL, Tremblay ME, Joseph B. Microglial subtypes: diversity within the microglial community. EMBO J. 2019;38:e101997. doi: 10.15252/embj.2019101997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 239.Stuckey SM, Ong LK, Collins-Praino LE, Turner RJ. Neuroinflammation as a key driver of secondary neurodegeneration following stroke? Int J Mol Sci. 2021;22:13101. doi: 10.3390/ijms222313101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.Sun LL, Zhang YX, Liu E, Ma QY, Anatol M, Han HB, Yan JH. The roles of astrocyte in the brain pathologies following ischemic stroke. Brain Injury. 2019;33:712–716. doi: 10.1080/02699052.2018.1531311. [DOI] [PubMed] [Google Scholar]
- 241.Sun M, Chen X, Yin YX, Gao Y, Zhang L, Chen B, Ji Y, Fukunaga K, Han F, Lu YM. Role of pericyte-derived SENP1 in neuronal injury after brain ischemia. CNS Neurosci Ther. 2020;26:815–828. doi: 10.1111/cns.13398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242.Sun MS, Jin H, Sun X, Huang S, Zhang FL, Guo ZN, Yang Y. Free radical damage in ischemia-reperfusion injury: an obstacle in acute ischemic stroke after revascularization therapy. Oxid Med Cell Longev. 2018;2018:3804979. doi: 10.1155/2018/3804979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.Sun Q, Xu X, Wang T, Xu Z, Lu X, Li X, Chen G. Neurovascular units and neural-glia networks in intracerebral hemorrhage: from mechanisms to translation. Transl Stroke Res. 2021;12:447–460. doi: 10.1007/s12975-021-00897-2. [DOI] [PubMed] [Google Scholar]
- 244.Surugiu R, Catalin B, Dumbrava D, Gresita A, Olaru DG, Hermann DM, Popa-Wagner A. Intracortical administration of the complement C3 receptor antagonist trifluoroacetate modulates microglia reaction after brain injury. Neural Plast. 20192019:1071036. doi: 10.1155/2019/1071036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Sweeney MD, Zhao Z, Montagne A, Nelson AR, Zlokovic BV. Blood-brain barrier: from physiology to disease and back. Physiol Rev. 2019;99:21–78. doi: 10.1152/physrev.00050.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246.Szalay G, Martinecz B, Lénárt N, Környei Z, Orsolits B, Judák L, Császár E, Fekete R, West BL, Katona G, Rózsa B, Dénes Á. Microglia protect against brain injury and their selective elimination dysregulates neuronal network activity after stroke. Nat Commun. 2016;7:11499. doi: 10.1038/ncomms11499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Takano T, Tian GF, Peng W, Lou N, Libionka W, Han X, Nedergaard M. Astrocyte-mediated control of cerebral blood flow. Nat Neurosci. 2006;9:260–267. doi: 10.1038/nn1623. [DOI] [PubMed] [Google Scholar]
- 248.Takano T, Oberheim N, Cotrina ML, Nedergaard M. Astrocytes and ischemic injury. Stroke. 2009;40:S8–12. doi: 10.1161/STROKEAHA.108.533166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Tang YJ, Li L, Sun LQ, Yu JS, Hu Z, Lian KQ, Cao G, Dai JX. In vivo two-photon calcium imaging in dendrites of rabies virus-labeled V1 corticothalamic neurons. Neurosci Bull. 2020;36:545–553. doi: 10.1007/s12264-019-00452-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250.Theer P, Hasan MT, Denk W. Two-photon imaging to a depth of 1000 microm in living brains by use of a Ti: Al2O3 regenerative amplifier. Opt Lett. 2003;28:1022–1024. doi: 10.1364/ol.28.001022. [DOI] [PubMed] [Google Scholar]
- 251.Tong L, Hill RA, Damisah EC, Murray KN, Yuan P, Bordey A, Grutzendler J. Imaging and optogenetic modulation of vascular mural cells in the live brain. Nat Protoc. 2021;16:472–496. doi: 10.1038/s41596-020-00425-w. [DOI] [PubMed] [Google Scholar]
- 252.Tóth L, Szöllősi D, Kis-Petik K, Adorján I, Erdélyi F, Kálmán M. The first postlesion minutes: an in vivo study of extravasation and perivascular astrocytes following cerebral lesions in various experimental mouse models. J Histochem Cytochem. 2019;67:29–39. doi: 10.1369/0022155418788390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Trachtenberg JT, Chen BE, Knott GW, Feng G, Sanes JR, Welker E, Svoboda K. Long-term in vivo imaging of experience-dependent synaptic plasticity in adult cortex. Nature. 2002;420:788–794. doi: 10.1038/nature01273. [DOI] [PubMed] [Google Scholar]
- 254.Turc G, Bhogal P, Fischer U, Khatri P, Lobotesis K, Mazighi M, Schellinger PD, Toni D, de Vries J, White P, Fiehler J. European Stroke Organisation (ESO)-European Society for Minimally Invasive Neurological Therapy (ESMINT) guidelines on mechanical thrombectomy in acute ischemic stroke. J Neurointerv Surg. 2019;11:535–538. doi: 10.1136/neurintsurg-2018-014568. [DOI] [PubMed] [Google Scholar]
- 255.Tvrdik P, Kearns KN, Sharifi KA, Sluzewski MF, Acton ST, Kalani MYS. Calcium Imaging of Microglial Network Activity in Stroke. Methods Mol Biol. 2019;2034:267–279. doi: 10.1007/978-1-4939-9658-2_19. [DOI] [PubMed] [Google Scholar]
- 256.Ueki H, Wang IH, Zhao D, Gunzer M, Kawaoka Y. Multicolor two-photon imaging of in vivo cellular pathophysiology upon influenza virus infection using the two-photon IMPRESS. Nat Protoc. 2020;15:1041–1065. doi: 10.1038/s41596-019-0275-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 257.Underly RG, Levy M, Hartmann DA, Grant RI, Watson AN, Shih AY. Pericytes as inducers of rapid, matrix metalloproteinase-9-dependent capillary damage during ischemia. J Neurosci. 2017;37:129–140. doi: 10.1523/JNEUROSCI.2891-16.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258.Underly RG, Shih AY. Rapid, nitric oxide synthesis-dependent activation of MMP-9 at pericyte somata during capillary ischemia in vivo. Front Physiol. 2020;11:619230. doi: 10.3389/fphys.2020.619230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 259.Vagal A, Aviv R, Sucharew H, Reddy M, Hou QH, Michel P, Jovin T, Tomsick T, Wintermark M, Khatri P. Collateral clock is more important than time clock for tissue fate a natural history study of acute ischemic strokes. Stroke. 2018;49:2102–2107. doi: 10.1161/STROKEAHA.118.021484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 260.Valkonen K, Martinez-Majander N, Sibolt G, Tiainen M, Raty S, Kokkonen T, Lappalainen K, Curtze S. Recanalization treatment for acute ischemic stroke:3-month outcome before and after implementation of routine endovascular treatment. eNeurologicalSci. 2022;27:100394. doi: 10.1016/j.ensci.2022.100394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 261.Vanlandewijck M, He LQ, Mae MAA, Andrae J, Ando K, Del Gaudio F, Nahar K, Lebouvier T, Lavina B, Gouveia L, Sun Y, Raschperger E, Rasanen M, Zarb Y, Mochizuki N, Keller A, Lendahl U, Betsholtz C. A molecular atlas of cell types and zonation in the brain vasculature. Nature. 2018;554:475–480. doi: 10.1038/nature25739. [DOI] [PubMed] [Google Scholar]
- 262.Vérant P, Ricard C, Serduc R, Vial JC, van der Sanden B. In vivo staining of neocortical astrocytes via the cerebral microcirculation using sulforhodamine B. J Biomed Opt. 2008a;13:064028. doi: 10.1117/1.3041163. [DOI] [PubMed] [Google Scholar]
- 263.Vérant P, Serduc R, van der Sanden B, Chantal R, Ricard C, Coles JA, Vial JC. Subtraction method for intravital two-photon microscopy: intraparenchymal imaging and quantification of extravasation in mouse brain cortex. J Biomed Opt. 2008b;13:011002. doi: 10.1117/1.2870083. [DOI] [PubMed] [Google Scholar]
- 264.Verkhratsky A, Nedergaard M. Physiology of astroglia. Physiol Rev. 2018;98:239–389. doi: 10.1152/physrev.00042.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 265.Vernieri F, Pasqualetti P, Matteis M, Passarelli F, Troisi E, Rossini PM, Caltagirone C, Silvestrini M. Effect of collateral blood flew and cerebral vasomotor reactivity on the outcome of carotid artery occlusion. Stroke. 2001;32:1552–1558. doi: 10.1161/01.str.32.7.1552. [DOI] [PubMed] [Google Scholar]
- 266.Volterra A, Meldolesi J. Astrocytes, from brain glue to communication elements: The revolution continues. Nat Rev Neurosci. 2005;6:626–640. doi: 10.1038/nrn1722. [DOI] [PubMed] [Google Scholar]
- 267.Wake H, Moorhouse AJ, Jinno S, Kohsaka S, Nabekura J. Resting microglia directly monitor the functional state of synapses in vivo and determine the fate of ischemic terminals. J Neurosci. 2009;29:3974–3980. doi: 10.1523/JNEUROSCI.4363-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 268.Walter HL, van der Maten G, Antunes AR, Wieloch T, Ruscher K. Treatment with AMD3100 attenuates the microglial response and improves outcome after experimental stroke. J Neuroinflammation. 2015;12:24. doi: 10.1186/s12974-014-0232-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 269.Wang S, Liu J, Goh CC, Ng LG, Liu B. NIR-II-excited intravital two-photon microscopy distinguishes deep cerebral and tumor vasculatures with an ultrabright NIR-I AIE luminogen. Adv Mater. 2019;31:e1904447. doi: 10.1002/adma.201904447. [DOI] [PubMed] [Google Scholar]
- 270.Wendling O, Bornert JM, Chambon P, Metzger D. Efficient temporally-controlled targeted mutagenesis in smooth muscle cells of the adult mouse. Genesis. 2009;47:14–18. doi: 10.1002/dvg.20448. [DOI] [PubMed] [Google Scholar]
- 271.Wicks EE, Ran KR, Kim JE, Xu R, Lee RP, Jackson CM. The translational potential of microglia and monocyte-derived macrophages in ischemic stroke. Front Immunol. 2022;13:897022. doi: 10.3389/fimmu.2022.897022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 272.Wieghofer P, Prinz M. Genetic manipulation of microglia during brain development and disease. Biochim Biophys Acta. 2016;1862:299–309. doi: 10.1016/j.bbadis.2015.09.019. [DOI] [PubMed] [Google Scholar]
- 273.Williamson MR, Franzen RL, Fuertes CJA, Dunn AK, Drew MR, Jones TA. A window of vascular plasticity coupled to behavioral recovery after stroke. J Neurosci. 2020;40:7651–7667. doi: 10.1523/JNEUROSCI.1464-20.2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 274.Williamson MR, Fuertes CJA, Dunn AK, Drew MR, Jones TA. Reactive astrocytes facilitate vascular repair and remodeling after stroke. Cell Rep. 2021;35:109048. doi: 10.1016/j.celrep.2021.109048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 275.Wollenweber FA, Tiedt S, Alegiani A, Alber B, Bangard C, Berrouschot J, Bode FJ, Boeckh-Behrens T, Bohner G, Bormann A, Braun M, Dorn F, Eckert B, Flottmann F, Hamann GF, Henn KH, Herzberg M, Kastrup A, Kellert L, Kraemer C, et al. Functional outcome following stroke thrombectomy in clinical practice. Stroke. 2019;50:2500–2506. doi: 10.1161/STROKEAHA.119.026005. [DOI] [PubMed] [Google Scholar]
- 276.Wu T, Liao J, Yu J, Gao Y, Li H, Wu J, Xia X, Shi K, Zheng W. In vivo label-free two-photon excitation autofluorescence microscopy of microvasculature using a 520 nm femtosecond fiber laser. Opt Lett. 2020;45:2704–2707. doi: 10.1364/OL.394242. [DOI] [PubMed] [Google Scholar]
- 277.Xie L, Kang H, Xu Q, Chen MJ, Liao Y, Thiyagarajan M, O'Donnell J, Christensen DJ, Nicholson C, Iliff JJ, Takano T, Deane R, Nedergaard M. Sleep drives metabolite clearance from the adult brain. Science. 2013;342:373–377. doi: 10.1126/science.1241224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 278.Xu HT, Pan F, Yang G, Gan WB. Choice of cranial window type for in vivo imaging affects dendritic spine turnover in the cortex. Nat Neurosci. 2007;10:549–551. doi: 10.1038/nn1883. [DOI] [PubMed] [Google Scholar]
- 279.Xu SB, Lu JN, Shao AW, Zhang JH, Zhang JM. Glial cells: role of the immune response in ischemic stroke. Front Immunol. 2020;11:294. doi: 10.3389/fimmu.2020.00294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 280.Yan W, Huang Y, Wang L, Li J, Guo Y, Yang Z, Qu J. Multi-color two-photon microscopic imaging based on a single-wavelength excitation. Biosensors (Basel) 2022;12:307. doi: 10.3390/bios12050307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 281.Yang CJ, Hawkins KE, Dore S, Candelario-Jalil E. Neuroinflammatory mechanisms of blood-brain barrier damage in ischemic stroke. Am J Physiol Cell Physiol. 2019;316:C135–153. doi: 10.1152/ajpcell.00136.2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 282.Yang G, Pan F, Parkhurst CN, Grutzendler J, Gan WB. Thinned-skull cranial window technique for long-term imaging of the cortex in live mice. Nat Protoc. 2010;5:201–208. doi: 10.1038/nprot.2009.222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 283.Yang T, Dai YJ, Chen G, Cui SS. Dissecting the dual role of the glial scar and scar-forming astrocytes in spinal cord injury. Front Cell Neurosci. 2020;14:78. doi: 10.3389/fncel.2020.00078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 284.Yang WJ, Carrillo-Reid L, Bando Y, Peterka DS, Yuste R. Simultaneous two-photon imaging and two-photon optogenetics of cortical circuits in three dimensions. Elife. 2018;7:e32671. doi: 10.7554/eLife.32671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 285.Yao X, Derugin N, Manley GT, Verkman AS. Reduced brain edema and infarct volume in aquaporin-4 deficient mice after transient focal cerebral ischemia. Neurosci Lett. 2015;584:368–372. doi: 10.1016/j.neulet.2014.10.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 286.Ye ZJ, Wei L, Geng X, Wang X, Li ZH, Xiao LH. Mitochondrion-specific blinking fluorescent bioprobe for nanoscopic monitoring of mitophagy. ACS Nano. 2019;13:11593–11602. doi: 10.1021/acsnano.9b05354. [DOI] [PubMed] [Google Scholar]
- 287.Yoon JH, Lee ES, Jeong Y. In vivo imaging of the cerebral endothelial glycocalyx in mice. J Vasc Res. 2017;54:59–67. doi: 10.1159/000457799. [DOI] [PubMed] [Google Scholar]
- 288.Yoon S, Lee H, Hong JH, Lim YS, Choi W. Laser scanning reflection-matrix microscopy for aberration-free imaging through intact mouse skull. Nat Commun. 2020;11:5721. doi: 10.1038/s41467-020-19550-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 289.Zamanian JL, Xu L, Foo LC, Nouri N, Zhou L, Giffard RG, Barres BA. Genomic analysis of reactive astrogliosis. J Neurosci. 2012;32:6391–6410. doi: 10.1523/JNEUROSCI.6221-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 290.Zambach SA, Cai C, Helms HCC, Hald BO, Dong Y, Fordsmann JC, Nielsen RM, Hu J, Lønstrup M, Brodin B, Lauritzen MJ. Precapillary sphincters and pericytes at first-order capillaries as key regulators for brain capillary perfusion. Proc Natl Acad Sci U S A. 2021;118:e2023749118. doi: 10.1073/pnas.2023749118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 291.Zhang S, Murphy TH. Imaging the impact of cortical microcirculation on synaptic structure and sensory-evoked hemodynamic responses in vivo. PLoS Biol. 2007;5:e119. doi: 10.1371/journal.pbio.0050119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 292.Zhang S. Microglial activation after ischaemic stroke. Stroke Vasc Neurol. 2019;4:71–74. doi: 10.1136/svn-2018-000196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 293.Zhang ZH, Russell LE, Packer AM, Gauld OM, Hausser M. Closed-loop all-optical interrogation of neural circuits in vivo. Nat Methods. 2018;15:1037–1040. doi: 10.1038/s41592-018-0183-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 294.Zhao SJ, Wu SL, Jia QY, Huang L, Lan MH, Wang PF, Zhang WJ. Lysosome-targetable carbon dots for highly efficient photothermal/photodynamic synergistic cancer therapy and photoacoustic/two-photon excited fluorescence imaging. Chem Eng J. 2020 doi:10.1016/j.cej.2020.124212. [Google Scholar]
- 295.Zhao YJ, Yu TT, Zhang C, Li Z, Luo QM, Xu TH, Zhu D. Skull optical clearing window for in vivo imaging of the mouse cortex at synaptic resolution. Light Sci Appl 7:17153. Erratum in: Light Sci Appl. 2018;2018;7:6. doi: 10.1038/lsa.2017.153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 296.Zhou YX, Shao AW, Yao YH, Tu S, Deng YC, Zhang JM. Dual roles of astrocytes in plasticity and reconstruction after traumatic brain injury. Cell Commun Signal. 2020;18:62. doi: 10.1186/s12964-020-00549-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 297.Zong W, Wu R, Chen S, Wu J, Wang H, Zhao Z, Chen G, Tu R, Wu D, Hu Y, Xu Y, Wang Y, Duan Z, Wu H, Zhang Y, Zhang J, Wang A, Chen L, Cheng H. Miniature two-photon microscopy for enlarged field-of-view, multi-plane and long-term brain imaging. Nat Methods. 2021;18:46–49. doi: 10.1038/s41592-020-01024-z. [DOI] [PubMed] [Google Scholar]
- 298.Zonta M, Angulo MC, Gobbo S, Rosengarten B, Hossmann KA, Pozzan T, Carmignoto G. Neuron-to-astrocyte signaling is central to the dynamic control of brain microcirculation. Nat Neurosci. 2003;6:43–50. doi: 10.1038/nn980. [DOI] [PubMed] [Google Scholar]
- 299.Zou Z, Dong YS, Liu DD, Li G, Hao GZ, Gao X, Pan PY, Liang GB. MAP4K4 induces early blood-brain barrier damage in a murine subarachnoid hemorrhage model. Neural Regen Res. 2021;16:325–332. doi: 10.4103/1673-5374.290904. [DOI] [PMC free article] [PubMed] [Google Scholar]