

Abstract
Since the development of Sanger sequencing in 1977, sequencing technology has played a pivotal role in molecular biology research by enabling the interpretation of biological genetic codes. Today, nanopore sequencing is one of the leading third‐generation sequencing technologies. With its long reads, portability, and low cost, nanopore sequencing is widely used in various scientific fields including epidemic prevention and control, disease diagnosis, and animal and plant breeding. Despite initial concerns about high error rates, continuous innovation in sequencing platforms and algorithm analysis technology has effectively addressed its accuracy. During the coronavirus disease (COVID‐19) pandemic, nanopore sequencing played a critical role in detecting the severe acute respiratory syndrome coronavirus‐2 virus genome and containing the pandemic. However, a lack of understanding of this technology may limit its popularization and application. Nanopore sequencing is poised to become the mainstream choice for preventing and controlling COVID‐19 and future epidemics while creating value in other fields such as oncology and botany. This work introduces the contributions of nanopore sequencing during the COVID‐19 pandemic to promote public understanding and its use in emerging outbreaks worldwide. We discuss its application in microbial detection, cancer genomes, and plant genomes and summarize strategies to improve its accuracy.
Keywords: nanopore sequencing, SARS‐CoV‐2, COVID‐19, cancer, plant, genome, mutation, pandemic
Nanopore sequencing has been instrumental in managing the COVID‐19 pandemic. This review aims to increase public understanding of its role in identifying SARS‐CoV‐2 variants and tracking changes in regional epidemiology. Additionally, the review explores the use of nanopore sequencing in detecting other microbial outbreaks, as well as its applications in cancer and plant genomics.

1. INTRODUCTION
The concept of nanopore sequencing, where single‐stranded nucleic acids pass through a nanopore in a membrane under an electric field, was first proposed by David Deamer in the 1980s. 1 Despite initial skepticism, technological advances eventually made nanopore sequencing a reality. 2 Proteins such as α‐hemolysin from Staphylococcus aureus 3 , 4 , 5 and Mycobacterium smegmatis porin A (MspA) 6 , 7 were shown to distinguish the four bases on single‐stranded nucleotide molecules. The use of phi29 DNA polymerase slowed down the translocation of nucleic acid molecules through the nanopore, improving the signal‐to‐noise ratio. 8 , 9 Oxford Nanopore Technologies (ONT), founded in 2005 by Oxford professor Bayley and colleagues, 10 facilitated the commercialization of nanopore sequencing with the release of their MinION sequencer in 2014. 2
Currently, ONT has established a complete sequencing system process that includes advanced library preparation techniques combined with amplicon and other technologies, as well as numerous bioinformatics methods for analyzing and mining nanopore sequencing data. 2
Since 2019, COVID‐19 caused by severe acute respiratory syndrome coronavirus (SARS‐CoV‐2) has spread to over 200 countries worldwide. 11 As an RNA virus, SARS‐CoV‐2 continuously mutates during transmission, and the emergence of SARS‐CoV‐2 variants poses challenges for epidemic control. 12 Obtaining a complete SARS‐CoV‐2 genome is crucial for detecting mutations because sequence changes can reduce the sensitivity of SARS‐CoV‐2 detection techniques. 13 The first genome sequence of SARS‐CoV‐2 was obtained through metagenomic sequencing. 14 Nanopore sequencing has played a significant role in the COVID‐19 pandemic. 15 There is no doubt that nanopore sequencing has broad prospects, as demonstrated during the COVID‐19 pandemic. The portability, high efficiency, and low cost of nanopore sequencing make it particularly well suited for dynamically monitoring SARS‐CoV‐2 mutations and the spread of the COVID‐19 pandemic in different countries and regions. Currently, nanopore sequencing has been used for diagnostic sequencing of SARS‐CoV‐2, 16 genome sequencing, 17 and related research 18 of SARS‐CoV‐2. In the field of pathogenic microorganism detection, nanopore sequencing has been used not only for detecting SARS‐CoV‐2 but also for identifying, typing, and monitoring the transmission of newly emerging monkeypox virus (MPXV) 19 and norovirus (NoV). 20 Nanopore sequencing can also be used to detect microbial communities in human tissues such as the skin and intestinal tract, 21 , 22 as well as in environmental samples. 23 Its long single‐molecule long‐read sequencing capabilities make it widely used in cancer research 24 , 25 and clinical diagnosis and treatment. 26 , 27 Additionally, nanopore sequencing has unique value in resolving high‐quality plant genomes. 28 However, public concerns about the error rate of nanopore sequencing still greatly limit its promotion and use. In reality, nanopore sequencing is a very cost‐effective technology, 29 especially for underdeveloped regions. With the rapid iteration of ONT's sequencing platform and the development of new base‐calling algorithms, the accuracy of nanopore sequencing has greatly improved. Furthermore, its application during the COVID‐19 pandemic has proven that data obtained through nanopore sequencing is reliable. 30
In this review, we have detailed the specific advantages of nanopore sequencing technology and its contributions during the COVID‐19 pandemic, including rapid identification of SARS‐CoV‐2, genotyping, dynamic transmission monitoring, and elucidation of related mechanisms of SARS‐CoV‐2. This work introduces the use of nanopore sequencing for detecting pathogenic microorganisms other than SARS‐CoV‐2, including viruses and bacteria. It also covers the application of nanopore sequencing in cancer research and clinical practice, as well as in plant genomics. Finally, we have analyzed the reasons for the low accuracy of nanopore sequencing and strategies for improving it. By introducing the contributions of nanopore sequencing during the COVID‐19 pandemic and its potential in other fields, we hope to promote the widespread application of next‐generation sequencing (NGS) technology and contribute to controlling new pandemics, including COVID‐19 or possible future ones.
2. MOLECULAR MECHANISMS OF NANOPORE SEQUENCING
2.1. Principle and specific process
During nanopore sequencing, nanopores act as biosensors. 1 A certain number of nanopores are fixed on a resistive film, providing the only channel between both sides of the film. The ion solution on the cis side of the resistive film contacts the ion solution on the trans side through the nanopore. 1 Electrodes are set at both ends of the nanopore sequencer to form a stable electric field. Nucleic acids (including DNA and RNA) will move to the nanopores under the action of an electric field in the nanopore sequencer. 31 , 32 , 33 , 34 The process of nucleic acid molecules passing through the nanopores is pulled by motor proteins, which can control the speed of nucleic acid passing through the nanopore. 2 During the passage of nucleic acid through the nanopore, the charge of the nanopore changes, which in turn leads to a change in the flow of electrons on the resistive membrane. The electron flow caused by different bases or the quality of the modified molecular structure has unique characteristics. 2 This electronic signal is captured and recorded by the nanopore sequencer, which is then passed through an algorithmic program to determine the base type and obtain the sequencing result 2 (Figure 1).
FIGURE 1.

Schematic diagram of the basic principle and process of nanopore sequencing for SARS‐CoV‐2 detection. After the SARS‐CoV‐2 nucleic acid samples are collected, they can be added to the nanopore sequencer after simple library preparation, and the sequencing results can be obtained in real‐time (A). Motor proteins pull the nucleic acid through the nanopore, and the nucleic acid is detected and transmitted to the computer through the generated tiny current signal (B). The computer recognizes the base type by analyzing the current characteristic signal and converts the current signal into the base sequence (C).
2.2. Key components
The core of nanopore sequencing is the nanopore. 2 , 35 The nanopores used in initial experiments were derived from Staphylococcus aureus α‐hemolysin, a heptamer consisting of 14 antiparallel β‐strands with a diameter of approximately 2.6 nm. 36 Other biological nanopores with similar activities can also be used for nanopore sequencing. For example, the protein Aerolysin, a pore‐forming toxin secreted by Aeromonas hydrophila, 37 has a diameter of 1.0–1.7 nm and has been shown to enable sensitive single‐base–pair discrimination. 38 The outer membrane protein G of Escherichia coli is also a potential candidate, but its open and closed conformational states make it currently unsuitable for single‐molecule sensing. 39
MspA is currently the main choice for ONT's nanopore sequencing equipment. MspA is an octamer with a minimum inner diameter of 1 nm, which is narrower and more stable than the channel of α‐hemolysin, allowing it to have higher single‐nucleotide resolution. 40 The difference in ionic currents of the four bases through the MspA nanopore is significantly greater than that of α‐hemolysin, allowing for more distinct distinction between the four bases. 41 MspA also outperforms α‐hemolysin under extreme conditions, allowing it to adapt to harsh environments in emergencies. 42
Motor proteins are critical for regulating the rate of nucleic acid translocation. Translocation of nucleic acids too fast will increase the difficulty of clip identification. In early experiments in the development of nanopore sequencing, single‐stranded DNA passed through the nanopore at a rate of 1–10 bases per millisecond, making it difficult to detect the sequencing signal. 10 The unstable translocation rate also brings difficulties to subsequent data analysis. The phi29 DNA polymerase derived from bacteriophage was used as a molecular ratcheting system. 37 Pairing MspA with phi29 DNAP slows DNA translocation for single‐base discrimination. 37
2.3. Advantages and disadvantages brought by the sequencing mechanism
Nanopore sequencing's single‐molecule direct sequencing enables it to read ultra‐long DNA and RNA molecules in a timely manner, which cannot be achieved by first‐ and second‐generation sequencing. However, its disadvantages include a higher error rate (compared with previous generations of sequencing) and higher sample quality requirements. Despite this, nanopore sequencing is widely used in genome detection in fields such as microorganisms, tumors, animals and plants, especially during the COVID‐19 pandemic. Its advantages continue to emerge in various fields while its shortcomings become less obvious with continuous equipment updates, technology alliances, and analysis technology updates.
3. APPLICATION OF NANOPORE SEQUENCING TECHNOLOGY DURING THE COVID‐19 OUTBREAK
3.1. Detection of SARS‐CoV‐2 infection and the advantages of nanopore sequencing
3.1.1. SARS‐CoV‐2 infection detection technology
SARS‐CoV‐2 tests can be classified into two types: nucleic acid‐based and protein‐based tests 43 (Table 1). The primary methods for detecting SARS‐CoV‐2 nucleic acid are reverse transcription.
TABLE 1.
Main detection methods of SARS‐CoV‐2.
| Detection methods | Target | Advantages | Limitations | References | |
|---|---|---|---|---|---|
| Molecular tests | RT‐PCR | Nucleic acid | The gold standard for virus detection; high sensitivity; suitable for large‐scale screening | Requires thermal cycling; prone to false negative results; time consuming | 44 , 45 |
| RT‐LAMP | Nucleic acid | Amplifies DNA under isothermal conditions; fast; high magnification; colorimetric measurement with strong visibility | Requires specific primers | 46 , 47 | |
| Sequencing | Nucleic acid | High sensitivity; can specifically identify and differentiate a variety of coronaviruses and pathogenic microorganisms | Complicated operation; expensive | 48 | |
| Serological tests | ELISA | Antibody | High sensitivity; qualitative or quantitative; ability to test multiple samples; low sample collection requirements | Not suitable for diagnosing acute infections; difficult to apply to mass screening | 49 , 50 |
| RDT | Antigen or antibody | Field test; simple and fast | Lack of high‐quality assessment; only qualitative analysis; questionable reliability of diagnostic results, use for research only. | 51 , 52 | |
| Neutralization test | Antibody | Facilitate vaccine development | It needs to be carried out in a laboratory with a higher biosafety level | 53 | |
Current methods for detecting SARS‐CoV‐2 are mainly divided into two categories: nucleic acid‐based molecular tests and antigen‐ or antibody‐based serological tests. Molecular tests are divided into RT‐PCR, RT‐LAMP, and sequencing, and serological tests are divided into ELISA, RDT, and neutralization tests. RT‐PCR, reverse transcriptase‐polymerase chain reaction; RT‐LAMP, reverse transcription loop‐mediated isothermal amplification; ELISA, enzyme‐linked immunosorbent assay; RDT, rapid diagnostic test.
Polymerase chain reaction (RT‐PCR), 54 reverse transcription loop‐mediated isothermal amplification (RT‐LAMP), and sequencing for diagnosis. 43 RT‐PCR is the most commonly used method for detecting SARS‐CoV‐2 nucleic acid. 55 It involves extracting viral RNA and converting it into complementary DNA (cDNA) using reverse transcriptase. 56 , 57 The cDNA is then used as a template to amplify the target gene using specific primers designed according to the SARS‐CoV‐2 genome and fluorescently labeled probes, 58 allowing for the detection of SARS‐CoV‐2. 55 While RT‐PCR is considered the “gold standard” for COVID‐19 diagnosis, 59 it relies on the equipment and expertise of the testing center and the transportation time of samples can increase its time cost. 60 Additionally, its higher false negative rate can pose challenges in managing the COVID‐19 pandemic. 43 RT‐LAMP is an alternative technique that rapidly amplifies DNA through strand displacement by DNA polymerase after the initial synthesis of a dumbbell‐shaped DNA template. 61 As a substitute for RT‐PCR, 60 LAMP 62 has low requirements for specialized equipment and provides fast diagnosis, 63 making it one of the mainstream technologies for SARS‐CoV‐2 detection. 64 However, its limited detection sensitivity restricts its widespread application. 65 Sequencing for diagnosis involves obtaining the whole genome information of SARS‐CoV‐2 through metagenomic sequencing, 66 , 67 , 68 including mainstream NGS Illumina and nanopore sequencing. 69 , 70 Its advantage is its high specificity and ability to simultaneously identify other respiratory viruses, compensating for the high false positives of RT‐PCR. 16
Additionally, new molecular detection technologies are being developed or are expected to be used for SARS‐CoV‐2 nucleic acid detection. For instance, researchers have established a nucleic acid detection platform called specific high‐sensitivity enzyme reporter unlocking (SHERLOCK) by combining CRISPR–Cas enzymology with nucleic acid preamplification. 71 This platform can be used for nucleic acid detection of Zika virus and dengue virus (DENV) 72 , 73 and also shows potential for SARS‐CoV‐2 nucleic acid detection. 74 , 75 Other technologies such as 3D‐printing 76 and micro/nanorobots 77 can also be used for SARS‐CoV‐2 detection. Researchers have created a disposable gene‐sensor using 3D pen‐printed electrodes (3D‐PP), 78 where hybridization of ssDNA targeting the N gene of SARS‐CoV‐2 in the sensor to the SARS‐CoV‐2 RNA target affects the sensor signal. 79 Plasmonic‐magnetic nanorobots 77 are also available for SARS‐CoV‐2 RNA detection. This nanorobot is a plasmonic‐magnetic hybrid, composed of an Fe3O4 backbone and an outer surface of Au and Ag. When subjected to a transverse rotating magnetic field, it can actively flip and move with precise directionality. It is important to note that 3D printing 80 and micro/nanorobots 81 can also be utilized for the detection of SARS‐CoV‐2 antigens or antibodies. During the COVID‐19 pandemic, the global medical system has faced an unexpected crisis, prompting the application of 3D printing technology and the exploration of emerging technologies such as nanoscience in the medical field. 82 This has proven to be both creative and promising. 3D printing technology has made significant contributions to the health system by producing personal protective equipment, sample collectors, safety accessories, and even isolation wards. 76 Micro/nanorobots have advanced capabilities such as remote control, automatic propulsion, and precise positioning. 77 However, in comparison with the nanopore sequencing for SARS‐CoV‐2 detection, the current applications of 3D printing and micro/nanorobots are limited and cannot obtain genome information of SARS‐CoV‐2. 83 , 84 , 85 Additionally, cost considerations must be taken into account when compared with RT‐PCR for SARS‐CoV‐2 diagnosis. Nanopore sequencing can obtain SARS‐CoV‐2 genome information while detecting the virus and has been widely used during the COVID‐19 pandemic. 86 , 87 , 88 Below, we will outline several advantages of nanopore sequencing for SARS‐CoV‐2 detection.
3.1.2. Advantages of using nanopore sequencing for SARS‐CoV‐2 detection
Unlike RT‐PCR and RT‐LAMP, which are used for large‐scale population screening, sequencing has the advantage of obtaining the specific sequence of the virus strain. This is particularly useful during the widespread spread of SARS‐CoV‐2, where a large number of mutant strains have emerged and genome sequencing can aid in their identification and classification. Analyzing the similarity of the SARS‐CoV‐2 genome in different regions can also help track virus transmission. After the COVID‐19 pandemic began, researchers quickly identified this new coronavirus strain 14 through metagenomic RNA sequencing on the Illumina platform. A genome‐wide comparison revealed that the SARS‐CoV‐2 genome was 96% identical to bat coronavirus, suggesting its possible origin. 89 However, Illumina sequencing has drawbacks such as being time consuming, requiring complex data analysis, and being costly, 90 limiting its application in some countries or regions, especially those that are economically and medically disadvantaged. 29 These areas are also most likely to be affected by the COVID‐19 pandemic for a prolonged period and may produce more SARS‐CoV‐2 mutant strains. 91 There is an urgent need to popularize sequencing technology in these areas to better understand the mutation, 92 , 93 transmission, and pathogenicity of SARS‐CoV‐2. Fortunately, next‐generation nanopore sequencing technology has effectively addressed the shortcomings of Illumina sequencing in terms of efficiency, convenience, and cost. 94 , 95 , 96
Advantage 1: Real‐time and efficient. Nanopore sequencing can obtain the data required for sequencing in a few hours or less, 97 making it more efficient than Illumina sequencing. This is mainly due to three advantages. First, the library preparation procedure for nanopore sequencing is simple. Direct sequencing eliminates the need for reverse transcription, amplification, and other processes required for Illumina sequencing, simplifying library construction and saving time while avoiding errors that may be introduced by redundant steps. 98 When faced with low viral load, nanopore sequencing can also be combined with different amplification techniques to improve sequencing accuracy. Using a rapid barcoding kit, sublibrary construction based on amplification takes only ten minutes. 99 Second, nanopore sequencing generates data in real‐time. Nucleic acid sequence information is collected as the nucleic acid passes through the nanopore. 97 For COVID‐19 patient samples with high viral load (Ct 18–24), it takes only 1–2 h to generate data covering more than 90% of SARS‐CoV‐2. 99 Finally, advanced bioinformatics analysis techniques enable real‐time data analysis, 2 even for nonprofessionals. 100
Advantage 2: Convenient and affordable. ONT's MinION device is a portable, field‐deployable device 99 that facilitates the timely deployment of sequencing equipment after small‐scale outbreaks, particularly in economically disadvantaged communities that cannot afford to purchase large quantities of equipment. In terms of sequencing cost, the MinION device starts at only $1000. 101 ONT is also committed to reducing the cost of nanopore sequencing. They have launched Flongle, an adapter for MinION that performs one‐time sequencing on small flow cells for only $100, making it a suitable option for community SARS‐CoV‐2 testing. 102 A study comparing the cost of sequencing a single SARS‐CoV‐2 sample using the ONT and Illumina platforms found that the ONT platform costs only $10–40, significantly lower than the $150–250 required by the Illumina platform. This is partly due to the rapid barcoding library preparation method used by the ONT platform, which requires fewer reagents. 29 Although the base error rate of nanopore sequencing samples is higher than that of Illumina sequencing, 29 the high cost effectiveness of nanopore sequencing makes it worth promoting in economically disadvantaged areas.
Advantage 3: Long single‐molecule direct sequencing. Unlike second‐generation Illumina sequencing, nanopore sequencing has the unique advantage of long‐read direct sequencing of nucleic acid molecules. 90 Its expanded use can advance our understanding of SARS‐CoV‐2 epigenetics and transcriptomics. Specifically, it has two advantages: direct sequencing and long‐read sequencing. Traditional sequencing requires reverse transcription and PCR amplification before sequencing the SARS‐CoV‐2 genome sequence, 90 and additional specific chemical treatments are needed to capture RNA modifications for understanding SARS‐CoV‐2 epigenetics. 103 These operations may introduce errors, but nanopore sequencing directly avoids these limitations. For instance, researchers have used nanopore sequencing to reveal the role of m6A RNA modifications in SARS‐CoV‐2 replication. 104 , 105 Additionally, the long‐read characteristic of nanopore sequencing means that its single‐stranded nucleic acid sequencing length is much longer than that of Illumina sequencing. In the face of the complex SARS‐CoV‐2 transcriptome, long‐read technology can easily obtain full‐length transcripts, 106 allowing researchers to glimpse the subgenome landscape of SARS‐CoV‐2. 107
3.1.3. Nanopore sequencing is appropriate for detecting SARS‐CoV‐2
Low read accuracy is a well‐known limitation of nanopore sequencing, raising concerns about its accuracy in sequencing the SARS‐CoV‐2 genome. 108 A study evaluating the effectiveness of nanopore sequencing for genomic analysis of SARS‐CoV‐2 used the ONT and Illumina platforms to sequence 157 samples. The researchers found that although the error rate of nanopore sequencing was higher than that of Illumina sequencing, it still achieved high‐precision determination of consensus‐level sequence and had a sensitivity and precision greater than 99% for monitoring single nucleotide variants (SNVs). 15 Monitoring SNVs alone is sufficient for routine phylogenetic analysis, 30 making nanopore sequencing an effective alternative to Illumina sequencing with trustworthy SARS‐CoV‐2 information. While short‐read sequencing led by Illumina sequencing remains the gold standard for SARS‐CoV‐2, the efficiency and cost advantages of emerging nanopore sequencing cannot be ignored, particularly in less developed countries or regions.
3.2. Library construction for nanopore sequencing for SARS‐CoV‐2 detection
Unlike NGS technology, library preparation for nanopore sequencing does not require amplification, 109 just adding motor proteins and adapters. 35 In the detection of SARS‐CoV‐2, the abundance of viral nucleic acid is generally low, 110 so the combination of different amplification techniques for nanopore sequencing is beneficial to improve the sensitivity of sequencing. A study evaluating the performance of different sequencing technologies found that at low viral loads, amplicon‐based enrichment methods were the most sensitive technology. 111 The mainstream amplification method in nanopore sequencing is the same as the library construction method of next‐generation RNA sequencing, that is, reverse transcription of mRNA and amplify the double‐stranded cDNA obtained by reverse transcription, which is called Amplicon‐Oxford nanopore sequencing. 112 In addition, nanopore targeted sequencing (NTS) is another amplification technology. 113 Compared with amplicon‐based technology, NTS has higher detection efficiency and is more suitable for clinical detection of SARS‐CoV‐2. 16 Loop‐mediated isothermal amplification (LAMP) is another amplification method that has certain advantages in combination with nanopore sequencing 114 (Figure 2).
FIGURE 2.

Nanopore sequencing library preparation and workflow for SARS‐CoV‐2 detection. Due to the low nucleic acid abundance of SARS‐CoV‐2 in samples, realistic library preparation for nanopore sequencing usually requires amplification of viral nucleic acids. The purified viral RNA needs to be reverse transcribed to obtain cDNA before amplification (A). Afterward, the researchers amplified the cDNA by LamPORE, NTS, and Amplicon‐Oxford nanopore sequencing. LamPORE technology uses LAMP technology to amplify cDNA (B). Both NTS and Amplicon‐Oxford nanopore sequencing use PCR technology to amplify cDNA. The difference is that NTS specifically amplifies 12 targets in the SARS‐CoV‐2 gene, which is more efficient and suitable for the detection of SARS‐CoV‐2 (C). Amplicon‐Oxford nanopore sequencing amplifies a large number of SARS‐CoV‐2 gene fragments, and it is beneficial to obtain the complete SARS‐CoV‐2 gene sequence by evaluating the overlap between the amplicons (D). After the above‐mentioned amplification is completed, the basic library construction process of nanopore sequencing is also required. First, add A bases to both ends of the nucleic acid to be tested to make the blunt ends become sticky ends, and then add Y linkers and motor protein. Among them, the motor protein is used to guide the nucleic acid into the nanopore, and there is also a tether on the Y linker, which acts as an anchor when the nucleic acid passes through the nanopore (E). LAMP, loop‐mediated isothermal amplification; NTS, nanopore targeted sequencing; PCR, polymerase chain reaction.
3.2.1. Amplicon‐Oxford nanopore sequencing
In the case of low viral nucleic acid abundance, metagenomic sequencing methods require longer sequencing time and higher costs to analyze the sequence, and multiplex amplicons are a better choice for metagenomic sequencing library construction. 112 Amplicon technology can help ONT sequencing obtain a high‐quality SARS‐CoV‐2 genome, compensating for the higher error rate of ONT sequencing compared with Illumina sequencing. 112 Amplicon technology uses random primers to reverse‐transcribe purified RNA from patient samples to cDNA and performs multiple PCRs to generate the SARS‐CoV‐2 gene amplicon 115 (Figure 2). The researchers used Amplicon‐Oxford nanopore sequencing to examine 42 samples, of which two out of the three the samples had 100% accurate sequencing results. In contrast, another one out of the three the samples had erroneous sequencing results due to poor sample quality and a concentration of purified amplicon less than 1 ng/μL. This highlights a limitation of nanopore sequencing compared with Illumina sequencing: it requires a higher input nucleic acid concentration for sequencing. 2 In addition, increasing the length of the amplicon increases efficiency and reduces cost. 116 Specifically, using the long amplicon protocol (∼2 and ∼2.5 kb) was preferred over the short amplicon protocol (∼400 bp). 117
The ARTIC protocol is the most extensive amplicon‐based SARS‐CoV‐2 sequencing protocol. 118 It relies on reverse transcription amplification of the SARS‐CoV‐2 genome using tiling and multiplexing amplicon protocols. 119 Updates to this protocol (from version v1 to v4) address some of the primer set issues, such as primer aggregation, low coverage, and mutations in primer sites, and effectively reduce the cost of sequencing. 118 At the same time, optimization can be carried out based on the ARTIC protocol (Table 2). For example, some researchers have designed two new primers to specifically amplify the immunodominant part of the spike protein gene. They then used S‐Protein‐Typer to automatically analyze mutations in the spike protein gene at the amino acid level. This method of specifically amplifying the immunodominant gene of the S protein increases sensitivity to mutations in the spike protein gene while reducing costs. 120
TABLE 2.
Amplicon‐based SARS‐CoV‐2 sequencing protocols.
| Protocol | Public time | Optimized features | References | |
|---|---|---|---|---|
| ARTIC protocol | ARTIC V1 | 2020.01.22 | – | 125 |
| ARTIC V2 | – | Solving the problem of primers forming dimers | 125 | |
| Tiled‐ClickSeq | 2020.03.11 | Reduces generation of overlapping amplicons, increases sensitivity to RNA recombination, detects 5′‐UTR | 17 , 121 | |
| ARTIC V3 | 2020.03.24 | Increase coverage and reduce costs | 125 | |
| ARTIC V4 | 2020.06.18 | Use variant sequences to solve the problem of amplicon loss and low genome sequence coverage caused by mutation of primer binding sites | 125 , 126 | |
| S‐Protein‐Typer | 2020.12 | Identify S protein gene mutations | 127 | |
| "Midnight" 1200 bp amplicon split primer sets | 2021.12.16 | Streamline processes and increase efficiency | 122 | |
| – | 2022.06.06 | Improve efficiency | 119 | |
| New protocol | CoronaHiT | 2021.02.09 | Flexible and high throughput | 123 |
| Mini‐XT | 2022.03.03 | Low cost | 124 | |
Amplicon‐based SARS‐CoV‐2 sequencing protocols include updated versions of the mainstream ARTIC protocol, optimized versions based on the ARTIC protocol, and new protocols are being developed.
Compared with tiled amplicon technology, the ClickSeq method can detect 5'‐UTRs 17 , 121 that most previous tiled amplicon technology sequencing would miss. In addition, the 1200 bp amplicon split primer sets of the midnight method can shorten the sample processing time to one working day, which is valuable in small hospitals with small sample volumes. 122 Additionally, using the ARTIC protocol with the Rapid Barcoding Sequencing kit reduces sample library preparation by 70 min. 119
In addition, CoronaHiT and Mini‐XT are two new protocols for amplicon‐based sequencing of SARS‐CoV‐2. 123 , 124 Among them, the CoronaHiT protocol can provide the advantage of flexible high‐throughput, 123 and Mini‐XT is a small library preparation solution. By using acoustic liquid transfer, Mini‐XT greatly reduces the use of reagents and other consumables while ensuring sequence quality, which reduces the cost of library preparation 124 (Table 2).
3.2.2. Nanopore targeted sequencing
Despite the significant progress made by nanopore sequencing over Illumina sequencing in terms of cost and efficiency, it remains more expensive and less efficient than RT‐PCR. As a result, NTS shifted its focus from obtaining the full genome sequence of SARS‐CoV‐2 to diagnosing and monitoring the virus. 16 By using targeted sequencing on the nanopore sequencing platform, NTS was able to achieve high sequencing sensitivity while further reducing detection costs and time. 16 NTS allows subsequent sequencing of amplified fragments specific to SARS‐CoV‐2 on the ONT platform 16 (Figure 2). The specificity of NTS for SARS‐CoV‐2 reached 100% and compared with RT‐PCR, NTS had a higher recognition rate for positive infections. 16 At the same time, NTS can effectively recognize mutant nucleic acid sequences, which is beneficial to classify SARS‐CoV‐2 variants. 16 By adding primers for other respiratory viruses, NTS can not only detect SARS‐CoV‐2 infections but also simultaneously monitor more than 10 other respiratory viruses, including influenza A and B viruses and parainfluenza virus. 16 This allows for timely identification of patients with coinfections of SARS‐CoV‐2 and other respiratory viruses. As the global COVID‐19 pandemic continues 128 and emerging epidemics such as MPXV 129 threaten to place additional strain on medical systems in underdeveloped areas, 130 NTS offers a solution to overcome the high false negative rates of traditional RT‐PCR while also monitoring for coinfections. This improves diagnostic efficiency and reduces the risk of cross‐infection.
3.2.3. LamPORE
LamPORE is a new platform that combines LAMP technology with nanopore sequencing. 114 Specifically, LamPORE is a process for reverse transcription of purified RNA to generate cDNA, followed by LAMP amplification and subsequent sequencing on the nanopore sequencing platform 114 (Figure 2). Nanopore sequencing can improve the low sensitivity of LAMP technology, making LamPORE technology consistent with RT‐PCR in sensitivity. 114 Both prospective and retrospective studies have confirmed the accuracy of the LamPORE technology for the detection of SARS‐CoV‐2. 131 At the same time, LAMP technology brings the possibility of high‐throughput detection to nanopore sequencing. 114 One LamPORE device can complete the detection of 480 samples in 1 h. 132 In addition, LamPORE can be used to detect SARS‐CoV‐2 variants. 132
3.3. Bioinformatics tools to analyze nanopore sequencing data of SARS‐CoV‐2
Nanopore sequencing produces a unique FAST5 data format and its long‐read, direct sequencing features are not available in traditional short‐read sequencing. As a result, bioinformatics tools developed for traditional short‐read sequencing cannot meet the analysis requirements of nanopore sequencing. After the release of ONT's commercial sequencing equipment, researchers developed numerous analysis tools based on specific analysis needs. Following the outbreak of the COVID‐19 pandemic, many analytical tools suitable for SARS‐CoV‐2 monitoring were also developed. Their functions mainly focus on monitoring SARS‐CoV‐2 mutations or integrating operations to simplify and improve interactivity. In this section, we highlight newly developed bioinformatics analysis tools for these outbreaks (Table 3)
TABLE 3.
Bioinformatics analysis of nanopore sequencing.
| Category | Analysis method | Download URL | Function | References |
|---|---|---|---|---|
| Variant detection | VirPool | Using a probabilistic model to estimate the proportion of SARS‐CoV‐2 mutations present in wastewater | – | 133 |
| Variabel | Accurate identification of intrahost variants with low allele frequencies | www.gitlab.com/treangenlab/variabel | 134 , 135 | |
| – | Process FASTQ reads into spike gene consensus sequences to accurately call spike protein variants. | – | 136 | |
| S‐Protein‐Typer | Identification of SARS‐CoV‐2 variants by monitoring mutations in the spike protein gene | https://github.com/MassimoGregorioTotaro/s‐protein‐typer | 120 | |
| LDV‐Caller | Identifying SARS‐CoV‐2 variations from low‐depth sequencing data at reduced costs | – | 137 | |
| Simplified integration | ONTdeCIPHER | The process involves integrating data to reconstruct the consensus genome for each viral isolate. Variants and their effects are then identified, and the lineage is inferred. Finally, multisequence alignments and phylogenetic analyses are performed. | https://github.com/emiracherif/ONTdeCIPHER | 137 |
| MALVIRUS | Integrated, easy to install, easy to use for SARS‐CoV‐2 genotyping | https://algolab.github.io/MALVIRUS | 138 | |
| CalmBelt | Integrated and interactive interface for tracking virus spread and spotting potential spreading clusters | https://github.com/BioML‐CM/CalmBelt | 139 | |
| poreCov | Integration for genotyping and genome reconstruction of SARS‐CoV‐2 | https://github.com/replikation/poreCov | 140 | |
| EDGE COVID‐19 | This workflow uses an integrated and intuitive web‐based interface to automate data quality control, reference‐based genome variant and consensus calling for SARS‐CoV‐2, and lineage determination. | https://edge‐covid19.edgebioinformatics.org https://github.com/LANL‐Bioinformatics/EDGE/tree/SARS‐CoV2 | 141 | |
| NanoCoV19 | Integrated, efficient, surveillance, and lineage analysis for SARS‐CoV‐2 | ‐ | 142 | |
| Others | BugSplit | Sensitive and specific identification of SARS‐CoV‐2 through high‐precision taxonomic binning of metagenomic data | https://bugseq.com/academic | 143 |
| ReadItAndKeep | This fast and lightweight tool scans sequence data and retains only the data that maps to the viral genome. This prevents data leakage related to the human genome. | https://github.com/GenomePathogenAnalysisService/read‐it‐and‐keep | 144 | |
| LeTRS | By identifying leader‐TRS junctions, the abundance of sgmRNAs can be quantitatively measured to study the biological characteristics of SARS‐CoV‐2. | https://scicrunch.org/resolver/RRID:SCR_022138 | 145 | |
| Genopo | A smartphone can be used for portable offline analysis of DNA methylation in human genome samples. | https://mobilegenomics.ce.pdn.ac.lk/genopo/download.html | 100 | |
| InterARTIC | Simple interactive interface to help ordinary users reconstruct consensus genome sequences. | https://github.com/Psy‐Fer/interARTIC/ | 146 | |
| Prediction | Determine optimal sequencing run times, reduce costs, and improve data quality. | – | 147 | |
| AccuVIR | Genome assembly | – | 148 |
During the COVID‐19 pandemic, several bioinformatics methods have been proposed to optimize the results of nanopore sequencing of SARS‐CoV‐2. The application of deep learning or model prediction has reduced the difficulty of data analysis and, consequently, the cost of sequencing.
3.3.1. Application 1: variant detection
Due to higher error rates in nanopore sequencing, some low‐occurrence mutations may be masked. 135 This is important to consider when studying within‐host variation of SARS‐CoV‐2, as it may affect how hosts respond to treatment. 149 To address this issue, researchers have developed a variant call filtering tool called Variabel. This tool compares samples from different patients or from the same patient at different times to distinguish between true variants and sequencing errors. 135 An evaluation of patient‐derived datasets from multiple viruses, including SARS‐CoV‐2, showed that Variabel can accurately identify mutations with an allele frequency below 0.5. 134 , 135 Additionally, deep learning can also improve SARS‐CoV‐2 variant detection in ONT sequencing. Researchers trained a generative adversarial network called LDV‐Caller to analyze low‐depth sequencing results at a level comparable to high‐depth sequencing, greatly reducing the cost of whole‐genome sequencing. 137 Detecting new variants of SARS‐CoV‐2 is also important. The spike protein is key for SARS‐CoV‐2 to bind to host cells, 150 and a large number of mutations in the spike protein can lead to immune escape. 135 Nanopore sequencing can detect mutations in the spike protein gene individually, rather than using traditional whole‐genome sequencing. 136 This can be done by amplifying the gene of the immunodominant part of the S protein using PCR and analyzing it with tools like S‐Protein‐Typer. 120 Additionally, VirPool can estimate the proportion of SARS‐CoV‐2 variants in wastewater samples by analyzing nanopore sequencing data using probabilistic models. 133
3.3.2. Application 2: simplified integration
Analyzing ONT sequencing results requires a certain level of bioinformatics knowledge, which can limit its widespread use. 146 To address this, several interactive applications have been developed for free use on mobile phones 100 and computers. 140 , 141 , 146 These applications use intuitive graphical interfaces, workflows, and job scheduling systems to improve interactivity and make bioinformatics analysis techniques more accessible. 139 , 146 , 151 This allows even users without any bioinformatics background to analyze data from SARS‐CoV‐2 sequencing. 146 Initially, these tools were designed for rapid analysis of the complete genome sequence of SARS‐CoV‐2. 100 , 140 , 146 Now they can also perform rapid typing, lineage analysis, and phylogenetic analysis of SARS‐CoV‐2 by integrating established nomenclature assignments, GISAID clades, and PANGO lineages. 139 , 142 , 151 This helps regional public health systems respond quickly to COVID‐19 outbreaks..
3.4. Nanopore sequencing for analysis of SARS‐CoV‐2 epidemiological information
The ongoing COVID‐19 pandemic has had a significant socioeconomic impact worldwide. However, it has also promoted the use of sequencing technology to detect virus mutations, track the virus's origin and dynamic evolution, and predict epidemic transmission and transmission chains. 152 The large amount of SARS‐CoV‐2 genomic data generated in a short period of time has greatly contributed to the analysis of the virus's epidemiological information. 152 Nanopore sequencing has been widely used during the pandemic, with rapid whole‐genome sequencing and analysis contributing to public health decision‐making and early assessment of SARS‐CoV‐2 transmission. 153 Specifically, nanopore sequencing technology contributes to the prevention and control of the COVID‐19 pandemic in three main ways. 154 First, it can quickly obtain genome sequence information from SARS‐CoV‐2 samples, 155 allowing for timely monitoring and understanding of the invasion of foreign SARS‐CoV‐2 lineages and the diversity of SARS‐CoV‐2 genomes in a region. 156 Second, long‐term monitoring of SARS‐CoV‐2 lineages in the same area can help understand the dynamic replacement of lineages in that region. Different regions may have different mutation lineages of SARS‐CoV‐2 due to varying selection pressures, 157 facilitating exploration of the underlying mechanisms of SARS‐CoV‐2 evolution by understanding spatial heterogeneity during the pandemic. 158 Third, nanopore sequencing can be used to trace the origin and spread of SARS‐CoV‐2, including interregional and intracommunity transmission. Phylogenetic analysis of the sequenced SARS‐CoV‐2 genome and known SARS‐CoV‐2 sequence information from other regions can help determine the most likely source of infection in newly 159 emerged areas, 160 providing a reliable reference for rapid public decision‐making. 161 Additionally, nanopore sequencing can also help identify cases of SARS‐CoV‐2 reinfection. 162
3.4.1. Identification of SARS‐CoV‐2 variants
The results of nanopore sequencing are beneficial to understand the diversity of the SARS‐CoV‐2 genome within a specific region. Timely identification and sequencing of emerging SARS‐CoV‐2 mutations is beneficial for monitoring the evolutionary direction of the virus and predicting changes in viral infectivity and virulence. 163 The identification of emerging VOCs in areas where the virus is rapidly progressing can help control the spread of the outbreak. 164
The application of nanopore sequencing identified a large number of emerging SARS‐CoV‐2 mutations (Table 4), and also specifically identified some VOCs with high transmissibility and high virulence. 164 In addition, the identification of SARS‐CoV‐2 variants by nanopore sequencing can help understand the susceptibility of populations to different SARS‐CoV‐2 variants after vaccination. 165
TABLE 4.
Nanopore sequencing for identifying SARS‐CoV‐2 mutations.
| Region | Time | Lineage | Nucleotide changes | Amino acid changes/transcriptional changes | Samples | References |
|---|---|---|---|---|---|---|
| Hangzhou, China | 2020.1–2020.3 | – | 33 substitutions | 21 amino acid variants in S, N, ORF1a/b, and ORF3a | 29 | 166 |
| Tuscany, Italy | 2020.4 | B1.1 | 5 single nucleotide changes (at positions 241, 3037, 144,08, 19,839, and 23,403) and mutations of 3 consecutive nucleotides (GGG→AAC) at position 28881 | D614G mutation | 1 | 167 |
| 10 different districts, India | 2020.3–2020.6 | A2a, A4, and B | – | 38 amino acid substitutions, include 24 in ORF1ab, 5 in S protein, 2 in ORF 3a, 1 each in E, M, and ORF 7a, and 4 in N protein | 26 | 168 |
| Nanjing, China | 2020–2021 | B.1.1.7, B.1.351, P.1, and B.1.617.2 | 328 nucleotide mutation sites include 205 nonsynonymous mutations and 112 synonymous mutations | ORF1ab, S, ORF3a, and N | 41 | 169 |
| France | 2021.1 | B.1.640 | 46 nucleotide substitutions and 37 deletions | 30 amino acid substitutions and 12 deletions | 13 | 170 |
| Mumbai, India | 2020.8–2020.9, 2021.5–2021.10 | B.1.1.306, B.1.36, B.1.1.32, B.1.1.281, B.1.617.2, B.1.617.2, K417N, AY.120, AY.38, and AY.99 | 642 mutations | D616G mutation, spike(S) mutation, nucleocapsid mutation | 155 | 171 , 172 |
| Colombia | 2021.1–2021.4 | B.1.621 | – | I95I, Y144T, and Y145S | – | 173 |
| Bangladesh | 2021.6–2021.7 | B.1.617.2 and B.1.1.529 | Spike (S) gene deletion | Spike (S) mutation, T19R, L452R, T478K, P681R, and D950N | 52 | 174 , 175 |
| Colorado, America | 2021.6 | AY.1, AY.2, and AY.3 | ORF1b:V2354F and ORF7a:Q94* S112L | NSP15:V303F and Premature stop codon | 34 | 176 |
| Arkansa, America | – | – | T265I, A3529V, G18C, Q57H, S201G, R203K, and G204R | D615G mutation | 2 | 177 |
Nanopore sequencing can quickly obtain the specific gene sequence of SARS‐CoV‐2 and rapidly identify emerging mutant strains. By detecting nucleotide changes in mutant strains, possible viral amino acid changes can be inferred.
Currently, nanopore sequencing is used in several regions for the identification of SARS‐CoV‐2 VOCs. 178 During the first wave of SARS‐CoV‐2 in the UK, researchers tested 563 SARS‐CoV‐2 samples from Oxford using nanopore sequencing. They detected 479 SARS‐CoV‐2 genetic variants, more than half of which involved changes in the amino acid sequence of the encoded protein of the viral genome. 179 In addition, they found that the number of genetic variants of SARS‐CoV‐2 was significantly increased in summer samples compared with spring samples. 179 Following a localized outbreak of SARS‐CoV‐2 infection at a Canadian hospital, researchers performed nanopore sequencing on samples from the hospital. The results showed that mutations in the spike protein gene were associated with headache symptoms in patients. 180 , 181 In southern France, nanopore sequencing identified a novel SARS‐CoV‐2 variant, B.1.640.2, in which the spike protein appeared with substitutions of N501Y and E484K. 170 In Colombia, nanopore sequencing identified the B.1.621 lineage involving amino acid substitutions in the spike protein Y144T, Y145S, R346K, and so on, resulting in B.1.621 being a VOCs. 173 The researchers developed a protocol based on nanopore sequencing in Paraguay based on Spike protein variants, a RT‐PCR for detecting Spike receptor binding domain, which was developed to rapidly identify SARS‐CoV‐2 VOCs during the Paraguay outbreak. 99 , 182 After PCR identified the first four omicron variants in Germany, nanopore sequencing was used for rapid sequencing to confirm the PCR results. 183 In addition, nanopore sequencing also identified lineages such as B.1.1.7, B.1.1.318 in the Republic of Gabon 184 and lineages such as B.1.351 and P.1 in Nice, France. 185 Nanopore sequencing identified a large number of SARS‐CoV‐2 mutant sequences. Including identifying 25 sublineages of SARS‐CoV‐2 B.1.1.529 in Bangladesh between December 2021 and January 2022. 174 , 186 SARS‐CoV‐2 B.1.617.2 sublineages were identified in 12 children in Chittagong, Bangladesh from June 2021 to July 2021. 175
In addition to VOCs, nanopore sequencing also helped to discover variants of SARS‐CoV‐2 in different regions. For example, five SARS‐CoV‐2 variants 171 were identified in Mumbai, India, between August and September 2020. Eight SARS‐CoV‐2 variants were found in rural United States 177 and Mexico. 187 SARS‐CoV‐2 Siena‐1/2020 lineage and Cali‐01 lineage were discovered in Italy 167 and Colombia, 188 respectively, in March 2020. Also, SARS‐CoV‐2 genomes were sequenced from March–April 2020 in Hong Kong 189 and August–September 2020 in Malta. 190
3.4.2. Tracking regional SARS‐CoV‐2 lineage changes
The turnover of the SARS‐CoV‐2 lineage within the region may have resulted from viral evolution 168 or the continuous introduction of foreign dominant mutants. 191 Sequencing samples at different times in the region is beneficial to understand the dynamic replacement process of the SARS‐CoV‐2 lineage in the region 168 (Table 5).
TABLE 5.
SARS‐CoV‐2 lineage replacement and transmission routes in the analysis area using nanopore sequencing.
| Regional lineage replacement | Region | Time | Original lineage (%) | Replaced lineage (%) | Samples | References |
|---|---|---|---|---|---|---|
| Ribeirao Preto City, Brazil | 2020.3–2020.4/2021 | B1.1.33 (61.1%), B.1.1 (27.8%), and B.1.1.28 (11.1%) | P.1 (91%) and P.2 (9%) | 29 | 192 | |
| Sierra Leone | 2020.10/2021.6 | Second wave, R.1 | Third wave, B.1.617.2 | 65 | 193 | |
| Indonesia | 2021.1/2021.7 | First wave, B.1.466.2 | Second wave, AY.23, AY.24, AY.39, AY.42, AY.43, and AY | 202 | 194 | |
| Congo | 2020.3–2020.8/2021.2–2021.3/20216–2021.9 | The first and second waves, B.1.214.2 | Third wave, Delta (B.1.617.2) | 74 | 191 |
| Interregional spread | Region | Time | Lineage | Speculated source | Samples | References |
|---|---|---|---|---|---|---|
| North‐Rhine Westphalia, Germany | 2020.2–2020.3 | – | Netherlands, China | 55 | 195 | |
| Ecuador | 2020.3–2021.1 | HEE‐01 (B) | Netherlands | 4 | 196 | |
| HGSQ‐USFQ‐018 (B1) | United Kingdom | |||||
| HGSQ‐ USFQ‐007 HGSQ‐USFQ‐010 (B) | Scotland | |||||
| India | 2020.3 | A2a | Europe and America | 104 | 197 | |
| A3 | Middle East | |||||
| A4 | Southeast Asia (Indonesia, Thailand, and Malaysia), and Central Asia (Kyrgyzstan) | |||||
| Tunis | 2020.3 | B.1 | Morocco, Ghana, Uganda, and the Democratic Republic of Congo (DRC) | 2 | 127 | |
| A | China | |||||
| Hangzhou, China | 2020.1–2020.3 | – | – | 29 | 166 | |
| Baltimore‐Washington metropolitan area | 2020.3 | – | – | 114 | 198 | |
| Cape Town, South Africa | 2020.3–2020.10 | B.1.1.29 B.1.8 | Europe | 50 | 199 | |
| Netherlands | 2020.3 | – | Germany | 14 | 200 | |
| Islamabad, Pakistan | 2020.11–2021.2 | B.1.1.250 | United Kingdom Bangladesh | 4 | 201 | |
| 2020.11–2021.2 | B.1.261 | Saudi Arabia | 4 | 201 | ||
| Ukraine | 2021.7 | Delta | Central and Eastern European countries | 24 | 202 | |
| Sierra Leone | 2020.3–2021.8 | B.1.617.2 lineage (Delta variant) | England, USA, and Scotland | 130 | 193 | |
| Italian Province of Trento | 2020.3–2021.12 | B.1 B.1.1.29 | Lombardy | 253 | 203 | |
| Nanjing, China | 2020–2021 | B.1.1.7 | Italy, Philippines, and Argentina | 42 | 169 | |
| B.1.351 | Philippines | |||||
| P.1 | Argentina | |||||
| B.1.617.2 | Russia and England | |||||
| Pakistan | 2020.12–2021.2 | B.1.1.7 B.1.36 B.1.1.212 | United Kingdom | 35 | 204 | |
| Armenia | 2020–2022 | B.1.1.7 | Iran, Italy, New Zealand, Russia, Jordan, Germany, and India | 145 | 205 |
Obtaining the specific gene sequence of SARS‐CoV‐2 by nanopore sequencing is beneficial to analyze the epidemiological information of SARS‐CoV‐2. Including understanding the lineage replacement of different waves of SARS‐CoV‐2 epidemics in the region, and inferring the origin of the SARS‐CoV‐2 lineage in the region through phylogenetic analysis.
Nanopore sequencing results reveal SARS‐CoV‐2 evolutionary trends in central India, with the highly infectious spike protein D614G variant continuing to replace other lineages. 168 The Delta lineage predominated in SARS‐CoV‐2 infection in Indonesia. 194 A reduction in some SARS‐CoV‐2 lineages was associated with vaccination status in the Mumbai region of India. 172 In Sierra Leone, 206 the second and third waves of SARS‐CoV‐2 infection consisted of different lineages. Of these, the second wave was the R.1 lineage and the third was the B.1.617.2 lineage, which resulted from continued foreign introductions. 193 In the Democratic Republic of Congo, nanopore sequencing identified seven SARS‐CoV‐2 lineages in three waves and observed 289 variants, and phylogenetic analysis revealed multiple introductions of SARS‐CoV‐2 lineages from different origins. 191 Two waves of outbreaks in the Brazilian city of Ribeirao Preto were analyzed using nanopore sequencing. The results showed that the B1.1.33 lineage was the main lineage in the early stage of the epidemic, and the B.1.1 and B.1.1.28 lineages were also identified; while the second wave of the epidemic was dominated by the P.1 lineage, in addition to the P.2 lineage. 192 In the Karachi region of Pakistan, the A222V variant in the second wave coexisted with the highly contagious D614G variant in the first wave. 207
3.4.3. Mapping SARS‐CoV‐2 transmission
After nanopore sequencing of SARS‐CoV‐2 in the new outbreak area, phylogenetic analysis was performed between the SARS‐CoV‐2 data obtained by sequencing and the virus sequence information in the known region, and the origin of the new outbreak can be inferred. 198 At the same time, nanopore sequencing was also used to detect the SARS‐CoV‐2 outbreak in the community, including the early invasion of SARS‐CoV‐2, 208 the speed of transmission, 158 and the target of transmission. 209
Monitoring the spread of SARS‐CoV‐2 between regions: Sequencing the SARS‐CoV‐2 genomic lineage by nanopore technology is beneficial to understand the introduction events of foreign SARS‐CoV‐2 and the transmission route within the region (Table 5).
Sample sequences from Hangzhou, China, from January to March 2020, suggest multiple overseas origins of SARS‐CoV‐2. 166 Multiple foreign sources of the 2020–2021 outbreak in Nanjing, China were analyzed by nanopore sequencing, including Argentina, Italy, the Philippines, Russia, and the United Kingdom. 169 The nanopore sequencing results of the SARS‐CoV‐2 strain from the outbreak in Heinsberg, Germany, from February to March 2020, showed that it was only related to the SARS‐CoV‐2 strain from the Netherlands. 195 Subsequently, sequencing results of infection cases in the Dutch town of Sittard in early March 2020 showed that the outbreak may have originated in Heinsberg, 200 Germany, which borders Sittard. Nanopore sequencing of SARS‐CoV‐2 isolates from Ecuador, Africa, from the March 2020 outbreak to January 2021 revealed three distinct origins, including the Netherlands, the United Kingdom, and Scotland. 196 , 210 Nanopore sequencing of the SARS‐CoV‐2 genome in India revealed that the outbreak originated from the introduction from Europe, the United States, and Asia after March 2020. 197 More than a quarter of the samples in Maryland and Washington, DC, between March 11 and 31, 2020, were analyzed by nanopore sequencing. The results indicated that the outbreak came from multiple introductions at different locations worldwide. 198 Phylogenetic analysis of sample sequences from the South African outbreak from March to October 2020 revealed that at least nine independent introductions occurred early in the South African outbreak. 199 From March 2020 to August 2021, the SARS‐CoV‐2 lineage in Sierra Leone, Africa, showed that it may have originated from England, the United States, and Scotland. 193 From March 2020 to December 2020, an analysis of the viral genome from the province of Trentino in Italy showed that it was from the Lombardy region. 203 From November 2020 to February 2021, nanopore sequencing of the SARS‐CoV‐2 lineage in the surrounding area of Pakistan showed that the outbreak originated from the introduction of the United Kingdom and Bangladesh. 201 The July 2021 outbreak of the SARS‐CoV‐2 Delta variant in Ukraine mainly came from Middle Eastern and Eastern European countries. 202 Nanopore sequencing of SARS‐CoV‐2 strains in the Karachi region of Pakistan revealed that their outbreaks from December 2020 to February 2021 were related to the UK SARS‐CoV‐2 strain. 204 The 2020–2022 virus lineage in Armenia shows that it may have originated in Iran, Italy, New Zealand, Russia, Jordan, Germany, India, and so on. 205
Nanopore sequencing can be used to monitor urban wastewater and predict the spread of VOCs across a country. 211 In Italy, researchers collected 332 wastewater samples from 20 regions and autonomous provinces between October and November 2022 and sequenced them using both Sanger and nanopore sequencing. 211 They found that the proportion of samples containing mutations from the BQ.1 and BQ1.1 lineages increased significantly in November, reaching 43%. 211 As expected, the variant that was prevalent before being replaced by BQ.1/BQ.1.1 in late 2022 quickly became the dominant SARS‐CoV‐2 lineage in Italy. 211
By further analyzing existing sequencing data and studying the general patterns and differences of SARS‐CoV‐2 genomes in different regions, researchers can gain a better understanding of the virus's evolution and transmission modes. 212 One study analyzed 186,682 samples of SARS‐CoV‐2 isolates from different locations worldwide and classified them into 303 subgroups using PhenoGraph, an unsupervised learning classifier. 212 The origin of these subgroups was then inferred based on their distribution in different countries and regions. 212 Another study analyzed 444,478 SARS‐CoV‐2 genome data and identified 42 comutation modules, which refer to simultaneous mutations that occur when the virus mutates under environmental selection pressure. 213 The sample sources were then grouped based on these comutation modules to determine the phylogenetic relationship between groups. 213 The widespread use of sequencing technology worldwide is therefore helpful in inferring the origin and dynamic spread of the epidemic. The affordability and portability of nanopore sequencing will facilitate the adoption of sequencing technology, enriching the universality and extensiveness of sequencing data in databases.
Monitoring community transmission of SARS‐CoV‐2 through nanopore sequencing, researchers can understand the SARS‐CoV‐2 lineage in the community and observe the transmission process of SARS‐CoV‐2 in the community. 214 Nanopore sequencing can be used to detect SARS‐CoV‐2 gene fragments in social waste water, which facilitates early detection of SARS‐CoV‐2 variant invasion. 208 After the emergence of new variants of SARS‐CoV‐2, the researchers sampled sewage in the community and found several typical VOCs including Alpha, Delta, 164 and Omicron. 215 In Düsseldorf, Germany, nanopore sequencing results indicated a potential link between SARS‐CoV‐2 outbreaks in local populations and hospital outbreaks. 216 Using nanopore sequencing results, the researchers calculated that the doubling time of the SARS‐CoV‐2 BA.2 in dwellings was only 1.28 days. 158 The study by Bosnia and Herzegovina revealed the phenomenon of SARS‐CoV‐2 transmission between humans and pets. 209 The nanopore sequencing results showed that pets in the home of the confirmed patient were also infected with the same SARS‐CoV‐2 as the owner. 209 In a recent long‐term study, researchers monitored the concentration of SARS‐CoV‐2 RNA in wastewater from a university compound over a period of 2.5 years. Using nanopore sequencing, they were able to detect the genome of the Omicron variant strain in the samples. 217
3.4.4. Identification of SARS‐CoV‐2 reinfection
A rare case of SARS‐CoV‐2 reinfection has emerged in Colombia, with two infections identified by nanopore sequencing as lineages B.1 and B.1.1.269. 218 By analyzing the variation of the SARS‐CoV‐2 genome in the GISAID database, the researchers speculated that this patient had multiple SARS‐CoV‐2 variants. 219 They then sequenced 42 COVID‐19 patients using nanopore sequencing and ultimately found that nearly half of them had SARS‐CoV‐2 variant coinfection. 219 In addition, some COVID‐19 patients who had recovered and been discharged tested positive again for the virus using RT‐PCR, even though they were no longer carrying it. Nanopore sequencing revealed that these patients had highly degraded SARS‐CoV‐2 genomes, representing approximately 34.5% of the full genome. Researchers believe that these re‐positive cases may be due to the shedding of cells containing residual SARS‐CoV‐2 nucleic acid in the patient's body. This suggests that these recovered patients had very low infectivity, particularly through respiratory transmission. 162
3.5. SARS‐CoV‐2 infection and other infections
Metagenome sequencing provides the composition of all microorganisms in a sample. 220 Metagenome sequencing was performed in patient samples, enabling direct detection of SARS‐CoV‐2. 221 , 222 By employing nanopore sequencing technology for metagenomic analysis, we can obtain complete influenza virus sequence information directly from respiratory samples taken from patients. This provides the possibility to distinguish SARS‐CoV‐2 infection from other infections and to diagnose SARS‐CoV‐2 infection with other infections. 221
Oropharyngeal commensal microorganisms seem to have a certain impact on the severity of symptoms after host infection with SARS‐CoV‐2. 223 By using nanopore sequencing to analyze the full‐length 16S rRNA gene, researchers found that the oropharyngeal flora of patients infected with SARS‐CoV‐2 was dysregulated. Opportunistic pathogens such as Peptostreptococcus anaerobius and Pseudomonas stutzeri were found to be enriched in these patients. 224 This may eventually lead to coinfection in patients with COVID‐19, whose mycobacterial and mycoplasma enrichment in the nasopharynx is closely associated with subsequent chest pain and fever. 225 Notably, a recent study found that SARS‐CoV‐2 coinfection with L. mirabilis was common in asymptomatic COVID‐19 cases. 226
The influx of critically ill patients into intensive care units due to COVID‐19 has led to a sharp increase in personnel density. These patients often require invasive treatments or examinations, such as invasive ventilation, and are at risk of secondary infections. Nanopore sequencing can quickly identify and type these infections, aiding in the diagnosis and treatment of COVID‐19 patients with secondary infections. 227 Real‐time whole‐genome sequencing on the ONT platform can also monitor persistent infections and assess changes in drug resistance after treatment, providing guidance for medical staff to make appropriate treatment decisions for individual patients. 228 It is worth noting that the results of nanopore sequencing also revealed the risk of secondary infection with a drug‐resistant bacterial infection in COVID‐19 patients, in which a hospital in New York City found secondary infection of carbapenemase‐producing Enterobacterales (CPE) in the COVID‐19 outbreak. 229 Nanopore sequencing was used to rapidly genotype CPE, detect antibiotic resistance genes, and perform phylogenetic analysis in this infection. 229 Notably, clinical metagenomics using nanopore sequencing can identify pathogens and predict bacterial resistance in just 8 h, allowing for timely antibacterial treatment for patients. 227
Middle East respiratory syndrome coronavirus (MERS‐CoV) is a zoonotic disease that has been prevalent in the Middle East since 2012. MERS‐CoV and SARS‐CoV‐2 are both coronaviruses and patients may be coinfected with both viruses. 230 In the Middle East, a large number of MERS‐CoV‐infected patients have similar symptoms to SARS‐CoV‐2‐infected patients. 231 The combination of nanopore sequencing and amplicon technology helped to rapidly differentiate between the two infections. 231 By combining nanopore sequencing with multiplex isothermal amplification, SARS‐CoV‐2 can be differentiated from a variety of viruses including influenza A virus (IAV), and human adenovirus. 232 By combining nanopore sequencing with RT‐PCR technology, the researchers were able to simultaneously screen 41 viruses including MERS‐CoV 233 while identifying SARS‐CoV‐2.
3.6. Nanopore sequencing to study the molecular mechanism of SARS‐CoV‐2 pathogenicity
3.6.1. Direct sequencing of the SARS‐CoV‐2 transcriptome and epitranscriptome
When using nanopore sequencing for clinical diagnosis of SARS‐CoV‐2 infection or monitoring mutations, amplification techniques are often used in conjunction with sequencing due to the generally low virus titer in clinical samples. This helps reduce the error rate of sequencing and the complexity of follow‐up analysis. Research into the specific pathogenic mechanism of SARS‐CoV‐2 can better demonstrate the advantages of direct sequencing and long‐reads provided by nanopore sequencing.
Transcriptomic studies of SARS‐CoV‐2: Studying the SARS‐CoV‐2 transcriptome can reveal specific components of the virus's genome and clarify novel events in its transcriptional process. This can help us understand the pathogenic mechanism of SARS‐CoV‐2 and provide new possibilities for developing clinical tools for prevention, diagnosis, and treatment. 107 While Illumina sequencing has higher accuracy, 18 it is limited by its short reads. Nanopore sequencing, on the other hand, can perform long‐read direct sequencing, which is especially useful for capturing full‐length transcripts and identifying novel transcripts not recognized by short reads. 234 By combining nanopore and Illumina sequencing, researchers have uncovered the complex transcriptomic architecture of SARS‐CoV‐2, including subgenomic RNAs (sgRNAs), open reading frames (ORFs), and transcription‐regulatory sequences (TRSs). They also discovered that SARS‐CoV‐2 differs from other coronaviruses in its unconventional RNA joining events, which deviate from the classical TRS‐mediated mechanism 107 (Figure 3).
FIGURE 3.

Nanopore sequencing was used to study the pathological mechanism of SARS‐CoV‐2. Nanopore sequencing advances the understanding of RNA modifications and transcriptomics of SARS‐CoV‐2. In the study of RNA modification, nanopore sequencing identified 5′ capping sites as well as m6A methylation sites in the SARS‐CoV‐2 genome. The m6A modification has been implicated in SARS‐CoV‐2 replication, and the production of m6A modification requires METTL3, which is upregulated after SARS‐CoV‐2 infection (A). In the study of transcriptomics, through nanopore sequencing of Vero cells infected with SARS‐CoV‐2, the researchers learned about the different structures of sgRNAs and the template switch process in sgRNA production, which involves the production of positive and negative sgRNAs (E). In addition, nanopore sequencing facilitates the study of mutations in the spike protein gene involved in host cell recognition by SARS‐CoV‐2 (B). The long‐read feature of nanopore sequencing has also helped to obtain the complete genome of an animal model of SARS‐CoV‐2 infection (C). Nanopore sequencing identifies the nsp1 gene as a novel target for RT‐PCR detection of SARS‐CoV‐2 (D). METTL3, methyltransferase‐like 3; m6A, N6‐methyladenosine; nsp1, nonstructural protein 1; sgRNAs, subgenomic RNAs.
Nanopore sequencing has also revealed regulatory signatures of the SARS‐CoV‐2 subgenome. 107 , 235 Specifically, SARS‐CoV‐2 sgRNAs can form different structures. 235 and are generated through successive template switches that rely on RNA interactions. 107 Template switching can occur in both directions, including during positive‐strand synthesis. 107 Nanopore sequencing has provided insight into the role of the transcription complex in SARS‐CoV‐2 template switching 236 (Figure 3).
When replicating in vivo, SARS‐CoV‐2 may be forced to delete certain regions from its genome, which could affect its infectivity or responsiveness to vaccines. 237 , 238 Transcriptome analysis using nanopore sequencing has revealed the absence of a furin‐like cleavage site‐encoding portion in the transcriptome of the SARS‐CoV‐2 spike protein. 135 This cleavage site is used to split the spike protein into functional subunits, increasing the virus's infectivity. 239 One study reported its deletion in over half of the transcripts encoding the spike protein. 237
Host cells respond differently to SARS‐CoV‐2 infection. 240 Researchers sequenced infected and mock‐infected human lung adenocarcinoma cells (Calu‐3), human colon cancer cells (Caco‐2), and African green monkey kidney epithelial cells (Vero) using the ONT platform. They found that GSDMB and KPNA2 gene transcripts 240 were associated with SARS‐CoV‐2 infection in different cells. ONT sequencing also confirmed that SARS‐CoV‐2 did not integrate into the DNA of infected HEK293T cells 241 (Figure 3).
Studies on RNA modification in SARS‐CoV‐2: One benefit of nanopore direct sequencing is its ability to accurately identify epigenetic modifications on bases. As nucleic acid molecules pass through the nanopore, any modifications on the base will affect the nanopore's surface current. By recording changes in the current in real‐time, it is possible to identify RNA modifications of SARS‐CoV‐2 after analysis. 242 RNA sequencing results of SARS‐CoV‐2 using the ONT platform have revealed its epitranscriptomic landscape. 18 The results showed that the SARS‐CoV‐2 transcript has at least 41 RNA modification sites, including N6‐methyladenosine (m6A), 5‐methylcytosine methylation (5mC), and 2′‐O‐methylation (Nm). These modification sites most frequently occur on the AAGAA sequence. 18 Additionally, modified RNA has a shorter poly(A) tail than unmodified RNA. 18 Studies have also found that SARS‐CoV‐2 genomic RNA has more modifications at multiple sites compared with sgRNA, 243 and that sgRNA modifications remain stable during the infection process 243 (Figure 3). The abundance of epigenetic modifications in SARS‐CoV‐2 may be related to maintaining the stability of its viral RNA.
m6A modification is the most prevalent type of RNA modification and requires methyltransferase‐like 3 (METTL3) for its production. 244 After infection with SARS‐CoV‐2, Vero E6 cells exhibit increased expression of METTL3 and its translocation from the nucleus to the cytoplasm. 105 Using a method called mixed‐weight neural bagging (MWNB), nanopore sequencing has been able to identify m6A modifications with a high accuracy of 97.85%. 245 ONT sequencing results have shown that SARS‐CoV‐2 RNA contains m6A modifications and that removing these modifications inhibits the replication of SARS‐CoV‐2. 105 This provides an effective target for treating SARS‐CoV‐2 infections. 104 Additionally, an increase in m6A modifications in the SARS‐CoV‐2 gene has been found to be associated with immune evasion by SARS‐CoV‐2 246 (Figure 3).
Nanopore ReCappable Sequencing (NRCeq) is a new technique based on the ONT platform that can identify full‐length sgRNA of SARS‐CoV‐2. 247 Its strength lies in its ability to distinguish intact full‐length sgRNAs from degraded sgRNA fragments. 248 The 5’ end of sgRNA has a specific m7G cap structure, and full‐length sgRNA can be recognized by monitoring this cap. 249 NRCeq can identify full‐length sgRNA of SARS‐CoV‐2 and map its 5’‐capping site by replacing m7G caps with azido‐modified caps. 106 , 247
3.6.2. Development of a novel detection method for SARS‐CoV‐2
RT‐PCR is the most widely used molecular detection technology for monitoring SARS‐CoV‐2. However, the frequent mutations of the virus present a significant challenge for primer design. 250 When a mutation occurs in the SARS‐CoV‐2 sequence targeted by RT‐PCR primers, the primers may no longer effectively amplify the target sequence, resulting in decreased detection sensitivity. Nanopore sequencing identified a nonstructural protein 1 (nsp1) gene in the SARS‐CoV‐2 genome. 251 This gene was highly expressed in samples from individuals infected with SARS‐CoV‐2 of varying severity. 251 The researchers developed a new RT‐PCR assay targeting the nsp1 gene and validated it using 101 clinical samples, with a sensitivity of 93.1% and a specificity of 100%. This achieved the detection sensitivity of conventional RT‐PCR 251 (Figure 3).
3.6.3. Mutation detection of spike protein
The infection process of SARS‐CoV‐2 involves the binding of the spike protein to the host cell angiotensin‐converting enzyme 2 receptor. 150 , 252 The spike protein gene has been a key target for RT‐PCR detection of SARS‐CoV‐2. 253 The study of the spike protein is an important part of the mechanism study of SARS‐CoV‐2. Samples from the South American country from March to April 2020 were sequenced using ONT platform and showed that the D614G substitution in the spike protein gene is common in South America. 254 Another study sequenced samples from January to March 2020 using ONT platform and found the G22017T variant, which corresponds to the W152L mutation of the spike protein 255 (Figure 3).
3.6.4. Sequencing animal models of SARS‐CoV‐2
The Syrian hamster (Mesocricetus auratus) is an important animal model in SARS‐CoV‐2 research because SARS‐CoV‐2 isolates can cause severe lung damage in Syrian hamster lungs. The imaging characteristics of this lung injury resemble those seen in patients with pneumonia following infection with SARS‐CoV‐2. 256 Benefiting from the advantages of long‐read nanopore sequencing, the application of nanopore sequencing allowed researchers to generate higher‐quality Syrian hamster genome sequences, which would be more beneficial for analyzing host responses to SARS‐CoV‐2 infection 257 (Figure 3). Advancing nanopore sequencing technology for reference genomes of animal models such as macaques, ferrets, and cats 258 will aid in understanding the relationship between virus pathogenicity and genetic differences across species.
4. APPLICATIONS OF NANOPORE SEQUENCING IN OTHER MICROBIAL DETECTION
Nanopore sequencing has played an undeniable role in the COVID‐19 pandemic. However, its application in microbial detection extends beyond SARS‐CoV‐2 and COVID‐19 prevention and control. It has been used to detect the genomes of human infectious viruses such as seasonal influenza, 259 MPXV c, Zika, 260 and dengue, 261 as well as animal and plant viruses such as African swine fever virus, 262 Lily virus, 263 and Potato virus Y (PVY). 264 Nanopore sequencing is also used to detect bacteria through 16S rRNA gene sequencing or whole‐genome sequencing to analyze microbial communities in the intestinal tract, 21 nasal cavity, 265 and skin 22 of normal individuals or patients. It can also confirm the specific typing of patient‐infected strains and detect drug resistance‐associated gene mutations. 266 Additionally, nanopore sequencing has been applied to the detection of foodborne pathogens through the detection of specific genes or whole‐genome sequencing to identify pathogen types and predict their potential virulence characteristics. 267
4.1. Virus detection
4.1.1. Detection of viruses in clinical samples
Nanopore sequencing's advantages in clinical virus genome detection lie in its affordability, portability, efficiency, and long‐read capabilities. Its affordability and portability make it easy to popularize and promote, particularly in countries or regions with limited medical resources. This facilitates the timely identification of emerging pathogen types and helps prevent and control epidemics. Its ability to efficiently obtain complete viral genome data is also useful for detecting new virus mutants or tracking the spread of the virus. During the COVID‐19 pandemic, nanopore sequencing played an important role in preventing and controlling regional epidemics caused by other viruses. For example, in Indonesia, where over 100,000 people are infected with DENV annually, researchers used the portable nanopore MinION platform to sequence amplified DENV, improving epidemiological surveillance in resource‐poor settings. 261 Similarly, in Central Africa, where MPXV poses a threat to public health, one study evaluated real‐time sequencing of the MPXV genome using the MinION sequencer to better understand its transmission. The results showed that the sequencing data generated by nanopore sequencing were complete and sufficient for phylogenetic analysis, providing accurate information about the virus source. 268
Influenza viruses can cause seasonal epidemics that pose significant challenges to human health and public health systems. Accurate detection is crucial for patient management and epidemic control. A UK study reported the use of nanopore sequencing for influenza surveillance in a clinical setting, where researchers performed metagenomic sequencing on 180 patient samples. The results showed an 83% sensitivity and 93% specificity for detecting IAV. 268 Nanopore sequencing was also used to identify clonotypic diversity in the IAV genome and, when combined with Illumina sequencing, increased the accuracy of clonotype identification to 98.2%. 269 Frequent mutations in IAV may reduce RT‐PCR detection of gene targets, but nanopore sequencing can identify new diagnostic targets for IAV mutants. 270 By sequencing clinically infected patients, researchers identified PB2 and NS genes as new IAV diagnostic targets and found that using them for RT‐PCR detection had lower detection limits than traditional M genes. 270
4.1.2. Animal and plant virus detection
Plant and animal viruses can cause significant economic losses in agriculture and animal husbandry worldwide. 271 Symptoms of plant viral infections are often atypical and nonspecific. While traditional sequencing technology has been widely used for detecting and identifying plant viruses, it is not suitable for on‐site detection and can be costly. 272 Nanopore sequencing's portability and flexible throughput make it uniquely adaptable for plant virus detection. 271 It has been used to detect viruses in common economic crops such as potato, wheat, broad bean, 273 lily, 274 and jasmine. 275 For example, nanopore sequencing can distinguish between virus infection and nutrient deficiency in early‐stage wheat growth and accurately identify Wheat streak mosaic virus while revealing variations in its resistance gene Wsm2. 276 It is also used for genotyping PVY, with whole‐genome sequencing of different PVY types showing over 95% consistency with the consensus sequence obtained by Illumina sequencing. 264
Animal virus infections can spread rapidly and have severe socioeconomic consequences. Nanopore sequencing has been used to identify various animal viruses, including differentiating between three Capripox Viruses in outbreaks in the Middle East and Africa, 277 identifying African horse sickness in an outbreak in Thailand, 278 and tracking Enzootic Bovine Leukosis caused by Bovine leukemia virus transmission in US pastures. 279 It has also been used to reveal the complex transcriptional landscape of porcine reproductive and respiratory syndrome virus. 280
4.2. Detection of microbial populations
By using nanopore sequencing for metagenomic or 16S rRNA sequencing, researchers can analyze the diversity of microbial populations in the environment or human tissues. The portable and real‐time MinION sequencing system released by ONT is suitable for field biological exploration to study microbial communities and has been used in various locations, including the Antarctic, 23 Arctic, 23 tropical desert 281 regions, and even the International Space Station. Its resilience to harsh environments 282 and ability to provide power and data transfer over a single USB 3.0 connection 283 enabled real‐time sequencing on the International Space Station, reducing transportation costs and avoiding sample degradation during storage. 283
In a healthy physiological state, certain types and quantities of microorganisms live on the human body surface and body cavity. Imbalances in microbiota are linked to the development of various diseases. 284 , 285 While short‐read 16S rRNA sequencing has commonly been used to analyze microbial populations in humans, full‐length 16S rRNA sequencing using nanopore sequencing technology is an effective and low‐cost alternative. 286 , 287 It has been used for sequencing the endometrial, 287 skin, 22 and gut 286 microbiomes.
Metagenomic sequencing can reveal the specific composition and function of bacterial, viral, and eukaryotic mixtures in the microbiome. While traditional short‐read sequencing produces incomplete and fragmented microbial genomes, 21 long‐read nanopore sequencing can assemble contiguous bacterial isolate genomes. 21 Nanopore sequencing can also detect DNA methylation in bacteria and microbiomes. Researchers developed a method called nanodisco to identify reliable DNA methylation 288 when applied to individual bacteria or the small intestinal microbiome of mice.
5. APPLICATION OF NANOPORE SEQUENCING IN CANCER GENOMES
Cancer is characterized by genomic changes and its development is accompanied by progressive changes in the genome and epigenome. 289 , 290 Sequencing technology has deepened our understanding of cancer genome changes and the emergence of NGS technology and high‐performance computers has enabled comprehensive analysis of the cancer genome at both basic research and clinical levels. 289 Sequencing technology is widely used in basic cancer genome research and its deepening has promoted clinical applications such as personalized cancer therapies through molecular target detection or cancer biomarker screening. 291 Nanopore sequencing's advantages in cancer research lie in its ability to perform direct single‐molecule long‐read sequencing, capturing cancer transcriptome 24 , 25 and epigenetic changes. 292 Its long‐read capabilities make it more sensitive to structural variations (SVs) in cancer genomes 293 , 294 and better able to identify alternative splicing sites in gene expression. 295 , 296 In clinical applications, nanopore sequencing's portability and speed make it a useful tool for biopsy, 297 particularly liquid biopsy. 297 Its long‐read capabilities can capture special DNA fragments in blood that cannot be achieved by short‐read sequencing 298 (Figure 4).
FIGURE 4.
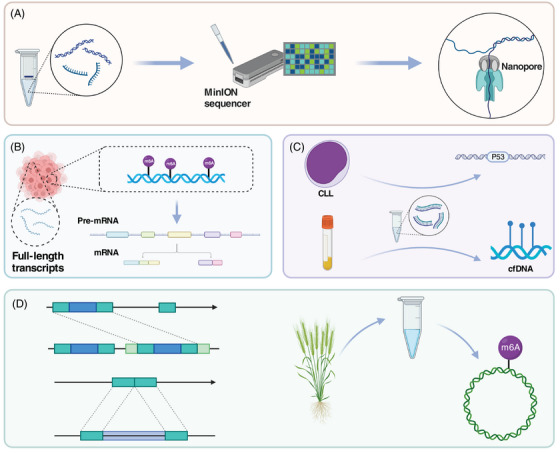
Nanopore sequencing for the detection of cancer and plant genomes. The exceptional performance of nanopore sequencing's single‐molecule long‐reads has made it a popular choice for sequencing cancer and plant genomes (A). In cancer research, nanopore sequencing is used to obtain transcripts and epigenetic modifications, as well as to understand the process of AS (B). Clinically, it can be used for gene analysis and liquid biopsy in patients (C). In plant research, nanopore sequencing is used to analyze and obtain high‐quality genomes (D). cfDNA, cell‐free DNA; CLL, chronic lymphocytic leukemia; m6A, N6‐methyladenosine.
5.1. Basic research on cancer
While RNA direct sequencing is possible on traditional sequencing platforms, 299 most RNA‐Seq experiments, including those for cancer transcriptomics, are performed on instruments that sequence converted DNA molecules. 300 This requires the preparation of RNA into cDNA. 300 Similarly, detection of epigenetic modifications is also indirect. 301 For example, DNA 5mC modification is detected through Bisulfite sequencing (BS‐Seq) and high‐throughput sequencing of m6A, the most common modification in eukaryotic mRNA, relies on m6A‐specific antibodies. 302 , 303 Nanopore sequencing can sequence RNA without PCR amplification and directly detect different types of epigenetic modifications, 301 avoiding complex sample handling procedures or potential bias from reverse transcription. Its long‐read capabilities help identify full‐length tumor transcripts, AS cleavage sites, and accurately locate SVs in the cancer genome. 25 One study used nanopore sequencing to directly sequence the transcriptomes of tumors and adjacent tissue from three hepatocellular carcinoma (HCC) patients and found 361 novel transcripts significantly associated with HCC patient prognosis. Another study analyzed the transcriptomes of 42 pairs of HCC and matched noncancerous liver samples using nanopore sequencing and detected 46,663 transcripts, one‐tenth of which were novel discoveries. 304 Nanopore sequencing revealed AS events in early stages of HCC carcinogenesis. 304 Another study used nanopore sequencing to perform whole‐genome sequencing of 11 HCC samples from Japan and identified 8004 insertions, 27 inversions, 6389 deletions, and 32 intrachromosomal translocations, and it also showed that long‐read sequencing identified more SVs 293 than short‐read sequencing.
5.2. Clinical research on cancer
In the era of precision oncology, the development of molecular target drugs allows for the identification of specific tumor types based on genomic changes and the administration of matching therapies. 305 Comprehensive assessment of a tumor patient's genome through sequencing can guide clinical treatment based on specific changes and help evaluate and predict cancer progression and prognosis. 306 Several studies have used nanopore sequencing to assess genomic changes in patients with hematological malignancies. 26 , 27 , 307 One study verified the feasibility of using nanopore sequencing to detect P53 gene mutations in chronic lymphocytic leukemia (CLL) patients and found that its accuracy reached 95–96% after correction by computer methods. 27 Notably, nanopore sequencing also identified a mutation not detected by Sanger sequencing. 27 Another study proposed a rapid mutation detection method for six acute myeloid leukemia (AML) hotspot genes based on nanopore sequencing. 26 After testing and validating using Sanger sequencing on 22 AML patients, one study demonstrated that nanopore sequencing results for the mutation status of hotspot genes were reliable and reproducible. 26
Clinical tumor biopsy, particularly liquid biopsy, is another promising application of nanopore sequencing. 308 Circulating cell‐free DNA (cfDNA) in blood carries informative features of its original tissue origin 298 and analyzing cfDNA in plasma is an effective and noninvasive strategy for obtaining tumor genomic information. 309 Nanopore sequencing can overcome the inconvenience and high equipment cost of NGS technology for clinical cfDNA detection. One study used nanopore sequencing to detect copy number variations in cfDNA in the plasma of lung cancer patients and healthy individuals, with results highly consistent with Illumina sequencing. 309 In terms of efficiency, nanopore sequencing can generate 2M reads within 3 h and complete the entire process, including analysis, in one working day. 309 It can also directly detect cfDNA methylation, a marker for early cancer screening. 310 Direct methylation detection by nanopore sequencing saves cost and time for sample processing before clinical testing while reducing sample degradation. 298 In addition to detecting cfDNA in blood, Bruzek et al. 311 used nanopore sequencing to detect cfDNA in cerebrospinal fluid (CSF) to determine the therapeutic effect of Pediatric high‐grade glioma. The detection of CSF samples showed a sensitivity of 85% and a specificity of 100%. 311 Furthermore, studies have reported the use of nanopore sequencing to detect SVs in clinical ovarian and prostate cancer tumor samples. 312
6. APPLICATION OF NANOPORE SEQUENCING IN PLANT GENOMES
Since the Arabidopsis thaliana (L.) Heynh genome was first sequenced and assembled using Sanger sequencing in 2000, sequencing technology has been widely used in botany. 313 Unlike animal genomes, plant genomes often contain a large number of repetitive sequences. For example, the wheat genome contains 90% repetitive sequences. 314 Additionally, the genome size of plants varies greatly. Genlisea tuberosa has the smallest genome at 61 Mb, 315 while the loblolly pine has the largest genome at 22 Gb. 315 Short‐read sequencing technology has difficulty spanning repetitive sequences and resolving polyploids in plant genomes. However, long‐read sequencing can overcome these obstacles and provides the possibility of analyzing complex plant genome sequences 314 (Figure 4).
Oxford nanopore sequencing, the most popular long‐read sequencing platform on the market, has been applied in the field of plant genome sequencing 28 and has resolved a series of high‐quality genomes. High‐quality reference genomes are crucial for exploring the evolutionary history of plants and for changing plant traits through artificial selection. 314 Oryza sativa is an important agricultural crop, but the lack of high‐quality reference genomes has hindered researchers' understanding of its evolutionary history. 316 Choi et al. 316 used nanopore sequencing to assemble the genomes of two circum‐basmati rice cultivars and found strong evidence of admixture between circum‐basmati and circum‐aus cultivars. This suggests that the genome composition of some circum‐basmati varieties can be traced back to japonica. 316 A. thaliana was the first important model species to obtain a full‐length genome through sequencing. However, due to the enrichment of highly repetitive elements, its new version of the reference genome still has missing or incorrect positions such as telomeres and centromeres. 317 , 318 Wang and colleagues 319 obtained an almost complete high‐quality genome of A. thaliana named Col‐XJTU. by combining nanopore sequencing with PacBio HiFi long‐read technology.
SVs are common in plant genomes and can directly affect plant traits such as crop yield and quality. 320 Long‐read sequencing is known to be more effective in identifying SVs, making it important to choose the right long‐read sequencing method for detecting plant SVs. In a comparison of the performance of nanopore sequencing and Pacific Biosciences (PacBio) sequencing in detecting pear SVs, Liu and colleagues 321 found that nanopore sequencing performed better and detected more SVs than PacBio sequencing. Another study used nanopore sequencing to identify 238,490 SVs from 100 different cultivars of wild and domesticated tomato. The researchers eventually identified hundreds of SV genes 322 that may broadly affect quantitative trait variation. By using genome editing technology to improve the SVs and expression levels of these genes, researchers were able to enhance tomato size, quantity, taste, and other traits. 322
RNA modifications play crucial and dynamic roles in regulating gene expression during plant embryonic development, flowering, and fruit ripening. Several studies have used nanopore sequencing to detect the most common modification in plant RNA, the m6A modification. 323 For example, Parker et al. 324 used nanopore sequencing to directly sequence wild‐type and mRNA methylation mutants from the model plant A. thaliana. This allowed them to map the complexity of mRNA processing and m6A modification in both wild‐type and mutant Arabidopsis thaliana. 324 Another study used nanopore direct RNA sequencing to detect m6A modifications in circular RNA from Phyllostachys edulis. This enabled them to pinpoint the location of m6A modifications in circular RNAs at single‐base resolution. 325
7. THE ACCURACY OF NANOPORE SEQUENCING
Despite its great potential in detecting SARS‐CoV‐2, the low read accuracy of nanopore sequencing remains a significant limitation. In this section, we provide an in‐depth explanation of the reasons for this low accuracy and potential strategies for improvement.
The high error rate of nanopore sequencing is primarily due to the low accuracy of base calling. 326 The basic principle of nanopore sequencing involves identifying the base composition or chemical modification of nucleic acid molecules by detecting the changes in current generated as they pass through the nanopore. However, the current changes caused by different bases are not as distinct as one might expect. From first‐generation to second‐generation sequencing, we have entered the era of high‐throughput sequencing, which has reduced accuracy to some extent. With third‐generation nanopore sequencing technology, single‐molecule sequencing is achieved on the basis of high‐throughput. It is understandable that during single‐molecule sequencing, the current change caused by a single base is very weak. Additionally, it is important to note that the original current signal recognized by current base‐callers does not contain only one base but multiple bases with an uncertain number. This signal is then decoded into the most likely set of bases. 326 , 327 Ideally, the original current signal of each recognition unit would differ by one base, but the speed at which nucleic acid passes through the nanopore is not constant, making it difficult to determine the type or position of a single base. 326 Finally, nanopore sequencing has difficulty identifying repetitive sequences of the same base 326 and homopolymers 97 that exceed the length of the nanopore reading head.
Several strategies can improve the identification accuracy of nanopore sequencing, including optimizing nanopores and motor proteins, performing multiple sequencing, and updating the base‐calling algorithm. 2 ONT has been optimizing nanopores and motor proteins. 2 By using biological nanopores and genetic engineering techniques, they can modify the size and charge location of the nanopore, ultimately affecting the number of bases read simultaneously. 327 Motor proteins control the speed at which nucleic acid strands pass through the nanopore. Maintaining a stable and slow passing speed helps the sensor recognize current changes and partially overcomes the problem of reading different numbers of bases at the same time. 327 For users who urgently need high‐accuracy sequencing using nanopore sequencing, performing multiple sequencing of a single nucleic acid to generate a consensus sequence can effectively improve the quality of sequencing data. 2 A study that conducted deep sequencing of E. coli strains with 30‐fold coverage on the MinION platform increased accuracy from 80 to 99.5%. 2 , 328 Additionally, developing new base‐calling algorithms is an effective way to improve the accuracy of nanopore sequencing. From past Markov model base‐calling algorithms to new‐generation neural network algorithms, the accuracy of nanopore sequencing combined with machine learning algorithms has increased to 98.3%. 329
8. CONCLUSIONS AND FUTURE PERSPECTIVES
Nanopore sequencing technology, as a third‐generation sequencing technology, has the characteristics of real‐time sequencing, long reads, and high efficiency. Compared with RT‐PCR, targeted nanopore sequencing is more sensitive for diagnostic sequencing of SARS‐CoV‐2. Nanopore sequencing is cheaper and faster than Illumina sequencing. Nanopore can identify novel mutants of SARS‐CoV‐2 and identify SARS‐CoV‐2 VOCs in outbreak areas. By analyzing the nanopore sequencing results, the researchers were able to trace the origin of SARS‐CoV‐2 variants in the epidemic region and keep abreast of the lineage replacement of SARS‐CoV‐2 in the region. Nanopore sequencing can also be used to diagnose SARS‐CoV‐2 infection and other infections, and to distinguish SARS‐CoV‐2 infection from other infections. Nanopore sequencing is also used to prevent and control local outbreaks of viruses such as MPXV and NoV, particularly in developing countries with limited medical resources. It is also used for sequencing the genomes of viruses that infect plants or animals and for detecting the diversity of microbial communities.
In addition, nanopore sequencing can perform long single molecule direct RNA sequencing and obtain RNA transcriptome and epitranscriptome, which is helpful for related research on the molecular mechanism of SARS‐CoV‐2 pathogenesis, as well as the occurrence and development of cancer or the acquisition of high‐quality genomes of plant genomes. mRNA vaccines are a type of vaccine for SARS‐CoV‐2. 330 The safety of mRNA vaccines is a concern 330 due to the occasional adverse reactions of unknown mechanisms with the use of mRNA vaccines. 331 Nanopore sequencing can carry out quality inspection of mRNA vaccines and ensure the safety of vaccines to a certain extent.
This article reviews the extensive application of nanopore sequencing technology in the COVID‐19 epidemic, and systematically summarizes the improvement of the ONT library preparation process and the relevant bioinformatics analysis of the mutation evolution of the SARS‐CoV‐2 virus. The use of nanopore sequencing in detecting other microbial pathogens, as well as its application in cancer and plant genomics, is also summarized. The rapid and convenient nature of nanopore sequencing is ideal for identifying regional SARS‐CoV‐2 VOCs and detecting the dynamic evolution of SARS‐CoV‐2 variants. Today, the global COVID‐19 outbreak has been effectively controlled, but regional outbreaks are still frequent. The application of nanopore sequencing will help to rapidly trace the origin of the COVID‐19 epidemic in the region and the extent of the epidemic in limited areas. At the same time, looking at the world's epidemic prevention and control forms, the highly contagious respiratory diseases caused by coronaviruses such as SARS and MERS have not disappeared. We can imagine that nanopore sequencing will play a huge role in this and the next global pandemic.
AUTHOR CONTRIBUTION
P. Z. and S. D. contributed to the conception, design, and final approval of the submitted version. Literature was collected and analyzed by P. Z. and S. D. P. Z., Y. D., C. Z., L. L., and S. D. contributed to the manuscript writing. P. Z., Y. D., C. Z., B. L., F. Z., and S. D. contributed to the graphic design, and all authors conceived and approved the final manuscript.
CONFLICT OF INTEREST STATEMENT
The authors declare that the study was conducted in the absence of any business or financial relationship that could be interpreted as a potential conflict of interest.
ETHICS STATEMENT
Not applicable.
ACKNOWLEDGMENTS
Figures were created through BioRender.com. This research was funded by the Hangzhou Agricultural and Social Development Research Project (20201203B152).
Zheng P, Zhou C, Ding Y, et al. Nanopore sequencing technology and its applications. MedComm. 2023;4:e316. 10.1002/mco2.316
DATA AVAILABILITY STATEMENT
Data sharing is not applicable to this article as no new data were created or analyzed in this study.
REFERENCES
- 1. Deamer D, Akeson M, Branton D. Three decades of nanopore sequencing. Nat Biotechnol. 2016;34(5):518‐524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Wang Y, Zhao Y, Bollas A, Wang Y, Au KF. Nanopore sequencing technology, bioinformatics and applications. Nat Biotechnol. 2021;39(11):1348‐1365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Bezrukov SM, Kasianowicz JJ. Current noise reveals protonation kinetics and number of ionizable sites in an open protein ion channel. Phys Rev Lett. 1993;70(15):2352‐2355. [DOI] [PubMed] [Google Scholar]
- 4. Walker B, Kasianowicz J, Krishnasastry M, Bayley H. A pore‐forming protein with a metal‐actuated switch. Protein Eng. 1994;7(5):655‐662. [DOI] [PubMed] [Google Scholar]
- 5. Stoddart D, Heron AJ, Mikhailova E, Maglia G, Bayley H. Single‐nucleotide discrimination in immobilized DNA oligonucleotides with a biological nanopore. Proc Nat Acad Sci USA. 2009;106(19):7702‐7707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Butler TZ, Pavlenok M, Derrington IM, Niederweis M, Gundlach JH. Single‐molecule DNA detection with an engineered MspA protein nanopore. Proc Nat Acad Sci USA. 2008;105(52):20647‐20652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Derrington IM, Butler TZ, Collins MD, et al. Nanopore DNA sequencing with MspA. Proc Nat Acad Sci USA. 2010;107(37):16060‐16065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Lieberman KR, Cherf GM, Doody MJ, Olasagasti F, Kolodji Y, Akeson M. Processive replication of single DNA molecules in a nanopore catalyzed by phi29 DNA polymerase. J Am Chem Soc. 2010;132(50):17961‐17972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Cherf GM, Lieberman KR, Rashid H, Lam CE, Karplus K, Akeson M. Automated forward and reverse ratcheting of DNA in a nanopore at 5‐Å precision. Nat Biotechnol. 2012;30(4):344‐348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Chen P, Sun Z, Wang J, et al. Portable nanopore‐sequencing technology: trends in development and applications. Front Microbiol. 2023;14:1043967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Safiabadi Tali SH, LeBlanc JJ, Sadiq Z, et al. Tools and techniques for severe acute respiratory syndrome coronavirus 2 (SARS‐CoV‐2)/COVID‐19 detection. Clin Microbiol Rev. 2021;34(3):e00228‐20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Wang M, Li M, Ren R, et al. International expansion of a novel SARS‐CoV‐2 mutant. J Virol. 2020;94(12):e00567‐20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Metzger C, Lienhard R, Seth‐Smith HMB, et al. PCR performance in the SARS‐CoV‐2 Omicron variant of concern? Swiss Med Wkly. 2021;151:w30120. [DOI] [PubMed] [Google Scholar]
- 14. Wu F, Zhao S, Yu B, et al. A new coronavirus associated with human respiratory disease in China. Nature. 2020;579(7798):265‐269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Bull RA, Adikari TN, Ferguson JM, et al. Analytical validity of nanopore sequencing for rapid SARS‐CoV‐2 genome analysis. Nat Commun. 2020;11(1):6272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Wang M, Fu A, Hu B, et al. Nanopore targeted sequencing for the accurate and comprehensive detection of SARS‐CoV‐2 and other respiratory viruses. Small (Weinheim an der Bergstrasse, Germany). 2020;16(32):e2002169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Jaworski E, Langsjoen RM, Mitchell B, et al. Tiled‐ClickSeq for targeted sequencing of complete coronavirus genomes with simultaneous capture of RNA recombination and minority variants. eLife. 2021;10:e68479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Kim D, Lee JY, Yang JS, Kim JW, Kim VN, Chang H. The architecture of SARS‐CoV‐2 transcriptome. Cell. 2020;181(4):914‐921.e10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Buenestado‐Serrano S, Herranz M, Palomino‐Cabrera R, et al. Rapid identification of relevant microbial strains by identifying multiple marker single nucleotide polymorphisms via amplicon sequencing: epidemic monkeypox virus as a proof of concept. Microbiol Spect.. 2023;11(1):e0419622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Xiong Q, Jiang H, Liu Z, et al. Untangling an AGS outbreak caused by the recombinant GII.12[P16] norovirus with nanopore sequencing. Front Cell Infect Microbiol. 2022;12:911563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Maghini DG, Moss EL, Vance SE, Bhatt AS. Improved high‐molecular‐weight DNA extraction, nanopore sequencing and metagenomic assembly from the human gut microbiome. Nat Protoc. 2021;16(1):458‐471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Rozas M, Brillet F, Callewaert C, Paetzold B. MinION™ nanopore sequencing of skin microbiome 16S and 16S‐23S rRNA gene amplicons. Front Cell Infect Microbiol. 2021;11:806476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Johnson SS, Zaikova E, Goerlitz DS, Bai Y, Tighe SW. Real‐time DNA sequencing in the antarctic dry valleys using the oxford nanopore sequencer. J Biomol Tech: JBT. 2017;28(1):2‐7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Cruz‐Garcia L, O'Brien G, Sipos B, et al. Generation of a transcriptional radiation exposure signature in human blood using long‐read nanopore sequencing. Radiat Res. 2020;193(2):143‐154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Fang Y, Chen G, Chen F, et al. Accurate transcriptome assembly by Nanopore RNA sequencing reveals novel functional transcripts in hepatocellular carcinoma. Cancer Sci. 2021;112(9):3555‐3568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Cumbo C, Minervini CF, Orsini P, et al. Nanopore targeted sequencing for rapid gene mutations detection in acute myeloid leukemia. Genes. 2019;10(12):1026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Minervini CF, Cumbo C, Orsini P, et al. TP53 gene mutation analysis in chronic lymphocytic leukemia by nanopore MinION sequencing. Diagn Pathol. 2016;11(1):96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Hofmann NR. Nanopore sequencing comes to plant genomes. Plant Cell. 2017;29(11):2677‐2678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Ranasinghe D, Jayadas TTP, Jayathilaka D, et al. Comparison of different sequencing techniques for identification of SARS‐CoV‐2 variants of concern with multiplex real‐time PCR. PLoS One. 2022;17(4):e0265220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Young BE, Fong SW, Chan YH, et al. Effects of a major deletion in the SARS‐CoV‐2 genome on the severity of infection and the inflammatory response: an observational cohort study. Lancet (London, England). 2020;396(10251):603‐611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Hu ZL, Huo MZ, Ying YL, Long YT. Biological nanopore approach for single‐molecule protein sequencing. Angew Chem Int Ed Engl. 2021;60(27):14738‐14749. [DOI] [PubMed] [Google Scholar]
- 32. Xu L, Seki M. Recent advances in the detection of base modifications using the Nanopore sequencer. J Hum Genet. 2020;65(1):25‐33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Żmieńko A, Satyr A. Nanopore sequencing and its application in biology. Postepy Biochem. 2020;66(3):193‐204. [DOI] [PubMed] [Google Scholar]
- 34. Lin B, Hui J, Mao H. Nanopore technology and its applications in gene sequencing. Biosensors. 2021;11(7):214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Zhou Y, Ren M, Zhang P, et al. Application of nanopore sequencing in the detection of foodborne microorganisms. Nanomaterials (Basel, Switzerland). 2022;12(9):1534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Ashkenasy N, Sánchez‐Quesada J, Bayley H, Ghadiri MR. Recognizing a single base in an individual DNA strand: a step toward DNA sequencing in nanopores. Angew Chem Int Ed Engl. 2005;44(9):1401‐1404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. MacKenzie M, Argyropoulos C. An introduction to nanopore sequencing: past, present, and future considerations. Micromachines. 2023;14(2):459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Cao C, Ying YL, Hu ZL, Liao DF, Tian H, Long YT. Discrimination of oligonucleotides of different lengths with a wild‐type aerolysin nanopore. Nat Nanotechnol. 2016;11(8):713‐718. [DOI] [PubMed] [Google Scholar]
- 39. Liang B, Tamm LK. Structure of outer membrane protein G by solution NMR spectroscopy. Proc Nat Acad Sci USA. 2007;104(41):16140‐16145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Faller M, Niederweis M, Schulz GE. The structure of a mycobacterial outer‐membrane channel. Science. 2004;303(5661):1189‐1192. [DOI] [PubMed] [Google Scholar]
- 41. Laszlo AH, Derrington IM, Gundlach JH. MspA nanopore as a single‐molecule tool: from sequencing to SPRNT. Methods (San Diego, Calif). 2016;105:75‐89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Abiola O, Angel JM, Avner P, et al. The nature and identification of quantitative trait loci: a community's view. Nat Rev Genet. 2003;4(11):911‐916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Zhou Y, Zhang L, Xie YH, Wu J. Advancements in detection of SARS‐CoV‐2 infection for confronting COVID‐19 pandemics. Lab Invest. 2022;102(1):4‐13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Wong ML, Medrano JF. Real‐time PCR for mRNA quantitation. BioTechniques. 2005;39(1):75‐85. [DOI] [PubMed] [Google Scholar]
- 45. Corman VM, Landt O, Kaiser M, et al. Detection of 2019 novel coronavirus (2019‐nCoV) by real‐time RT‐PCR. Euro Surveill. 2020;25(3):2000045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Notomi T, Okayama H, Masubuchi H, et al. Loop‐mediated isothermal amplification of DNA. Nucleic Acids Res. 2000;28(12):E63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Yan C, Cui J, Huang L, et al. Rapid and visual detection of 2019 novel coronavirus (SARS‐CoV‐2) by a reverse transcription loop‐mediated isothermal amplification assay. Clin Microbiol Infect. 2020;26(6):773‐779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Chen Q, Li J, Deng Z, Xiong W, Wang Q, Hu YQ. Comprehensive detection and identification of seven animal coronaviruses and human respiratory coronavirus 229E with a microarray hybridization assay. Intervirology. 2010;53(2):95‐104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Lisboa Bastos M, Tavaziva G, Abidi SK, et al. Diagnostic accuracy of serological tests for covid‐19: systematic review and meta‐analysis. BMJ (Clin Res Ed). 2020;370:m2516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Liu W, Liu L, Kou G, et al. Evaluation of nucleocapsid and spike protein‐based enzyme‐linked immunosorbent assays for detecting antibodies against SARS‐CoV‐2. J Clin Microbiol. 2020;58(6):e00461‐20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Khalaf K, Papp N, Chou JT, Hana D, Mackiewicz A, Kaczmarek M. SARS‐CoV‐2: pathogenesis, and advancements in diagnostics and treatment. Front Immunol. 2020;11:570927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Bruning AHL, Leeflang MMG, Vos J, et al. Rapid tests for influenza, respiratory syncytial virus, and other respiratory viruses: a systematic review and meta‐analysis. Clin Infect Dis. 2017;65(6):1026‐1032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Sholukh AM, Fiore‐Gartland A, Ford ES, et al. Evaluation of cell‐based and surrogate SARS‐CoV‐2 neutralization assays. J Clin Microbiol. 2021;59(10):e0052721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Lin Y, Dai Y, Liu Y, et al. Rapid PCR‐based nanopore adaptive sequencing improves sensitivity and timeliness of viral clinical detection and genome surveillance. Front Microbiol. 2022;13:929241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Yüce M, Filiztekin E, Özkaya KG. COVID‐19 diagnosis—a review of current methods. Biosens Bioelectron. 2021;172:112752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Pu R, Liu S, Ren X, et al. The screening value of RT‐LAMP and RT‐PCR in the diagnosis of COVID‐19: systematic review and meta‐analysis. J Virol Methods. 2022;300:114392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Teymouri M, Mollazadeh S, Mortazavi H, et al. Recent advances and challenges of RT‐PCR tests for the diagnosis of COVID‐19. Pathol Res Pract. 2021;221:153443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Aranha C, Patel V, Bhor V, Gogoi D. Cycle threshold values in RT‐PCR to determine dynamics of SARS‐CoV‐2 viral load: an approach to reduce the isolation period for COVID‐19 patients. J Med Virol. 2021;93(12):6794‐6797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Rahbari R, Moradi N, Abdi M. rRT‐PCR for SARS‐CoV‐2: analytical considerations. Clin Chim Acta. 2021;516:1‐7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Dao Thi VL, Herbst K, Boerner K, et al. A colorimetric RT‐LAMP assay and LAMP‐sequencing for detecting SARS‐CoV‐2 RNA in clinical samples. Sci Transl Med. 2020;12(556):eabc7075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Thompson D, Lei Y. Mini review: recent progress in RT‐LAMP enabled COVID‐19 detection. Sens Actuator A Phys. 2020;2(1):100017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Huang WE, Lim B, Hsu CC, et al. RT‐LAMP for rapid diagnosis of coronavirus SARS‐CoV‐2. Microb Biotechnol. 2020;13(4):950‐961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Rai P, Kumar BK, Deekshit VK, Karunasagar I, Karunasagar I. Detection technologies and recent developments in the diagnosis of COVID‐19 infection. Appl Microbiol Biotechnol. 2021;105(2):441‐455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Yang S, Pan X, Yuan D, Zeng P, Jia P. Cross‐disciplinary approaches to assist with nucleic acid testing for SARS‐CoV‐2. Appl Microbiol Biotechnol. 2021;105(16‐17):6291‐6299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Baek YH, Um J, Antigua KJC, et al. Development of a reverse transcription‐loop‐mediated isothermal amplification as a rapid early‐detection method for novel SARS‐CoV‐2. Emerg Microbes Infect. 2020;9(1):998‐1007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Talap J, Shen M, Yu L, Zeng S, Cai S. RT‐LAMP assay combining multi‐fluorescent probes for SARS‐CoV‐2 RNA detection and variant differentiation. Talanta. 2022;248:123644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Zhang Y, Tanner NA. Improving RT‐LAMP detection of SARS‐CoV‐2 RNA through primer set selection and combination. PLoS One. 2022;17(4):e0254324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Tamanaha E, Zhang Y, Tanner NA. Profiling RT‐LAMP tolerance of sequence variation for SARS‐CoV‐2 RNA detection. PLoS One. 2022;17(3):e0259610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Yang J, Hu X, Wang W, et al. RT‐LAMP assay for rapid detection of the R203M mutation in SARS‐CoV‐2 Delta variant. Emerg Microbes Infect. 2022;11(1):978‐987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Huang X, Tang G, Ismail N, Wang X. Developing RT‐LAMP assays for rapid diagnosis of SARS‐CoV‐2 in saliva. EBioMedicine. 2022;75:103736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Kellner MJ, Koob JG, Gootenberg JS, Abudayyeh OO, Zhang F. SHERLOCK: nucleic acid detection with CRISPR nucleases. Nat Protoc. 2019;14(10):2986‐3012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Gootenberg JS, Abudayyeh OO, Lee JW, et al. Nucleic acid detection with CRISPR‐Cas13a/C2c2. Science. 2017;356(6336):438‐442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Myhrvold C, Freije CA, Gootenberg JS, et al. Field‐deployable viral diagnostics using CRISPR‐Cas13. Science. 2018;360(6387):444‐448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Joung J, Ladha A, Saito M, et al. Point‐of‐care testing for COVID‐19 using SHERLOCK diagnostics. medRxiv. 2020. [Google Scholar]
- 75. Joung J, Ladha A, Saito M, et al. Detection of SARS‐CoV‐2 with SHERLOCK one‐pot testing. N Engl J Med. 2020;383(15):1492‐1494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Kumar KPA, Pumera M. 3D‐printing to mitigate COVID‐19 Pandemic. Adv Funct Mater. 2021;31(22):2100450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Kim J, Mayorga‐Martinez CC, Vyskočil J, Ruzek D, Pumera M. Plasmonic‐magnetic nanorobots for SARS‐CoV‐2 RNA detection through electronic readout. Appl Mater Today. 2022;27:101402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. STARSurg Collaborative and COVIDSurg Collaborative . Death following pulmonary complications of surgery before and during the SARS‐CoV‐2 pandemic. Br J Surg. 2021;108(12):1448‐1464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Crevillen AG, Mayorga‐Martinez CC, Vaghasiya JV, Pumera M. 3D‐printed SARS‐CoV‐2 RNA genosensing microfluidic system. Adv Mater Technol. 2022;7(6):2101121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Muñoz J, Pumera M. 3D‐Printed COVID‐19 immunosensors with electronic readout. J Chem Eng (Lausanne, Switzerland : 1996). 2021;425:131433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Mayorga‐Martinez CC, Vyskočil J, Novotný F, et al. Collective behavior of magnetic microrobots through immuno‐sandwich assay: On‐the‐fly COVID‐19 sensing. Appl Mater Today. 2022;26:101337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Gootenberg JS, Abudayyeh OO, Kellner MJ, Joung J, Collins JJ, Zhang F. Multiplexed and portable nucleic acid detection platform with Cas13, Cas12a, and Csm6. Science. 2018;360(6387):439‐444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Mallakpour S, Behranvand V, Hussain CM. Worldwide fight against COVID‐19 using nanotechnology, polymer science, and 3D printing technology. Polym Bull (Berlin, Germany). 2023;80(1):165‐183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Martin‐Noguerol T, Paulano‐Godino F, Menias CO, Luna A. Lessons learned from COVID‐19 and 3D printing. Am J Emerg Med. 2021;46:659‐660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85. Tino R, Moore R, Antoline S, et al. COVID‐19 and the role of 3D printing in medicine. 3D Print Med. 2020;6(1):11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Niranjan YC, Channabasavanna SG, Krishnapillai S, et al. The unprecedented role of 3D printing technology in fighting the COVID‐19 pandemic: a comprehensive review. Materials (Basel, Switzerland). 2022;15(19):6827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Hsiao WK, Lorber B, Paudel A. Can 3D printing of oral drugs help fight the current COVID‐19 pandemic (and similar crisis in the future)? Expert Opin Drug Deliv. 2020;17(7):899‐902. [DOI] [PubMed] [Google Scholar]
- 88. Perez‐Mañanes R, José SGS, Desco‐Menéndez M, et al. Application of 3D printing and distributed manufacturing during the first‐wave of COVID‐19 pandemic. Our experience at a third‐level university hospital. 3D Print Med. 2021;7(1):7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Zhou P, Yang XL, Wang XG, et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature. 2020;579(7798):270‐273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90. Pervez MT, Hasnain MJU, Abbas SH, Moustafa MF, Aslam N, Shah SSM. A comprehensive review of performance of next‐generation sequencing platforms. Biomed Res Int. 2022;2022:3457806. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 91. Tegally H, Wilkinson E, Giovanetti M, et al. Detection of a SARS‐CoV‐2 variant of concern in South Africa. Nature. 2021;592(7854):438‐443. [DOI] [PubMed] [Google Scholar]
- 92. Wang R, Qian C, Pang Y, et al. opvCRISPR: One‐pot visual RT‐LAMP‐CRISPR platform for SARS‐cov‐2 detection. Biosens Bioelectron. 2021;172:112766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93. Sharma A, Balda S, Apreja M, Kataria K, Capalash N, Sharma P. COVID‐19 diagnosis: current and future techniques. Int J Biol Macromol. 2021;193(Pt B):1835‐1844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94. Nandi SS, Lambe UP, Sawant SA, Gohil T, Deshpande J. Development of a RT‐LAMP assay for detection of SARS‐CoV‐2. Indian J Med Res. 2022;155(1):148‐155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95. El‐Kafrawy SA, El‐Daly MM, Hassan AM, Harakeh SM, Alandijany TA, Azhar EI. Rapid and reliable detection of SARS‐CoV‐2 using direct RT‐LAMP. Diagnostics (Basel, Switzerland). 2022;12(4):828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96. Sherrill‐Mix S, Hwang Y, Roche AM, et al. Detection of SARS‐CoV‐2 RNA using RT‐LAMP and molecular beacons. Genome Biol. 2021;22(1):169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Jain M, Olsen HE, Paten B, Akeson M. The Oxford Nanopore MinION: delivery of nanopore sequencing to the genomics community. Genome Biol. 2016;17(1):239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98. Head SR, Komori HK, LaMere SA, et al. Library construction for next‐generation sequencing: overviews and challenges. BioTechniques. 2014;56(2):61‐64, 66, 68, passim. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99. Brinkmann A, Ulm SL, Uddin S, et al. AmpliCoV: rapid whole‐genome sequencing using multiplex PCR amplification and real‐time oxford nanopore MinION sequencing enables rapid variant identification of SARS‐CoV‐2. Front Microbiol. 2021;12:651151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100. Samarakoon H, Punchihewa S, Senanayake A, et al. Genopo: a nanopore sequencing analysis toolkit for portable Android devices. Commun Biol. 2020;3(1):538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101. Ciuffreda L, Rodríguez‐Pérez H, Flores C. Nanopore sequencing and its application to the study of microbial communities. Comput Struct Biotechnol J. 2021;19:1497‐1511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102. Petersen LM, Martin IW, Moschetti WE, Kershaw CM, Tsongalis GJ. Third‐generation sequencing in the clinical laboratory: exploring the advantages and challenges of nanopore sequencing. J Clin Microbiol. 2019;58(1):e01315‐19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103. Wiener D, Schwartz S. The epitranscriptome beyond m(6)A. Nat Rev Genet. 2021;22(2):119‐131. [DOI] [PubMed] [Google Scholar]
- 104. Burgess HM, Depledge DP, Thompson L, et al. Targeting the m(6)A RNA modification pathway blocks SARS‐CoV‐2 and HCoV‐OC43 replication. Genes Dev. 2021;35(13‐14):1005‐1019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105. Zhang X, Hao H, Ma L, et al. Methyltransferase‐like 3 modulates severe acute respiratory syndrome coronavirus‐2 RNA N6‐methyladenosine modification and replication. mBio. 2021;12(4):e0106721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106. Ugolini C, Mulroney L, Leger A, et al. Nanopore ReCappable sequencing maps SARS‐CoV‐2 5' capping sites and provides new insights into the structure of sgRNAs. Nucleic Acids Res. 2022;50(6):3475‐3489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107. Wang D, Jiang A, Feng J, et al. The SARS‐CoV‐2 subgenome landscape and its novel regulatory features. Mol Cell. 2021;81(10):2135‐2147.e5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108. Tshiabuila D, Giandhari J, Pillay S, et al. Comparison of SARS‐CoV‐2 sequencing using the ONT GridION and the Illumina MiSeq. Bmc Genomics [Electronic Resource]. 2022;23(1):319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109. Kumar KR, Cowley MJ, Davis RL. Next‐generation sequencing and emerging technologies. Semin Thromb Hemost. 2019;45(7):661‐673. [DOI] [PubMed] [Google Scholar]
- 110. Mohamadian M, Chiti H, Shoghli A, Biglari S, Parsamanesh N, Esmaeilzadeh A. COVID‐19: virology, biology and novel laboratory diagnosis. J Gene Med. 2021;23(2):e3303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111. Charre C, Ginevra C, Sabatier M, et al. Evaluation of NGS‐based approaches for SARS‐CoV‐2 whole genome characterisation. Virus Evol. 2020;6(2):veaa075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112. Yan Y, Wu K, Chen J, et al. Rapid acquisition of high‐quality SARS‐CoV‐2 genome via amplicon‐oxford nanopore sequencing. Virol Sin. 2021;36(5):901‐912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113. Fu Y, Chen Q, Xiong M, et al. Clinical performance of nanopore targeted sequencing for diagnosing infectious diseases. Microbiol Spectr. 2022;10(2):e0027022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114. Peto L, Rodger G, Carter DP, et al. Diagnosis of SARS‐CoV‐2 infection with LamPORE, a high‐throughput platform combining loop‐mediated isothermal amplification and nanopore sequencing. J Clin Microbiol. 2021;59(6):e03271‐20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115. Moore SC, Penrice‐Randal R, Alruwaili M, et al. Amplicon‐based detection and sequencing of SARS‐CoV‐2 in nasopharyngeal swabs from patients with COVID‐19 and identification of deletions in the viral genome that encode proteins involved in interferon antagonism. Viruses. 2020;12(10):1164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116. Freed NE, Vlková M, Faisal MB, Silander OK. Rapid and inexpensive whole‐genome sequencing of SARS‐CoV‐2 using 1200 bp tiled amplicons and Oxford Nanopore Rapid Barcoding. Biol Methods Protoc. 2020;5(1):bpaa014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117. Brejová B, Boršová K, Hodorová V, et al. Nanopore sequencing of SARS‐CoV‐2: comparison of short and long PCR‐tiling amplicon protocols. PLoS One. 2021;16(10):e0259277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118. Lambisia AW, Mohammed KS, Makori TO, et al. Optimization of the SARS‐CoV‐2 ARTIC Network V4 primers and whole genome sequencing protocol. Front Med. 2022;9:836728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119. Arévalo MT, Karavis MA, Katoski SE, et al. A rapid, whole genome sequencing assay for detection and characterization of novel coronavirus (SARS‐CoV‐2) clinical specimens using nanopore sequencing. Front Microbiol. 2022;13:910955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120. Wagner GE, Totaro MG, Volland A, et al. A novel high‐throughput nanopore‐sequencing‐based strategy for rapid and automated s‐protein typing of SARS‐CoV‐2 variants. Viruses. 2021;13(12):2548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121. Jaworski E, Langsjoen RM, Mitchell B, et al. Tiled‐ClickSeq for targeted sequencing of complete coronavirus genomes with simultaneous capture of RNA recombination and minority variants. bioRxiv. 2021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122. Pembaur A, Sallard E, Weil PP, Ortelt J, Ahmad‐Nejad P, Postberg J. Simplified point‐of‐care full SARS‐CoV‐2 genome sequencing using nanopore technology. Microorganisms. 2021;9(12):2598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123. Baker DJ, Aydin A, Le‐Viet T, et al. CoronaHiT: high‐throughput sequencing of SARS‐CoV‐2 genomes. Genome Med. 2021;13(1):21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124. Fuchs M, Radulescu C, Tang M, et al. Mini‐XT, a miniaturized tagmentation‐based protocol for efficient sequencing of SARS‐CoV‐2. J Transl Med. 2022;20(1):105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125. Gaucher J, Vial G, Montellier E, et al. Intermittent hypoxia rewires the liver transcriptome and fires up fatty acids usage for mitochondrial respiration. Front Med. 2022;9:829979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126. Davis JJ, Long SW, Christensen PA, et al. Analysis of the ARTIC version 3 and version 4 SARS‐CoV‐2 primers and their impact on the detection of the G142D amino acid substitution in the spike protein. Microbiol Spectr. 2021;9(3):e0180321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127. Handrick S, Bestehorn‐Willmann M, Eckstein S, et al. Whole genome sequencing and phylogenetic classification of Tunisian SARS‐CoV‐2 strains from patients of the Military Hospital in Tunis. Virus Genes. 2020;56(6):767‐771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128. Kang K, Xu X, Dong R, Tian G. The risk perception of COVID‐19 and pandemic‐related behaviors: a moderated mediation model of political trust and self‐efficacy. Psychol Health Med. 2023:1‐14. [DOI] [PubMed] [Google Scholar]
- 129. Candela C, Raccagni AR, Bruzzesi E, et al. Human monkeypox experience in a tertiary level hospital in Milan, Italy, between May and October 2022: epidemiological features and clinical characteristics. Viruses. 2023;15(3):667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130. Malik S, Asghar M, Waheed Y. Mitigation measures to control the expected Mpox outbreak in a developing country—Pakistani scenario. Vaccines. 2023;11(3):502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131. Ptasinska A, Whalley C, Bosworth A, et al. Diagnostic accuracy of loop‐mediated isothermal amplification coupled to nanopore sequencing (LamPORE) for the detection of SARS‐CoV‐2 infection at scale in symptomatic and asymptomatic populations. Clin Microbiol Infect. 2021;27(9):1348.e1‐1348.e7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132. Morsli M, Anani H, Bréchard L, et al. LamPORE SARS‐CoV‐2 diagnosis and genotyping: a preliminary report. J Clin Virol. 2021;138:104815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133. Gafurov A, Baláž A, Amman F, et al. VirPool: model‐based estimation of SARS‐CoV‐2 variant proportions in wastewater samples. BMC Bioinf. 2022;23(1):551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134. Liu Y, Kearney J, Mahmoud M, Kille B, Sedlazeck FJ, Treangen TJ. Rescuing low frequency variants within intra‐host viral populations directly from Oxford nanopore sequencing data. bioRxiv. 2021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135. Liu Y, Kearney J, Mahmoud M, Kille B, Sedlazeck FJ, Treangen TJ. Rescuing low frequency variants within intra‐host viral populations directly from Oxford nanopore sequencing data. Nat Commun. 2022;13(1):1321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136. Liao YC, Chen FJ, Chuang MC, et al. High‐integrity sequencing of spike gene for SARS‐CoV‐2 variant determination. Int J Mol Sci. 2022;23(6):3257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137. Yang H, Gu F, Zhang L, Hua XS. Using generative adversarial networks for genome variant calling from low depth ONT sequencing data. Sci Rep. 2022;12(1):8725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138. Ciccolella S, Denti L, Bonizzoni P, Della Vedova G, Pirola Y, Previtali M. MALVIRUS: an integrated application for viral variant analysis. BMC Bioinf. 2022;22(Suppl 15):625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139. Yingtaweesittikul H, Ko K, Abdul Rahman N, Tan SYL, Nagarajan N, Suphavilai C. CalmBelt: Rapid SARS‐CoV‐2 Genome Characterization for Outbreak Tracking. Front Med. 2021;8:790662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140. Brandt C, Krautwurst S, Spott R, et al. poreCov—an easy to use, fast, and robust workflow for SARS‐CoV‐2 genome reconstruction via nanopore sequencing. Front Genet. 2021;12:711437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141. Lo CC, Shakya M, Connor R, et al. EDGE COVID‐19: a web platform to generate submission‐ready genomes from SARS‐CoV‐2 sequencing efforts. Bioinformatics. 2022;38(10):2700‐2704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142. Lang J. NanoCoV19: An analytical pipeline for rapid detection of severe acute respiratory syndrome coronavirus 2. Front Genet. 2022;13:1008792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143. Chandrakumar I, Gauthier NPG, Nelson C, et al. BugSplit enables genome‐resolved metagenomics through highly accurate taxonomic binning of metagenomic assemblies. Commun Biol. 2022;5(1):151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144. Hunt M, Swann J, Constantinides B, Fowler PW, Iqbal Z. ReadItAndKeep: rapid decontamination of SARS‐CoV‐2 sequencing reads. Bioinformatics. 2022;38(12):3291‐3293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145. Dong X, Penrice‐Randal R, Goldswain H, et al. Analysis of SARS‐CoV‐2 known and novel subgenomic mRNAs in cell culture, animal model, and clinical samples using LeTRS, a bioinformatic tool to identify unique sequence identifiers. GigaScience. 2022;11:giac045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146. Ferguson JM, Gamaarachchi H, Nguyen T, et al. InterARTIC: an interactive web application for whole‐genome nanopore sequencing analysis of SARS‐CoV‐2 and other viruses. Bioinformatics. 2021;38(5):1443‐1446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147. Valencia‐Valencia DE, Lopez‐Alvarez D, Rivera‐Franco N, et al. PredictION: a predictive model to establish the performance of Oxford sequencing reads of SARS‐CoV‐2. PeerJ. 2022;10:e14425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148. Yu R, Cai D, Sun Y. AccuVIR: an ACCUrate VIRal genome assembly tool for third‐generation sequencing data. Bioinformatics. 2023;39(1):btac827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149. Pathak AK, Mishra GP, Uppili B, et al. Spatio‐temporal dynamics of intra‐host variability in SARS‐CoV‐2 genomes. Nucleic Acids Res. 2022;50(3):1551‐1561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150. Jackson CB, Farzan M, Chen B, Choe H. Mechanisms of SARS‐CoV‐2 entry into cells. Nat Rev Mol Cell Biol. 2022;23(1):3‐20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151. Cherif E, Thiam FS, Salma M, et al. ONTdeCIPHER: an amplicon‐based nanopore sequencing pipeline for tracking pathogen variants. Bioinformatics. 2022;38(7):2033‐2035. [DOI] [PubMed] [Google Scholar]
- 152. Chiara M, D'Erchia AM, Gissi C, et al. Next generation sequencing of SARS‐CoV‐2 genomes: challenges, applications and opportunities. Briefings Bioinf. 2021;22(2):616‐630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153. Sun X, Song L, Yang W, et al. Nanopore sequencing and its clinical applications. Methods Mol Biol. 2020;2204:13‐32. [DOI] [PubMed] [Google Scholar]
- 154. Ewing AD, Smits N, Sanchez‐Luque FJ, et al. Nanopore sequencing enables comprehensive transposable element epigenomic profiling. Mol Cell. 2020;80(5):915‐928.e5. [DOI] [PubMed] [Google Scholar]
- 155. Lee I, Razaghi R, Gilpatrick T, et al. Simultaneous profiling of chromatin accessibility and methylation on human cell lines with nanopore sequencing. Nat Methods. 2020;17(12):1191‐1199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156. Stevanovski I, Chintalaphani SR, Gamaarachchi H, et al. Comprehensive genetic diagnosis of tandem repeat expansion disorders with programmable targeted nanopore sequencing. Sci Adv. 2022;8(9):eabm5386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157. Tegally H, San JE, Cotten M, et al. The evolving SARS‐CoV‐2 epidemic in Africa: Insights from rapidly expanding genomic surveillance. Science. 2022;378(6615):eabq5358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158. Cheng VCC, Ip JD, Chu AWH, et al. Rapid spread of severe acute respiratory syndrome coronavirus 2 (SARS‐CoV‐2) omicron subvariant BA.2 in a single‐source community outbreak. Clin Infect Dis. 2022;75(1):e44‐e49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159. Sahajpal NS, Mondal AK, Ananth S, et al. Clinical validation of a multiplex PCR‐based detection assay using saliva or nasopharyngeal samples for SARS‐Cov‐2, influenza A and B. Sci Rep. 2022;12(1):3480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160. Wang Y, Chen D, Zhu C, et al. Genetic surveillance of five SARS‐CoV‐2 clinical samples in henan province using nanopore sequencing. Front Immunol. 2022;13:814806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161. Wang M, Fu A, Hu B, et al. Nanopore targeted sequencing for the accurate and comprehensive detection of SARS‐CoV‐2 and other respiratory viruses. Small. 2021;17(32):e2104078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162. Lu J, Peng J, Xiong Q, et al. Clinical, immunological and virological characterization of COVID‐19 patients that test re‐positive for SARS‐CoV‐2 by RT‐PCR. EBioMedicine. 2020;59:102960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163. Harvey WT, Carabelli AM, Jackson B, et al. SARS‐CoV‐2 variants, spike mutations and immune escape. Nat Rev Microbiol. 2021;19(7):409‐424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164. Barbé L, Schaeffer J, Besnard A, et al. SARS‐CoV‐2 whole‐genome sequencing using oxford nanopore technology for variant monitoring in wastewaters. Front Microbiol. 2022;13:889811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165. Andeweg SP, Vennema H, Veldhuijzen I, et al. Elevated risk of infection with SARS‐CoV‐2 Beta, Gamma, and Delta variant compared to Alpha variant in vaccinated individuals. Sci Transl Med. 2022:eabn4338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166. Li J, Wang H, Mao L, et al. Rapid genomic characterization of SARS‐CoV‐2 viruses from clinical specimens using nanopore sequencing. Sci Rep. 2020;10(1):17492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167. Cusi MG, Pinzauti D, Gandolfo C, Anichini G, Pozzi G, Santoro F. Whole‐genome sequence of SARS‐CoV‐2 isolate siena‐1/2020. Microbiol Resour Announc. 2020;9(39):e00944‐20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168. Sharma S, Dash PK, Sharma SK, et al. Emergence and expansion of highly infectious spike protein D614G mutant SARS‐CoV‐2 in central India. Sci Rep. 2021;11(1):18126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169. Zhao N, Zhou N, Fan H, et al. Mutations and phylogenetic analyses of SARS‐CoV‐2 among imported COVID‐19 from abroad in Nanjing, China. Front Microbiol. 2022;13:851323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170. Colson P, Delerce J, Burel E, et al. Emergence in southern France of a new SARS‐CoV‐2 variant harbouring both N501Y and E484K substitutions in the spike protein. Arch Virol. 2022;167(4):1185‐1190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171. Nilgiriwala K, Mandal A, Patel G, et al. Genome sequences of five SARS‐CoV‐2 variants from mumbai, india, obtained by nanopore sequencing. Microbiol Resour Announc. 2021;10(15):e00231‐21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172. Nilgiriwala K, Kadam P, Patel G, et al. Genomics of postvaccination SARS‐CoV‐2 infections during the Delta dominated second wave of COVID‐19 pandemic, from Mumbai Metropolitan Region (MMR), India. J Med Virol. 2022;94(9):4206‐4215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173. Laiton‐Donato K, Franco‐Muñoz C, Álvarez‐Díaz DA, et al. Characterization of the emerging B.1.621 variant of interest of SARS‐CoV‐2. Infect Genet Evol. 2021;95:105038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174. Afrad MH, Khan MH, Rahman SIA, et al. Genome sequences of 15 SARS‐CoV‐2 sublineage B.1.617.2 Strains in Bangladesh. Microbiol Resour Announc. 2021;10(28):e0056021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175. Mannan A, Mehedi HMH, Rob MA, et al. Genome Sequences of SARS‐CoV‐2 Sublineage B.1.617.2 strains from 12 children in Chattogram, Bangladesh. Microbiol Resour Announc. 2021;10(42):e0091221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176. Brinkac L, Diepold S, Mitchell S, et al. SARS‐CoV‐2 Delta variant isolates from vaccinated individuals. Bmc Genomics [Electronic Resource]. 2022;23(1):417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177. Jenjaroenpun P, Wanchai V, Ono‐Moore KD, et al. Two SARS‐CoV‐2 genome sequences of isolates from rural U.S. patients harboring the D614G mutation, obtained using nanopore sequencing. Microbiol Resour Announc. 2020;10(1):e01109‐20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178. Stüder F, Petit JL, Engelen S, Mendoza‐Parra MA. Real‐time SARS‐CoV‐2 diagnostic and variants tracking over multiple candidates using nanopore DNA sequencing. Sci Rep. 2021;11(1):15869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179. Munis AM, Andersson M, Mobbs A, Hyde SC, Gill DR. Genomic diversity of SARS‐CoV‐2 in Oxford during United Kingdom's first national lockdown. Sci Rep. 2021;11(1):21484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180. Paré B, Rozendaal M, Morin S, et al. Patient health records and whole viral genomes from an early SARS‐CoV‐2 outbreak in a Quebec hospital reveal features associated with favorable outcomes. PLoS One. 2021;16(12):e0260714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181. Paré B, Rozendaal M, Morin S, et al. Genomic epidemiology and associated clinical outcomes of a SARS‐CoV‐2 outbreak in a general adult hospital in Quebec. medRxiv : the preprint server for health sciences. 2021. [Google Scholar]
- 182. Martinez M, Nguyen PV, Su M, et al. SARS‐CoV‐2 variants in paraguay: detection and surveillance with an economical and scalable molecular protocol. Viruses. 2022;14(5):873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183. Dächert C, Muenchhoff M, Graf A, et al. Rapid and sensitive identification of omicron by variant‐specific PCR and nanopore sequencing: paradigm for diagnostics of emerging SARS‐CoV‐2 variants. Med Microbiol Immunol (Berl). 2022;211(1):71‐77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184. Manouana GP, Nzamba Maloum M, Bikangui R, et al. Emergence of B.1.1.318 SARS‐CoV‐2 viral lineage and high incidence of alpha B.1.1.7 variant of concern in the Republic of Gabon. Int J Infect Dis. 2022;114:151‐154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185. Rios G, Lacoux C, Leclercq V, et al. Monitoring SARS‐CoV‐2 variants alterations in nice neighborhoods by wastewater nanopore sequencing. Lancet Reg Health Eur. 2021;10:100202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186. Bin Manjur OH, Afrad MH, Khan MH, et al. Genome sequences of 25 SARS‐CoV‐2 sublineage B.1.1.529 omicron strains in Bangladesh. Microbiol Resour Announc. 2022;11(4):e0011922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187. Laamarti M, Chemao‐Elfihri MW, Kartti S, et al. Genome sequences of six SARS‐CoV‐2 strains isolated in Morocco, Obtained Using Oxford Nanopore MinION technology. Microbiol Resour Announc. 2020;9(32):e00767‐20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188. Lopez‐Alvarez D, Parra B, Cuellar WJ. Genome sequence of SARS‐CoV‐2 isolate Cali‐01, from Colombia, Obtained Using Oxford Nanopore MinION sequencing. Microbiol Resour Announc. 2020;9(26):e00573‐20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189. Au CH, Chan WS, Lam HY, et al. Genome sequences of SARS‐CoV‐2 strains detected in Hong Kong. Microbiol Resour Announc. 2020;9(31):e00697‐20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190. Biazzo M, Madeddu S, Elnifro E, et al. Genome sequences of 10 SARS‐CoV‐2 viral strains obtained by nanopore sequencing of nasopharyngeal swabs in Malta. Microbiol Resour Announc. 2021;10(4):e01375‐20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191. Ntagereka PB, Oyola SO, Baenyi SP, et al. Whole‐genome sequencing of SARS‐CoV‐2 reveals diverse mutations in circulating Alpha and Delta variants during the first, second, and third waves of COVID‐19 in South Kivu, east of the Democratic Republic of the Congo. Int J Infect Dis. 2022;122:136‐143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192. Slavov SN, Giovanetti M, Dos Santos Bezerra R, et al. Molecular surveillance of the on‐going SARS‐COV‐2 epidemic in Ribeirao Preto City, Brazil. Infect Genet Evol. 2021;93:104976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193. Lin L, Zhang J, Rogers J, et al. The dynamic change of SARS‐CoV‐2 variants in Sierra Leone. Infect Genet Evol. 2022;98:105208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194. Prasetyoputri A, Dharmayanthi AB, Iryanto SB, et al. The dynamics of circulating SARS‐CoV‐2 lineages in Bogor and surrounding areas reflect variant shifting during the first and second waves of COVID‐19 in Indonesia. PeerJ. 2022;10:e13132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195. Walker A, Houwaart T, Wienemann T, et al. Genetic structure of SARS‐CoV‐2 reflects clonal superspreading and multiple independent introduction events, North‐Rhine Westphalia, Germany, February and March 2020. Euro Surveill. 2020;25(22):2000746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196. Marquez S, Prado‐Vivar B, Guadalupe JJ, et al. Genome sequencing of the first SARS‐CoV‐2 reported from patients with COVID‐19 in Ecuador. medRxiv : the preprint server for health sciences. 2020. [Google Scholar]
- 197. Kumar P, Pandey R, Sharma P, et al. Integrated genomic view of SARS‐CoV‐2 in India. Wellcome Open Res. 2020;5:184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198. Thielen PM, Wohl S, Mehoke T, et al. Genomic diversity of SARS‐CoV‐2 during early introduction into the Baltimore‐Washington metropolitan area. JCI insight. 2021;6(6):e144350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199. Engelbrecht S, Delaney K, Kleinhans B, et al. Multiple early introductions of SARS‐CoV‐2 to Cape Town, South Africa. Viruses. 2021;13(3):526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200. van Hensbergen M, den Heijer CDJ, Wolffs P, et al. COVID‐19: first long‐term care facility outbreak in the Netherlands following cross‐border introduction from Germany, March 2020. BMC Infect Dis. 2021;21(1):418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201. Tamim S, Trovao NS, Thielen P, et al. Genetic and evolutionary analysis of SARS‐CoV‐2 circulating in the region surrounding Islamabad, Pakistan. Infect Genet Evol. 2021;94:105003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202. Yakovleva A, Kovalenko G, Redlinger M, et al. Tracking SARS‐COV‐2 variants using nanopore sequencing in Ukraine in summer 2021. Res Sq. 2021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203. Bianco L, Moser M, Silverj A, et al. On the origin and propagation of the COVID‐19 outbreak in the Italian Province of Trento, a Tourist Region of Northern Italy. Viruses. 2022;14(3):580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204. Nasir A, Bukhari AR, Trovão NS, et al. Evolutionary history and introduction of SARS‐CoV‐2 Alpha VOC/B.1.1.7 in Pakistan through international travelers. Virus Evol. 2022;8(1):veac020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205. Avetyan D, Hakobyan S, Nikoghosyan M, et al. Molecular analysis of SARS‐CoV‐2 lineages in armenia. Viruses. 2022;14(5):1074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206. Borrega R, Nelson DKS, Koval AP, et al. Cross‐reactive antibodies to SARS‐CoV‐2 and MERS‐CoV in pre‐COVID‐19 blood samples from Sierra Leoneans. Viruses. 2021;13(11):2325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207. Mukry SN, Ahmed S, Raza Bukhari A, et al. Sequencing of SARS‐CoV‐2 in local transmission cases through Oxford Nanopore MinION platform from Karachi Pakistan. J Infect Dev Ctries. 2021;15(10):1376‐1383. [DOI] [PubMed] [Google Scholar]
- 208. Dharmadhikari T, Rajput V, Yadav R, et al. High throughput sequencing based direct detection of SARS‐CoV‐2 fragments in wastewater of Pune, West India. Sci Total Environ. 2022;807(Pt 3):151038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209. Goletic S, Goletic T, Softic A, et al. The evidence of SARS‐CoV‐2 human‐to‐pets transmission in household settings in Bosnia and Herzegovina. Front Genet. 2022;13:839205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210. Márquez S, Prado‐Vivar B, José Guadalupe J, et al. SARS‐CoV‐2 genome sequencing from COVID‐19 in Ecuadorian patients: a whole country analysis. medRxiv : the preprint server for health sciences. 2021. [Google Scholar]
- 211. La Rosa G, Brandtner D, Bonanno Ferraro G, et al. Wastewater surveillance of SARS‐CoV‐2 variants in October‐November 2022 in Italy: detection of XBB.1, BA.2.75 and rapid spread of the BQ.1 lineage. Sci Total Environ. 2023;873:162339. [DOI] [PubMed] [Google Scholar]
- 212. Yang ZK, Pan L, Zhang Y, Luo H, Gao F. Data‐driven identification of SARS‐CoV‐2 subpopulations using PhenoGraph and binary‐coded genomic data. Briefings Bioinf. 2021;22(6):bbab307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213. Qin L, Ding X, Li Y, Chen Q, Meng J, Jiang T. Co‐mutation modules capture the evolution and transmission patterns of SARS‐CoV‐2. Briefings Bioinf. 2021;22(6):bbab222. [DOI] [PubMed] [Google Scholar]
- 214. Løvestad AH, Jørgensen SB, Handal N, Ambur OH, Aamot HV. Investigation of intra‐hospital SARS‐CoV‐2 transmission using nanopore whole‐genome sequencing. J Hosp Infect. 2021;111:107‐116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215. Rasmussen LD, Richter SR, Midgley SE, Franck KT. Detecting SARS‐CoV‐2 Omicron B.1.1.529 variant in wastewater samples by using nanopore sequencing. Emerg Infect Dis. 2022;28(6):1296‐1298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216. Walker A, Houwaart T, Finzer P, et al. Characterization of severe acute respiratory syndrome coronavirus 2 (SARS‐CoV‐2) infection clusters based on integrated genomic surveillance, outbreak analysis and contact tracing in an urban setting. Clin Infect Dis. 2022;74(6):1039‐1046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217. Wang T, Wang C, Myshkevych Y, Mantilla‐Calderon D, Talley E, Hong PY. SARS‐CoV‐2 wastewater‐based epidemiology in an enclosed compound: A 2.5‐year survey to identify factors contributing to local community dissemination. Sci Total Environ. 2023;875:162466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218. Ramírez JD, Muñoz M, Ballesteros N, et al. Phylogenomic evidence of reinfection and persistence of SARS‐CoV‐2: first report from Colombia. Vaccines. 2021;9(3):282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219. Li Y, Jiang Y, Li Z, et al. Both simulation and sequencing data reveal coinfections with multiple SARS‐CoV‐2 variants in the COVID‐19 pandemic. Comput Struct Biotechnol J. 2022;20:1389‐1401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220. Gu W, Miller S, Chiu CY. Clinical metagenomic next‐generation sequencing for pathogen detection. Annu Rev Pathol. 2019;14:319‐338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221. Mostafa HH, Fissel JA, Fanelli B, et al. Metagenomic next‐generation sequencing of nasopharyngeal specimens collected from confirmed and suspect COVID‐19 patients. mBio. 2020;11(6):e01969‐20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222. Lewandowski K, Xu Y, Pullan ST, et al. Metagenomic nanopore sequencing of influenza virus direct from clinical respiratory samples. J Clin Microbiol. 2019;58(1):e00963‐19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223. Bradley ES, Zeamer AL, Bucci V, et al. Oropharyngeal microbiome profiled at admission is predictive of the need for respiratory support among COVID‐19 patients. Front Microbiol. 2022;13:1009440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224. Shi YL, He MZ, Han MZ, et al. Characterization of altered oropharyngeal microbiota in hospitalized patients with mild SARS‐CoV‐2 infection. Front Cell Infect Microbiol. 2022;12:824578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225. Prasad P, Mahapatra S, Mishra R, et al. Long‐read 16S‐seq reveals nasopharynx microbial dysbiosis and enrichment of Mycobacterium and Mycoplasma in COVID‐19 patients: a potential source of co‐infection. Mol Omics. 2022;18(6):490‐505. [DOI] [PubMed] [Google Scholar]
- 226. Jitvaropas R, Mayuramart O, Sawaswong V, Kaewsapsak P, Payungporn S. Classification of salivary bacteriome in asymptomatic COVID‐19 cases based on long‐read nanopore sequencing. Exp Biol Med (Maywood, NJ). 2022:15353702221118091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227. Charalampous T, Alcolea‐Medina A, Snell LB, et al. Evaluating the potential for respiratory metagenomics to improve treatment of secondary infection and detection of nosocomial transmission on expanded COVID‐19 intensive care units. Genome Med. 2021;13(1):182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228. Snell LB, Alcolea‐Medina A, Charalampous T, et al. Real‐time whole genome sequencing to guide patient‐tailored therapy of severe acute respiratory syndrome coronavirus 2 infection. Clin Infect Dis. 2023;76(6):1125‐1128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229. Gomez‐Simmonds A, Annavajhala MK, McConville TH, et al. Carbapenemase‐producing Enterobacterales causing secondary infections during the COVID‐19 crisis at a New York City hospital. J Antimicrob Chemother. 2021;76(2):380‐384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230. de Wit E, van Doremalen N, Falzarano D, Munster VJ. SARS and MERS: recent insights into emerging coronaviruses. Nat Rev Microbiol. 2016;14(8):523‐534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231. Aljabr W, Alruwaili M, Penrice‐Randal R, et al. Amplicon and metagenomic analysis of Middle East Respiratory Syndrome (MERS) coronavirus and the microbiome in patients with severe MERS. mSphere. 2021;6(4):e0021921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232. Bi C, Ramos‐Mandujano G, Tian Y, et al. Simultaneous detection and mutation surveillance of SARS‐CoV‐2 and multiple respiratory viruses by rapid field‐deployable sequencing. Med (New York, NY). 2021;2(6):689‐700.e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233. Brinkmann A, Uddin S, Ulm SL, et al. RespiCoV: Simultaneous identification of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS‐CoV‐2) and 46 respiratory tract viruses and bacteria by amplicon‐based Oxford‐Nanopore MinION sequencing. PLoS One. 2022;17(3):e0264855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234. Garalde DR, Snell EA, Jachimowicz D, et al. Highly parallel direct RNA sequencing on an array of nanopores. Nat Methods. 2018;15(3):201‐206. [DOI] [PubMed] [Google Scholar]
- 235. Yang SL, DeFalco L, Anderson DE, et al. Comprehensive mapping of SARS‐CoV‐2 interactions in vivo reveals functional virus‐host interactions. Nat Commun. 2021;12(1):5113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236. Liang J, Shi J, Chen S, et al. How the replication and transcription complex functions in jumping transcription of SARS‐CoV‐2. Front Genet. 2022;13:904513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237. Davidson AD, Williamson MK, Lewis S, et al. Characterisation of the transcriptome and proteome of SARS‐CoV‐2 reveals a cell passage induced in‐frame deletion of the furin‐like cleavage site from the spike glycoprotein. Genome Med. 2020;12(1):68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238. Liu Z, Zheng H, Lin H, et al. Identification of common deletions in the spike protein of severe acute respiratory syndrome coronavirus 2. J Virol. 2020;94(17):e00790‐20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 239. Coutard B, Valle C, de Lamballerie X, Canard B, Seidah NG, Decroly E. The spike glycoprotein of the new coronavirus 2019‐nCoV contains a furin‐like cleavage site absent in CoV of the same clade. Antiviral Res. 2020;176:104742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240. Chang JJ, Gleeson J, Rawlinson D, et al. Long‐read RNA sequencing identifies polyadenylation elongation and differential transcript usage of host transcripts during SARS‐CoV‐2 in vitro infection. Front Immunol. 2022;13:832223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241. Smits N, Rasmussen J, Bodea GO, et al. No evidence of human genome integration of SARS‐CoV‐2 found by long‐read DNA sequencing. Cell Rep. 2021;36(7):109530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242. van Dijk EL, Jaszczyszyn Y, Naquin D, Thermes C. The third revolution in sequencing technology. Trends Genet: TIG. 2018;34(9):666‐681. [DOI] [PubMed] [Google Scholar]
- 243. Chang JJ, Rawlinson D, Pitt ME, et al. Transcriptional and epi‐transcriptional dynamics of SARS‐CoV‐2 during cellular infection. Cell Rep. 2021;35(6):109108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244. Xu P, Ge R. Roles and drug development of METTL3 (methyltransferase‐like 3) in anti‐tumor therapy. Eur J Med Chem. 2022;230:114118. [DOI] [PubMed] [Google Scholar]
- 245. Liu R, Ou L, Sheng B, et al. Mixed‐weight neural bagging for detecting m(6)A modifications in SARS‐CoV‐2 RNA sequencing. IEEE Trans Biomed Eng. 2022;69(8):2557‐2568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246. Campos JHC, Maricato JT, Braconi CT, Antoneli F, Janini LMR, Briones MRS. Direct RNA sequencing reveals SARS‐CoV‐2 m6A sites and possible differential DRACH motif methylation among variants. Viruses. 2021;13(11):2108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247. Mulroney L, Wulf MG, Schildkraut I, et al. Identification of high‐confidence human poly(A) RNA isoform scaffolds using nanopore sequencing. RNA (New York, NY). 2022;28(2):162‐176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248. Workman RE, Tang AD, Tang PS, et al. Nanopore native RNA sequencing of a human poly(A) transcriptome. Nat Methods. 2019;16(12):1297‐1305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249. Walker AP, Fan H, Keown JR, Knight ML, Grimes JM, Fodor E. The SARS‐CoV‐2 RNA polymerase is a viral RNA capping enzyme. Nucleic Acids Res. 2021;49(22):13019‐13030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250. Li D, Zhang J, Li J. Primer design for quantitative real‐time PCR for the emerging coronavirus SARS‐CoV‐2. Theranostics. 2020;10(16):7150‐7162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251. Chan WM, Ip JD, Chu AW, et al. Identification of nsp1 gene as the target of SARS‐CoV‐2 real‐time RT‐PCR using nanopore whole‐genome sequencing. J Med Virol. 2020;92(11):2725‐2734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 252. Xia X. Domains and functions of spike protein in Sars‐Cov‐2 in the context of vaccine design. Viruses. 2021;13(1):109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253. Kumavath R, Barh D, Andrade BS, et al. The spike of SARS‐CoV‐2: uniqueness and applications. Front Immunol. 2021;12:663912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254. Franco‐Muñoz C, Álvarez‐Díaz DA, Laiton‐Donato K, et al. Substitutions in spike and nucleocapsid proteins of SARS‐CoV‐2 circulating in South America. Infect Genet Evol. 2020;85:104557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255. Ip JD, Kok KH, Chan WM, et al. Intra‐host non‐synonymous diversity at a neutralizing antibody epitope of SARS‐CoV‐2 spike protein N‐terminal domain. Clin Microbiol Infect. 2021;27(9):1350.e1‐1350.e5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 256. Imai M, Iwatsuki‐Horimoto K, Hatta M, et al. Syrian hamsters as a small animal model for SARS‐CoV‐2 infection and countermeasure development. Proc Nat Acad Sci USA. 2020;117(28):16587‐16595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 257. Harris RA, Raveendran M, Lyfoung DT, et al. Construction of a new chromosome‐scale, long‐read reference genome assembly for the Syrian hamster, Mesocricetus auratus. GigaScience. 2022;11:giac039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258. Cleary SJ, Pitchford SC, Amison RT, et al. Animal models of mechanisms of SARS‐CoV‐2 infection and COVID‐19 pathology. Br J Pharmacol. 2020;177(21):4851‐4865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 259. Xu Y, Lewandowski K, Downs LO, et al. Nanopore metagenomic sequencing of influenza virus directly from respiratory samples: diagnosis, drug resistance and nosocomial transmission, United Kingdom, 2018/19 influenza season. Euro Surveill. 2021;26(27):2000004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 260. Quick J, Grubaugh ND, Pullan ST, et al. Multiplex PCR method for MinION and Illumina sequencing of Zika and other virus genomes directly from clinical samples. Nat Protoc. 2017;12(6):1261‐1276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 261. Stubbs SCB, Blacklaws BA, Yohan B, et al. Assessment of a multiplex PCR and nanopore‐based method for dengue virus sequencing in Indonesia. Virology journal. 2020;17(1):24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262. Jia L, Jiang M, Wu K, et al. Nanopore sequencing of African swine fever virus. Sci China Life Sci. 2020;63(1):160‐164. [DOI] [PubMed] [Google Scholar]
- 263. Ambike S, Cheng CC, Feuerherd M, et al. Targeting genomic SARS‐CoV‐2 RNA with siRNAs allows efficient inhibition of viral replication and spread. Nucleic Acids Res. 2022;50(1):333‐349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 264. Della Bartola M, Byrne S, Mullins E. Characterization of potato virus Y isolates and assessment of nanopore sequencing to detect and genotype potato viruses. Viruses. 2020;12(4):478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 265. Heikema AP, Horst‐Kreft D, Boers SA, et al. Comparison of illumina versus nanopore 16S rRNA gene sequencing of the human nasal microbiota. Genes. 2020;11(9):1105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 266. Smith C, Halse TA, Shea J, et al. Assessing nanopore sequencing for clinical diagnostics: a comparison of next‐generation sequencing (NGS) methods for Mycobacterium tuberculosis. J Clin Microbiol. 2020;59(1):e00583‐20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 267. Imanian B, Donaghy J, Jackson T, et al. The power, potential, benefits, and challenges of implementing high‐throughput sequencing in food safety systems. NPJ Sci Food. 2022;6(1):35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 268. Vandenbogaert M, Kwasiborski A, Gonofio E, et al. Nanopore sequencing of a monkeypox virus strain isolated from a pustular lesion in the Central African Republic. Sci Rep. 2022;12(1):10768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 269. Cao Y, Liu H, Yan Y, Liu W, Liu D, Li J. Discriminating clonotypes of influenza A virus genes by nanopore sequencing. Int J Mol Sci. 2021;22(18):10069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 270. Yip CC, Chan WM, Ip JD, et al. Nanopore sequencing reveals novel targets for detection and surveillance of human and avian influenza A viruses. J Clin Microbiol. 2020;58(5):e02127‐19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 271. Sun K, Liu Y, Zhou X, et al. Nanopore sequencing technology and its application in plant virus diagnostics. Front Microbiol. 2022;13:939666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 272. Roossinck MJ. Deep sequencing for discovery and evolutionary analysis of plant viruses. Virus Res. 2017;239:82‐86. [DOI] [PubMed] [Google Scholar]
- 273. Maina S, Norton SL, McQueen VL, King S, Rodoni B. Oxford nanopore sequencing reveals an exotic broad bean mottle virus genome within Australian grains post‐entry quarantine. Microbiol Resour Announc. 2022;11(7):e0021122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 274. Lee HJ, Cho IS, Jeong RD. Nanopore metagenomics sequencing for rapid diagnosis and characterization of lily viruses. Plant Pathol J. 2022;38(5):503‐512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 275. Amoia SS, Minafra A, Nicoloso V, Loconsole G, Chiumenti M. A new jasmine virus C isolate identified by nanopore sequencing is associated to yellow mosaic symptoms of jasminum officinale in Italy. Plants (Basel, Switzerland). 2022;11(3):309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 276. Fellers JP, Webb C, Fellers MC, Shoup Rupp J, De Wolf E. Wheat virus identification within infected tissue using nanopore sequencing technology. Plant Dis. 2019;103(9):2199‐2203. [DOI] [PubMed] [Google Scholar]
- 277. Eltom KH, Althoff AC, Hansen S, et al. Differentiation of capripox viruses by nanopore sequencing. Vaccines. 2021;9(4):351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 278. Toh X, Wang Y, Rajapakse MP, et al. Use of nanopore sequencing to characterize African horse sickness virus (AHSV) from the African horse sickness outbreak in Thailand in 2020. Transbound Emerg Dis. 2022;69(3):1010‐1019. [DOI] [PubMed] [Google Scholar]
- 279. Pavliscak LA, Nirmala J, Singh VK, et al. Tracing viral transmission and evolution of bovine leukemia virus through long read Oxford nanopore sequencing of the proviral genome. Pathogens (Basel, Switzerland). 2021;10(9):1191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 280. Zhang R, Wang P, Ma X, et al. Nanopore‐based direct RNA‐sequencing reveals a high‐resolution transcriptional landscape of porcine reproductive and respiratory syndrome virus. Viruses. 2021;13(12):2531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 281. Latorre‐Pérez A, Gimeno‐Valero H, Tanner K, Pascual J, Vilanova C, Porcar M. A round trip to the desert: in situ nanopore sequencing informs targeted bioprospecting. Front Microbiol. 2021;12:768240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 282. Sutton MA, Burton AS, Zaikova E, et al. Radiation tolerance of nanopore sequencing technology for life detection on Mars and Europa. Sci Rep. 2019;9(1):5370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 283. Burton AS, Stahl SE, John KK, et al. Off earth identification of bacterial populations using 16S rDNA nanopore sequencing. Genes. 2020;11(1):76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 284. Manos J. The human microbiome in disease and pathology. APMIS. 2022;130(12):690‐705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 285. Liu S, Gao J, Zhu M, Liu K, Zhang HL. Gut microbiota and dysbiosis in Alzheimer's disease: implications for pathogenesis and treatment. Mol Neurobiol. 2020;57(12):5026‐5043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 286. Szoboszlay M, Schramm L, Pinzauti D, Scerri J, Sandionigi A, Biazzo M. Nanopore is preferable over illumina for 16S amplicon sequencing of the gut microbiota when species‐level taxonomic classification, accurate estimation of richness, or focus on rare taxa is required. Microorganisms. 2023;11(3):804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 287. Oberle A, Urban L, Falch‐Leis S, et al. 16S rRNA long‐read nanopore sequencing is feasible and reliable for endometrial microbiome analysis. Reprod Biomed Online. 2021;42(6):1097‐1107. [DOI] [PubMed] [Google Scholar]
- 288. Tourancheau A, Mead EA, Zhang XS, Fang G. Discovering multiple types of DNA methylation from bacteria and microbiome using nanopore sequencing. Nat Methods. 2021;18(5):491‐498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 289. Nakagawa H, Fujita M. Whole genome sequencing analysis for cancer genomics and precision medicine. Cancer Sci. 2018;109(3):513‐522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 290. Hanahan D. Hallmarks of cancer: new dimensions. Cancer Discov. 2022;12(1):31‐46. [DOI] [PubMed] [Google Scholar]
- 291. Vockley JG, Niederhuber JE. Diagnosis and treatment of cancer using genomics. BMJ (Clinical research ed). 2015;350:h1832. [DOI] [PubMed] [Google Scholar]
- 292. Leger A, Amaral PP, Pandolfini L, et al. RNA modifications detection by comparative Nanopore direct RNA sequencing. Nat Commun. 2021;12(1):7198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 293. Fujimoto A, Wong JH, Yoshii Y, et al. Whole‐genome sequencing with long reads reveals complex structure and origin of structural variation in human genetic variations and somatic mutations in cancer. Genome Med. 2021;13(1):65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 294. Aganezov S, Goodwin S, Sherman RM, et al. Comprehensive analysis of structural variants in breast cancer genomes using single‐molecule sequencing. Genome Res. 2020;30(9):1258‐1273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 295. Qu H, Wang Z, Zhang Y, et al. Long‐read nanopore sequencing identifies mismatch repair‐deficient related genes with alternative splicing in colorectal cancer. Dis Markers. 2022;2022:4433270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 296. You Y, Clark MB, Shim H. NanoSplicer: accurate identification of splice junctions using Oxford Nanopore sequencing. Bioinformatics. 2022;38(15):3741‐3748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 297. Guo Y, Li H, Chen H, et al. Metagenomic next‐generation sequencing to identify pathogens and cancer in lung biopsy tissue. EBioMedicine. 2021;73:103639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 298. Katsman E, Orlanski S, Martignano F, et al. Detecting cell‐of‐origin and cancer‐specific methylation features of cell‐free DNA from Nanopore sequencing. Genome Biol. 2022;23(1):158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 299. Ozsolak F, Platt AR, Jones DR, et al. Direct RNA sequencing. Nature. 2009;461(7265):814‐818. [DOI] [PubMed] [Google Scholar]
- 300. Hrdlickova R, Toloue M, Tian B. RNA‐Seq methods for transcriptome analysis. Wiley Interdiscip Rev RNA. 2017;8(1). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 301. Zhao LY, Song J, Liu Y, Song CX, Yi C. Mapping the epigenetic modifications of DNA and RNA. Protein Cell. 2020;11(11):792‐808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 302. Meyer KD, Saletore Y, Zumbo P, Elemento O, Mason CE, Jaffrey SR. Comprehensive analysis of mRNA methylation reveals enrichment in 3' UTRs and near stop codons. Cell. 2012;149(7):1635‐46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 303. Dominissini D, Moshitch‐Moshkovitz S, Schwartz S, et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A‐seq. Nature. 2012;485(7397):201‐206. [DOI] [PubMed] [Google Scholar]
- 304. Kiyose H, Nakagawa H, Ono A, et al. Comprehensive analysis of full‐length transcripts reveals novel splicing abnormalities and oncogenic transcripts in liver cancer. PLos Genet. 2022;18(8):e1010342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 305. Prasad V, Fojo T, Brada M. Precision oncology: origins, optimism, and potential. Lancet Oncol. 2016;17(2):e81‐e86. [DOI] [PubMed] [Google Scholar]
- 306. Shaw KRM, Maitra A. The status and impact of clinical tumor genome sequencing. Annu Rev Genomics Hum Genet. 2019;20:413‐432. [DOI] [PubMed] [Google Scholar]
- 307. Minervini CF, Cumbo C, Redavid I, et al. Nanopore sequencing approach for immunoglobulin gene analysis in chronic lymphocytic leukemia. Sci Rep. 2021;11(1):17668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 308. Yu SCY, Deng J, Qiao R, et al. Comparison of single molecule, real‐time sequencing and nanopore sequencing for analysis of the size, end‐motif, and tissue‐of‐origin of long cell‐free DNA in plasma. Clin Chem. 2023;69(2):168‐179. [DOI] [PubMed] [Google Scholar]
- 309. Martignano F, Munagala U, Crucitta S, et al. Nanopore sequencing from liquid biopsy: analysis of copy number variations from cell‐free DNA of lung cancer patients. Mol Cancer. 2021;20(1):32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 310. Klein EA, Richards D, Cohn A, et al. Clinical validation of a targeted methylation‐based multi‐cancer early detection test using an independent validation set. Ann Oncol. 2021;32(9):1167‐1177. [DOI] [PubMed] [Google Scholar]
- 311. Bruzek AK, Ravi K, Muruganand A, et al. Electronic DNA analysis of CSF cell‐free tumor DNA to quantify multi‐gene molecular response in pediatric high‐grade glioma. Clin Cancer Res. 2020;26(23):6266‐6276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 312. Valle‐Inclan JE, Stangl C, de Jong AC, et al. Optimizing nanopore sequencing‐based detection of structural variants enables individualized circulating tumor DNA‐based disease monitoring in cancer patients. Genome Med. 2021;13(1):86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 313. Li FW, Harkess A. A guide to sequence your favorite plant genomes. Appl Plant Sci. 2018;6(3):e1030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 314. Li C, Lin F, An D, Wang W, Huang R. Genome sequencing and assembly by long reads in plants. Genes. 2017;9(1):6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 315. Michael TP, VanBuren R. Progress, challenges and the future of crop genomes. Curr Opin Plant Biol. 2015;24:71‐81. [DOI] [PubMed] [Google Scholar]
- 316. Choi JY, Lye ZN, Groen SC, et al. Nanopore sequencing‐based genome assembly and evolutionary genomics of circum‐basmati rice. Genome Biol. 2020;21(1):21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 317. Kawakatsu T, Huang SC, Jupe F, et al. Epigenomic diversity in a global collection of Arabidopsis thaliana accessions. Cell. 2016;166(2):492‐505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 318. Long Q, Rabanal FA, Meng D, et al. Massive genomic variation and strong selection in Arabidopsis thaliana lines from Sweden. Nat Genet. 2013;45(8):884‐890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 319. Wang B, Yang X, Jia Y, et al. High‐quality Arabidopsis thaliana genome assembly with nanopore and HiFi long reads. Genomics Proteomics Bioinformatics. 2022;20(1):4‐13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 320. Gabur I, Chawla HS, Snowdon RJ, Parkin IAP. Connecting genome structural variation with complex traits in crop plants. Theor Appl Genet. 2019;132(3):733‐750. [DOI] [PubMed] [Google Scholar]
- 321. Liu Y, Zhang M, Wang R, et al. Comparison of structural variants detected by PacBio‐CLR and ONT sequencing in pear. Bmc Genomics [Electronic Resource]. 2022;23(1):830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 322. Alonge M, Wang X, Benoit M, et al. Major impacts of widespread structural variation on gene expression and crop improvement in tomato. Cell. 2020;182(1):145‐161.e23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 323. Liang Z, Riaz A, Chachar S, Ding Y, Du H, Gu X. Epigenetic modifications of mRNA and DNA in plants. Mol Plant. 2020;13(1):14‐30. [DOI] [PubMed] [Google Scholar]
- 324. Parker MT, Knop K, Sherwood AV, et al. Nanopore direct RNA sequencing maps the complexity of Arabidopsis mRNA processing and m(6)A modification. eLife. 2020;9:e49658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 325. Wang Y, Wang H, Xi F, et al. Profiling of circular RNA N(6) ‐methyladenosine in moso bamboo (Phyllostachys edulis) using nanopore‐based direct RNA sequencing. J Integr Plant Biol. 2020;62(12):1823‐1838. [DOI] [PubMed] [Google Scholar]
- 326. Senol Cali D, Kim JS, Ghose S, Alkan C, Mutlu O. Nanopore sequencing technology and tools for genome assembly: computational analysis of the current state, bottlenecks and future directions. Briefings Bioinf. 2019;20(4):1542‐1559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 327. de Lannoy C, de Ridder D, Risse J. The long reads ahead: de novo genome assembly using the MinION. F1000Research. 2017;6:1083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 328. Loman NJ, Quick J, Simpson JT. A complete bacterial genome assembled de novo using only nanopore sequencing data. Nat Methods. 2015;12(8):733‐735. [DOI] [PubMed] [Google Scholar]
- 329. Wan YK, Hendra C, Pratanwanich PN, Göke J. Beyond sequencing: machine learning algorithms extract biology hidden in Nanopore signal data. Trends Genet: TIG. 2022;38(3):246‐257. [DOI] [PubMed] [Google Scholar]
- 330. Fathizadeh H, Afshar S, Masoudi MR, et al. SARS‐CoV‐2 (Covid‐19) vaccines structure, mechanisms and effectiveness: a review. Int J Biol Macromol. 2021;188:740‐750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 331. Pardi N, Hogan MJ, Porter FW, Weissman D. mRNA vaccines—a new era in vaccinology. Nat Rev Drug Discov. 2018;17(4):261‐279. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data sharing is not applicable to this article as no new data were created or analyzed in this study.