

Abstract
Bone regeneration therapy is clinically important, and targeted regulation of endoplasmic reticulum (ER) stress is important in regenerative medicine. The processing of proteins in the ER controls cell fate. The accumulation of misfolded and unfolded proteins occurs in pathological states, triggering ER stress. ER stress restores homeostasis through three main mechanisms, including protein kinase-R-like ER kinase (PERK), inositol-requiring enzyme 1ɑ (IRE1ɑ) and activating transcription factor 6 (ATF6), collectively known as the unfolded protein response (UPR). However, the UPR has both adaptive and apoptotic effects. Modulation of ER stress has therapeutic potential for numerous diseases. Repair of bone defects involves both angiogenesis and bone regeneration. Here, we review the effects of ER stress on osteogenesis and angiogenesis, with emphasis on ER stress under high glucose (HG) and inflammatory conditions, and the use of ER stress inducers or inhibitors to regulate osteogenesis and angiogenesis. In addition, we highlight the ability for exosomes to regulate ER stress. Recent advances in the regulation of ER stress mediated osteogenesis and angiogenesis suggest novel therapeutic options for bone defects.
Keywords: Endoplasmic reticulum stress, Unfolded protein response, Osteogenesis, Angiogenesis, High glucose, Inflammation, Exosome, Bone defects
Introduction
As the elderly population has increased globally, so has the number of patients with clinical bone defects [1, 2]. Patients with diabetes mellitus have a high incidence of bone defects [3]. Diabetes mellitus impairs bone regeneration and biomechanics in newly regenerated bone, which may be related to metabolic disorders and dysfunction of mitochondrial function and macrophage polarization induced by blood glucose fluctuations, leading to the production of reactive oxygen species (ROS), which creates an inflammatory microenvironment at the site of bone defect [4, 5]. At present, in the field of bone tissue engineering, increasing studies indicate that the functional polarization of macrophages can be adjusted by various modified hydrogels and 3D bioprinting of multicell-laden scaffolds, so as to promote the repair of diabetic bone defects [6, 7]. The latest research shows that stem cell therapy can also be a therapeutic target, bone marrow-derived macrophage (BMDM) -derived exosomal miRNA can affect bone marrow mesenchymal stem cell (BMSCs) differentiation, providing effective methods and potential therapeutic targets for the treatment of diabetic bone defects [8]. However, the role of BMD-derived exosomal miRNA in diabetes and their communication with BMSCs remains unknown. The specific mechanisms underlying impaired bone repair and regeneration in diabetic conditions remain to be investigated.
Bone regeneration requires both osteogenesis and angiogenesis [9]. The mechanisms of bone regeneration include membrane-internalized bone and cartilage-internalized bone [10]. Bone regeneration requires the participation of osteoblasts, osteoclasts, and chondrocytes. Endothelial cells (ECs) promote angiogenesis, thereby contributing to bone regeneration [11]– [13]. Angiogenesis depends on the coordination of pro- and anti-angiogenic factors [14]. Vascular endothelial growth factor (VEGF) and fibroblast growth factor (FGF) were the earliest identified pro-angiogenic factors [15]. They drive EC proliferation, migration, and differentiation to promote angiogenesis [16].
The cellular and molecular mechanisms of angiogenesis and osteogenesis in bone regeneration have been investigated. However, ER stress pathway has received little attention. ER stress is related to many human diseases [17]. Drugs targeting ER stress have been developed [18]. ER stress is a double-edged sword that determines whether cells survive or die [19]. Appropriate ER stress restores cellular homeostasis by activating adaptive cellular adaptive programs, whereas excessive ER stress induces cell death by triggering apoptosis [20].
ER stress has dual roles in the regulation of osteogenesis and angiogenesis. Here, we review the effect of ER stress on osteogenesis and angiogenesis, including the link between HG, inflammation and ER stress signaling pathways. As a subclass of extracellular vesicles, exosomes come from a wide range of sources, can be secreted by almost all kinds of cells, and exist in various body fluids [21]. ER stress can promote exosome formation and release [22–24]. We also reviewed that exosomes from different sources promote osteogenesis and angiogenesis. Therefore, it is possible that ER stress serves as a downstream signaling pathway for exosomes to regulate osteogenesis and angiogenesis.
Working principle of ER stress
Occurrence of ER stress
The ER is the site of protein synthesis and processing [25], the largest intracellular organelle [26]. Proteins tend to enter the ER in an unfolded form, where they begin to fold. However, folding of proteins in the ER is inefficient (< 20%) [27], so protein quality control requires a balance between protein folding and degradation [28]. ER quality control (ERQC) identifies and eliminates misfolded proteins to maintain cellular homeostasis [29]. However, suppression of ERQC by environmental and genetic factors leads to increased protein misfolding [30] and accumulation of misfolded or unfolded proteins in the ER, leading to ER stress [31, 32]. Therefore, ER stress is an important cellular defense mechanism and is vital for maintaining ER homeostasis.
ER stress signaling pathways
ER stress can be classified as the UPR, ER overload response, and sterol regulatory cascade [33]. UPR occurs when a signal of misfolded ER proteins is transmitted to the nucleus [34]. Ischemia [35, 36], HG [37], and other pathological states activate the ER stress signaling pathway. The UPR is a signal transduction pathway that transmits information about protein folding to the nucleus and cytoplasm to restore ER homeostasis [38] and relieve ER stress [39]. In 1977 glucose-regulated protein (GPR) was discovered [40]. GPR promotes the correct folding of proteins in the ER [41, 42], linking glucose induction to protein misfolding. In 1988, Kozutsumi et al. [42] proposed a signaling transduction pathway activated by ER stress. The mammalian UPR pathway was first identified in yeast [43, 44] and is coordinated by three ER transmembrane sensor proteins: protein kinase-R-like ER kinase (PERK), inositol-requiring enzyme 1ɑ (IRE1ɑ), and activating transcription factor 6 (ATF6). It dynamically regulates ER protein folding to maintain ER homeostasis (Fig. 1) [45, 46]. In a non-stressed state, the ER chaperone immunoglobulin heavy-chain binding protein (BiP)/G protein coupled receptor 78 (GPR78) binds to the ER domain to stabilize ATF6 disulfides [38, 47], and PERK and IRE1ɑ bind to BiP and are inactivated [30]. However, in ER stress, BiP dissociates and binds unfolded or misfolded proteins and perform protein folding [48], activating ER receptors [49]. The IRE1ɑ-X-box binding protein (XBP1), PERK-eukaryotic initiation factor 2ɑ (eIF2ɑ), and ATF6 signaling pathways induce the UPR and restore ER stability [50, 51]. IRE1ɑ is the most evolutionarily conserved factor in the UPR [30, 38]. Activated PERK phosphorylates eIF2ɑ, attenuating protein translation to relieve the ER load under stress, and promotes ATF4 translation [30, 45]. The PERK signaling pathway is associated with a series of immune metabolic diseases [52, 53], including tumors [54–56]. ATF6 disulfide is decreased by protein disulfide isomerase (PDI) activity [30], and full-length ATF6 (ATF6p90) monomer increases and is transferred to the Golgi apparatus, where it is cleaved by the site 1 protease (S1P) and site 2 protease (S2P) to release an N-terminal transcriptionally active 50 kDa fragment (ATF6p50) [38, 57]. ATF6p50 is transported to the nucleus to perform functions such as protein folding [38]. ATF6 also maintains the stability of viral proteins [57] and homeostasis in normally developing tissues and organs [58].
Fig. 1.

Major UPR pathways initiated in the ER [38]. RIDD: regulated IRE1ɑ-dependent decay; TRAF: tumor necrosis factor receptor associated factor; ERAD: ER-associated protein degradation; PP1: protein phosphatase 1; CreP: constitutive repressor of eIF2ɑ phosphorylation; DR5: death receptor 5; TXNIP: thioredoxin-interacting protein; IP3R: inositol-1,4,5-triphosphate receptor; BI-1: Bax inhibitor-1; GADD34: growth arrest and DNA damage inducible gene 34. Hetz C, Zhang K, Kaufman RJ. Mechanisms, regulation, and functions of the unfolded protein response. Nat Rev Mol Cell Biol. 2020;21(8):421–38.
Copyright© The Authors 2020. Published by Springer Ltd
ER stress pathways: a double-edged sword
After the occurrence of ER stress, misfolded or unfolded proteins that accumulate in the ER are eliminated through two primary degradation pathways: ER-associated degradation (ERAD) and autophagy [59]. ERAD is activated in response to ER stress, as it maintains ER homeostasis by eliminating misfolded proteins from the ER and preventing their accumulation [60]. The UPR controls cell fate [17, 19, 20, 61]. A prolonged UPR indicates non-recovery from ER stress, and adaptive output cannot compensate for the pressure in the ER, and the UPR induces apoptosis [20]. Sustained activation of ATF4 in combination with CCAAT-enhancer-binding protein homologous protein (CHOP) induces apoptosis [34]. Thus, the dual role of the PERK-eIF2ɑ axis is vital for coordinating translation and protein balance. There are three main mechanisms (Fig. 1): the IRE1ɑ/ASK1 (apoptosis signal regulating kinase 1)/JNK (c-jun kinase) pathway [62], caspase-12-dependent pathway [63], and growth arrest and DNA damage-inducible 153 (CHOP/GADD153) pathway [64, 65]. CHOP, a key apoptotic factor, upregulates ROS, triggers calcium (Ca2+) release, and promotes transcription, constituting a positive-feedback loop that triggers apoptosis [34]. It also downregulates the anti-apoptotic protein B-cell lymphoma 2 (Bcl-2) to induce apoptosis [66]. Despite advances in research on the mechanism of ER stress, the balance between pro-survival and pro-death UPR signals remains unclear, and the full extent of ER stress's role in different stages of disease is yet to be fully elucidated. Future research is necessary to answer these key questions.
ER stress pathways in orthopedics
Osteoblasts play a critical role in bone formation and reconstruction by synthesizing new collagen. Because osteoblasts secrete a significant amount of extracellular matrix proteins, they are particularly vulnerable to ER stress-induced dysfunction. Targeted ER stress therapy can be used to treat orthopedic diseases. Liu et al. [67] discovered that IL-1β can induce excessive ER stress in chondrocytes, leading to chondrocyte apoptosis and subsequent cartilage degradation, which accelerates the progression of osteoarthritis (OA). Inhibition of ER stress by the IRE1ɑ pathway suppresses chondrocyte apoptosis, thus mitigating the progression of OA [68]. Sim et al. [69] found that the function of ERAD, which is regulated by ER stress, was reduced in patients with OA, leading to the accumulation of misfolded proteins and cartilage loss. Enhanced ERAD activity is necessary for cartilage formation and maintenance. The activation of PERK and ATF4 is involved in the inducing the stress response protein sestrin2 under ER stress after spinal cord injury (SCI) [70]. Inhibiting ER stress through overexpression of sestrin2 promotes functional recovery and neuronal survival, indicating its potential as a therapeutic target for SCI repair. Huang et al. [71] found that inhibition of neuronal apoptosis mediated by ER stress can reduce apoptosis and protects neurons. ER stress has potential to be a new target for treating SCI; Metastasis of osteosarcoma cells can be inhibited by knocking out secretion-associated Ras-related GTPase 1A (SAR1A), a key regulator of ER homeostasis [72]. Moreover, ER stress can induce hypertrophic chondrocyte dysfunction, which may be a potential cause of osteogenesis imperfecta (OI) [73]. Nevertheless, studies have demonstrated that downstream ER stress is necessary to maintain Ol bone integrity to a certain extent. Reducing ER stress alone may not be sufficient to rescue Ol phenotype and may even exacerbate it [74]. Although we have known that ER stress is part of the mechanism of OI disease, how to improve OI bone strength by regulating ER stress remains to be studied.
The repair and regeneration of bone defects caused by trauma, tumor, infection and other factors have been significant clinical challenges. If ER stress can be precisely regulated to an appropriate level through bone tissue engineering or stem cell therapy, it could help in the rapid regeneration of bone tissue. Xiang et al. [75] modulated the expression of osteogenic proteins through the PERK-eIF2ɑ-ATF4 pathway of appropriate ER stress by Ca2+ changes mediated by biphasic calcium phosphate, a classic bone void filler. Zheng et al. [76] used the osteogenesis-promoting drug HA15 to target HSPA5 to inhibit excessive ER stress and ultimately promote osteogenesis and angiogenesis in rabbit bone defect models. Future studies can use the involvement of the ER stress pathway in the regulation of osteogenesis and angiogenesis as a starting point through cell experiments, investigate the precise mechanism by which ER stress regulates osteogenesis and angiogenesis, and seek more possible therapeutic targets and interventions in the pathogenesis of bone defect from the level of gene regulation, bringing good news to the majority of patients.
Effects of ER stress pathways on osteogenesis
Appropriate ER stress contributes to osteogenic differentiation
Bone morphogenetic proteins (BMPs) are implicated in osteogenic differentiation and ectopic bone formation [77]. BMP2 and BMP9 induce ER stress to promote the differentiation of BMSCs into osteoblasts [78–81]. UPR signaling is an essential regulator of bone development [82, 83].
The three UPR signaling pathways are linked to the promotion of osteogenic differentiation by ER stress. Kazuhisa et al. [84] discovered Osterix (Osx), a transcription factor necessary for bone formation. Ten years later, Stavroula [85] identified Osx as a target gene of XBP1, linking ER stress and osteogenesis. The IRE1ɑ-XBP1 signaling pathway promotes not only osteoblast maturation by promoting Osx transcription [86] but also bone regeneration via myostatin mRNA decay [87]. ATF4 is a key transcription factor for osteoblast differentiation and bone formation [88, 89]. Activation of the PERK-eIF2ɑ-ATF4 pathway promotes the expression of genes required for osteogenesis [90] and induces osteogenic differentiation [91, 92] and type I collagen secretion, which are essential for neonatal bone development and osteogenic differentiation [93]. Won-Gu et al. [94] showed that BMP2 stimulates osteoblast differentiation by regulating osteocalcin gene expression via the ER stress-activated ATF6 pathway [58]. Although the three UPR signaling pathways are implicated in osteogenesis, the underlying mechanisms are unclear.
Excessive ER stress induces osteoblast apoptosis
Excessive ER stress inhibits osteogenic differentiation and induces their apoptosis [91, 95, 96], which is an important mechanism of osteoporosis [97]. The effect may be related to the overexpression of CHOP caused by excessive ER stress [98], and there are sex differences in sensitivity to CHOP [99]. Overexpression of CHOP reduces alkaline phosphatase activity and calcified bone nodule formation [100], and initiates osteoblast apoptosis, inhibits bone formation, and induces osteopenia [98, 100].
ER stress-mediated osteoblast apoptosis is driven by an increase in the intracellular Ca2+ concentration [101]. An increased intracellular Ca2+ disrupts Ca2+ homeostasis, leading to Ca2+ overload [102] and excessive ER stress103 and inducing osteoblast apoptosis [104, 105]. Furthermore, micronutrients such as cadmium [106], fluorine [107, 108], and iron [109] initiate the ER stress apoptosis pathway by increasing intracellular Ca2+. Therefore, controlling intracellular Ca2+ has therapeutic potential for micronutrient-induced osteoporosis. We summarize the effects of ER stress inducers on osteogenic differentiation in Table 1.
Table 1.
ER stress inducers used to modulate osteogenic differentiation
| ER stress inducer | Pathway | Stress degree | Up/down | Mechanism | Refs. |
|---|---|---|---|---|---|
| TNF-ɑ | JNK | Excessive | Down | Inhibit osteogenic differentiation of BMSCs | [128] |
| Curcumin | ATF6 | Appropriate | Up | Promote osteogenic differentiation of C3H10T1/2 cells | [135] |
| METTL3 | – | Excessive | Down | induce osteoblast apoptosis | [136] |
| CDs | PERK-eIF2ɑ-ATF4 | Appropriate | Up | promote pre-osteoblast differentiation in vitro and bone regeneration in vivo | [137] |
| PIs | IRE1ɑ-XBP1 | Approriate | Up | promote osteogenic differentiation | [138] |
| Melatonin | PERK-eIF2ɑ-ATF4 | Excessive | Down | induce apoptosis in hFOB 1.19 human osteoblastic cells | [139] |
| AGEs | IRE1ɑ | Excessive | Down | induce apoptosis in osteoblastic MC3T3-E1 and human osteoblastic hFOB 1.9 cells | [140] |
| TNF-ɑ | PERK | Excessive | Down | inhibit osteogenic differentiation of PDLSCs | [141] |
| HA15 | PERK-eIF2ɑ-ATF4 | Appropriate | Up | promote osteogenic differentiation in vitro, and attenuate estrogen deficiency-induced bone loss in vivo | [142] |
| MNT | PERK-eIF2ɑ-ATF4 | Appropriate | Up | Promote osteogenic differentiation of stem cells | [143] |
| PA | CHOP/Caspase-12 /JNK | Excessive | Down | induce apoptosis in osteoblastic MC3T3-E1 cells | [144] |
| FTO | A positive feedback loop with pAMPK | Appropriate | Up | promote osteogenic differentiation of C3H10T1/2 cells | [145] |
| Metallic wear debris | – | Excessive | Down | induce osteoblast apoptosis | [146] |
TNF-ɑ: tumor necrosis factor-ɑ; METTL3: methyltransferase-like 3; CDs: carbon dots; PI: proteasome inhibitor; AGE: advanced glycation end product; PDLSC: periodontal ligament stem cell; MNT: micro-/nano-topography; PA: palmitate; FTO: fat mass and obesity associated
Regulation of ER stress pathways to interfere with osteogenesis
GCs induce osteoblast apoptosis by activating ER stress pathways
A normal concentration of glucose does not activate ER stress [110], but chronic HG induces pancreatic β cells to continuously secrete Ca2+ to activate ER stress [37], thus inhibiting osteogenic differentiation in a glucose concentration-dependent manner [110].
Since 1984, glucocorticoids (GCs) have been used for variety of immune-related diseases [111]. However, long-term use of GCs increases the incidence of osteonecrosis, among which osteonecrosis of the femoral head (ONFH) is the most common [112]. Although the mechanism of GC-induced ONFH is unclear, GCs can activate ER stress and promote the production of ROS, thereby inducing apoptosis in osteoblasts [113, 114], particularly in the proximal femur [115]. This may be a mechanism of ONFH.
The PERK-eIF2ɑ-ATF4-CHOP pathway is implicated in GC-induced osteoblast apoptosis [116]. Therefore, controlling this pathway could ameliorate GC-induced osteoblast apoptosis. The plant compound geniposide (GEN) [117], 4-phenylbutyric acid (4-PBA) [117, 118], the PERK phosphorylation inhibitor GSK2656157 [116], and melatonin [119] can block PERK downstream signaling and significantly inhibit ER stress, thereby attenuating GC-induced osteoblast apoptosis. GEN together with the plant compound paeoniflorin activate autophagy in vivo and in vitro, thus suppressing GC-induced apoptosis [115, 120]. 4-PBA downregulates ATF4 and reduces mutant type I collagen [121], whereas salubrinal (inhibitor of eIF2ɑ dephosphorylation) upregulates ATF4 [35, 122]. Both regulate the eIF2ɑ pathway, thereby reducing ER stress to promote osteogenesis. Unfortunately, salubrinal has no effect on osteoblast apoptosis induced by high-dose GC [114].
Regulation of ER stress pathways on osteogenesis under inflammatory conditions
Long-term inflammatory responses can affect stem cells' ability to repair [123]. Tumor necrosis factor ɑ (TNF-ɑ)-induced inflammation has been reported to inhibit osteogenic differentiation of BMSCs [124], possibly because ER stress-activated nuclear factor κB (NF-κB) translocates into the nucleus to promote the transcription of other pro-inflammatory cytokines [125] and osteolysis [126]. Xue et al. [96] found for the first time that long-term chronic inflammation reduces the expression of lysine acetyltransferase 6B (KAT6B, also known as MORF), which leads to continuous activation of PERK signaling pathway downstream of ER stress, and reduces the osteogenic differentiation ability of periodontal ligament stem cells (PDLSCs).
Subsequently, Li et al. [127] used low-intensity pulse ultrasound to up-regulate the osteogenic effect of PDLSCs under inflammatory conditions through UPR. Zhao et al. [128] demonstrated that JNK pathway activated by ER stress mediates TNF-ɑ-induced inflammation in BMSCs. These studies have confirmed that inhibiting ER stress can effectively reduce inflammatory response and enhance the osteogenic differentiation ability of stem cells, which may provide new insights for improving stem cell osteogenic differentiation and treating inflammatory bone diseases such as osteoporosis, so that inhibiting ER stress under inflammatory conditions to promote osteogenesis has great potential.
Exosomes regulate osteogenesis by activating ER stress pathways
Studies have shown that miRNA from exosomes of different cellular origins can enter recipient cells and then regulate the expression of genes associated with osteogenesis at the translational level to regulate osteogenesis [129]. We have also reviewed the use of exosome-derived non-coding RNAs for osteogenesis before [130]. However, whether exosomes promote osteogenesis by regulating ER stress is unclear. Platelet-rich plasma (PRP) has been widely used in clinical repair of bone and soft tissue injuries. Recent studies have shown that PRP contains a large number of extracellular vesicles [131]. Tao et al. [132] found that PRP-derived exosomes (PRP-Exos) binds to related receptors, promotes Akt phosphorylation, activates β-catenin to promote osteogenesis, and activates Bcl-2 to inhibit GC-induced apoptosis and ER stress (Fig. 2). Exosomes show great potential in PRP repair tissues, which is closely related to downstream ER stress pathways. Wang et al. [133] reported that miR-485–5p modified exosomes inhibit ER stress and alleviate chondrocyte apoptosis for the treatment of OA. Liao et al. [134] demonstrated that BMSCs-derived exosomes (BMSCs-Exos) can improve the apoptosis of nucleus pulposus cells induced by ER stress. Can BMSCs-Exos attenuate osteoblast apoptosis by inhibiting excessive ER stress? This may be a new mechanism of exosome promoting osteogenesis, which needs to be verified by future experiments.
Fig. 2.
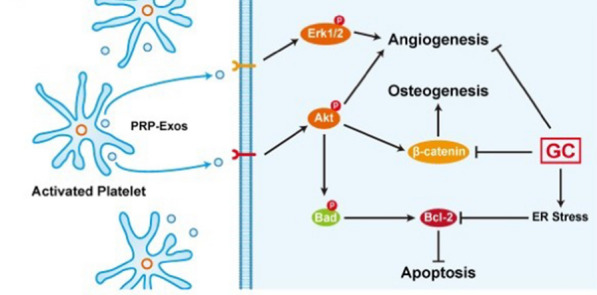
PRP-Exos rescued cells from GC-induced apoptosis via the Akt/Bcl-2 pathway [132]. Tao SC, Yuan T, Rui BY, Zhu ZZ, Guo SC, Zhang CQ. Exosomes derived from human platelet-rich plasma prevent apoptosis induced by glucocorticoid-associated endoplasmic reticulum stress in rat osteonecrosis of the femoral head via the Akt/Bad/Bcl-2 signal pathway. Theranostics. 2017;7(3):733–50.
Copyright© The Authors 2017. Published by Ivyspring International Publisher
Effects of ER stress pathways on angiogenesis
Appropriate ER stress contributes to angiogenesis
ER stress promotes the differentiation of monocytes into ECs, leading to angiogenesis [147, 148], suggesting that ER stress can promote angiogenesis. The angiogenic effect of ER stress is mediated by regulation of angiogenic factors by the UPR. Appropriate ER stress triggers the production of angiogenic factors [149, 150]; however, the mechanism is unclear.
Three UPR signaling pathways bind to regulatory regions of VEGFA, and jointly drive VEGFA transcription [151, 152]. ER stress initiates angiogenesis signaling via UPR-mediated upregulation of VEGFA [153, 154]. The inducible ER chaperone oxygen-regulated protein 150 (OPR150) promotes the expression of VEGFA in pathological conditions and is a potential target for regulating angiogenesis [155]. Under ER stress, the IRE1ɑ-XBP1 pathway promotes tumor angiogenesis [156]– [158], the PERK-ATF4 pathway promotes bone angiogenesis [153, 159], and the ATF6 pathway promotes embryonic angiogenesis [160], by upregulating VEGFA. Binet et al. [161] reported a pro-angiogenic role for the UPR in diseases characterized by pathological vascular abnormalities. Therefore, targeted regulation of angiogenesis through the UPR has therapeutic potential for vascular necrotizing diseases. VEGFA can spontaneously increase in acute myocardial ischemia, inducing intracellular Ca2+ overload and activating ER stress in a positive-feedback loop [162]. Excessive ER stress induces BMSCs apoptosis [128], and VEGFA stimulates the differentiation of BMSCs into ECs, thus protecting BMSCs and promoting angiogenesis [163]. Increased spontaneous VEGFA production also promotes compensatory angiogenesis through the ROS-ER stress-autophagy axis [162].
The UPR also regulates vascular growth factors. For example, ER stress promotes angiogenesis by upregulating interleukin 8 (IL-8) [164], FGF2 [36], placental growth factor (PIGF) [165], and granulocyte–macrophage colony stimulating factor (GM-CSF) [166] via different transcriptional mechanisms. However, pentraxin 3 (PTX3) has a high affinity for FGF2 and can inhibit its angiogenesis [167, 168], but Ma et al. [169] found that the ATF4 pathway activates SMAD-specific E3 ubiquitin ligase 2 and leads to PTX3 degradation, thus promoting angiogenesis. Philippe et al. [36] demonstrated that the PERK pathway activates the translation of dependent internal ribosome entry site (IRES), thereby promoting the expression of the angiogenic factors VEGFA and FGF2. These studies have suggested potential therapeutic targets for ischemia in vascular necrotizing diseases.
Excessive ER stress impairs angiogenesis
Excessive ER stress impairs angiogenesis not only by reducing the transcription of pro-angiogenetic growth factors such as VEGFA [170– 172] and PIGF [173] but also by activating negative angiogenic regulators such as delta-like 4 (DLL4) IRES [174]. Excessive ER stress can induce apoptosis of ECs [175–178], thus suppressing their angiogenesis [171]. Maamoun et al. [179] showed that ER stress causes EC dysfunction, suggesting that targeting ER stress could promote angiogenesis (Table 2). The ER stress-mediated decreased expression of angiogenic genes is related to age [180].
Table 2.
ER stress inhibitors used to promote angiogenesis
| ER stress inhibitor | Pathway | Animal model | Mechanism | Refs. |
|---|---|---|---|---|
| GB | CHOP, GPR78, caspase-12 | Rat | Promote perforator flap angiogenesis | [210] |
| Quercetin | ATF6/GPR78 | N/A | Protect HBMECs and promote angiogenesis | [211] |
| Vitamin D | GPR78, JNK1, eIF2ɑ, XBP-1 | N/A | Protect HUVECs and promote angiogenesis | [212] |
| PTP1B inhibition | PI3K/Akt | N/A | Protect HUVECs, activate eNOS and promote angiogenesis | [171] |
| SFN | ATF6/GPR78 | Chick | Promote embryo angiogenesis | [213] |
| Naringin | GPR78, CHOP, caspase-12, Cyt.c | Rat | Protect VECs and promote angiogenesis | [214] |
| Salubrinal | eIF2ɑ-ATF4-GPR78 | Rat | Promote HUVESs, upregulate VEGFA and promote angiogenesis | [35] |
| HO-1 | BiP, PERK-eIF2ɑ-ATF4 | N/A | Alleviate HG-induced HUVECs apoptosis and promote angiogenesis | [172] |
| PCB2 | PERK, IRE1ɑ and ATF6 | Mouse | Alleviate HG-induced ECs dysfunction and promote angiogenesis | [192] |
| streptozotocin | CHOP | Mouse | Alleviate HG-induced APCs dysfunction and promote vascular repair | [193] |
| GSK2656157 | PERK | Mouse | Alleviate GCs-induced ECs apoptosis and promote angiogenesis | [116] |
GB: Ginkgolide B; N/A: no animal; HBMECs: human brain microvascular endothelial cells; HUVECs: human umbilical endothelial cells; PTP1B: protein tyrosine phosphatase 1B; eNOS: endothelial nitric oxide synthase; SFN: sulforaphane; Cyt.c: cytochrome c; VECs: vascular endothelial cells; HO-1: Hemeoxygenase-1; PCB2: procyanidin B2; APCs: angiogenic progenitor cells
The anti-angiogenic effect of ER stress also has benefits, such as inhibiting cancer progression [181]. ER stress can induce the expression of miR-153, which inhibits angiogenesis by two mechanisms, suggesting a novel therapeutic strategy for breast cancer [182].
Regulation of ER stress pathways to interfere with angiogenesis
HG impairs angiogenesis by activating ER stress pathways
In diabetic retinopathy (DR), HG damages normal blood vessels and causes abnormal neovascularization [183, 184]. ER stress is closely related to retinal angiogenesis [185]. Wang et al. [186] showed that regulation of ER stress can inhibit abnormal neovascularization. However, whether damaged normal blood vessels can be restored by regulating ER stress is unknown.
HG rapidly activates ER stress in ECs [187] and angiogenic progenitor cells (APCs) [188], leading to microvascular EC dysfunction and impair angiogenesis [189]. ECs have a greater apoptotic effect under GC induction than do other cells [190, 191]. Gao et al. [116] demonstrated that GCs induce EC apoptosis by activating ER stress, leading to microvascular damage. Alleviating the ER stress induced by HG can counteract HG-induced EC apoptosis [172], thus restoring angiogenesis [192] and enhancing vascular repair by circulating angiogenic cells (CACs) [193] (Table 2). Inhibition of ER stress can prevent vascular damage by upregulating pro-angiogenic factors and downregulating anti-angiogenic factors [194]. Wang et al. [195] found that an atypical UPR pathway mediated by IRE1ɑ regulates miRs, thereby protecting the pro-angiogenic growth factor angiopoietin 1 (ANGPT1) from miR attack under HG conditions and promoting bone marrow–derived progenitor cell (BMPC) angiogenesis. Therefore, targeting ER stress is the key to reversing HG-induced vascular injury.
Regulation of ER stress pathways on angiogenesis under inflammatory conditions
In recent years, ER stress pathways secondary to inflammation have become new targets for intracellular therapy. ER stress can induce nucleotide-binding domain and leucine-rich repeat containing (NLRP3) inflammasome through PERK and IRElα pathways, regulate the release of inflammatory cytokines, and trigger inflammatory response [196]. Wang et al. [197] demonstrated that there is a positive feedback loop between interleukin-17A (IL-17A) and ER stress, and that inhibition of ER stress or IL-17A can reduce the neovascularization area of DR. At present, inhibition of ER stress can alleviate inflammation and inhibit angiogenesis, which has been proved in both cell and animal experiments [186, 198]. Although ER stress pathway shows great potential in anti-inflammatory and anti-vascular therapy, more in-depth mechanism studies are needed before clinical trials.
Exosomes regulate angiogenesis by activating ER stress pathways
Exosomes promote angiogenesis by inducing the regeneration of damaged blood vessels by inhibiting EC apoptosis and promoting their angiogenic activity [199–202]. Tumor cell-derived exosomes deliver miR-25-3p to ECs, thereby disrupting ECs integrity, increasing vascular permeability, and promoting angiogenesis, thereby promoting tumor metastasis [203]. Based on the role of ER stress in numerous pathological conditions, whether exosomes promote angiogenesis by regulating ER stress is a topic of interest. Tao et al. [132] have found that PRP-Exos activates the Akt pathway under ER stress, releasing multiple growth factors and promoting angiogenesis (Fig. 2).
Exosomes have a dual regulatory effect on angiogenesis. Angiogenesis can be inhibited by exosomes. For example, exosomal circular RNAs (circRNAs) act as signal carriers to trigger EC dysfunction [204], exosomes can enhance the inhibitory effect of the anti-angiogenic peptide KV11 on pathological retinal angiogenesis [205], and circulating exosomal miR-20b-5p is transferred to vascular ECs to inhibit the regeneration of diabetic damaged blood vessels [206]. Wang et al. showed that ER-stressed HN4 cell-derived exosomes modified by miR-424–5p inhibit angiogenesis by HUVECs [207].
Exosomes from different sources have different regulatory effects on angiogenesis under ER stress. Until now, studies on exosomes promoting angiogenesis by activating ER stress have focused on exosomes of tumor cell origin. Lin et al. [208] demonstrated that after knocking down PERK in HUVEC, HeLa cell-derived exosomes can significantly improve HUVEC proliferation. We know that BMSCs-Exos have great potential in promoting angiogenesis [209], but whether ER stress may be a downstream pathway and whether we can enhance the ability of BMSCs-Exos to promote angiogenesis by regulating ER stress needs to be demonstrated in future studies.
Potential interventions related to ER stress pathways
Because the ER controls protein synthesis and degradation, ER stress is used clinically to restore myogenic differentiation to treat uremic sarcopenia [215]. Moreover, clinical trials by Bella et al. [216] suggested that ER stress may play a key role in the pathogenesis of amyotrophic lateral sclerosis by altering the regulation of protein balance, and that molecules acting on functional control of the UPR pathway may be beneficial in slowing disease progression, but subgroup analyses were not performed in this study. Therefore, this effect on targeting ER stress is considered exploratory. Besides, drugs targeting the IRE1ɑ-XBP1 pathway can inhibit vascular smooth muscle apoptosis, thereby alleviating aortic dissection [217]. Dexmedetomidine pretreatment can effectively protect myocardial ischemia–reperfusion-induced acute kidney injury by inhibiting ER stress [218].
Regulation of ER-related signaling pathways is most commonly used in the treatment of tumor diseases. ER stress is an essential intermediate targeting pathway in tumor therapy. Activation of ER stress can increase the cytotoxicity of photodynamic therapy to tumor cells [219]. Chemotherapy can increase tumor (sarcoma and gastric cancer) sensitivity by activating ER stress [220, 221]. Use of some chemotherapy drugs is limited by their toxicity. However, drugs that inhibit ER stress have reduced toxicity, and can be used in chemotherapy for cancer [222, 223]. Basic research by Varone et al. [224] showed that ISRIB (a small molecule that inhibits the action of phosphorylated eIF2ɑ) increases ER protein load, reactivates protein synthesis in damaged protein homeostasis, and ultimately promotes tumor cytotoxicity. ISRIB offers a new treatment option that can effectively inhibit tumor progression in conditions with impaired protein balance.
Although the mechanism of ER stress has been relatively clear, the current research on the intervention effect of ER stress in many diseases such as different types of diabetes and its complications is far from enough. Regulating a key signaling pathway node in the complex process of ER stress to affect the occurrence and development of diseases is an important target for drug therapy exploration, which has important clinical guiding value and practical significance. Further large-scale and long-term studies are needed to confirm the clinical benefits of this new pharmacological protocol, which may provide a promising therapeutic approach for targeted therapies for a number of diseases in the clinic.
Conclusion and perspective
In regenerative medicine, bone defects can be improved by promoting angiogenesis and osteogenesis. Our research has focused on inducing the regeneration of dead blood vessels and bone. ER stress is involved in many diseases. ER stress is a double-edged sword; its activation can promote cell generation, but excessive activation can induce apoptosis. ER stress plays a dual role in osteogenesis and angiogenesis, and thereby determines cell fate. Here we systematically reviewed the effect of ER stress on osteogenesis and angiogenesis. ER stress can be activated in pathological conditions such as HG and inflammation, or by inducers, and is inactivated by inhibitors. Therefore, regulation of ER stress has potential as a therapeutic target to promote osteogenesis and angiogenesis. Although regulating ER stress stimulates osteogenesis and angiogenesis, the mechanism is unclear. Efforts should focus on unraveling the mechanisms underlying the roles of ER stress in osteogenesis and angiogenesis.
Acellular therapy, such as exosome-mediated regulation of ER stress, is a focus of research. BMSCs-Exos have great potential for osteogenesis and angiogenesis, and we propose to hypothesize that ER stress can act as a downstream pathway for their regulation. Our future studies will further clarify the mechanism by which BMSCs-Exos promote angiogenesis and bone regeneration by regulating ER stress. Further research on the mechanism of ER stress regulating osteogenesis and angiogenesis will be helpful for the repair of bone defects.
Acknowledgements
Not applicable.
Abbreviations
- ER
Endoplasmic reticulum
- UPR
Unfolded protein response
- HG
High glucose
- ECs
Endothelial cells
- VEGF
Vascular endothelial growth factor
- FGF
Fibroblast growth factor
- ERQC
ER quality control
- GPR
Glucose-regulated protein
- PERK
Protein kinase-R-like ER kinase
- IRE1ɑ
Inositol-requiring enzyme 1ɑ
- ATF6
Activating transcription factor 6
- BiP
Immunoglobulin heavy-chain binding protein
- GPR78
G protein coupled receptor 78
- XBP1
X-box binding protein
- eIF2ɑ
Eukaryotic initiation factor 2ɑ
- PDI
Protein disulfide isomerase
- ATF6p90
Full-length ATF6
- S1P
Site 1 protease
- S2P
Site 2 protease
- ATF6p50
50 KDa fragment
- CHOP
CCAAT-enhancer-binding protein homologus protein
- ASK1
Apoptosis signal regulating kinase 1
- JNK
C-jun kinase
- GADD153
Growth arrest and DNA damage-inducible 153
- ROS
Reactive oxygen species
- Ca2 +
Calcium
- Bcl-2
B-cell lymphoma 2
- RIDD
Regulated IRE1ɑ-dependent decay
- TRAF
Tumor necrosis factor receptor associated factor
- ERAD
ER-associated protein degradation
- PP1
Protein phosphatase 1
- CreP
Constitutive repressor of eIF2ɑ phosphorylation
- DR5
Death receptor 5
- TXNIP
Thioredoxin-interacting protein
- IP3R
Inositol-1,4,5-triphosphate receptor
- BI-1
Bax inhibitor-1
- GADD34
Growth arrest and DNA damage inducible gene 34
- OA
Osteoarthritis
- IDD
Intervertebral disc degeneration
- SCI
Spinal cord injury
- SAR1A
Secretion-associated Ras-related GTPase 1A
- OI
Osteogenesis imperfecta
- BMPs
Bone morphogenetic proteins
- BMSCs
Bone marrow mesenchymal stem cells
- Osx
Osterix
- GCs
Glucocorticoids
- ONFH
Osteonecrosis of the femoral head
- GEN
Geniposide
- 4-PBA
4-Phenylbutyric acid
- PRP-Exos
Exosomes derived from platelet-rich plasma
- TNF-ɑ
Tumor necrosis factor-ɑ
- METTL3
Methyltransferase-like 3
- CDs
Carbon dots
- PI
Proteasome inhibitor
- AGE
Advanced glycation end product
- PDLSC
Periodontal ligament stem cell
- MNT
Micro-/nano-topography
- PA
Palmitate
- FTO
Fat mass and obesity associated
- OPR150
Oxygen-regulated protein 150
- IL-8
Interleukin 8
Author contributions
TYW and TL developed the concept, discussed the ideas, and wrote the manuscript. All authors made substantial, direct and intellectual contribution to the review. All authors read and approved the final manuscript.
Funding
This work was supported by grants from the National Natural Science Foundation of China (grant numbers 82272489, 82203588); Mount Taishan Scholar Project Special Fund; Qingdao Traditional Chinese Medicine Science and Technology Project (2021-zyym28); and the Science and Technology Development Project of Shandong Geriatric Society (LKJGG2021W082). The funding body played no role in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
Availability of data and materials
Not applicable.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors have declared that no competing interest exists.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Bharadwaz A, Jayasuriya AC. Osteogenic differentiation cues of the bone morphogenetic protein-9 (BMP-9) and its recent advances in bone tissue regeneration. Mater Sci Eng C Mater Biol Appl. 2021;120:111748. doi: 10.1016/j.msec.2020.111748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Yuan B, Wang L, Zhao R, Yang X, Yang X, Zhu X, Liu L, Zhang K, Song Y, Zhang X. A biomimetically hierarchical polyetherketoneketone scaffold for osteoporotic bone repair. Sci Adv. 2020;6:eabc4704. doi: 10.1126/sciadv.abc4704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Jiao H, Xiao E, Graves DT. Diabetes and its effect on bone and fracture healing. Curr Osteoporos Rep. 2015;13:327–335. doi: 10.1007/s11914-015-0286-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ceriello A, Esposito K, Piconi L, Ihnat MA, Thorpe JE, Testa R, Boemi M, Giugliano D. Oscillating glucose is more deleterious to endothelial function and oxidative stress than mean glucose in normal and type 2 diabetic patients. Diabetes. 2008;57:1349–1354. doi: 10.2337/db08-0063. [DOI] [PubMed] [Google Scholar]
- 5.Cai F, Liu Y, Liu K, Zhao R, Chen W, Yusufu A, Liu Y. Diabetes mellitus impairs bone regeneration and biomechanics. J Orthop Surg Res. 2023;18:169. doi: 10.1186/s13018-023-03644-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sun X, Ma Z, Zhao X, Jin W, Zhang C, Ma J, Qiang L, Wang W, Deng Q, Yang H, Zhao J, Liang Q, Zhou X, Li T, Wang J. Three-dimensional bioprinting of multicell-laden scaffolds containing bone morphogenic protein-4 for promoting M2 macrophage polarization and accelerating bone defect repair in diabetes mellitus. Bioact Mater. 2021;6:757–769. doi: 10.1016/j.bioactmat.2020.08.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wu Z, Bai J, Ge G, Wang T, Feng S, Ma Q, Liang X, Li W, Zhang W, Xu Y, Guo K, Cui W, Zha G, Geng D. Regulating macrophage polarization in high glucose microenvironment using lithium-modified bioglass-hydrogel for diabetic bone regeneration. Adv Healthc Mater. 2022;11:e2200298. doi: 10.1002/adhm.202200298. [DOI] [PubMed] [Google Scholar]
- 8.Zhang D, Wu Y, Li Z, Chen H, Huang S, Jian C, Yu A. MiR-144-5p, an exosomal miRNA from bone marrow-derived macrophage in type 2 diabetes, impairs bone fracture healing via targeting Smad1. J Nanobiotechnology. 2021;19:226. doi: 10.1186/s12951-021-00964-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Rather HA, Jhala D, Vasita R. Dual functional approaches for osteogenesis coupled angiogenesis in bone tissue engineering. Mater Sci Eng C Mater Biol Appl. 2019;103:109761. doi: 10.1016/j.msec.2019.109761. [DOI] [PubMed] [Google Scholar]
- 10.Schott NG, Friend NE, Stegemann JP. Coupling osteogenesis and vasculogenesis in engineered orthopedic tissues. Tissue Eng Part B Rev. 2021;27:199–214. doi: 10.1089/ten.teb.2020.0132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Grellier M, Bordenave L, Amedee J. Cell-to-cell communication between osteogenic and endothelial lineages: implications for tissue engineering. Trends Biotechnol. 2009;27:562–571. doi: 10.1016/j.tibtech.2009.07.001. [DOI] [PubMed] [Google Scholar]
- 12.Qin Y, Sun R, Wu C, Wang L, Zhang C. Exosome: a novel approach to stimulate bone regeneration through regulation of osteogenesis and angiogenesis. Int J Mol Sci. 2016;17:712. doi: 10.3390/ijms17050712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gerber HP, Vu TH, Ryan AM, Kowalski J, Werb Z, Ferrara N. VEGF couples hypertrophic cartilage remodeling, ossification and angiogenesis during endochondral bone formation. Nat Med. 1999;5:623–628. doi: 10.1038/9467. [DOI] [PubMed] [Google Scholar]
- 14.Huang Z, Bao SD. Roles of main pro- and anti-angiogenic factors in tumor angiogenesis. World J Gastroenterol. 2004;10:463–470. doi: 10.3748/wjg.v10.i4.463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Shing Y, Folkman J, Sullivan R, Butterfield C, Murray J, Klagsbrun M. Heparin affinity: purification of a tumor-derived capillary endothelial cell growth factor. Science. 1984;223:1296–1299. doi: 10.1126/science.6199844. [DOI] [PubMed] [Google Scholar]
- 16.Lamalice L, Le Boeuf F, Huot J. Endothelial cell migration during angiogenesis. Circ Res. 2007;100:782–794. doi: 10.1161/01.RES.0000259593.07661.1e. [DOI] [PubMed] [Google Scholar]
- 17.Oakes SA, Papa FR. The role of endoplasmic reticulum stress in human pathology. Annu Rev Pathol. 2015;10:173–194. doi: 10.1146/annurev-pathol-012513-104649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Marciniak SJ, Chambers JE, Ron D. Pharmacological targeting of endoplasmic reticulum stress in disease. Nat Rev Drug Discov. 2022;21:115–140. doi: 10.1038/s41573-021-00320-3. [DOI] [PubMed] [Google Scholar]
- 19.Lin JH, Li H, Yasumura D, Cohen HR, Zhang C, Panning B, Shokat KM, Lavail MM, Walter P. IRE1 signaling affects cell fate during the unfolded protein response. Science. 2007;318:944–949. doi: 10.1126/science.1146361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Shore GC, Papa FR, Oakes SA. Signaling cell death from the endoplasmic reticulum stress response. Curr Opin Cell Biol. 2011;23:143–149. doi: 10.1016/j.ceb.2010.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.van der Pol E, Böing AN, Harrison P, Sturk A, Nieuwland R. Classification, functions, and clinical relevance of extracellular vesicles. Pharmacol Rev. 2012;64:676–705. doi: 10.1124/pr.112.005983. [DOI] [PubMed] [Google Scholar]
- 22.Kanemoto S, Nitani R, Murakami T, Kaneko M, Asada R, Matsuhisa K, Saito A, Imaizumi K. Multivesicular body formation enhancement and exosome release during endoplasmic reticulum stress. Biochem Biophys Res Commun. 2016;480:166–172. doi: 10.1016/j.bbrc.2016.10.019. [DOI] [PubMed] [Google Scholar]
- 23.Gurunathan S, Kim JH. Graphene oxide enhances biogenesis and release of exosomes in human ovarian cancer cells. Int J Nanomedicine. 2022;17:5697–5731. doi: 10.2147/IJN.S385113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Gurunathan S, Kang MH, Jeyaraj M, Kim JH. Palladium nanoparticle-induced oxidative stress, endoplasmic reticulum stress, apoptosis, and immunomodulation enhance the biogenesis and release of exosome in human leukemia monocytic cells (THP-1) Int J Nanomedicine. 2021;16:2849–2877. doi: 10.2147/IJN.S305269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Anelli T, Sitia R. Protein quality control in the early secretory pathway. EMBO J. 2008;27:315–327. doi: 10.1038/sj.emboj.7601974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Romero-Brey I, Bartenschlager R. Endoplasmic reticulum: the favorite intracellular niche for viral replication and assembly. Viruses. 2016;8:160. doi: 10.3390/v8060160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Smith MH, Ploegh HL, Weissman JS. Road to ruin: targeting proteins for degradation in the endoplasmic reticulum. Science. 2011;334:1086–1090. doi: 10.1126/science.1209235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Dokladny K, Myers OB, Moseley PL. Heat shock response and autophagy–cooperation and control. Autophagy. 2015;11:200–213. doi: 10.1080/15548627.2015.1009776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Shin YJ, Vavra U, Strasser R. Proper protein folding in the endoplasmic reticulum is required for attachment of a glycosylphosphatidylinositol anchor in plants. Plant Physiol. 2021;186:1878–1892. doi: 10.1093/plphys/kiab181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wiseman RL, Mesgarzadeh JS, Hendershot LM. Reshaping endoplasmic reticulum quality control through the unfolded protein response. Mol Cell. 2022;82:1477–1491. doi: 10.1016/j.molcel.2022.03.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Burman A, Tanjore H, Blackwell TS. Endoplasmic reticulum stress in pulmonary fibrosis. Matrix Biol. 2018;68–69:355–365. doi: 10.1016/j.matbio.2018.03.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Tabas I, Ron D. Integrating the mechanisms of apoptosis induced by endoplasmic reticulum stress. Nat Cell Biol. 2011;13:184–190. doi: 10.1038/ncb0311-184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Pahl HL. Signal transduction from the endoplasmic reticulum to the cell nucleus. Physiol Rev. 1999;79:683–701. doi: 10.1152/physrev.1999.79.3.683. [DOI] [PubMed] [Google Scholar]
- 34.Hu H, Tian M, Ding C, Yu S. The C/EBP homologous protein (CHOP) transcription factor functions in endoplasmic reticulum stress-induced apoptosis and microbial infection. Front Immunol. 2018;9:3083. doi: 10.3389/fimmu.2018.03083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Liu D, Zhang Y, Li X, Li J, Yang S, Xing X, Fan G, Yokota H, Zhang P. eIF2alpha signaling regulates ischemic osteonecrosis through endoplasmic reticulum stress. Sci Rep. 2017;7:5062. doi: 10.1038/s41598-017-05488-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Philippe C, Dubrac A, Quelen C, Desquesnes A, Van Den Berghe L, Segura C, Filleron T, Pyronnet S, Prats H, Brousset P, Touriol C. PERK mediates the IRES-dependent translational activation of mRNAs encoding angiogenic growth factors after ischemic stress. Sci Signal. 2016;9:ra44. doi: 10.1126/scisignal.aaf2753. [DOI] [PubMed] [Google Scholar]
- 37.Wang Y, Gao L, Li Y, Chen H, Sun Z. Nifedipine protects INS-1 beta-cell from high glucose-induced ER stress and apoptosis. Int J Mol Sci. 2011;12:7569–7580. doi: 10.3390/ijms12117569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hetz C, Zhang K, Kaufman RJ. Mechanisms, regulation and functions of the unfolded protein response. Nat Rev Mol Cell Biol. 2020;21:421–438. doi: 10.1038/s41580-020-0250-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Chen X, Cubillos-Ruiz JR. Endoplasmic reticulum stress signals in the tumour and its microenvironment. Nat Rev Cancer. 2021;21:71–88. doi: 10.1038/s41568-020-00312-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Shiu RP, Pouyssegur J, Pastan I. Glucose depletion accounts for the induction of two transformation-sensitive membrane proteinsin Rous sarcoma virus-transformed chick embryo fibroblasts. Proc Natl Acad Sci USA. 1977;74:3840–3844. doi: 10.1073/pnas.74.9.3840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Dorner AJ, Wasley LC, Kaufman RJ. Increased synthesis of secreted proteins induces expression of glucose-regulated proteins in butyrate-treated Chinese hamster ovary cells. J Biol Chem. 1989;264:20602–20607. doi: 10.1016/S0021-9258(19)47105-6. [DOI] [PubMed] [Google Scholar]
- 42.Kozutsumi Y, Segal M, Normington K, Gething MJ, Sambrook J. The presence of malfolded proteins in the endoplasmic reticulum signals the induction of glucose-regulated proteins. Nature. 1988;332:462–464. doi: 10.1038/332462a0. [DOI] [PubMed] [Google Scholar]
- 43.Cox JS, Shamu CE, Walter P. Transcriptional induction of genes encoding endoplasmic reticulum resident proteins requires a transmembrane protein kinase. Cell. 1993;73:1197–1206. doi: 10.1016/0092-8674(93)90648-A. [DOI] [PubMed] [Google Scholar]
- 44.Cox JS, Walter P. A novel mechanism for regulating activity of a transcription factor that controls the unfolded protein response. Cell. 1996;87:391–404. doi: 10.1016/S0092-8674(00)81360-4. [DOI] [PubMed] [Google Scholar]
- 45.Adamson B, Norman TM, Jost M, Cho MY, Nunez JK, Chen Y, Villalta JE, Gilbert LA, Horlbeck MA, Hein MY, Pak RA, Gray AN, Gross CA, Dixit A, Parnas O, Regev A, Weissman JS. A multiplexed single-cell CRISPR screening platform enables systematic dissection of the unfolded protein response. Cell. 2016;167:1867–1882 e1821. doi: 10.1016/j.cell.2016.11.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Keestra-Gounder AM, Byndloss MX, Seyffert N, Young BM, Chavez-Arroyo A, Tsai AY, Cevallos SA, Winter MG, Pham OH, Tiffany CR, de Jong MF, Kerrinnes T, Ravindran R, Luciw PA, McSorley SJ, Baumler AJ, Tsolis RM. NOD1 and NOD2 signalling links ER stress with inflammation. Nature. 2016;532:394–397. doi: 10.1038/nature17631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Preissler S, Ron D. Early events in the endoplasmic reticulum unfolded protein response. Cold Spring Harb Perspect Biol. 2019;11:a033894. doi: 10.1101/cshperspect.a033894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Behnke J, Feige MJ, Hendershot LM. BiP and its nucleotide exchange factors Grp170 and Sil1: mechanisms of action and biological functions. J Mol Biol. 2015;427:1589–1608. doi: 10.1016/j.jmb.2015.02.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Cnop M, Toivonen S, Igoillo-Esteve M, Salpea P. Endoplasmic reticulum stress and eIF2alpha phosphorylation: the Achilles heel of pancreatic beta cells. Mol Metab. 2017;6:1024–1039. doi: 10.1016/j.molmet.2017.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Urra H, Dufey E, Avril T, Chevet E, Hetz C. Endoplasmic reticulum stress and the hallmarks of cancer, trends. Cancer. 2016;2:252–262. doi: 10.1016/j.trecan.2016.03.007. [DOI] [PubMed] [Google Scholar]
- 51.Lebeaupin C, Vallee D, Hazari Y, Hetz C, Chevet E, Bailly-Maitre B. Endoplasmic reticulum stress signalling and the pathogenesis of non-alcoholic fatty liver disease. J Hepatol. 2018;69:927–947. doi: 10.1016/j.jhep.2018.06.008. [DOI] [PubMed] [Google Scholar]
- 52.Chen S, Henderson A, Petriello MC, Romano KA, Gearing M, Miao J, Schell M, Sandoval-Espinola WJ, Tao J, Sha B, Graham M, Crooke R, Kleinridders A, Balskus EP, Rey FE, Morris AJ, Biddinger SB. Trimethylamine N-oxide binds and activates PERK to promote metabolic dysfunction. Cell Metab. 2019;30:1141–1151.e1145. doi: 10.1016/j.cmet.2019.08.021. [DOI] [PubMed] [Google Scholar]
- 53.Bettigole SE, Glimcher LH. Endoplasmic reticulum stress in immunity. Annu Rev Immunol. 2015;33:107–138. doi: 10.1146/annurev-immunol-032414-112116. [DOI] [PubMed] [Google Scholar]
- 54.Chen Y, Mi Y, Zhang X, Ma Q, Song Y, Zhang L, Wang D, Xing J, Hou B, Li H, Jin H, Du W, Zou Z. Dihydroartemisinin-induced unfolded protein response feedback attenuates ferroptosis via PERK/ATF4/HSPA5 pathway in glioma cells. J Exp Clin Cancer Res. 2019;38:402. doi: 10.1186/s13046-019-1413-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Mohamed E, Sierra RA, Trillo-Tinoco J, Cao Y, Innamarato P, Payne KK, de Mingo Pulido A, Mandula J, Zhang S, Thevenot P, Biswas S, Abdalla SK, Costich TL, Hanggi K, Anadon CM, Flores ER, Haura EB, Mehrotra S, Pilon-Thomas S, Ruffell B, Munn DH, Cubillos-Ruiz JR, Conejo-Garcia JR, Rodriguez PC. The unfolded protein response mediator pERK governs myeloid cell-driven immunosuppression in tumors through inhibition of STING signaling. Immunity. 2020;52:668–682 e667. doi: 10.1016/j.immuni.2020.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Cubillos-Ruiz JR, Bettigole SE, Glimcher LH. Tumorigenic and immunosuppressive effects of endoplasmic reticulum stress in cancer. Cell. 2017;168:692–706. doi: 10.1016/j.cell.2016.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Jheng JR, Lau KS, Lan YW, Horng JT. A novel role of ER stress signal transducer ATF6 in regulating enterovirus A71 viral protein stability. J Biomed Sci. 2018;25:9. doi: 10.1186/s12929-018-0412-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Hillary RF, FitzGerald U. A lifetime of stress: ATF6 in development and homeostasis. J Biomed Sci. 2018;25:48. doi: 10.1186/s12929-018-0453-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Guerriero CJ, Brodsky JL. The delicate balance between secreted protein folding and endoplasmic reticulum-associated degradation in human physiology. Physiol Rev. 2012;92:537–576. doi: 10.1152/physrev.00027.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Hwang J, Qi L. Quality control in the endoplasmic reticulum: crosstalk between ERAD and UPR pathways. Trends Biochem Sci. 2018;43:593–605. doi: 10.1016/j.tibs.2018.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Hetz C, Papa FR. The unfolded protein response and cell fate control. Mol Cell. 2018;69:169–181. doi: 10.1016/j.molcel.2017.06.017. [DOI] [PubMed] [Google Scholar]
- 62.Corazzari M, Rapino F, Ciccosanti F, Giglio P, Antonioli M, Conti B, Fimia GM, Lovat PE, Piacentini M. Oncogenic BRAF induces chronic ER stress condition resulting in increased basal autophagy and apoptotic resistance of cutaneous melanoma. Cell Death Differ. 2015;22:946–958. doi: 10.1038/cdd.2014.183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Zuo S, Kong D, Wang C, Liu J, Wang Y, Wan Q, Yan S, Zhang J, Tang J, Zhang Q, Lyu L, Li X, Shan Z, Qian L, Shen Y, Yu Y. CRTH2 promotes endoplasmic reticulum stress-induced cardiomyocyte apoptosis through m-calpain. EMBO Mol Med. 2018;10:e8237. doi: 10.15252/emmm.201708237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Oyadomari S, Mori M. Roles of CHOP/GADD153 in endoplasmic reticulum stress. Cell Death Differ. 2004;11:381–389. doi: 10.1038/sj.cdd.4401373. [DOI] [PubMed] [Google Scholar]
- 65.Yamaguchi H, Wang HG. CHOP is involved in endoplasmic reticulum stress-induced apoptosis by enhancing DR5 expression in human carcinoma cells. J Biol Chem. 2004;279:45495–45502. doi: 10.1074/jbc.M406933200. [DOI] [PubMed] [Google Scholar]
- 66.McCullough KD, Martindale JL, Klotz LO, Aw TY, Holbrook NJ. Gadd153 sensitizes cells to endoplasmic reticulum stress by down-regulating Bcl2 and perturbing the cellular redox state. Mol Cell Biol. 2001;21:1249–1259. doi: 10.1128/MCB.21.4.1249-1259.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Liu Y, Zhu H, Yan X, Gu H, Gu Z, Liu F. Endoplasmic reticulum stress participates in the progress of senescence and apoptosis of osteoarthritis chondrocytes. Biochem Biophys Res Commun. 2017;491:368–373. doi: 10.1016/j.bbrc.2017.07.094. [DOI] [PubMed] [Google Scholar]
- 68.Zhu Z, Gao S, Chen C, Xu W, Xiao P, Chen Z, Du C, Chen B, Gao Y, Wang C, Liao J, Huang W. The natural product salicin alleviates osteoarthritis progression by binding to IRE1alpha and inhibiting endoplasmic reticulum stress through the IRE1alpha-IkappaBalpha-p65 signaling pathway. Exp Mol Med. 2022;54:1927–1939. doi: 10.1038/s12276-022-00879-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Sim HJ, Cho C, Kim HE, Hong JY, Song EK, Kwon KY, Jang DG, Kim SJ, Lee HS, Lee C, Kwon T, Yang S, Park TJ. Augmented ERAD (ER-associated degradation) activity in chondrocytes is necessary for cartilage development and maintenance. Sci Adv. 2022;8:eabl4222. doi: 10.1126/sciadv.abl4222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Li Y, Zhang J, Zhou K, Xie L, Xiang G, Fang M, Han W, Wang X, Xiao J. Elevating sestrin2 attenuates endoplasmic reticulum stress and improves functional recovery through autophagy activation after spinal cord injury. Cell Biol Toxicol. 2021;37:401–419. doi: 10.1007/s10565-020-09550-4. [DOI] [PubMed] [Google Scholar]
- 71.Huang Z, Gong J, Lin W, Feng Z, Ma Y, Tu Y, Cai X, Liu J, Lv C, Lv X, Wu Q, Lu W, Zhao J, Ying Y, Li S, Ni W, Chen H. Catalpol as a component of Rehmannia glutinosa protects spinal cord injury by inhibiting endoplasmic reticulum stress-mediated neuronal apoptosis. Front Pharmacol. 2022;13:860757. doi: 10.3389/fphar.2022.860757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Zhan F, Deng Q, Chen Z, Xie C, Xiang S, Qiu S, Tian L, Wu C, Ou Y, Chen J, Xu L. SAR1A regulates the RhoA/YAP and autophagy signaling pathways to influence osteosarcoma invasion and metastasis. Cancer Sci. 2022 doi: 10.1111/cas.15551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Scheiber AL, Guess AJ, Kaito T, Abzug JM, Enomoto-Iwamoto M, Leikin S, Iwamoto M, Otsuru S. Endoplasmic reticulum stress is induced in growth plate hypertrophic chondrocytes in G610C mouse model of osteogenesis imperfecta. Biochem Biophys Res Commun. 2019;509:235–240. doi: 10.1016/j.bbrc.2018.12.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Duran I, Zieba J, Csukasi F, Martin JH, Wachtell D, Barad M, Dawson B, Fafilek B, Jacobsen CM, Ambrose CG, Cohn DH, Krejci P, Lee BH, Krakow D. 4-PBA treatment improves bone phenotypes in the Aga2 mouse model of osteogenesis imperfecta. J Bone Miner Res. 2022;37:675–686. doi: 10.1002/jbmr.4501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Xiang Z, Wu Q, Wang Y, Wang P, He Y, Li J. eIF2alpha-ATF4 pathway activated by a change in the calcium environment participates in BCP-mediated bone regeneration. ACS Biomater Sci Eng. 2021;7:3256–3268. doi: 10.1021/acsbiomaterials.0c01802. [DOI] [PubMed] [Google Scholar]
- 76.Zheng C, Attarilar S, Li K, Wang C, Liu J, Wang L, Yang J, Tang Y. 3D-printed HA15-loaded β-tricalcium phosphate/poly (lactic-co-glycolic acid) bone tissue scaffold promotes bone regeneration in rabbit radial defects. Int J Bioprinting. 2021;7:317. doi: 10.18063/ijb.v7i1.317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Strong AL, Spreadborough PJ, Dey D, Yang P, Li S, Lee A, Haskins RM, Grimm PD, Kumar R, Bradley MJ, Yu PB, Levi B, Davis TA. BMP ligand trap ALK3-Fc attenuates osteogenesis and heterotopic ossification in blast-related lower extremity trauma. Stem Cells Dev. 2021;30:91–105. doi: 10.1089/scd.2020.0162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Chen L, Zou X, Zhang RX, Pi CJ, Wu N, Yin LJ, Deng ZL. IGF1 potentiates BMP9-induced osteogenic differentiation in mesenchymal stem cells through the enhancement of BMP/Smad signaling. BMB Rep. 2016;49:122–127. doi: 10.5483/BMBRep.2016.49.2.228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Moore KA, Hollien J. The unfolded protein response in secretory cell function. Annu Rev Genet. 2012;46:165–183. doi: 10.1146/annurev-genet-110711-155644. [DOI] [PubMed] [Google Scholar]
- 80.Zhang J, Weng Y, Liu X, Wang J, Zhang W, Kim SH, Zhang H, Li R, Kong Y, Chen X, Shui W, Wang N, Zhao C, Wu N, He Y, Nan G, Chen X, Wen S, Zhang H, Deng F, Wan L, Luu HH, Haydon RC, Shi LL, He TC, Shi Q. Endoplasmic reticulum (ER) stress inducible factor cysteine-rich with EGF-like domains 2 (Creld2) is an important mediator of BMP9-regulated osteogenic differentiation of mesenchymal stem cells. PLoS ONE. 2013;8:e73086. doi: 10.1371/journal.pone.0073086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Doron B, Abdelhamed S, Butler JT, Hashmi SK, Horton TM, Kurre P. Transmissible ER stress reconfigures the AML bone marrow compartment. Leukemia. 2019;33:918–930. doi: 10.1038/s41375-018-0254-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Horiuchi K, Tohmonda T, Morioka H. The unfolded protein response in skeletal development and homeostasis. Cell Mol Life Sci. 2016;73:2851–2869. doi: 10.1007/s00018-016-2178-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Hughes A, Oxford AE, Tawara K, Jorcyk CL, Oxford JT. Endoplasmic reticulum stress and unfolded protein response in cartilage pathophysiology; contributing factors to apoptosis and osteoarthritis. Int J Mol Sci. 2017;18:665. doi: 10.3390/ijms18030665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Nakashima K, Zhou X, Kunkel G, Zhang Z, Deng JM, Behringer RR, de Crombrugghe B. The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone formation. Cell. 2002;108:17–29. doi: 10.1016/S0092-8674(01)00622-5. [DOI] [PubMed] [Google Scholar]
- 85.Kousteni S. Osterix finds its master. EMBO Rep. 2011;12:382–383. doi: 10.1038/embor.2011.61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Tohmonda T, Miyauchi Y, Ghosh R, Yoda M, Uchikawa S, Takito J, Morioka H, Nakamura M, Iwawaki T, Chiba K, Toyama Y, Urano F, Horiuchi K. The IRE1alpha-XBP1 pathway is essential for osteoblast differentiation through promoting transcription of Osterix. EMBO Rep. 2011;12:451–457. doi: 10.1038/embor.2011.34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.He S, Fu T, Yu Y, Liang Q, Li L, Liu J, Zhang X, Zhou Q, Guo Q, Xu D, Chen Y, Wang X, Chen Y, Liu J, Gan Z, Liu Y. IRE1alpha regulates skeletal muscle regeneration through myostatin mRNA decay. J Clin Invest. 2021 doi: 10.1172/JCI143737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Shioi A, Ikari Y. Plaque calcification during atherosclerosis progression and regression. J Atheroscler Thromb. 2018;25:294–303. doi: 10.5551/jat.RV17020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Yang X, Matsuda K, Bialek P, Jacquot S, Masuoka HC, Schinke T, Li L, Brancorsini S, Sassone-Corsi P, Townes TM, Hanauer A, Karsenty G. ATF4 is a substrate of RSK2 and an essential regulator of osteoblast biology; implication for Coffin-Lowry syndrome. Cell. 2004;117:387–398. doi: 10.1016/S0092-8674(04)00344-7. [DOI] [PubMed] [Google Scholar]
- 90.Saito A, Ochiai K, Kondo S, Tsumagari K, Murakami T, Cavener DR, Imaizumi K. Endoplasmic reticulum stress response mediated by the PERK-eIF2(alpha)-ATF4 pathway is involved in osteoblast differentiation induced by BMP2. J Biol Chem. 2011;286:4809–4818. doi: 10.1074/jbc.M110.152900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Shi M, Song W, Han T, Chang B, Li G, Jin J, Zhang Y. Role of the unfolded protein response in topography-induced osteogenic differentiation in rat bone marrow mesenchymal stem cells. Acta Biomater. 2017;54:175–185. doi: 10.1016/j.actbio.2017.03.018. [DOI] [PubMed] [Google Scholar]
- 92.Yang SY, Wei FL, Hu LH, Wang CL. PERK-eIF2alpha-ATF4 pathway mediated by endoplasmic reticulum stress response is involved in osteodifferentiation of human periodontal ligament cells under cyclic mechanical force. Cell Signal. 2016;28:880–886. doi: 10.1016/j.cellsig.2016.04.003. [DOI] [PubMed] [Google Scholar]
- 93.Wei J, Sheng X, Feng D, McGrath B, Cavener DR. PERK is essential for neonatal skeletal development to regulate osteoblast proliferation and differentiation. J Cell Physiol. 2008;217:693–707. doi: 10.1002/jcp.21543. [DOI] [PubMed] [Google Scholar]
- 94.Jang WG, Kim EJ, Kim DK, Ryoo HM, Lee KB, Kim SH, Choi HS, Koh JT. BMP2 protein regulates osteocalcin expression via Runx2-mediated Atf6 gene transcription. J Biol Chem. 2012;287:905–915. doi: 10.1074/jbc.M111.253187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Park SJ, Kim KJ, Kim WU, Oh IH, Cho CS. Involvement of endoplasmic reticulum stress in homocysteine-induced apoptosis of osteoblastic cells. J Bone Miner Metab. 2012;30:474–484. doi: 10.1007/s00774-011-0346-9. [DOI] [PubMed] [Google Scholar]
- 96.Xue P, Li B, An Y, Sun J, He X, Hou R, Dong G, Fei D, Jin F, Wang Q, Jin Y. Decreased MORF leads to prolonged endoplasmic reticulum stress in periodontitis-associated chronic inflammation. Cell Death Differ. 2016;23:1862–1872. doi: 10.1038/cdd.2016.74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Li H, Li D, Ma Z, Qian Z, Kang X, Jin X, Li F, Wang X, Chen Q, Sun H, Wu S. Defective autophagy in osteoblasts induces endoplasmic reticulum stress and causes remarkable bone loss. Autophagy. 2018;14:1726–1741. doi: 10.1080/15548627.2018.1483807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Pereira RC, Stadmeyer LE, Smith DL, Rydziel S, Canalis E. CCAAT/enhancer-binding protein homologous protein (CHOP) decreases bone formation and causes osteopenia. Bone. 2007;40:619–626. doi: 10.1016/j.bone.2006.09.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Wu CT, Chen YW, Su YH, Chiu CY, Guan SS, Yang RS, Liu SH. Gender difference of CCAAT/enhancer binding protein homologous protein deficiency in susceptibility to osteopenia. J Orthop Res. 2019;37:942–947. doi: 10.1002/jor.24264. [DOI] [PubMed] [Google Scholar]
- 100.Shirakawa K, Maeda S, Gotoh T, Hayashi M, Shinomiya K, Ehata S, Nishimura R, Mori M, Onozaki K, Hayashi H, Uematsu S, Akira S, Ogata E, Miyazono K, Imamura T. CCAAT/enhancer-binding protein homologous protein (CHOP) regulates osteoblast differentiation. Mol Cell Biol. 2006;26:6105–6116. doi: 10.1128/MCB.02429-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Yan Y, Wang G, Luo X, Zhang P, Peng S, Cheng X, Wang M, Yang X. Endoplasmic reticulum stress-related calcium imbalance plays an important role on Zinc oxide nanoparticles-induced failure of neural tube closure during embryogenesis. Environ Int. 2021;152:106495. doi: 10.1016/j.envint.2021.106495. [DOI] [PubMed] [Google Scholar]
- 102.Vassalle M, Lin C-I. Calcium overload and cardiac function. J Biomed Sci. 2004;11:542–565. doi: 10.1007/BF02256119. [DOI] [PubMed] [Google Scholar]
- 103.Mohsin AA, Thompson J, Hu Y, Hollander J, Lesnefsky EJ, Chen Q. Endoplasmic reticulum stress-induced complex I defect: central role of calcium overload. Arch Biochem Biophys. 2020;683:108299. doi: 10.1016/j.abb.2020.108299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Cheng Z, Liu Y, Ma M, Sun S, Ma Z, Wang Y, Yu L, Qian X, Sun L, Zhang X, Liu Y, Wang Y. Lansoprazole-induced osteoporosis via the IP3R- and SOCE-mediated calcium signaling pathways. Mol Med. 2022;28:21. doi: 10.1186/s10020-022-00448-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Cui C, Lin T, Gong Z, Zhu Y. Relationship between autophagy, apoptosis and endoplasmic reticulum stress induced by melatonin in osteoblasts by septin7 expression. Mol Med Rep. 2020;21:2427–2434. doi: 10.3892/mmr.2020.11063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Liu W, Xu C, Ran D, Wang Y, Zhao H, Gu J, Liu X, Bian J, Yuan Y, Liu Z. CaMK mediates cadmium induced apoptosis in rat primary osteoblasts through MAPK activation and endoplasmic reticulum stress. Toxicology. 2018;406–407:70–80. doi: 10.1016/j.tox.2018.06.002. [DOI] [PubMed] [Google Scholar]
- 107.Li X, Meng L, Wang F, Hu X, Yu Y. Sodium fluoride induces apoptosis and autophagy via the endoplasmic reticulum stress pathway in MC3T3-E1 osteoblastic cells. Mol Cell Biochem. 2019;454:77–85. doi: 10.1007/s11010-018-3454-1. [DOI] [PubMed] [Google Scholar]
- 108.Wang J, Zhao Y, Cheng X, Li Y, Xu H, Manthari RK, Wang J. Effects of different Ca(2+) level on fluoride-induced apoptosis pathway of endoplasmic reticulum in the rabbit osteoblast in vitro. Food Chem Toxicol. 2018;116:189–195. doi: 10.1016/j.fct.2018.04.013. [DOI] [PubMed] [Google Scholar]
- 109.Che J, Lv H, Yang J, Zhao B, Zhou S, Yu T, Shang P. Iron overload induces apoptosis of osteoblast cells via eliciting ER stress-mediated mitochondrial dysfunction and p-eIF2alpha/ATF4/CHOP pathway in vitro. Cell Signal. 2021;84:110024. doi: 10.1016/j.cellsig.2021.110024. [DOI] [PubMed] [Google Scholar]
- 110.Tan J, Zhou Y, Luo J, Wu X, Liu H, Wang W, Li Z, Zhong M, Wu L, Li X. High glucose inhibits the osteogenic differentiation of periodontal ligament stem cells in periodontitis by activating endoplasmic reticulum stress. Ann Transl Med. 2022;10:204. doi: 10.21037/atm-22-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Cain DW, Cidlowski JA. Immune regulation by glucocorticoids. Nat Rev Immunol. 2017;17:233–247. doi: 10.1038/nri.2017.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Chang C, Greenspan A, Gershwin ME. The pathogenesis, diagnosis and clinical manifestations of steroid-induced osteonecrosis. J Autoimmun. 2020;110:102460. doi: 10.1016/j.jaut.2020.102460. [DOI] [PubMed] [Google Scholar]
- 113.Liu W, Zhao Z, Na Y, Meng C, Wang J, Bai R. Dexamethasone-induced production of reactive oxygen species promotes apoptosis via endoplasmic reticulum stress and autophagy in MC3T3-E1 cells. Int J Mol Med. 2018;41:2028–2036. doi: 10.3892/ijmm.2018.3412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Sato AY, Tu X, McAndrews KA, Plotkin LI, Bellido T. Prevention of glucocorticoid induced-apoptosis of osteoblasts and osteocytes by protecting against endoplasmic reticulum (ER) stress in vitro and in vivo in female mice. Bone. 2015;73:60–68. doi: 10.1016/j.bone.2014.12.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Huang J, Ye Y, Xiao Y, Ren Q, Zhou Q, Zhong M, Jiao L, Wu L. Geniposide ameliorates glucocorticoid-induced osteoblast apoptosis by activating autophagy. Biomed Pharmacother. 2022;155:113829. doi: 10.1016/j.biopha.2022.113829. [DOI] [PubMed] [Google Scholar]
- 116.Gao Y, Zhu H, Wang Q, Feng Y, Zhang C. Inhibition of PERK signaling prevents against glucocorticoid-induced endotheliocyte apoptosis and osteonecrosis of the femoral head. Int J Biol Sci. 2020;16:543–552. doi: 10.7150/ijbs.35256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Xiao Y, Ren Q, Zheng Y, Zhang S, Ouyang J, Jiao L, Tang C, Li L, Shi W, Wang M, Zhang S, Zhang D, Zhong B, Peng F, Chen Z, Wu L. Geniposide ameliorated dexamethasone-induced endoplasmic reticulum stress and mitochondrial apoptosis in osteoblasts. J Ethnopharmacol. 2022;291:115154. doi: 10.1016/j.jep.2022.115154. [DOI] [PubMed] [Google Scholar]
- 118.Feng Y, Zhang R, Wang YR, Chen F, Luo Q, Cai C, Jiao Y, Xue P. Inhibition of endoplasmic reticulum stress by 4-phenyl butyric acid presents therapeutic effects on periodontitis: experimental studies in vitro and in rats. Stem Cells Int. 2021;2021:6618943. doi: 10.1155/2021/6618943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Zhou R, Ma Y, Tao Z, Qiu S, Gong Z, Tao L, Zhu Y. Melatonin inhibits glucose-induced apoptosis in osteoblastic cell line through PERK-eIF2alpha-ATF4 pathway. Front Pharmacol. 2020;11:602307. doi: 10.3389/fphar.2020.602307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Yang L, Liu S, Mu S, Guo R, Zhou L, Fu Q. Paeoniflorin attenuates dexamethasone-induced apoptosis of osteoblast cells and promotes bone formation via regulating AKT/mTOR/autophagy signaling pathway. Evid Based Complement Alternat Med. 2021;2021:6623464. doi: 10.1155/2021/6623464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Duangchan T, Tawonsawatruk T, Angsanuntsukh C, Trachoo O, Hongeng S, Kitiyanant N, Supokawej A. Amelioration of osteogenesis in iPSC-derived mesenchymal stem cells from osteogenesis imperfecta patients by endoplasmic reticulum stress inhibitor. Life Sci. 2021;278:119628. doi: 10.1016/j.lfs.2021.119628. [DOI] [PubMed] [Google Scholar]
- 122.Li J, Yang S, Li X, Liu D, Wang Z, Guo J, Tan N, Gao Z, Zhao X, Zhang J, Gou F, Yokota H, Zhang P. Role of endoplasmic reticulum stress in disuse osteoporosis. Bone. 2017;97:2–14. doi: 10.1016/j.bone.2016.12.009. [DOI] [PubMed] [Google Scholar]
- 123.Tang J, Wu T, Xiong J, Su Y, Zhang C, Wang S, Tang Z, Liu Y. Porphyromonas gingivalis lipopolysaccharides regulate functions of bone marrow mesenchymal stem cells. Cell Prolif. 2015;48:239–248. doi: 10.1111/cpr.12173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Kuang W, Zheng L, Xu X, Lin Y, Lin J, Wu J, Tan J. Dysregulation of the miR-146a-Smad4 axis impairs osteogenesis of bone mesenchymal stem cells under inflammation. Bone Res. 2017;5:17037. doi: 10.1038/boneres.2017.37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Victor P, Sarada D, Ramkumar KM. Crosstalk between endoplasmic reticulum stress and oxidative stress: focus on protein disulfide isomerase and endoplasmic reticulum oxidase 1. Eur J Pharmacol. 2021;892:173749. doi: 10.1016/j.ejphar.2020.173749. [DOI] [PubMed] [Google Scholar]
- 126.Jin S, Park JY, Hong JM, Kim TH, Shin HI, Park EK, Kim SY. Inhibitory effect of (-)-epigallocatechin gallate on titanium particle-induced TNF-α release and in vivo osteolysis. Exp Mol Med. 2011;43:411–418. doi: 10.3858/emm.2011.43.7.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Li H, Deng Y, Tan M, Feng G, Kuang Y, Li J, Song J. Low-intensity pulsed ultrasound upregulates osteogenesis under inflammatory conditions in periodontal ligament stem cells through unfolded protein response. Stem Cell Res Ther. 2020;11:215. doi: 10.1186/s13287-020-01732-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Zhao X, Zhang G, Wu L, Tang Y, Guo C. Inhibition of ER stress-activated JNK pathway attenuates TNF-alpha-induced inflammatory response in bone marrow mesenchymal stem cells. Biochem Biophys Res Commun. 2021;541:8–14. doi: 10.1016/j.bbrc.2020.12.101. [DOI] [PubMed] [Google Scholar]
- 129.Ma S, Zhang Y, Li S, Li A, Li Y, Pei D. Engineering exosomes for bone defect repair. Front Bioeng Biotechnol. 2022;10:1091360. doi: 10.3389/fbioe.2022.1091360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Ren YZ, Ding SS, Jiang YP, Wen H, Li T. Application of exosome-derived noncoding RNAs in bone regeneration: opportunities and challenges. World J Stem Cells. 2022;14:473–489. doi: 10.4252/wjsc.v14.i7.473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Qu M, Zou X, Fang F, Wang S, Xu L, Zeng Q, Fan Z, Chen L, Yue W, Xie X, Pei X. Platelet-derived microparticles enhance megakaryocyte differentiation and platelet generation via miR-1915-3p. Nat Commun. 2020;11:4964. doi: 10.1038/s41467-020-18802-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Tao SC, Yuan T, Rui BY, Zhu ZZ, Guo SC, Zhang CQ. Exosomes derived from human platelet-rich plasma prevent apoptosis induced by glucocorticoid-associated endoplasmic reticulum stress in rat osteonecrosis of the femoral head via the Akt/Bad/Bcl-2 signal pathway. Theranostics. 2017;7:733–750. doi: 10.7150/thno.17450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Wang Y, Fan A, Lu L, Pan Z, Ma M, Luo S, Liu Z, Yang L, Cai J, Yin F. Exosome modification to better alleviates endoplasmic reticulum stress induced chondrocyte apoptosis and osteoarthritis. Biochem Pharmacol. 2022;206:115343. doi: 10.1016/j.bcp.2022.115343. [DOI] [PubMed] [Google Scholar]
- 134.Liao Z, Luo R, Li G, Song Y, Zhan S, Zhao K, Hua W, Zhang Y, Wu X, Yang C. Exosomes from mesenchymal stem cells modulate endoplasmic reticulum stress to protect against nucleus pulposus cell death and ameliorate intervertebral disc degeneration in vivo. Theranostics. 2019;9:4084–4100. doi: 10.7150/thno.33638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Son HE, Kim EJ, Jang WG. Curcumin induces osteoblast differentiation through mild-endoplasmic reticulum stress-mediated such as BMP2 on osteoblast cells. Life Sci. 2018;193:34–39. doi: 10.1016/j.lfs.2017.12.008. [DOI] [PubMed] [Google Scholar]
- 136.Kong Y, Zhang Y, Cai Y, Li D, Yi B, Xu Q. METTL3 mediates osteoblast apoptosis by regulating endoplasmic reticulum stress during LPS-induced inflammation. Cell Signal. 2022;95:110335. doi: 10.1016/j.cellsig.2022.110335. [DOI] [PubMed] [Google Scholar]
- 137.Jin N, Jin N, Wang Z, Liu L, Meng L, Li D, Li X, Zhou D, Liu J, Bu W, Sun H, Yang B. Osteopromotive carbon dots promote bone regeneration through the PERK-eIF2alpha-ATF4 pathway. Biomater Sci. 2020;8:2840–2852. doi: 10.1039/D0BM00424C. [DOI] [PubMed] [Google Scholar]
- 138.Zhang D, De Veirman K, Fan R, Jian Q, Zhang Y, Lei L, Evans H, Wang Y, Lei L, Wang B, Williamson RA, Chantry A, He P, Li A, De Raeve H, Vanderkerken K, He A, Hu J. ER stress arm XBP1s plays a pivotal role in proteasome inhibition-induced bone formation. Stem Cell Res Ther. 2020;11:516. doi: 10.1186/s13287-020-02037-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Meng X, Zhu Y, Tao L, Zhao S, Qiu S. Periostin has a protective role in melatonininduced cell apoptosis by inhibiting the eIF2alphaATF4 pathway in human osteoblasts. Int J Mol Med. 2018;41:1003–1012. doi: 10.3892/ijmm.2017.3300. [DOI] [PubMed] [Google Scholar]
- 140.Suzuki R, Fujiwara Y, Saito M, Arakawa S, Shirakawa JI, Yamanaka M, Komohara Y, Marumo K, Nagai R. Intracellular accumulation of advanced glycation end products induces osteoblast apoptosis via endoplasmic reticulum stress. J Bone Miner Res. 2020;35:1992–2003. doi: 10.1002/jbmr.4053. [DOI] [PubMed] [Google Scholar]
- 141.Tan J, Zhou L, Xue P, An Y, Luo L, Zhang R, Wu G, Wang Y, Zhu H, Wang Q. Tumor necrosis factor-alpha attenuates the osteogenic differentiation capacity of periodontal ligament stem cells by activating PERK signaling. J Periodontol. 2016;87:e159–171. doi: 10.1902/jop.2016.150718. [DOI] [PubMed] [Google Scholar]
- 142.Han C, Xie K, Yang C, Zhang F, Liang Q, Lan C, Chen J, Huang K, Liu J, Li K, Tang Y, Wang L. HA15 alleviates bone loss in ovariectomy-induced osteoporosis by targeting HSPA5. Exp Cell Res. 2021;406:112781. doi: 10.1016/j.yexcr.2021.112781. [DOI] [PubMed] [Google Scholar]
- 143.Mengqi S, Wen S, Boxin Z, Minni L, Yan Z, Qun W, Yumei Z. Micro/nano topography with altered nanotube diameter differentially trigger endoplasmic reticulum stress to mediate bone mesenchymal stem cell osteogenic differentiation. Biomed Mater. 2020;16:015024. doi: 10.1088/1748-605X/abbfee. [DOI] [PubMed] [Google Scholar]
- 144.Jing L, Jia XW. Lycium barbarum polysaccharide arbitrates palmitate-induced apoptosis in MC3T3E1 cells through decreasing the activation of ERSmediated apoptosis pathway. Mol Med Rep. 2018;17:2415–2421. doi: 10.3892/mmr.2017.8128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Son HE, Min HY, Kim EJ, Jang WG. Fat mass and obesity-associated (FTO) stimulates osteogenic differentiation of C3H10T1/2 cells by inducing mild endoplasmic reticulum stress via a positive feedback loop with p-AMPK. Mol Cells. 2020;43:58–65. doi: 10.14348/molcells.2019.0136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Yang F, Tang J, Dai K, Huang Y. Metallic wear debris collected from patients induces apoptosis in rat primary osteoblasts via reactive oxygen speciesmediated mitochondrial dysfunction and endoplasmic reticulum stress. Mol Med Rep. 2019;19:1629–1637. doi: 10.3892/mmr.2019.9825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Niu J, Wang K, Zhelyabovska O, Saad Y, Kolattukudy PE. MCP-1-induced protein promotes endothelial-like and angiogenic properties in human bone marrow monocytic cells. J Pharmacol Exp Ther. 2013;347:288–297. doi: 10.1124/jpet.113.207316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Roy A, Kolattukudy PE. Monocyte chemotactic protein-induced protein (MCPIP) promotes inflammatory angiogenesis via sequential induction of oxidative stress, endoplasmic reticulum stress and autophagy. Cell Signal. 2012;24:2123–2131. doi: 10.1016/j.cellsig.2012.07.014. [DOI] [PubMed] [Google Scholar]
- 149.Soczewski E, Grasso E, Gallino L, Hauk V, Fernandez L, Gori S, Paparini D, Perez Leiros C, Ramhorst R. Immunoregulation of the decidualization program: focus on the endoplasmic reticulum stress. Reproduction. 2020;159:R203–R211. doi: 10.1530/REP-19-0391. [DOI] [PubMed] [Google Scholar]
- 150.Pereira ER, Liao N, Neale GA, Hendershot LM. Transcriptional and post-transcriptional regulation of proangiogenic factors by the unfolded protein response. PLoS ONE. 2010;5:e12521. doi: 10.1371/journal.pone.0012521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Ghosh R, Lipson KL, Sargent KE, Mercurio AM, Hunt JS, Ron D, Urano F. Transcriptional regulation of VEGF-A by the unfolded protein response pathway. PLoS ONE. 2010;5:e9575. doi: 10.1371/journal.pone.0009575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Miyagi H, Kanemoto S, Saito A, Asada R, Iwamoto H, Izumi S, Kido M, Gomi F, Nishida K, Kiuchi Y, Imaizumi K. Transcriptional regulation of VEGFA by the endoplasmic reticulum stress transducer OASIS in ARPE-19 cells. PLoS ONE. 2013;8:e55155. doi: 10.1371/journal.pone.0055155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Castranova D, Davis AE, Lo BD, Miller MF, Paukstelis PJ, Swift MR, Pham VN, Torres-Vazquez J, Bell K, Shaw KM, Kamei M, Weinstein BM. Aminoacyl-transfer RNA synthetase deficiency promotes angiogenesis via the unfolded protein response pathway. Arterioscler Thromb Vasc Biol. 2016;36:655–662. doi: 10.1161/ATVBAHA.115.307087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Loeuillard E, El Mourabit H, Lei L, Lemoinne S, Housset C, Cadoret A. Endoplasmic reticulum stress induces inverse regulations of major functions in portal myofibroblasts during liver fibrosis progression. Biochim Biophys Acta Mol Basis Dis. 1864;2018:3688–3696. doi: 10.1016/j.bbadis.2018.10.008. [DOI] [PubMed] [Google Scholar]
- 155.Ozawa K, Kondo T, Hori O, Kitao Y, Stern DM, Eisenmenger W, Ogawa S, Ohshima T. Expression of the oxygen-regulated protein ORP150 accelerates wound healing by modulating intracellular VEGF transport. J Clin Investig. 2001;108:41–50. doi: 10.1172/JCI200111772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Drogat B, Auguste P, Nguyen DT, Bouchecareilh M, Pineau R, Nalbantoglu J, Kaufman RJ, Chevet E, Bikfalvi A, Moenner M. IRE1 signaling is essential for ischemia-induced vascular endothelial growth factor-A expression and contributes to angiogenesis and tumor growth in vivo. Cancer Res. 2007;67:6700–6707. doi: 10.1158/0008-5472.CAN-06-3235. [DOI] [PubMed] [Google Scholar]
- 157.Wu SX, Ye SS, Hong YX, Chen Y, Wang B, Lin XJ, Lin X. Hepatitis B virus small envelope protein promotes hepatocellular carcinoma angiogenesis via endoplasmic reticulum stress signaling to upregulate the expression of vascular endothelial growth factor A. J Virol. 2022;96:e0197521. doi: 10.1128/jvi.01975-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Zeng L, Xiao Q, Chen M, Margariti A, Martin D, Ivetic A, Xu H, Mason J, Wang W, Cockerill G, Mori K, Li JY, Chien S, Hu Y, Xu Q. Vascular endothelial cell growth-activated XBP1 splicing in endothelial cells is crucial for angiogenesis. Circulation. 2013;127:1712–1722. doi: 10.1161/CIRCULATIONAHA.112.001337. [DOI] [PubMed] [Google Scholar]
- 159.Zhu K, Jiao H, Li S, Cao H, Galson DL, Zhao Z, Zhao X, Lai Y, Fan J, Im HJ, Chen D, Xiao G. ATF4 promotes bone angiogenesis by increasing VEGF expression and release in the bone environment. J Bone Miner Res. 2013;28:1870–1884. doi: 10.1002/jbmr.1958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Soczewski E, Gori S, Paparini D, Grasso E, Fernandez L, Gallino L, Schafir A, Irigoyen M, Lobo TF, Salamone G, Mattar R, Daher S, Perez Leiros C, Ramhorst R. VIP conditions human endometrial receptivity by privileging endoplasmic reticulum stress through ATF6alpha pathway. Mol Cell Endocrinol. 2020;516:110948. doi: 10.1016/j.mce.2020.110948. [DOI] [PubMed] [Google Scholar]
- 161.Binet F, Sapieha P. ER stress and angiogenesis. Cell Metab. 2015;22:560–575. doi: 10.1016/j.cmet.2015.07.010. [DOI] [PubMed] [Google Scholar]
- 162.Zou J, Fei Q, Xiao H, Wang H, Liu K, Liu M, Zhang H, Xiao X, Wang K, Wang N. VEGF-A promotes angiogenesis after acute myocardial infarction through increasing ROS production and enhancing ER stress-mediated autophagy. J Cell Physiol. 2019;234:17690–17703. doi: 10.1002/jcp.28395. [DOI] [PubMed] [Google Scholar]
- 163.Li J, Li Z, Wang C, Li Z, Xu H, Hu Y, Tan Z, Zhang F, Liu C, Yang M, Wang Y, Jin Y, Peng Z, Biswas S, Zhu L. The regulatory effect of VEGF-Ax on rat bone marrow mesenchymal stem cells' angioblastic differentiation and its proangiogenic ability. Stem Cells Dev. 2020;29:667–677. doi: 10.1089/scd.2019.0198. [DOI] [PubMed] [Google Scholar]
- 164.Gargalovic PS, Imura M, Zhang B, Gharavi NM, Clark MJ, Pagnon J, Yang WP, He A, Truong A, Patel S, Nelson SF, Horvath S, Berliner JA, Kirchgessner TG, Lusis AJ. Identification of inflammatory gene modules based on variations of human endothelial cell responses to oxidized lipids. Proc Natl Acad Sci USA. 2006;103:12741–12746. doi: 10.1073/pnas.0605457103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Vandewynckel YP, Laukens D, Devisscher L, Bogaerts E, Paridaens A, Van den Bussche A, Raevens S, Verhelst X, Van Steenkiste C, Jonckx B, Libbrecht L, Geerts A, Carmeliet P, Van Vlierberghe H. Placental growth factor inhibition modulates the interplay between hypoxia and unfolded protein response in hepatocellular carcinoma. BMC Cancer. 2016;16:9. doi: 10.1186/s12885-015-1990-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Ying R, Li SW, Chen JY, Zhang HF, Yang Y, Gu ZJ, Chen YX, Wang JF. Endoplasmic reticulum stress in perivascular adipose tissue promotes destabilization of atherosclerotic plaque by regulating GM-CSF paracrine. J Transl Med. 2018;16:105. doi: 10.1186/s12967-018-1481-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.O'Neill CL, Guduric-Fuchs J, Chambers SE, O'Doherty M, Bottazzi B, Stitt AW, Medina RJ. Endothelial cell-derived pentraxin 3 limits the vasoreparative therapeutic potential of circulating angiogenic cells. Cardiovasc Res. 2016;112:677–688. doi: 10.1093/cvr/cvw209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Rusnati M, Camozzi M, Moroni E, Bottazzi B, Peri G, Indraccolo S, Amadori A, Mantovani A, Presta M. Selective recognition of fibroblast growth factor-2 by the long pentraxin PTX3 inhibits angiogenesis. Blood. 2004;104:92–99. doi: 10.1182/blood-2003-10-3433. [DOI] [PubMed] [Google Scholar]
- 169.Ma M, Yang W, Cai Z, Wang P, Li H, Mi R, Jiang Y, Xie Z, Sui P, Wu Y, Shen H. SMAD-specific E3 ubiquitin ligase 2 promotes angiogenesis by facilitating PTX3 degradation in MSCs from patients with ankylosing spondylitis. Stem Cells. 2021;39:581–599. doi: 10.1002/stem.3332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Yu H, Wen K, Zhou X, Zhang Y, Yan Z, Fu H, Zhu J, Zhu Y. Role of unfolded protein response in genital malformation/damage of male mice induced by flutamide. Hum Exp Toxicol. 2020;39:1690–1699. doi: 10.1177/0960327120937049. [DOI] [PubMed] [Google Scholar]
- 171.Abdelsalam S, Pasha M, El-Gamal H, Hasan M, Elrayess M, Zeidan A, Korashy H, Agouni A. Protein tyrosine phosphatase 1B inhibition improves endoplasmic reticulum stress-impaired endothelial cell angiogenic response: a critical role for cell survival. Mol Med Rep. 2021;24:1. doi: 10.3892/mmr.2021.12304. [DOI] [PubMed] [Google Scholar]
- 172.Maamoun H, Zachariah M, McVey JH, Green FR, Agouni A. Heme oxygenase (HO)-1 induction prevents endoplasmic reticulum stress-mediated endothelial cell death and impaired angiogenic capacity. Biochem Pharmacol. 2017;127:46–59. doi: 10.1016/j.bcp.2016.12.009. [DOI] [PubMed] [Google Scholar]
- 173.Lokeswara AW, Hiksas R, Irwinda R, Wibowo N. Preeclampsia: from cellular wellness to inappropriate cell death, and the roles of nutrition. Front Cell Dev Biol. 2021;9:726513. doi: 10.3389/fcell.2021.726513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Jaud M, Philippe C, Van Den Berghe L, Segura C, Mazzolini L, Pyronnet S, Laurell H, Touriol C. The PERK Branch of the Unfolded Protein Response Promotes DLL4 Expression by Activating an Alternative Translation Mechanism. Cancers (Basel) 2019;11:142. doi: 10.3390/cancers11020142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Tang V, Fu S, Rayner BS, Hawkins CL. 8-Chloroadenosine induces apoptosis in human coronary artery endothelial cells through the activation of the unfolded protein response. Redox Biol. 2019;26:101274. doi: 10.1016/j.redox.2019.101274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Zeng L, Zampetaki A, Margariti A, Pepe AE, Alam S, Martin D, Xiao Q, Wang W, Jin ZG, Cockerill G, Mori K, Li YS, Hu Y, Chien S, Xu Q. Sustained activation of XBP1 splicing leads to endothelial apoptosis and atherosclerosis development in response to disturbed flow. Proc Natl Acad Sci U S A. 2009;106:8326–8331. doi: 10.1073/pnas.0903197106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Huang J, Wan L, Lu H, Li X. High expression of active ATF6 aggravates endoplasmic reticulum stressinduced vascular endothelial cell apoptosis through the mitochondrial apoptotic pathway. Mol Med Rep. 2018;17:6483–6489. doi: 10.3892/mmr.2018.8658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Zachariah M, Maamoun H, Milano L, Rayman MP, Meira LB, Agouni A. Endoplasmic reticulum stress and oxidative stress drive endothelial dysfunction induced by high selenium. J Cell Physiol. 2020;236:4348–4359. doi: 10.1002/jcp.30175. [DOI] [PubMed] [Google Scholar]
- 179.Maamoun H, Abdelsalam SS, Zeidan A, Korashy HM, Agouni A. Endoplasmic reticulum stress: a critical molecular driver of endothelial dysfunction and cardiovascular disturbances associated with diabetes. Int J Mol Sci. 2019;20:1658. doi: 10.3390/ijms20071658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Shimoda Y, Matsuo K, Ono K, Soma Y, Ueyama T, Matoba S, Yamada H, Ikeda K. Aging differentially alters the expression of angiogenic genes in a tissue-dependent manner. Biochem Biophys Res Commun. 2014;446:1243–1249. doi: 10.1016/j.bbrc.2014.03.098. [DOI] [PubMed] [Google Scholar]
- 181.Bae H, Lee JY, Song G, Lim W. Fucosterol suppresses the progression of human ovarian cancer by inducing mitochondrial dysfunction and endoplasmic reticulum stress. Mar Drugs. 2020;18:261. doi: 10.3390/md18050261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Liang H, Xiao J, Zhou Z, Wu J, Ge F, Li Z, Zhang H, Sun J, Li F, Liu R, Chen C. Hypoxia induces miR-153 through the IRE1alpha-XBP1 pathway to fine tune the HIF1alpha/VEGFA axis in breast cancer angiogenesis. Oncogene. 2018;37:1961–1975. doi: 10.1038/s41388-017-0089-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Madonna R, Giovannelli G, Confalone P, Renna FV, Geng YJ, De Caterina R. High glucose-induced hyperosmolarity contributes to COX-2 expression and angiogenesis: implications for diabetic retinopathy. Cardiovasc Diabetol. 2016;15:18. doi: 10.1186/s12933-016-0342-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Anitha RE, Janani R, Peethambaran D, Baskaran V. Lactucaxanthin protects retinal pigment epithelium from hyperglycemia-regulated hypoxia/ER stress/VEGF pathway mediated angiogenesis in ARPE-19 cell and rat model. Eur J Pharmacol. 2021;899:174014. doi: 10.1016/j.ejphar.2021.174014. [DOI] [PubMed] [Google Scholar]
- 185.Zhang SX, Ma JH, Bhatta M, Fliesler SJ, Wang JJ. The unfolded protein response in retinal vascular diseases: implications and therapeutic potential beyond protein folding. Prog Retin Eye Res. 2015;45:111–131. doi: 10.1016/j.preteyeres.2014.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Wang Y, Wang L, Guo H, Peng Y, Nie D, Mo J, Ye L. Knockdown of MALAT1 attenuates high-glucose-induced angiogenesis and inflammation via endoplasmic reticulum stress in human retinal vascular endothelial cells. Biomed Pharmacother. 2020;124:109699. doi: 10.1016/j.biopha.2019.109699. [DOI] [PubMed] [Google Scholar]
- 187.Chen Y, Wang JJ, Li J, Hosoya KI, Ratan R, Townes T, Zhang SX. Activating transcription factor 4 mediates hyperglycaemia-induced endothelial inflammation and retinal vascular leakage through activation of STAT3 in a mouse model of type 1 diabetes. Diabetologia. 2012;55:2533–2545. doi: 10.1007/s00125-012-2594-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Bhatta M, Ma JH, Wang JJ, Sakowski J, Zhang SX. Enhanced endoplasmic reticulum stress in bone marrow angiogenic progenitor cells in a mouse model of long-term experimental type 2 diabetes. Diabetologia. 2015;58:2181–2190. doi: 10.1007/s00125-015-3643-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Lai DW, Lin KH, Sheu WH, Lee MR, Chen CY, Lee WJ, Hung YW, Shen CC, Chung TJ, Liu SH, Sheu ML. TPL2 (therapeutic targeting tumor progression locus-2)/ATF4 (activating transcription factor-4)/SDF1alpha (chemokine stromal cell-derived factor-alpha) axis suppresses diabetic retinopathy. Circ Res. 2017;121:e37–e52. doi: 10.1161/CIRCRESAHA.117.311066. [DOI] [PubMed] [Google Scholar]
- 190.El Zaoui I, Behar-Cohen F, Torriglia A. Glucocorticoids exert direct toxicity on microvasculature: analysis of cell death mechanisms. Toxicol Sci. 2015;143:441–453. doi: 10.1093/toxsci/kfu243. [DOI] [PubMed] [Google Scholar]
- 191.Yu QS, Guo WS, Cheng LM, Lu YF, Shen JY, Li P. Glucocorticoids significantly influence the transcriptome of bone microvascular endothelial cells of human femoral head. Chin Med J (Engl) 2015;128:1956–1963. doi: 10.4103/0366-6999.160564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Nie X, Tang W, Zhang Z, Yang C, Qian L, Xie X, Qiang E, Zhao J, Zhao W, Xiao L, Wang N. Procyanidin B2 mitigates endothelial endoplasmic reticulum stress through a PPARdelta-dependent mechanism. Redox Biol. 2020;37:101728. doi: 10.1016/j.redox.2020.101728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Bhatta M, Chatpar K, Hu Z, Wang JJ, Zhang SX. Reduction of endoplasmic reticulum stress improves angiogenic progenitor cell function in a mouse model of type 1 diabetes. Cell Death Dis. 2018;9:467. doi: 10.1038/s41419-018-0501-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Amin A, Choi SK, Galan M, Kassan M, Partyka M, Kadowitz P, Henrion D, Trebak M, Belmadani S, Matrougui K. Chronic inhibition of endoplasmic reticulum stress and inflammation prevents ischaemia-induced vascular pathology in type II diabetic mice. J Pathol. 2012;227:165–174. doi: 10.1002/path.3960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Wang JM, Qiu Y, Yang ZQ, Li L, Zhang K. Inositol-requiring enzyme 1 facilitates diabetic wound healing through modulating MicroRNAs. Diabetes. 2017;66:177–192. doi: 10.2337/db16-0052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Bronner DN, Abuaita BH, Chen X, Fitzgerald KA, Nuñez G, He Y, Yin XM, O'Riordan MX. Endoplasmic reticulum stress activates the inflammasome via NLRP3- and caspase-2-driven mitochondrial damage. Immunity. 2015;43:451–462. doi: 10.1016/j.immuni.2015.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Wang Y, Gao S, Gao S, Li N, Xie B, Shen X. Blocking the interaction between interleukin-17A and endoplasmic reticulum stress in macrophage attenuates retinal neovascularization in oxygen-induced retinopathy. Cell Biosci. 2021;11:82. doi: 10.1186/s13578-021-00593-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Ozge G, Karaca U, Savran M, Usta G, Gulle K, Sevimli M, Cankara FN, Asci H. Salubrinal ameliorates inflammation and neovascularization via the caspase 3/Enos signaling in an alkaline-induced rat corneal neovascularization model. Medicina (Kaunas) 2023;59:323. doi: 10.3390/medicina59020323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Chen L, Wang Y, Li S, Zuo B, Zhang X, Wang F, Sun D. Exosomes derived from GDNF-modified human adipose mesenchymal stem cells ameliorate peritubular capillary loss in tubulointerstitial fibrosis by activating the SIRT1/eNOS signaling pathway. Theranostics. 2020;10:9425–9442. doi: 10.7150/thno.43315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Huang P, Wang L, Li Q, Tian X, Xu J, Xu J, Xiong Y, Chen G, Qian H, Jin C, Yu Y, Cheng K, Qian L, Yang Y. Atorvastatin enhances the therapeutic efficacy of mesenchymal stem cells-derived exosomes in acute myocardial infarction via up-regulating long non-coding RNA H19. Cardiovasc Res. 2020;116:353–367. doi: 10.1093/cvr/cvz139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Liao Z, Chen Y, Duan C, Zhu K, Huang R, Zhao H, Hintze M, Pu Q, Yuan Z, Lv L, Chen H, Lai B, Feng S, Qi X, Cai D. Cardiac telocytes inhibit cardiac microvascular endothelial cell apoptosis through exosomal miRNA-21-5p-targeted cdip1 silencing to improve angiogenesis following myocardial infarction. Theranostics. 2021;11:268–291. doi: 10.7150/thno.47021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Olejarz W, Kubiak-Tomaszewska G, Chrzanowska A, Lorenc T. Exosomes in angiogenesis and anti-angiogenic therapy in cancers. Int J Mol Sci. 2020;21:5840. doi: 10.3390/ijms21165840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Zeng Z, Li Y, Pan Y, Lan X, Song F, Sun J, Zhou K, Liu X, Ren X, Wang F, Hu J, Zhu X, Yang W, Liao W, Li G, Ding Y, Liang L. Cancer-derived exosomal miR-25-3p promotes pre-metastatic niche formation by inducing vascular permeability and angiogenesis. Nat Commun. 2018;9:5395. doi: 10.1038/s41467-018-07810-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Xiong L, Chen L, Wu L, He W, Chen D, Peng Z, Li J, Zhu X, Su L, Li Y, Gong Y, Xiao H. Lipotoxicity-induced circGlis3 impairs beta cell function and is transmitted by exosomes to promote islet endothelial cell dysfunction. Diabetologia. 2022;65:188–205. doi: 10.1007/s00125-021-05591-4. [DOI] [PubMed] [Google Scholar]
- 205.Dong X, Lei Y, Yu Z, Wang T, Liu Y, Han G, Zhang X, Li Y, Song Y, Xu H, Du M, Yin H, Wang X, Yan H. Exosome-mediated delivery of an anti-angiogenic peptide inhibits pathological retinal angiogenesis. Theranostics. 2021;11:5107–5126. doi: 10.7150/thno.54755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Xiong Y, Chen L, Yan C, Zhou W, Endo Y, Liu J, Hu L, Hu Y, Mi B, Liu G. Circulating Exosomal miR-20b-5p inhibition restores Wnt9b signaling and reverses diabetes-associated impaired wound healing. Small. 2020;16:e1904044. doi: 10.1002/smll.201904044. [DOI] [PubMed] [Google Scholar]
- 207.Wang Z, Jiao P, Zhong Y, Ji H, Zhang Y, Song H, Du H, Ding X, Wu H. The endoplasmic reticulum-stressed head and neck squamous cell carcinoma cells induced exosomal miR-424-5p inhibits angiogenesis and migration of humanumbilical vein endothelial cells through LAMC1-mediated Wnt/beta-catenin signaling pathway. Cell Transplant. 2022;31:9636897221083549. doi: 10.1177/09636897221083549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Lin Y, Zhang C, Xiang P, Shen J, Sun W, Yu H. Exosomes derived from HeLa cells break down vascular integrity by triggering endoplasmic reticulum stress in endothelial cells. J Extracell Vesicles. 2020;9:1722385. doi: 10.1080/20013078.2020.1722385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Nazari-Shafti TZ, Neuber S, Garcia Duran A, Xu Z, Beltsios E, Seifert M, Falk V, Stamm C. Human mesenchymal stromal cells and derived extracellular vesicles: Translational strategies to increase their proangiogenic potential for the treatment of cardiovascular disease. Stem Cells Transl Med. 2020;9:1558–1569. doi: 10.1002/sctm.19-0432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Lin D, Wu H, Zhou Z, Tao Z, Jia T, Gao W. Ginkgolide B improves multiterritory perforator flap survival by inhibiting endoplasmic reticulum stress and oxidative stress. J Invest Surg. 2021;34:610–616. doi: 10.1080/08941939.2019.1676483. [DOI] [PubMed] [Google Scholar]
- 211.Li MT, Ke J, Guo SF, Wu Y, Bian YF, Shan LL, Liu QY, Huo YJ, Guo C, Liu MY, Liu YJ, Han Y. The protective effect of quercetin on endothelial cells injured by hypoxia and reoxygenation. Front Pharmacol. 2021;12:732874. doi: 10.3389/fphar.2021.732874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Haas MJ, Jafri M, Wehmeier KR, Onstead-Haas LM, Mooradian AD. Inhibition of endoplasmic reticulum stress and oxidative stress by vitamin D in endothelial cells. Free Radic Biol Med. 2016;99:1–10. doi: 10.1016/j.freeradbiomed.2016.07.020. [DOI] [PubMed] [Google Scholar]
- 213.Wang G, Nie JH, Bao Y, Yang X. Sulforaphane rescues ethanol-suppressed angiogenesis through oxidative and endoplasmic reticulum stress in chick embryos. J Agric Food Chem. 2018;66:9522–9533. doi: 10.1021/acs.jafc.8b02949. [DOI] [PubMed] [Google Scholar]
- 214.Shangguan WJ, Zhang YH, Li ZC, Tang LM, Shao J, Li H. Naringin inhibits vascular endothelial cell apoptosis via endoplasmic reticulum stress and mitochondrialmediated pathways and promotes intraosseous angiogenesis in ovariectomized rats. Int J Mol Med. 2017;40:1741–1749. doi: 10.3892/ijmm.2017.3160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Jheng JR, Chen YS, Ao UI, Chan DC, Huang JW, Hung KY, Tarng DC, Chiang CK. The double-edged sword of endoplasmic reticulum stress in uremic sarcopenia through myogenesis perturbation. J Cachexia Sarcopenia Muscle. 2018;9:570–584. doi: 10.1002/jcsm.12288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Dalla Bella E, Bersano E, Antonini G, Borghero G, Capasso M, Caponnetto C, Chiò A, Corbo M, Filosto M, Giannini F, Spataro R, Lunetta C, Mandrioli J, Messina S, Monsurrò MR, Mora G, Riva N, Rizzi R, Siciliano G, Silani V, Simone I, Sorarù G, Tugnoli V, Verriello L, Volanti P, Furlan R, Nolan JM, Abgueguen E, Tramacere I, Lauria G. The unfolded protein response in amyotrophic later sclerosis: results of a phase 2 trial. Brain. 2021;144:2635–2647. doi: 10.1093/brain/awab167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Zhang W, Wang M, Gao K, Zhong X, Xie Y, Dai L, Liu W, Liu Y, He X, Li S, Madhusudhan T, Wang H, Zeng H. Pharmacologic IRE1alpha kinase inhibition alleviates aortic dissection by decreasing vascular smooth muscle cells apoptosis. Int J Biol Sci. 2022;18:1053–1064. doi: 10.7150/ijbs.63593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Tang C, Hu Y, Gao J, Jiang J, Shi S, Wang J, Geng Q, Liang X, Chai X. Dexmedetomidine pretreatment attenuates myocardial ischemia reperfusion induced acute kidney injury and endoplasmic reticulum stress in human and rat. Life Sci. 2020;257:118004. doi: 10.1016/j.lfs.2020.118004. [DOI] [PubMed] [Google Scholar]
- 219.Szokalska A, Makowski M, Nowis D, Wilczynski GM, Kujawa M, Wojcik C, Mlynarczuk-Bialy I, Salwa P, Bil J, Janowska S, Agostinis P, Verfaillie T, Bugajski M, Gietka J, Issat T, Glodkowska E, Mrowka P, Stoklosa T, Hamblin MR, Mroz P, Jakobisiak M, Golab J. Proteasome inhibition potentiates antitumor effects of photodynamic therapy in mice through induction of endoplasmic reticulum stress and unfolded protein response. Cancer Res. 2009;69:4235–4243. doi: 10.1158/0008-5472.CAN-08-3439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Chen W, Zou P, Zhao Z, Chen X, Fan X, Vinothkumar R, Cui R, Wu F, Zhang Q, Liang G, Ji J. Synergistic antitumor activity of rapamycin and EF24 via increasing ROS for the treatment of gastric cancer. Redox Biol. 2016;10:78–89. doi: 10.1016/j.redox.2016.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Chiu HW, Tseng YC, Hsu YH, Lin YF, Foo NP, Guo HR, Wang YJ. Arsenic trioxide induces programmed cell death through stimulation of ER stress and inhibition of the ubiquitin-proteasome system in human sarcoma cells. Cancer Lett. 2015;356:762–772. doi: 10.1016/j.canlet.2014.10.025. [DOI] [PubMed] [Google Scholar]
- 222.Gao L, Yuan P, Wei Y, Fu Y, Hou Y, Li P, Chen Y, Ruan Y, Zhou N, Zheng X, Feng W. Total flavonoids of Selaginella tamariscina (P. Beauv.) Spring ameliorates doxorubicin-induced cardiotoxicity by modulating mitochondrial dysfunction and endoplasmic reticulum stress via activating MFN2/PERK. Phytomedicine. 2022;100:154065. doi: 10.1016/j.phymed.2022.154065. [DOI] [PubMed] [Google Scholar]
- 223.Wang F, Han L. Upregulation of serum and glucocorticoid-regulated kinase 1 (SGK1) ameliorates doxorubicin-induced cardiotoxic injury, apoptosis, inflammation and oxidative stress by suppressing glucose regulated protein 78 (GRP78)-mediated endoplasmic reticulum stress. Bioengineered. 2022;13:844–855. doi: 10.1080/21655979.2021.2013109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Varone E, Decio A, Barbera MC, Bolis M, Di Rito L, Pisati F, Giavazzi R, Zito E. Endoplasmic reticulum oxidoreductin 1-alpha deficiency and activation of protein translation synergistically impair breast tumour resilience. Br J Pharmacol. 2022;179:5180–5195. doi: 10.1111/bph.15927. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.