

Summary
The properties of dorsal root ganglia (DRG) neurons that innervate the distal colon are poorly defined, hindering our understanding of their roles in normal physiology and gastrointestinal disease. Here, we report genetically defined subsets of colon innervating DRG neurons with diverse morphologic and physiologic properties. Four colon innervating DRG neuron populations are mechanosensitive and exhibit distinct force thresholds to colon distension. The highest threshold population, selectively labeled using Bmpr1b genetic tools, is necessary and sufficient for behavioral responses to high colon distension, which is partly mediated by the mechanosensory ion channel Piezo2. This Aδ-HTMR population mediates behavioral over-reactivity to colon distension caused by inflammation in a model of inflammatory bowel disease. Thus, like cutaneous DRG mechanoreceptor populations, colon innervating mechanoreceptors exhibit distinct anatomical and physiological properties and tile force threshold space, and genetically defined colon innervating HTMRs mediate pathophysiological responses to colon distension revealing a target population for therapeutic intervention.
Graphical Abstract
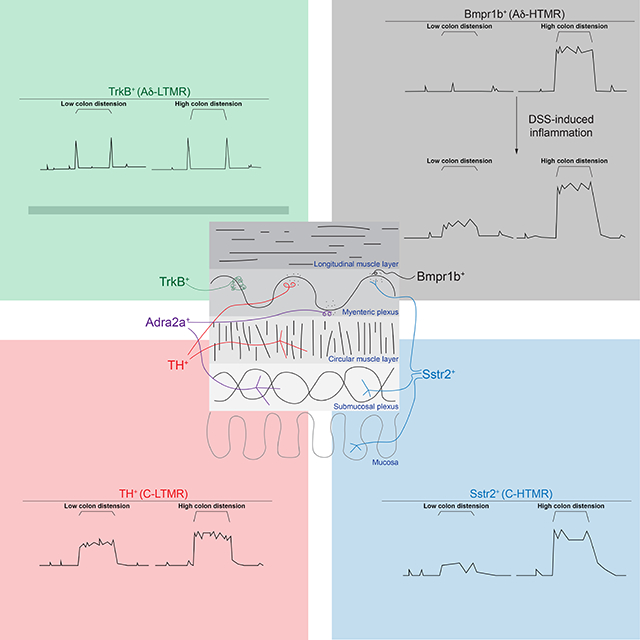
In Brief:
Distinct subtypes of colon innervating mechanoreceptors exhibit distinct anatomical and physiological properties and tile force threshold space. The highest threshold subtype is hyper-reactive to colonic distension during inflammation.
Introduction
Colon mechanosensation is critical for water resorption, motility through the GI tract, defecation, and, under pathologic distension, pain perception1–3. Sensory neurons extrinsic to the gut that innervate the distal colon and mediate these responses are those with cell bodies within the dorsal root ganglia (DRG), the DRG afferents. In addition to innervating internal organs, DRG afferents provide sensory innervation to the skin, where they have been extensively studied4. Low threshold mechanoreceptors (LTMRs) respond to gentle, innocuous mechanical forces5–10, whereas high threshold mechanoreceptors (HTMRs) start to respond at higher mechanical forces, and sometimes, although not always, encode force into the noxious range11,12. Cutaneous DRG afferents can also be subdivided by conduction velocity, rate of adaption to sustained skin indentation, morphology, and genetic diversity, which has been used to create mouse genetic tools to label and manipulate the subtypes5–10,12–15.
Colon innervating DRG afferents have been categorized based on ex vivo physiologic recordings, or anatomical or genetic analysis16–29. Specifically, colonic DRG afferents have been defined by their response to probing at low or high mechanical forces, mucosal stroking, and circumferential wall stretch19,30, and their conduction velocities are in the Aδ or C-fiber range3,19,31–33. Moreover, a range of morphologically distinct colon innervating DRG neuron types have been identified through anterograde tracing25. These include endings that wrap around myenteric cell bodies (intraganglionic varicose endings (IGVE)), endings that branch into the circular or longitudinal muscle layers (intramuscular arrays (IMA)), and endings that terminate within the submucosa, extend through the mucosa, or associate with blood vessels25. A large portion of colon innervating DRG neurons express calcitonin gene related peptide alpha (CGRP), encoded by Calca24,25, and have thus been classified as peptidergic.
Peptidergic DRG afferents that innervate the skin have either Aδ- or C-fibers34,35 and can respond to capsaicin, mustard oil, pruritogens, and others chemicals36 and/or high force mechanical stimuli11,37. Responding to noxious stimuli, including supraphysiologic distension and inflammation, is an important and clinically relevant role of colonic DRG afferents. Intracolonic inflammation leads to both behavioral38 and physiologic hypersensitivity to colonic distension38–40, but the identity of DRG afferent subtypes that mediate this response is unknown.
Single cell sequencing has revealed a large transcriptional heterogeneity within the DRG, with at least fifteen transcriptionally distinct sensory neuron subtypes13,27,28,41–43, however the identity and functions of the DRG neuron subtypes that innervate the colon are unclear. Here, we used mouse molecular genetic approaches, morphological analyses, functional imaging, electrophysiology, and behavior to determine the properties and functional diversity of colon innervating DRG afferents and their roles in physiologic and pathophysiologic mechanosensation.
Results
Colon innervating DRG afferents have distinct morphologic and physiologic properties
Key to understanding sensation of the colon in both physiologic and pathologic states is appreciating the morphological and physiological properties of colon-innervating sensory neuron subtypes and the functions they subserve. In addition to DRG afferents, which provide extrinsic sensory innervation mainly to the distal gut, the GI tract has an extensive intrinsic system, the enteric nervous system (ENS), as well as additional extrinsic sensory and pre-ganglionic parasympathetic innervation provided by the vagus nerve, mainly to the upper GI tract, pre-ganglionic parasympathetic (splanchnic) innervation to the lower GI tract, and sympathetic nervous system (SNS) effector neurons terminating throughout the GI tract44. To establish mouse genetic tools for interrogating DRG sensory neuron innervation of the distal GI tract, we characterized labeling of peripheral nervous system (PNS) subdivisions in mouse lines commonly used to manipulate DRG afferents, such as those in which a recombinase is expressed under control of the Advillin45,46 and PIRT genes47.
We first attempted to clarify whether Advillin expression is time dependent in different parts of the PNS, perhaps allowing for DRG labeling while excluding other PNS populations by inducing recombinase expression at distinct development time points. 4-hydroxytamoxifen (4-HT) treatment of Advillin-CreER; R26LSL-tdTomato mice promoted Cre-mediated tdTomato reporter expression in DRG and SNS neuronal populations regardless of the age 4-HT was given, and labeled the ENS when given at P5 but excluded the ENS if 4-HT was given at earlier time points (Figure S1A–H, P–Q). ENS labeling was defined by tdTomato+ myenteric cell bodies, while SNS labeling was identified by tdTomato+ cell bodies in sympathetic ganglia. In the skin and colon, control staining with anti-beta tubulin III (TUJ1) labeled all neurons (extrinsic and intrinsic), TH staining labeled all sympathetic endings and a subset of TH+ DRG afferents, and CGRP staining labeled both CGRPα+ DRG afferents and CGRPβ+ ENS neurons, as CGRP antibodies cannot distinguish between the two isoforms. Intersecting Flp recombinase expressed from the Advillin gene locus (AdvillinFlp) with Cre recombinase expressed in cells caudal to cervical level 2 (Cdx2-Cre) and a dual-recombinase tdTomato line (R26FSF-LSL-tdTomato; Ai65) labeled both the DRG and SNS and excluded the ENS48 (Figure S1I–J, P–Q). PIRTCre, another commonly used DRG line, also robustly labeled the ENS (Figure S1K, P).
Since Advillin expression (regardless of developmental time point or when intersected with Cdx2-Cre) and PIRTCre did not allow for selective DRG labeling, we developed an alternate strategy using the Phox2B-Flp mouse line49, in which Flp recombinase is expressed in the ENS, SNS, and nodose ganglia, but not in the DRG (Figure S1L–M, P–Q). Utilizing Phox2B-Flp in conjunction with Advillin-CreER for a Cre-on, Flp-off genetic labeling approach (Advillin-CreER; Phox2B-Flp; R26FRT-LSL-ChR2-YFP-FRT) led to YFP expression in cells in which Cre recombinase was expressed under control of the Advillin gene, but not in cells in which Flp recombinase was expressed under the control of the Phox2B gene. Indeed, in these mice YFP expression was observed in virtually all DRG neurons but not in the sympathetic ganglia, ENS neurons, or vagal afferents, as evidenced by lack of labeling in the nucleus tractus solitarius (NTS) (Figure 1A and S1N–O, Q). This strategy confirmed that DRG afferents densely innervate the distal colon: most DRG neurons terminate within the myenteric plexus level, with fewer fibers found within the submucosal plexus level (Figure 1A, S1R). Additionally, this work defined genetic tools for selectively labeling colon innervating DRG neurons, which we subsequently used in conjunction with mouse lines that by themselves labeled other PNS neuronal populations.
Figure 1: DRG afferents with distinct morphological and physiological properties innervate the distal colon and mediate Piezo2-dependent behavioral responses to colon distension.
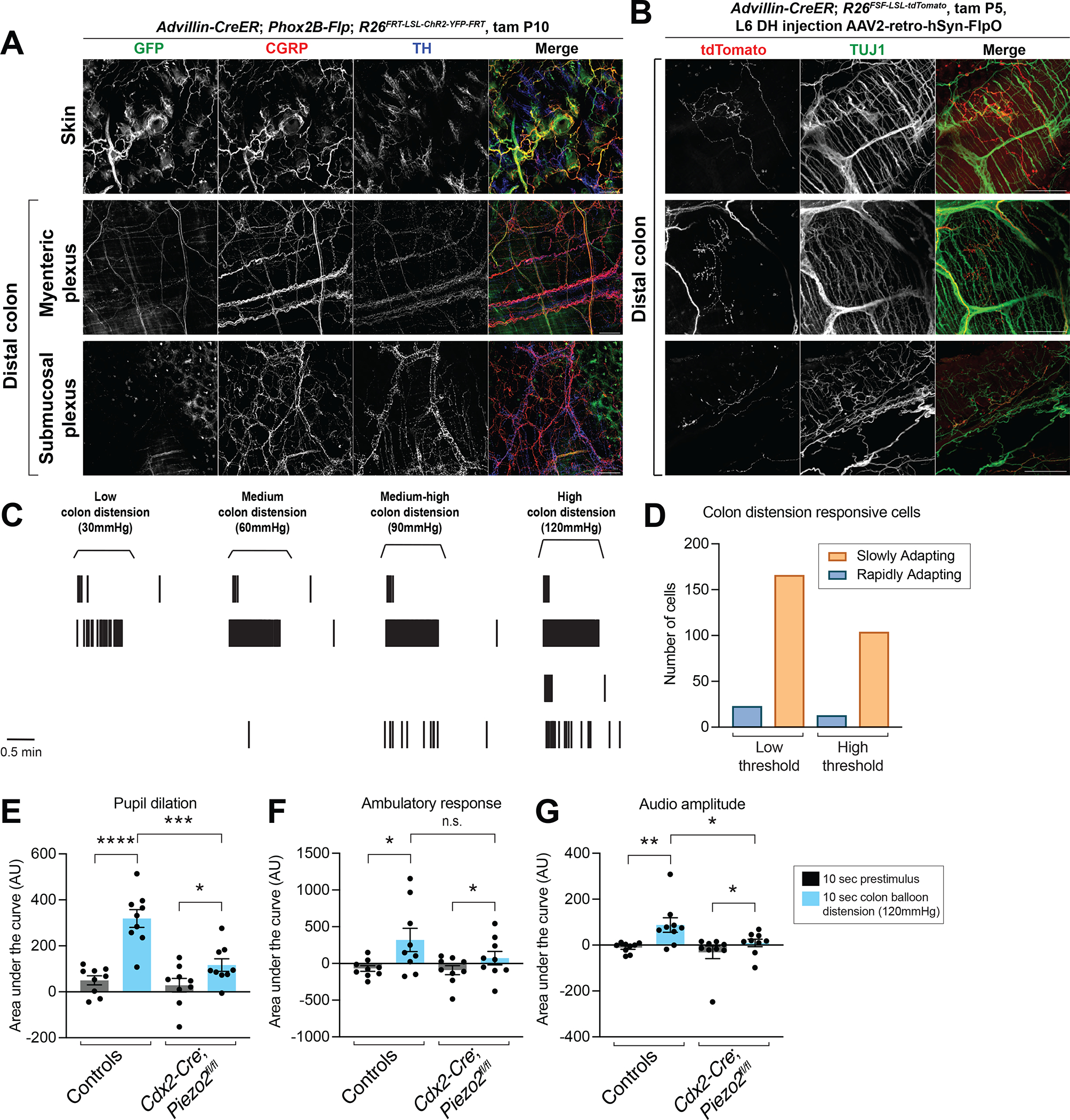
A: Whole mount immunostaining of skin and distal colon from Advillin-CreER; Phox2B-Flp; R26FRT-LSL-ChR2-YFP-FRT mice (1mg tamoxifen at P10; sacrificed at 6 weeks). Note that the GFP antibody labels YFP-expressing cells.
B: AAV2-retro-hSyn-FlpO virus injection into the L6 dorsal horn (DH) of Advillin-CreER; R26FSF-LSL-tdTomato mice to sparsely label DRG afferent terminals in the distal colon (1mg tamoxifen at P5; sacrificed at 6 weeks). All scale bars are 100 μm unless otherwise specified.
C: Example raster plots of single units from L6-S1 DRG multielectrode array (MEA) recordings. The top unit is classified as low-threshold rapidly adapting, followed by a low-threshold slowly adapting unit; the third unit is high-threshold rapidly adapting; the bottom unit is high-threshold slowly adapting.
D: Quantification of the number of units in each category from L6-S1 DRG MEA recordings (data from N = 14 mice, ~4 MEA penetrations per DRG per mouse).
E-G: The behavioral response to intracolonic high-threshold balloon distension in Cdx2-Cre; Piezo2fl/fl animals, measured by pupil dilation (E), movement (F), and vocalizations (G) (N = 9 mice per group; paired and unpaired t test for pupil dilation and accelerometer data; Mann-Whitney and Wilcoxon matched-pairs signed rank test for vocalization data). The following symbols are used in this and subsequent figures for P values: n.s., not significant; *, P < 0.05; **, P < 0.01; ***, P < 0.001; ****, P < 0.0001.
To assess the morphological diversity of DRG afferents that innervate the colon, we injected retro-AAV2-Flp virus into the L6-S1 dorsal horn of Advillin-CreER; R26FSF-LSL-tdTomato mice, such that tdTomato was expressed exclusively and sparsely in DRG afferents. The DRG afferent terminals in the colon exhibited multiple distinct morphologies, including IGVEs, IMAs, and endings that project through the mucosa towards the epithelium (Figure 1B). These ending morphologies reflect those that have been observed previously25.
We also assessed the diversity of physiological response properties of colon innervating DRG afferents. Colon innervating DRG afferents have distinct physiological responses in ex vivo preparations20,24,50–52, but their in vivo physiologic responses to colon distension have not been described. In vivo DRG multielectrode array (MEA) recordings revealed four categories of physiologic responses to colon distension: low-threshold rapidly adapting (RA); high-threshold RA; low-threshold slowly adapting (SA); and high-threshold SA (Figure 1C). The more common response types were SA, while the RA types were less common, although this may be due to a technical limitation of the assay, as RA neurons spike only a few times at the distension onset and/or offset and may therefore be more difficult to distinguish (Figure 1D).
We next assessed the role of Piezo2, the main mechanosensory ion channel in cutaneous DRG afferents53, in colon mechanosensation. To this end, we developed a behavioral assay for colon distension responses in awake animals. When distending an intra-colonic balloon to high force, mice exhibited an increase in pupil dilation, mobility, and vocalizations (Figure 1E–1G). This may reflect a pain response, as a robust increase in pupil dilation, indicating a change in autonomic tone, movement, and vocalizations (including in the ultrasonic range) would be expected with a noxious stimulus. Piezo2 ablation in cells caudal to cervical level 2 using Cdx2-Cre; Piezo2flox/flox mice, including DRG afferents (and colon epithelium48) but not enteric neurons (Figure S1I, P), decreased, but did not completely eliminate, the colon distension behavioral response. These findings indicate that Piezo2 is responsible for most of the behavioral response to high-threshold colon mechanosensation (Figure 1E–1G).
Five genetically-defined subtypes of DRG afferents innervate the distal colon
The findings here and from prior work20,24,25,50–52 that colon innervating DRG afferents exhibit a range of morphologies and physiologic colon distension responses implies an underlying afferent subtype heterogeneity. Therefore, to gain genetic access to colon innervating DRG neuron subtypes, we used tools that label a range of genetically defined DRG neurons, which were developed to explore the properties and functions of cutaneous DRG neuron populations. We focused on tools that label Aδ- and C-fiber DRG neuron subtypes because previous work has established that heavily myelinated neurons with Aβ conduction velocity do not innervate visceral organs3,19,31–33. We used mouse intersectional genetic tools to selectively label MrgD+, TrkB+, and Tyrosine Hydroxylase (TH)+ DRG neurons7,8,54,55, which label cutaneous C-fiber HTMR, Aδ-LTMR, and C-LTMR DRG neuron populations, respectively. To label MrgD+ DRG afferents while excluding the ENS, we employed the Cre-on, Flp-off strategy (MrgprdCreER; Phox2B-Flp; R26FRT-LSL-tdTomato-FRT) and found that tdTomato-labeled MrgD+ neurons do not innervate the colon, stomach, or duodenum (Figure 2A, S2A, O–P). On the other hand, intersecting TrkBCreER and AdvillinFlp mice labeled DRG neurons while excluding the SNS and ENS, and this approach revealed that a TrkB+ DRG neuron population innervates the distal colon, stomach, and duodenum (Figure 2B, K, S2B, O–P), though we cannot fully exclude some contribution of vagal fibers with this intersection in the upper GI tract. To label TH+ DRG neurons while excluding SNS and nodose labeling, we used the Cre-on, Flp-off strategy with THCreER and Phox2B-Flp (THCreER; Phox2B-Flp; R26FRT-LSL-tdTomato-FRT) mice and found that tdTomato+ TH+ DRG afferents innervate the distal colon and stomach (though the innervation to the stomach was considerably sparser than the colon), but not the duodenum (Figure 2C, K, S2C, O–P).
Figure 2: Five transcriptionally distinct DRG afferent subtypes that innervate the colon.
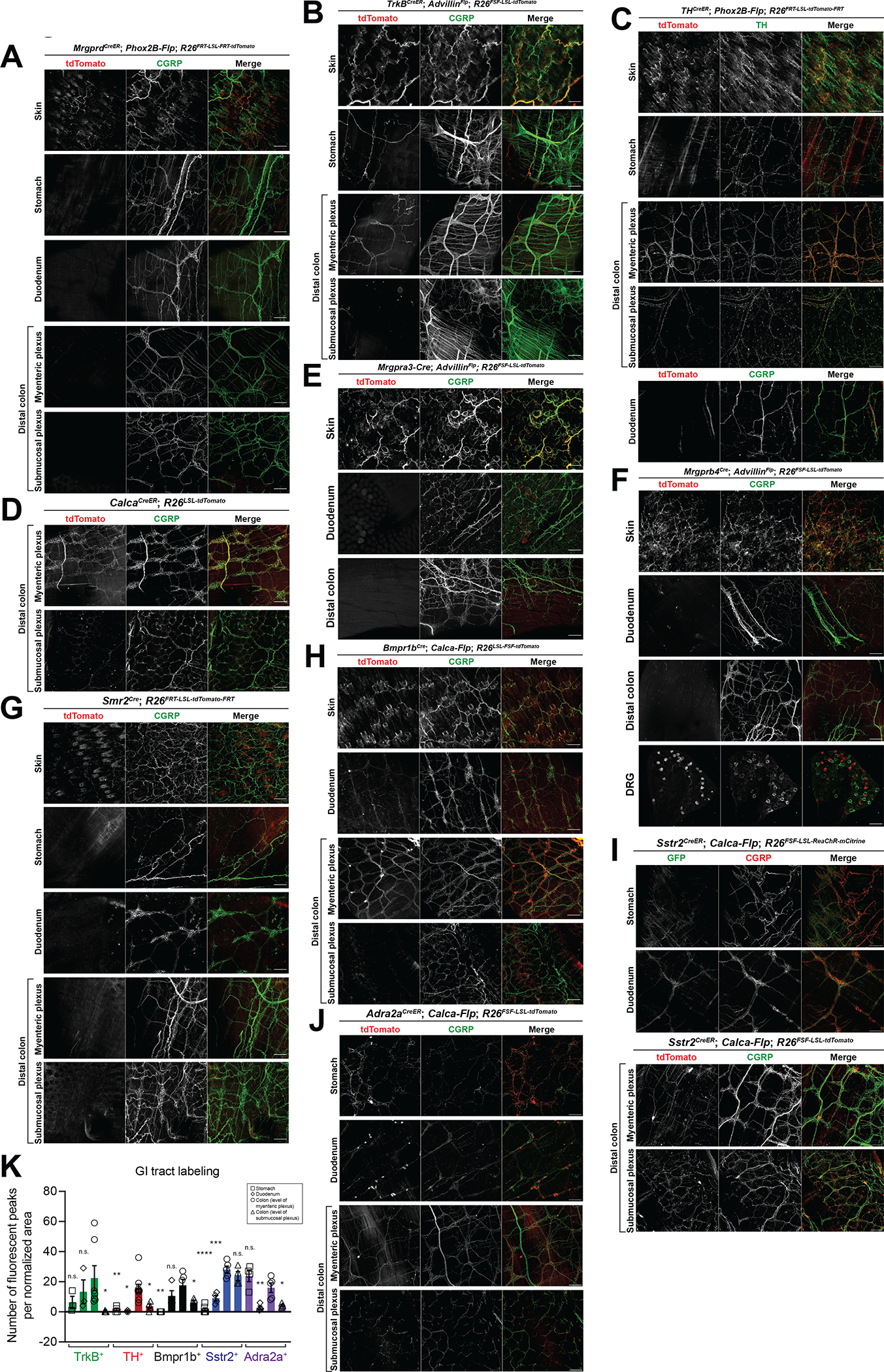
Whole mount immunostaining in skin, stomach, duodenum, distal colon, or DRGs from (A) MrgprdCreER; Phox2B-Flp; R26FRT-LSL-tdTomato-FRT mice (1mg tamoxifen at P7); (B) TrkBCreER; AdvillinFlp; R26FSF-LSL-tdTomato mice (0.5mg tamoxifen at P5); (C) THCreER; Phox2B-Flp; R26FRT-LSL-tdTomato-FRT mice (0.5mg tamoxifen at P23); (D) CalcaCreER; R26LSL-tdTomato mice (1mg tamoxifen at P11); (E) Mrgpra3-Cre; AdvillinFlp; R26FSF-LSL-tdTomato mice; (F) Mrgprb4Cre; AdvillinFlp; R26FSF-LSL-tdTomato mice; (G) Smr2Cre; R26FRT-LSL-tdTomato-FR mice T; (H) Bmpr1bCre; Calca-Flp; R26FSF-LSL-tdTomato mice; (I) Sstr2CreER; Calca-Flp; R26FSF-LSL-ReaChR-mCitrine mice (2mg tamoxifen at P21) (top 2 panels); Sstr2CreER; Calca-Flp; R26FSF-LSL-tdTomato mice (0.5mg tamoxifen at P22) (bottom 2 panels); and (J) Adra2aCreER; Calca-Flp; R26FSF-LSL-tdTomato mice (4mg tamoxifen at P14 and P16). Animals were sacrificed at 3–6 weeks. All scale bars are 100 μm unless otherwise specified.
K: Quantification of density of fibers labeling the stomach, duodenum, or colon (at the level of either the myenteric or submucosal plexus) in the five colon-innervating populations. Quantified from at least 3 images from at least N = 2 animals. Comparisons are stomach, duodenum, or colon (submucosal plexus) each compared to colon (myenteric plexus) using an unpaired t test.
We next focused on CGRP expressing DRG neuron subtypes, because CGRP+ DRG afferents innervate the colon25. Multiple lines were used to characterize the CGRP population, including two BAC transgenics (Calca-Flp and Calca-GFP) and two Calca knock-ins (CalcaCreER and CalcaLox-GFP-STOP-Lox-hDTR). These lines preferentially labeled CGRP+ DRG afferents with no SNS labeling and limited off-target ENS labeling (Figure 2D, S2D–G). Of the CGRP+ subsets of DRG neurons defined in Sharma et al.13, MrgA3+ neurons (and MrgB4+ neurons, which are a subset of the broader MrgA3+ population) and Smr2+ neurons do not innervate the GI tract (Figure 2E–G). Conversely, three subtypes of CGRP+ DRG afferents, defined by Bmpr1b, Sstr2, or Adra2a expression13, did innervate the colon (Figure 2H–K). The Bmpr1b+ population also innervated the duodenum but not the stomach (Figure 2H, K). The Sstr2+ population innervated both the stomach and duodenum, though less densely than the colon (Figure 2I, K). The Adra2a+ population innervated the colon and stomach to similar degrees, with less innervation to the duodenum (Figure 2J–K). For these three CGRP+ DRG neuron subsets, we used Bmpr1bCre 13 and the newly developed Sstr2CreER and Adra2aCreER alleles43 and either intersection with Calca-Flp or the Cre-on, Flp-off strategy with Phox2B-Flp to confirm that the ENS and SNS were mostly excluded (Figure 2H–J, S2H–P). Notably, based on prior nodose ganglia RNA sequencing data sets56, Bmpr1b, Sstr2, and Adra2a are not expressed in the nodose, and thus labeling observed in the GI tract with these lines likely represents DRG but not nodose innervation. It is also noteworthy that most genetically defined DRG neuron subtypes that innervate the colon had, as a population, the same general pattern observed when all DRG afferents were labeled, namely, dense innervation along the myenteric but not submucosal plexus, except the Sstr2+ subtype, which exhibited a similar density of innervation at the level of the myenteric and submucosal plexuses (Figure 2B–C, H–K).
Finally, we performed in situ hybridization in DRGs to quantify the density of overlap between the colon-innervating populations. We found no overlap between the TrkB+, TH+, and CGRP+ populations, and little overlap between the Bmpr1b+, Sstr2+, and Adra2a+ populations (Figure S2Q–S). Thus, the TrkB+, TH+, Sstr2+, Adra2a+, and Bmpr1b+ transcriptionally defined subtypes of DRG neurons have axons that innervate the distal colon, whereas MrgD+, MrgA3+, and Smr2+ DRG neurons do not. Genetic access to five transcriptionally defined DRG subtypes enables physiological, morphological, and functional analysis of these populations.
Genetic subtypes of DRG afferents have distinct morphologies in the distal colon
We and others have found that DRG afferents that innervate the distal colon exhibit a range of morphologies25. We hypothesized that each of the five transcriptionally defined DRG subtypes that innervate the colon exhibits a particular morphology and that collectively these subtypes account for much or all of the full range of DRG neuron morphologies. To test this, we performed sparse genetic labeling using mice harboring TrkBCreER, THCreER, Sstr2CreER, Adra2aCreER alleles and a newly generated Bmpr1bCreER allele (Figure S3A) and treated with a low dose of tamoxifen to label and visualize the endings of individual neurons for each of the five colon innervating DRG neuron subtypes. This analysis revealed that TrkB+ DRG afferents terminate exclusively in the myenteric plexus, with IGVE endings that wrap around ENS neuron cell bodies (individual TrkB+ DRG neurons wrapping around an average of 16 myenteric plexus neuron cell bodies) (Figure 3A, F–G). TH+ DRG neurons, on the other hand, either branched into the myenteric plexus with few myenteric wrappings or into the circular muscle layer, forming previously defined intramuscular array (IMA) endings (Figure 3B, F–G). Intersecting Bmpr1bCreER with Calca-Flp and a dual recombinase fluorescent reporter (R26FSF-LSL-tdTomato) allowed for sparse labeling of Bmpr1b+ DRG neurons (Figure S3B). As with TrkB+ DRG neurons, individual tdTomato labeled Bmpr1b+ neurons formed IGVEs, although with far fewer branches and wrappings around myenteric cell bodies; individual Bmpr1b+ DRG neurons typically wrapped around only 1–2 cell bodies (Figure 3C, F–G). tdTomato labeled Sstr2+ DRG neuron endings terminated in the myenteric plexus, submucosal plexus, or mucosa, in each case by branching but without any myenteric neuron cell body wrappings, consistent with previously described “internodal strands” (Figure 3D, F–G). tdTomato labeled Adra2a+ DRG neuron endings terminated either within the myenteric or submucosal plexuses, with few myenteric neuron cell body wrappings (Figure 3E–G). When comparing across the five different subtypes, TrkB+ endings were the most elaborate, with the longest length, most abundant branches, and most wrappings around myenteric cell bodies (Figure 3G–J). In summary, as observed for skin innervating DRG neurons subtypes43, each of the five transcriptionally distinct, colon innervating DRG neuron subtypes exhibits a unique distal colon termination pattern and morphology, and collectively these colonic endings account for the range of morphologies observed in our random labeling experiments.
Figure 3: Five DRG neuron subtypes with distinct morphologies innervate the distal colon.

A-E: Tracings of whole mount immunofluorescence from (A) TrkBCreER; AdvillinFlp; R26FSF-LSL-tdTomato mice (0.005mg tamoxifen at P5); (B) THCreER; Phox2B-Flp; R26FRT-LSL-tdTomato-FRT mice (0.1mg tamoxifen at P24); (C) Bmpr1bCreER; Calca-Flp; R26FSF-LSL-tdTomato mice (1–3mg tamoxifen at P21); (D) Sstr2CreER; Calca-Flp; R26FSF-LSL-tdTomato mice (0.05mg tamoxifen at P21); and (E) Adra2aCreER; Calca-Flp; R26FSF-LSL-tdTomato mice (1mg tamoxifen P21–25 and 4mg 8 weeks), all treated with low amounts of tamoxifen to achieve sparse labeling. Animals were sacrificed after P21 and at least 2 weeks after last tamoxifen dose. All scale bars are 100 μm unless otherwise specified.
F: Graph summarizing ending termination location in the colon wall for the genetically labeled subtypes.
G: Graph quantifying myenteric cell body wrappings observed with individual IGVEs by genetic subtype. Each point represents an individual ending.
H-J: Graphs quantifying total ending length (H), width (I), or branches (J). Each point represents an individual ending.
K: Diagram showing the colonic morphologies of distinct DRG afferent subtypes.
At least four genetic subtypes of DRG afferents tile mechanical distension force space
We hypothesized that the transcriptionally and morphologically distinct colon innervating DRG neuron subtypes are also physiologically distinct. To determine which, if any, of the colon innervating DRG neuron subtypes respond to colon distension, we used the mouse genetic tools to express a GCaMP reporter selectively in each colon innervating subtype for in vivo L6-S1 DRG calcium imaging experiments (Figure 4A). Four of the five DRG neuron subtypes were assessed in this analysis; due to the sparse nature of labeling with the Adra2aCreER line, we were unable to perform functional experiments with this subtype. The TrkB+ DRG afferents responded to colon distension starting at low forces and in a RA manner (Figure 4B, S4A–B). Interestingly, the same colon responding TrkB+ neurons also often responded to back hairy skin pinch (Figure 4B). To ensure the colon responses were not due to slight movement of the skin during balloon distension of the colon, lidocaine was inserted into the colon. While colon distension responses were eliminated following lidocaine, skin pinch responses remained intact for the same neurons (Figure S4C–E). The TH+ DRG afferents responded to low-medium force colon distension in a SA manner (Figure 4C, S4F–G).
Figure 4: Four DRG neuron subtypes display distinct responses to colon distension and collectively tile force threshold space.
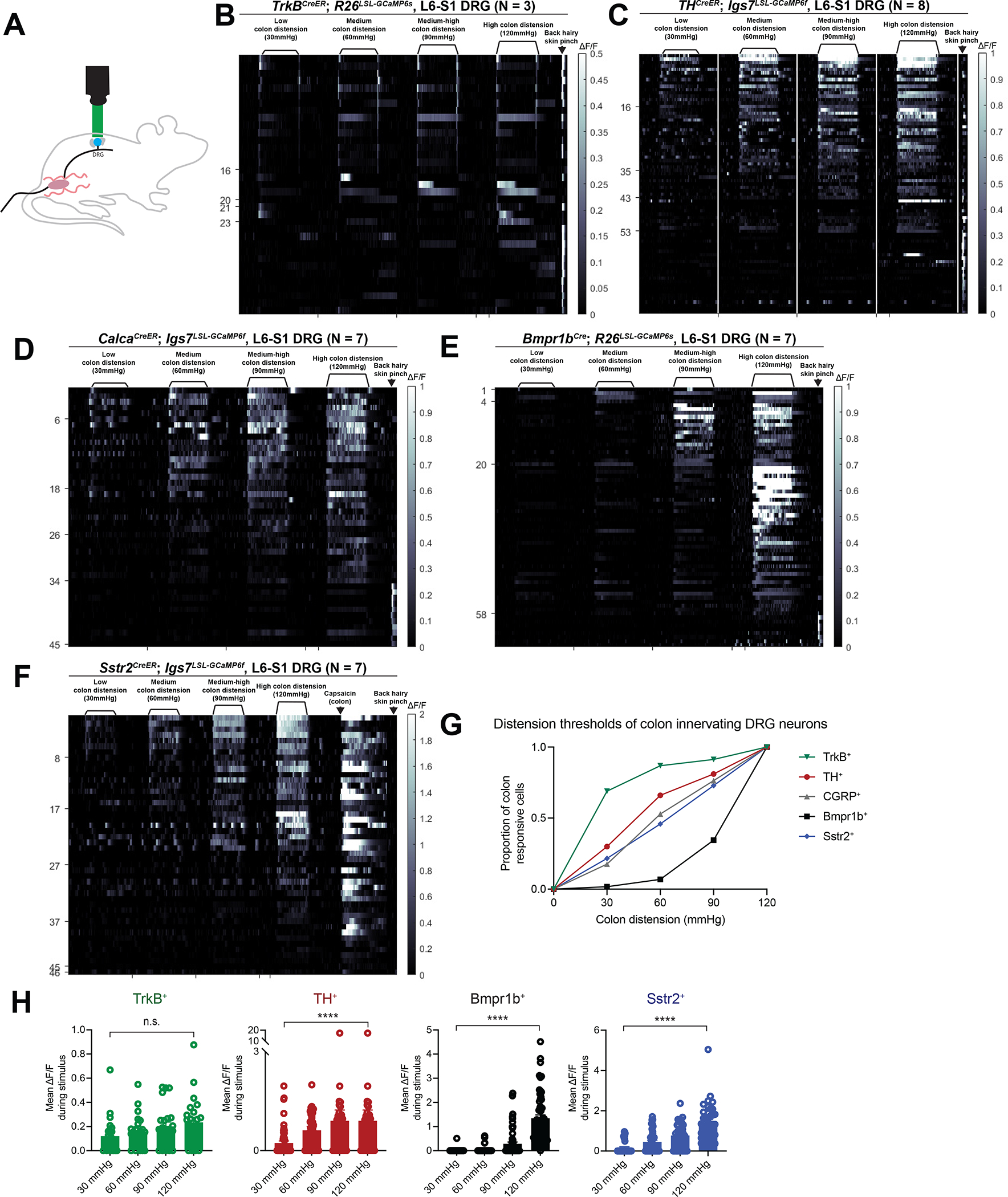
A: A schematic for the DRG calcium imaging set up.
B-F: In vivo DRG calcium imaging responses to colon distension in (B) TrkBCreER; R26LSL-GCaMP6s mice (N = 3; 0.5mg tamoxifen at P5); (C) THCreER; Igs7LSL-GCaMP6f mice (N = 8; 1–2mg tamoxifen at P21); (D) CalcaCreER; Igs7LSL-GCaMP6f mice (N = 7; 0.5 mg tamoxifen at P8–9); (E) Bmpr1bCre; R26LSL-GCaMP6s mice (N = 7); (F) and Sstr2CreER; Igs7LSL-GCaMP6f mice (N = 7; 1mg tamoxifen at P24).
G: Graph depicting the distension thresholds of all colon innervating populations based on calcium imaging.
H: Mean calcium activation (ΔF/F) during the distension stimulus for each cell type from calcium imaging experiments. Analyses were performed using repeated-measures ANOVAs and comparisons between low (30mmHg) and high (120mmHg) forces are plotted. See Table S1 for additional statistical details.
DRG calcium imaging from the entire CGRP+ population indicated that this transcriptionally and morphologically heterogenous population of DRG neurons respond mostly in a SA manner and to higher forces than either the TrkB+ or TH+ populations (Figure 4D, S4H–I). The Bmpr1b+ subtype of CGRP+ neurons also responded in a high-threshold SA manner (Figure 4E, S4J–K), although with a higher threshold than the CGRP population (Figure 4D, E, G). It is notable that the TrkB+, TH+, and Bmpr1b+ populations have force thresholds and physiologic response properties that are relatively comparable to their genetic counterparts in the skin7,8,11,13. Finally, the Sstr2+ CGRP subtype, which expresses TrpV1, responded to medium-high force colon distension (Figure 4F, S4L–M). Consistent with their expression of TrpV1, intracolonic capsaicin activated the Sstr2+ afferents, but not Bmpr1b+ afferents, which do not express appreciable levels of TrpV1 (Figure 4F, S4N–O).
To assess the extent to which these four colon-innervating populations encode force into the higher range, we calculated mean calcium activation (ΔF/F) from the calcium imaging data during the stimulus period. Consistent with their designation as an LTMR, the TrkB+ units did not have significant changes in intracellular calcium levels with increasing forces (Figure 4H). Interestingly, while robustly responding at the lowest threshold, the TH+ units exhibited increased mean calcium signals with increasing forces when comparing 30mmHg to 120mmHg, though notably no significant increase when comparing 60mmHg to 120mmHg (Figure 4H). The Bmpr1b+ and Sstr2+ populations had significantly increased intracellular calcium levels with increasing forces, as would be expected for HTMRs (Figure 4H).
We next expressed an opsin in each distension-sensitive subtype and performed in vivo L6-S1 DRG electrophysiology using MEAs and optotagging with intracolonic light stimulation (Figure 5A–B). Consistent with the GCaMP experiments, all opto-tagged TrkB+ units responded to the lowest colon distension forces and in a RA manner (Figure 5C–E, S5D). The optogenetic latency of this population was approximately 10ms, their waveform was narrow, and their cell body size was large, consistent with A-fiber neurons (Figure 5D, F–G, S5A–C). Also consistent with the GCaMP experiments, all opto-tagged TH+ units responded to the lowest distension forces and in a SA manner (Figure 5C–E, S5D). In comparison to the TrkB+ neurons, TH+ neurons exhibited a longer optogenetic latency, which was in the C-fiber range, a wider action potential waveform, and a small cell body size. Thus, like their cutaneous counterparts, colon innervating TrkB+ neurons can be classified as Aδ-LTMRs, whereas colon innervating TH+ neurons are C-LTMRs (Figure 5D, F–G, S5A–C).
Figure 5: In vivo electrophysiological response properties of four colon innervating DRG neuron populations.
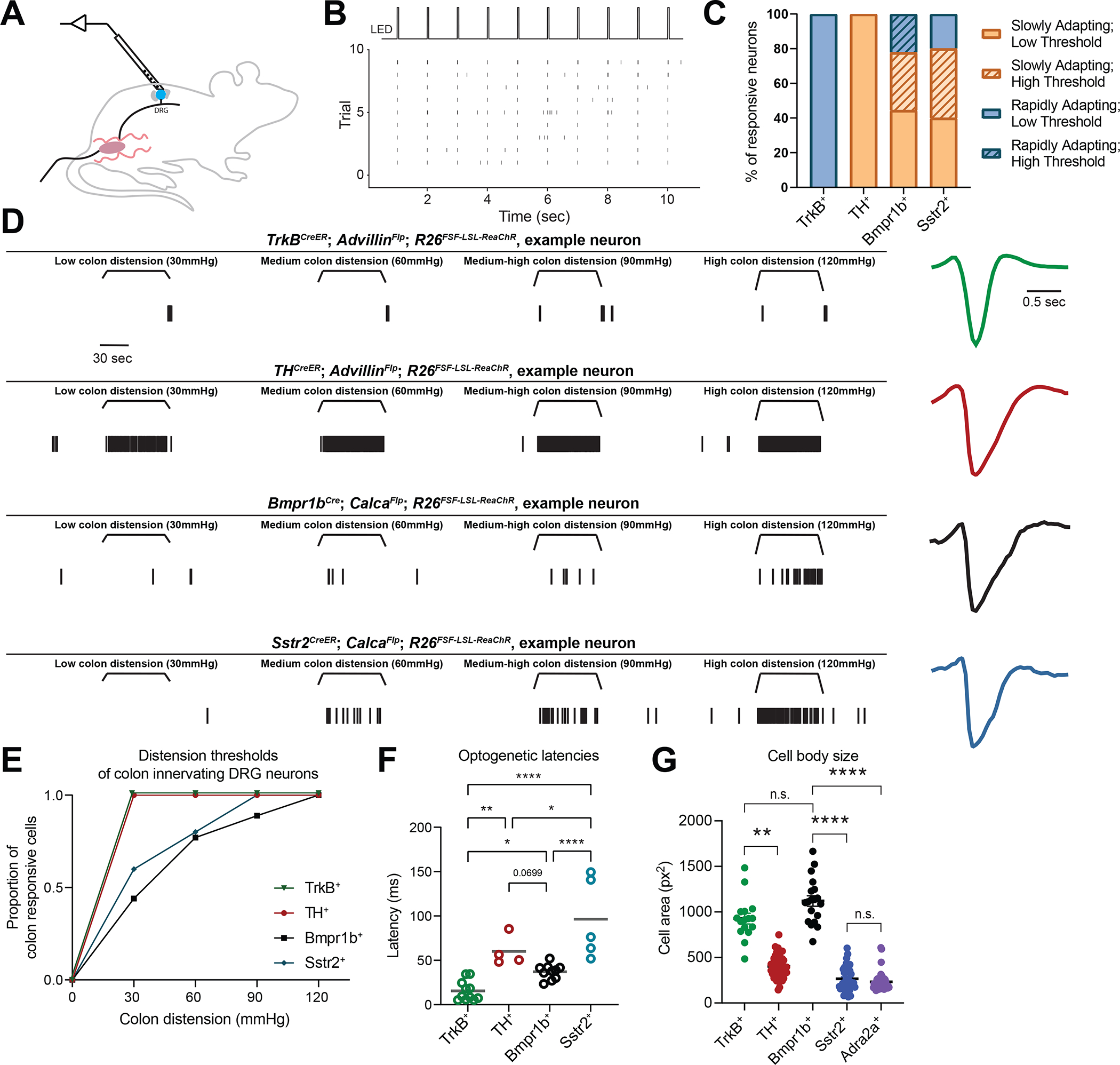
A: A schematic of the DRG multielectrode array (MEA) set up.
B: Example raster plot showing light-evoked responses of an opto-tagged neuron.
C: Percent of neurons with different electrophysiological responses to colon distension in each genetic subtype.
D: In vivo electrophysiological responses of one representative example neuron from each genetic class. Raster plot on the left; normalized extracellular action potential waveform on the right.
E: Graph depicting the distension thresholds of all colon innervating populations based on MEA recordings.
F: Optogenetic latency by genetic subtype. Each point represents one neuron. TrkB+ units in green (n = 7 cells from N = 6 mice); TH+ units in red (n = 4 cells from N = 8 mice); Bmpr1b+ units in black (n = 10 cells from N = 5 mice); and Sstr2+ units in blue (n = 5 cells from N = 5 mice). Comparisons performed using a one-way ANOVA [F(3, 26)= 4.518, p <0.0001] with post-hoc Holm-Šídák’s tests.
G: Cell size quantification of each of the colon-innervating from ISH. TrkB+ (n = 16 cells from N = 3 DRG), TH+ (n = 48 cells from N = 3 DRG), Bmpr1b+ (n = 18 cells from N = 3 DRG), Sstr2+ (n = 41 cells from N = 3 DRG), and TH+ (n = 42 cells from N = 3 DRG). Analyses were performed using a one-way ANOVA and relevant comparisons are plotted. See Table S1 for additional statistical details.
Analogous to the GCaMP measurements, many opto-tagged Bmpr1b+ units started to respond to colon distension at higher forces and all responded in a SA manner (Figure 5C–E, S5D). Interestingly, some Bmpr1b+ units responded to lower thresholds, perhaps suggesting heterogeneity in this subtype. This population had an optogenetic latency that was shorter than the TH+ C-LTMRs but longer than the TrkB+ Aδ-LTMRs, although with a wide waveform (Figure 5D, F, S5A–C), supporting an Aδ designation, like their cutaneous counterparts. Consistent with this, Bmpr1b+ cell bodies were considerably larger than the C fiber populations (TH+, Sstr2+, and Adra2a+) and not significantly different from the TrkB+ population, further supporting that colon-innervating Bmpr1b+ afferents are Aδ-HTMRs (Figure 5G). MEA recordings also confirmed that many Sstr2+ neurons begin to respond at higher thresholds, and all responded in a SA manner with long optogenetic latencies and wide action potential (AP) waveforms, which, together with the small cell body size, indicate that these are C-fiber neurons (C-HTMRs) (Figure 5C–G, S5A–D). Notably, both HTMR populations responded to lower distension forces in the MEA recordings compared with the calcium imaging experiments, which is likely due to the higher sensitivity of physiologic recordings. To determine the extent to which these four populations continue to encode distension forces into high force range, we performed a firing rate analysis of the MEA recording data. TrkB+ units did not appreciably increase their firing rate with increasing forces, consistent with their function as an LTMR (Figure S5E). As observed in the calcium imaging experiments, TH+ units appeared to increase their firing rates with increasing forces, though due to the small sample size statistical analyses were not performed on the firing rates from this population (Figure S5E). Conversely, the Bmpr1b+ and Sstr2+ populations exhibited increased firing rates at higher forces, indicating that these populations encode into the higher force range, in line with their function as HTMRs, with Bmpr1b+ neurons displaying the highest total firing rates (Figure S5E). Thus, while colon innervating TrkB+ neurons are Aδ-LTMRs and TH+ neurons are C-LTMRs, Bmpr1b+ and Sstr2+ neurons are Aδ-HTMR and C-HTMR populations, respectively. Moreover, these four morphologically and physiologically distinct colon mechanoreceptor subtypes tile mechanosensory force threshold space, suggesting a population code to explain behavioral and perceptual responses to colon distension (Figure 4G, 5E).
Bmpr1b+ Aδ-HTMRs are necessary and sufficient for sensing high force colon distension
We next asked whether selective activation of any or all genetic subpopulations of DRG afferents is sufficient to mediate a behavioral response. We inserted an LED probe into the colon of mice expressing an opsin, either Channelrhodopsin-2 (ChR2) or a red-shifted variant of Channelrhodopsin (ReaChR), in each of the four physiologically defined subtypes and recorded pupil dilation (indicative of a change in autonomic tone), ambulatory movement, and vocalizations in response to a light stimulus. In control experiments, light in the colon of wild-type animals did not lead to an appreciable response, while light on the skin of the left hindpaw led to a small amount of pupil constriction, presumably due to ambient light from the LED probe directly stimulating the retina, but no change in movement or vocalizations (Figure 6A–C and S6A–C).
Figure 6: The Bmpr1b+ Aδ-HTMR population is necessary and sufficient to drive a behavioral response to colon stimulation.
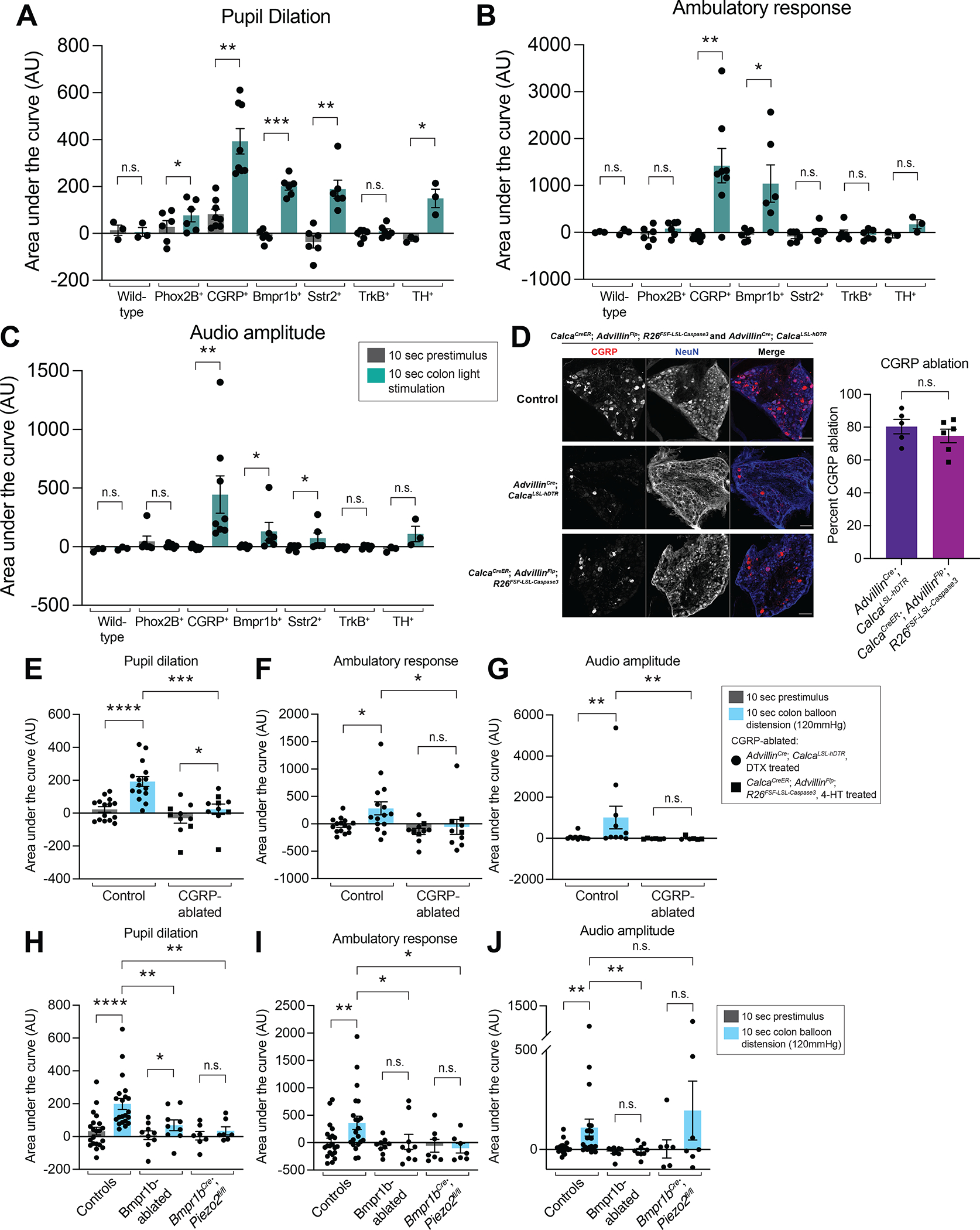
A-C: Behavioral responses (pupil dilation (A), movement (B), and vocalizations (C)) to colonic optogenetic stimulation of wild-type mice (N = 3), Phox2B-Flp; R26FSF-mCitrine-ReaChR mice (N = 6), Calca-Flp; R26FSF-ReaChR or CalcaCreER; R26FRT-LSL-ChR2-YFP-FRT (1mg tamoxifen at P12) mice (N = 8), Bmpr1bCre; Calca-Flp; R26FSF-LSL-mCitrine-ReaChR mice (N = 6), Sstr2CreER; Calca-Flp; R26FSF-LSL-mCitrine-ReaChR or Sstr2CreER; R26LSL-mCitrine-ReaChR mice (N = 6; 2mg tamoxifen at P21), TrkBCreER; AdvillinFlp; R26FSF-LSL-mCitrine-ReaChR mice (N = 6; 0.5mg tamoxifen at P5), and THCreER; Phox2B-Flp; R26FRT-LSL-ChR2-YFP-FRT mice (N = 3; 1mg tamoxifen at P9). All comparisons are paired t tests except for the vocalization data for the Phox2B-Flp; R26FSF-mCitrine-ReaChR mice, Calca-Flp; R26FSF-ReaChR or CalcaCreER; R26FRT-LSL-ChR2-YFP-FRT mice, Bmpr1bCre; Calca-Flp; R26FSF-LSL-mCitrine-ReaChR mice, and Sstr2CreER; Calca-Flp; R26FSF-LSL-mCitrine-ReaChR or Sstr2CreER; R26LSL-mCitrine-ReaChR mice, which were analyzed using the Wilcoxon matched-pairs signed rank test.
D: DRG whole mount immunofluoresence in control and CGRP-ablated animals (either AdvillinCre; CalcaLSL-hDTR, DTX treated or AdvillinFlp; CalcaCreER; R26FSF-LSL-Caspase3, 2mg tam daily P21-P23). Quantification of the percent CGRP ablation with the two methods shown in the right panel (N = 5 animals for AdvillinCre; CalcaLSL-hDTR, DTX treated and N = 6 animals for AdvillinFlp; CalcaCreER; R26FSF-LSL-Caspase3, 2mg tam daily P21-P23, unpaired t test for comparison).
E-G: Behavioral responses (pupil dilation (E), movement (F), and vocalizations (G)) to high force colon balloon distension in CGRP-ablated animals, compared to controls. Each point represents the average of data from an individual animal. (N = 15 for controls for pupil dilation and accelerometer and 10 for vocalization; N = 10 for CGRP-ablated (6 of which are AdvillinCre; CalcaLSL-hDTR, DTX treated (black circles) and 4 of which AdvillinFlp; CalcaCreER; R26FSF-LSL-Caspase3, 2mg tam daily P21-P23 (black squares) for pupil dilation and accelerometer and N = 6 for CGRP-ablated (2 of which are AdvillinCre; CalcaLSL-hDTR, DTX treated (black circles) and 4 of which AdvillinFlp; CalcaCreER; R26FSF-LSL-Caspase3, 2mg tam daily P21-P23 (black squares) for vocalization (note there are fewer animals for the vocalization data because a subset of the animals were run in a previous version of the setup with a different microphone). Comparisons for the pupil dilation and accelerometer data are paired t tests for the control, Wilcoxon matched-pairs signed rank tests for the CGRP-ablated, and Mann Whitney tests for the unpaired comparisons. Comparisons for the vocalization data are Wilcoxon matched-pairs signed rank tests for the paired comparisons and a Mann Whitney test for the unpaired comparison).
H-J: Behavioral responses (pupil dilation (H), movement (I), and vocalizations (J)) to high force colon balloon distension in Bmpr1b-ablated or Bmpr1bCre; Piezo2fl/fl animals, compared to controls. Each point represents the average of data from an individual animal. (N = 21 for controls; N = 9 for Bmpr1b-ablated; N = 7 for Bmpr1bCre; Piezo2fl/fl. Comparisons for pupil dilation data are Wilcoxon matched-pairs signed rank tests for the control and Bmpr1bCre; Piezo2fl/fl paired groups, a t test for the Bmpr1b-ablated paired groups, and Mann Whitney tests for unpaired groups. Comparisons for accelerometer and vocalization data are Wilcoxon matched-pairs signed rank test for paired groups and Mann Whitney tests for unpaired groups).
We found that colonic or skin light stimulation of animals expressing ReaChR in TrkB+ Aδ-LTMRs, using TrkBCreER; AdvillinFlp; R26FSF-LSL-ReaChR-mCitrine mice, did not elicit any obvious behavioral response (Figure 6A–C and S6A–C). Colonic light stimulation in animals expressing ChR2 in the TH+ C-LTMRs, using THCreER; Phox2B-Flp; R26FRT-LSL-ChR2-YFP-FRT mice, increased pupil dilation without affecting vocalizations or movement (Figure 6A–C). This was distinct from the skin light stimulation behavioral response in these animals, in which there was no increase in pupil dilation, movement, or vocalizations (Figure S6A–C). On the other hand, optogenetic-activation of CGRP+ DRG afferents, using CalcaCreER; R26LSL-ChR2-YFP or Calca-Flp; R26FSF-ReaChR mice, led to a strong behavioral response, with significant increases in pupil dilation, movement, and vocalizations during light stimulation of both the colon (Figure 6A–C) and skin (Figure S6A–C). Increases in pupil dilation, movement, and vocalizations may together indicate a nocifensive response, consistent with the known function of CGRP+ DRG afferents in the skin.
We next tested the individual CGRP subsets. As with the entire CGRP+ afferent population, optical activation of the Bmpr1b+ Aδ-HTMR subset of CGRP neurons, from both the colon and skin of Bmpr1bCre; Calca-Flp; R26FSF-LSL-mCitrine-ReaChR mice, was sufficient to drive increases in pupil dilation, movement, and vocalizations. This response was less pronounced than colon light stimulation of the entire CGRP+ population (Figure 6A–C and S6A–C), which includes both the Bmpr1b+ Aδ-HTMRs and Sstr2+ C-HTMRs. Conversely, selective activation of the Sstr2+ C-HTMR colon-innervating subtype, using Sstr2CreER; Calca-Flp; R26FSF-LSL-mCitrine-ReaChR or Sstr2CreER; R26LSL-mCitrine-ReaChR mice, led to significant increases in pupil dilation and vocalizations, but not movement, whereas skin activation led to increases in pupil dilation, vocalizations, and movement (Figure 6A–C and S6A–C). Notably, these responses were not due to activation of the SNS, ENS, or vagal afferents, as colon light activation in control animals expressing ReaChR in these populations, but not in the DRG (using Phox2B-Flp; R26FSF-ReaChR mice), led to a slight increase in pupil dilation without a change in vocalizations or movement (Figure 6A–C). Thus, optogenetic activation of Bmpr1b+ Aδ-HTMRs, and to a lesser extent Sstr2+ C-HTMRs, evoked behavioral responses analogous to the high colon distension response (Figure 1E–1G), whereas TrkB+ Aδ-LTMRs and TH+ C-LTMRs did not.
Since CGRP+ HTMRs respond to high force mechanical distension of the colon and they evoke robust behavioral responses when optogenetically activated, we aimed to define the role of the entire CGRP population in high-threshold colon mechanosensation. To this end, we ablated the entire CGRP population using two methods. First, we expressed the human diphtheria toxin receptor (hDTR), a well-established ablation method, exclusively in CGRP-expressing DRG afferents (AdvillinCre; CalcaLox-GFP-STOP-Lox-hDTR) (Figure 6D). While this method was often efficient, it required 21 days of low-dose diphtheria toxin (DTX) injection, as using higher amounts of diphtheria toxin (DTX) in fewer injections was toxic in control animals (data not shown). To circumvent this challenge and provide a complementary approach, we generated a new mouse line in which pro-Caspase3 and the protease necessary for its cleavage and subsequent activation are expressed only in the presence of both Cre and Flp recombinases, such that apoptosis is induced in a specific population (R26FSF-LSL-Caspase3) (Figure S6D). Using this novel mouse line, CGRP-expressing DRG afferents were ablated (CalcaCreER; AdvillinFlp; R26FSF-LSL-Caspase3) to a similar degree as with hDTR expression (Figure 6D). Ablating the CGRP-expressing afferents through either method significantly decreased behavioral responses to high colon distension (Figure 6E–G), indicating that one or more CGRP subpopulation is necessary for high-threshold colon mechanosensation.
We speculated that loss of the Bmpr1b-expressing CGRP population underlies some or all of the CGRP ablation phenotype. Bmpr1b+ Aδ-HTMRs were ablated by expressing human diphtheria toxin receptor (hDTR) in these neurons using Bmpr1bCre; CalcaLox-GFP-STOP-Lox-hDTR mice and treating them with DTX (Figure S6E). Indeed, ablation of Bmpr1b+ Aδ-HTMRs significantly decreased the high colon distension evoked behavioral response, indicating that this population is necessary for sensing high colon distension (Figure 6H–J). We also asked whether Piezo2 within Bmpr1b+ Aδ-HTMRs mediates behavioral responses to high force distension using Bmpr1bCre; Piezo2flox/flox mice in which Piezo2 was deleted in all Bmpr1b+ cells (Figure S6F). Piezo2 loss significantly decreased the high colon distension behavioral response (Figure 6H–J). Piezo2 deletion in the Bmpr1b+ population did not eliminate the physiologic response to high threshold colon distension in DRG calcium imaging but did significantly reduce the mean calcium activation response to high threshold distension (Figure S6G–I). These findings imply that Piezo2 expressed in Bmpr1b+ Aδ-HTMRs contributes to but is not fully responsible for the high colon distension response, analogous to noxious, high-threshold mechanosensation in the skin57.
Bmpr1b+ HTMRs mediate inflammation-induced hypersensitivity in the colon
We next explored the role of Bmpr1b+ Aδ-HTMRs in colon physiology and pathophysiology. Under physiological circumstances, it is thought that DRG afferents impact motility through the GI tract58. We measured distal colon motility and found that Bmpr1b+ Aδ-HTMR ablation significantly increased lower GI tract motility time, indicating these neurons contribute to motility under normal physiological conditions (Figure 7A). We also asked whether these Aδ-HTMRs mediate responses to other, non-mechanical stimuli. In an open field assay, ablation of the entire CGRP+ DRG neuron population did not significantly alter behavioral responses to colonic mustard oil, a TrpA1 agonist (Figure S7A–C). Thus, the Bmpr1b+ Aδ-HTMRs, which are a subset of the CGRP+ population and do not appreciably express TrpA113, while important for distension responses and motility, do not mediate the colonic mustard oil response.
Figure 7: Bmpr1b+ Aδ-HTMRs are necessary for colon physiology and inflammation-induced colonic mechanical hypersensitivity.
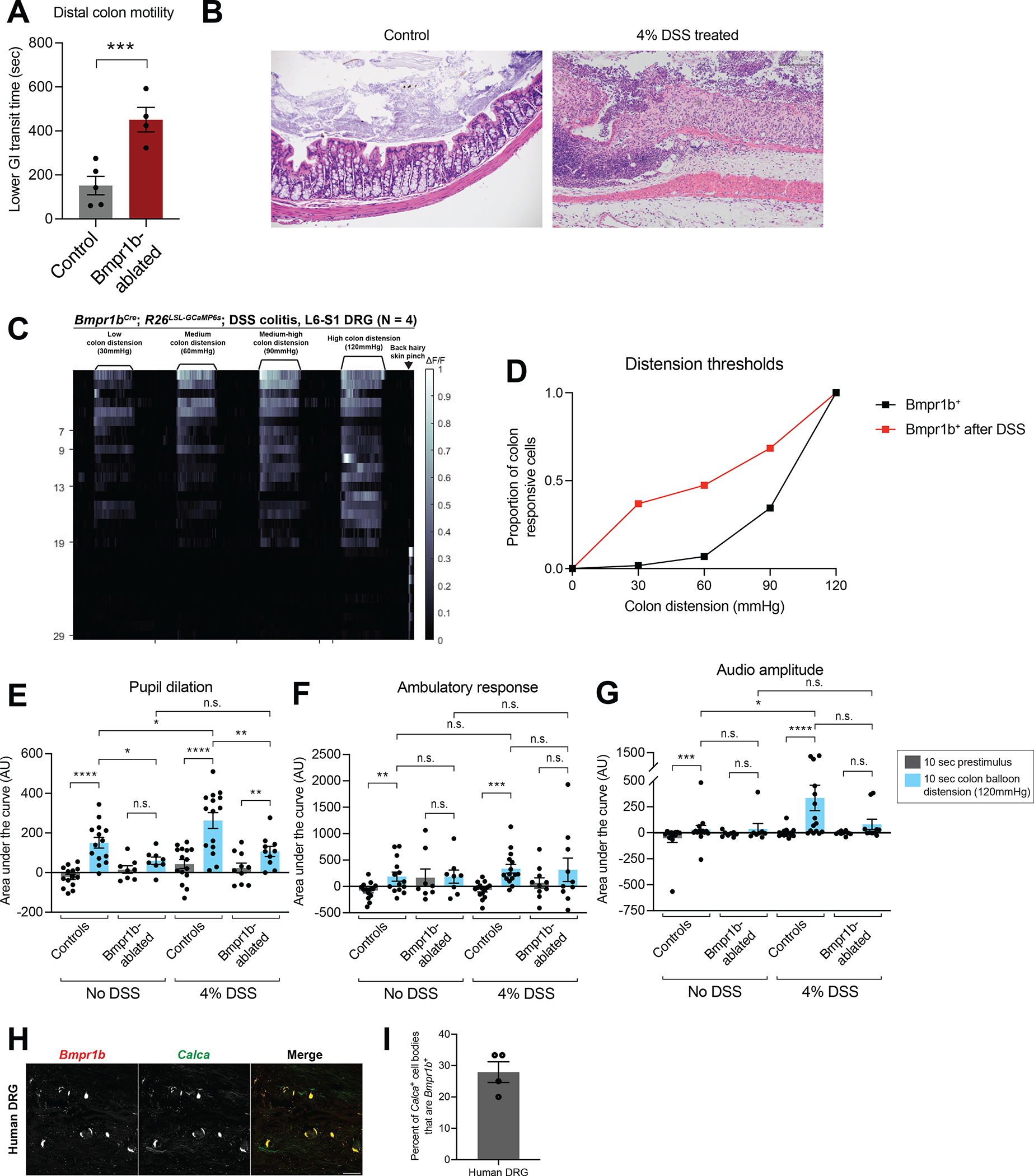
A: Distal colon motility in controls and Bmpr1b+ neuron-ablated animals.
B: Colon hematoxylin and eosin (H&E) staining in an example control and DSS treated animal. Scale bar is 200 μm.
C: In vivo DRG calcium imaging responses to colon distension in Bmpr1bCre; R26LSL-GCaMP6s mice following DSS treatment (N = 4).
D: Distension thresholds based on DRG calcium imaging of the Bmpr1b+ population in control or DSS treated animals. Data from Bmpr1bCre; R26LSL-GCaMP6s (N = 7 for control; N = 4 for DSS treated). The control data are replotted from the data shown in Figure 4G.
E-G: Behavioral responses (pupil dilation (E), movement (F), and vocalizations (G)) to high force colon balloon distension in Bmpr1b+ neuron-ablated mice or controls. Each point represents the average of data from an individual animal. (N = 14 control without DSS; N = 8 Bmpr1b-ablated without DSS; N = 15 control DSS-treated; N = 10 Bmpr1b-ablated DSS-treated. Comparisons for pupil dilation and accelerometer data are paired and unpaired t tests. Comparisons for vocalization data are Wilcoxon matched-pairs signed rank test for paired groups and Mann Whitney tests for unpaired groups.)
H: ISH of a human DRG reveals that some Calca+ DRG neurons are Bmpr1b+. The white arrow indicates a Bmpr1b+; Calca+ cell body. Note that the bright signal in all channels represents lipofuscin, which is commonly observed in ISH of human DRGs.
I: Quantification of the percent of Calca+ cell bodies in the human DRG that are Bmpr1b+. Data from N = 4 images.
Previous reports indicate that colon inflammation causes behavioral over-reactivity and physiologic hypersensitivity to colon distension, though the identity of the DRG neuron population that mediates this response has remained elusive. Because Bmpr1b+ Aδ-HTMRs are both necessary and sufficient to mediate a nocifensive-like behavioral response, we tested their role in inflammation-induced hypersensitivity. Indeed, the Bmpr1b+ Aδ-HTMRs were more sensitive to distension in the setting of inflammation as compared with controls, based on calcium imaging experiments in Bmpr1bCre; R26LSL-GCaMP6s DRGs after dextran sodium sulfate (DSS)-induced inflammation (Figure 7B–D). Furthermore, Bmpr1b+ Aδ-HTMR ablation (using Bmpr1bCre; CalcaLox-GFP-STOP-Lox-hDTR mice) reduced behavioral hypersensitivity to distension caused by DSS (Figure 7E–G, S7C). Control animals (untreated or DSS-treated) exhibited an increase in pupil dilation, movement, and vocalizations over their baseline in response to intracolonic balloon distension (Figure 7E–G). Ablated animals (untreated or DSS-treated) did not have an increased response over their baseline, apart from the DSS-treated Bmpr1b-ablated animals, which had an increase in pupil dilation only over their baseline, although this was significantly decreased in comparison to controls (Figure 7E–G). Additionally, while control animals displayed a significant increase in pupil dilation and vocalizations after DSS treatment, indicative of behavioral over-reactivity, Bmpr1b-ablated animals exhibited no change after DSS treatment (Figure 7E–G). To investigate if this hypersensitivity of Bmpr1b+ Aδ-HTMRs could be due to changes in expression of TrpV1 in the setting of inflammation, we performed ISH against TrpV1 in control and DSS-treated animals. We found that there was minimal TrpV1 expression in the Bmpr1b+ neurons in control animals and no change in TrpV1 expression in these cells following DSS treatment (Figure S7D–E). Finally, asked whether Bmpr1b+; CGRP+ neurons are observed in humans, which has been suggested by recent human DRG single cell sequencing analyses59. Indeed, we found that approximately 30% of CGRP+ DRG neurons in the human DRG express Bmpr1b (Figure 7H–I).
Taken together, gain of function and loss of function analyses indicate that Bmpr1b+ Aδ-HTMRs mediate pathophysiological responses to colon distension after chemical induced inflammation.
Discussion
Colon mechanosensation is important in both GI physiology and pathophysiology. However, lack of genetic access to colon innervating sensory neurons has hindered our understanding colonic sensory neurons and their functions. Here, we labeled distinct transcriptionally defined subtypes of DRG afferents, while excluding the other sources of innervation to the GI tract, thus enabling selective genetic access to colon innervating sensory neuron subtypes. We characterized nearly all Aδ and C fibers populations that have been identified in single cell sequencing experiments and found five colon innervating populations. Notably, due to the extensive nature of our search, it is likely that we have identified a majority of colon innervating DRG populations, though there could be additional populations that remain unidentified or for which we lack mouse tools.
Of these five subtypes, the TrkB+ colon innervating DRGs form elaborate IGVEs, respond to low threshold colon distension in a RA manner with short optogenetic latencies, establishing their colonic function at Aδ-LTMRs. The TH+ colon innervating DRG neurons form either circular muscle IMAs or less elaborate IGVEs, respond to low force distension in a SA manner, and have longer optogenetic latencies, signifying their colonic function as C-LTMRs. We also established the properties of three CGRP+ subtypes in the colon. The Bmpr1b+ CGRP subtype, which form simple IGVEs with 1–2 myenteric wrappings, respond to high threshold colon distension in a SA manner, and have slightly faster optogenetic latencies than the C-LTMRs and large cell bodies, implying their function as Aδ-HTMRs. The Sstr2+ CGRP subtype form endings that branch into the myenteric plexus, submucosa, or mucosa, respond to high threshold colon distension and intracolonic capsaicin in a SA manner, and have optogenetic latencies that are slower than the C-LTMRs, establishing their function as C-HTMRs. Finally, Adra2a+ CGRP neurons form IGVEs with few myenteric wrappings or branch into the submucosal layer, however, their physiological properties remain elusive and will need to be determined in future studies.
As in the skin, the colon is innervated by both LTMRs and HTMRs. Remarkably, there is a conservation of properties of three of the four subtypes that innervate the colon and skin. Thus, like Aδ-LTMRs that innervate hairy skin7, the TrkB+ neurons that innervate the colon are Aδ-LTMRs and respond in a RA-manner, whereas the TH+ afferents in the colon exhibit low force thresholds and slow conduction velocities in the C-fiber range, thus resembling C-LTMRs that innervate hairy skin8. Likewise, the Bmpr1b+ CGRP afferents are Aδ-HTMRs, similar to their cutaneous counterparts11,13. This conservation of response properties despite distinct end organ morphologies and environments in the skin and colon implies that the genetic composition of a mechanosensory neuron type dictates its force thresholds and adaptation to sustained stimuli.
Conversely, the TrpV1+ DRG afferents are thought to be mechanically insensitive but heat-responsive in the skin, and TrpV1 ablation leads to deficits in heat but not mechanical sensitivity60. Colon innervating, TrpV1+, Sstr2+ C-HTMRs, on the other hand, respond to both high-threshold colon distension as well as intracolonic capsaicin. Multiple possibilities may explain this dichotomy, including distinct microenvironments of the skin and colon or genetic heterogeneity within the Sstr2-expressing population.
Like their cutaneous counterparts, we find that most mechanosensitivity of colon innervating DRG afferents is mediated by Piezo2. However, some of the high-threshold force sensitivity of colon innervating DRG afferents is Piezo2-independent, implying that either a high-threshold mechanosensitive ion channel or tissue damage responses mediated by other signals.
We found that a subset of the TrkB+ Aδ-LTMRs respond to both colonic distension and skin pinch, implying that some of these neurons may innervate both organs. Interestingly, in an accompanying paper by Servin-Vences et al.61, recordings from Piezo2+ DRG neurons were done and neurons that respond to both skin and colon stimulation was not observed. Multiple explanations may account for this discrepancy, including that our calcium imaging experiments were performed with different Cre driver lines. While we recorded from DRG neuron subtypes separately, Servin-Vences et al. recorded from the majority of DRG neurons simultaneously, which may have obscured the relatively small number of rapidly adapting TrkB+ Aδ-LTMRs. Other experimental details, such as recording at different DRGs (L6/S1 in the present paper and S2 in Servin-Vences et al.) or stimulating the skin in different locations could also account for these differences.
Interestingly, Bmpr1b+ Aδ-HTMR ablation prolongs colon motility, implicating this HTMR population in normal physiologic function. It is notable that this DRG afferent’s role in the distal colon appears to be necessary to sense contents and increase expulsion, though future work will be needed to characterize its role in other parts of the GI tract.
Finally, while it is well-established that colon inflammation, such as in the setting of inflammatory bowel disease, leads to mechanical hypersensitivity, the genetic identity, morphologic and physiologic properties, and functions of intrinsic and extrinsic neurons responsible for this behavioral change have remained elusive. We found that Bmpr1b+ Aδ-HTMRs become more sensitive and mediate pathophysiological responses to colon distension after inflammation and may therefore represent a clinically relevant therapeutic target for treating colon pain. Further work is needed to deeply probe the genetic expression profiles of Bmpr1b+ Aδ-HTMRs and other DRG afferent populations13 to uncover potential molecular targets to treat abdominal pain.
Limitations of the study
Here, we identify five colon innervating DRG populations. While we have extensively characterized these populations, additional studies will be needed to understand if the subtypes for which we do not have specific mouse tools or if other, not yet identified subtypes exist and innervate the colon. One subpopulation expressing Adra2a was described anatomically but, due to the sparse labeling with the Adra2aCreER line, was not characterized functionally. Future work is needed to understand its role in colon sensation. Finally, while we characterized the responses of these populations to colon mechanosensation, additional work is needed to understand their responses to other colonic stimuli.
STAR Methods
Resource availability
Lead contact
Further information and requests for resources and reagents should be directed to and will be fulfilled by the lead contact, David Ginty (david_ginty@hms.harvard.edu).
Material availability
All new reagents will be made available upon request.
Data and code availability
All data reported in this study will be shared by the lead contact upon request.
All original code is available in the paper’s supplemental information and will be shared by the lead author upon request.
Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.
Experimental model and study participant details
Animals
All mouse experiments in this study were approved by the National Institutes of Health and the Harvard Medical School IACUC. Experiments followed the ethical guidelines outlined in the NIH Guide for the care and use of laboratory animals (https://grants.nih.gov/grants/olaw/guide-for-the-care-and-use-of-laboratory-animals.pdf). Mice were housed under standard conditionsand given chow and water ad libitum.
Bmpr1bT2a-CreER and R26FSF-LSL-Caspase3 mice were generated using standard CRISPR-based homologous recombination techniques in embryonic stem (ES) cells. Chimeras were generated by blastocyst injection and subsequent germline transmission was confirmed by tail PCR. The neo selection cassette was excised using a Flp-deleter strain in the case of the Bmpr1bT2a-CreER and a R26phiC31 strain in the case of the R26FSF-LSL-Caspase3 mice.
The following mice are available from Jax: Advillin-CreERT2 (Stock No. 032027), AdvillinCre (Stock No. 032536), Piezo2flox (Stock No. 027720), Phox2B-Flp (Stock No. 022407), Cdx2-Cre (Stock No. 009352), MrgprdCreER (Stock No. 031286), R26FSF-LSL-tdTomato (Ai65; Stock No. 021875), R26LSL-tdTomato (Stock No. 007914), R26FSF-tdTomato (derived from Ai65; Stock No. 021875), R26FRT-LSL-ChR2-YFP-FRT (Ai32; Stock No. 012569), R26FRT-LSL-tdTomato-FRT (Ai9; Stock No. 007909), R26FSF-LSL-ReaChR (Stock No. 024846), R26LSL-GCaMP6s (Ai96; Stock No. 028866), and Igs7LSL-GCaMP6f (Ai148; Stock No. 030328). Mice generated in the Ginty lab (and available at JAX now or in the near future) are: AdvillinFlp (Choi et al, 2020)62, THCreER (Stock No. 025614)14, Calca-Flp (Choi et al, 2020)62, Bmpr1bCre (Sharma et al, 2020)13, Sstr2CreER (Qi et al, 2023)43, Smr2Cre (Qi et al, 2023)43, Adra2aCreER (Qi et al, 2023)43, and TrkBCreER (Rutlin et al, 2014)7. Other mice used in the study have been previously described: CalcaCreER (Song et al, 2012)63, CalcaLox-GFP-STOP-Lox-hDTR (McCoy et al, 2012)36, Calca-GFP (MMRRC, Stock No. 011187-UCD), Mrgpra3-Cre (Han et al, 2013)64, Mrgprb4CreER (Vrontou et al, 2013)65, and PIRTCre (Patil et al, 2019)47.
Animals were maintained on mixed C57bl/J6, 129S1/SvImJ and CD1 backgrounds and included both males and females. All experiments with wild-type animals were conducted with mice on the C57Bl/6J background obtained from Jackson Laboratory. All animals used were immunocompetent. Some animals were first used for behavioral experiments and subsequently used for anatomy or physiologic recordings. Animals were drug naïve. Some animals were not test naïve; for example, some were used for establishing the parameters of behavioral assays and subsequently used in the final version of the behavioral assay.
Method details
Tamoxifen treatment
Tamoxifen was dissolved in ethanol (20 mg/ml). It was then mixed with an equal volume of sunflower seed oil (Sigma) and vortexed for 20 minutes. The mixture was subsequently centrifuged under vacuum for 30 minutes to remove the ethanol. The solution was kept at −80 °C. It was delivered by oral gavage to pregnant females for embryonic treatment, or by intraperitoneal (IP) injection for postnatal treatment. For all analyses, the plug date was designated as E0.5 and the date of birth as P0.
Mice were treated with tamoxifen at the following timepoints (by genotype) unless otherwise specified: Advillin-CreER (P21–25); MrgprdCreER (P6-P9); TrkBCreER (P3–5); THCreER (P21); CalcaCreER (P11); Sstr2CreER (P21); Adra2CreER (P14 and P16); Bmpr1bCreER (P21). Animals were sacrificed at least more than 2 weeks after tamoxifen treatment.
Whole mount immunofluorescence
Immunofluorescence experiments were done in animals between 3–12 weeks old. The whole-mount immunofluorescence (IF) was performed as previously described7 for skin samples. The protocol was adapted for optimized use in the GI tract. Mice (P21–42) were euthanized with isoflurane and perfused with 5ml PBS followed by 10–20 ml of 4% paraformaldehyde (PFA) in PBS at room temperature (RT). Skin and the GI tract were dissected from perfused mice. Hair was removed from the skin using commercial depilatory cream (NAIR, Church and Dwight Co.; Princeton, NJ) for 0.5–1 min and washed with water. The colon was dissected open and the luminal contents were removed. Skin and colon were post-fixed in Zamboni’s fixation buffer at 4°C for 12–24 hours. Thereafter, the samples were washed with PBS for 3×5 minutes, followed by PBST (PBS with 1% Triton X-100) every 20 minutes for 5–7 hours. A small piece of each tissue was cut (0.5mm × 0.5mm) and incubated in primary antibodies diluted in blocking solution (75% 1% PBST, 20% DMSO, 5% Normal Goat Serum (NGS) (Vector Labs, S-1000) or Normal Donkey Serum (NDS) (Jackson Immuno, 005-000-121)) for 3–4 days on a rocking platform at RT. The samples were thereafter washed again in PBST for 5–7 hours every 20 minutes and then incubated with secondary antibodies in blocking solution on rocking platform for 2 days at RT. The samples were again washed in PBST for 5–7 hours every 20 minutes. The samples were subsequently dehydrated: 1 wash for 10 minutes in 50% methanol (MeOH), 1 wash for 10 minutes in 80% MeOH, followed by 3 washes for 20 minutes in 100% MeOH. The samples were stored in 100% MeOH at −20°C. To image, the samples were cleared in BABB (1 part Benzyl Alcohol: 2 parts Benzyl Benzoate) for 5 minutes, and then mounted on slides using BABB as the mounting medium.
Tissue section immunofluorescence
Mice were perfused as above. Spinal columns containing spinal cords, DRGs, and sympathetic chain were dissected, post-fixed at 4°C in a PFA (4% in PBS) solution for 12–24 hours, and subsequently washed 3×5 minutes in PBS. For DRG and spinal cord IF, the spinal cord and DRGs were dissected out after fixation. For DRG, spinal cord, and sympathetic chain IF, the spinal column was left intact. Tissues were cryoprotected in 30% sucrose in PBS at 4°C for two nights and thereafter embedded in OCT and frozen at −20°C. Tissues were sectioned at 20–30μm with a cryostat. Slides were rehydrated with PBS 3×5 minutes and then blocked with PBS containing 0.1% Triton X-100 and 5% NGS or NDS for 20–60 minutes at room temperature. Tissue sections were incubated with primary antibodies diluted in PBS with 5% NGS or NDS overnight at 4°C. The following day, sections were washed 4×5 minutes in PBS containing 0.02% Triton X-100 and then incubated with secondary antibodies diluted 1:500 in PBS with 5% NGS or NDS for 1 hour at RT. The sections were washed again 4×5 minutes in PBS containing 0.02% Triton X-100 and mounted with fluoromount-G (Southern Biotech).
Primary antibodies used for both whole mount and tissue section IF include: chicken anti-GFP (GFP-1020, Aves Labs; 1:1000), goat anti-GFP (G8965-01E; US Biological Life Sciences; 1:500), rabbit anti-DsRed (632496; Clontech; 1:500), goat anti-mCherry (AB0040-200, Acris, 1:500), rabbit anti-CGRP (Immunostar, 24112, 1:500), chicken anti-NF200 (Aves Labs, NFH, 1:1000), rabbit anti-TUJ1 (Biolegend, 802001, 1:500), sheep anti-TH (Sigma, AB1542; 1:500), rabbit anti-TH (Sigma; AB152; 1:500), and mouse anti-NeuN (Sigma; MAB377; 1:500).
Secondary antibodies used for both whole mount and tissue section IF included: Alexa 488, 546 or 647 conjugated goat anti-chicken antibodies, Alexa 488, 546 or 647 conjugated goat anti-rabbit antibodies, Alexa 488, 546 or 647 conjugated donkey anti-goat antibodies, Alexa 488, 546 or 647 conjugated donkey anti-rabbit antibodies, Alexa 488, 546 or 647 conjugated donkey anti-goat antibodies. All secondary antibodies above were purchased from Life Technologies, except Alexa 488/647 conjugated donkey anti-chicken antibodies, which were purchased from Jackson ImmunoResearch.
For all anatomy experiments, data was collected from at least N = 2 animals. All scale bars represent 100 μm unless otherwise specified. Note that for any anatomical analysis, the DRGs are typically lumbar level, and the sympathetic ganglia are typically thoracolumbar.
In situ hybridization
RNA fluorescence in situ hybridization (RNA-FISH) was performed as previously described13. Briefly, individual DRGs were rapidly dissected from mice and subsequently frozen in dry-ice-cooled 2-metylbutane and stored at −80°C until they were sectioned. Human L5 DRGs were ordered from AnaBios. DRG were sectioned at a thickness of 20 μm and RNAs were detected with RNAscope (Advanced Cell Diagnostics) using the manufacturer’s protocol. The following probes were used: Mm-Bmpr1b (Cat#: 533941), Mm-Sstr2 (Cat#: 437681), Mm-Adra2a (Cat#: 425341), Mm-TrkB (NTRK2) (Cat#: 423611), Mm-TH (Cat#: 317621), Mm-Calca TV2-TV3 (Cat#: 420361), Mm-Piezo2 E43–45 (Cat#: 439971), Mm-Trpv1 (Cat#: 313338), Hs-Bmpr1b (Cat#: 468031), Hs-Calca (Cat#: 605551), GCaMP6s (Cat#: 557091), and tdTomato (Cat#: 317041).
Tissue processing for H&E
Animals were perfused and colons were dissected as above, although the colons were not dissected open, and their luminal contents were left intact. Colons were post-fixed at 4°C in 4% PFA for 12–24 hours, and subsequently washed 3×5 minutes in PBS. Tissues were embedded in paraffin and sectioned. Thereafter, slides were stained with standard hematoxylin and eosin (H&E) staining techniques and imaged on a light microscope (Olympus BX63).
Single neuron morphological analysis
For sparse labeling of single neurons, mice were treated with approximately 1/10th or 1/100th the usual tam dose for each subtype. The tissues were processed by whole mount immunofluorescence. The endings were traced in ImageJ using Single Neurite Tracer (SNT). Parameters such as process length, width, and number of branches were calculated using SNT.
Bmpr1b+ and CGRP+ neuron ablation analysis
To assess the percent of Bmpr1b+ neurons ablated per DRG in Bmpr1bCre; CalcaLSL-hDTR DRGs, we first used in situ hybridization to confirm in two animals that > 75% of the Bmpr1b+ cells were ablated. We also calculated the average percent of Calca+ cells that were also Bmpr1b+ per DRG in controls (average across N = 3 DRGs). For most animals, we used immunofluorescence to quantify the number of CGRP+ cells over the total number of cells per DRG (total number of cells quantified by NeuN staining). The average percent of Bmpr1b+ cells out of the total Calca+ population was then used to calculate the amount of Bmpr1b+ cell ablation per DRG. This was compared between controls and ablated animals.
To assess the percent of CGRP+ cells ablated per DRG in AdvillinCre; CalcaLSL-hDTR or CalcaCreER; AdvillinFlp; R26FSF-LSL-Caspase3 DRGs, immunofluorescence was used to quantify the number of CGRP+ cells over the total number of cells per DRG (total number of cells quantified by NeuN staining). This was compared between ablated animals and an average of multiple controls to calculate percent ablation. Note that ablation efficiency for the R26FSF-LSL-Caspase3 line was best with the allele homozygous (CalcaCreER; AdvillinFlp; R26FSF-LSL-Caspase3/FSF-LSL-Caspase3). Animals were only included in behavioral analyses if they had > 75% ablation.
Piezo2 deletion and TrpV1 quantification
To quantify Piezo2 deletion in Bmpr1bCre; Piezo2fl/fl or Bmpr1bCre; R26GCaMP6S; Piezo2fl/fl animals or TrpV1 expression in control and DSS-treated wild-type animals, DRGs were processed for in situ hybridization. All experiments had control and knockout or DSS-treated slides processed on the same day, with the same antibody conditions. Samples were also imaged on the same day using the same microscope settings, and all processing of the images was consistent between control and knock-out or DSS slides. For Bmpr1bCre; Piezo2fl/fl or control and DSS-treated DRGs, regions of interest (ROIs) were drawn in ImageJ around Bmpr1b positive cells. The total fluorescence intensity in the Piezo2 or TrpV1 channel of each of these regions was calculated in ImageJ and divided by the cell area to quantify the Piezo2 expression per genotype or TrpV1 expression per treatment condition. For the Bmpr1bCre; R26GCaMP6S; Piezo2fl/fl DRGs, the ROIs were drawn around the GCaMP6S cells and processed in an analogous fashion.
Cell size quantification
To quantify cell size in each colon-innervating subtype wild-type DRGs were processed for in situ hybridization (from N = 3 mice). Samples were imaged with the same microscope settings and all processing of the images was consistent between genotypes. ROIs were drawn in ImageJ around TrkB+, TH+, Bmpr1b+, Sstr2+, or Adra2a+ cells and the area (in px2) of each ROI was calculated using ImageJ.
ENS and SNS labeling quantification
To quantify ENS labeling, the total number of myenteric cell bodies labeled using a control stain (such as TUJ1) was counted. Then, the number of myenteric cell bodies that were labeled by the mouse line of interest was counted and a percent of total myenteric cell bodies per image was calculated. Similarly, for SNS labeling, the total number of sympathetic neuron cell bodies labeled using a TH control stain was counted. Then, the number of sympathetic cell bodies that were labeled by the mouse line of interest was counted and a percent of total SNS cell bodies per image was calculated.
Density labeling
To calculate the density of fibers at the level of myenteric vs. submucosal plexus, the image was opened in ImageJ and a diagonal line was drawn across the image from the bottom left corner to the top right corner. This line represents the normalized area of each image. The plot profile function in ImageJ was used to calculate the fluorescence intensity in the normalized area. A custom program in Matlab was used to calculate the number of peaks of fluorescence over a background threshold of 50 (to exclude background signal) and the number of peaks across the line was reported. This number represents the number of fibers that cross the diagonal line across each image, and thus provided an estimate of the density of fibers across the normalized area of an image.
In vivo calcium imaging and electrophysiology
Mice were anesthetized with urethane (0.5–1.5 mg/g) and placed on a bite bar (Kopf Instruments) on a custom heated recording platform. Temperature was monitored throughout the procedure with a temperature controller (TC-344B, Warner Instruments) and maintained at 35.5–37.5°C with a thermoelectric heater (C3200-6145, Honeywell) embedded in castable cement (Aremco). To expose L6-S1 DRGs, an incision was made over the spine (T10-S2) and the overlying tissue was retracted, exposing the vertebral canal. The vertebral canal was secured with custom spinal clamps (made in the Harvard Medical School Neurobiology Department Machine Shop) and the bone dorsal to the target DRG was removed using forceps.
The exposed L6-S1 DRG was then used for calcium imaging with a Zeiss Axio Examiner microscope or multiunit electrophysiological recordings using an MEA probe (H4-32ch, Cambridge Neurotech) that was inserted into the DRG for recording.
A balloon attached to a manometer (Cole Parmer) was inserted into the colon of the mice. For all calcium imaging and electrophysiology experiments, each colon distension measurement involved a 1-minute baseline, 1 minute with the balloon distended to the specified pressure, and a 1-minute period after deflation for a total 3-minute recording. Prior to insertion, the balloon measured 2 mm in diameter (for reference, stool pellets from a 12-week-old mouse were also on average 2 +/− 0 mm in diameter). With the balloon inserted prior to inflation, the colon diameter measured 2.67 +/− 0.23 mm. The colon diameter with balloon inserted increased to 3.33 +/− 0.47 mm at 30 mmHg, 4 +/− 0.41 mm at 60 mmHg, 4.42 +/− 0.31 mm at 90 mmHg, and 5 +/− 0 mm at 120 mmHg of balloon distension (all measurements are mean +/− standard deviation across N = 3 animals). For hairy skin pinch measurements, the pinch was performed using forceps after a 10-second baseline and with a 10 second period after stimulus, for a total 20-second recording.
For colon optogenetic stimulation in MEA experiments, the MEA probe was left in place in the DRG and the colon balloon was replaced with an LED probe (ThorLabs; 470 nm, 17.2 mW (Min) Fiber-Coupled LED) inserted 1cm into the colon. Opto-responsive units were determined by delivering brief (5–20ms) pulses of blue light (60mW) to the colon. The first spike to each light pulse was used to determine the optogenetic latency.
Spike sorting
Spike sorting was performed as previously described69,70. Briefly, open-source software (JRCLUST version 3.2.2) was used to automatically sort action potentials into clusters, manually refine clusters, and classify clusters as single or multi units. The voltage traces were filtered with a differentiation filter of order 3. Frequency outliers were removed with a threshold of 10 median absolute deviations (MADs). Action potentials were detected with a threshold of 4.5 times the standard deviation of the noise. Action potentials with similar times across sites within 60 mm were merged and action potentials were then sorted into clusters with a density-based-clustering algorithm (clustering by fast search and find of density peaks) with cutoffs for log10® at −3 and log10(d) at 0.6. Clusters with a waveform correlation greater than 0.99 were automatically merged and outlier spikes (> 6.5 MADs) were removed from each cluster.
Manual cluster curation was performed with JRCLUST split and merge tools. Single units were classified based on waveform consistency, spike amplitude, clear refectory periods (interspike intervals of > 1ms), and waveform separability with nearby clusters. Only clusters determined to be single units were used. Spike times for single units were exported and analyzed in Python (3.8.5).
Calcium imaging analysis
The motion correction of the calcium imaging videos was performed in ImageJ using the moco plugin71. Individual responding ROIs were manually segmented. Measured intensity for each ROI was further processed using a custom MATLAB code. Bleach correction was performed before calculating ΔF/F. Response was defined as 2.5 times over the standard deviation of the baseline. In the heatmap plot, cells of the same subtype were sorted by the maximum response.
To estimate the relative proportions of each population as a total of colon innervating DRG neurons, we calculated from the calcium imaging data that on average the TrkB+ population had 7.7 colon-responsive cells per DRG, the TH+ population had 6.6 colon-responsive cells per DRG, the Bmpr1b+ population had 8.3 colon-responsive cells per DRG, and the Sstr2+ population had 6.4 colon-responsive cells per DRG. If we estimate that these four populations represent 80% of colon-innervating neurons in L6-S1 DRG (accounting for the Adra2a+ population and possibly other, unknown, but more rare subtypes), then the TrkB+ population would represent approximately 21% of all colon-innervating cells; the TH+ population would represent ~18% of all colon-innervating cells, the Bmpr1b+ population would be ~23% of all colon-innervating cells, and the Sstr2+ population would be ~18% of all colon-innervating cells.
Dorsal horn injections
Mice (P21–P28) were anesthetized by continuous inhalation of isoflurane (1.5–2.5%) using an isoflurane vaporizer (VetEquip). Laminectomies were performed to expose the lumbar spinal cord at L6, and a total of 450 nl of AAV virus (AAV2-retro-hSyn-FlpO, Boston Children’s Core; titer 2.06×1014 gc/ml) was directly injected into two or three adjacent spots in the dorsal spinal cord using pulled glass pipettes (Wiretrol II, Drummond) and a microsyringe pump injector (UMP3, World Precision Instruments).
Behavioral testing
Male and female mice of mixed genetic background were used for all behavior experiments. Testing was done with mice between 6–12 weeks of age. All animals were group housed and controls for most experiments were littermates. The exceptions to this were three of the controls in the Cdx2-Cre; Piezo2flox/flox balloon distension experiments and wild-type optogenetic experiments, for which wild-type C57BL/6J animals were used. For all behavioral assays, mice were habituated to the behavior room in their home cage for 20 minutes each day, prior to either a habituation or test day.
Headplating
For all behavior experiments in which pupil dilation, movement, and vocalizations were recorded, the mice were headplated. For this, mice were anaesthetized with 1.5–2% isoflurane, the scalp was removed, the skull was dried, and a titanium headplate was affixed to the skull using dental cement (Metabond). The mice were treated with sustained release buprenorphine (0.1 mg/kg) after the procedure. Mice were allowed at least 5–7 days to recover after the procedure prior to undergoing behavioral testing.
Colon distension and optogenetic behavioral testing
The behavioral setup for colon distension and optogenetic testing involved a bar to which the headplate could attach mounted next to a custom tube, to body fix the animals. The tube was mounted on foam to allow some vibration with movement of the mouse. The accelerometer was attached to the outside of the tube. A video camera (digital USB 2.0 CMOS) was mounted to record the eye and a microphone which could record in the ultrasonic range (Bat Conservation and Management, Inc.) was mounted at the front of the setup. The setup was placed in a sound-attenuating box (Med. Associates, Inc.) to decrease ambient noise.
For habituation, the mice were head-fixed and body-fixed in the setup for 5 minutes on the first day and 10 minutes on the second day. An investigator held their tail in place to re-capitulate the day of the assay. The behavioral test was performed after 2 days of habituation.
For colon distension testing, the intracolonic balloon was built as previously described72. The balloon, which was connected to a manometer (Cole Parmer) was inserted into the colon of the mice and secured to the tail with tape. An investigator lightly held the tail in place to avoid balloon movement causing damage to the colon. Each distension trial was 5 minutes in total and included a 1-minute baseline, followed by four balloon inflations (lasting 10 seconds each) at each minute mark. The balloon was inflated to 2.4PSI within 2 seconds and held at 2.4PSI for the 10 second stimulus duration. Each animal was tested for at least 1 trial. If an animal displayed stressful behaviors such as kicking during the baseline period, then the trial was repeated and the trial with the least amount of baseline stress was used for the final analysis.
For colon optogenetic testing, an LED (1 mm in diameter) was inserted 1cm into the colon and secured to the tail with tape. An investigator lightly held the tail in place to avoid LED movement causing damage to the colon. Each trial was 5 minutes in total, with a 1-minute baseline, followed by four 10-second blue light stimulations at 10Hz. For animals expressing Channelrhodopsin2 (ChR2), 60mW power was used, while for animals expressing Red-shifted variant of Channelrhodopsin (ReaChR), 36mW power was used. For each animal, the optogenetic stimulation was repeated three times and the average of the three trials was used for each animal.
After colon optogenetic activation, the LED was removed from the colon, cleaned, and optogenetic stimulation of the back left paw was performed with the same experimental paradigm as colon stimulation.
The blue LED (Doric Lenses) was connected to a programmable LED driver (Doric Lenses). Optogenetic stimulation was controlled by the combination of custom programs written in Bonsai software and Doric Neuroscience Studio (4.1.5.2) through an Arduino circuit board (Uno, Arduino) and custom sketches written in Arduino software (1.8.7).
Pupil dilation, movement, and vocalization analysis
The pupil dilation analysis was performed as previously described62. Briefly, pupils were recorded at 30 frames s−1 using a digital USB 2.0 CMOS video camera mounted near one eye. An infrared illuminator was used to obtain clear images of the pupil. The pupil diameter was tracked at 10Hz using a custom program written in Bonsai software.
Movement (ambulatory response) data was collected using an accelerometer (SparkFun ADXL335) connected to a data acquisition card (National Instruments). The movement in the X, Y, and Z axes was summed at each time point for the total movement. The data were collected at 20Hz using a custom program written in Bonsai software and analyzed using a custom program written in Matlab.
Vocalizations (including in the ultrasonic range) were recorded using one of two microphones (UltraVox or Bat Conservation and Management, Inc.). The amplitude and frequency of sound were recorded at 20Hz using a custom program written in Bonsai software and analyzed using a custom program written in Matlab.
The average baseline pupil diameter, ambulatory response, or audio amplitude was the average of those values during the 1-minute baseline period before the beginning of the first optogenetic stimulation or balloon distension. The value at each time point was divided by the average of the 1-minute baseline to obtain a normalized value. This value was subtracted from 1 and multiplied by 100 to obtain a percent change in pupil dilation, movement, or vocalizations. The area under the curve (AUC) in the 10 second prestimulus time period and 10 second stimulus period were calculated using a custom program written in Matlab for each stimulation. The average prestimulus and stimulus values were calculated for each mouse (across either four balloon distensions in one five minute trial for the balloon assay, or 12 total optogenetic stimulations across three trials for the optogenetic activation assay).
Diphtheria toxin (DTX) mediated ablation
Bmpr1bCre; CalcaLSL-hDTR or AdvillinCre; CalcaLSL-hDTR mice were treated every day for 21 days with intraperitoneal (IP) injections of DTX (0.175 μg/g total over 21 injections) starting at age 6 weeks. Animals were tested in behavioral assays 18 days after completing DTX injections. Ablation efficiency was calculated as described above. Data shown are from animals with > 75% ablation.
Colon motility assay
Colon motility was assessed as previously described73–75. Briefly, the mice were fasted overnight with free access to water. Mice were lightly anesthetized with isoflurane and a lubricated 3mm glass bead was gently inserted 2cm into the colon using a slender glass rod (McMaster Carr) with a demarcation at 2cm. Mice were then placed in an empty cage with no bedding and the time to bead expulsion was recorded.
Colonic mustard oil behavioral testing
Colonic mustard oil behavioral testing was performed as previously described76. Briefly, on day 1, animals were habituated for 20 minutes in the chambers, which were 10cm × 10cm × 10cm acrylic chambers with one clear wall and three black walls. During this 20-minute period on day 1, animals were tracked with an overhead camera to record their baseline movement in the open field assay. On day 2, animals were again habituated for 20 minutes to the chamber.
The behavioral test was performed on day 3. Prior to the start of the assay, animals were anesthetized briefly with isoflurane. 50μl of either vehicle (70% ethanol dissolved in sterile saline) or 1% mustard oil in vehicle (Sigma) was inserted ~1cm into the distal colon using a 24G reusable oral gavage needle (Fine Science Tools (FST)). Animals were then placed into the chamber and given 5 minutes to recover from the brief anesthesia. Their movement on an overhead camera as well as their behaviors on a camera mounted in front of the test chamber were recorded for 20 minutes.
For the distance traveled analysis, the mouse centroid was tracked, and speed and velocity of mouse locomotion were analyzed at 2 Hz with video files taken using an overhead camera and a custom program written in Bonsai software62. The distance traveled on day 3 (the day of the assay) was normalized to distance traveled at baseline (day 1) for each animal. Abdominal licking and contractions were analyzed manually using video files filmed by the camera mounted in front of the test chambers by investigators who were blinded to genotype.
Dextran sodium sulfate (DSS)-induced colitis
To induce colitis, mice were treated with a solution containing dextran sodium sulfate (DSS; 4–5%) (MP Biomedical) dissolved in the drinking water. Controls were allowed access to regular drinking water. Animals were weighed on day 0 (the day when DSS was added to the drinking water). On each day of DSS treatment, weight and stool characteristics were recorded for each animal. This allowed for monitoring of the disease activity index (DAI), as previously described77. Animals were allowed access to 4–5% DSS in the drinking water for 6 days, followed by 1 day of regular drinking water. The behavioral and physiological experiments were performed on day 7, and the animals were perfused immediately thereafter for histological analysis.
Quantification and statistical analyses
Statistical analyses were performed using GraphPad Prism (Version 8, GraphPad Software). The number of mice and the statistical tests used for individual experiments are included in the figure legends. Both non-parametric tests and parametric tests were used, depending on data normality, for comparing two independent groups (Mann-Whitney U test or Student’s t test) or two related groups (Wilcoxon matched-pairs signed rank test or paired Student’s t test) and multiple groups (Kruskal-Wallis H test or one-way ANOVA). The following symbols are used in the figure legends for P values: n.s., not significant; *, P < 0.05; **, P < 0.01; ***, P < 0.001; ****, P < 0.0001. For all analyses, power analyses were conducted to calculate the ideal sample size for each experiment following experiments using 3–5 mice in each behavioral assay. Mice were randomly allocated into experimental groups when possible. Experiments were replicated in multiple animals (at least N = 2 for anatomy and at least N = 3 for behavior and physiologic experiments). For balloon distension experiments, if control animals did not respond to the balloon distension, the data (for both controls and experimental animals) was not included. All graphs show mean +/− standard error of the mean (SEM). For histological analyses, images were analyzed by investigators who were blinded to genotype whenever possible. For the behavioral assays, approximately half of the initial experiments were performed by experimenters who were not blinded to genotype, and the remainder of the experiments were performed by experimenters who were blinded to genotype, when possible. The data for all behavioral experiments were analyzed by experimenters who were blinded to genotype. Blinding was not used for the collection of data in electrophysiological recordings, but the data were analyzed automatically using Matlab or Python with the same scripts run for each experimental group.
Supplementary Material
A-M: Skin and colon whole mount immunofluorescence or DRG, superior cervical ganglia (SCG), sympathetic chain, or spinal cord immunofluorescence in (A-C) Advillin-CreER; R26LSL-tdTomato mice treated with 1mg tamoxifen at P5; (D-F) Advillin-CreER; R26LSL-tdTomato mice treated with 0.5mg tamoxifen at P1; (G-H) Advillin-CreER; R26LSL-tdTomato mice treated with 3mg tamoxifen at E12.5; (I-J) AdvillinFlp; Cdx2-Cre; R26FSF-LSL-tdTomato; (K) PIRTCre; R26LSL-tdTomato; (L-M) Phox2B-Flp; R26FSF-ReaChR-mCitrine; or (N) Advillin-CreER; Phox2B-Flp; R26FRT-LSL-ChR2-YFP-FRT animals treated with 1mg tamoxifen P21–25. All scale bars are 100 μm unless otherwise specified.
O: Brainstem immunofluorescence in Advillin-CreER; Phox2B-Flp; R26FRT-LSL-ChR2-YFP-FRT treated with 1mg tamoxifen P21–25 (area postrema (AP), nucleus tractus solitarius (NTS), dorsal column nucleus (DCN), spinal trigeminal nucleus (SPV)). Scale bar is 500 μm. Animals were sacrificed at 3–6 weeks.
P: ENS labeling quantification (percent of myenteric cell bodies labeled) across the different mouse lines. 3 separate images were quantified from at least N = 2 animals.
Q: SNS labeling quantification (percent of sympathetic cell bodies labeled) across the different mouse lines. Images quantified from at least N = 2 separate sympathetic ganglia.
R: Quantification of the fiber labeling density at the level of the myenteric vs. submucosal plexus in the Advillin-CreER; Phox2B-Flp; R26FRT-LSL-ChR2-YFP-FRT animals. At least 3 separate images were quantified from N = 3 animals. Compared using an unpaired t test.
A-F: Skin and colon whole mount immunofluorescence or DRG and sympathetic chain immunofluorescence in (A) MrgprdCreER; Phox2B-Flp; R26FRT-LSL-tdTomato-FRT mice (1mg tamoxifen at P7); (B) TrkBCreER; AdvillinFlp; R26FSF-LSL-ReaChR-mCitrine mice (0.5mg tamoxifen at P5); (C) THCreER; Phox2B-Flp; R26FRT-LSL-tdTomato-FRT mice (0.5–1mg tamoxifen at P11–14); (D) Calca-GFP; (E) CalcaFlp; R26FSF-tdTomato; (F) CalcaLox-GFP-STOP-Lox-hDTR mice.
G: ENS labeling quantification (percent of myenteric cell bodies labeled) across the different Calca mouse lines. At least 2 separate images were quantified from at least N = 2 animals.
H-N: DRG and sympathetic chain immunofluorescence in (H-I) Smr2Cre; R26FRT-LSL-tdTomato-FRT mice; (J) Bmpr1bCre; AdvillinFlp; R26FSF-LSL-tdTomato mice; (K-M) Sstr2CreER; Phox2B-Flp; R26FRT-LSL-ChR2-YFP-FRT mice (1mg tamoxifen at P13); and (N) Adra2CreER; Calca-Flp; R26FSF-LSL-tdTomato mice (4mg tamoxifen at P14 and P16). All scale bars are 100 μm unless otherwise specified. Animals were sacrificed at 3–6 weeks.
O: ENS labeling quantification (percent of myenteric cell bodies labeled) across the different mouse lines. At least 3 separate images were quantified from at least N = 2 animals.
P: SNS labeling quantification (percent of sympathetic cell bodies labeled) across the different mouse lines. Images quantified from at least N = 2 separate sympathetic ganglia.
Q: In situ hybridization in a wild-type L6 DRG showing that the TrkB+, TH+, and Calca+ populations are nonoverlapping.
R: In situ hybridization in a wild-type L6 DRG showing that the three colon-innervating CGRP+ subtypes (Bmprb1b+, Sstr2+, and Adra2a+) populations are minimally overlapping.
S: Quantification of the percent overlap between the different colon-innervating populations based on ISH in L6-S1 DRGs. At least 6 separate DRGs were quantified from N = 3 animals.
A: Bmpr1bCreER targeting vector schematic.
B: DRG in situ hybridization in Bmpr1bCreER; Calca-Flp; R26FSF-LSL-mCitrine-ReaChR mice treated with 3 mg tamoxifen at P21 and sacrificed at least fourteen days later.
A: Calcium traces from an example TrkB+ neuron.
B: Graph with distension thresholds of all TrkB+ responsive cells (data from N = 3 mice).
C: Calcium traces showing response to colon distension or skin pinch in an example TrkB+ neuron before and after intra-colonic lidocaine.
D: Maximum fluorescence intensity in the 10 seconds prior and 1 minute during intracolonic balloon distension (120mmHg) before or after intracolonic lidocaine in TrkB+ cells (each point respresents an individual neuron; N = 3 animals, 2 TrkBCreER; R26LSL-GCaMP6s mice and 1 TrkBCreER; R26LSL-GCaMP6s; Piezo2fl/+ mouse).
E: Maximum fluorescence intensity in the 10 seconds prior and during back hairy skin pinch before or after intracolonic lidocaine in TrkB+ neurons (each point represents an individual neuron; N = 3 animals, 2 TrkBCreER; R26LSL-GCaMP6s mice and 1 TrkBCreER; R26LSL-GCaMP6s; Piezo2fl/+ mouse).
F: Calcium traces depicting colon distension responses of an example TH+ neuron.
G: Graph with distension thresholds of all TH+ responsive neurons recorded (data from N = 8 mice).
H: Calcium traces from an example CGRP+ neuron.
I: Graph with distension thresholds of all CGRP+ responsive neurons (data from N = 7 mice).
J: Calcium traces from an example Bmpr1b+ neuron.
K: Graph with distension thresholds of all Bmpr1b+ responsive neurons (data from N = 7 mice).
L: Calcium traces from an example Sstr2+ neuron.
M: Graph with distension thresholds of all Sstr2+ responsive neurons (data from N = 7 mice).
N: In vivo DRG calcium imaging responses to colon distension and intracolonic capsaicin in Bmpr1bCre; R26LSL-GCaMP6s (N = 2) animals.
O: Table summarizing key genes expressed in each of the DRG subtypes that innervate the colon, based on DRG single cell RNA sequencing13. X indicates expression; low indicates low expression; - indicates not expressed.
A: Raw action potential (AP) waveforms of each DRG afferent subtype (mean +/− SEM).
B: Normalized AP waveforms of each DRG afferent subtype (mean +/− SEM).
C: Example autocorrelograms of each DRG afferent subtype.
D: Raster plot with all opto-tagged units, organized by genotype. TrkB+ units in green (n = 7 cells from N = 6 mice); TH+ units in red (n = 3 cells from N = 8 mice); Bmpr1b+ units in black (n = 9 cells from N = 5 mice); and Sstr2+ units in blue (n = 5 cells from N = 5 mice).
E: Firing rates of each DRG afferent subtype during the on, sustained (sus), and off periods at each colonic distension force. Repeated-measures ANOVAs were performed across distension periods and comparisons between low (30mmHg) and high (120mmHg) forces are plotted. Statistical tests were not performed for the TH+ afferents due to low sample size. See Table S1 for additional statistical details.
A-C: Behavioral responses (pupil dilation (A), movement (B), and vocalizations (C)) to skin (back hindpaw and leg) optogenetic stimulation in wild-type mice (N = 3), Phox2B-Flp; R26FSF-mCitrine-ReaChR mice (N = 6), Calca-Flp; R26FSF-ReaChR or CalcaCreER; R26FRT-LSL-ChR2-YFP-FRT (1mg tamoxifen at P12) mice (N = 8), Bmpr1bCre; Calca-Flp; R26FSF-LSL-mCitrine-ReaChR mice (N = 6), Sstr2CreER; Calca-Flp; R26FSF-LSL-mCitrine-ReaChR or Sstr2CreER; R26LSL-mCitrine-ReaChR mice (N = 6; 2mg tamoxifen at P21), TrkBCreER; AdvillinFlp; R26FSF-LSL-mCitrine-ReaChR mice (N = 5; 0.5mg tamoxifen at P5), and THCreER; Phox2B-Flp; R26FRT-LSL-ChR2-YFP-FRT mice (N = 3; 1mg tamoxifen at P9). All comparisons are paired t tests except the accelerometer data for Sstr2CreER; Calca-Flp; R26FSF-LSL-mCitrine-ReaChR mice and and THCreER; Phox2B-Flp; R26FRT-LSL-ChR2-YFP-FRT mice and vocalization data for Calca-Flp; R26FSF-ReaChR or CalcaCreER; R26FRT-LSL-ChR2-YFP-FRT mice, Sstr2CreER; Calca-Flp; R26FSF-LSL-mCitrine-ReaChR or Sstr2CreER; R26LSL-mCitrine-ReaChR mice, and TrkBCreER; AdvillinFlp; R26FSF-LSL-mCitrine-ReaChR mice, which were analyzed using the Wilcoxon matched-pairs signed rank test.
D: Schematic of the targeting construct for the R26FSF-LSL-Caspase3 mice.
E: DRG in situ hybridization in control and Bmpr1b-ablated animals.
F-G: Quantification of Piezo2 deletion efficiency based on DRG in situ hybridization in control and Bmpr1bCre; Piezo2fl/fl or Bmpr1bCre; R26LSL-GCaMP6s; Piezo2fl/fl mice (comparisons are unpaired t tests).
H: Mean calcium activation (ΔF/F) during high threshold colon distension (120 mmHg) in responsive cells from control and Bmpr1bCre; R26LSL-GCaMP6s; Piezo2fl/fl DRGs (analyzed using the Mann-Whitney test). Note that the control neuron measurements are data that is replotted from Figure 4.
I: In vivo DRG calcium imaging responses to colon distension in Bmpr1bCre; R26LSL-GCaMP6s; Piezo2fl/fl (N = 7) animals.
A-B: Behavioral response to intracolonic mustard oil in control and CGRP-ablated (AdvillinCre; CalcaLSL-hDTR) animals. All comparisons are unpaired t tests (N = 17 for controls, vehicle treated; N = 16 for controls, mustard oil treated; N = 8 for CGRP-ablated, vehicle treated; N = 7 for CGRP-ablated, mustard oil treated).
C: Percent CGRP ablation in CGRP-ablated (AdvillinCre; CalcaLSL-hDTR) and Bmpr1b-ablated (Bmpr1bCre; CalcaLSL-hDTR). Each point represents one animal, which was calculated from the average of 5 DRGs per animal. For CGRP-ablated experiments, only animals with >75% CGRP ablation were included in analyses (animals in Figure S7A–B). For Bmpr1b-ablated experiments, only animals > 22% CGRP ablation (correlated to > 75% Bmpr1b ablation) were included in analyses (animals in Figure 6D–F, 7A, E–G).
D: ISH to look at levels of TrpV1 expression in Bmpr1b+ DRG neurons in control and DSS treated animals.
E: Quantification of TrpV1 expression from ISH in in Bmpr1b+ DRG neurons in control and DSS treated animals (79 cells from n = 6 DRG from N = 3 animals for control and 55 cells from n = 6 DRG from N = 3 animals for DSS-treated; compared using a Mann Whitney test).
Key Resource Table.
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Antibodies | ||
| Goat polyclonal anti-mCherry | Sicgen | Cat# AB0040; RRID:AB_2333093 |
| Goat polyclonal anti-GFP | US Biological Life Sciences | Cat# G8965-01E |
| Chicken polyclonal anti-GFP | Aves Labs | Cat# GFP-1020; RRID:AB_10000240 |
| Rabbit monoclonal anti-dsRed | Clontech | Cat# 632496; RRID:AB_10013483 |
| Rabbit polyclonal anti-CGRP | Immunostar | Cat# 24112; RRID:AB_572217 |
| Chicken polyclonal anti-NF200 | Aves Labs | Cat# NFH; RRID:AB_2313552 |
| Rabbit polyclonal anti-TUJ1 | Biolegend | Cat# 802001; RRID:AB_2564645 |
| Sheep polyclonal anti-TH | Sigma | Cat# AB1542; RRID:AB_90755 |
| Rabbit polyclonal anti-TH | Sigma | Cat# AB152 |
| Mouse monoclonal anti-NeuN | Sigma | Cat# MAB377; RRID:AB_2298772 |
| Donkey anti-Goat IgG Alexa Fluor 546 | Thermo Fisher Scientific | Cat# A-11056; RRID:AB_2534103 |
| Donkey anti-Rabbit IgG Alexa Fluor 488 | Thermo Fisher Scientific | Cat# A-21206; RRID:AB_2535792 |
| Donkey anti-Chicken IgG Alexa Fluor 488 | Jackson ImmunoResearch Labs | Cat# 703-545-155; RRID:AB_2340375 |
| Donkey anti-Goat IgG Alexa Fluor 488 | Thermo Fisher Scientific | Cat# A-11055; RRID:AB_2534102 |
| Donkey anti-Rabbit IgG Alexa Fluor 546 | Thermo Fisher Scientific | Cat# A-10040; RRID:AB_2534016 |
| Goat anti-Mouse IgG1 Alexa Fluor 647 | Thermo Fisher Scientific | Cat #A-21240; RRID:AB_2535809 |
| Bacterial and virus strains | ||
| AAV2-retro-hSyn-FlpO | Boston Children’s viral core | N/A |
| Chemicals, peptides, and recombinant proteins | ||
| Tamoxifen | Sigma | Cat# T5648-1g |
| Sunflower seed oil | Sigma | Cat# S5007 |
| Urethane | Sigma | Cat# U2500 |
| Isofluorane | Henry Schein | Cat# 029405 |
| Normal Goat Serum (NGS) | Vector Labs | Cat# S-1000 |
| Normal Donkey Serum (NDS) | Jackson Immuno | Cat# 005-000-121 |
| Mustard oil (Allyl isothiocyanate) | Sigma-Aldrich | Cat# 377430-5G |
| Capsaicin | Sigma-Aldrich | Cat# M2028 |
| Dextran sodium sulfate (DSS) | Fisher | Cat# MP216011025 |
| Diphtheria toxin (DTX) | Sigma | Cat# D0564 |
| Paraformaldehyde, reagent grade, crystalline | Millipore Sigma | Cat# P6148-500G |
| Critical commercial assays | ||
| RNAscope Fluorescent Multiplex Assay | ACD Bio | Cat# 320850 |
| RNAscope Protease III & Protease IV Reagents | ACD Bio | Cat# 322340 |
| RNAscope Probe Diluent | ACD Bio | Cat# 300041 |
| Experimental models: Organisms/Strains | ||
| Mouse: C57Bl/6J | The Jackson Laboratory | Stock No. 000664; RRID:IMSR_JAX:000664 |
| Mouse: Advillin-CreERT2 | The Jackson Laboratory | Stock No. 032027; RRID:IMSR_JAX:032027 |
| Mouse: AdvillinCre | The Jackson Laboratory | Stock No. 032536; RRID:IMSR_JAX:032536 |
| Mouse: Piezo2flox | The Jackson Laboratory | Stock No. 027720; RRID:IMSR_JAX:027720 |
| Mouse: Phox2B-Flp | The Jackson Laboratory | Stock No. 022407; RRID:IMSR_JAX:022407 |
| Mouse: Cdx2-Cre | The Jackson Laboratory | Stock No. 009352; RRID:IMSR_JAX:009352 |
| Mouse: MrgprdCreER | The Jackson Laboratory | Stock No. 031286; RRID:IMSR_JAX:031286 |
| Mouse: R26FSF-LSL-tdTomato (Ai65) | The Jackson Laboratory | Stock No. 021875; RRID:IMSR_JAX:021875 |
| Mouse: R26LSL-tdTomato (Ai14) | The Jackson Laboratory | Stock No. 007914; RRID:IMSR_JAX:007914 |
| Mouse: R26FSF-tdTomato (derived from Ai65) | The Jackson Laboratory | Stock No. 021875; RRID:IMSR_JAX:021875 |
| Mouse: R26FRT-LSL-ChR2-YFP-FRT (Ai32) | The Jackson Laboratory | Stock No. 012569; RRID:IMSR_JAX:012569 |
| Mouse: R26FSF-LSL-ReaChR | The Jackson Laboratory | Stock No. 024846; RRID:IMSR_JAX:024846 |
| Mouse: R26FRT-LSL-tdTomato-FRT (Ai9) | The Jackson Laboratory | Stock No. 007909; RRID:IMSR_JAX:007909 |
| Mouse: R26LSL-GCaMP6s (Ai96) | The Jackson Laboratory | Stock No. 028866; RRID:IMSR_JAX:028866 |
| Mouse: Igs7LSL-GCaMP6f (Ai148) | The Jackson Laboratory | Stock No. 030328; RRID:IMSR_JAX:030328 |
| Mouse: AdvillinFlp | (Choi et al, 2020)62 | N/A |
| Mouse: THCreER | The Jackson Laboratory | Stock No. 025614; RRID:IMSR_JAX:025614 |
| Mouse: Calca-Flp | (Choi et al, 2020)62 | N/A |
| Mouse: Bmpr1bMCre | (Sharma et al, 2020)13 | N/A |
| Mouse: Sstr2CreER | (Qi et al, 2023)43 | N/A |
| Mouse: Smr2Cre | (Qi et al, 2023)43 | N/A |
| Mouse: Adra2aCreER | (Qi et al, 2023)43 | N/A |
| Mouse: TrkBCreER | The Jackson Laboratory | Stock No. 027214; MGI:5616440 |
| Mouse: CalcaCreER | (Song et al, 2012)63 | N/A |
| Mouse: CalcaLox-GFP-STOP-Lox-hDTR | (McCoy et al, 2012)36 | N/A |
| Mouse: Calca-GFP | MMRRC | Stock No. 011187-UCD |
| Mouse: Mrgpra3-Cre | (Han et al, 2013)64 | N/A |
| Mouse: Mrgprb4CreER | (Vrontou et al, 2013)65 | N/A |
| Mouse: PIRTCre | (Patil et al, 2019)47 | N/A |
| Oligonucleotides | ||
| Mm-Piezo2 E43–45 | ACD Bio | Cat#: 439971 |
| Mm-Bmpr1b | ACD Bio | Cat#: 533941 |
| Mm-Calca TV2-TV3 | ACD Bio | Cat#: 420361 |
| Mm-Sstr2 | ACD Bio | Cat#: 437681 |
| Mm-Adra2a | ACD Bio | Cat#: 425341 |
| Mm-TrkB (NTRK2) | ACD Bio | Cat#: 423611 |
| Mm-TH | ACD Bio | Cat#: 317621 |
| Mm-Trpv1 | ACD Bio | Cat#: 313338 |
| Hs-Bmpr1b | ACD Bio | Cat#: 468031 |
| Hs-Calca | ACD Bio | Cat#: 605551 |
| GCaMP6s | ACD Bio | Cat#: 557091 |
| tdTomato | ACD Bio | Cat#: 317041 |
| Software and Algorithms | ||
| ImageJ/FIJI | NIH | https://ImageJ.nih.gov/ij/; RRID:SCR_003070 |
| GraphPad-PRISM | GraphPad | N/A |
| JRCLUST | (Jun et al, 2017)66 | https://github.com/ JaneliaSciComp/JRCLUST |
| MATLAB | Mathworks | https://www.mathworks.com/products/MATLAB.html; RRID:SCR 001622 |
| Python | (Rossum et al, 1994)67 | https://www.python.org/ |
| Bonsai | (Lopes et al, 2015)68 | https://bonsai-rx.org/ |
| Other | ||
| Multielectrode arrays | Cambridge Neurotech | H4–32ch |
| LED probe | ThorLabs | 470 nm, 17.2 mW (Min) Fiber-Coupled LED |
| Video camera | Fisher Scientific | Digital USB 2.0 CMOS |
| Ultrasonic microphone | Bat Conservation and Management, Inc. | Pettersson M500 microphone |
| Accelerometer | SparkFun | ADXL335 |
| Glass rods | McMaster Carr | 1357T117 |
| Human L5 DRG | AnaBios | N/A |
Highlights:
Five genetic subtypes of DRG sensory afferents innervate the distal colon
These subtypes have diverse ending morphologies in the colon
Four subtypes have distinct physiologic response properties to colon distension
One subtype (Bmpr1b+) is hyper-reactive to colonic distension during inflammation
Acknowledgements
We thank P. Gorelik and M. DeLisle for technical assistance, J. Peng for input on human DRG ISH, and Ginty lab members for comments on the manuscript. We thank Caiying Guo and the Janelia Transgenic Core Facility for generating Bmpr1bCreER and R26FSF-LSL-Caspase3 mouse lines. We thank X. Dong for PIRTCre mice. This work was supported by NIH grants T32-GM007753 and 1-R25-AI147393-01 (R.L.W), DP2NS127278 (N.S.), NS097344 and AT011447 (D.D.G.), an NEI P30 Core Grant for Vision Research EY012196 (O.M.), Klingenstein-Simons Foundation (N.S.), Whitehall Foundation (N.S.), The Tan-Yang Center for Autism Research (D.D.G.), and the Lefler Center for Neurodegenerative Disorders (D.D.G.). D.D.G. is an HHMI investigator. This article is subject to HHMI’s Open Access to Publications policy. HHMI lab heads have previously granted a nonexclusive CC BY 4.0 license to the public and a sublicensable license to HHMI in their research articles. Pursuant to those licenses, the author-accepted manuscript of this article can be made freely available under a CC BY 4.0 license immediately upon publication.
Footnotes
Declaration of interests
The authors declare no competing interests.
Inclusion and diversity
We support inclusive, diverse, and equitable conduct of research.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Heitmann PT, Vollebregt PF, Knowles CH, Lunniss PJ, Dinning PG, and Scott SM (2021). Understanding the physiology of human defaecation and disorders of continence and evacuation. Nat Rev Gastroenterol Hepatol 18, 751–769. 10.1038/s41575-021-00487-5. [DOI] [PubMed] [Google Scholar]
- 2.Kim M, Heo G, and Kim SY (2022). Neural signalling of gut mechanosensation in ingestive and digestive processes. Nat Rev Neurosci 23, 135–156. 10.1038/s41583-021-00544-7. [DOI] [PubMed] [Google Scholar]
- 3.Gebhart GF, and Bielefeldt K (2016). Physiology of Visceral Pain. Compr Physiol 6, 1609–1633. 10.1002/cphy.c150049. [DOI] [PubMed] [Google Scholar]
- 4.Handler A, and Ginty DD (2021). The mechanosensory neurons of touch and their mechanisms of activation. Nat Rev Neurosci 22, 521–537. 10.1038/s41583-021-00489-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bai L, Lehnert BP, Liu J, Neubarth NL, Dickendesher TL, Nwe PH, Cassidy C, Woodbury CJ, and Ginty DD (2015). Genetic Identification of an Expansive Mechanoreceptor Sensitive to Skin Stroking. Cell 163, 1783–1795. 10.1016/j.cell.2015.11.060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Neubarth NL, Emanuel AJ, Liu Y, Springel MW, Handler A, Zhang Q, Lehnert BP, Guo C, Orefice LL, Abdelaziz A, et al. (2020). Meissner corpuscles and their spatially intermingled afferents underlie gentle touch perception. Science 368. 10.1126/science.abb2751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Rutlin M, Ho CY, Abraira VE, Cassidy C, Woodbury CJ, and Ginty DD (2014). The Cellular and Molecular Basis of Direction Selectivity of A delta-LTMRs. Cell 159, 1640–1651. 10.1016/j.cell.2014.11.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Li LS, Rutlin M, Abraira VE, Cassidy C, Kus L, Gong SC, Jankowski MP, Luo WQ, Heintz N, Koerber HR, et al. (2011). The Functional Organization of Cutaneous Low-Threshold Mechanosensory Neurons. Cell 147, 1615–1627. 10.1016/j.cell.2011.11.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Luo W, Enomoto H, Rice FL, Milbrandt J, and Ginty DD (2009). Molecular identification of rapidly adapting mechanoreceptors and their developmental dependence on ret signaling. Neuron 64, 841–856. 10.1016/j.neuron.2009.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lewin GR, and McMahon SB (1991). Physiological properties of primary sensory neurons appropriately and inappropriately innervating skeletal muscle in adult rats. J Neurophysiol 66, 1218–1231. 10.1152/jn.1991.66.4.1218. [DOI] [PubMed] [Google Scholar]
- 11.Ghitani N, Barik A, Szczot M, Thompson JH, Li C, Le Pichon CE, Krashes MJ, and Chesler AT (2017). Specialized Mechanosensory Nociceptors Mediating Rapid Responses to Hair Pull. Neuron 95, 944–954 e944. 10.1016/j.neuron.2017.07.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Perl ER (1968). Myelinated afferent fibres innervating the primate skin and their response to noxious stimuli. J Physiol 197, 593–615. 10.1113/jphysiol.1968.sp008576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Sharma N, Flaherty K, Lezgiyeva K, Wagner DE, Klein AM, and Ginty DD (2020). The emergence of transcriptional identity in somatosensory neurons. Nature 577, 392–398. 10.1038/s41586-019-1900-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Abraira VE, Kuehn ED, Chirila AM, Springel MW, Toliver AA, Zimmerman AL, Orefice LL, Boyle KA, Bai L, Song BJ, et al. (2017). The Cellular and Synaptic Architecture of the Mechanosensory Dorsal Horn. Cell 168, 295–310 e219. 10.1016/j.cell.2016.12.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Liu Y, Rutlin M, Huang S, Barrick CA, Wang F, Jones KR, Tessarollo L, and Ginty DD (2012). Sexually dimorphic BDNF signaling directs sensory innervation of the mammary gland. Science 338, 1357–1360. 10.1126/science.1228258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Brierley SM, Jones RC 3rd, Xu L, Gebhart GF, and Blackshaw LA (2005). Activation of splanchnic and pelvic colonic afferents by bradykinin in mice. Neurogastroenterol Motil 17, 854–862. 10.1111/j.1365-2982.2005.00710.x. [DOI] [PubMed] [Google Scholar]
- 17.Page AJ, Brierley SM, Martin CM, Martinez-Salgado C, Wemmie JA, Brennan TJ, Symonds E, Omari T, Lewin GR, Welsh MJ, and Blackshaw LA (2004). The ion channel ASIC1 contributes to visceral but not cutaneous mechanoreceptor function. Gastroenterology 127, 1739–1747. 10.1053/j.gastro.2004.08.061. [DOI] [PubMed] [Google Scholar]
- 18.Castro J, Harrington AM, Garcia-Caraballo S, Maddern J, Grundy L, Zhang J, Page G, Miller PE, Craik DJ, Adams DJ, and Brierley SM (2017). alpha-Conotoxin Vc1.1 inhibits human dorsal root ganglion neuroexcitability and mouse colonic nociception via GABAB receptors. Gut 66, 1083–1094. 10.1136/gutjnl-2015-310971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Brierley SM, Hibberd TJ, and Spencer NJ (2018). Spinal Afferent Innervation of the Colon and Rectum. Front Cell Neurosci 12, 467. 10.3389/fncel.2018.00467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lynn PA, Olsson C, Zagorodnyuk V, Costa M, and Brookes SJ (2003). Rectal intraganglionic laminar endings are transduction sites of extrinsic mechanoreceptors in the guinea pig rectum. Gastroenterology 125, 786–794. [DOI] [PubMed] [Google Scholar]
- 21.Spencer NJ, Kerrin A, Zagorodnyuk VP, Hennig GW, Muto M, Brookes SJ, and McDonnell O (2008). Identification of functional intramuscular rectal mechanoreceptors in aganglionic rectal smooth muscle from piebald lethal mice. Am J Physiol Gastrointest Liver Physiol 294, G855–867. 10.1152/ajpgi.00502.2007. [DOI] [PubMed] [Google Scholar]
- 22.Song X, Chen BN, Zagorodnyuk VP, Lynn PA, Blackshaw LA, Grundy D, Brunsden AM, Costa M, and Brookes SJ (2009). Identification of medium/high-threshold extrinsic mechanosensitive afferent nerves to the gastrointestinal tract. Gastroenterology 137, 274–284, 284 e271. 10.1053/j.gastro.2009.02.061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Humenick A, Chen BN, Wiklendt L, Spencer NJ, Zagorodnyuk VP, Dinning PG, Costa M, and Brookes SJ (2015). Activation of intestinal spinal afferent endings by changes in intra-mesenteric arterial pressure. J Physiol 593, 3693–3709. 10.1113/JP270378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hibberd TJ, Kestell GR, Kyloh MA, Brookes SJ, Wattchow DA, and Spencer NJ (2016). Identification of different functional types of spinal afferent neurons innervating the mouse large intestine using a novel CGRPalpha transgenic reporter mouse. Am J Physiol Gastrointest Liver Physiol 310, G561–573. 10.1152/ajpgi.00462.2015. [DOI] [PubMed] [Google Scholar]
- 25.Spencer NJ, Kyloh M, and Duffield M (2014). Identification of different types of spinal afferent nerve endings that encode noxious and innocuous stimuli in the large intestine using a novel anterograde tracing technique. PLoS One 9, e112466. 10.1371/journal.pone.0112466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Schuster DJ, Dykstra JA, Riedl MS, Kitto KF, Honda CN, McIvor RS, Fairbanks CA, and Vulchanova L (2013). Visualization of spinal afferent innervation in the mouse colon by AAV8-mediated GFP expression. Neurogastroenterol Motil 25, e89–100. 10.1111/nmo.12057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hockley JRF, Taylor TS, Callejo G, Wilbrey AL, Gutteridge A, Bach K, Winchester WJ, Bulmer DC, McMurray G, and Smith ESJ (2018). Single-cell RNAseq reveals seven classes of colonic sensory neuron. Gut. 10.1136/gutjnl-2017-315631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Meerschaert KA, Adelman PC, Friedman RL, Albers KM, Koerber HR, and Davis BM (2020). Unique Molecular Characteristics of Visceral Afferents Arising from Different Levels of the Neuraxis: Location of Afferent Somata Predicts Function and Stimulus Detection Modalities. J Neurosci 40, 7216–7228. 10.1523/JNEUROSCI.1426-20.2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Smith-Anttila CJA, Mason EA, Wells CA, Aronow BJ, Osborne PB, and Keast JR (2020). Identification of a Sacral, Visceral Sensory Transcriptome in Embryonic and Adult Mice. eNeuro 7. 10.1523/ENEURO.0397-19.2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Feng B, and Gebhart GF (2011). Characterization of silent afferents in the pelvic and splanchnic innervations of the mouse colorectum. Am J Physiol Gastrointest Liver Physiol 300, G170–180. 10.1152/ajpgi.00406.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Nadelhaft I, and Vera PL (1991). Neurons labelled after the application of tracer to the distal stump of the transected hypogastric nerve in the rat. J Auton Nerv Syst 36, 87–96. 10.1016/0165-1838(91)90104-b. [DOI] [PubMed] [Google Scholar]
- 32.Sengupta JN, and Gebhart GF (1994). Characterization of mechanosensitive pelvic nerve afferent fibers innervating the colon of the rat. J Neurophysiol 71, 2046–2060. 10.1152/jn.1994.71.6.2046. [DOI] [PubMed] [Google Scholar]
- 33.Shea VK, Cai R, Crepps B, Mason JL, and Perl ER (2000). Sensory fibers of the pelvic nerve innervating the Rat’s urinary bladder. J Neurophysiol 84, 1924–1933. 10.1152/jn.2000.84.4.1924. [DOI] [PubMed] [Google Scholar]
- 34.McCarthy PW, and Lawson SN (1990). Cell type and conduction velocity of rat primary sensory neurons with calcitonin gene-related peptide-like immunoreactivity. Neuroscience 34, 623–632. 10.1016/0306-4522(90)90169-5. [DOI] [PubMed] [Google Scholar]
- 35.Basbaum AI, Bautista DM, Scherrer G, and Julius D (2009). Cellular and molecular mechanisms of pain. Cell 139, 267–284. 10.1016/j.cell.2009.09.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.McCoy ES, Taylor-Blake B, and Zylka MJ (2012). CGRPalpha-expressing sensory neurons respond to stimuli that evoke sensations of pain and itch. PLoS One 7, e36355. 10.1371/journal.pone.0036355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lawson SN, Crepps B, and Perl ER (2002). Calcitonin gene-related peptide immunoreactivity and afferent receptive properties of dorsal root ganglion neurones in guinea-pigs. J Physiol 540, 989–1002. 10.1113/jphysiol.2001.013086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Feng B, La JH, Tanaka T, Schwartz ES, McMurray TP, and Gebhart GF (2012). Altered colorectal afferent function associated with TNBS-induced visceral hypersensitivity in mice. Am J Physiol Gastrointest Liver Physiol 303, G817–824. 10.1152/ajpgi.00257.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Hughes PA, Brierley SM, Martin CM, Brookes SJ, Linden DR, and Blackshaw LA (2009). Post-inflammatory colonic afferent sensitisation: different subtypes, different pathways and different time courses. Gut 58, 1333–1341. 10.1136/gut.2008.170811. [DOI] [PubMed] [Google Scholar]
- 40.Gao X, Han S, Huang Q, He SQ, Ford NC, Zheng Q, Chen Z, Yu S, Dong X, and Guan Y (2021). Calcium imaging in population of dorsal root ganglion neurons unravels novel mechanisms of visceral pain sensitization and referred somatic hypersensitivity. Pain 162, 1068–1081. 10.1097/j.pain.0000000000002096. [DOI] [PubMed] [Google Scholar]
- 41.Usoskin D, Furlan A, Islam S, Abdo H, Lonnerberg P, Lou D, Hjerling-Leffler J, Haeggstrom J, Kharchenko O, Kharchenko PV, et al. (2015). Unbiased classification of sensory neuron types by large-scale single-cell RNA sequencing. Nat Neurosci 18, 145–153. 10.1038/nn.3881. [DOI] [PubMed] [Google Scholar]
- 42.Zeisel A, Hochgerner H, Lonnerberg P, Johnsson A, Memic F, van der Zwan J, Haring M, Braun E, Borm LE, La Manno G, et al. (2018). Molecular Architecture of the Mouse Nervous System. Cell 174, 999–1014 e1022. 10.1016/j.cell.2018.06.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Qi L, Iskols M, Shi D, Reddy P, Walker C, Lezgiyeva K, Voisin T, Pawlak M, Kuchroo VK, Chiu I, et al. (2023). A DRG genetic toolkit reveals molecular, morphological, and functional diversity of somatosensory neuron subtypes. bioRxiv, 2023.2004.2022.537932. 10.1101/2023.04.22.537932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Furness JB, Poole DP, Cho HJ, Callaghan BP, and Rivera LR (2016). The innervation of the gastrointestinal tract. In Yamada’s Textbook of Gastroenterology, Sixth Edition, (Wiley John & Sons Ltd), pp. 239–258. [Google Scholar]
- 45.Zhou X, Wang L, Hasegawa H, Amin P, Han BX, Kaneko S, He Y, and Wang F (2010). Deletion of PIK3C3/Vps34 in sensory neurons causes rapid neurodegeneration by disrupting the endosomal but not the autophagic pathway. Proc Natl Acad Sci U S A 107, 9424–9429. 10.1073/pnas.0914725107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Lau J, Minett MS, Zhao J, Dennehy U, Wang F, Wood JN, and Bogdanov YD (2011). Temporal control of gene deletion in sensory ganglia using a tamoxifen-inducible Advillin-Cre-ERT2 recombinase mouse. Mol Pain 7, 100. 10.1186/1744-8069-7-100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Patil MJ, Meeker S, Bautista D, Dong X, and Undem BJ (2019). Sphingosine-1-phosphate activates mouse vagal airway afferent C-fibres via S1PR3 receptors. J Physiol 597, 2007–2019. 10.1113/JP277521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Hinoi T, Akyol A, Theisen BK, Ferguson DO, Greenson JK, Williams BO, Cho KR, and Fearon ER (2007). Mouse model of colonic adenoma-carcinoma progression based on somatic Apc inactivation. Cancer Res 67, 9721–9730. 10.1158/0008-5472.CAN-07-2735. [DOI] [PubMed] [Google Scholar]
- 49.Hirsch MR, d’Autreaux F, Dymecki SM, Brunet JF, and Goridis C (2013). A Phox2b::FLPo transgenic mouse line suitable for intersectional genetics. Genesis 51, 506–514. 10.1002/dvg.22393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Guo T, Bian Z, Trocki K, Chen L, Zheng G, and Feng B (2019). Optical recording reveals topological distribution of functionally classified colorectal afferent neurons in intact lumbosacral DRG. Physiol Rep 7, e14097. 10.14814/phy2.14097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Brierley SM, Jones RC 3rd, Gebhart GF, and Blackshaw LA (2004). Splanchnic and pelvic mechanosensory afferents signal different qualities of colonic stimuli in mice. Gastroenterology 127, 166–178. [DOI] [PubMed] [Google Scholar]
- 52.Lynn PA, and Blackshaw LA (1999). In vitro recordings of afferent fibres with receptive fields in the serosa, muscle and mucosa of rat colon. J Physiol 518, 271–282. 10.1111/j.1469-7793.1999.0271r.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Ranade SS, Woo SH, Dubin AE, Moshourab RA, Wetzel C, Petrus M, Mathur J, Begay V, Coste B, Mainquist J, et al. (2014). Piezo2 is the major transducer of mechanical forces for touch sensation in mice. Nature 516, 121–125. 10.1038/nature13980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Zylka MJ, Rice FL, and Anderson DJ (2005). Topographically distinct epidermal nociceptive circuits revealed by axonal tracers targeted to Mrgprd. Neuron 45, 17–25. 10.1016/j.neuron.2004.12.015. [DOI] [PubMed] [Google Scholar]
- 55.Dussor G, Zylka MJ, Anderson DJ, and McCleskey EW (2008). Cutaneous sensory neurons expressing the Mrgprd receptor sense extracellular ATP and are putative nociceptors. J Neurophysiol 99, 1581–1589. 10.1152/jn.01396.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Kupari J, Haring M, Agirre E, Castelo-Branco G, and Ernfors P (2019). An Atlas of Vagal Sensory Neurons and Their Molecular Specialization. Cell Rep 27, 2508–2523 e2504. 10.1016/j.celrep.2019.04.096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Murthy SE, Loud MC, Daou I, Marshall KL, Schwaller F, Kuhnemund J, Francisco AG, Keenan WT, Dubin AE, Lewin GR, and Patapoutian A (2018). The mechanosensitive ion channel Piezo2 mediates sensitivity to mechanical pain in mice. Sci Transl Med 10. 10.1126/scitranslmed.aat9897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Someya S, Nagao M, Shibata C, Tanaka N, Sasaki H, Kikuchi D, Miyachi T, Naitoh T, and Unno M (2015). Intracolonic Administration of the TRPA1 Agonist Allyl Isothiocyanate Stimulates Colonic Motility and Defecation in Conscious Dogs. J Gastrointest Surg 19, 1342–1349. 10.1007/s11605-015-2813-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Yu H, Usoskin D, Nagi SS, Hu Y, Kupari J, Bouchatta O, Cranfill SL, Su Y, Lv Y, Song H, et al. (2023). Single-Soma Deep RNA sequencing of Human DRG Neurons Reveals Novel Molecular and Cellular Mechanisms Underlying Somatosensation. bioRxiv. 10.1101/2023.03.17.533207. [DOI] [Google Scholar]
- 60.Cavanaugh DJ, Lee H, Lo L, Shields SD, Zylka MJ, Basbaum AI, and Anderson DJ (2009). Distinct subsets of unmyelinated primary sensory fibers mediate behavioral responses to noxious thermal and mechanical stimuli. Proc Natl Acad Sci U S A 106, 9075–9080. 10.1073/pnas.0901507106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Servin-Vences MR, Lam RM, Koolen A, Wang Y, Saade DN, Loud M, Kacmaz H, Frausto S, Zhang Y, Beyder A, et al. (2023). PIEZO2 in somatosensory neurons controls gastrointestinal transit. Cell. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Choi S, Hachisuka J, Brett MA, Magee AR, Omori Y, Iqbal NU, Zhang D, DeLisle MM, Wolfson RL, Bai L, et al. (2020). Parallel ascending spinal pathways for affective touch and pain. Nature 587, 258–263. 10.1038/s41586-020-2860-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Song H, Yao E, Lin C, Gacayan R, Chen MH, and Chuang PT (2012). Functional characterization of pulmonary neuroendocrine cells in lung development, injury, and tumorigenesis. Proc Natl Acad Sci U S A 109, 17531–17536. 10.1073/pnas.1207238109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Han L, Ma C, Liu Q, Weng HJ, Cui Y, Tang Z, Kim Y, Nie H, Qu L, Patel KN, et al. (2013). A subpopulation of nociceptors specifically linked to itch. Nat Neurosci 16, 174–182. 10.1038/nn.3289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Vrontou S, Wong AM, Rau KK, Koerber HR, and Anderson DJ (2013). Genetic identification of C fibres that detect massage-like stroking of hairy skin in vivo. Nature 493, 669–673. 10.1038/nature11810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Jun JJ, Mitelut C, Lai C, Gratiy SL, Anastassiou CA, and Harris TD (2017). Real-time spike sorting platform for high-density extracellular probes with ground-truth validation and drift correction. BioRxiv, 101030. [Google Scholar]
- 67.Van Rossum G, and Drake FL Jr (1994). Python reference manual. [Google Scholar]
- 68.Lopes G, Bonacchi N, Frazao J, Neto JP, Atallah BV, Soares S, Moreira L, Matias S, Itskov PM, Correia PA, et al. (2015). Bonsai: an event-based framework for processing and controlling data streams. Front Neuroinform 9, 7. 10.3389/fninf.2015.00007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Chirila AM, Rankin G, Tseng SY, Emanuel AJ, Chavez-Martinez CL, Zhang D, Harvey CD, and Ginty DD (2022). Mechanoreceptor signal convergence and transformation in the dorsal horn flexibly shape a diversity of outputs to the brain. Cell. 10.1016/j.cell.2022.10.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Emanuel AJ, Lehnert BP, Panzeri S, Harvey CD, and Ginty DD (2021). Cortical responses to touch reflect subcortical integration of LTMR signals. Nature 600, 680–685. 10.1038/s41586-021-04094-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Dubbs A, Guevara J, and Yuste R (2016). moco: Fast Motion Correction for Calcium Imaging. Front Neuroinform 10, 6. 10.3389/fninf.2016.00006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Christianson JA, and Gebhart GF (2007). Assessment of colon sensitivity by luminal distension in mice. Nat Protoc 2, 2624–2631. 10.1038/nprot.2007.392. [DOI] [PubMed] [Google Scholar]
- 73.Chandrasekharan BP, Kolachala VL, Dalmasso G, Merlin D, Ravid K, Sitaraman SV, and Srinivasan S (2009). Adenosine 2B receptors (A(2B)AR) on enteric neurons regulate murine distal colonic motility. FASEB J 23, 2727–2734. 10.1096/fj.09-129544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.France M, Bhattarai Y, Galligan JJ, and Xu H (2012). Impaired propulsive motility in the distal but not proximal colon of BK channel beta1-subunit knockout mice. Neurogastroenterol Motil 24, e450–459. 10.1111/j.1365-2982.2012.01981.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Coates MD, Johnson AC, Greenwood-Van Meerveld B, and Mawe GM (2006). Effects of serotonin transporter inhibition on gastrointestinal motility and colonic sensitivity in the mouse. Neurogastroenterol Motil 18, 464–471. 10.1111/j.1365-2982.2006.00792.x. [DOI] [PubMed] [Google Scholar]
- 76.Laird JM, Martinez-Caro L, Garcia-Nicas E, and Cervero F (2001). A new model of visceral pain and referred hyperalgesia in the mouse. Pain 92, 335–342. [DOI] [PubMed] [Google Scholar]
- 77.Park YH, Kim N, Shim YK, Choi YJ, Nam RH, Choi YJ, Ham MH, Suh JH, Lee SM, Lee CM, et al. (2015). Adequate Dextran Sodium Sulfate-induced Colitis Model in Mice and Effective Outcome Measurement Method. J Cancer Prev 20, 260–267. 10.15430/JCP.2015.20.4.260. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
A-M: Skin and colon whole mount immunofluorescence or DRG, superior cervical ganglia (SCG), sympathetic chain, or spinal cord immunofluorescence in (A-C) Advillin-CreER; R26LSL-tdTomato mice treated with 1mg tamoxifen at P5; (D-F) Advillin-CreER; R26LSL-tdTomato mice treated with 0.5mg tamoxifen at P1; (G-H) Advillin-CreER; R26LSL-tdTomato mice treated with 3mg tamoxifen at E12.5; (I-J) AdvillinFlp; Cdx2-Cre; R26FSF-LSL-tdTomato; (K) PIRTCre; R26LSL-tdTomato; (L-M) Phox2B-Flp; R26FSF-ReaChR-mCitrine; or (N) Advillin-CreER; Phox2B-Flp; R26FRT-LSL-ChR2-YFP-FRT animals treated with 1mg tamoxifen P21–25. All scale bars are 100 μm unless otherwise specified.
O: Brainstem immunofluorescence in Advillin-CreER; Phox2B-Flp; R26FRT-LSL-ChR2-YFP-FRT treated with 1mg tamoxifen P21–25 (area postrema (AP), nucleus tractus solitarius (NTS), dorsal column nucleus (DCN), spinal trigeminal nucleus (SPV)). Scale bar is 500 μm. Animals were sacrificed at 3–6 weeks.
P: ENS labeling quantification (percent of myenteric cell bodies labeled) across the different mouse lines. 3 separate images were quantified from at least N = 2 animals.
Q: SNS labeling quantification (percent of sympathetic cell bodies labeled) across the different mouse lines. Images quantified from at least N = 2 separate sympathetic ganglia.
R: Quantification of the fiber labeling density at the level of the myenteric vs. submucosal plexus in the Advillin-CreER; Phox2B-Flp; R26FRT-LSL-ChR2-YFP-FRT animals. At least 3 separate images were quantified from N = 3 animals. Compared using an unpaired t test.
A-F: Skin and colon whole mount immunofluorescence or DRG and sympathetic chain immunofluorescence in (A) MrgprdCreER; Phox2B-Flp; R26FRT-LSL-tdTomato-FRT mice (1mg tamoxifen at P7); (B) TrkBCreER; AdvillinFlp; R26FSF-LSL-ReaChR-mCitrine mice (0.5mg tamoxifen at P5); (C) THCreER; Phox2B-Flp; R26FRT-LSL-tdTomato-FRT mice (0.5–1mg tamoxifen at P11–14); (D) Calca-GFP; (E) CalcaFlp; R26FSF-tdTomato; (F) CalcaLox-GFP-STOP-Lox-hDTR mice.
G: ENS labeling quantification (percent of myenteric cell bodies labeled) across the different Calca mouse lines. At least 2 separate images were quantified from at least N = 2 animals.
H-N: DRG and sympathetic chain immunofluorescence in (H-I) Smr2Cre; R26FRT-LSL-tdTomato-FRT mice; (J) Bmpr1bCre; AdvillinFlp; R26FSF-LSL-tdTomato mice; (K-M) Sstr2CreER; Phox2B-Flp; R26FRT-LSL-ChR2-YFP-FRT mice (1mg tamoxifen at P13); and (N) Adra2CreER; Calca-Flp; R26FSF-LSL-tdTomato mice (4mg tamoxifen at P14 and P16). All scale bars are 100 μm unless otherwise specified. Animals were sacrificed at 3–6 weeks.
O: ENS labeling quantification (percent of myenteric cell bodies labeled) across the different mouse lines. At least 3 separate images were quantified from at least N = 2 animals.
P: SNS labeling quantification (percent of sympathetic cell bodies labeled) across the different mouse lines. Images quantified from at least N = 2 separate sympathetic ganglia.
Q: In situ hybridization in a wild-type L6 DRG showing that the TrkB+, TH+, and Calca+ populations are nonoverlapping.
R: In situ hybridization in a wild-type L6 DRG showing that the three colon-innervating CGRP+ subtypes (Bmprb1b+, Sstr2+, and Adra2a+) populations are minimally overlapping.
S: Quantification of the percent overlap between the different colon-innervating populations based on ISH in L6-S1 DRGs. At least 6 separate DRGs were quantified from N = 3 animals.
A: Bmpr1bCreER targeting vector schematic.
B: DRG in situ hybridization in Bmpr1bCreER; Calca-Flp; R26FSF-LSL-mCitrine-ReaChR mice treated with 3 mg tamoxifen at P21 and sacrificed at least fourteen days later.
A: Calcium traces from an example TrkB+ neuron.
B: Graph with distension thresholds of all TrkB+ responsive cells (data from N = 3 mice).
C: Calcium traces showing response to colon distension or skin pinch in an example TrkB+ neuron before and after intra-colonic lidocaine.
D: Maximum fluorescence intensity in the 10 seconds prior and 1 minute during intracolonic balloon distension (120mmHg) before or after intracolonic lidocaine in TrkB+ cells (each point respresents an individual neuron; N = 3 animals, 2 TrkBCreER; R26LSL-GCaMP6s mice and 1 TrkBCreER; R26LSL-GCaMP6s; Piezo2fl/+ mouse).
E: Maximum fluorescence intensity in the 10 seconds prior and during back hairy skin pinch before or after intracolonic lidocaine in TrkB+ neurons (each point represents an individual neuron; N = 3 animals, 2 TrkBCreER; R26LSL-GCaMP6s mice and 1 TrkBCreER; R26LSL-GCaMP6s; Piezo2fl/+ mouse).
F: Calcium traces depicting colon distension responses of an example TH+ neuron.
G: Graph with distension thresholds of all TH+ responsive neurons recorded (data from N = 8 mice).
H: Calcium traces from an example CGRP+ neuron.
I: Graph with distension thresholds of all CGRP+ responsive neurons (data from N = 7 mice).
J: Calcium traces from an example Bmpr1b+ neuron.
K: Graph with distension thresholds of all Bmpr1b+ responsive neurons (data from N = 7 mice).
L: Calcium traces from an example Sstr2+ neuron.
M: Graph with distension thresholds of all Sstr2+ responsive neurons (data from N = 7 mice).
N: In vivo DRG calcium imaging responses to colon distension and intracolonic capsaicin in Bmpr1bCre; R26LSL-GCaMP6s (N = 2) animals.
O: Table summarizing key genes expressed in each of the DRG subtypes that innervate the colon, based on DRG single cell RNA sequencing13. X indicates expression; low indicates low expression; - indicates not expressed.
A: Raw action potential (AP) waveforms of each DRG afferent subtype (mean +/− SEM).
B: Normalized AP waveforms of each DRG afferent subtype (mean +/− SEM).
C: Example autocorrelograms of each DRG afferent subtype.
D: Raster plot with all opto-tagged units, organized by genotype. TrkB+ units in green (n = 7 cells from N = 6 mice); TH+ units in red (n = 3 cells from N = 8 mice); Bmpr1b+ units in black (n = 9 cells from N = 5 mice); and Sstr2+ units in blue (n = 5 cells from N = 5 mice).
E: Firing rates of each DRG afferent subtype during the on, sustained (sus), and off periods at each colonic distension force. Repeated-measures ANOVAs were performed across distension periods and comparisons between low (30mmHg) and high (120mmHg) forces are plotted. Statistical tests were not performed for the TH+ afferents due to low sample size. See Table S1 for additional statistical details.
A-C: Behavioral responses (pupil dilation (A), movement (B), and vocalizations (C)) to skin (back hindpaw and leg) optogenetic stimulation in wild-type mice (N = 3), Phox2B-Flp; R26FSF-mCitrine-ReaChR mice (N = 6), Calca-Flp; R26FSF-ReaChR or CalcaCreER; R26FRT-LSL-ChR2-YFP-FRT (1mg tamoxifen at P12) mice (N = 8), Bmpr1bCre; Calca-Flp; R26FSF-LSL-mCitrine-ReaChR mice (N = 6), Sstr2CreER; Calca-Flp; R26FSF-LSL-mCitrine-ReaChR or Sstr2CreER; R26LSL-mCitrine-ReaChR mice (N = 6; 2mg tamoxifen at P21), TrkBCreER; AdvillinFlp; R26FSF-LSL-mCitrine-ReaChR mice (N = 5; 0.5mg tamoxifen at P5), and THCreER; Phox2B-Flp; R26FRT-LSL-ChR2-YFP-FRT mice (N = 3; 1mg tamoxifen at P9). All comparisons are paired t tests except the accelerometer data for Sstr2CreER; Calca-Flp; R26FSF-LSL-mCitrine-ReaChR mice and and THCreER; Phox2B-Flp; R26FRT-LSL-ChR2-YFP-FRT mice and vocalization data for Calca-Flp; R26FSF-ReaChR or CalcaCreER; R26FRT-LSL-ChR2-YFP-FRT mice, Sstr2CreER; Calca-Flp; R26FSF-LSL-mCitrine-ReaChR or Sstr2CreER; R26LSL-mCitrine-ReaChR mice, and TrkBCreER; AdvillinFlp; R26FSF-LSL-mCitrine-ReaChR mice, which were analyzed using the Wilcoxon matched-pairs signed rank test.
D: Schematic of the targeting construct for the R26FSF-LSL-Caspase3 mice.
E: DRG in situ hybridization in control and Bmpr1b-ablated animals.
F-G: Quantification of Piezo2 deletion efficiency based on DRG in situ hybridization in control and Bmpr1bCre; Piezo2fl/fl or Bmpr1bCre; R26LSL-GCaMP6s; Piezo2fl/fl mice (comparisons are unpaired t tests).
H: Mean calcium activation (ΔF/F) during high threshold colon distension (120 mmHg) in responsive cells from control and Bmpr1bCre; R26LSL-GCaMP6s; Piezo2fl/fl DRGs (analyzed using the Mann-Whitney test). Note that the control neuron measurements are data that is replotted from Figure 4.
I: In vivo DRG calcium imaging responses to colon distension in Bmpr1bCre; R26LSL-GCaMP6s; Piezo2fl/fl (N = 7) animals.
A-B: Behavioral response to intracolonic mustard oil in control and CGRP-ablated (AdvillinCre; CalcaLSL-hDTR) animals. All comparisons are unpaired t tests (N = 17 for controls, vehicle treated; N = 16 for controls, mustard oil treated; N = 8 for CGRP-ablated, vehicle treated; N = 7 for CGRP-ablated, mustard oil treated).
C: Percent CGRP ablation in CGRP-ablated (AdvillinCre; CalcaLSL-hDTR) and Bmpr1b-ablated (Bmpr1bCre; CalcaLSL-hDTR). Each point represents one animal, which was calculated from the average of 5 DRGs per animal. For CGRP-ablated experiments, only animals with >75% CGRP ablation were included in analyses (animals in Figure S7A–B). For Bmpr1b-ablated experiments, only animals > 22% CGRP ablation (correlated to > 75% Bmpr1b ablation) were included in analyses (animals in Figure 6D–F, 7A, E–G).
D: ISH to look at levels of TrpV1 expression in Bmpr1b+ DRG neurons in control and DSS treated animals.
E: Quantification of TrpV1 expression from ISH in in Bmpr1b+ DRG neurons in control and DSS treated animals (79 cells from n = 6 DRG from N = 3 animals for control and 55 cells from n = 6 DRG from N = 3 animals for DSS-treated; compared using a Mann Whitney test).
Data Availability Statement
All data reported in this study will be shared by the lead contact upon request.
All original code is available in the paper’s supplemental information and will be shared by the lead author upon request.
Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.