

Abstract
Multiple Myeloma (MM) remains an incurable plasma cell neoplasm. Although little is known about the etiology of MM, several metabolic risk factors such as obesity, diabetes mellitus, diet, and the human intestinal microbiome have been linked to the pathogenesis of MM. In this article, we provide a detailed review of dietary and microbiome factors involved in the pathogenesis of MM and their impact on outcomes. Concurrent with treatment advancements that have improved survival in MM, focused efforts are needed to reduce the burden of MM as well as improve MM specific and overall outcomes once MM is diagnosed. The findings presented in this review will provide a comprehensive guide on the evidence available to date of the impact of dietary and other lifestyle interventions on the gut microbiome and on MM incidence, outcomes, and quality of life. Data generated from such studies can help formulate evidence-based guidelines for healthcare providers to counsel individuals at risk such as those with Monoclonal Gammopathy of Undetermined Significance (MGUS) and Smoldering Multiple Myeloma (SMM) as well as MM survivors with respect to their dietary habits.
INTRODUCTION
Multiple myeloma (MM), the second most common hematologic malignancy, is characterized by neoplastic proliferation of plasma cells. The Surveillance, Epidemiology, and End Results Program estimates about 34,920 new MM cases were diagnosed in 2021 and, in 2019 an estimated 159,787 people were living with MM in the US [1]. Although MM remains incurable for the majority, treatments continue to improve longevity and a subset of patients may be cured. As MM patients live longer, it is increasingly important to address dietary and lifestyle factors that contribute to morbidity and mortality.
The Third Expert Report on diet, nutrition, physical activity, and cancer risk provides updated objective evidence for the role of diet and nutrition in cancer [2]. An online survey of 421 plasma cell disorder patients revealed that 82% of patients had dietary questions after diagnosis and 57% reported that their dietary questions were not addressed by their oncologists. Among the patients who received dietary guidance from their oncologists, 94% attempted to follow it reflecting the positive influence oncologists can have [3].
Dietary nutrient absorption and metabolism are facilitated by the gut microbiota. Our understanding of this ecosystem of >1013 microorganisms in health and disease has been significantly improved with the availability of next-generation sequencing approaches evaluating conserved 16 S rRNA regions to profile the microbial composition and shotgun metagenomics to study its function [4, 5]. The microbiome plays key roles in absorption and energy utilization from soluble fiber, fats, proteins, and vitamins [6]. Nutritional intake, in turn, directly impacts the human microbiome. In addition to participating in the absorption of nutrients, the microbiota also serves a role in immunologic stimuli and regulation of cytokine secretion in intestinal epithelial cells by activation of signal transduction pathways [5, 7]. There is emerging data on the influence of microbiota alterations in the development of certain malignancies through promotion of inflammation and other complex host-microbiome interactions [5].
In this review, we will discuss the available evidence for diet and microbiome as potential modifiers of natural history of MM and outline mechanisms by which these factors may influence the process of myelomagenesis or outcomes after diagnosis. In addition, this review highlights the gap in knowledge and care delivery in aspects of care that are highly relevant to patients.
OVERALL DIETARY PATTERNS AND RISK OF MM
Diet is a major modifiable risk factor for cancer and several studies have assessed the association between diet and MM [8–10]. Diet is greatly influenced by racial, socioeconomic, and other lifestyle factors, which can potentially confound various associations. Majority of the studies outlined below thus, adjusted for combinations of age, gender, race, BMI, total energy intake, physical activity, smoking and alcohol consumption.
Two studies evaluated associations between such dietary patterns and risk of MM. A pooled analysis of two large prospective studies (EPIC-Oxford and Oxford-Vegetarian), categorized participants into meat eaters, fish eaters, vegetarians, and vegans based on a food-frequency questionnaire. The analysis demonstrated significantly lower risk for cancer overall in fish eaters (Relative Risk, RR 0.88, 95% Confidence Interval, CI: 0.80–0.97), vegetarians (RR 0.89, 95% CI: 0.83–0.96) and vegans (RR 0.81, 95% CI: 0.66–0.98) compared to meat eaters. This difference was even more pronounced for MM in vegetarians and vegans (RR 0.23, 95% CI: 0.09–0.59) and in fish eaters (RR 0.80, 95% CI: 0.34–1.76) as compared to meat eaters [11] (Table 1A). The large prospective Nurses’ Health Study and Health Professionals Follow-up Study revealed a statistically significant 16% increase in MM risk in men with the Empirical Dietary Inflammatory Pattern (EDIP) (Hazard Ratio, HR 1.16, 95% CI: 1.02–1.32 per 1-SD increase) [12].
Table 1.
Association of Dietary Patterns with Multiple Myeloma.
| A | |||||
|---|---|---|---|---|---|
| Population | Meat eaters | Fish eaters | Vegetarians | Vegans | |
| Number | 32,491 | 8612 | 18,298 | 2246 | Key AJCN 2014 [11] |
| Cancer cases | 3275 | 520 | 1098 | 105 | |
| Relative risk reduction | 100 | 88 | 89 | 81 | |
| Multiple myeloma cases | 53 (0.2%) | 7 (0.08%) | 5 (0.03%) | ||
| Relative risk reduction | 100 | 80 | 23 | ||
| B | |||||
| Diet | Hazard Ratio (per 1 SD↑) | Confidence interval | P value | Study | |
| Multiple Myeloma in Males | |||||
| Unhealthy Dietary Patterns | |||||
| Empirical Dietary Inflammatory Pattern (EDIP) | 1.16 | 1.02–1.32 | Lee et al. JNCI Can Spec 2019 [12] | ||
| Multiple Myeloma Specific Mortality | |||||
| Healthy Dietary Patterns | |||||
| Alternate Healthy Eating Index-2010 (AHEI) | 0.76 | 0.67–0.87 | <0.001 | Lee et al. Int J Cancer 2020 [13] | |
| Alternate Mediterranean diet | 0.85 | 0.75–0.97 | 0.01 | ||
| Dietary Approaches to Stop Hypertension (DASH) diet | 0.85 | 0.76–0.95 | 0.006 | ||
| Prudent diet | 0.76 | 0.66–0.87 | <0.001 | ||
| Unhealthy Dietary Patterns | |||||
| Western diet | 1.24 | 1.07–1.44 | 0.005 | ||
| Empirical Dietary Index for Insulin Resistance (EDIR) | 1.16 | 1.02–1.33 | 0.03 | ||
| Empirical Dietary Index for Hyperinsulinemia (EDIH) | 1.17 | 1.01–1.35 | 0.03 | ||
A:Oxford Vegetarian study and EPIC-Oxford cohort and Multiple Myeloma by Key et al. B:Dietary Patterns and Multiple Myeloma by Lee et al.
A pooled prospective survival analyses of 423 MM patients from the NHS and HPFS cohorts (345 total deaths, 295 MM related deaths) demonstrated 15–24% lower MM-specific mortality for presumed healthy pre-diagnosis plant based dietary patterns (HR range 0.76–0.85 per 1-SD increase in scores). Simultaneously they also saw a 16–24% higher MM-specific mortality in unhealthy pre-diagnosis dietary patterns (HR range 1.16–1.24, per 1-SD increase in scores) [13]. These findings suggest that pre-diagnosis high insulinemic or inflammatory diets are associated with higher MM risk and increased risk of MM related death whereas plant-based dietary habits may reduce risk of development of MM and risk of death once MM develops (Table 1B). The strength of this study lies in the ability to evaluate multiple dietary patterns, and it is the largest dietary study to date in MM. However, enrollment of a primarily white health professional cohort, lack of post-diagnosis dietary data and limited clinical data regarding risk and treatments were some limitations. Additional studies are needed to confirm these findings in larger and more diverse populations.
Plant-based diets are high in soluble fiber with higher levels of bacteria from Bacteroidetes, Lactobacilli, Bifidobacteria and Clostridium clusters XIVa and IV [14]. The latter groups contain Eubacterium, Roseburia, Faecalibacterium, which degrade fiber and produce short-chain fatty acids (SCFAs), such as butyrate, acetate, and propionate [6, 15]. Western diets are lower in soluble fiber resulting in less SCFA production. SCFAs have important roles in the immune response, preventing inflammatory disease states, and are associated with improved MM outcomes [16, 17]. Several studies describing the microbiome of untreated MM patients have, however, offered conflicting observations regarding the role of SCFA producing bacteria which we discuss later in the review.
INDIVIDUAL DIETARY COMPONENTS AND RISK OF MM
Multiple studies have evaluated the impact of specific dietary factors on MM. We have summarized these associations as well as their relevant effects on the gut microbiome in Table 2 and potential mechanisms are illustrated in Fig. 1.
Table 2.
Overview of studies and risk of plasma cell disorders.
| A | ||||||||
|---|---|---|---|---|---|---|---|---|
| Study Cohort/Location | Study Design | N | Dietary Factor | Risk | Consumption patterns | Risk | 95% CI | Study |
| Age, Gene/Environment Susceptibility - Reykjavik study | Prospective Cohort | 5764 (575 MGUS) | Fruits | MGUS | Fruit ≥ 3/wk in adolescent period | OR 0.62 | 0.41–0.95 | Thordardottir et al. Plos One 2018 [18] |
| 5764 (575 MGUS) | Fruits | MGUS → MM | Fruit ≥ 3 wk; after 50 years (late life) | HR 0.34 | 0.13–0.89 | |||
| Institute of Hematology, University Clinical Center, Belgrade, Yugoslavia | Case-control | 100 MM; 100 controls | Fruits | MM | Fruit (4–7/wk) | OR 0.6 | 0.3–1.1 | Vlajinac et al. Neoplasma 2003 [20] |
| Yale Comprehensive Cancer Center’s Rapid Case Ascertainment Shared Resource and Connecticut Tumor Registry, Connecticut, USA | Case-control | 173 MM; 670 controls | Citrus Fruits | MM | 1/mo | OR: 1.0 | Hosgood et al. Cancer Causes Control 2007 [10] | |
| 1/wk | 0.6 | 0.4–1.0 | ||||||
| 2/wk | 0.7 | 0.4–1.3 | ||||||
| >2/wk | 0.9 | 0.5–1.5 | ||||||
| Cancer registries from the Georgia Center for Cancer Statistics, the Metropolitan Detroit Cancer Surveillance System and the New Jersey Cancer registry, USA | Case-control | 539 MM; 1989 controls | Citrus Fruits | MM | Quartiles of consumption: 1 | OR:1 | Brown et al. Cancer Causes Control 2001 [9] | |
| 2 | 1.3 | 1.0–1.8 | ||||||
| 3 | 1.4 | 1.1–2.0 | ||||||
| 4 | 1.4 | 1.0–1.9 | ||||||
| Yale Comprehensive Cancer Center’s Rapid Case Ascertainment Shared Resource and Connecticut Tumor Registry, Connecticut, USA | Case-control | 173 MM; 670 controls | Cruciferous Vegetables | MM | <1/ wk | OR 1 | Hosgood et al. Cancer Causes Control 2007 [10] | |
| 6/mo | 0.5 | 0.3–0.9 | ||||||
| 2/wk | 0.8 | 0.5–1.3 | ||||||
| > 2/wk | 0.5 | 0.3–0.8 | ||||||
| Tomatoes | MM | 2–3 month | OR:1 | |||||
| 1/ week | 0.6 | 04–0.9 | ||||||
| 2/week | 0.5 | 0.3–0.8 | ||||||
| > 2/week | 0.2 | 0.1–0.6 | ||||||
| Cancer registries from the Georgia Center for Cancer Statistics, the Metropolitan Detroit Cancer Surveillance System and the New Jersey Cancer registry, United States | Case-control | 539 MM; 1989 controls | Cruciferous Vegetables | MM | Quartiles of consumption: 1 | 1 | Brown et al. Cancer Causes Control 2001 [9] | |
| 2 | 0.9 | 0.7–1.1 | ||||||
| 3 | 0.8 | 0.6–1.0 | ||||||
| 4 | 0.7 | 0.6–0.99 | ||||||
| Institute of Hematology, University Clinical Center, Belgrade, Yugoslavia | Case-control | 100 MM; 100 controls | Vegetables | MM | Vegetable-rich diets (4–7/wk) | OR:0.4 | 0.1–1.0 | Vlajinac et al. Neoplasma 2003 [20] |
| Age, Gene/Environment Susceptibility - Reykjavik study | Prospective Cohort | 5764 (575 MGUS) | Whole wheat bread | MGUS | Whole wheat bread ≥5 /wk in midlife | OR:0.75 | 0.57–0.99 | Thordardottir et al. Plos One 2018 [18] |
| Integrated case-control studies conducted in Northern Italy | Case-control | 120 MM; 7990 controls | Whole grains | MM | Whole grains (>3 days/ week) | OR:0.5 | 0.2–1.1 | Chatenoud et al. Int J Cancer 1998 [28] |
| Series of hospital-based case-control studies conducted in Northern Italy | Case-control | 185 MM; 10,058 controls | Whole grains | MM | Whole grains (>3 days/ week) | OR:0.60 | 0.3–1.0 | La Vecchia et al. Proc Nutr Soc 2003 [29] |
| Yale Comprehensive Cancer Center’s Rapid Case Ascertainment Shared Resource and Connecticut Tumor Registry, Connecticut, USA | Case-control | 173 MM; 670 controls | Fish | MM | 1/mo | OR:1.0 | Hosgood et al. Cancer Causes Control 2007 [10] | |
| 1–3/mo | 0.5 | 0.3–0.7 | ||||||
| 4/mo | 0.8 | 0.5–1.3 | ||||||
| >4/mo | 0.4 | 0.2–0.7 | ||||||
| Yale Comprehensive Cancer Center’s Rapid Case Ascertainment Shared Resource and Connecticut Tumor Registry, Connecticut, USA | Case-control | 173 MM; 670 controls | Shellfish | MM | 1/mo | OR:1.0 | Hosgood et al. Cancer Causes Control 2007 [10] | |
| 1–2/mo | 0.6 | 0.3–1.0 | ||||||
| 3/mo | 0.9 | 0.6–1.3 | ||||||
| >3/mo | 0.5 | 0.3–0.9 | ||||||
| Cancer registries from the Georgia Center for Cancer Statistics, the Metropolitan Detroit Cancer Surveillance System and the New Jersey Cancer registry, USA | Case-control | 539 MM; 1989 controls | Fish | MM | Quartiles of consumption: Low | 1 | Brown et al. Cancer Causes Control 2001 [9] | |
| Medium | 0.8 | 0.6–1.0 | ||||||
| High | 0.7 | 0.5–0.9 | ||||||
| Canadian National Enhanced Cancer Surveillance System (NECSS) | Case-control | 287 MM; 4175 controls | Fish | MM | Servings/week: <0.5 | OR:0.87 | 0.6–1.23 | Fritschi et al. Cancer Epidemiology, Biomarkers & Prevention 2004 [35] |
| 0.5 − <4 | 0.68 | 0.47–1.0 | ||||||
| 4+ | 0.76 | 0.5–1.16 | ||||||
| % fat from fresh fish: Q2 | OR:0.87 | 0.62–1.22 | ||||||
| Q3 | 0.65 | 0.46–0.92 | ||||||
| Q4 | 0.64 | 0.45–0.90 | ||||||
| % total energy from fresh fish: Q2 | OR:0.84 | 0.60–1.17 | ||||||
| Q3 | 0.61 | 0.43–0.87 | ||||||
| Q4 | 0.69 | 0.49–0.97 | ||||||
| Integrated case-control studies conducted in Northern Italy | Case-control | 120 MM; 7990 controls | Fish | MM | Servings/week: <1 | OR: 0.9 | 0.6–1.3 | Fernandez et al. Am J Clin Nutr 1999 [36] |
| 1 | 0.5 | 0.3–0.9 | ||||||
| ≥2 | 0.9 | 0.7–1.1 | ||||||
| 5 major hospitals in Northwest China | Case-control | 220 MM; 220 controls | Fish | MM | Never | OR: 1.0 | Wang et al. Cancer Causes Control 2015 [37] | |
| 1–3/wk | 0.79 | 0.37–1.68 | ||||||
| 1–2/wk | 0.86 | 0.31–2.39 | ||||||
| ≥ 3/wk | 0.77 | 0.30–1.95 | ||||||
| Integrated case-control studies conducted in Northern Italy | Case-control | 120 MM; 7990 controls | Meat | MM | Intake Categories: Intermediate | OR: 0.7 | 0.4–1.1 | Tavani et al. Int J Cancer 2000 [40] |
| High | 1.4 | 0.9–2.2 | ||||||
| Increment of 1 portion/ day | 1.6 | 1.1–2.2 | ||||||
| Institute of Hematology, University Clinical Center, Belgrade, Yugoslavia | Case-control | 100 MM; 100 controls | Meat | MM | Consumption: 7 times/week | OR: 1.9 | 0.9–3.9 | Vlajinac et al. Neoplasma 2003 [20] |
| Cancer registries from the Georgia Center for Cancer Statistics, the Metropolitan Detroit Cancer Surveillance System and the New Jersey Cancer registry, USA | Case-control | 539 MM; 1989 controls | Egg intake in men | MM | Quartiles of consumption: 1 | OR: 1.0 | Brown et al. Cancer Causes Control 2001 [9] | |
| 2 | 1.4 | 0.9–2.2 | ||||||
| 3 | 2.3 | 1.5–3.5 | ||||||
| 4 | 2.1 | 1.4–3.6 | ||||||
| Yale Comprehensive Cancer Center’s Rapid Case Ascertainment Shared Resource and Connecticut Tumor Registry, Connecticut, United States | Case-control | 173 MM; 670 controls | Dairy Products | MM | Cream Soups <1/mo | 1 | Hosgood et al. Cancer Causes Control 2007 [10] | |
| 1/mo | 0.7 | 0.4–1.1 | ||||||
| 2–3/mo | 0.9 | 0.5–1.7 | ||||||
| >3/mo | 2.5 | 1.3–4.6 | ||||||
| Icecreams/Milkshakes <1/mo | OR: 1.0 | |||||||
| 1/mo | 0.6 | 0.3–1.0 | ||||||
| 2–3/mo | 0.9 | 0.6–1.5 | ||||||
| >3/mo | 1.5 | 1.0–2.3 | ||||||
| Pudding/ Custard/ Flan <1/mo | 1.0 | |||||||
| 1/mo | 0.7 | 0.4–1.1 | ||||||
| 2–3/mo | 0.6 | 0.3–1.1 | ||||||
| >3/mo | 2.4 | 1.4–4.0 | ||||||
| Yogurt <1/mo | 1.0 | |||||||
| 1/mo | 1.4 | 0.8–2.5 | ||||||
| 2–3/mo | 0.7 | 0.4–1.2 | ||||||
| >3/mo | 1.0 | 0.7–1.6 | ||||||
| Institute of Hematology, University Clinical Center, Belgrade, Yugoslavia | Case-control | 100 MM; 100 controls | Yogurt | MM | 4–7/week | OR: 3.1 | 1.6–6.0 | Vlajinac et al. Neoplasma 2003 [20] |
| B | ||||||||
| Study Cohort/Location | Study Design | N | Supplement Factor | Risk | Consumption patterns | Risk | 95% CI | Study |
| Veterans Affair nationwide database, USA | 1889 MM; 528 Vitamin-D deficient | Vitamin D serum levels | MM survival | Overall: Normal levels | Median OS: 3.91 years | 3.59–4.38 | Yellapragada et al. Blood Adv 2020 [60] | |
| Deficiency (<20 ng/ml) | 3.10 years | 2.73–3.52 | ||||||
| Whites: Normal levels | 3.87 years | 3.59–4.42 | ||||||
| Deficiency | 2.71 years | 2.18–3.47 | ||||||
| Blacks: Normal levels | 3.95 years | 3.25–5.35 | ||||||
| Deficiency | 3.54 years | 2.99–5.52 | ||||||
| Yale Comprehensive Cancer Center’s Rapid Case Ascertainment Shared Resource and Connecticut Tumor Registry, Connecticut, USA | Case-control | 173 MM; 670 controls | Vitamin A dietary intake | MM | In take (IU/day): <5486 | OR: 1 | Hosgood et al. Cancer Causes Control 2007 [10] | |
| 5489–7545 | 0.3 | 0.2–0.4 | ||||||
| 7546–10,625 | 0.2 | 0.1–0.4 | ||||||
| >10,625 | 0.2 | 0.1–0.4 | ||||||
| Cancer registries from the Georgia Center for Cancer Statistics, the Metropolitan Detroit Cancer Surveillance System and the New Jersey Cancer registry, USA | Case-control | 539 MM; 1989 controls | Vitamin C supplements | MM | Whites | OR: 0.6 | 0.5–0.9 | Brown et al. Cancer Causes Control 2001 [9] |
| Blacks | 0.8 | 0.5–1.4 | ||||||
| C | ||||||||
| Study Cohort/Location | Study Design | N | Lifestyle Factor | Risk | Consumption patterns | Risk | 95% CI | Study |
| No language restriction - international | Meta-analysis of 16 case-control studies and 10 cohort studies | 3921 cases and 19,594 controls;3167 incident cases in total cohort of 2,557,649 subjects | Alcohol | MM | Ever consumption of alcohol | Pooled RR: 0.88 | 0.79–0.99 | Psaltopoulou et al. Leuk Lymphoma 2015 [90] |
| Ever wine drinker | Pooled RR: 0.77 | 0.67–0.89 | ||||||
| National Institutes of Health (NIH)-AARP Diet and Health Study | Prospective Cohort | 499,292 (1312 MM) | Alcohol | MM | Men and women combined | HR: 0.89 | 0.76–1.03 | Santo et al. Int J Cancer 2019 [91] |
| None | ||||||||
| <1 drink/week | 1.0 | |||||||
| <1 drink/day | 0.91 | 0.78–1.05 | ||||||
| 1–1.9 drinks/day | 0.88 | 0.73–1.07 | ||||||
| 2–2.9 drinks/day | 0.7 | 0.50–0.98 | ||||||
| 3–3.9 drinks/day | 0.72 | 0.49–1.07 | ||||||
| ≥4 drinks/day | 0.79 | 0.60–1.04 | ||||||
| Per 10 g/day increase | 0.96 | 0.93–0.99 | ||||||
| Yale Comprehensive Cancer Center’s Rapid Case Ascertainment Shared Resource and Connecticut Tumor Registry, Connecticut, USA | Case-control | 173 MM; 670 controls | Alcohol | MM | Ever beer drinker | OR: 0.5 | 0.3–0.8 | Hosgood et al. Cancer Causes Control 2007 [10] |
| Ever wine drinker | 0.6 | 0.4–0.9 | ||||||
| Ever liquor drinker | 0.7 | 0.5–1.2 | ||||||
| Frequency of consumption of alcohol (liquor, mixed drinks, wine, beer): <2/mo | OR: 1 | |||||||
| 2–12/mo | 0.4 | 0.2–0.6 | ||||||
| 3–7/wk | 0.6 | 0.4–1.1 | ||||||
| >1/day | 0.3 | 0.5–0.8 | ||||||
| Swedish construction workers (Sweden) | Prospective Cohort | 334,957 (1322 incident neoplasms; 236 MM) | Smoking | MM | Smoking Status: Never | RR: 1 | Adami et al. Cancer Causes Control 1998 [95] | |
| Ex-Smoker | 1.1 | 0.8–1.5 | ||||||
| Current | 1 | 0.8–1.4 | ||||||
| Pooled analysis – international | Case-control | 2670 MM; 11,913 controls | Smoking | MM | Smoking Status: Never | OR: 1 | Andreotti et al. Cancer Epidemiol Biomakers Prev 2015 [96] | |
| Ever Smoker | 0.95 | 0.87–1.05 | ||||||
| Current Smoker | 0.82 | 0.73–0.93 | ||||||
| Former Smoker | 1.3 | 0.92–1.14 | ||||||
| Institute of Hematology, University Clinical Center, Belgrade, Yugoslavia | Case-control | 100 MM; 100 controls | Smoking | MM | ≥25 cigarettes/day | OR: 6.9 | 1.3–34.9 | Vlajinac et al. Neoplasma 2003 [20] |
A:Dietary studies. B:Supplement studies. C:Lifestyle studies.
Fig. 1. A beneficial dietary pattern may improve outcomes through modifying the microbiome, metabolome, and immune microenvironment in plasma cell disorders.
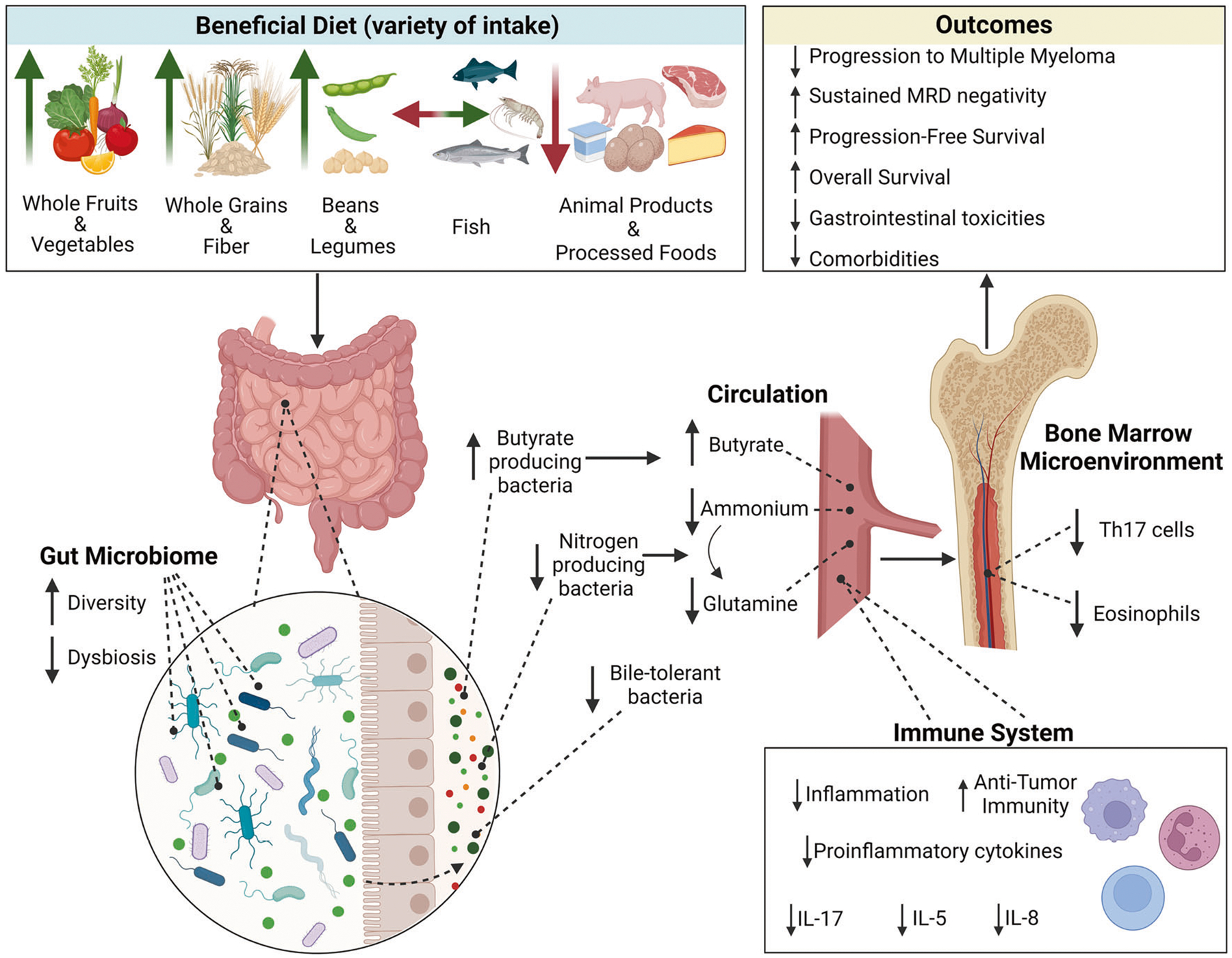
A beneficial diet, outlined in the figure consists of regular consumption of a variety of whole grains, fiber, beans and legumes fruits and vegetables, especially cruciferous vegetables, while limiting intake of animal products. The hypothesized mechanism is outlined above, showcasing that a beneficial diet modifies the gut microbiome and immune microenvironment and thus influences outcomes related to progression and survival in plasma cell disorders as well as reduces toxicities and comorbidities. With consumption of a beneficial diet, the composition and diversity of the gut microbiome improves with increasing short-chain fatty acid producers and decreasing nitrogen producing bacteria and bile-tolerant bacteria. Created with BioRender.com.
Fruits and vegetables
In the AGES-Reykjavik prospective cohort study, fruit consumption (≥3 times per week during the adolescent period) was associated with reduced risk of MGUS (Odds Ratio, OR 0.62, 95% CI: 0.41–0.95). Additionally, fruit consumption during late life was associated with lower risk of progression of MGUS to MM (HR 0.34, 95% CI: 0.13–0.89) [18]. A recent case control study using the National Health and Nutrition Examination Survey (NHANES) showed low intake of fruits and vegetables was associated with lower odds of MGUS [19]. In contrast, three case-control studies did not show significant associations. A trend was noted between fruit consumption 4–7 times/week and MM risk (OR 0.6, 95% CI: 0.3–1.1) [20]. Whereas for citrus fruits, one study showed a trend towards decreased MM risk (>2/week vs 1/month) (OR 0.9, 95% CI: 0.5–1.5, Ptrend = 0.22) [10] and another study showed a non-significant trend towards increased MM risk (OR range for lowest to highest quartile 1.0–1.4; Ptrend = 0.144) [9].
These three case control studies outlined above showed reduced MM risk with increased consumption of vegetables. One study showed reduced MM risk with cruciferous vegetables (>2 times/week vs <1/week) (OR 0.5, 95% CI: 0.3–0.8, Ptrend =0.01) and tomatoes (>2 times/week vs 2–3 times/month) (OR 0.2, 95% CI: 0.1–0.6, Ptrend = 0.002) [10]. The other study demonstrated a similar protective effect of cruciferous vegetables (OR range for lowest to highest intake quartile 1.0–0.7; Ptrend =0.045) [9], as well as overall vegetables 4–7 times/week (OR 0.4, 95% CI: 0.1–1.0) [20].
Fruits are rich in dietary fiber and polyphenols. Dietary fiber is composed of soluble and insoluble carbohydrates including pectin and inulin among others. Pectin makes up 35% of fruit fiber cell wall and has shown to increase butyrate producing microbiota species such as Clostridium cluster XIV [21]. Inulin, a natural storage polysaccharide, present in fruits like bananas, has been shown to increase Faecalibacterium prausnitzii, another butyrate producing bacteria [22]. Additionally higher levels of SCFA have been seen in individuals adhering to a Mediterranean plant forward diet [23]. Vegetables also have a similar favorable impact on the gut microbiome. Cruciferous vegetables are known to contain isothiocyanates, dithiothiones, glucobrassicin and indoles, as well as flavonoids which inhibit cancer development in laboratory animals [9, 24–26]. They also contain vitamin C, an antioxidant that reduces endogenous formation of oxygen-free radicals [27]. Tomatoes, contain lycopene, a compound with anticancer properties [10].
These findings suggest an association between higher fruit consumption and reduced risk of development of MGUS and progression of MGUS to MM and higher consumption of vegetables especially cruciferous vegetables and tomatoes and reduced risk of MM. One possible mechanism could be a positive impact on butyrate producing bacteria (Fig. 1).
Whole grains and fiber
The AGES-Reykjavik prospective cohort study also demonstrated that midlife intake of whole wheat bread ≥5 times per week was associated with lower risk of MGUS (OR 0.75, 95% CI: 0.57–0.99) [18]. The NHANES study similarly showed that low intake of whole grains and high intake of refined sugars and sugar sweetened beverages was associated with the risk of MGUS [19]. An integrated case control study in Northern Italy showed that higher frequency of whole grain food intake (>3 days/week) is associated with reduced MM risk (OR 0.5, 95% CI: 0.2–1.1) [28]. Updated results from this study showed consistent reduced MM risk (OR 0.6, 95% CI: 0.3–1.0) [29]. A smaller case-control study outlined above found no association with bread, grains, or cereals [9].
Whole grains include the bran, endosperm and germ and thus have higher quantities of insoluble and soluble fiber, nutrients and micronutrients like zinc, magnesium, vitamin B6, niacin, antioxidants like vitamin E, phenolic acid and lignans like phytoestrogens when compared with refined grains [29]. Whole grains also have a lower digestion rate and glycemic index with consequent lower rise in insulin and insulin-like growth factor-1 (IGF-1) levels [29]. Both insulin and IGF-1 have been implicated in the pathogenesis of MM by promoting MM cell growth [30]. Indigestible carbohydrate is a major substrate for microbes in the GI tract [31]. A systematic review of 40 studies (1308 participants) demonstrated an increase in microbiome diversity and/or abundance following intact cereal fiber consumption. The effects were apparent from 24 h to 52 weeks and even seen with increases as low as 6–8 g daily [32]. Phyla such as Bacteroidetes, Firmicutes and Actinobacteria and species such as Bifidobacteria, Lactobacillus, Bacteroides and Prevotella significantly increased [32]. Studies have also shown an increase in plasma ferulic acid, a phenolic compound with antioxidant properties [33]. Additionally, a study (14 adults) showed a significant increase in total SCFA levels following consumption all bran cereal [34].
These studies reveal several beneficial effects of whole grains on gut microbiota with consequent increase in phenolic acids and SCFAs (Fig. 1).
Fish
Most case control studies have shown an inverse association with MM risk and fish consumption. In the Connecticut women’s study, fresh fish intake >4 times/month (OR 0.4, 95% CI: 0.2–0.7, Ptrend < 0.001) and shellfish intake >3 times/month (OR 0.5, 95% CI: 0.3–0.9, Ptrend = 0.05) were inversely associated with MM risk [10]. A similar protective effect of fish consumption for MM risk was observed (OR 0.7, 95% CI 0.5–0.9; Ptrend 0.012 for high vs low tertile of fish consumption) in the above-mentioned case control study [9]. A population-based case-control study revealed a protective effect of consumption of higher proportion of total energy or fat intake from fresh fish (OR 0.64, 95% CI: 0.45–0.90, Ptrend < 0.01 for highest vs lowest quartile) [35]. Another integrated case-control study (120 MM cases, 7990 controls) had similar results (OR 0.5, 95% CI: 0.3–0.9 for ≥2 servings/week) [36]. A meta-analysis of 5 case-control studies found a significant inverse relation between the highest versus lowest category of fish consumption and MM risk (RR = 0.65, 95% CI: 0.46–0.91) with relatively high heterogeneity (I2 = 55.6%) [37]. This meta-analysis also suggested a significant non-linear dose-response relationship for this association (p for nonlinearity 0.0041) with the lowest risk linked to once weekly fish consumption [37]. Although seafood is considered healthy, ocean pollution due to microplastics, methyl-mercury, polychlorinated biphenyls and other toxins have made frequent consumption potentially hazardous and may explain the nonlinear dose relationship [38].
Fish and fish oils contain polyunsaturated omega-3 essential fatty acids (PUFAs) that are mainly responsible for the protective effect of fresh fish. They competitively inhibit arachidonic acid utilization in the production of eicosanoids, which have been implicated in tumor progression [35]. They also limit cell growth in mouse MM cell lines in vitro. Additionally, fish oil suppresses T-and B- cell proliferation, enhances apoptosis and reduces IL-6 production (a promoter of MM cell growth) [9]. PUFAs have been shown to decrease ratio of Firmicutes/Bacteroidetes, and increase abundance of butyrate-producing bacterial genera such as Bifidobacterium, Lachnospira, Roseburia and Lactobacillus [39]. Fish also contains vitamin D which decreases risk for MM as discussed below.
Thus, fish consumption maybe protective through multiple mechanisms (Fig. 1).
Meat
There was no statistically significant association between intake of red meat and MM risk (OR for the highest vs lowest tertile 1.4, 95% CI: 0.9–2.2) in a case-control study conducted in Northern Italy [40]. The case-control study from Belgrade, Yugoslavia, suggested that MM cases more frequently consumed meat (7 times/week) when compared with controls (OR 1.9, 95% CI: 0.9–3.9) [20]. Additionally, the EPIC-Oxford Cohort and NHS and HPFS studies showed a reduced MM risk in those eating a plant-based diet and increased risk in meat eaters and those eating a Western diet suggesting that meat intake impacts MM risk [11, 12]. Red meat alters gut microbiome composition which can modulate tumor suppressor and onco-genes. Additionally, components of meat including N-nitroso compounds, heterocyclic amines, and heme [41] have direct effects on cancer risk. Red meat is associated with an alkylating signature in colorectal cancer tumors [42] (Fig. 1).
Eggs
There is no definitive data for egg consumption and MM risk. One small case control study from Georgia, Detroit and New Jersey outlined above showed increased MM risk with egg consumption in men (OR for highest vs lowest quartile 2.1, 95% CI 1.4–3.6; Ptrend = 0.008) [9].
Dairy
The case control study from Connecticut outlined above demonstrated a higher risk for MM with the highest intake quartiles of cream soups (OR 2.5, 95% CI 1.3–4.6, Ptrend = 0.01), ice-creams and milkshakes (OR 1.5, 95% CI: 1.0–2.3, Ptrend = 0.01), pudding and custard (OR 2.4, 95% CI: 1.4–4.0, Ptrend < 0.001) [10]. Consumption of yogurt in the case-control study from Belgrade, Yugoslavia was also associated with increased MM risk (OR 3.1, 95% CI 1.6–6.0) [20]. This higher MM risk may be attributed to higher IGF-1 and insulin levels associated with dairy consumption which has been associated with MM pathogenesis [30, 43, 44].
SUPPLEMENTS AND RISK OF MM
Curcumin
Curcumin, from curcuma longa (turmeric), is a promising natural compound being investigated as a chemo preventive agent. A randomized, double-blind, placebo-controlled cross over study in 25 MGUS/SMM patients utilizing 4 g dose of curcumin with cross over at 3 months followed by an 8 g extension study, demonstrated a statistically significant decrease in the free kappa to free lambda light chain ratio (rFLC, −35% and −36% for 4 g and 8 g dose) and non-statistically significant reduction in involved free light chain, (iFLC, −8% and −10% for 4 g and 8 g dose) and the difference between clonal and nonclonal light chain (dFLC, −9% and −11% for 4 g and 8 g dose) [45]. Statistically significant reductions in total serum and random urinary protein concentrations were also seen [45]. Updated results from long-term follow up of 13 MGUS/SMM patients taking curcumin (4–8 g daily) for 3–9 years showed a change in paraproteins (decrease in 5, slight increase in 3 and rest stable), bone marrow plasma cells (decrease in 4, increase in 2 and rest stable) and reduction in free light chain ratio in 3 patients [46, 47]. Although these preliminary studies suggest a possible long-term benefit for a small subset of patients, additional larger and more mechanism focused studies are needed to define appropriate recommendations for plasma cell disorder patients. Curcumin may inhibit MM proliferation through downregulation of IL-6 and NF-kB as well as suppression of RANKL signaling [48] and has been shown to modify gut microbiota [49]. Supplementation increased bacterial diversity and led to distinct microbiome changes in healthy subjects suggesting that these changes may drive its anti-inflammatory or anti-cancer properties [50]. Two randomized clinical trials are currently underway investigating curcumin supplementation on the gut microbiota in plasma cell disorders. The NUTRIVENTION-2 trial will evaluate 2-week butyrate changes in SMM patients on a curcumin supplement arm, while the NUTRIVENTION-3 trial will evaluate butyrate changes in 12 weeks in MGUS/SMM patients on both curcumin and algae omega-3 supplements [51, 52]. Further research into underlying mechanisms on efficacy such as its effect on the microbiome in plasma cell disorder patients are needed.
Vitamin D
Vitamin D deficiency is the most common nutritional deficiency, and its incidence is higher in MM than in healthy adults. Vitamin D is an immunomodulatory agent that plays a role in regulation of calcium and phosphate homeostasis. Foods like fatty fish and mushrooms are naturally rich sources although a wide range of fortified foods are also available. It is synthesized endogenously after exposure to sunlight [53]. It inhibits growth of MM cells by cell cycle arrest, down regulation of Bcl-2 (an anti-apoptotic protein) and increase in caspase 3 protease activity (apoptosis regulator) [54, 55].
Vitamin D deficiency (<20 ng/ml) and insufficiency (21–29 ng/ml) have been associated with many skeletal and non-skeletal disorders. Several studies have investigated the impact of vitamin D deficiency in MM considering the high risk of skeletal fractures in this population [56–63]. One study in MM patients revealed deficiency in 40% (<14.4 ng/ml), and insufficiency in 35% (14.4–30 ng/ml) [56]. In newly diagnosed MM patients, vitamin D deficiency (<20 ng/ml) correlated with higher C-reactive protein(2.40 vs 0.84 mg/L, p = 0.02), creatinine (1.75 vs 1.24 mg/dL, p = 0.03) and, lower albumin (3.12 vs 3.39 g/dL, p = 0.003) [57]. Vitamin D deficiency correlated with the International Staging System stage, Stage I (16%), Stage II (20%) and Stage III (37%) (p = 0.03) [57]. Vitamin D deficiency (<10 ng/ml) was associated with higher bone marrow plasma cell percentage [62].
In the Veterans Affairs nationwide database, vitamin D deficient patients had worse median overall survival (OS)(3.10 years, 95%CI 2.73–3.52 vs 3.91 years, 95% CI 3.59–4.38; p = 0.002) and higher mortality risk (HR 1.24; p = 0.002) [60]. This OS difference was mainly detected in white patients whereas no difference was noted in African Americans. MM patients post autologous stem cell transplant (ASCT) with low vitamin D levels (≤21 ng/ml) had inferior progression free survival (p = 0.041) and OS (p = 0.049) compared to those with vitamin D levels >21 ng/ml [63]. Several small studies show that deficiency is associated with increased incidence of peripheral neuropathy in SMM, MM patients [64, 65] and increased severity in MM treated with bortezomib and/or thalidomide [58]. Underlying mechanisms include depletion of nerve growth factor and reduced vitamin D neuroprotective effects such as the ability to lower intracellular calcium levels, protection of nerve cells against reactive oxygen species damage and decreased inflammation [65].
Additionally, 25-hydroxy vitamin D deficiency (<25 nmol/L) was associated with statistically significant higher M protein (33 ± 16 g/L), higher urinary deoxypyridinoline excretion (marker of bone turnover) (9.8 ± 5 nmol/mmol Cr) and lower DEXA T-score of femoral neck (−2.4 × 1) in 108 MM patients [59].
Thus, vitamin D deficiency/insufficiency is present in about 75% of MM patients and is associated with higher inflammation, ISS stage, neuropathy, inferior PFS and OS and negative impact on myeloma activity, bone turnover and bone mineral density. Larger population-based prospective studies are warranted to confirm these findings and to objectively measure the benefit of supplementation.
Because vitamin D supplementation modulates the composition of the gut microbiota [66], it would also be interesting to investigate how the microbiota of MM patients is modified by vitamin D.
Vitamin A and C
Dietary intake of Vitamin A and not supplementation was associated with decreased MM risk for all quartiles of vitamin A (Ptrend < 0.001) [10]. Numerous epidemiologic studies have found statistically significant cancer protective effects of vitamin C or vitamin C rich foods [67]. In MM, a case control study showed a statistically significant reduction in MM risk with use of vitamin C supplements in whites (OR 0.6, 95% CI: 0.5–0.9) and a nonsignificant reduction in blacks (OR 0.8, 95% CI: 0.5–1.4). Also, a significant negative risk trend with duration of supplementation was observed for whites (Ptrend < 0.05) [9].
The role of vitamin C as an antioxidant is well known. Concurrent administration with bortezomib abrogates bortezomib mediated inhibition of proteosome activity in human cancer cell lines [68] possibly through direct binding between the hydroxyl group of vitamin C and boronic acid of bortezomib [68]. This was also seen in MM xenograft mouse models [69]. Taken together, these data suggest a benefit of foods rich in vitamin C and a potential for negative effects of vitamin C supplementation in the context of bortezomib treatment.
Omega-3 fatty acids
The PUFAs, eicosapentaenoic acid (EPA), alpha-linolenic acid (ALA), and docosahexaenoic acid (DHA) play important physiologic roles. EPA and ALA have cytotoxic activity on SP 2/0 mouse MM cells in vitro possibly through generation of reactive oxygen species and lipid peroxides [70]. EPA and DHA, induced concentration and time-dependent potent apoptotic effects in plasma cells from MM patients with little to no effects on healthy donor cells [71], at least partly through caspase activation [71]. There was enhanced bortezomib induced cell death when human MM cell lines were pre-treated with DHA or EPA but decreased cell death with concomitant use of bortezomib with DHA or EPA suggesting the importance of timing [72]. Similarly, DHA and EPA increased dexamethasone sensitivity in MM cells through the p53/miR-34a axis and Bcl-2 suppression [73].
A small phase II prospective study (25 newly diagnosed patients) showed that concomitant administration of a tablet containing DHA, α-lipoic acid, vitamin C and vitamin E twice daily for 6 months with bortezomib could prevent chemotherapy-induced neuropathy or its worsening, preserve quality of life and avoid treatment interruption [74].
Additionally, omega-3 PUFAs can exert a positive effect by reversing the altered/dysregulated microbiota composition and increasing butyrate producing bacteria and the production of SCFAs [75] although its effects on the microbiome in MM has not been studied.
Thus, omega-3 fatty acids have anti-inflammatory effects and inhibit growth of MM cell lines [71, 72, 76, 77]. Perhaps the link between fish consumption and a decreased risk of MM may be secondary to omega-3 fatty acids and vitamin D present in fish [37]. Besides fish, other plant based dietary sources of omega 3 include flaxseeds, chia seeds, walnuts, algae, seaweed, and brussels sprouts. However, there have been no interventional clinical trials to date to guide physicians in using PUFAs as an adjunctive therapy for MM management. Two randomized clinical trials will be evaluating changes in stool butyrate levels with algae omega 3 supplementation. The NUTRIVENTION-2 trial will evaluate stool butyrate changes at 2 weeks in SMM patients on an omega 3 supplement arm, while the NUTRIVENTION-3 trial will evaluate stool butyrate changes in 12 weeks in MGUS/SMM patients on algae omega 3 and curcumin supplements [51, 52].
Probiotics
Despite limited data on probiotic supplements, many patients take these supplements without their physicians’ advice. One study surveying 499 cancer patients showed that 28.5% of patients were taking probiotics, many in conjunction with chemotherapy. Less than half of those probiotics were based on their physician’s recommendation. This study also revealed that up to 50% of MM patients used probiotics. There was preference for probiotic preparations consisting predominantly of Lactobacillus and Bifidobacterium strains. A self-reported benefit was described by 61% of patients on chemotherapy, 35% reported no effect and 8.5% reported negative side effects [78].
Systematic reviews suggest that probiotics may prevent diarrhea, acute respiratory tract infections and improve cardiometabolic parameters in non-cancer populations [79]. Some studies suggest that butyrate producing probiotics may promote gut bacterial diversity and play an anti-inflammatory role [80]. In metastatic renal cell carcinoma, a bifidogenic live bacterial product augmented checkpoint inhibitor response [81]. Whereas in melanoma patients on checkpoint inhibitors, sufficient dietary fiber intake without probiotics was associated with improved progression free survival compared to sufficient fiber with probiotics [82]. There is limited data to suggest that Bifidobacterium species reduce adverse effects of chemotherapy, immunotherapy, and radiation therapy by inhibiting proinflammatory cytokines [83]. However, a study of a single species probiotic supplement with Bifidobacterium infantis showed no significant changes in gut microbiota composition or diversity [84]. Though probiotic use is frequent there has been no interventional clinical study to date in the MM population. Therefore, there is not enough data to currently recommend probiotic supplements in MM patients given the known risks of bacteremia from probiotic strains [85] and incomplete post-antibiotic gut microbiome recovery [86, 87]. Further clinical studies are warranted to systematically study the risks and benefits of probiotic use on the microbiome in MM and on MM risk and outcomes and one study in SMM is under development [51].
However, dietary probiotics in the form of highly fermented foods such as kombucha, kimchi and yogurt increased microbiome diversity and decreased markers of inflammation in a 17-week study of 18 patients [88]. At this time for microbiome health, fermented foods may be considered until further studies on probiotics confirm benefit.
OTHER LIFESTYLE FACTORS AND RISK OF MM
Alcohol
Results in MM contradict the results seen for overall cancer risk. A meta-analysis examining the association between alcohol consumption and MM risk showed that ever consumption of alcohol was associated with a reduced MM risk (pooled RR 0.88, 95% CI: 0.79–0.99), especially for wine (pooled RR 0.77, 95% CI: 0.67–0.89 for ever drinkers). This inverse association was significant in females (pooled RR 0.79, 95% CI: 0.69–0.89) [89]. The NIH-AARP Diet and Health Study showed similar inverse association (Ptrend = 0.01) [90]. In this study, compared to light drinkers (<1 drink/week), those who consumed 2–2.9 drinks/day had a statistically significant reduction in MM risk (HR 0.70, 95% CI: 0.50–0.98) [90]. Similar results were seen in a small case control study as well [10].
This paradox in MM to alcohol’s increased overall cancer risk may be due to factors such as impairment of the immune system in heavy drinkers but enhancement of cellular and immune responses in light to moderate drinkers [91]. Additionally, low alcohol consumption improves insulin sensitivity which in turn decreases diabetes and obesity which are related to plasma cell disorders [30, 90]. Another possible hypothesis is that red wine contains antioxidants such as resveratrol (polyphenol in the skin of grapes) which induce apoptosis by inhibiting signal transducer and activator of transcription 3 in human myeloma cell lines [92]. Indirectly, resveratrol modulates the gut microbiota and suppresses the induction of T helper 17 (Th17) lymphocytes [93], an immune cell population that, as discussed below, is relevant in MM pathogenesis.
Smoking
A large prospective study of Swedish construction workers showed no association between smoking with risk of developing leukemia, lymphoma, or MM [94]. Another adequately powered large, pooled analysis of 9 case-control studies demonstrated no significant association between ever smokers, current smokers or former smokers and MM risk compared with never smokers [95]. However, a case-control study from Belgrade, Yugoslavia demonstrated that smoking ≥25 cigarettes/day significantly increased MM risk (OR 6.9, 95% CI: 1.3–34.9) [20]. Thus, there is no clear link between smoking and MM risk.
The effects of alcohol and smoking on the intestinal microbiome have been studied [96, 97] however, data regarding its implications on cancer and specifically MM are lacking.
MICROBIOME
Diet substantially alters the gut microbiome within short periods of time (even 24 h) and additionally may affect disease outcomes, providing a rationale for investigating this relationship in plasma cell disorders [15, 98]. Fecal concentrations of SCFAs such as butyrate and acetate from carbohydrate metabolism are higher on a plant-based than an animal-based diet; while an animal-based diet have a higher proportion of branched-chain fatty acids such as isovalerate and isobutyrate from amino acid fermentation [15, 99]. Diet impacts two major factors of the gut microbiome—diversity and composition. Numerous studies have evaluated these microbiome factors in plasma cell disorders. We summarize the evidence in the next section.
Plasma cell disorders are associated with progressive intestinal dysbiosis when compared to healthy controls
A retrospective study of 40 newly diagnosed MM patients and 17 controls in China evaluated the composition and diversity of intestinal microbiota [5]. In addition to observing that the α diversity (species diversity within a single sample) was significantly decreased in the MM group compared to healthy controls, the evaluation of the composition of intestinal microbiota between MM patients and healthy controls revealed higher levels of Bacteroides, Faecalibacterium and Roseburia genera. Furthermore, the levels of Clostridium leptum of which F. Prausnitzii is the dominant species significantly correlated with ISS stage (R = 0.138, p = 0.017) suggesting a higher abundance of butyrate production associates with more advanced MM [5]. However, two other case-control studies assessing stool samples from MGUS or MM patients relative to healthy controls revealed a qualitatively higher median microbial diversity in patients with MGUS or MM, but a relatively lower abundance of butyrate producing bacteria relative to healthy controls [100, 101]. These studies did not evaluate dietary intake at the time of stool sample acquisition, which may have contributed to some of the differing results. Nevertheless, dysbiosis appears to be a feature of MM, supporting the need for additional larger cohort studies with concurrent evaluation of both dietary intake and microbiome specimens.
Microbiota driven progression of MM
An in vivo mouse study revealed that Prevotella heparinolytica, a commensal microbe found in both human and murine microbiota, can accelerate progression of MM by promoting differentiation of Th17 cells colonizing the intestinal tract and their migration to the bone marrow of transgenic Vk*MYC mice [102]. Lack of IL-17, or alteration of intestinal microbiota in Vk*MYC mice delayed the development of MM. Treatment of these mice with antibodies against IL-17, IL-17RA and IL-5 reduced accumulation of Th17 cells and eosinophils in the bone marrow and delayed disease progression. It was also observed that higher levels of IL-17 in bone marrow of human patients with smoldering myeloma predicted faster disease progression [102]. An observational, non-interventional study is ongoing to demonstrate that in humans a correlation exists between BM levels of IL-17 and composition of the gut microbiota in SMM or MM patients (NCT05712967). Because sodium chloride drives the expansion of Th17 cells which are at the basis of several immune mediated diseases and tumors like MM [103], it would be important to investigate the impact of salt-rich diets on MM. In humans, a comparison between the microbiota of 19 newly diagnosed MM patients and 18 healthy controls demonstrated significant enrichment of nitrogen-recycling bacteria such as Klebsiella and Streptococcus in MM patients. Fecal microbiota transplantation into 5TGM1 mice suggested that nitrogen recycling bacteria enriched in MM accelerated MM progression. Klebsiella pneumoniae was found to promote MM progression via de novo synthesis of glutamine (facilitated by nitrogen capture) in mice [101]. Of relevance, glutamine metabolism promotes Th17 cells [104] and mice fed with glutamine-deficient diet demonstrated slower MM progression further supporting the notion that targeted dietary interventions can modify MM natural history [101].
Microbiome changes during stem cell transplantation
Several studies have looked at changes in fecal microbiota diversity in patients undergoing allogeneic and ASCT [105–107]. A large study of lymphoma, MM, and amyloidosis ASCT recipients revealed lower pre-transplant diversity when compared with healthy controls which further decreased during transplantation [105]. A decreased risk of death and progression was observed with above-median fecal microbiota diversity in the peri-engraftment period [105].
Other studies have shown similar association between decreased oral microbiome diversity and increased risk of post-transplant GI complications as well [106, 107].
In allogenic SCT patients, Eubacterium limosum was associated with reduced risk of relapse or disease progression [108] and decreased diversity was associated with higher risk of transplant-related and graft-versus-host disease related deaths [16].
Pre-transplant dietary analysis in a prospective study (30 MM patients post melphalan 200 mg/m2 ASCT) revealed deficiencies in fiber, vegetable, and whole grain intake below national guidelines with possible consequences to the gut microbiome [109]. This study also confirmed findings of significant loss of gut microbiome diversity post transplantation driven by decreases in Blautia, Ruminococcus, and Faecalibacterium genera and suggested an association between these diversity changes and exposure to intravenous antibiotics used for treatment of neutropenic fever [109]. A retrospective analysis of 142 MM patients who received ASCT demonstrated a decreased median PFS (2.38 vs 6.58 years, p = 0.00003) and decreased median OS (7.43 vs 17.39 years, p = <0.0001) in patients treated with antibiotics compared with those who were not [110]. Fluoroquinolone use and beta-lactam use predicted for decreased PFS and OS. Multivariate analysis showed increased progression risk with beta-lactam use (HR 2.25, 95% CI: 1.31–3.85) and increased mortality risk with antibiotic treatment (HR 5.70, 95% CI: 1.34–34.29). However, it is unclear if the worse outcomes noted in those treated with antibiotics were related to antibiotic mediated alterations in gut microbiome or were merely reflective of a sicker population. These findings warrant larger studies to assess the impact of antibiotics on microbial diversity and outcomes post-transplant.
Sustained MRD negativity associated with intestinal microbiota
The intestinal microbiota plays a role in influencing treatment related efficacy and toxicity [111]. In 34 MM patients on lenalidomide maintenance, a higher relative abundance of butyrate producers Eubacterium Hallii and Faecalibacterium Prausnitzii were seen in minimal residual disease (MRD) negative compared with MRD positive patients [112]. Updated analysis showed that a higher α-diversity (P = 0.0035), relative abundance of butyrate producers (P = 0.025), and stool butyrate concentration (P = 0.037) at 3 months was associated with sustained MRD negativity [17]. These results suggest an association between intestinal microbiota composition and deeper treatment responses.
Dietary changes lead to intestinal microbiome changes
Microbiome diversity.
Prior studies described above indicate the importance of gut microbial diversity in improved patient outcomes such as GI toxicities and survival. The Earth Microbiome Project included over 10 000 individuals and showed an increased gut microbiome diversity and abundances of sub-operational taxonomic units such as Lachnospiraceae, Ruminococcaceae and F. prausnitzii in those who ate over 30 different plants per week compared to those who ate fewer than 10 plants per week [113]. This suggested the significance of a diet abundant in a variety of plant foods. In MM patients on lenalidomide maintenance the Dietary Flavonoid Diversity Index (variety of phytochemicals) was associated with butyrate concentration (P = 0.008) and gut microbiome diversity (P = 0.03) [17]. An ongoing study is evaluating the effect of prebiotic supplementation (resistant starch) in MM and lymphoma patients post ASCT on gut microbiome diversity (NCT05135351).
Microbiome composition.
In these lenalidomide maintenance patients, healthier dietary proteins based on Healthy Eating Index 2015 (from seafood and plants), correlated with butyrate at 3 months (P = 0.009) and sustained MRD negativity (P = 0.05). Consumption of dietary flavonoids (phytochemicals with antioxidant effects) correlated with stool butyrate concentration (anthocyanidins P = 0.01, flavones P = 0.01, and flavanols P = 0.02) as well suggesting the importance of healthier plant-based diet in gut microbiome changes and long-term responses [17].
There is growing evidence showing that diet can considerably influence microbiome composition. The PREDICT 1 trial, a single-arm, single-blinded intervention study, including a discovery cohort in the UK and a validation cohort in the US, analyzed 1098 baseline and 105 follow-up microbiome samples to investigate the interactions between diet, microbiome and cardiometabolic blood biomarkers [114]. This study demonstrated close associations between microbial composition and various dietary indices including the Healthy Food Diversity index (HFD), the Healthy/Unhealthy Plant-based Dietary Indices (hPDI/uPDI) and Healthy Eating Index (HEI) in discovery and validation cohorts [114]. A study of 9 healthy individuals each completing 5 days on two diet arms (plant-based diet and animal-based diet) showed that diet can rapidly and reproducibly alter the human gut microbiome community structure [15]. Animal-based diet increased the abundance of bile-tolerant microbes (Alistipes, Bilophila and Bacteroides) and decreased levels of Firmicutes (Roseburia, Eubacterium rectale and Ruminococcus bromii) that metabolize dietary plant polysaccharides. The stool SCFAs acetate and butyrate were increased on the plant-based diet and isovalerate and isobutyrate were increased on the animal-based diet [15]. Butyrate is known to directly affect growth and differentiation of colonic epithelial cells and exert a predominantly inhibitory effect on colorectal cancers. Accumulation of butyrate within cancerous colonic cells leads to inhibition of histone deacetylase (HDAC) thereby promoting apoptosis and cell death [115]. Sodium butyrate, an HDAC inhibitor, was shown to lead to cell cycle arrest, induce apoptosis and decrease survival of human MM cell lines in a dose-dependent manner [116]. Butyrate also favors bone formation through a complex mechanism that acts on dendritic cells and CD4+ T cells, which eventually stimulate CD8+ T cells to ignite osteoblastogenesis via Wnt10b [117, 118]. Interestingly, HDAC inhibition also leads to the expansion of regulatory T cells at the expense of Th17 cells [119]. Conversely, butyrate promoted the acquisition of memory phenotype of antigen activated CD8+ T cells [120] with potent anti-tumor immune responses [82]. Butyrate also has anti-inflammatory effects such as reducing expression of IL-8 and NF-κB [17]. Thus, the gut microbiota, through the production of metabolites like SCFAs can substantially impact the course of MM. This potential relationship between butyrate producers and treatment response in MM needs to be explored further.
Interventional clinical trials.
Dietary changes produce predictable shifts in host microbial composition, and this has led to a rising interest in developing nutritional strategies to modulate the gut microbiota [98]. A pilot nutrition-based intervention study (NUTRIVENTION) in patients with elevated BMI and MGUS or SMM has completed enrollment and prelim results show the feasibility of a whole food, plant-based diet for weight loss and adherence at 12 weeks from enrollment (NCT04920084) [99]. There was also an improvement in biomarkers of MM progression such as insulin resistance as well as an increase in relative abundance of butyrate producers and stool butyrate levels [99]. A larger randomized study of a whole food plant-based diet when compared to omega-3 and curcumin supplements and placebo supplements is currently enrolling (NCT05640843) [52]. This study will set the stage for larger lifestyle-based trials targeting prevention and survivorship in patients diagnosed with plasma cell disorders [51, 52].
While data for intermittent fasting is not available in plasma cell disorders, a randomized trial of prolonged nightly fasting in patients with MGUS and SMM and elevated BMI is ongoing (NCT05565638). A study of the fasting mimicking diet which includes 5 days of a plant-based calorie restricted diet with every cycle of chemotherapy was conducted in patients with a variety of cancers including MM and it showed improvements in metabolic markers as well as reduced peripheral blood immunosuppressive cells and increased effector cells [121].
Conclusions and future research directions.
In conclusion, this review outlines the current associations between diet, gut microbiome, and MM. MM risk was lower in those with plant-based dietary patterns when compared with meat-eaters. Consumption of fruits, vegetables especially cruciferous vegetables, whole grains, seafood were associated with reduced MM risk (Tables 3 and 4). A high incidence of vitamin D deficiency was observed in MM patients and was found to impact the clinical presentation, myeloma activity, bone turnover and outcomes (PFS and OS) in MM patients. Microbiome dysbiosis is seen in MM and is associated with progression, treatment related efficacy and toxicity. Current studies are underway to evaluate the effect of dietary and supplement interventions on the gut microbiome [51, 52, 99].
Table 3.
Summary of evidence for plasma cell disorders.
| Overall Diet | |
|---|---|
| Dietary Patterns [8–13] |
|
| |
| |
| |
| |
| Individual Dietary Components | |
| Fruit [9, 10, 18, 20] |
|
| |
| Vegetables [9, 10, 20] |
|
| |
| |
| Whole grain [9, 18, 28, 29] |
|
| |
| Fish [9, 10, 35–37] |
|
| |
| Meat [20, 40] |
|
| Eggs [9] |
|
| Dairy [10, 20] |
|
| Supplements | |
| Curcumin [45–47] |
|
| |
| |
| Vitamin D [56–63] |
|
| |
| Vitamin A [10] |
|
| Vitamin C [9, 67] |
|
| |
| Omega-3 fatty acids [75, 76] |
|
| |
| Lifestyle Factors | |
| Alcohol [10, 89, 90] |
|
| |
| **These associations are contradictory to alcohol’s increased overall cancer risk** | |
| Smoking [20, 94, 95] |
|
Table 4.
Overview of dietary recommendations for multiple myeloma and other plasma cell disorders.
| Dietary Recommendations | Canadian Dietary Guidelines 2019 | American Institute for Cancer Research (AICR) Recommendations | Recommendations based on limited Myeloma-Specific data | ||
|---|---|---|---|---|---|
| Recommended Dietary Patterns | Among protein foods, consume plant-based more often. | At least 2/3 of plate should be plant-based foods and majority of diet should consist of plant foods | Plant-based dietary patterns are associated with reduced risk of development of MM; incorporate plant-based foods whenever possible | ||
| There is association between Mediterranean-style diets and decreased CVD risk | An integrated approach to considering the evidence shows that most diets that are protective against cancer are rich in foods of plant origin. | ||||
| Plant Food Items | |||||
| Fruits | Eat plenty of vegetables and fruits while avoiding those with added sugars, sodium, or saturated fat | Eat a variety of fruit and vegetables; at least 15 oz of a variety of non-starchy vegetables and fruit | Consume a variety of fruits; including citrus fruits (orange, lemon, lime, grapefruit, etc.) | ||
| Vegetables | Consume a wide variety of vegetables; including tomatoes and cruciferous vegetables (arugula, bok choy, broccoli, brussel sprouts, cabbage, kale, turnips, etc.) | ||||
| Whole Grains & Fiber | Eat a variety of whole grains. Whole grains should be prioritized over refined grains | Consume a diet that provides at least 30 g per day of fiber from food sources; include whole grains in most meals | Consume a variety of whole grains and fiber-rich foods | ||
| Animal Food Items | |||||
| Fish | Fish and shellfish are considered healthful protein foods choose fresh or frozen that have not been battered or deep-fried | Aim for one serving (8 oz) once weekly | When consuming fish, choose fish highest in omega-3 fatty acids and low in contaminants into your regular diet | ||
| Meat | Plant-based protein foods can provide more fiber and less saturated fat than other types of protein foods such as meat and eggs, thus one should choose plant-based options more often. When choosing meat and poultry, choose lean cuts, skinless poultry, without rich sauces, and prepared with little or no added sodium or saturated fat. | Limit consumption of red and processed meat. No more than three portions/week (12–18 oz total) of red meat | Limit red meat consumption and overall meat consumption. Incorporate plant-based foods whenever possible | ||
| Eggs | At least 2/3 of plate should be plant-based foods and majority of diet should consist of plant foods An integrated approach to considering the evidence shows that most diets that are protective against cancer are rich in foods of plant origin. | Limit overall egg consumption and incorporate plant-based foods whenever possible | |||
| Dairy | Plant-based protein foods can provide more fiber and less saturated fat than other types of protein foods such as dairy. When choosing dairy products, choose lower fat and unsweetened. | Limit overall dairy consumption, especially high-fat dairy and incorporate plant-based foods whenever possible | |||
| Beverages | |||||
| Alcohol** | There are health risks associated with alcohol consumption. If you drink, have no more than 10 drinks a week for women and 15 drinks a week for men*. If you do not drink alcohol, you are not encouraged to start. |
Limit alcohol consumption; for cancer prevention, it’s best not to drink alcohol | Moderate consumption is associated with reduced risk of MM progression | ||
| Vitamins | |||||
| Vitamin D | Aim to meet nutritional needs through diet alone. High-dose supplements are not recommended for cancer prevention | Regularly check Vitamin D levels and supplement if deficient | |||
| Vitamin C | |||||
| Eat a diet rich in Vitamin C foods, avoiding high dose Vitamin C supplementation | |||||
Canadian Dietary Guidelines: food-guide.canada.ca.
AICR Dietary Recommendations: https://www.wcrf.org/diet-and-cancer/cancer-prevention-recommendations/.
Based upon this review of the literature.
One serving of alcohol is 12 oz of beer/cider/cooler, or 5 oz. wine, or 1.5 oz. of distilled alcohol.
The factors outlined above are largely modifiable and may thus lower risk for MM or improve outcomes post diagnosis. However, larger prospective clinical trials and preclinical studies are needed to confirm these findings. This review provides the framework for future research efforts in this area which would enable formation of guidelines to reduce the MM burden and improve the quality of life for MM survivors.
ACKNOWLEDGEMENTS
UAS received research support from the American Society of Hematology Clinical Research Training Institute, NIH/NCI Cancer Center Support Grant P30CA008748, Parker Institute for Cancer Immunotherapy, International Myeloma Society, Paula and Rodger Riney Foundation, TREC Training Workshop R25CA203650 (PI: Melinda Irwin), NCI MSK Paul Calabresi Career Development Award for Clinical Oncology K12 CA184746, HealthTree Foundation, and the Allen Foundation Inc. MB has received funding from Associazione Italiana per la Ricerca sul Cancro (AIRC) under IG2018- ID. 21808. MB was also supported by a grant from the Leukemia & Lymphoma Society (# 6618–21). AML received research support from NIH/NCI Cancer Center Support Grant P30CA008748, NCI 1R01CA249981–01, Sawiris Family Fund, and Paula and Rodger Riney Foundation.
COMPETING INTERESTS
UAS reports grants from NIH/NCI Cancer Center Support Grant P30CA008748, MSK Paul Calabresi Career Development Award for Clinical Oncology K12CA184746, Paula and Rodger Riney Foundation, Allen Foundation Inc, Parker Institute for Cancer Immunotherapy at MSK, HealthTree Foundation, and International Myeloma Society as well as non-financial support from American Society of Hematology Clinical Research Training Institute, TREC Training Workshop R25CA203650 (PI: Melinda Irwin). UAS also reports research funding support from Celgene/BMS, Janssen, Plantable, Sabinsa pharmaceuticals, VeggieDoctor and M and M labs to the institution, non-financial research support; personal fees from ACCC, MashUp MD, Janssen Biotech, Sanofi, BMS, MJH LifeSciences, Intellisphere, Phillips Gilmore Oncology Communications, and RedMedEd outside the submitted work. MB has received honoraria from Bristol Meyers Squibb; Co-owner of the patent # EP18209623.0 - Strategies to improve colonization and expression of Prevotella melaninogenica in the gut of patients affected by IL-17-mediated diseases. AML reports grants from Bristol Myers Squibb and Genentech; grants, personal fees, and non-financial support from Pfizer; and grants and personal fees from Janssen outside the submitted work. AML has a patent for US20150037346A1 licensed and with royalties paid from Serametrix, Inc. No disclosures were reported by the other authors.
REFERENCES
- 1.Surveillance, Epidemiology, and End Results (SEER) Program (www.seer.cancer.gov) SEER*Stat Database: Incidence - SEER Research Data, 12 Registries, Nov 2021 Sub (1992–2019) - Linked To County Attributes - Time Dependent (1990–2019) Income/Rurality, 1969–2020 Counties, National Cancer Institute, DCCPS, Surveillance Research Program, released April 2022, based on the November 2021 submission.
- 2.Clinton SK, Giovannucci EL, Hursting SD. The world cancer research fund/American institute for cancer research third expert report on diet, nutrition, physical activity, and cancer: impact and future directions. J Nutr. 2020;150:663–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Malik MA, Sweeney NW, Jafri M, Derkach A, Chmielewski C, Adintori PA, et al. Nutrition perceptions, needs and practices among patients with plasma cell disorders. Blood Cancer J. 2022;12:70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Almeida A, Mitchell AL, Boland M, Forster SC, Gloor GB, Tarkowska A, et al. A new genomic blueprint of the human gut microbiota. Nature. 2019;568:499–504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Zhang B, Gu J, Liu J, Huang B, Li J. Fecal microbiota taxonomic shifts in Chinese multiple myeloma patients analyzed by quantitative polimerase chain reaction (QPCR) and 16S rRNA high-throughput sequencing. Med Sci Monit Int Med J Exp Clin Res. 2019;25:8269–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Vernocchi P, Del Chierico F, Putignani L. Gut microbiota metabolism and interaction with food components. Int J Mol Sci. 2020;21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Honda K, Littman DR. The microbiota in adaptive immune homeostasis and disease. Nature. 2016;535:75–84. [DOI] [PubMed] [Google Scholar]
- 8.Zhang FF, Cudhea F, Shan Z, Michaud DS, Imamura F, Eom H, et al. Preventable cancer burden associated with poor diet in the United States. JNCI Cancer Spectr. 2019;3:pkz034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Brown LM, Gridley G, Pottern LM, Baris D, Swanso CA, Silverman DT, et al. Diet and nutrition as risk factors for multiple myeloma among blacks and whites in the United States. Cancer Causes Control CCC. 2001;12:117–25. [DOI] [PubMed] [Google Scholar]
- 10.Hosgood HD 3rd, Baris D, Zahm SH, Zheng T, Cross AJ. Diet and risk of multiple myeloma in Connecticut women. Cancer Causes Control CCC. 2007;18:1065–76. [DOI] [PubMed] [Google Scholar]
- 11.Key TJ, Appleby PN, Crowe FL, Bradbury KE, Schmidt JA, Travis RC. Cancer in British vegetarians: updated analyses of 4998 incident cancers in a cohort of 32,491 meat eaters, 8612 fish eaters, 18,298 vegetarians, and 2246 vegans. Am J Clin Nutr. 2014;100:378s–85s. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lee DH, Fung TT, Tabung FK, Colditz GA, Ghobrial IM, Rosner BA, et al. Dietary pattern and risk of multiple myeloma in two large prospective US cohort studies. JNCI Cancer Spectr. 2019;3:pkz025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lee DH, Fung TT, Tabung FK, Marinac CR, Devore EE, Rosner BA, et al. Pre-diagnosis dietary pattern and survival in patients with multiple myeloma. Int J Cancer. 2020;147:1823–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Nagpal R, Shively CA, Register TC, Craft S, Yadav H. Gut microbiome-Mediterranean diet interactions in improving host health. F1000Res. 2019;8:699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.David LA, Maurice CF, Carmody RN, Gootenberg DB, Button JE, Wolfe BE, et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature. 2014;505:559–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Peled JU, Gomes ALC, Devlin SM, Littmann ER, Taur Y, Sung AD, et al. Microbiota as predictor of mortality in allogeneic hematopoietic-cell transplantation. N Engl J Med. 2020;382:822–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Shah UA, Maclachlan KH, Derkach A, Salcedo M, Barnett K, Caple J, et al. Sustained minimal residual disease negativity in multiple myeloma is associated with stool butyrate and healthier plant-based diets. Clin Cancer Res. 2022; 28:5149–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Thordardottir M, Lindqvist EK, Lund SH, Costello R, Burton D, Steingrimsdottir L, et al. Dietary intake is associated with risk of multiple myeloma and its precursor disease. PLoS One. 2018;13:e0206047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Joseph JM, Tang L, Hillengass J, Moysich K, Landgren O, Usmani S, et al. Low intake of fruits and vegetables and high intake of processed meats and juices are associated with risk of mgus in the national health and nutrition examination survey (NHANES) database. Blood. 2022;140:12556–8. [Google Scholar]
- 20.Vlajinac HD, Pekmezović TD, Adanja BJ, Marinković JM, Kanazir MS, Suvajdzić ND, et al. Case-control study of multiple myeloma with special reference to diet as risk factor. Neoplasma. 2003;50:79–83. [PubMed] [Google Scholar]
- 21.Bang SJ, Kim G, Lim MY, Song EJ, Jung DH, Kum JS, et al. The influence of in vitro pectin fermentation on the human fecal microbiome. AMB Express. 2018;8:98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ramirez-Farias C, Slezak K, Fuller Z, Duncan A, Holtrop G, Louis P. Effect of inulin on the human gut microbiota: stimulation of Bifidobacterium adolescentis and Faecalibacterium prausnitzii. Br J Nutr. 2009;101:541–50. [DOI] [PubMed] [Google Scholar]
- 23.Garcia-Mantrana I, Selma-Royo M, Alcantara C, Collado MC. Shifts on gut microbiota associated to mediterranean diet adherence and specific dietary intakes on general adult population. Front Microbiol. 2018;9:890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Zhang Y, Kensler TW, Cho CG, Posner GH, Talalay P. Anticarcinogenic activities of sulforaphane and structurally related synthetic norbornyl isothiocyanates. Proc Natl Acad Sci USA. 1994;91:3147–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bondonno NP, Dalgaard F, Kyrø C, Murray K, Bondonno CP, Lewis JR, et al. Flavonoid intake is associated with lower mortality in the Danish Diet Cancer and Health Cohort. Nat Commun. 2019;10:3651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kopustinskiene DM, Jakstas V, Savickas A, Bernatoniene J. Flavonoids as anticancer agents. Nutrients. 2020;12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Block G Vitamin C and cancer prevention: the epidemiologic evidence. Am J Clin Nutr. 1991;53:270s–82s. [DOI] [PubMed] [Google Scholar]
- 28.Chatenoud L, Tavani A, La Vecchia C, Jacobs DR Jr., Negri E, Levi F, et al. Whole grain food intake and cancer risk. Int J Cancer. 1998;77:24–8. [DOI] [PubMed] [Google Scholar]
- 29.La Vecchia C, Chatenoud L, Negri E, Franceschi S. Session: whole cereal grains, fibre and human cancer wholegrain cereals and cancer in Italy. Proc Nutr Soc. 2003;62:45–9. [DOI] [PubMed] [Google Scholar]
- 30.Parikh R, Tariq SM, Marinac CR, Shah UA. A comprehensive review of the impact of obesity on plasma cell disorders. Leukemia. 2022;36:301–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Seal CJ, Courtin CM, Venema K, de Vries J. Health benefits of whole grain: effects on dietary carbohydrate quality, the gut microbiome, and consequences of processing. Compr Rev Food Sci Food Saf. 2021;20:2742–68. [DOI] [PubMed] [Google Scholar]
- 32.Jefferson A, Adolphus K. The effects of intact cereal grain fibers, including wheat bran on the gut microbiota composition of healthy adults: a systematic review. Front Nutr. 2019;6:33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Vitaglione P, Mennella I, Ferracane R, Rivellese AA, Giacco R, Ercolini D, et al. Whole-grain wheat consumption reduces inflammation in a randomized controlled trial on overweight and obese subjects with unhealthy dietary and lifestyle behaviors: role of polyphenols bound to cereal dietary fiber. Am J Clin Nutr. 2015;101:251–61. [DOI] [PubMed] [Google Scholar]
- 34.Neacsu M, McMonagle J, Fletcher RJ, Hulshof T, Duncan SH, Scobbie L, et al. Availability and dose response of phytophenols from a wheat bran rich cereal product in healthy human volunteers. Mol Nutr Food Res. 2017;61. [DOI] [PubMed] [Google Scholar]
- 35.Fritschi L, Ambrosini GL, Kliewer EV, Johnson KC. Dietary fish intake and risk of leukaemia, multiple myeloma, and non-Hodgkin lymphoma. Cancer epidemiology, biomarkers & prevention: a publication of the American Association for Cancer Research, cosponsored by the American Society of Preventive. Oncology. 2004;13:532–7. [PubMed] [Google Scholar]
- 36.Fernandez E, Chatenoud L, La Vecchia C, Negri E, Franceschi S. Fish consumption and cancer risk. Am J Clin Nutr. 1999;70:85–90. [DOI] [PubMed] [Google Scholar]
- 37.Wang YZ, Wu QJ, Zhu J, Wu L. Fish consumption and risk of myeloma: a meta-analysis of epidemiological studies. Cancer Causes Control CCC. 2015;26:1307–14. [DOI] [PubMed] [Google Scholar]
- 38.Landrigan PJ, Stegeman JJ, Fleming LE, Allemand D, Anderson DM, Backer LC, et al. Human health and ocean pollution. Ann Glob Health. 2020;86:151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Parolini C Effects of fish n-3 PUFAs on intestinal microbiota and immune system. Mar Drugs. 2019;17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Tavani A, La Vecchia C, Gallus S, Lagiou P, Trichopoulos D, Levi F, et al. Red meat intake and cancer risk: a study in Italy. Int J Cancer. 2000;86:425–8. [DOI] [PubMed] [Google Scholar]
- 41.Abu-Ghazaleh N, Chua WJ, Gopalan V. Intestinal microbiota and its association with colon cancer and red/processed meat consumption. J Gastroenterol Hepatol. 2021;36:75–88. [DOI] [PubMed] [Google Scholar]
- 42.Gurjao C, Zhong R, Haruki K, Li YY, Spurr LF, Lee-Six H, et al. Discovery and features of an alkylating signature in colorectal cancer. Cancer Discov. 2021;11:2446–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Allen NE, Appleby PN, Davey GK, Kaaks R, Rinaldi S, Key TJ. The associations of diet with serum insulin-like growth factor I and its main binding proteins in 292 women meat-eaters, vegetarians, and vegans. Cancer Epidemiol, Biomarkers Prev. 2002;11:1441–8. [PubMed] [Google Scholar]
- 44.Knuppel A, Fensom GK, Watts EL, Gunter MJ, Murphy N, Papier K, et al. Circulating insulin-like growth factor-i concentrations and risk of 30 cancers: prospective analyses in UK Biobank. Cancer Res. 2020;80:4014–21. [DOI] [PubMed] [Google Scholar]
- 45.Golombick T, Diamond TH, Manoharan A, Ramakrishna R. Monoclonal gammopathy of undetermined significance, smoldering multiple myeloma, and curcumin: a randomized, double-blind placebo-controlled cross-over 4g study and an open-label 8g extension study. Am J Hematol. 2012;87:455–60. [DOI] [PubMed] [Google Scholar]
- 46.Golombick T, Diamond TH, Manoharan A, Ramakrishna R. Long-term follow-up of curcumin treated MGUS/SMM patients–an updated single centre experienceJ. Hematol Med Oncol. 2017;2. [Google Scholar]
- 47.Golombick T, Diamond TH, Manoharan A, Ramakrishna R. Long term use of curcumin in two smoldering multiple myeloma patients. J Hematol Malig. 2013;3:18–32. [Google Scholar]
- 48.Bharti AC, Donato N, Singh S, Aggarwal BB. Curcumin (diferuloylmethane) down-regulates the constitutive activation of nuclear factor-kappa B and IkappaBalpha kinase in human multiple myeloma cells, leading to suppression of proliferation and induction of apoptosis. Blood. 2003;101:1053–62. [DOI] [PubMed] [Google Scholar]
- 49.Scazzocchio B, Minghetti L, D’Archivio M. Interaction between gut microbiota and curcumin: a new key of understanding for the health effects of curcumin. Nutrients. 2020;12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Peterson CT, Vaughn AR, Sharma V, Chopra D, Mills PJ, Peterson SN, et al. Effects of turmeric and curcumin dietary supplementation on human gut microbiota: a double-blind, randomized, placebo-controlled pilot study. J Evid Based Integr Med. 2018;23:2515690X18790725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Castro F, Sweeney NW, Derkach A, Traore K, Anuraj A, Guttentag L, et al. Microbial changes in response to a plant-based diet and/or supplements in SMM patients: a national multi-arm randomized prospective telehealth study via healthtree: the nutrition prevention (NUTRIVENTION-2) study. Blood. 2022;140:13079–81. [Google Scholar]
- 52.Shah UA, Castro F, Anuraj A, Schach E, Derkach A, Joseph NS, et al. A randomized placebo controlled study of a plant-based dietary versus supplement versus placebo intervention in patients with monoclonal gammopathy of undetermined significance (MGUS) and smoldering multiple myeloma (SMM) -the nutrition prevention (NUTRIVENTION-3) study. Blood. 2022;140:5052–5. [Google Scholar]
- 53.Burwick N Vitamin D and plasma cell dyscrasias: reviewing the significance. Ann Hematol. 2017;96:1271–7. [DOI] [PubMed] [Google Scholar]
- 54.Park WH, Seol JG, Kim ES, Hyun JM, Jung CW, Lee CC, et al. Induction of apoptosis by vitamin D3 analogue EB1089 in NCI-H929 myeloma cells via activation of caspase 3 and p38 MAP kinase. Br J Haematol. 2000;109:576–83. [DOI] [PubMed] [Google Scholar]
- 55.Park WH, Seol JG, Kim ES, Jung CW, Lee CC, Binderup L, et al. Cell cycle arrest induced by the vitamin D(3) analog EB1089 in NCI-H929 myeloma cells is associated with induction of the cyclin-dependent kinase inhibitor p27. Exp Cell Res. 2000;254:279–86. [DOI] [PubMed] [Google Scholar]
- 56.Badros A, Goloubeva O, Terpos E, Milliron T, Baer MR, Streeten E. Prevalence and significance of vitamin D deficiency in multiple myeloma patients. Br J Haematol. 2008;142:492–4. [DOI] [PubMed] [Google Scholar]
- 57.Ng AC, Kumar SK, Rajkumar SV, Drake MT. Impact of vitamin D deficiency on the clinical presentation and prognosis of patients with newly diagnosed multiple myeloma. Am J Hematol. 2009;84:397–400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Wang J, Udd KA, Vidisheva A, Swift RA, Spektor TM, Bravin E, et al. Low serum vitamin D occurs commonly among multiple myeloma patients treated with bortezomib and/or thalidomide and is associated with severe neuropathy. Support Care Cancer. 2016;24:3105–10. [DOI] [PubMed] [Google Scholar]
- 59.Diamond T, Golombick T, Manoharan A. Vitamin D status may effect the skeletal complications of multiple myeloma. Am J Hematol. 2010;85:302–3. [DOI] [PubMed] [Google Scholar]
- 60.Yellapragada SV, Fillmore NR, Frolov A, Zhou Y, Dev P, Yameen H, et al. Vitamin D deficiency predicts for poor overall survival in white but not African American patients with multiple myeloma. Blood Adv. 2020;4:1643–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Mohr SB, Gorham ED, Garland CF, Grant WB, Garland FC, Cuomo RE. Are low ultraviolet B and vitamin D associated with higher incidence of multiple myeloma? J Steroid Biochem Mol Biol. 2015;148:245–52. [DOI] [PubMed] [Google Scholar]
- 62.Lauter B, Schmidt-Wolf IG. Prevalence, supplementation, and impact of vitamin D deficiency in multiple myeloma patients. Cancer Invest. 2015;33:505–9. [DOI] [PubMed] [Google Scholar]
- 63.Eicher F, Mansouri Taleghani B, Schild C, Bacher U, Pabst T. Reduced survival after autologous stem cell transplantation in myeloma and lymphoma patients with low vitamin D serum levels. Hematol Oncol. 2020;38:523–30. [DOI] [PubMed] [Google Scholar]
- 64.Nath K, Ganeshalingam V, Ewart B, Heyer E, Watt K, Birchley A, et al. A retrospective analysis of the prevalence and clinical outcomes of vitamin D deficiency in myeloma patients in tropical Australia. Support Care Cancer. 2020;28:1249–54. [DOI] [PubMed] [Google Scholar]
- 65.Oortgiesen BE, Kroes JA, Scholtens P, Hoogland J, Dannenberg-de Keijzer P, Siemes C, et al. High prevalence of peripheral neuropathy in multiple myeloma patients and the impact of vitamin D levels, a cross-sectional study. Support Care Cancer. 2022;30:271–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Naderpoor N, Mousa A, Fernanda Gomez Arango L, Barrett HL, Dekker Nitert M, de Courten B. Effect of vitamin D supplementation on faecal microbiota: a randomised clinical trial. Nutrients. 2019;11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Block G Epidemiologic evidence regarding vitamin C and cancer. Am J Clin Nutr. 1991;54:1310s–4s. [DOI] [PubMed] [Google Scholar]
- 68.Zou W, Yue P, Lin N, He M, Zhou Z, Lonial S, et al. Vitamin C inactivates the proteasome inhibitor PS-341 in human cancer cells. Clin Cancer Res Off J Am Assoc Cancer Res. 2006;12:273–80. [DOI] [PubMed] [Google Scholar]
- 69.Perrone G, Hideshima T, Ikeda H, Okawa Y, Calabrese E, Gorgun G, et al. Ascorbic acid inhibits antitumor activity of bortezomib in vivo. Leukemia. 2009;23:1679–86. [DOI] [PubMed] [Google Scholar]
- 70.Kumar GS, Das UN. Free radical-dependent suppression of growth of mouse myeloma cells by alpha-linolenic and eicosapentaenoic acids in vitro. Cancer Lett. 1995;92:27–38. [DOI] [PubMed] [Google Scholar]
- 71.Mortaz E, Moloudizargari M, Khosravi A, Asghari MH, Movassaghi M, Varahram M, et al. EPA and DHA have selective toxicity for PBMCs from multiple myeloma patients in a partly caspase-dependent manner. Clin Nutr. 2020;39:2137–43. [DOI] [PubMed] [Google Scholar]
- 72.Chen J, Garssen J, Redegeld F. The efficacy of bortezomib in human multiple myeloma cells is enhanced by combination with omega-3 fatty acids DHA and EPA: Timing is essential. Clin Nutr. 2021;40:1942–53. [DOI] [PubMed] [Google Scholar]
- 73.Dai X, Li M, Geng F. Omega-3 polyunsaturated fatty acids eicosapentaenoic acid and docosahexaenoic acid enhance dexamethasone sensitivity in multiple myeloma cells by the p53/miR-34a/Bcl-2 Axis. Biochem (Mosc). 2017;82:826–33. [DOI] [PubMed] [Google Scholar]
- 74.Maschio M, Zarabla A, Maialetti A, Marchesi F, Giannarelli D, Gumenyuk S, et al. The effect of docosahexaenoic acid and α-Lipoic acid as prevention of bortezomib-related neurotoxicity in patients with multiple myeloma. Integr Cancer Ther. 2019;18:1534735419888584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Costantini L, Molinari R, Farinon B, Merendino N. Impact of Omega-3 fatty acids on the gut microbiota. Int J Mol Sci. 2017;18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Abdi J, Garssen J, Faber J, Redegeld FA. Omega-3 fatty acids, EPA and DHA induce apoptosis and enhance drug sensitivity in multiple myeloma cells but not in normal peripheral mononuclear cells. J Nutr Biochem. 2014;25:1254–62. [DOI] [PubMed] [Google Scholar]
- 77.D’Eliseo D, Di Renzo L, Santoni A, Velotti F. Docosahexaenoic acid (DHA) promotes immunogenic apoptosis in human multiple myeloma cells, induces autophagy and inhibits STAT3 in both tumor and dendritic cells. Genes Cancer. 2017;8:426–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Ciernikova S, Mego M, Semanova M, Wachsmannova L, Adamcikova Z, Stevurkova V, et al. Probiotic survey in cancer patients treated in the out-patient department in a comprehensive cancer center. Integr Cancer Ther. 2017;16:188–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Valdes AM, Walter J, Segal E, Spector TD. Role of the gut microbiota in nutrition and health. BMJ. 2018;361:k2179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Boesmans L, Valles-Colomer M, Wang J, Eeckhaut V, Falony G, Ducatelle R, et al. Butyrate producers as potential next-generation probiotics: safety assessment of the administration of butyricicoccus pullicaecorum to healthy volunteers. mSystems. 2018;3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Dizman N, Meza L, Bergerot P, Alcantara M, Dorff T, Lyou Y, et al. Nivolumab plus ipilimumab with or without live bacterial supplementation in metastatic renal cell carcinoma: a randomized phase 1 trial. Nat Med. 2022;28:704–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Spencer CN, McQuade JL, Gopalakrishnan V, McCulloch JA, Vetizou M, Cogdill AP, et al. Dietary fiber and probiotics influence the gut microbiome and melanoma immunotherapy response. Science. 2021;374:1632–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Badgeley A, Anwar H, Modi K, Murphy P, Lakshmikuttyamma A. Effect of probiotics and gut microbiota on anti-cancer drugs: Mechanistic perspectives. Biochim Biophys Acta Rev Cancer. 2021;1875:188494. [DOI] [PubMed] [Google Scholar]
- 84.Washburn RL, Sandberg D, Gazdik, Stofer MA. Supplementation of a single species probiotic does not affect diversity and composition of the healthy adult gastrointestinal microbiome. Hum Nutr Metab. 2022;28:200148. [Google Scholar]
- 85.Yelin I, Flett KB, Merakou C, Mehrotra P, Stam J, Snesrud E, et al. Genomic and epidemiological evidence of bacterial transmission from probiotic capsule to blood in ICU patients. Nat Med. 2019;25:1728–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Suez J, Zmora N, Zilberman-Schapira G, Mor U, Dori-Bachash M, Bashiardes S, et al. Post-antibiotic gut mucosal microbiome reconstitution is impaired by probiotics and improved by autologous FMT. Cell. 2018;174:1406–23.e16. [DOI] [PubMed] [Google Scholar]
- 87.Brevi A, Cogrossi LL, Lorenzoni M, Mattorre B, Bellone M. The insider: impact of the gut microbiota on cancer immunity and response to therapies in multiple myeloma. Front Immunol. 2022;13:845422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Wastyk HC, Fragiadakis GK, Perelman D, Dahan D, Merrill BD, Yu FB, et al. Gut-microbiota-targeted diets modulate human immune status. Cell. 2021; 184:4137–53.e14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Psaltopoulou T, Sergentanis TN, Sergentanis IN, Karadimitris A, Terpos E, Dimopoulos MA. Alcohol intake, alcoholic beverage type and multiple myeloma risk: a meta-analysis of 26 observational studies. Leuk Lymphoma. 2015;56:1484–501. [DOI] [PubMed] [Google Scholar]
- 90.Santo L, Liao LM, Andreotti G, Purdue MP, Hofmann JN. Alcohol consumption and risk of multiple myeloma in the NIH-AARP Diet and Health Study. Int J Cancer. 2019;144:43–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Díaz LE, Montero A, González-Gross M, Vallejo AI, Romeo J, Marcos A. Influence of alcohol consumption on immunological status: a review. Eur J Clin Nutr. 2002;56:S50–3. [DOI] [PubMed] [Google Scholar]
- 92.Boissy P, Andersen TL, Abdallah BM, Kassem M, Plesner T, Delaissé JM. Resveratrol inhibits myeloma cell growth, prevents osteoclast formation, and promotes osteoblast differentiation. Cancer Res. 2005;65:9943–52. [DOI] [PubMed] [Google Scholar]
- 93.Alrafas HR, Busbee PB, Nagarkatti M, Nagarkatti PS. Resveratrol modulates the gut microbiota to prevent murine colitis development through induction of Tregs and suppression of Th17 cells. J Leukoc Biol. 2019;106:467–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Adami J, Nyrén O, Bergström R, Ekbom A, Engholm G, Englund A, et al. Smoking and the risk of leukemia, lymphoma, and multiple myeloma (Sweden). Cancer Causes Control. 1998;9:49–56. [DOI] [PubMed] [Google Scholar]
- 95.Andreotti G, Birmann BM, Cozen W, De Roos AJ, Chiu BC, Costas L, et al. A pooled analysis of cigarette smoking and risk of multiple myeloma from the international multiple myeloma consortium. Cancer Epidemiol Biomarkers Prev. 2015;24:631–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Engen PA, Green SJ, Voigt RM, Forsyth CB, Keshavarzian A. The Gastrointestinal microbiome: alcohol effects on the composition of intestinal microbiota. Alcohol Res. 2015;37:223–36. [PMC free article] [PubMed] [Google Scholar]
- 97.Savin Z, Kivity S, Yonath H, Yehuda S. Smoking and the intestinal microbiome. Arch Microbiol. 2018;200:677–84. [DOI] [PubMed] [Google Scholar]
- 98.Singh RK, Chang HW, Yan D, Lee KM, Ucmak D, Wong K, et al. Influence of diet on the gut microbiome and implications for human health. J Transl Med. 2017;15:73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Shah UA, Derkach A, Castro F, Anuraj A, Blaslov J, Tran L, et al. A pilot plant based dietary intervention in MGUS and SMM patients with elevated BMI is feasible and associated with improvements in metabolic and microbiome biomarkers of progression. Blood. 2022;140:5066–9. [Google Scholar]
- 100.Antoine Pepeljugoski C, Morgan G, Braunstein M. Analysis of intestinal microbiome in multiple myeloma reveals progressive dysbiosis compared to MGUS and healthy individuals. Blood. 2019;134:3076. [Google Scholar]
- 101.Jian X, Zhu Y, Ouyang J, Wang Y, Lei Q, Xia J, et al. Alterations of gut microbiome accelerate multiple myeloma progression by increasing the relative abundances of nitrogen-recycling bacteria. Microbiome. 2020;8:74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Calcinotto A, Brevi A, Chesi M, Ferrarese R, Garcia Perez L, Grioni M, et al. Microbiota-driven interleukin-17-producing cells and eosinophils synergize to accelerate multiple myeloma progression. Nat Commun. 2018;9:4832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Bellone M, Brevi A, Huber S. Microbiota-propelled T helper 17 cells in inflammatory diseases and cancer. Microbiol Mol Biol Rev. 2020;84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Johnson MO, Wolf MM, Madden MZ, Andrejeva G, Sugiura A, Contreras DC, et al. Distinct regulation of Th17 and Th1 cell differentiation by glutaminase-dependent metabolism. Cell. 2018;175:1780–95.e19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Khan N, Lindner S, Gomes ALC, Devlin SM, Shah GL, Sung AD, et al. Fecal microbiota diversity disruption and clinical outcomes after auto-HCT: a multi-center observational study. Blood. 2021;137:1527–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.El Jurdi N, Filali-Mouhim A, Salem I, Retuerto M, Dambrosio NM, Baer L, et al. Gastrointestinal microbiome and mycobiome changes during autologous transplantation for multiple myeloma: results of a prospective pilot study. Biol Blood Marrow Transplant. 2019;25:1511–9. [DOI] [PubMed] [Google Scholar]
- 107.Laheij A, Raber-Durlacher JE, Koppelmans RGA, Huysmans M, Potting C, van Leeuwen SJM, et al. Microbial changes in relation to oral mucositis in autologous hematopoietic stem cell transplantation recipients. Sci Rep. 2019;9:16929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Peled JU, Devlin SM, Staffas A, Lumish M, Khanin R, Littmann ER, et al. Intestinal microbiota and relapse after hematopoietic-cell transplantation. J Clin Oncol Off J Am Soc Clin Oncol. 2017;35:1650–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.D’Angelo C, Sudakaran S, Asimakopoulos F, Hematti P, El-Gamal D, Safdar N, et al. Perturbation of the gut microbiome and association with outcomes following autologous stem cell transplantation in patients with multiple myeloma. Leuk Lymphoma. 2023;64:87–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.McKenna M, Feinman R, Ahn J, Wang S, Gourna Paleoudis E, Vesole DH, et al. Severe infections and antibiotic use negatively impact progression free and overall survival of multiple myeloma patients undergoing autologous stem cell transplantation. Blood. 2019;134:5510. [Google Scholar]
- 111.Alkharabsheh O, Sidiqi MH, Aljama MA, Gertz MA, Frankel AE. The human microbiota in multiple myeloma and proteasome inhibitors. Acta Haematol. 2020;143:118–23. [DOI] [PubMed] [Google Scholar]
- 112.Pianko MJ, Devlin SM, Littmann ER, Chansakul A, Mastey D, Salcedo M, et al. Minimal residual disease negativity in multiple myeloma is associated with intestinal microbiota composition. Blood Adv. 2019;3:2040–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.McDonald D, Hyde E, Debelius JW, Morton JT, Gonzalez A, Ackermann G, et al. American gut: an open platform for citizen science microbiome research. mSystems. 2018;3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Asnicar F, Berry SE, Valdes AM, Nguyen LH, Piccinno G, Drew DA, et al. Microbiome connections with host metabolism and habitual diet from 1,098 deeply phenotyped individuals. Nat Med. 2021;27:321–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Encarnação JC, Abrantes AM, Pires AS, Botelho MF. Revisit dietary fiber on colorectal cancer: butyrate and its role on prevention and treatment. Cancer Metastasis Rev. 2015;34:465–78. [DOI] [PubMed] [Google Scholar]
- 116.Yao R, Han D, Sun X, Fu C, Wu Q, Yao Y, et al. Histone deacetylase inhibitor NaBut suppresses cell proliferation and induces apoptosis by targeting p21 in multiple myeloma. Am J Transl Res. 2017;9:4994–5002. [PMC free article] [PubMed] [Google Scholar]
- 117.Li JY, Yu M, Pal S, Tyagi AM, Dar H, Adams J, et al. Parathyroid hormone-dependent bone formation requires butyrate production by intestinal microbiota. J Clin Invest. 2020;130:1767–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Tyagi AM, Yu M, Darby TM, Vaccaro C, Li JY, Owens JA, et al. The microbial metabolite butyrate stimulates bone formation via T regulatory cell-mediated regulation of WNT10B expression. Immunity. 2018;49:1116–31.e7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Arpaia N, Campbell C, Fan X, Dikiy S, van der Veeken J, deRoos P, et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature. 2013;504:451–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Bachem A, Makhlouf C, Binger KJ, de Souza DP, Tull D, Hochheiser K, et al. Microbiota-derived short-chain fatty acids promote the memory potential of antigen-activated CD8(+) T cells. Immunity. 2019;51:285–97.e5. [DOI] [PubMed] [Google Scholar]
- 121.Vernieri C, Fuca G, Ligorio F, Huber V, Vingiani A, Iannelli F, et al. Fasting-mimicking diet is safe and reshapes metabolism and antitumor immunity in patients with cancer. Cancer Discov. 2022;12:90–107. [DOI] [PMC free article] [PubMed] [Google Scholar]