

Abstract
Since the first 70 years of reporting cancer chemotherapy, malignant tumors have been the second most common cause of death in children and adults. Currently, the commonly used anti-cancer methods include surgery, chemotherapy, radiotherapy, and immunotherapy. Although these treatment methods could alleviate cancer, they lead to different forms of side effects and have no particularly significant effect on prolonging the patients’ life span. Glycyrrhizic acid (GL), a native Chinese herbal extract, has a wide range of pharmacological effects, such as anti-cancer, anti-inflammatory, antioxidant, and immune regulation. In this review, the anti-cancer effects and mechanisms of GL are summarized in various cancers. The inhibition of GL on chemotherapy-induced side effects, including hepatotoxicity, nephrotoxicity, genotoxicity, neurotoxicity and pulmonary toxicity, is highlighted. Therefore, GL may be a promising and ideal drug for cancer therapy.
Keywords: cancer, glycyrrhizic acid, anti-cancer, side effects, mechanisms
1 Introduction
Cancer has afflicted the multicellular organism for more than 200 million years, and the ancestors of modern humans were plagued by cancer more than a million years ago (Hausman, 2019). As the second most natural cause of death in children and adults (Gilbertson, 2011), each type of cancer has its particular age and form of occurrence, occurring in different proportions and in distinct genders. Until now, people have expended gigantic efforts to understand the origin of cancer cells, the formation of cancer tissue, and the mechanisms of their spread and recurrence, but this disease remains a mystery. A growing body of research suggests that the change in the incidence rate of cancer may be caused by more subtle alterations in the cell hierarchy (Lim et al., 2009; Molyneux et al., 2010). The formation of cancer probably involves three pathways: 1) spreading from the origin of growth to adjacent cells or tissues; 2) spreading from lymph nodes to local lymph nodes; 3) spreading to various tissues and organs throughout the body through blood circulation (Wong and Hynes, 2006).
Although the medical field has made some progress in cancer treatment after more than half a century of research, cancer is still deemed as a human tragedy. There are many treatment methods for cancer based on its type and progression stage. The traditional treatment methods for cancer include surgery, chemotherapy and radiotherapy. With the development of the times and technology, new cancer treatment methods, such as target-specific therapies and immunotherapy, are increasingly applied in clinical practice (Liu et al., 2021). However, these treatment methods all have certain limitations. Surgical treatment is highly traumatic and it cannot completely eliminate cancer cells. The effect of radiotherapy will be not ideal when tumors are not sensitive to radiation and tumors are of systemic metastasis. Among them, chemotherapy for malignant tumors has the strongest cancer killing effect (Safarzadeh et al., 2014; Jain et al., 2016). However, its toxic effects on healthy cells, multiple drug resistance (MDR) after permanent treatment and low bioavailability have enormously limited its clinical application (Rastogi et al., 2014). After these treatments, the survival time of cancer patients has been prolonged, but a great proportion of patients still experience cancer recurrence and cannot achieve permanently survival (Yin et al., 2021). Therefore, seeking novel methods for cancer treatment remains a crucial aspect of medical research.
2 Glycyrrhizic acid
2.1 The structure of GL
Glycyrrhizic acid (GL), a triterpenoid saponin, is the main active component and sweet component of the extract from the root of Glycyrrhiza uralensis Fisch. GL is structurally composed of two molecules of glucuronic acid and one molecule of glycyrrhetinic acid (GA) (Figure 1). GL is metabolized to GA under the action of gut bacteria. In addition, GL is metabolized in the intestine or be transformed via enzymolysis to GA-3-O-mono-β-d-glucuronide (GAMG), a distal glucuronic acid hydrolysate of GL with higher bioavailability and stronger physiological functions (Zuo et al., 2023).
FIGURE 1.
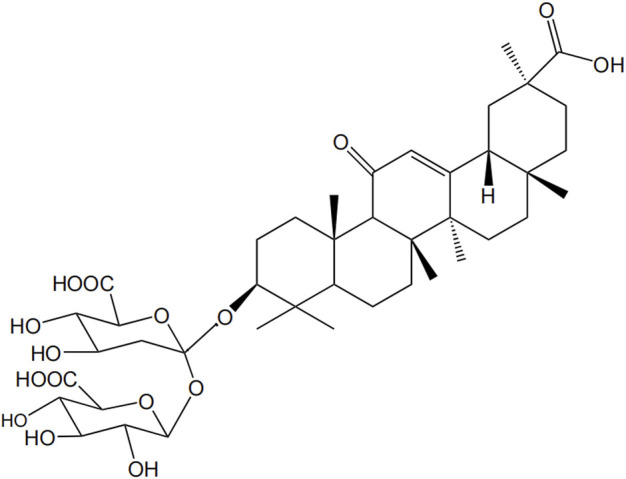
Structural formula of GL.
2.2 The functions of GL
Accumulating studies have demonstrated that GL shows multiple pharmacological activities, such as anti-inflammatory (Liu et al., 2018; Maione et al., 2019), anti-tumor (Zhang et al., 2015; Chang et al., 2019; Tsai et al., 2020), anti-viral (Cinatl et al., 2003; Fu et al., 2016), antioxidant (Xu et al., 2018), immunoregulation (Bernela et al., 2016; Han et al., 2017) and liver protection (Huo et al., 2020) etc. GL mainly inhibits the expression of nuclear factor kappa-B (NF-κB) pathway, thereupon then restraining the encoding of important genes such as inflammatory cytokines, anti-apoptotic factors and cyclooxygenase-2 (COX2) (Li S. et al., 2014), achieving anti-inflammatory and anti-tumor effects. Thus, GL is commonly used in treating various cancers in clinical practice (Stecanella et al., 2021; Zuo et al., 2022), as well as in alleviating liver, lung, and kidney damage caused by cancer chemotherapy (Orazizadeh et al., 2014; Qu et al., 2017). Meanwhile, it also has a protective effect on brain tissue in cases of global cerebral ischemia, brain injury caused by cerebral hemorrhage, and focal ischemia (Xiong et al., 2016).
In addition, GL could facilitate the entry of other drugs into cells through interaction with cell membranes due to its strong hydrophilicity and lipophilicity (Selyutina and Polyakov, 2019). Studies have shown that even at high concentrations, GL still has pharmacological tolerance in rats and humans, and long-term administration has no significant toxic or side effects (van Rossum et al., 1999; Bi et al., 2023; Li et al., 2023). Therefore, it is often used as a blending agent for other drugs to diminish toxicity and enhance drug efficacy (Radwant and Aboul-Enein, 2002; Chen et al., 2009).
3 Roles of GL in cancer therapy
GL inhibits the occurrence and development of cancer by inducing apoptosis pathways in cancer cells. It shows a series of anti-cancer related pharmacological activities, such as broad-spectrum anti-cancer ability, anti-chemotherapy and radiotherapy induced tissue toxicity, absorption enhancement and anti-multiple drug resistance (MDR) mechanism (Wakamatsu et al., 2007; Ajazuddin et al., 2014). Multiple studies have shown that GL acts as an inhibitor of cell signal transduction molecules, angiogenesis inhibitors, tumor related cytokine inhibitors, efficiently and low-toxicity inhibiting the migration and invasion of several types of cancer cells (Kohlschutter et al., 2008; Smolarczyk et al., 2012). The following sections will specifically highlight the anti-cancer mechanism of GL and its inhibition on the side effects of cancer treatment.
3.1 Anti-cancer effects of GL
GL has been reported to have inhibitory effects on various cancers, such as leukemia (Chueh et al., 2012; Bruserud et al., 2015; Shustik et al., 2017), malignant glioma (Li S. et al., 2014), colon cancer (Yang et al., 2015; Jiang et al., 2016), lung cancer (Khan et al., 2013; Huang et al., 2014) etc. The application of GL in various cancers was summarized in Table 1.
TABLE 1.
Summary of anti-cancer mechanisms of GL.
| Cancer type | Models | Administration method and dosage of GL | The efficacy of GL | The anticancer mechanism of GL | References |
|---|---|---|---|---|---|
| Colon Cancer | Wistar rats | Orally, 15 mg/kg for 15 weeks | Anti-tumor; Anti-inflammation | Reducing the expression of Ki-67, proliferating cell nuclear antigen (PCNA), NF-kB, COX-2, iNOS, VEGF; enhancing the expression of p53, connexin-43, Bcl-2, survivin, and caspase-3 | Khan et al. (2018) |
| Balb/c mice | intraperitoneally, 50 or 100 mg/day for 20 days combined with bacteria overexpressing β-glucuronidase | Anti-tumor | Inhibiting tumor growth, increasing apoptosis rate | Afkhami-Poostchi et al. (2020) | |
| Gastric Cancer | KATO III cells | 3 mg/mL for 72 h | Anti-tumor | Inhibiting the growth of KATO III cells; Inducing apoptosis | Hibasami et al. (2005) |
| MGC-803 cells | 1 mg/mL for 24 and 48 h | Anti-tumor | Down-regulating the expression of G1 phase related proteins; Inhibiting phosphorylation of the PI3K/AKT pathway; Downregulating the expression of Bcl-1, survivin and p65, and Up-regulating the expression of Bax and the cleavage of PARP | Wang et al. (2020) | |
| AGS cells | 200 µM for 4 h | Antibacterial infection | Inhibiting the expression of HMGB1; Restoring autolysosomal degradation function | Khan et al. (2023) | |
| Leukemia | WEHI-3 cells | 200, 250, 300, 350 and 400 μM GL and for 24 and 48 h | Anti-tumor | Increasing the levels of ROS and the activity of caspase-3; decreasing the mitochondrial membrane potential (ΔΨm); leading to G0/G1 phase arrest, DNA damage and breakage | Chueh et al. (2012) |
| BALB/c mice suffering TF-1 cells | 100 mg/kg body weight (peritumorally once every other day) | Anti-tumor | Reducing the activity of AKT, mTOR, and STAT3 in tumors; Attenuating expression of cyclin D1, survivin and increasing cleaved caspase-3, cleaved PARP expression | He et al. (2015) | |
| K562 cells/EL-4 lymphoma-bearing C57BL/6J mice | 1.0–4.0 mM for 48 h/50 or 500 mg/kg/day injected i.p. for 4 days | Anti-tumor | Enhancing the levels of apoptosis in K562 cells; Decreasing the tumor burden in mice | Hostetler et al. (2017) | |
| Glioblastoma | human glioblastoma U251 cells | 0, 1, 2, 4 mM for 1, 2 or 4 days | Anti-tumor; Anti-inflammation | Inhibiting the proliferation; Down-regulating the expression of p65 | Li et al. (2014) |
| Lung Cancer | A549 cells/A549 cell xenograft in nude mice | 1, 2 mM for 3 days/50 mg/kg every 2 days for 8 weeks | Anti-tumor | Inhibiting the growth of A549 cells and inducting apoptosis; Inhibiting expression of TxAS | Huang et al. (2014) |
| HCC827 cell xenograft in nude mice | 100 mg/kg for 2 weeks | Anti-tumor | Inhibiting the growth of HCC827 cells; Targeting JAK/STAT/HMGB1 signaling | Wu et al. (2018) | |
| Hepatoma | HepG2 and PLC/PRF/5 | 0.5, 1, and 2 mM for 48 h | Anti-tumor | Inhibiting tumor growth through inducing differentiation and repressing stemness; Enhancing the anti-tumor effects of sorafenib | Cai et al. (2019) |
| SK-Hep1 and Hep3B cells; SK-Hep1/luc2 tumor-bearing mice | 40 or 80 μM for 48 h; 50 mg/kg/day by intraperitoneal injection for 7 days | Anti-tumor | Inhibiting tumor cell growth, cell invasion, and expression of AKT, ERK, EGFR phosphorylation, anti-apoptotic and metastatic proteins; triggering caspase-8/9-mediated apoptosis | Tsai et al. (2020) | |
| Melanoma | Murine B16F10 melanoma; human A375 melanoma cells | 5, 10, 20, 30, 40, 50, 100 ug/mL for 24 h | Anti-tumor | Down-regulating the expression of Treg specific marker, pSTAT3, COX2, PGE2, Bcl2; Up-regulating the expression of Bax and inducing cell apoptosis; Inhibiting of the pSTAT3-mediated Immunosuppressive function of Tregs and MDSCs | Juin et al. (2020) |
| Mice inoculated with B16F10 melanoma cells | 10 mg/kg for 1, 3, 5 and 7 days | Anti-tumor | Inhibiting the pulmonary metastases of B16 melanoma | Kobayashi et al. (2002) |
3.1.1 Anti-colon cancer
Colon cancer, a disease with high incidence, easy migration and difficult cure, constantly develops drug resistance during chemotherapy, increasing the difficulty of treatment (Liang et al., 2020). Khan et al. (2018) found that GL administration inhibited the 1,2-dimethylhydrazine-induced colon tumorigenesis in Wistar rats through reducing the expression of Ki-67, proliferating cell nuclear antigen (PCNA), NF-kB, cyclooxygenase-2(COX-2), inducible nitric oxide synthase (iNOS), and vascular endothelial growth factor (VEGF) and enhancing the expression of p53, connexin-43, B-cell lymphoma-2 (Bcl-2), survivin, and caspase-3, suggesting the chemopreventive potential of GL against colon cancer. Simultaneously, upregulation of the expression of tumor suppressor protein p53 activates casepase-3, thereby inducing tumor cell apoptosis (Yi et al., 2016). In addition, GL reduced drug resistance by combining with first-line anti-cancer drugs. Some theories suggest that GL may enhance the absorption of paclitaxel-loaded GL micelles in the jejunum and colon by inhibiting p-glycoprotein (Yang et al., 2015). Interestingly, Afkhami-Poostchi et al. (2020) applied bacteria-directed enzyme prodrug therapy to convert GL to glycyrrhetinic acid and found that combined treatment of bacteria overexpressing β-glucuronidase and GL more greatly inhibited tumor growth when compared with sole GL treatment.
3.1.2 Anti-gastric cancer
Gastric cancer is the fifth leading cause of cancer-related deaths worldwide, with approximately half of cases occurring in developing countries (Ang and Fock, 2014; Karimi et al., 2014; den Hoed and Kuipers, 2016). GL could inhibit the growth of stomach cancer KATO III cells and induce the fragmentation of DNA to oligonucleosomal-sized fragments, suggesting that GL induced apoptosis (Hibasami et al., 2005). Wang et al. (2020) reported that GL induced apoptosis MGC-803 cells of inhibiting phosphorylation of the PI3K/AKT pathway, downregulating the expression of Bcl-1, survivin and p65, and upregulating the expression of Bax and the cleavage of poly (ADP-ribose) polymerase (PARP). Cell cycle arrest could cause inhibition of cell proliferation. A study has shown that GL inhibited the proliferation of MGC-803 cells, a kind of gastric cancer cells, by inducing G1/s phase arrest because GL treatment downregulated the levels of several G1 phase-related proteins (cyclin D1, D2, D3, E1, and E2) (Wang et al., 2020). In a recent study, Khan et al. (2023) reported that GL treatment inhibited helicobacter pylori infection in AGS cells (gastric cancer cells) via inhibiting high mobility group box1 (HMGB1) and inducing autolysosomal degradation function.
3.1.3 Anti-leukemia
Leukemia is one of the causes of cancer-related deaths in humans, and is an invasive malignant tumor, that is, produced by the rapid growth of abnormal white blood cells (Lee et al., 2007). GL treatment increased the levels of reactive oxygen species (ROS) and the activity of caspase-3 and decreased the mitochondrial membrane potential (ΔΨm) in WEHI-3 cells, as well as led to G0/G1 phase arrest, DNA damage and breakage in a dose-dependent manner (Chueh et al., 2012). GL could inhibit TF-1 cells proliferation in vitro and reduce the volume of TF-1 tumor via inhibiting the activation of AKT/mTOR/signal transducer and activator of transcription 3 (STAT3) signaling pathway, attenuating the expression of cyclin D1 and survivin and increasing the cleavage of caspase-3 and PARP (He et al., 2015). In addition, GL reversed multidrug resistance in human leukemia cell line CEM/ADR 5000 (Zhou and Wink, 2018). Based on these anti-leukemia capabilities, researches on the rationality of GL combination therapy are gradually being carried out. A study has shown that co-treatment with GL and imatinib (a first-line drug) not only enhanced the levels of apoptosis greatly in K562 cells (chronic myeloid leukemia), but also decreased the tumor burden significantly in EL-4 lymphoma-bearing C57BL/6J mice (Hostetler et al., 2017).
3.1.4 Anti-glioblastoma
Gliomas are the most common brain tumor of the central nervous system, approximately accounting for 35%–50% of adult intracranial tumors. It is worth noting that malignant gliomas account for about 60% of gliomas (Penas-Prado et al., 2012). Research has shown that GL inhibited the proliferation of human glioblastoma U251 cells in a time- and dose-dependent manners via down-regulating the expression of p65 (Li S. et al., 2014). NF-κB is a key transcription factor involved in the pathological processes of various human diseases, controlling multiple genes involved in the development of diffuse gliomas and promoting the growth of high-grade gliomas (Kanzawa et al., 2003; Wang et al., 2004). GL inhibited NF-κB pathway via downregulating the expression of p65 protein, encoding of important genes such as anti-inflammatory cytokines, COX-2 and iNOS (Li S. et al., 2014). Dipotassium glycyrrhizinate, a dipotassium salt of GL, also showed an anti-proliferative effect via inducing apoptosis and an anti-migratory effect in U251 and U138MG cells through upregulating the levels of miR-4443 and miR-3620, which are responsible for the post-transcriptional inhibition of NF-κB (Bonafe et al., 2022a).
3.1.5 Anti-lung cancer
Lung cancer is the most common malignant tumor worldwide and the main cause of death for cancer patients. Huang et al. (2014) found that GL inhibited the growth of A549 cells by induction of apoptosis, but had no effects on NCI-H23 cells, another lung adenocarcinoma cell line. They further reported that GL suppressed the expression and activity of thromboxane synthase (TxAS) in A549 cells and clarified the anti-tumor effect of GL in lung adenocarcinoma cells is dependent on inhibition of TxAS (Huang et al., 2014). Importantly, treatment with 50 mg/kg GL for 8 weeks significantly inhibited the growth of xenograft of lung adenocarcinoma cells in vivo (Huang et al., 2014). Deng et al. (2017) reported that the effect of GL treatment alone is comparable to combination of GL and cisplatin in a mouse lung adenocarcinoma model, suggesting the clinical application of GL may decrease the dosage of cisplatin, thereby reducing the side effects of chemotherapy. In HCC827 cells, a non-small cell lung cancer cell line, GL inhibited the migration and invasion of cancer cells via targeting JAK/STAT/HMGB1 signaling (Wu et al., 2018).
3.1.6 Anti-hepatoma
Liver cancer is a common malignant tumor with extremely high mortality rate, ranking third among global cancer mortality rates (Sung et al., 2021). Through the research on the prevalence of liver cancer, it is found that the incidence rate of liver cancer in most countries around the world is still rising year by year. Hepatocellular carcinoma (HCC) has poor differentiation ability and proliferate indefinitely. The dedifferentiation of HCC contributes to malignant progression, characterized by significant morphological changes and loss of liver function (Sun et al., 2011). A recent study has shown that GL administration led to a decrease in stem cell pluripotency and induced the differentiation in HCC in vitro and in vivo by targeting c-Jun N-terminal kinase 1 (JNK1) (Cai et al., 2019); Importantly, blockage of JNK1 mitigate the degree of malignancy of HCC (Cai et al., 2019). In addition, GL combination effectively enhanced the anti-tumor effects of sorafenib, an inhibitor of multi kinases, in HCC treatment (Cai et al., 2019). Tsai et al. (2020) reported that GL not only inhibited dramatically the tumor cell growth and invasion, as well as the phosphorylation of extracellular-signal-regulated kinase (ERK), Akt (Ser473), epidermal growth factor receptor (EGFR) and the anti-apoptotic and metastatic proteins, but also triggered markedly caspase-8/9-mediated apoptosis in HCC in vitro and in vivo. The combination of GL could reverse the resistance of cisplatin in hepatocellular carcinoma cells via inhibiting of MDR-associated proteins (Wakamatsu et al., 2007). Covalent conjugation of GL with polyethyleneimine increased significantly the gene transfection efficiency and superior selectivity for HepG2 cells, suggesting the potential clinical application in vivo (Cao et al., 2019). Several studies reported that co-delivery of GL enhanced the therapeutic efficacy of doxorubicin for hepatocellular carcinoma in vitro and in vivo (Wang et al., 2019; Yang et al., 2019), suggesting the significant clinical application value.
3.1.7 Anti-melanoma
Melanoma is a highly malignant tumor with pigmented cell, melanocyte, accounting for less than 5% of all skin cancers but 80% of skin cancer-related deaths (Bertolotto, 2013). The incidence of malignant melanoma has been steadily increasing globally over the past few decades (Godar, 2011). The immunosuppressive tumor microenvironment (TME) has been identified as a major barrier to evoke an anti-tumor response in melanoma. Moreover, immunosuppressive TME is directly connected with the high activation of T-regulatory cells (Tregs) and myeloid-derived suppressor cells (MDSCs) function. GL downregulated the expression of the anti-apoptotic factor Bcl 2, upregulated the expression of the proinflammatory factor Bax and enhanced the activity of caspase-9 and caspase-3, indicating that GL inhibited the proliferation of melanoma cells by inducing apoptosis (Juin et al., 2020). Meanwhile, GL incubation effectively reduced the expression of Treg specific markers (Forkhead Box P3, glucocorticoid-induced TNFR-related protein and cytotoxic T lymphocyte antigen 4), phospho-STAT3, COX-2 and prostaglandin E2 in melanoma cells, finally limiting the progression of melanoma (Juin et al., 2020). Furthermore, studies showed that GL inhibited the metastasis of melanoma cells by regulating T helper type 2 (Th2) cell resistance, interfering with further the dissemination of melanoma (Kobayashi et al., 2002; Su et al., 2017). Dipotassium of GL inhibited the metastases of melanoma Cells into brain (Bonafe et al., 2022b).
To summarize, GL exerts the anti-tumor activity via inhibiting cell proliferation, inducing apoptosis and resulting in cell cycle arrest in various tumors (Figure 2).
FIGURE 2.

Anti-cancer mechanism diagram of GL. GL mainly plays an important role in various cancers by inhibiting the proliferation of cancer cells, inducing cell apoptosis through its anti-inflammatory, antioxidant, and immune regulatory effects, and inducing cell cycle arrest.
3.2 Inhibiting the side effects of cancer treatment
Most anti-cancer drugs could cause inevitable damage to normal cells during treatment. Therefore, the protection of normal tissues and organs becomes the primary goal of improving the life quality of patients. An increasing number of studies have shown that GL has a strong therapeutic effect on liver, lung and kidney injury caused by chemotherapy or radiotherapy. In addition to the above anti-cancer capabilities, GL could significantly reduce side effects occurring during chemotherapy, especially hepatotoxicity (Li J. Y. et al., 2014). In addition, GL also has better therapeutic effects on nephrotoxicity (Ju et al., 2017), genotoxicity (Arjumand and Sultana, 2011), neurotoxicity (Klein et al., 2023), pulmonary toxicity (Zhu et al., 2021) and other events occurring during cancer treatment. In the following section, we will review the inhibition and potential mechanisms of GL on these side effects (Figure 3).
FIGURE 3.

Mechanism diagram of GL reducing toxic side effects. GL could inhibit a series of side effects such as nephrotoxicity, hepatotoxicity, genotoxicity, neurotoxicity and lung toxicity in cancer treatment by inhibiting the release of inflammatory factors, inhibiting apoptosis and clearing ROS. Fundamentally speaking, GL mainly reduces a series of tissue damage in the process of tumor treatment through its anti-inflammatory and antioxidant properties, thus effectively reducing the side effects of tumor treatment.
3.2.1 Inhibition of hepatotoxicity
Liver is the most important organ for the metabolism of chemical drugs. After metabolized in the liver, a large number of metabolites accumulates in the liver and lead to liver damage and hepatotoxicity. The action mechanism is by inducing the liver lipid peroxidation, resulting in imbalance in redox status, at the same time high levels of ROS-mediated oxidative stress reaction cause liver tissue cell apoptosis and inflammation damage, eventually lead to hepatotoxicity (Waseem et al., 2015). GL administration for 3 h before high-dose methotrexate administration significantly increased hepatic enzyme levels (Mano et al., 2023), suggesting that optimal administration of GL could avoid pharmacokinetic interactions with methotrexate and exerts a hepatoprotective effects. Hepatotoxicity is a common toxic side effect of many chemotherapeutic drugs. Kishimoto et al. (2021) reported that 39 of 118 patients with acute leukemia (33%) developed grade 3–4 hepatotoxicity after combined administration of intravenous monoammonium glycyrrhizinate and methotrexate. As one of the hepatoprotective compounds, GL is commonly used in the treatment of acute and chronic liver injury, viral hepatitis, hepatic steatosis, liver fibrosis, liver cancer and other diseases. Moreover, GL has various pharmacological effects, such as anti-inflammatory, neuroprotection, antiviral, anti-tumor, antioxidant, and hepatoprotective activities (Li J. Y. et al., 2014). It could reduce the content of ROS by reducing lipid peroxidation (Kiso et al., 1984) and increasing the activity of superoxide dismutase (SOD), glutathione peroxidase (GPx) and catalase (Kao et al., 2009). Furthermore, GL inhibits tripterygium-glycoside-tablet-induced acute liver injury by regulating pyruvate kinase M2 and reducing oxidative, inflammation stress and apoptosis (Wang et al., 2022). In addition, GL could significantly reduce the steatosis and necrosis of hepatocytes, inhibit interstitial inflammation and liver fibrosis, and promote cell regeneration (Li J. Y. et al., 2014).
3.2.2 Inhibition of nephrotoxicity
Chemotherapy is one of the main cancer treatments. In cancer treatment, traditional chemotherapeutic drugs are the first-line drugs for the treatment of several malignancies, but they also cause kidney toxicity. In addition to traditional cytotoxic drugs, molecular targeted drugs could also affect kidney function and cause nephrotoxicity, which limits the efficacy of treatment and affects the life quality and overall survival of patients (Santos et al., 2020). Studies have shown that the nephrotoxicity induced by some anti-cancer drugs may be closely related to the oxidative stress and inflammatory responses. GL has antioxidant and anti-inflammatory properties, which could effectively inhibit pro-inflammatory substances (NF-κB, TNF-α, IL-1β, IL-6, and HMGB 1), increase levels of nuclear factor erythroid 2-related factor 2 (Nrf 2) and heme oxygenase-1 (HO-1), and restore the activity of antioxidant enzyme and GSH/GSSG ratio (Wu et al., 2015), reduce lipid peroxidation (Arjumand and Sultana, 2011), suggesting that GL effectively inhibits renal inflammation and oxidative stress, and relieve chemical kidney injury. In gentamicin-induced renal injury, GL may inhibit gentamicin-induced ROS generation or scavenge ROS before it reaches the cellular target tissue, while GL also restores the expression of aquaporin 2, thus improving renal defects in rats with gentamicin-induced acute renal failure (Sohn et al., 2003). Moreover, GL could also ameliorate cisplatin-induced renal injury and inhibit nephrotoxicity through reducing ROS-mediated p53 activation and promoting p21 expression in HK-2 cells (Ju et al., 2017). The renal protective effects of GL may be related to upregulation of Nrf2 and downregulation of NF-κB in the kidney of BALB/c mice (Wu et al., 2015).
3.2.3 Inhibition of genotoxicity
Most of the anti-tumor drugs have genotoxic effects that contribute to growth inhibition. Exposure to genotoxins causes an increased risk of carcinogenic and teratogenicity (Arjumand and Sultana, 2011). These genotoxic and oncogenic potential may lead to the formation of secondary cancers (Said Salem et al., 2017). Some studies have indicated that many drug-induced genotoxicity and chromosomal instability are closely related to the parameters of oxidative stress. ROS act directly on intracellular components, including lipids, proteins and DNA, and disrupt their structures. The chemoprotective agents could exert their antigenotoxic effects through one or variety of mechanisms, such as inhibiting the formation of reactive carcinogenic metabolites, inducing enzymes that detoxify carcinogens, scavenging ROS, inhibiting cell proliferation and regulating cell apoptosis (Arjumand and Sultana, 2011). GL has anti-inflammatory, anti-oxidant, anti-cancer, and immunomodulatory effects (Akamatsu et al., 1991; Kelloff et al., 1994; Wakamatsu et al., 2007) and is considered as a possible chemopreventive agent. Increasing genotoxic researches have demonstrated that GL is not only nonteratogenic and nonmutagenic, but also have anti-genotoxic property (Isbrucker and Burdock, 2006). Arjumand and Sultana (2011) reported that GL administration significantly reversed the genotoxicity induced by cisplatin, including a decrease in DNA fragmentation and increases in the content of glutathione and activities of the anti-oxidant enzymes (catalase, glutathione peroxidase, glutathione reductase, quinone reductase and glutathione-S-transferase).
3.2.4 Inhibition of neurotoxicity
Cancer therapies could cause a wide range of neurologic adverse effects and may cause a significant increase in morbidity and mortality of cancer patients (Dietrich, 2020). Some chemotherapeutic drugs, such as taxanes (paclitaxel and docetaxel), platinum compounds (cisplatin, oxaliplatin and carboplatin), and vinca alkaloids (vinblastine and vincristine), have strong neurotoxicity and could easily induce a peripheral neuropathy (CIPN) (Klein and Lehmann, 2021). CIPN is associated with a length-dependent axonal sensory neuropathy in taxol-induced neuropathy (Staff et al., 2017). Paclitaxel easy access to and accumulate in the dorsal root ganglia, which will cause numbness and pain in hands and feet (Cavaletti et al., 2000). Paclitaxel induces disruption of axonal transport through microtubule stabilization, changes in mitochondrial morphology and function, and inflammatory responses, leading to axonal symmetry damage and loss of nerve fibers, and subsequent neurotoxicity (Niznansky et al., 2022). Increasing studies have suggested that GL could provide neuroprotection in nerve system due to its strong anti-inflammatory, anti-apoptosis, antioxidant and autophagy regulation properties (Kim et al., 2012; Yang et al., 2018; Gendy et al., 2023; Shan et al., 2023). A recent study has shown that GL alleviated paclitaxel-induced neurotoxicity in vitro and in vivo by inhibiting the neuronal uptake mediated by organic anion transport peptides (OATPs), which are the main neuronal transporters of paclitaxel (Klein et al., 2023).
3.2.5 Inhibition of pulmonary toxicity
Some chemotherapeutic agents, such as bleomycin, could cause severe pulmonary fibrosis and produce pulmonary toxicity (van der Schoot et al., 2016). GL could alleviate benzo(a)pyrene exposure-induced lung injury in rats via ameliorating the detoxification and antioxidant function of lung (Qamar et al., 2012). HMGB1 is a cytokine-like protein found in the nucleus of all cells and has multiple functions in inflammation, infection, tissue damage, cell apoptosis, and immune response (Qu et al., 2019). Furthermore, HMGB1 could not only act as a proinflammatory factor to directly involve in tissue damage (Qin et al., 2006), but also induce lung fibrosis through NF-κB-mediated release of transforming growth factor beta1 (TGF-β1) (Wang et al., 2017). As an inhibitor of HMGB 1, GL treatment reduced the inflammation and fibrosis by inhibiting the mitogen-activated protein kinase (MAPK) inflammatory signaling and Smad3 fibrotic signaling pathway (Zhu et al., 2021), thereby alleviating the lung toxicity induced by bleomycin, which is clinically used to treated various tumors (Gederaas et al., 2023).
4 Conclusion and expectation
With the development of modern science and technology, the discovery of plant pharmacological components was promoted. GL, as the main active ingredient of the licorice extract, has a wide range of pharmacological activities. Notably, increasing literature have demonstrated that GL shows inhibitory effects on various cancers by inhibiting cell proliferation, inducing apoptosis and resulting in cell cycle arrest through multitudinous mechanisms. What’s more, GL could reduce effectively the side effects of cancer treatment via inhibition of chemotherapy-induced renal toxicity, liver toxicity, genotoxicity, neurotoxicity and pulmonary toxicity. Given the important roles of GL in cancer treatment, the application of GL alone in the cancer chemotherapy or combined use with other anti-tumor drugs will have very bright application prospects.
Besides above side effects induced in the chemotherapy of cancer, ototoxicity is another common side effect which limits the clinical use of chemotherapeutics (Basirat et al., 2023). Many chemotherapeutics could affect the inner ear or auditory nerve, leading to hearing loss. For example, Cisplatin is widely used as a chemotherapeutic drug with a high rate of ototoxicity (an average incidence of more than 60%) (Arwanda et al., 2023). Inflammation and oxidative stress may be closely related to the ototoxicity of Cisplatin (Ramkumar et al., 2021). However, the role of GL in combating ototoxicity induced by chemotherapy is largely unclear. Therefore, it is expected that GL will help solving the hearing loss caused by ototoxic drugs in the future. What’s more, the mechanisms for GL inhibiting various side effects are not presently clarified accurately. Thus, more studies should be performed to elucidate the protective mechanisms.
GL shows stronger actions in cancer treatment, but some factors may limit its clinical application. The human body has multiple reactions to GL and there are significant individual differences. However, the reasons for the differences are not yet clear, and the proportion of sensitive populations has not been determined. On the basis of different administration methods and drug concentrations, electrolyte imbalance, edema, elevated blood pressure and false aldosterone symptoms may occur (Gomez-Sanchez and Gomez-Sanchez, 1992; Johns, 2009; Celik et al., 2012). Studies have shown that the risk of toxicity after oral administration is much lower than intravenous or intraperitoneal administration, but we should still be particularly careful to avoid high doses or long-term ingestion of GL (Wang and Nixon, 2001). What’s more, people with diseases such as heart and kidney problems, hypertension may be more susceptible to the adverse effects of GL (Ruszymah et al., 1995). In addition, the content of GL in different varieties of liquorice varies greatly, and the yield depends on the source of plants, which also limits greatly its development and application. Han et al. (2021) recommended that regulating biosynthesis pathway of GL through environmental stimuli would provide a new idea for obtaining high-quality GL. Although side effects of GL have been reported, their incidence can be avoided with reasonable medication. The most important thing is that its potential for treating various diseases is worthy of recognition.
Funding Statement
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study is financially supported by National College Student Innovation Training Program of China (202310354023) and Jiaxing University Student Research and Training Program (8517221143).
Author contributions
YZ: Writing–original draft. ZS: Writing–original draft. JX: Resources, Software, Writing–original draft. YL: Resources, Software, Writing–original draft. JH: Resources, Software, Writing–original draft. JJ: Writing–review and editing. XZ: Conceptualization, Writing–review and editing. LL: Conceptualization, Writing–review and editing.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
- Afkhami-Poostchi A., Mashreghi M., Iranshahi M., Matin M. M. (2020). Use of a genetically engineered E. coli overexpressing ÿ-glucuronidase accompanied by glycyrrhizic acid, a natural and anti-inflammatory agent, for directed treatment of colon carcinoma in a mouse model. Int. J. Pharm. 579, 119159. 10.1016/j.ijpharm.2020.119159 [DOI] [PubMed] [Google Scholar]
- Ajazuddin, , Alexander A., Qureshi A., Kumari L., Vaishnav P., Sharma M., et al. (2014). Role of herbal bioactives as a potential bioavailability enhancer for Active Pharmaceutical Ingredients. Fitoterapia 97, 1–14. 10.1016/j.fitote.2014.05.005 [DOI] [PubMed] [Google Scholar]
- Akamatsu H., Komura J., Asada Y., Niwa Y. (1991). Mechanism of anti-inflammatory action of glycyrrhizin: effect on neutrophil functions including reactive oxygen species generation. Planta Med. 57 (2), 119–121. 10.1055/s-2006-960045 [DOI] [PubMed] [Google Scholar]
- Ang T. L., Fock K. M. (2014). Clinical epidemiology of gastric cancer. Singap. Med. J. 55 (12), 621–628. 10.11622/smedj.2014174 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arjumand W., Sultana S. (2011). Glycyrrhizic acid: a phytochemical with a protective role against cisplatin-induced genotoxicity and nephrotoxicity. Life Sci. 89 (13-14), 422–429. 10.1016/j.lfs.2011.06.016 [DOI] [PubMed] [Google Scholar]
- Arwanda M. D., Haryuna T. S., Adriztina I., Khalid K. (2023). Curcumin prevents ototoxicity induced by cisplatin as evaluated with OAE. Iran. J. Otorhinolaryngol. 35 (129), 189–197. 10.22038/IJORL.2023.71786.3452 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Basirat U., Bin Tariq U., Moeen N., Jawhar Z. H., Shoja S. J., Kareem A. K., et al. (2023). A systematic review of the chemo/radioprotective effects of melatonin against ototoxic adverse effects induced by chemotherapy and radiotherapy. Curr. Pharm. Des. 29 (15), 1218–1229. 10.2174/1381612829666230503145707 [DOI] [PubMed] [Google Scholar]
- Bernela M., Ahuja M., Thakur R. (2016). Enhancement of anti-inflammatory activity of glycyrrhizic acid by encapsulation in chitosan-katira gum nanoparticles. Eur. J. Pharm. Biopharm. 105, 141–147. 10.1016/j.ejpb.2016.06.003 [DOI] [PubMed] [Google Scholar]
- Bertolotto C. (2013). Melanoma: from melanocyte to genetic alterations and clinical options. Sci. (Cairo) 2013, 635203. 10.1155/2013/635203 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bi X., Yang L., Lin Y., Deng W., Jiang T., Zhang L., et al. (2023). Efficacy and safety of glycyrrhizic acid in treatment of autoimmune hepatitis. Am. J. Chin. Med. 51 (2), 391–405. 10.1142/S0192415X23500209 [DOI] [PubMed] [Google Scholar]
- Bonafe G. A., Dos Santos J. S., Fernandes A., Ziegler J. V., Marson F. A. L., Rocha T., et al. (2022a). Anti-migratory effect of dipotassium glycyrrhizinate on glioblastoma cell lines: microarray data for the identification of key MicroRNA signatures. Front. Oncol. 12, 819599. 10.3389/fonc.2022.819599 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonafe G. A., Dos Santos J. S., Ziegler J. V., Marson F. A. L., Rocha T., Ortega M. M. (2022b). Dipotassium glycyrrhizinate on melanoma cell line: inhibition of cerebral metastases formation by targeting NF-kB genes-mediating MicroRNA-4443 and MicroRNA-3620-dipotassium glycyrrhizinate effect on melanoma. Int. J. Mol. Sci. 23 (13), 7251. 10.3390/ijms23137251 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bruserud O., Nepstad I., Hauge M., Hatfield K. J., Reikvam H. (2015). STAT3 as a possible therapeutic target in human malignancies: lessons from acute myeloid leukemia. Expert Rev. Hematol. 8 (1), 29–41. 10.1586/17474086.2015.971005 [DOI] [PubMed] [Google Scholar]
- Cai S., Bi Z., Bai Y., Zhang H., Zhai D., Xiao C., et al. (2019). Glycyrrhizic acid-induced differentiation repressed stemness in hepatocellular carcinoma by targeting c-jun N-terminal kinase 1. Front. Oncol. 9, 1431. 10.3389/fonc.2019.01431 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao M., Gao Y., Zhan M., Qiu N., Piao Y., Zhou Z., et al. (2019). Glycyrrhizin acid and glycyrrhetinic acid modified polyethyleneimine for targeted DNA delivery to hepatocellular carcinoma. Int. J. Mol. Sci. 20 (20), 5074. 10.3390/ijms20205074 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cavaletti G., Cavalletti E., Oggioni N., Sottani C., Minoia C., D'Incalci M., et al. (2000). Distribution of paclitaxel within the nervous system of the rat after repeated intravenous administration. Neurotoxicology 21 (3), 389–393. [PubMed] [Google Scholar]
- Celik M. M., Karakus A., Zeren C., Demir M., Bayarogullari H., Duru M., et al. (2012). Licorice induced hypokalemia, edema, and thrombocytopenia. Hum. Exp. Toxicol. 31 (12), 1295–1298. 10.1177/0960327112446843 [DOI] [PubMed] [Google Scholar]
- Chang H. Y., Chen S. Y., Wu C. H., Lu C. C., Yen G. C. (2019). Glycyrrhizin attenuates the process of epithelial-to-mesenchymal transition by modulating HMGB1 initiated novel signaling pathway in prostate cancer cells. J. Agric. Food Chem. 67 (12), 3323–3332. 10.1021/acs.jafc.9b00251 [DOI] [PubMed] [Google Scholar]
- Chen L., Yang J., Davey A. K., Chen Y. X., Wang J. P., Liu X. Q. (2009). Effects of diammonium glycyrrhizinate on the pharmacokinetics of aconitine in rats and the potential mechanism. Xenobiotica 39 (12), 955–963. 10.3109/00498250903271997 [DOI] [PubMed] [Google Scholar]
- Chueh F. S., Hsiao Y. T., Chang S. J., Wu P. P., Yang J. S., Lin J. J., et al. (2012). Glycyrrhizic acid induces apoptosis in WEHI-3 mouse leukemia cells through the caspase- and mitochondria-dependent pathways. Oncol. Rep. 28 (6), 2069–2076. 10.3892/or.2012.2029 [DOI] [PubMed] [Google Scholar]
- Cinatl J., Morgenstern B., Bauer G., Chandra P., Rabenau H., Doerr H. W. (2003). Glycyrrhizin, an active component of liquorice roots, and replication of SARS-associated coronavirus. Lancet 361 (9374), 2045–2046. 10.1016/s0140-6736(03)13615-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- den Hoed C. M., Kuipers E. J. (2016). Gastric cancer: how can we reduce the incidence of this disease? Curr. Gastroenterol. Rep. 18 (7), 34. 10.1007/s11894-016-0506-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deng Q. P., Wang M. J., Zeng X., Chen G. G., Huang R. Y. (2017). Effects of glycyrrhizin in a mouse model of lung adenocarcinoma. Cell Physiol. Biochem. 41 (4), 1383–1392. 10.1159/000467897 [DOI] [PubMed] [Google Scholar]
- Dietrich J. (2020). Neurotoxicity of cancer therapies. Contin. (Minneap Minn) 26 (6), 1646–1672. 10.1212/CON.0000000000000943 [DOI] [PubMed] [Google Scholar]
- Fu X., Wang Z., Li L., Dong S., Li Z., Jiang Z., et al. (2016). Novel chemical ligands to ebola virus and marburg virus nucleoproteins identified by combining affinity mass spectrometry and metabolomics approaches. Sci. Rep. 6, 29680. 10.1038/srep29680 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gederaas O. A., Sharma A., Mbarak S., Sporsheim B., Hogset A., Bogoeva V., et al. (2023). Proteomic analysis reveals mechanisms underlying increased efficacy of bleomycin by photochemical internalization in bladder cancer cells. Mol. Omics 2023. 10.1039/d2mo00337f [DOI] [PubMed] [Google Scholar]
- Gendy A. M., El-Sadek H. M., Amin M. M., Ahmed K. A., El-Sayed M. K., El-Haddad A. E., et al. (2023). Glycyrrhizin prevents 3-nitropropionic acid-induced neurotoxicity by downregulating HMGB1/TLR4/NF-κB p65 signaling, and attenuating oxidative stress, inflammation, and apoptosis in rats. Life Sci. 314, 121317. 10.1016/j.lfs.2022.121317 [DOI] [PubMed] [Google Scholar]
- Gilbertson R. J. (2011). Mapping cancer origins. Cell 145 (1), 25–29. 10.1016/j.cell.2011.03.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Godar D. E. (2011). Worldwide increasing incidences of cutaneous malignant melanoma. J. Skin. Cancer 2011, 858425. 10.1155/2011/858425 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomez-Sanchez E. P., Gomez-Sanchez C. E. (1992). Central hypertensinogenic effects of glycyrrhizic acid and carbenoxolone. Am. J. Physiol. 263 (6), E1125–E1130. 10.1152/ajpendo.2006.263.6.E1125 [DOI] [PubMed] [Google Scholar]
- Han S., Sun L., He F., Che H. (2017). Anti-allergic activity of glycyrrhizic acid on IgE-mediated allergic reaction by regulation of allergy-related immune cells. Sci. Rep. 7 (1), 7222. 10.1038/s41598-017-07833-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han Y. X., Jia Q. J., Yang D. F., Chai W. G., Zhang X. M., He Q. L., et al. (2021). Current advances in environmental stimuli regulating the glycyrrhizic acid biosynthesis pathway. Fitoterapia 151, 104860. 10.1016/j.fitote.2021.104860 [DOI] [PubMed] [Google Scholar]
- Hausman D. M. (2019). What is cancer? Perspect. Biol. Med. 62 (4), 778–784. 10.1353/pbm.2019.0046 [DOI] [PubMed] [Google Scholar]
- He S. Q., Gao M., Fu Y. F., Zhang Y. N. (2015). Glycyrrhizic acid inhibits leukemia cell growth and migration via blocking AKT/mTOR/STAT3 signaling. Int. J. Clin. Exp. Pathol. 8 (5), 5175–5181. [PMC free article] [PubMed] [Google Scholar]
- Hibasami H., Iwase H., Yoshioka K., Takahashi H. (2005). Glycyrrhizin induces apoptosis in human stomach cancer KATO III and human promyelotic leukemia HL-60 cells. Int. J. Mol. Med. 16 (2), 233–236. 10.3892/ijmm.16.2.233 [DOI] [PubMed] [Google Scholar]
- Hostetler B. J., Uchakina O. N., Ban H., McKallip R. J. (2017). Treatment of hematological malignancies with glycyrrhizic acid. Anticancer Res. 37 (3), 997–1004. 10.21873/anticanres.11409 [DOI] [PubMed] [Google Scholar]
- Huang R. Y., Chu Y. L., Jiang Z. B., Chen X. M., Zhang X., Zeng X. (2014). Glycyrrhizin suppresses lung adenocarcinoma cell growth through inhibition of thromboxane synthase. Cell Physiol. Biochem. 33 (2), 375–388. 10.1159/000356677 [DOI] [PubMed] [Google Scholar]
- Huo X., Meng X., Zhang J., Zhao Y. (2020). Hepatoprotective effect of different combinations of 18α-and 18β-Glycyrrhizic acid against CCl4-induced liver injury in rats. Biomed. Pharmacother. 122, 109354. 10.1016/j.biopha.2019.109354 [DOI] [PubMed] [Google Scholar]
- Isbrucker R. A., Burdock G. A. (2006). Risk and safety assessment on the consumption of Licorice root (Glycyrrhiza sp), its extract and powder as a food ingredient, with emphasis on the pharmacology and toxicology of glycyrrhizin. Regul. Toxicol. Pharmacol. 46 (3), 167–192. 10.1016/j.yrtph.2006.06.002 [DOI] [PubMed] [Google Scholar]
- Jain A. S., Dhawan V. V., Sarmento B., Nagarsenker M. S. (2016). In vitro and ex vivo evaluations of lipid anti-cancer nanoformulations: insights and assessment of bioavailability enhancement. AAPS PharmSciTech 17 (3), 553–571. 10.1208/s12249-016-0522-2 [DOI] [PubMed] [Google Scholar]
- Jiang L., Cheng Q., Zhang B., Zhang M. (2016). IL-13 induces the expression of 11βHSD2 in IL-13Rα2 dependent manner and promotes the malignancy of colorectal cancer. Am. J. Transl. Res. 8 (2), 1064–1072. [PMC free article] [PubMed] [Google Scholar]
- Johns C. (2009). Glycyrrhizic acid toxicity caused by consumption of licorice candy cigars. CJEM 11 (1), 94–96. 10.1017/s1481803500010988 [DOI] [PubMed] [Google Scholar]
- Ju S. M., Kim M. S., Jo Y. S., Jeon Y. M., Bae J. S., Pae H. O., et al. (2017). Licorice and its active compound glycyrrhizic acid ameliorates cisplatin-induced nephrotoxicity through inactivation of p53 by scavenging ROS and overexpression of p21 in human renal proximal tubular epithelial cells. Eur. Rev. Med. Pharmacol. Sci. 21 (4), 890–899. [PubMed] [Google Scholar]
- Juin S. K., Ghosh S., Majumdar S. (2020). Glycyrrhizic acid facilitates anti-tumor immunity by attenuating Tregs and MDSCs: an immunotherapeutic approach. Int. Immunopharmacol. 88, 106932. 10.1016/j.intimp.2020.106932 [DOI] [PubMed] [Google Scholar]
- Kanzawa T., Ito H., Kondo Y., Kondo S. (2003). Current and future gene therapy for malignant gliomas. J. Biomed. Biotechnol. 2003 (1), 25–34. 10.1155/S1110724303209013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kao T. C., Shyu M. H., Yen G. C. (2009). Neuroprotective effects of glycyrrhizic acid and 18beta-glycyrrhetinic acid in PC12 cells via modulation of the PI3K/Akt pathway. J. Agric. Food Chem. 57 (2), 754–761. 10.1021/jf802864k [DOI] [PubMed] [Google Scholar]
- Karimi P., Islami F., Anandasabapathy S., Freedman N. D., Kamangar F. (2014). Gastric cancer: descriptive epidemiology, risk factors, screening, and prevention. Cancer Epidemiol. Biomarkers Prev. 23 (5), 700–713. 10.1158/1055-9965.EPI-13-1057 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelloff G. J., Boone C. W., Crowell J. A., Steele V. E., Lubet R., Sigman C. C. (1994). Chemopreventive drug development: perspectives and progress. Cancer Epidemiol. Biomarkers Prev. 3 (1), 85–98. [PubMed] [Google Scholar]
- Khan R., Khan A. Q., Lateef A., Rehman M. U., Tahir M., Ali F., et al. (2013). Glycyrrhizic acid suppresses the development of precancerous lesions via regulating the hyperproliferation, inflammation, angiogenesis and apoptosis in the colon of Wistar rats. PLoS One 8 (2), e56020. 10.1371/journal.pone.0056020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khan R., Rehman M. U., Khan A. Q., Tahir M., Sultana S. (2018). Glycyrrhizic acid suppresses 1,2-dimethylhydrazine-induced colon tumorigenesis in Wistar rats: alleviation of inflammatory, proliferation, angiogenic, and apoptotic markers. Environ. Toxicol. 33 (12), 1272–1283. 10.1002/tox.22635 [DOI] [PubMed] [Google Scholar]
- Khan U., Karmakar B. C., Basak P., Paul S., Gope A., Sarkar D., et al. (2023). Glycyrrhizin, an inhibitor of HMGB1 induces autolysosomal degradation function and inhibits Helicobacter pylori infection. Mol. Med. 29 (1), 51. 10.1186/s10020-023-00641-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim S. W., Jin Y., Shin J. H., Kim I. D., Lee H. K., Park S., et al. (2012). Glycyrrhizic acid affords robust neuroprotection in the postischemic brain via anti-inflammatory effect by inhibiting HMGB1 phosphorylation and secretion. Neurobiol. Dis. 46 (1), 147–156. 10.1016/j.nbd.2011.12.056 [DOI] [PubMed] [Google Scholar]
- Kishimoto K., Hasegawa D., Uemura S., Nakamura S., Kozaki A., Saito A., et al. (2021). Effect of monoammonium glycyrrhizinate on the development of hepatotoxicity after initial intrathecal chemotherapy for leukemia. Anticancer Res. 41 (12), 6231–6236. 10.21873/anticanres.15443 [DOI] [PubMed] [Google Scholar]
- Kiso Y., Tohkin M., Hikino H., Hattori M., Sakamoto T., Namba T. (1984). Mechanism of antihepatotoxic activity of glycyrrhizin. I: effect on free radical generation and lipid peroxidation. Planta Med. 50 (4), 298–302. 10.1055/s-2007-969714 [DOI] [PubMed] [Google Scholar]
- Klein I., Isensee J., Wiesen M. H. J., Imhof T., Wassermann M. K., Muller C., et al. (2023). Glycyrrhizic acid prevents paclitaxel-induced neuropathy via inhibition of OATP-mediated neuronal uptake. Cells 12 (9), 1249. 10.3390/cells12091249 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klein I., Lehmann H. C. (2021). Pathomechanisms of paclitaxel-induced peripheral neuropathy. Toxics 9 (10), 229. 10.3390/toxics9100229 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kobayashi M., Fujita K., Katakura T., Utsunomiya T., Pollard R. B., Suzuki F. (2002). Inhibitory effect of glycyrrhizin on experimental pulmonary metastasis in mice inoculated with B16 melanoma. Anticancer Res. 22 (6C), 4053–4058. [PubMed] [Google Scholar]
- Kohlschutter J., Michelfelder S., Trepel M. (2008). Drug delivery in acute myeloid leukemia. Expert Opin. Drug Deliv. 5 (6), 653–663. 10.1517/17425247.5.6.653 [DOI] [PubMed] [Google Scholar]
- Lee S. J., Kim K. H., Park J. S., Jung J. W., Kim Y. H., Kim S. K., et al. (2007). Comparative analysis of cell surface proteins in chronic and acute leukemia cell lines. Biochem. Biophys. Res. Commun. 357 (3), 620–626. 10.1016/j.bbrc.2007.03.191 [DOI] [PubMed] [Google Scholar]
- Li J. Y., Cao H. Y., Liu P., Cheng G. H., Sun M. Y. (2014a). Glycyrrhizic acid in the treatment of liver diseases: literature review. Biomed. Res. Int. 2014, 872139. 10.1155/2014/872139 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li M., Xiang L., Li Y. (2023). Efficacy and safety of compound glycyrrhizin in the patients with vitiligo: a systematic review and meta-analysis. Expert Rev. Clin. Pharmacol. 16 (6), 601–611. 10.1080/17512433.2023.2213887 [DOI] [PubMed] [Google Scholar]
- Li S., Zhu J. H., Cao L. P., Sun Q., Liu H. D., Li W. D., et al. (2014b). Growth inhibitory in vitro effects of glycyrrhizic acid in U251 glioblastoma cell line. Neurol. Sci. 35 (7), 1115–1120. 10.1007/s10072-014-1661-4 [DOI] [PubMed] [Google Scholar]
- Liang G., Zhu Y., Ali D. J., Tian T., Xu H., Si K., et al. (2020). Engineered exosomes for targeted co-delivery of miR-21 inhibitor and chemotherapeutics to reverse drug resistance in colon cancer. J. Nanobiotechnology 18 (1), 10. 10.1186/s12951-019-0563-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lim E., Vaillant F., Wu D., Forrest N. C., Pal B., Hart A. H., et al. (2009). Aberrant luminal progenitors as the candidate target population for basal tumor development in BRCA1 mutation carriers. Nat. Med. 15 (8), 907–913. 10.1038/nm.2000 [DOI] [PubMed] [Google Scholar]
- Liu J., Pandya P., Afshar S. (2021). Therapeutic advances in oncology. Int. J. Mol. Sci. 22 (4), 2008. 10.3390/ijms22042008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu W., Huang S., Li Y., Li Y., Li D., Wu P., et al. (2018). Glycyrrhizic acid from licorice down-regulates inflammatory responses via blocking MAPK and PI3K/Akt-dependent NF-κB signalling pathways in TPA-induced skin inflammation. Medchemcomm 9 (9), 1502–1510. 10.1039/c8md00288f [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maione F., Minosi P., Di Giannuario A., Raucci F., Chini M. G., De Vita S., et al. (2019). Long-lasting anti-inflammatory and antinociceptive effects of acute ammonium glycyrrhizinate administration: pharmacological, biochemical, and docking studies. Molecules 24 (13), 2453. 10.3390/molecules24132453 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mano Y., Abe K., Takahashi M., Higurashi T., Kawano Y., Miyazaki S., et al. (2023). Optimal administration of glycyrrhizin avoids pharmacokinetic interactions with high-dose methotrexate and exerts a hepatoprotective effect. Anticancer Res. 43 (4), 1493–1501. 10.21873/anticanres.16298 [DOI] [PubMed] [Google Scholar]
- Molyneux G., Geyer F. C., Magnay F. A., McCarthy A., Kendrick H., Natrajan R., et al. (2010). BRCA1 basal-like breast cancers originate from luminal epithelial progenitors and not from basal stem cells. Cell Stem Cell 7 (3), 403–417. 10.1016/j.stem.2010.07.010 [DOI] [PubMed] [Google Scholar]
- Niznansky L., Osinova D., Kuruc R., Hengerics Szabo A., Szoradova A., Masar M., et al. (2022). Natural taxanes: from plant composition to human pharmacology and toxicity. Int. J. Mol. Sci. 23 (24), 15619. 10.3390/ijms232415619 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orazizadeh M., Fakhredini F., Mansouri E., Khorsandi L. (2014). Effect of glycyrrhizic acid on titanium dioxide nanoparticles-induced hepatotoxicity in rats. Chem. Biol. Interact. 220, 214–221. 10.1016/j.cbi.2014.07.001 [DOI] [PubMed] [Google Scholar]
- Penas-Prado M., Armstrong T. S., Gilbert M. R. (2012). Glioblastoma. Handb. Clin. Neurol. 105, 485–506. 10.1016/B978-0-444-53502-3.00004-5 [DOI] [PubMed] [Google Scholar]
- Qamar W., Khan R., Khan A. Q., Rehman M. U., Lateef A., Tahir M., et al. (2012). Alleviation of lung injury by glycyrrhizic acid in benzo(a)pyrene exposed rats: probable role of soluble epoxide hydrolase and thioredoxin reductase. Toxicology 291 (1-3), 25–31. 10.1016/j.tox.2011.10.012 [DOI] [PubMed] [Google Scholar]
- Qin S., Wang H., Yuan R., Li H., Ochani M., Ochani K., et al. (2006). Role of HMGB1 in apoptosis-mediated sepsis lethality. J. Exp. Med. 203 (7), 1637–1642. 10.1084/jem.20052203 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qu B., Xing R., Wang H., Chen X., Ge Q., Peng D., et al. (2017). Multiple effects of magnesium isoglycyrrhizinate on the disposition of docetaxel in docetaxel-induced liver injury. Xenobiotica 47 (4), 290–296. 10.1080/00498254.2016.1185195 [DOI] [PubMed] [Google Scholar]
- Qu L., Chen C., Chen Y., Li Y., Tang F., Huang H., et al. (2019). High-mobility group box 1 (HMGB1) and autophagy in acute lung injury (ALI): a review. Med. Sci. Monit. 25, 1828–1837. 10.12659/MSM.912867 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Radwant M. A., Aboul-Enein H. Y. (2002). The effect of oral absorption enhancers on the in vivo performance of insulin-loaded poly(ethylcyanoacrylate) nanospheres in diabetic rats. J. Microencapsul. 19 (2), 225–235. 10.1080/02652040110081406 [DOI] [PubMed] [Google Scholar]
- Ramkumar V., Mukherjea D., Dhukhwa A., Rybak L. P. (2021). Oxidative stress and inflammation caused by cisplatin ototoxicity. Antioxidants (Basel) 10 (12), 1919. 10.3390/antiox10121919 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rastogi V., Yadav P., Bhattacharya S. S., Mishra A. K., Verma N., Verma A., et al. (2014). Carbon nanotubes: an emerging drug carrier for targeting cancer cells. J. Drug Deliv. 2014, 670815. 10.1155/2014/670815 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruszymah B. H., Nabishah B. M., Aminuddin S., Khalid B. A. (1995). Effects of glycyrrhizic acid on right atrial pressure and pulmonary vasculature in rats. Clin. Exp. Hypertens. 17 (3), 575–591. 10.3109/10641969509037425 [DOI] [PubMed] [Google Scholar]
- Safarzadeh E., Sandoghchian Shotorbani S., Baradaran B. (2014). Herbal medicine as inducers of apoptosis in cancer treatment. Adv. Pharm. Bull. 4 (1), 421–427. 10.5681/apb.2014.062 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Said Salem N. I., Noshy M. M., Said A. A. (2017). Modulatory effect of curcumin against genotoxicity and oxidative stress induced by cisplatin and methotrexate in male mice. Food Chem. Toxicol. 105, 370–376. 10.1016/j.fct.2017.04.007 [DOI] [PubMed] [Google Scholar]
- Santos M. L. C., de Brito B. B., da Silva F. A. F., Botelho A., de Melo F. F. (2020). Nephrotoxicity in cancer treatment: an overview. World J. Clin. Oncol. 11 (4), 190–204. 10.5306/wjco.v11.i4.190 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Selyutina O. Y., Polyakov N. E. (2019). Glycyrrhizic acid as a multifunctional drug carrier - from physicochemical properties to biomedical applications: a modern insight on the ancient drug. Int. J. Pharm. 559, 271–279. 10.1016/j.ijpharm.2019.01.047 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shan Y., Liu P., Zhou Y., Ding X., Liu H., Yang J. (2023). Prenatal sevoflurane exposure impairs the learning and memory of rat offspring via HMGB1-induced NLRP3/ASC inflammasome activation. ACS Chem. Neurosci. 14 (4), 699–708. 10.1021/acschemneuro.2c00620 [DOI] [PubMed] [Google Scholar]
- Shustik C., Bence-Bruckler I., Delage R., Owen C. J., Toze C. L., Coutre S. (2017). Advances in the treatment of relapsed/refractory chronic lymphocytic leukemia. Ann. Hematol. 96 (7), 1185–1196. 10.1007/s00277-017-2982-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smolarczyk R., Cichon T., Matuszczak S., Mitrus I., Lesiak M., Kobusinska M., et al. (2012). The role of Glycyrrhizin, an inhibitor of HMGB1 protein, in anticancer therapy. Arch. Immunol. Ther. Exp. Warsz. 60 (5), 391–399. 10.1007/s00005-012-0183-0 [DOI] [PubMed] [Google Scholar]
- Sohn E. J., Kang D. G., Lee H. S. (2003). Protective effects of glycyrrhizin on gentamicin-induced acute renal failure in rats. Pharmacol. Toxicol. 93 (3), 116–122. 10.1034/j.1600-0773.2003.930302.x [DOI] [PubMed] [Google Scholar]
- Staff N. P., Grisold A., Grisold W., Windebank A. J. (2017). Chemotherapy-induced peripheral neuropathy: a current review. Ann. Neurol. 81 (6), 772–781. 10.1002/ana.24951 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stecanella L. A., Bitencourt A. P. R., Vaz G. R., Quarta E., Silva Junior J. O. C., Rossi A. (2021). Glycyrrhizic acid and its hydrolyzed metabolite 18β-glycyrrhetinic acid as specific ligands for targeting nanosystems in the treatment of liver cancer. Pharmaceutics 13 (11), 1792. 10.3390/pharmaceutics13111792 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Su X., Wu L., Hu M., Dong W., Xu M., Zhang P. (2017). Glycyrrhizic acid: A promising carrier material for anticancer therapy. Biomed. Pharmacother. 95, 670–678. 10.1016/j.biopha.2017.08.123 [DOI] [PubMed] [Google Scholar]
- Sun W., Ding J., Wu K., Ning B. F., Wen W., Sun H. Y., et al. (2011). Gankyrin-mediated dedifferentiation facilitates the tumorigenicity of rat hepatocytes and hepatoma cells. Hepatology 54 (4), 1259–1272. 10.1002/hep.24530 [DOI] [PubMed] [Google Scholar]
- Sung H., Ferlay J., Siegel R. L., Laversanne M., Soerjomataram I., Jemal A., et al. (2021). Global cancer statistics 2020: gLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 71 (3), 209–249. 10.3322/caac.21660 [DOI] [PubMed] [Google Scholar]
- Tsai J. J., Pan P. J., Hsu F. T., Chung J. G., Chiang I. T. (2020). Glycyrrhizic acid modulates apoptosis through extrinsic/intrinsic pathways and inhibits protein kinase B- and extracellular signal-regulated kinase-mediated metastatic potential in hepatocellular carcinoma in vitro and in vivo . Am. J. Chin. Med. 48 (1), 223–244. 10.1142/S0192415X20500123 [DOI] [PubMed] [Google Scholar]
- van der Schoot G. G. F., Westerink N. L., Lubberts S., Nuver J., Zwart N., Walenkamp A. M. E., et al. (2016). Variation in the HFE gene is associated with the development of bleomycin-induced pulmonary toxicity in testicular cancer patients. Eur. J. Cancer 59, 134–141. 10.1016/j.ejca.2016.02.013 [DOI] [PubMed] [Google Scholar]
- van Rossum T. G., Vulto A. G., Hop W. C., Brouwer J. T., Niesters H. G., Schalm S. W. (1999). Intravenous glycyrrhizin for the treatment of chronic hepatitis C: a double-blind, randomized, placebo-controlled phase I/II trial. J. Gastroenterol. Hepatol. 14 (11), 1093–1099. 10.1046/j.1440-1746.1999.02008.x [DOI] [PubMed] [Google Scholar]
- Wakamatsu T., Nakahashi Y., Hachimine D., Seki T., Okazaki K. (2007). The combination of glycyrrhizin and lamivudine can reverse the cisplatin resistance in hepatocellular carcinoma cells through inhibition of multidrug resistance-associated proteins. Int. J. Oncol. 31 (6), 1465–1472. 10.3892/ijo.31.6.1465 [DOI] [PubMed] [Google Scholar]
- Wang H., Ge X., Qu H., Wang N., Zhou J., Xu W., et al. (2020). Glycyrrhizic acid inhibits proliferation of gastric cancer cells by inducing cell cycle arrest and apoptosis. Cancer Manag. Res. 12, 2853–2861. 10.2147/CMAR.S244481 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang H., Wang H., Zhang W., Huang H. J., Liao W. S., Fuller G. N. (2004). Analysis of the activation status of Akt, NFkappaB, and Stat3 in human diffuse gliomas. Lab. Invest. 84 (8), 941–951. 10.1038/labinvest.3700123 [DOI] [PubMed] [Google Scholar]
- Wang Q., Huang Y., Li Y., Zhang L., Tang H., Zhang J., et al. (2022). Glycyrrhizic acid mitigates tripterygium-glycoside-tablet-induced acute liver injury via PKM2 regulated oxidative stress. Metabolites 12 (11), 1128. 10.3390/metabo12111128 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Q. S., Gao L. N., Zhu X. N., Zhang Y., Zhang C. N., Xu D., et al. (2019). Co-delivery of glycyrrhizin and doxorubicin by alginate nanogel particles attenuates the activation of macrophage and enhances the therapeutic efficacy for hepatocellular carcinoma. Theranostics 9 (21), 6239–6255. 10.7150/thno.35972 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Q., Wang J., Wang J., Hong S., Han F., Chen J., et al. (2017). HMGB1 induces lung fibroblast to myofibroblast differentiation through NF‑κB‑mediated TGF‑β1 release. Mol. Med. Rep. 15 (5), 3062–3068. 10.3892/mmr.2017.6364 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Z. Y., Nixon D. W. (2001). Licorice and cancer. Nutr. Cancer 39 (1), 1–11. 10.1207/S15327914nc391_1 [DOI] [PubMed] [Google Scholar]
- Waseem M., Bhardwaj M., Tabassum H., Raisuddin S., Parvez S. (2015). Cisplatin hepatotoxicity mediated by mitochondrial stress. Drug Chem. Toxicol. 38 (4), 452–459. 10.3109/01480545.2014.992437 [DOI] [PubMed] [Google Scholar]
- Wong S. Y., Hynes R. O. (2006). Lymphatic or hematogenous dissemination: how does a metastatic tumor cell decide? Cell Cycle 5 (8), 812–817. 10.4161/cc.5.8.2646 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu C. H., Chen A. Z., Yen G. C. (2015). Protective effects of glycyrrhizic acid and 18β-glycyrrhetinic acid against cisplatin-induced nephrotoxicity in BALB/c mice. J. Agric. Food Chem. 63 (4), 1200–1209. 10.1021/jf505471a [DOI] [PubMed] [Google Scholar]
- Wu X., Wang W., Chen Y., Liu X., Wang J., Qin X., et al. (2018). Glycyrrhizin suppresses the growth of human NSCLC cell line HCC827 by downregulating HMGB1 level. Biomed. Res. Int. 2018, 6916797. 10.1155/2018/6916797 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiong X., Gu L., Wang Y., Luo Y., Zhang H., Lee J., et al. (2016). Glycyrrhizin protects against focal cerebral ischemia via inhibition of T cell activity and HMGB1-mediated mechanisms. J. Neuroinflammation 13 (1), 241. 10.1186/s12974-016-0705-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu C., Liang C., Sun W., Chen J., Chen X. (2018). Glycyrrhizic acid ameliorates myocardial ischemic injury by the regulation of inflammation and oxidative state. Drug Des. Devel Ther. 12, 1311–1319. 10.2147/DDDT.S165225 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang F. H., Zhang Q., Liang Q. Y., Wang S. Q., Zhao B. X., Wang Y. T., et al. (2015). Bioavailability enhancement of paclitaxel via a novel oral drug delivery system: paclitaxel-loaded glycyrrhizic acid micelles. Molecules 20 (3), 4337–4356. 10.3390/molecules20034337 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang G., Li J., Cai Y., Yang Z., Li R., Fu W. (2018). Glycyrrhizic acid alleviates 6-hydroxydopamine and corticosterone-induced neurotoxicity in SH-SY5Y cells through modulating autophagy. Neurochem. Res. 43 (10), 1914–1926. 10.1007/s11064-018-2609-5 [DOI] [PubMed] [Google Scholar]
- Yang T., Lan Y., Cao M., Ma X., Cao A., Sun Y., et al. (2019). Glycyrrhetinic acid-conjugated polymeric prodrug micelles co-delivered with doxorubicin as combination therapy treatment for liver cancer. Colloids Surf. B Biointerfaces 175, 106–115. 10.1016/j.colsurfb.2018.11.082 [DOI] [PubMed] [Google Scholar]
- Yi W., Xiao E., Ding R., Luo P., Yang Y. (2016). High expression of fibronectin is associated with poor prognosis, cell proliferation and malignancy via the NF-κB/p53-apoptosis signaling pathway in colorectal cancer. Oncol. Rep. 36 (6), 3145–3153. 10.3892/or.2016.5177 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yin W., Wang J., Jiang L., James Kang Y. (2021). Cancer and stem cells. Exp. Biol. Med. (Maywood) 246 (16), 1791–1801. 10.1177/15353702211005390 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang H., Huang Q., Zhai J., Zhao Y. N., Zhang L. P., Chen Y. Y., et al. (2015). Structural basis for 18-beta-glycyrrhetinic acid as a novel non-GSH analog glyoxalase I inhibitor. Acta Pharmacol. Sin. 36 (9), 1145–1150. 10.1038/aps.2015.59 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou J. X., Wink M. (2018). Reversal of multidrug resistance in human colon cancer and human leukemia cells by three plant extracts and their major secondary metabolites. Med. (Basel) 5 (4), 123. 10.3390/medicines5040123 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu Z. H., Li X., He L. F., Cai H. F., Ye B., Wu Z. M. (2021). Glycyrrhizic acid, as an inhibitor of HMGB1, alleviates bleomycin-induced pulmonary toxicity in mice through the MAPK and Smad3 pathways. Immunopharmacol. Immunotoxicol. 43 (4), 461–470. 10.1080/08923973.2021.1939371 [DOI] [PubMed] [Google Scholar]
- Zuo J., Meng T., Wang Y., Tang W. (2023). A review of the antiviral activities of glycyrrhizic acid, glycyrrhetinic acid and glycyrrhetinic acid monoglucuronide. Pharm. (Basel) 16 (5), 641. 10.3390/ph16050641 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zuo Z., He L., Duan X., Peng Z., Han J. (2022). Glycyrrhizic acid exhibits strong anticancer activity in colorectal cancer cells via SIRT3 inhibition. Bioengineered 13 (2), 2720–2731. 10.1080/21655979.2021.2001925 [DOI] [PMC free article] [PubMed] [Google Scholar]