

Abstract
Background/Aims:
Exercise training enhances vasodilatation to vascular endothelial growth factor (VEGF165) in collateral-dependent coronary arterioles. Interaction of VEGF receptor 2 (VEGFR-2) and the non-tyrosine-kinase receptor, neuropilin-1 has been reported to potentiate VEGF165-mediated signaling. In the current study, we tested the hypotheses that neuropilin-1 mediates the exercise-enhanced VEGF165-mediated vasodilatation in collateral-dependent arterioles and that neuropilin-1 and/or VEGFR-2 protein levels are increased in these arterioles.
Methods:
Ameroid occluders were surgically placed around the proximal left circumflex coronary artery of miniature swine. Eight weeks after surgery, the animals were randomized into sedentary or exercise training (treadmill run; 5 days/week; 14 weeks) protocols. Coronary arterioles (~100 μm diameter) were isolated from both collateral-dependent and control (left anterior descending) myocardial regions and studied by in vitro videomicroscopy or frozen for immunoblot analysis.
Results:
Exercise-enhanced VEGF165-mediated vasodilatation in collateral-dependent arterioles was reversed by inhibition of the VEGF165-neuropilin-1 interaction. VEGF121, which does not interact with neuropilin-1, induced similar vasodilatation in arterioles from all treatment groups. Immunoblot revealed significantly elevated VEGFR-1, VEGFR-2 and neuropilin-1 protein levels in collateral-dependent arterioles of exercise-trained pigs.
Conclusions:
Neuropilin-1 plays a vital role in the exercise-enhanced VEGF165-mediated vasodilatation of collateral-dependent coronary arterioles and is associated with increased neuropilin-1 receptor protein levels.
Keywords: Coronary artery disease, Coronary microcirculation, Exercise, Vascular endothelial growth
Introduction
Vascular endothelial growth factor (VEGF) is an endothelial-cell-specific mitogen that induces physiologically significant vascular growth and remodeling (neovascularization) under conditions of myocardial and peripheral ischemia [1–5]. VEGF-stimulated neovascularization enhances collateral vessel development and improves regional blood flow and myocardial function in a similar model of ischemic heart disease [6]. VEGF has also been shown to mediate endothelium-dependent vasodilatation in coronary arterioles via stimulation of nitric oxide [7, 8]. We previously reported that exercise training enhances the vasodilatory response to VEGF165 in collateral-dependent coronary arterioles from a porcine model of chronic coronary artery occlusion [9]. This adaptation to exercise training was dependent on nitric oxide, cyclooxygenase and tyrosine kinase receptor activity since the enhanced vasodilatation was abolished by inhibitors of nitric oxide synthase, prostaglandins, and tyrosine kinase [9].
Alternative exon splicing of the VEGF gene results in the generation of multiple isoforms, including VEGF121, VEGF145, VEGF165, VEGF183, VEGF189, and VEGF206. The VEGF121, VEGF165, and VEGF189 isoforms are expressed constitutively in cardiac tissue of numerous species. VEGF165 has been reported to be the predominant isoform in cardiac tissue from humans [10], dogs [11] and pigs [12], while the expression of VEGF189 has been reported to be substantial for hearts from rats, mice and guinea pigs [13, 14]. Interestingly, others [11] suggest that the highly homologous nucleotide sequences between VEGF183 and VEGF189 result in published levels for VEGF189 that are probably representative of a combination of signals from VEGF183 and VEGF189, and thus inflated.
The biological effects of VEGF are mediated through two high-affinity tyrosine-kinase membrane receptors associated with endothelial cells, VEGF receptor 1 (VEGFR-1; Flt-1) and VEGF receptor 2 (VEGFR-2; KDR/Flk-1) [15], although VEGFR-2 has been reported to be the primary mediator of VEGF-induced biological responses in vascular endothelial cells [16]. Furthermore, recent studies indicate that the non-tyrosine-kinase, cell surface receptor, neuropilin-1, enhances the binding of VEGF165 to VEGFR-2 in vascular endothelial cells, increasing the potency of VEGF165 [17–20]. VEGFR-2 and neuropilin-1 do not interact directly but are bridged by VEGF165, forming a ternary complex [21]. Neuropilin-1 does not appear to confer signaling activity in response to VEGF165 when VEGFR-2 is inhibited or absent [18, 19]. Because neuropilin-1 binds exon 7 of VEGF165, the absence of this poly-nucleotide sequence in VEGF121 eliminates the interaction of VEGF121 with neuropilin-1 [18, 22].
Experimental evidence suggests that neuropilin-1 plays a key role in embryonic vessel formation [23, 24] including VEGF-induced endothelial migration and sprout formation [18, 25]. However, the role that neuropilin-1 plays in VEGF165-stimulated vasomotor responsiveness has not been addressed, although studies have revealed that overexpression of neuropilin-1 in embryonic mice is lethal and produces several cardiovascular anomalies, including blood vessel dilation [23, 25]. For the present study, we tested the hypothesis that neuropilin-1 mediates the exercise-enhanced VEGF165-mediated vasodilatation observed in collateral-dependent arterioles and that neuropilin-1 and/or VEGFR-2 receptor protein levels are increased in collateral-dependent arterioles of exercise-trained pigs. The effect of exercise training on VEGF receptor protein levels has not been evaluated previously in ischemic or normally perfused myocardium. Thus, the clinical implications of these novel studies are considerable and may provide important insights into adaptations in signaling pathways associated with the therapeutic intervention of exercise training.
Materials and Methods
Animal Protocols
Animal protocols were approved by Texas A&M University Institutional Animal Care and Use Committee. All protocols and methods conformed to the National Institutes of Health (NIH) Guide for the Care and Use of Laboratory Animals (DHHS Publication No. (NIH) 85–23, Office of Science and Health Reports, Bethesda, Md., USA).
Porcine Model of Chronic Coronary Occlusion and Collateral Development
Chronic coronary occlusion and collateral development were induced in adult female Yucatan miniature swine as previously described [9, 26]. Briefly, animals were sedated with glycopyrrolate (0.004 mg/kg) and midazolam (0.5 mg/kg) after which surgical anesthesia was achieved with ketamine (20 mg/kg). Glycopyrrolate, midazolam, and ketamine were administered intramuscularly (i.m.). Following intubation, anesthesia was maintained with 3% isoflurane and 97% O2 during the sterile surgical procedure. A left lateral thoracotomy was performed and an ameroid occluder (Research Instruments SW, Calif., USA), 2.5–3.5 mm inner diameter, was placed around the proximal left circumflex coronary artery. The diameter of the occluder was selected upon visual inspection of the artery to provide a secure, but nonconstrictive fit. During and following surgery, animals received buprenorphine (0.01 mg/kg i.v.) every 4 h on day of surgery and ketofen (2.2 mg/kg i.m.) in the evening of and day after surgery for pain relief. Antibiotics (Naxcel, 4 mg/kg i.m.) were administered immediately prior to surgery and for 5 days postoperatively. During surgery, pancuronium bromide (0.1 mg/kg), bretylium tosylate (5 mg/kg), lidocaine (1 mg/kg), and atropine (0.05 mg/kg) were administered intravenously as clinically indicated.
Training Procedures
Following 8 weeks of postoperative recovery, pigs were randomly assigned to sedentary or exercise training groups. Animals in both groups were fed once daily and allowed water ad libitum. Exercise-trained animals were subjected to a progressive treadmill exercise program, 5 days/week for 14 weeks, used extensively by our laboratory [9, 26, 27], while sedentary animals remained confined to their pens. By week 12 of the progressive exercise program, animals were running 85 min/day, 5 days/week as described in detail previously [26].
Training Efficacy/Oxidative Enzyme Capacity
Effectiveness of the exercise training program was determined by comparing skeletal muscle oxidative enzyme capacity and heart-to-body weight ratio of the exercise-trained and sedentary animals. Following euthanasia of the animals, samples were taken from the deltoid and triceps brachii (long, medial and lateral heads) muscles, frozen in liquid N2, and stored at −80°C until processed. Citrate synthase activity was measured from whole muscle homogenate using spectrophotometric analysis as previously described [9, 26, 27].
Isolated Microvessel Preparation
Following completion of the protocols, the animals were anesthetized with ketamine (30 mg/kg i.m.) and pentobarbital sodium (35 mg/kg i.v.). The heart was removed via left lateral thoracotomy and placed in ice-cold (0–4°C) Krebs bicarbonate buffer. Myocardial tissue samples were obtained from both the collateral-dependent region of the left ventricular wall and a control region from the left ventricular wall perfused by the nonoccluded left anterior descending coronary artery. Tissue samples were placed in a chilled (4°C) dissection chamber containing physiological saline solution (PSS).
Subepicardial arteriolar branches (~100 μm internal diameter; ~1–1.5 mm axial length) were dissected from surrounding myocardium, and transferred to a Lucite chamber containing PSS-albumin equilibrated to room temperature (~25°C). Each end of the arteriole was cannulated with a glass micropipette (~40 μm diameter) and secured with 11–0 ophthalmic suture (Alcon). The chamber was transferred to an inverted microscope (Olympus IX70) for visualization. The microscope was coupled to a video camera (Panasonic BP310), a CCTV monitor (Panasonic), a video micrometer (Microcirculation Research Institute, TAMUSHSC) for video dimensional analysis, and a data acquisition system (Macintosh/MacLab).
Each micropipette was connected to a hydrostatic pressure reservoir containing filtered PSS-albumin and adjusted to set the intraluminal pressure of the arteriole at 40 mm Hg without allowing flow through the vessel lumen. Leaks were detected by pressurizing the arteriole to 40 mm Hg, then verifying that the intraluminal diameter remained constant when the valve to the reservoir system was closed. Only arterioles that were free of leaks were studied. The vessel chamber bath (PSS-albumin) was gradually warmed and maintained at 37°C for the duration of the experiment. Luminal diameter was monitored continuously throughout the experiment.
Experimental Protocols
Arterioles underwent a 1-hour equilibration period at 40 mm Hg during which time the vessels established a stable basal tone. Arterioles were further preconstricted with endothelin-1 until a preconstriction level of ~20–60% maximal diameter was attained. Once the arteriolar diameter stabilized, VEGF concentration-response relationships were determined by cumulative additions of concentrated stock solutions directly to the tissue bath. VEGF concentration was increased when the response to the previous concentration had stabilized. Arteriolar responses to VEGF165 and VEGF121, as well as VEGF165 in the presence of a synthetic peptide inhibitor (details in Drugs and Solutions) designed to inhibit the interaction of VEGF165 and neuropilin-1 were assessed. While a subset of the VEGF165 data have been published previously [4], these data were collected as part of a large study that included collection of the VEGF121 and VEGF165 plus peptide inhibitor data reported in this paper and thus are most appropriate for inclusion in this study.
Since vessels subjected to repeated exposures of VEGF may exhibit tachyphylaxis [8, 28], each arteriole underwent only a single concentration-response curve to VEGF, either in the absence or presence of selective inhibitors. For experiments in which an inhibitor was present, arterioles were incubated with the inhibitor until a steady-state diameter was attained (~20 min). At completion of the experimental protocol, maximal (passive) intraluminal diameter of the arteriole in response to sodium nitroprusside (100 μm) was measured at 40 mm Hg intraluminal pressure.
Immunoblots
Arterioles (~100 μm diameter; ~12–15 mm total length) were isolated from both the nonoccluded and collateral-dependent myocardial regions, quick-frozen and stored at −80 °C for later immunoblot analysis of VEGFR-1, VEGFR-2, phosphorylated VEGFR-2 (pTyr1175) and neuropilin-1. Arterioles were homogenized in 25 μl of 2× sample buffer (Tris-Cl, pH 6.8, 126 mm; glycerol, 12.6%; 2-mercaptoethanol, 1.44 m; bromophenol blue 0.004%; SDS 5%) by repetitive freeze-thaw cycles and vortex. Arteriole lysate (5 μg total protein) was subjected to SDS-polyacrylamide gel electrophoresis (7.5% gel) and transferred to polyvinylidene fluoride membranes. Membranes were cut and blocked for 2 h at 25°C in Tris-buffered saline containing 5% non-fat dry milk and 0.1% Tween-20, and incubated overnight at 4°C in blocking buffer with primary VEGFR-1 (sc-316; 1:200), VEGFR-2 (sc-504; 1:750), neuropilin-1 (sc-5541; 1:100) (all from Santa Cruz Bio-technology), pVEGFR-2 (pTyr1175; Cell Signaling Technology No. 2478; 1:250), and β-actin (Novus Biologicals NB600–501; 1:10,000) antibodies. After washing, membranes were incubated with the appropriate horseradish-peroxidase-conjugated species-specific anti-IgG (VEGFR-1, VEGFR-2 and pVEGFR-2, 1:20,000; neuropilin-1 and β-actin, 1: 50,000) for 2 h at 25°C. Peroxidase activity was detected using SuperSignal West Dura Substrate (Pierce). Protein bands were quantified using a Luminescent Image Analyzer and Multigauge software 2.3 (LAS-3000; Fuji Photo Film Company, Japan). Normalization for potential loading differences was accomplished using ratios of the densitometry signals for proteins of interest to β-actin.
Drugs and Solutions
The presence of exon 6 of placenta growth factor (PlGF)152 and exon 7 of VEGF165 are required for the ability of these respective growth factors to bind neuropilin-1 [22, 29]. Because of the relative homology between exon 6 of PlGF152 and exon 7 of VEGF165, a synthetic peptide corresponding to the first 16 amino acid residues encoded by exon 6 of PlGF152 has been well established to inhibit the binding of both PlGF152 and VEGF165 to neuropilin-1 [17, 29]. Thus, the PlGF152 exon-6 peptide used in these previous studies was synthesized for the present studies by Sigma Genosys (The Woodlands, Tex., USA), dissolved in DMSO as a stock solution, and subsequent dilutions were prepared in PSS; final concentration of DMSO in the vessel bath was 0.01%. Vehicle control studies indicated that this final concentration of DMSO did not alter resting diameter, vessel tone or vasodilator function. Data from the literature indicate that the concentration of peptide inhibitor (10 μM) used in our studies effectively and specifically inhibits the VEGF165-neuropilin interaction without inhibition of the binding of VEGF165 to VEGFR-2, as observed at higher concentrations [17]. VEGF165 and VEGF121 were purchased from R&D Systems and prepared in PSS-albumin.
Chemicals were obtained from Sigma unless indicated otherwise. PSS solution used in all isolated arteriole studies contained (mM): 145 NaCl, 4.7 KCl, 2.0 CaCl2, 1.17 MgSO4, 1.2 NaH2 PO4, 5.0 glucose, 2.0 pyruvate, 0.02 EDTA and 3.0 Mops buffer. For PSS-albumin, 1% bovine serum albumin (US Biologicals) was added to PSS. Krebs solution contained (mM): 118.3 NaCl, 4.7 KCl, 2.5 CaCl2, 1.2 MgSO4, 1.2 KH2 PO4, 25 NaHCO3, and 11.1 glucose, was maintained at 4°C and aerated with 95% O2 and 5% CO2. All solutions were adjusted to pH 7.4.
Data Analysis
Citrate synthase activity of skeletal muscle and heart-to-body weight ratios were compared using Student’s unpaired t test. Vasodilatory responses to VEGF are presented as the percentage increase in internal diameter relative to the maximal possible dilation [(DSS – DB)/(DP – DB)] 100, where DB is the endothelin-pre-constricted baseline diameter, DSS is the steady-state diameter attained at each concentration of VEGF, and DP is the passive (maximal) diameter of the arteriole. When more than one coronary arteriole from the collateral-dependent or nonoccluded regions of a given animal was used in identical protocols, the responses from these vessels were averaged before data analyses were conducted. VEGF concentration-response curves of arterioles were analyzed using two-way repeated measures analysis of variance. Fisher’s test for least significant difference was used to discriminate for differences between individual points. Immunoblot data were compared using one-way analysis of variance and planned orthogonal contrasts for post-hoc analysis [30]. For all analyses, significance was defined as p ≤ 0.05. Data are represented as mean ± standard error of the mean.
Results
Training Efficacy
The effectiveness of the 14-week exercise training program was demonstrated by significant increases in both skeletal muscle oxidative capacity and heart-to-body weight ratio in exercise-trained compared with sedentary animals. Citrate synthase activity was increased (p < 0.05) in the long (26.5 ± 1.9 vs. 19.0 ± 1.4 μmol/min/g), medial (36.5 ± 1.6 vs. 23.7 ± 1.7 μmol/min/g) and lateral (26.7 ± 1.8 vs. 18.9 ± 0.9 μmol/min/g) heads of the triceps brachii muscle and the deltoid muscle (30.1 ± 1.0 vs. 22.7 ± 1.0 μmol/min/g) in exercise-trained (n = 24) compared with sedentary (n = 23) animals, respectively. Heart-to-body weight ratio also increased (p < 0.05) in exercise-trained (n = 24) compared with sedentary (n = 24) pigs (5.79 ± 0.14 vs. 4.78 ± 0.13 g/kg).
Characteristics of Arterioles
The characteristics of the coronary arterioles from each treatment group are summarized in table 1. Maximal (passive) intraluminal diameters of cannulated coronary arterioles measured at 40 mm Hg intraluminal pressure in the presence of 100 μM nitroprusside were not significantly different between treatment groups. The level of preconstriction (% of maximal intraluminal diameter) was similar between coronary arterioles of all treatment groups. The concentration of endothelin-1 required to attain this level of preconstriction was not significantly different between groups.
Table 1.
Characteristics of coronary arterioles
| Treatment group | Maximal diameter μm | Endothelin [log M] | Preconstriction % |
|---|---|---|---|
|
| |||
| LAD (control) | |||
| SED (n = 26) | 113 ± 6 | −9.94 ± 0.05 | 38 ± 4 |
| EX (n = 28) | 113 ± 7 | −9.91 ± 0.07 | 42 ± 3 |
| LCX (Coll-dep) | |||
| SED (n = 24) | 119 ± 6 | −9.86 ± 0.07 | 39 ± 4 |
| EX (n = 29) | 123 ± 6 | −9.81 ± 0.08 | 49 ± 4 |
LAD = Left anterior descending artery; LCX = left circumflex artery; Coll-dep = collateral-dependent; SED = sedentary; EX = exercise-trained. Values are means 8 SEM. n = number of arterioles studied. No significant differences exist.
VEGF165- and VEGF121-Mediated Concentration-Response Curves
Concentration-response curves for VEGF165 and VEGF121 were compared in arterioles isolated from the collateral-dependent region distal to chronic occlusion and from the nonoccluded region of sedentary and exercise-trained animals. These data demonstrate that arterioles from the collateral-dependent region of exercise-trained animals display significantly enhanced VEGF165-mediated vasodilatation compared with collateral-dependent arterioles from sedentary animals and arterioles from the nonoccluded region in both sedentary and exercise-trained animals (fig. 1A). Interestingly, VEGF165-mediated dilatation was not altered by occlusion or exercise training alone since arterioles from both the collateral-dependent region of sedentary animals and the nonoccluded region of exercise-trained animals responded similarly to arterioles of the nonoccluded region of sedentary pigs.
Fig. 1.
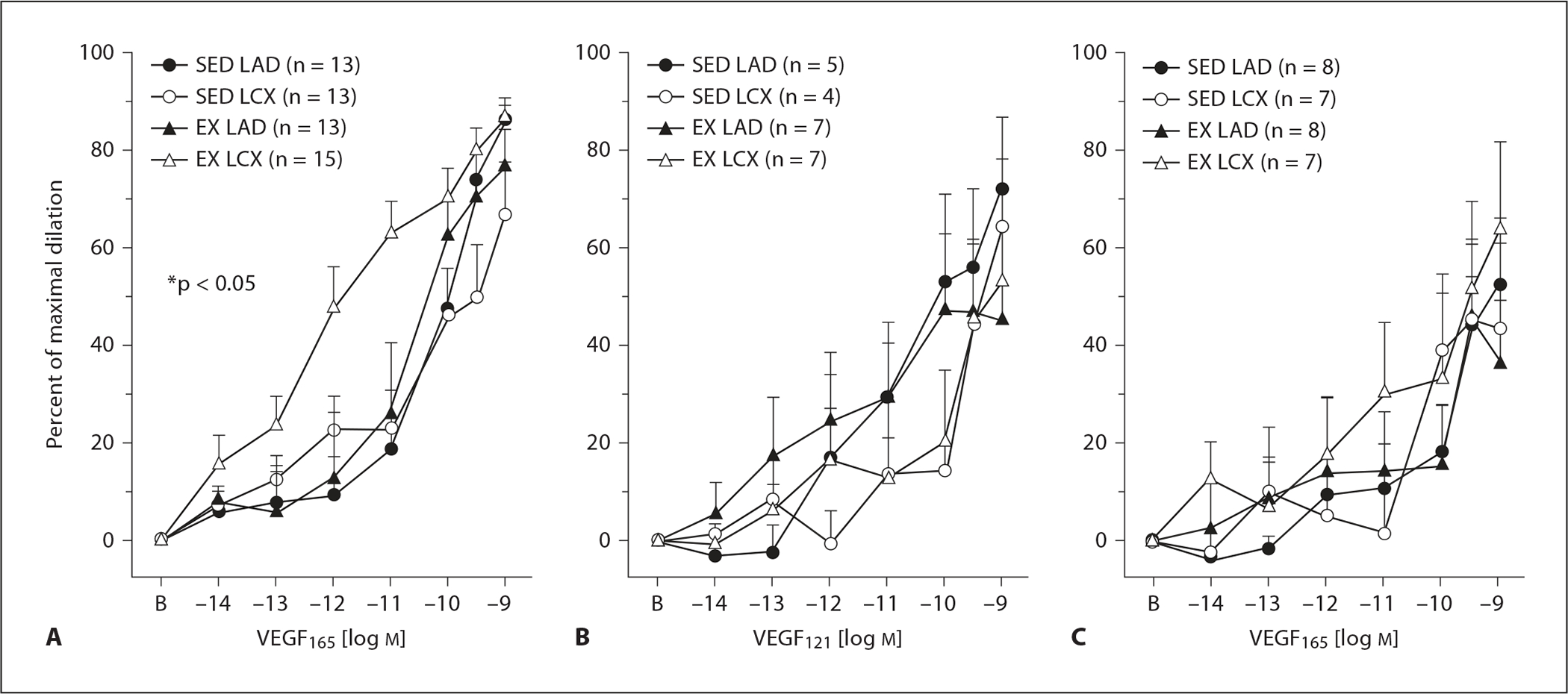
VEGF-mediated vasodilatation responses of coronary arterioles isolated from collateral-dependent and nonoccluded myocardial regions of hearts subjected to chronic occlusion. A VEGF165-induced vasodilatation was significantly enhanced in collateral-dependent arterioles of exercise-trained compared with other treatment groups. B VEGF121-mediated vasodilatation was not altered by chronic occlusion and/or exercise training. C Enhanced VEGF165-mediated vasodilatation in the collateral-dependent arterioles of exercise-trained pigs was abolished in the presence of a synthetic peptide inhibitor of the VEGF165-neuropilin-1 interaction. SED = Sedentary; EX = exercise-trained; LAD = arterioles from nonoccluded control region; LCX = arterioles from collateral-dependent region. Values are means ± SEM of the number of animals in parentheses. * Significantly different from other treatment groups.
In contrast to our findings with VEGF165, the vasodilatory response to VEGF121 was not enhanced in the collateral-dependent region of exercise-trained animals (fig. 1B). VEGF121-mediated vasodilatation tended to be modestly attenuated in arterioles from the collateral-dependent region compared with those from the nonoccluded region of both sedentary and exercise-trained pigs, but these differences were not statistically significant. The lack of an observed increase in VEGF121-mediated dilatation in the collateral-dependent arteriole of exercise-trained swine compelled us to examine the role of neuropilin-1 in VEGF165-mediated vasodilatation.
Role of Neuropilin-1 in VEGF165-Mediated Vasodilatation
To determine the contribution of neuropilin-1 to VEGF165-mediated vasodilatation, we utilized the synthetic peptide inhibitor of VEGF165-neuropilin-1 binding. Treatment of arterioles with the peptide inhibitor did not alter the basal tone or the endothelin-1 concentration required for preconstriction. Inhibition of the VEGF165-neuropilin-1 interaction significantly attenuated VEGF165-induced vasodilatation of arterioles from all treatment groups (compare fig. 1A and C). Importantly, the attenuation of VEGF165-mediated vasodilatation was most profound in collateral-dependent arterioles of exercise-trained pigs so that the difference between the collateral-dependent arterioles of exercise-trained animals and the other treatment groups observed in figure 1A was abolished after inhibition of the VEGF165-neuropilin-1 interaction (fig. 1C). The significant attenuation of VEGF165-mediated dilatation in the presence of the synthetic peptide was statistically similar among arterioles from the nonoccluded region of exercise-trained animals and both the nonoccluded and collateral-dependent region of sedentary animals.
VEGFR-1, VEGFR-2, pVEGFR-2 and Neuropilin-1 Receptor Protein Levels
Immunoblot analysis identified two proteins in coronary arteriolar lysate that reacted with the VEGFR-2 antibody. Previous data examining VEGFR-2 protein suggests that these are an immature, nonglycosylated soluble form (~150 kDa) and a mature, fully glycosylated cell surface form (~230 kDa) [31] of VEGFR-2. Densitometric analysis of receptor protein levels revealed that collateral-dependent arterioles of exercise-trained animals exhibit significantly increased neuropilin-1 (fig. 2B) and membrane and soluble forms of VEGFR-2 (fig. 2C, D). In contrast, pVEGFR-2 (pTyr1175) was not altered by occlusion or exercise training (fig. 2E). VEGFR-1 protein levels were also significantly increased in collateral-dependent arterioles of exercise-trained animals compared with other treatment groups (fig. 2F) but to a lesser extent than VEGFR-2 and neuropilin-1.
Fig. 2.
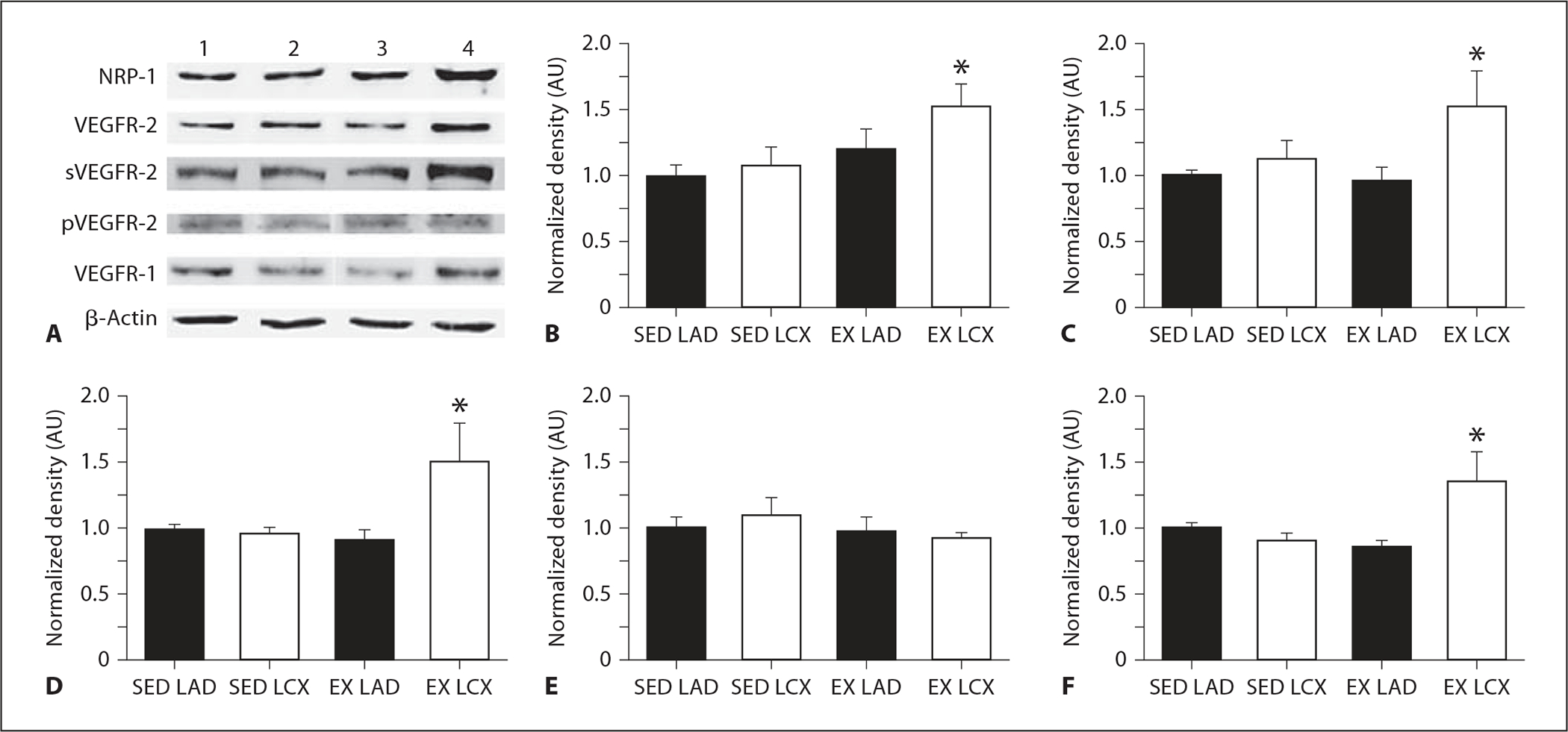
Effect of chronic occlusion and exercise training on neuropilin-1, VEGFR-2, pVEGFR-2 and VEGFR-1 receptor protein levels as determined by immunoblot. Proteins of interest were quantified by densitometry analysis, normalized to β-actin, and expressed relative to SED LAD density. SED = Sedentary; EX = exercise-trained; LAD = arterioles from nonoccluded control region; LCX = arterioles from collateral-dependent region. Values are means ± SEM from arterioles of sedentary (n = 6) and exercise-trained (n = 6) animals. * Significantly different from other treatment groups. A Representative immunoblot images for proteins of interest from arterioles of 1 sedentary and 1 exercise-trained animal. Lanes 1–4 are SED LAD, SED LCX, EX LAD, and EX LCX, respectively. Protein levels for neuropilin-1 (B), cell surface VEGFR-2 (C), the soluble, immature form of VEGFR-2 (D), pVEGFR-2 (pY1175; E), and VEGFR-1 (F).
Discussion
These studies provide the first evidence that neuropilin-1 contributes to VEGF165-mediated vasodilatation in any type of vasculature. Specifically, we demonstrated that inhibition of the VEGF165-neuropilin-1 interaction significantly attenuated VEGF165-induced vasodilatation of both nonoccluded and collateral-dependent coronary arterioles from sedentary and exercise-trained animals. Importantly, the attenuation of VEGF165-mediated vasodilatation was most profound in collateral-dependent arterioles of exercise-trained pigs so that the observed difference between the collateral-dependent arterioles of exercise-trained animals and the other treatment groups was abolished after inhibition of the VEGF165-neuropilin-1 interaction. These data suggest that together, chronic occlusion and exercise training stimulate an adaptive response which enhances the role of neuropilin-1 in VEGF165-mediated vasodilatation of coronary arterioles; an adaptation not observed with occlusion or exercise training alone.
In contrast to our findings with VEGF165, the vasodilatory response to VEGF121 was not altered by chronic occlusion and/or exercise training. These data indicate that the exercise training-enhanced VEGF-induced vasodilatation in the underlying setting of chronic occlusion is specific to VEGF165. Although VEGF121 and VEGF165 have similar affinity for VEGFR-2, biochemical assay indicates that VEGF165 is a more potent activator of VEGFR-2-mediated biological activity [32, 33]. This higher potency of VEGF165 at VEGFR-2 has been attributed to the formation of ternary complexes containing neuropilin-1, VEGFR-2 and VEGF165, which markedly increases VEGF165 binding to VEGFR-2 [17, 19, 20]. Furthermore, neuropilin-1 does not bind VEGF121 due to the absence of the neuropilin-binding domain in VEGF121 [18, 33]. It is interesting that despite an increase in VEGFR-2 protein, VEGF121-mediated vasodilatation was not augmented in the collateral-dependent arterioles of exercise-trained swine. This finding is difficult to interpret, but taken together with our finding that the enhanced VEGF165-mediated dilatation was abolished by inhibition of the VEGF165-neuropilin-1 interaction despite elevated VEGFR-2 protein, suggests that the increased VEGFR-2 protein alone was not sufficient to enhance VEGF-mediated vasodilatation without the potentiating effects of neuropilin-1.
We also report the novel finding that VEGFR-2 and W neuropilin-1 protein levels are significantly increased in collateral-dependent arterioles of exercise-trained pigs. These studies are the first to examine the effect of exercise training and/or chronic occlusion on protein levels for VEGFR-2 and neuropilin-1 in the coronary microvasculature. Our data suggest that increased receptor protein levels contribute to the enhanced VEGF165-mediated vasodilatation observed in the collateral-dependent arterioles of exercise-trained animals. Chronic ischemia in rat skeletal muscle has been shown to produce increased VEGFR-2 protein levels 1 week after iliac artery ligation, which progressively recovered to below control levels 5 weeks after ligation [34]. However, subsequent muscular activity, produced by electrical stimulation of the muscle fibers distal to ligation, re-established increased VEGFR-2 protein levels [34], suggesting in our study that stimuli associated with exercise (muscular activity) were necessary to maintain elevated receptor protein levels in tissue distal to occlusion. Whether the effect of exercise training in the collateral-dependent vasculature in our study was associated with altered hemodynamic forces or ischemia secondary to the increased blood flow demand with each exercise bout is unknown. Indeed, clinical and experimental studies have documented that both diseased patients and animal models display persistent myocardial ischemia and dysfunction in the collateral-dependent region during exercise [35–37]. Our laboratory has not completed studies to evaluate the assertion that myocardial ischemia persists in the collateral-dependent region during exercise; however, Roth et al. [37] previously demonstrated in a comparable porcine model of chronic occlusion and exercise training that while adherence to an exercise training program following occlusion produces improvements in collateral blood flow, regional myocardial ischemia and contractile dysfunction during exercise persist.
While inhibition of the neuropilin-1-VEGF165 interaction had the most profound effect in arterioles from the collateral-dependent region of exercise-trained swine, the peptide inhibitor also significantly attenuated VEGF165-mediated vasodilatation in arterioles from the other treatment groups. These data suggest neuropilin-1 plays a role in VEGF165-mediated dilatation in coronary arterioles even under control, nonoccluded conditions. Interestingly, the attenuation was statistically similar in arterioles from the nonoccluded and collateral-dependent regions of sedentary animals and the nonoccluded region of the exercise-trained animals, indicating that the contribution of neuropilin-1 is not altered by occlusion or exercise training alone. It is also intriguing to note that the neuropilin-1 receptor protein levels were very similar among arterioles from these treatment groups, closely corresponding with the comparable functional effect of the peptide inhibitor on the vasodilatory response. Taken together, these data suggest that the enhanced VEGF165-mediated vasodilatation may be attributable primarily to the increased neuropilin-1 receptor protein levels in the collateral-dependent arterioles of exercise-trained animals.
It is interesting to note that we also observed an increase in protein levels of both VEGFR-1 and the immature, soluble form of VEGFR-2 in arterioles isolated from the collateral-dependent region of exercise-trained swine. The function of VEGFR-1 remains a topic of debate, but is proposed to act as a decoy receptor to negatively regulate the activity of VEGF at the vascular endothelium rather than as a mitogenic signal transduction receptor [38]. It is important to note that the relative increase in VEGFR-1 in arterioles of the collateral-dependent region of exercise-trained swine was not as profound as that observed with VEGFR-2 or neuropilin-1. In contrast to a proposed negative role of VEGFR-1 in VEGF signaling, others report that the heterodimerization of VEGFR-1 and VEGFR-2 isoforms and subsequent signal transduction at these receptors produced the most efficient signaling through pathways that ultimately increase nitric oxide production, whereas VEGFR-2 homodimers were less efficient and VEGFR-1 homodimers were least efficient [39]. Taken together with our previous observation that the enhanced VEGF165-mediated vasodilatation in collateral-dependent arterioles of exercise-trained animals is reversed by nitric oxide synthase inhibition [9], our observation that both VEGFR-1 and VEGFR-2 protein levels were increased in arterioles from this region suggests that the formation of VEGFR-1/VEGFR-2 heterodimers may contribute to the observed increase in dilation. Interestingly, VEGF165-induced prostacyclin production requires heterodimerization of VEGFR-1 and VEGFR-2 and is potentiated by the coexpression of neuropilin-1 [40], suggesting the concomitant increase in all three receptor protein levels in our study has the potential to contribute to enhanced VEGF165-mediated vasodilatation.
Similarly, the function of soluble VEGFR-2 has not been fully investigated; however, initial studies suggest that soluble VEGFR-2 retains ligand-binding ability and thereby may act as a natural endogenous inhibitor of VEGF signaling [41]. Indeed, intravenous administration of recombinant adenovirus encoding soluble VEGFR-2 has been shown to significantly attenuate ischemia-induced angiogenesis and associated tissue perfusion in a hindlimb ischemia model [42], as well as reducing VEGF-mediated neovascularization in corneal micropocket assays [43]. On the other hand, these studies determined the effect of circulating soluble VEGFR-2 at concentrations in vast excess of physiological plasma levels [41]. Furthermore, the effect of circulating soluble VEGFR-2 may differ from intracellular soluble VEGFR-2 measured in our studies. The potential role of soluble VEGFR-2 as an endogenous inhibitor of VEGF signaling suggests that the simultaneous increase in both membrane-bound and soluble VEGFR-2 observed in our studies is paradoxical. However, we propose that these exercise-induced adaptations may provide cells in the collateral-dependent myocardium with an intrinsic ability to regulate VEGF signaling; presumably a system of checks and balances. For instance, differences in binding affinities of the receptors for VEGF or cellular localization of the receptors may preferentially regulate the activation of these receptors depending on the local concentration of VEGF.
We previously reported that exercise training-enhanced VEGF165-mediated vasodilatation in collateral-dependent coronary arterioles was reversed by nitric oxide synthase inhibition [9], suggesting that increased nitric oxide production contributed to the enhanced vasodilatory response. Despite the existence of multiple phosphorylation sites on VEGFR-2, phosphorylated Tyr1175 has been shown to interact with signaling pathways that result in the activation of endothelial nitric oxide synthase and subsequent nitric oxide production [44, 45]. While we anticipated that increased pTyr1175 might contribute to the observed increase in VEGF165-mediated dilation, our data suggest that phosphorylated Tyr1175 is not altered by occlusion and/or exercise training under basal conditions. Whether phosphorylation levels of Tyr1175 in response to VEGF165 stimulation would be different after occlusion and exercise training remains to be determined.
Sellke et al. [8] previously reported increased VEGFR-2 mRNA expression as well as enhanced VEGF-mediated vasodilatory responses in collateral-dependent coronary arterioles of untrained pigs approximately 7–9 weeks following ameroid placement. We speculate that our disparate findings in collateral-dependent arterioles of sedentary animals may be attributable to more extensive collateral development in our longer duration protocol (22 weeks) and thus, potentially reduced stimuli for upregulation of the VEGF signaling pathway. It is also interesting to consider whether exercise-enhanced VEGFR-2 and neuropilin-1 protein levels in our study would persist with longer-term exercise training after structural adaptations are complete or if these increased protein levels are necessary to maintain the increased level of capillarity/collateralization.
Limitations
She specific mechanisms by which neuropilin-1 potentiates VEGF165-mediated vasodilatation in the collateral-dependent arterioles from exercise-trained animals remain to be identified. We have previously demonstrated that both nitric oxide and prostaglandins contribute to the exercise-enhanced VEGF165-mediated vasodilatation in arterioles from the collateral-dependent region [9]. Others [40] have reported that the binding of VEGF165 to neuropilin-1 promotes prostacyclin synthesis in bovine aortic endothelial cells, while the effects on nitric oxide production were not evaluated. As neuropilin-1 does not confer signaling activity of its own [18, 19], it is likely that all of the vasodilatory pathways that contribute to VEGF165-mediated vasodilatation also contribute to the potentiation of the VEGF165 conferred by neuropilin-1. Thus, we speculate that both nitric oxide and prostacyclin are cellular mechanisms by which neuropilin-1 potentiates VEGF165-mediated vasodilatation in our studies. Other vasodilatory pathways, such as endothelial-derived hyperpolarization factors, may also contribute.
Conclusions and Clinical Implications
Previous studies have demonstrated a significant role for neuropilin-1 in events associated with VEGF-mediated angiogenesis and vasculogenesis [18, 23, 24, 46]. These studies provided the basis for our interest in determining the role of neuropilin-1 in VEGF-mediated vasodilatation. We provide evidence that coronary arterioles isolated from both nonoccluded and collateral-dependent myocardial regions display significant dependence on neuropilin-1 for VEGF165-induced vasodilatation, with the most marked dependence observed in collateral-dependent arterioles of exercise-trained swine. We propose that this increased contribution of neuropilin-1 is an adaptive response to persistent myocardial ischemia and/or hemodynamic forces in the collateral-dependent myocardium of both diseased patients and animal models during periods of increased metabolic demand such as exercise. Indeed, even following a prolonged exercise training program, subjects continue to display persistent myocardial ischemia and contractile dysfunction during exercise with no additional myocardial necrosis [37, 47]. Taken together, our data suggest that neuropilin-1 contributes significantly to VEGF165-mediated vasodilatation in coronary arterioles from control and collateral-dependent myocardial regions. Furthermore, the observed exercise training-enhanced VEGF165-induced vasodilatation in the collateral-dependent region closely corresponds with increased neuropilin-1 receptor protein levels.
Acknowledgments
The authors greatly appreciate the technical and surgical expertise of Mildred Mattox and the technical contributions of Erin Ashmore. These studies were supported by research funds from the National Institutes of Health, R01-HL064931 (CLH and JLP) and Centers for Disease Control and Prevention, CDC-623086 (GAL).
References
- 1.Lopez JJ, Laham RJ, Stamler A, Pearlman JD, Bunting S, Kaplan A, Carrozza JP, Sellke FW, Simons M: VEGF administration in chronic myocardial ischemia in pigs. Cardiovasc Res 1998;40:272–281. [DOI] [PubMed] [Google Scholar]
- 2.Pearlman JD, Hibberd MG, Chuang ML, Harada K, Lopez JJ, Gladstone SR, Friedman M, Sellke FW, Simons M: Magnetic resonance mapping demonstrates benefits of VEGF-induced myocardial angiogenesis. Nat Med 1995;1:1085–1089. [DOI] [PubMed] [Google Scholar]
- 3.Harada K, Friedman M, Lopez JJ, Wang SY, Li J, Prasad PV, Pearlman JD, Edelman ER, Sellke FW, Simons M: Vascular endothelial growth factor administration in chronic myocardial ischemia. Am J Physiol Heart Circ Physiol 1996;270:H1791–H1802. [DOI] [PubMed] [Google Scholar]
- 4.Banai S, Jaklitsch MT, Shou M, Lazarous DF, Scheinowitz M, Biro S, Epstein SE, Unger EF: Angiogenic-induced enhancement of collateral blood flow to ischemic myocardium by vascular endothelial growth factor in dogs. Circulation 1994;89:2183–2189. [DOI] [PubMed] [Google Scholar]
- 5.Takeshita S, Zheng LP, Brogi E: Therapeutic angiogenesis. A single intraarterial bolus of vascular endothelial growth factor augments revascularization in a rabbit ischemic hind limb model. J Clin Invest 1994;93:662–670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sato K, Wu T, Laham RJ, Johnson RB, Doug-las P, Li J, Sellke FW, Bunting S, Simons M, Post MJ: Efficacy of intracoronary or intravenous VEGF165 in a pig model of chronic myocardial ischemia. J Am Coll Cardiol 2001;37:616–623. [DOI] [PubMed] [Google Scholar]
- 7.Métais C, Li JY, Li J, Simons M, Sellke FW: Effects of coronary artery disease on expression and microvascular response to VEGF. Am J Physiol Heart Circ Physiol 1998;44:H1411–H1418. [DOI] [PubMed] [Google Scholar]
- 8.Sellke FW, Wang SY, Stamler A, Lopez JJ, Li J, Simons M: Enhanced microvascular relaxations to VEGF and bFGF in chronically ischemic porcine myocardium. Am J Physiol 1996;271:H713–H720. [DOI] [PubMed] [Google Scholar]
- 9.Fogarty JA, Muller-Delp JM, Delp MD, Mattox ML, Laughlin MH, Parker JL: Exercise training enhances vasodilation responses to vascular endothelial growth factor in porcine coronary arterioles exposed to chronic coronary occlusion. Circulation 2004;109:664–670. [DOI] [PubMed] [Google Scholar]
- 10.De Boer RA, Henning RH, Tio RA, Pinto YM, Brouwer RMHJ, Ploeg RJ, Bohm M, Van Gilst WH, Van Veldhuisen DJ: Identification of a specific pattern of downregulation in expression of isoforms of vascular endothelial growth factor in dilated cardiomyopathy. Heart 2002;88:412–414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Jingjing L, Srinivasan B, Bian X, Downey HF, Roque RS: Vascular endothelial growth factor is increased following coronary artery occlusion in the dog heart. Mol Cell Biochem 2000;214:23–30. [DOI] [PubMed] [Google Scholar]
- 12.Sharma HS, Tang ZH, Gho BCG, Verdouw PD: Nucleotide sequence and expression of the porcine vascular endothelial growth factor. Biochim Biophys Acta 1995; 1260:235–238. [DOI] [PubMed] [Google Scholar]
- 13.Ng YS, Rohan R, Sunday ME, Demello DE, D’Amore PA: Differential expression of VEGF isoforms in mouse during development and in the adult. Dev Dyn 2001;220:112–121. [DOI] [PubMed] [Google Scholar]
- 14.LaDoux A, Frelin C: Hypoxia is a strong inducer of vascular endothelial growth factor mRNA expression in the heart. Biochem Biophys Res Comm 1993;195:1005–1010. [DOI] [PubMed] [Google Scholar]
- 15.Neufeld G, Cohen T, Gengrinovitch S, Poltorak Z: Vascular endothelial growth factor (VEGF) and its receptors. FASEB J 1999;13:9–22. [PubMed] [Google Scholar]
- 16.Wei W, Jin H, Chen ZW, Zioncheck TF, Yim APC, He GW: Vascular endothelial growth factor-induced nitric oxide- and PGI2 -dependent relaxation in human internal mammary arteries: a comparative study with KDR and Flt-1 selective mutants. J Cardiovasc Pharmacol 2004;44:615–621. [DOI] [PubMed] [Google Scholar]
- 17.Whitaker GB, Limberg BJ, Rosenbaum JS: Vascular endothelial growth factor receptor-2 and neuropilin-1 form a receptor complex that is responsible for the differential signaling potency of VEGF165 and VEGF121. J Biol Chem 2001;276:25520–25531. [DOI] [PubMed] [Google Scholar]
- 18.Soker S, Takashima S, Miao HQ, Neufeld G, Klagsbrun M: Neuropilin-1 is expressed by endothelial and tumor cells as an isoform-specific receptor for vascular endothelial growth factor. Cell 1998;92:735–745. [DOI] [PubMed] [Google Scholar]
- 19.Soker S, Miao HQ, Nomi M, Takashima S, Klagsbrun M: VEGF165 mediates formation of complexes containing VEGFR-2 and neuropilin-1 that enhance VEGF165 -receptor binding. J Cell Biochem 2002;85:357–368. [DOI] [PubMed] [Google Scholar]
- 20.Mac Gabhann F, Popel AS: Differential binding of VEGF isoforms to VEGF receptor 2 in the presence of neuropilin-1: a computational model. Am J Physiol Heart Circ Physiol 2005;288:H2851–H2860. [DOI] [PubMed] [Google Scholar]
- 21.Fuh G, Garcia KC, de Vos AM: The interaction of neuropilin-1 with vascular endothelial growth factor and its receptor flt-1. J Biol Chem 2000;275:26690–26695. [DOI] [PubMed] [Google Scholar]
- 22.Soker S, Fidder H, Neufeld G, Klagsbrun M: Characterization of novel vascular endothelial growth factor (VEGF) receptors on tumor cells that bind VEGF165 via its exon 7-encoded domain. J Biol Chem 1996;271:5761–5767. [DOI] [PubMed] [Google Scholar]
- 23.Kitsukawa T, Shimono A, Kawakami A, Kondoh H, Fujisawa H: Overexpression of a membrane protein, neuropilin, in chimeric mice causes anomalies in the cardiovascular system, nervous system and limbs. Development 1995;121:4309–4318. [DOI] [PubMed] [Google Scholar]
- 24.Kawasaki T, Kitsukawa T, Bekku Y, Matsuda 2Y, Sanbo, Yagi T, Fujisawa H: A requirement for neuropilin-1 in embryonic vessel formation. Development 1999;126:4895–4902. [DOI] [PubMed] [Google Scholar]
- 25.Miao HQ, Lee P, Lin H, Soker S, Klagsbrun M: Neuropilin-1 expression by tumor cells promotes tumor angiogenesis and progression. FASEB J 2000;14:2532–2539. [DOI] [PubMed] [Google Scholar]
- 26.Heaps CL, Sturek M, Rapps JA, Laughlin MH, Parker JL: Exercise training restores adenosine-induced relaxation in coronary arteries distal to chronic occlusion. Am J Physiol Heart Circ Physiol 2000;278:H1984–H1992. [DOI] [PubMed] [Google Scholar]
- 27.Heaps CL, Mattox ML, Kelly KA, Meininger CJ, Parker JL: Exercise training increases basal tone in arterioles distal to chronic coronary occlusion. Am J Physiol Heart Circ Physiol 2006;290:H1128–H1135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lopez JJ, Laham RJ, Carrozza JP, Tofukuji M, Sellke FW, Bunting S, Simons M: Hemodynamic effects of intracoronary VEGF delivery: evidence of tachyphylaxis and NO dependence of response. Am J Physiol Heart Circ Physiol 1997;273:H1317–H1323. [DOI] [PubMed] [Google Scholar]
- 29.Migdal M, Huppertz B, Tessler S, Comforti 2A, Shibuya M, Reich R, Baumann H, Neufeld G: Neuropilin-1 is a placenta growth factor2 receptor. J Biol Chem 1998;273:22272–22278. [DOI] [PubMed] [Google Scholar]
- 30.Field A: Discovering Statistics Using SPSS, ed 2. Thousand Oaks, Sage Publications, 2005. [Google Scholar]
- 31.Takahashi T, Shibuya M: The 230-kDa mature form of KDR/Flk-1 (VEGF receptor-2) activates the PLC-gamma pathway and partially induces mitotic signals in NIH3T3 fibroblasts. Oncogene 1997;14:2079–2089. [DOI] [PubMed] [Google Scholar]
- 32.Keyt BA, Berleau LT, Nguyen HV, Chen H, Heinsohn H, Vandlen R, Ferrara N: The carboxyl-terminal domain (111–165) of vascular endothelial growth factor is critical for its mitogenic potency. J Biol Chem 1996;271:7788–7795. [DOI] [PubMed] [Google Scholar]
- 33.Soker S, Gollamudi-Payne S, Fidder H, Charmahelli H, Klagsbrun M: Inhibition of vascular endothelial growth factor (VEGF)-induced endothelial cell proliferation by a peptide corresponding to exon 7-encoded domain of VEGF165. J Biol Chem 1997;272:31582–31588. [DOI] [PubMed] [Google Scholar]
- 34.Milkiewicz M, Hudlicka O, Verhaeg J, Egg-inton S, Brown MD: Differential expression of Flk-1 and Flt-1 in rat skeletal muscle in response to chronic ischaemia: favourable effect of muscle activity. Clin Sci 2003;105:473–482. [DOI] [PubMed] [Google Scholar]
- 35.Kolibash AJ, Bush CA, Wepsic RA, Schroeder DP, Tetalman MR, Lewis RP: Coronary collateral vessels: spectrum of physiologic capabilities with respect to providing rest and stress myocardial perfusion, maintenance of left ventricular function and protection against infarction. Am J Cardiol 1982;50:230–238. [DOI] [PubMed] [Google Scholar]
- 36.Roth DM, Maruoka Y, Rogers J, White FC, Longhurst JC, Bloor CM: Development of coronary collateral circulation in left circumflex ameroid-occluded swine myocardium. Am J Physiol 1987;25:H1279–H1288. [DOI] [PubMed] [Google Scholar]
- 37.Roth DM, White FC, Nichols ML, Dobbs SL, Longhurst JC, Bloor CM: Effects of chronic exercise on regional myocardial function and coronary collateral development after gradual coronary artery occlusion in pigs. Circulation 1990;82:1778–1789. [DOI] [PubMed] [Google Scholar]
- 38.Park JE, Chen HH, Winer J, Houck KA, Ferrara N: Placenta growth factor. Potentiation of vascular endothelial growth factor bioactivity, in vitro and in vivo, and high affinity binding to Flt-1 but not to Flk-1/KDR. J Biol Chem 1994;269:25646–25654. [PubMed] [Google Scholar]
- 39.Huang K, Andersson C, Roomans GM, Ito N, Claesson-Welsh L: Signaling properties of VEGF receptor-1 and −2 homo- and heterodimers. Int J Biochem Cell Biol 2001;33:315–324. [DOI] [PubMed] [Google Scholar]
- 40.Neagoe PE, Lemieux C, Sirois MG: Vascular endothelial growth factor (VEGF)-A165-induced prostacyclin synthesis requires the activation of VEGF receptor-1 and −2 heterodimer. J Biol Chem 2005;280:9904–9912. [DOI] [PubMed] [Google Scholar]
- 41.Ebos JM, Bocci G, Man S, Thorpe PE, Hicklin DJ, Zhou D, Jia X, Kerbel RS: A naturally occurring soluble form of vascular endothelial growth factor receptor 2 detected in mouse and human plasma. Mol Cancer Res 2004;2:315–326. [PubMed] [Google Scholar]
- 42.Jacobi J, Tam BY, Wu G, Hoffman J, Cooke JP, Kuo CJ: Adenoviral gene transfer with soluble vascular endothelial growth factor receptors impairs angiogenesis and perfusion in a murine model of hindlimb ischemia. Circulation 2004;110:2424–2429. [DOI] [PubMed] [Google Scholar]
- 43.Kuo CJ, Farnebo F, Yu EY, Christofferson R, Swearingen RA, Carter R, von Recum HA, Yuan J, Kamihara J, Flynn E, D’Amato R, Folkman J, Mulligan RC: Comparative evaluation of the antitumor activity of antiangiogenic proteins delivered by gene transfer. Proc Natl Acad Sci USA 2001;98:4605–4610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Takahashi T, Yamaguchi S, Chida K, Shibu - ya M: A single autophosphorylation site on KDR/Flk-1 is essential for VEGF-A-dependent activation of PLC-gamma and DNA synthesis in vascular endothelial cells. EMBO J 2001;20:2768–2778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Holmqvist K, Cross MJ, Rolny C, Hagerkvist R, Rahimi N, Matsumoto T, Claesson-Welsh L, Welsh M: The adaptor protein shb binds to tyrosine 1175 in vascular endothelial growth factor (VEGF) receptor-2 and regulates VEGF-dependent cellular migration. J Biol Chem 2004;279:22267–22275. [DOI] [PubMed] [Google Scholar]
- 46.Miao HQ, Soker S, Feiner L, Alonso JL, Rap-er JA, Klagsbrun M: Neuropilin-1 mediates collapsin-1/semaphorin III inhibition of endothelial cell motility: functional competition of collapsin-1 and vascular endothelial growth factor-165. J Cell Biol 1999;146:233–242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Belardinelli R, Georgiou D, Ginzton L, Cianci G, Purcaro A: Effects of moderate exercise training on thallium uptake and contractile response to low-dose dobutamine of dysfunctional myocardium in patients with ischemic cardiomyopathy. Circulation 1998;97:553–561. [DOI] [PubMed] [Google Scholar]