

Abstract
Objectives:
Numerous genome-wide association studies (GWASs) have identified CACNA1C as one of the top risk genes for schizophrenia. As a necessary post-GWAS follow-up, here, we focused on this risk gene, carefully investigated its novel risk variants for schizophrenia, and explored their potential functions.
Methods:
We analyzed four independent samples (including three European and one African-American) comprising 5,648 cases and 6,936 healthy subjects to identify replicable SNP-schizophrenia associations. The potential regulatory effects of schizophrenia-risk alleles on CACNA1C mRNA expression in 16 brain regions (n=348), gray matter volumes (GMVs) of five subcortical structures (n=34,431), and surface areas (SA) and thickness (TH) of 34 cortical regions (n=36,936) were also examined.
Results:
A novel 17-variant block across introns 36–45 of CACNA1C was significantly associated with schizophrenia in the same effect direction across at least two independent samples (1.8×10−4≤p≤0.049). Most risk variants within this block showed significant associations with CACNA1C mRNA expression (1.6×10−3≤p≤0.050), GMVs of subcortical structures (0.016≤p≤0.048), cortical SA (0.010≤p≤0.050) and TH (0.004≤p≤0.050) in multiple brain regions.
Conclusion:
We have identified a novel and functional risk variant block at CACNA1C for schizophrenia, providing further evidence for the important role of this gene in the pathogenesis of schizophrenia.
Keywords: CACNA1C, schizophrenia, eQTL, gray matter volume, cortical surface area, cortical thickness
1. Introduction
Numerous genome-wide association studies (GWASs) have reported the α−1C subunit of the L-type voltage-gated calcium channel gene (CACNA1C, 12p13.3, Chr12:1948979–2677376 (b36), between CACNA2D4 and ITFG2-AS1) as one of the top risk genes for schizophrenia. Many risk variants at CACNA1C have been associated with schizophrenia, some of which were replicated across multiple independent studies1–12. However, this gene remains to be comprehensively investigated; in particular, the functional roles of most risk variants have yet to be thoroughly explored.
To build on GWAS findings, it is necessary to continue focusing on this risk gene. Recently, we examined 847 SNPs at CACNA1C, and identified a 20-SNP block at intron 3 that was consistently associated with schizophrenia across seven independent cohorts (2.5×10−17≤p≤0.049). This block was also significantly associated with CACNA1C mRNA expression in the brain across three independent European cohorts (5.1×10−12≤p≤8.3×10−3), and could be tagged by the most significant risk SNP, rs1006737 (meta-analysis of 17 studies: p=1.62×10−42)13. Moreover, rs1006737 was found to be significantly negatively associated with the gray matter volume (GMV) of thalamus (p=0.010), the surface area (SA) of isthmus cingulate cortex (p=0.013), and the thickness (TH) of transverse temporal and superior temporal sulcus cortexes (0.005≤p≤0.043)13. These pieces of evidence supported the most promising role of this variant block in schizophrenia risk.
However, CACNA1C is a large gene that is 730kb-long with 58 exons, and might harbor other missing risk variant blocks for schizophrenia. Therefore, in the present study, we aimed to identify those missing risk blocks independent of the previously-reported one, and explore their potential, and maybe distinct, regulatory effects on CACNA1C mRNA expression in the brain, subcortical GMVs, as well as cortical SA and TH, in order to complete the map of all risk variant blocks for schizophrenia across entire CACNA1C.
Usually, a GWAS examines variants across genome unbiasly, and thus is subject to an overly-conservative Bonferronic correction (α=5×10−8) to control for false positives, which may lead to the loss of some important information, e.g., missing critical findings with 5×10−8<p<10−5, because, for most common diseases such as schizophrenia that involves many common variants with small effect sizes, most risk effects of individual variants are difficult to reach this significance level. The present study, as a post-GWAS follow-up, aimed to preserve the information as loosely as possible (α=0.05), but control for false positives as strictly as possible using an alternative strategy, i.e., a series of replication and functional validation steps (detailed below). We examined replicable SNP-schizophrenia associations across multiple cohorts to ensure their robustness, and then demonstrated them to be biological, but not only statistical, associations by examining a series of potential regulatory effects on CACNA1C mRNA expression and brain structure.
2. Materials and Methods
2.1. Subjects
Four independent samples, comprising three European and one African-American population, were used for SNP-schizophrenia association analysis. Sample #1 was sourced from the GAIN dataset (dbGaP access number: phs000021.v3.p2), consisting of 1,350 European patients with schizophrenia and 1,378 healthy European controls. Sample #2 was obtained from the Ashkenazi Jewish dataset (phs000448.v1.p1), comprising 1,044 Jewish patients with schizophrenia and 2,052 healthy Jewish controls. Sample #3 was sourced from the Bulgarian Schizophrenia Trio Sequencing Study (phs000687.v1.p1), consisting of a total of 1826 European parent-offspring trio subjects, of which 621 were offspring with schizophrenia. Sample #4 was sourced from the GAIN dataset (phs000021.v3.p2), comprising 1,195 African-American patients with schizophrenia and 954 healthy African-American controls. Samples #1 and #2 were also examined in the previous study13.
All subjects included in the study were 18 years or older. Affected subjects met DSM-IV criteria for schizophrenia14. Subjects with neurological disorders, substance use disorders, or mental retardation were excluded. Controls were free of schizophrenia, schizoaffective disorder, bipolar disorder, major depressive disorder, and psychotic symptoms such as auditory hallucination and persecutory delusion. Informed written consents were obtained from all subjects before participating in the study, which was reviewed and approved by the Human Investigation Committee of Yale University. Detailed demographic data for these samples have been published in previous studies15–18.
2.2. SNP-disease association analysis
Genotyping, imputation (using IMPUTE219), and data cleaning has been described in details earlier13. The allele frequencies of SNPs were compared between individuals with schizophrenia and controls using the Fisher exact test as implemented in the program PLINK20, to examine the SNP-disease associations.
Although the Bonferroni correction is a commonly used method to correct for multiple comparisons, it may miss some weak but valuable SNP-disease associations. To avoid overly-conservative correction and preserve the information of modest SNP-disease associations, we used a replication and functional validation design. This approach allowed us to control for false positives by requiring the associations to be replicable in the same effect direction across at least two independent cohorts and the risk SNPs to be functionally validated. Only then, we considered them as statistically significant associations if the p-values for associations were close to 0.05.
2.3. cis-acting expression quantitative trait locus (cis-eQTL) analysis
To investigate the potential regulatory effects of schizophrenia-risk variants on the CACNA1C mRNA expression in human postmortem brains, we conducted cis-eQTL analysis on two independent cohorts: a UK European cohort (n=138) (BRAINEAC dataset)21 and a European-American cohort (n=210) (GTEx dataset)22. All subjects were free of significant neurodegenerative and neuropsychiatric disorders.
In the UK European cohort, we analyzed a total of 10 brain regions, including the cerebellar cortex, prefrontal cortex, hippocampus, medulla, occipital cortex, putamen, substantia nigra, temporal cortex, thalamus, and intralobular white matter. In the European-American cohort, we analyzed a total of 11 brain regions, including the basal ganglia (putamen, caudate, nucleus accumbens, and substantia nigra), limbic system [anterior cingulate cortex (BA24), amygdala, hippocampus, and hypothalamus], prefrontal cortex (Brodmann Area or BA9), cerebellum, and cerebellar hemisphere. To compare the normalized mRNA expression levels between different alleles of each variant, we used t-tests.
2.4. Regulatory effect of risk variants on the GMVs of subcortical structures
The potential regulatory effects of schizophrenia-risk variants on the GMVs of several brain regions were analyzed in a European sample (n=34,431; ENIGMA2 dataset)23 using multiple linear regression analysis. Specifically, we focused on the nucleus accumbens, amygdala, brainstem, caudate, and thalamus. All subjects were free of neurodegenerative and neuropsychiatric disorders.
2.5. Regulatory effect of risk variants on cortical surface area and thickness
A total of 36,936 subjects, including 33,992 European (23,909 from 49 ENIGMA cohorts and 10,083 from the UK Biobank) and 2,944 non-European participants (eight cohorts)24, were analyzed. Measures of cortical SA and TH were derived from in vivo whole brain T1-weighted MRI scans using FreeSurfer25. The potential regulatory effects of schizophrenia-risk variants on 70 traits (total SA, average TH, and the SA and TH of 34 cortical regions averaged across right and left hemispheres) were analyzed using multiple linear regression. The analysis adjusted for the effects of sex, linear and nonlinear age effects, interactions between age and sex, ancestry (the first four MDS components), diagnostic status (when the cohort followed a case-control design), MRI acquisition orientation, scanner (when multiple scanners were used at the same site), and global measure (total SA or average TH).
3. Results
3.1. A variant block across introns 36–45 of CACNA1C was significantly associated with schizophrenia across multiple samples
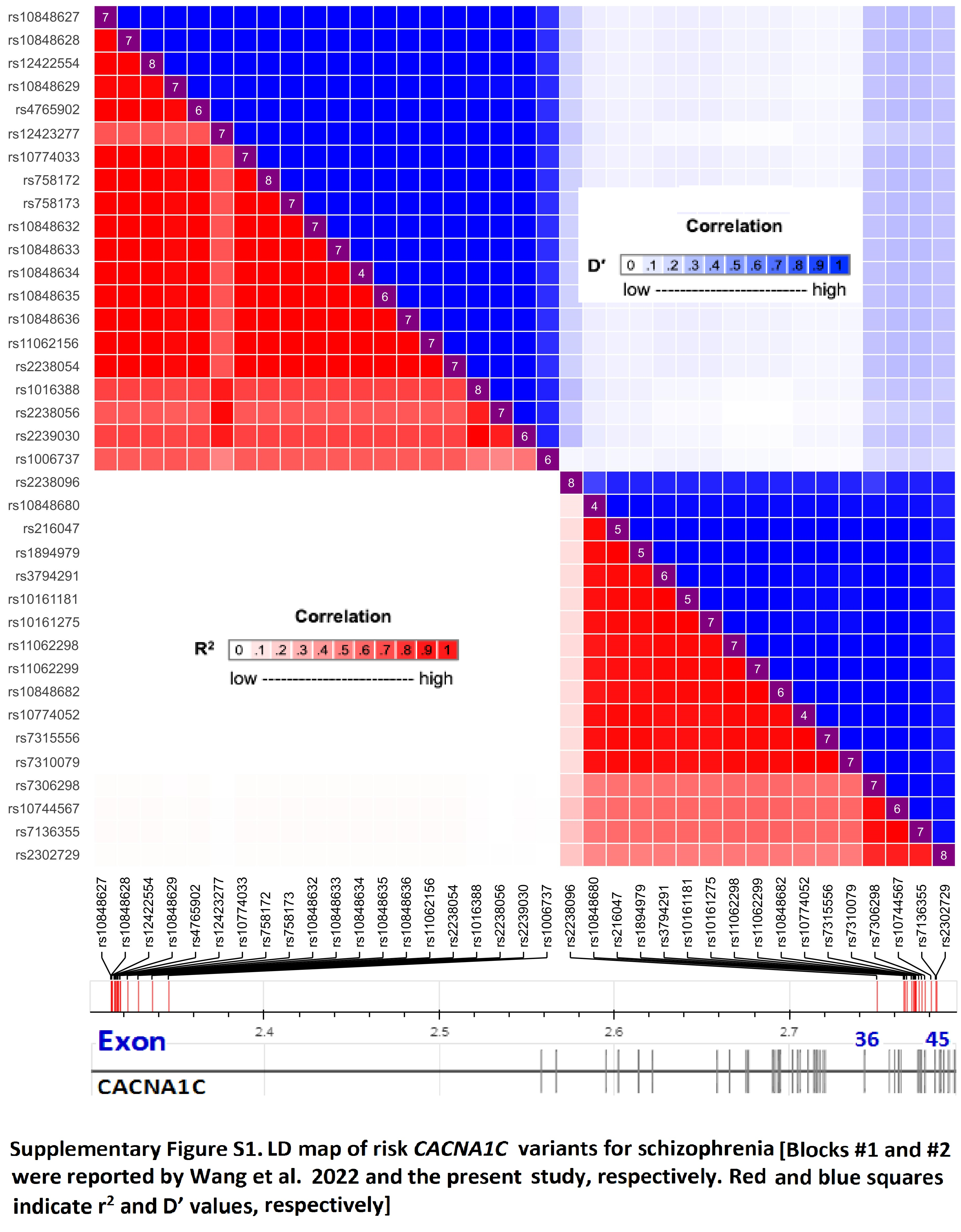
Independent of the previously-reported risk variant block at intron 3 mentioned earlier13, we identified a novel 17-variant block (within block: D’>0.85)13 spanning introns 36–45 of CACNA1C (Supplementary Figure S1). This new variant block was significantly associated with schizophrenia in at least two of the five independent samples we studied (1.8×10−4≤p≤0.049; Table 1). The risk allele of each variant had a significantly higher frequency in the cases (or “transmitted” group) than in the controls (or “untransmitted” group), and this was consistent across the samples. When further defining a block with R2>0.85, this 17-variant block could be classified into three sub-blocks, i.e., classes #1, #2 and #3 harboring one, twelve and four SNPs, respectively (Figure S1; Table 1). Based on the three identified classes of alleles, we were able to distinguish between different functional patterns, as described below.
Table 1.
p values for SNP-schizophrenia and SNP-mRNA expression associations
| “CACNA1C SNP - Schizophrenia” associations. | “SNP - CACNA1C mRNA” associations | Effect | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Europeans | Africans | Europeans | Europeans | direction | |||||||||||||
| Sample #1 | #2 | #3 | #4 | BRAINEAC | GTEx | of risk | |||||||||||
| Genomic | Risk | Case=1351 | 1044 | 621 (offspring) | Risk | 1195 | Effect | CRBL | FCTX | OCTX | TCTX | Effect | AMYL | ACC | or effect | ||
| Class | SNP | position (b36) | allele | Control=1378 | 2052 | 1205 (parents) | allele | 954 | allele | 138 | 138 | 138 | 138 | allele | 210 | 210 | alleles |
| #1 | rs2238096 | 2620174 | a | - | >0.05 | 0.021 | A | 0.047 | a | >0.05 | 1.6×10−3 | 0.022 | 0.050 | - | - | - | ↑↑- |
| #2 | rs10848680 | 2635439 | G | 0.005 | - | 0.020 | - | G | 0.020 | >0.05 | >0.05 | >0.05 | A | 0.050 | >0.05 | ↑↑↓ | |
| #2 | rs216047 | 2636121 | T | 1.8×10−4 | - | 0.013 | - | T | 0.024 | >0.05 | >0.05 | >0.05 | C | 0.035 | >0.05 | ↑↑↓ | |
| #2 | rs1894979 | 2637260 | G | 2.3×10−4 | - | 0.004 | - | G | 0.023 | >0.05 | >0.05 | >0.05 | A | 0.050 | >0.05 | ↑↑↓ | |
| #2 | rs3794291 | 2639926 | C | 0.048 | - | 0.007 | - | C | 0.026 | >0.05 | >0.05 | >0.05 | T | 0.035 | >0.05 | ↑↑↓ | |
| #2 | rs10161181 | 2640991 | A | - | >0.05 | 0.001 | a | 0.011 | A | 0.025 | >0.05 | >0.05 | >0.05 | - | - | - | ↑↑- |
| #2 | rs10161275 | 2641279 | T | 0.046 | >0.05 | 0.009 | - | T | 0.026 | >0.05 | >0.05 | >0.05 | - | - | - | ↑↑- | |
| #2 | rs11062298 | 2641970 | A | 0.039 | - | 0.001 | a | 0.015 | A | 0.029 | >0.05 | >0.05 | >0.05 | C | 0.050 | >0.05 | ↑↑↓ |
| #2 | rs11062299 | 2641999 | T | >0.05 | 0.011 | 0.002 | >0.05 | T | 0.029 | >0.05 | >0.05 | >0.05 | C | 0.050 | >0.05 | ↑↑↓ | |
| #2 | rs10848682 | 2642017 | C | >0.05 | 0.011 | 0.002 | - | C | 0.029 | >0.05 | >0.05 | >0.05 | G | 0.050 | >0.05 | ↑↑↓ | |
| #2 | rs10774052 | 2642432 | C | 0.039 | - | 0.002 | >0.05 | C | 0.036 | >0.05 | >0.05 | >0.05 | - | - | - | ↑↑- | |
| #2 | rs7315556 | 2644116 | G | >0.05 | 0.009 | 0.002 | - | G | 0.037 | >0.05 | >0.05 | >0.05 | A | 0.037 | >0.05 | ↑↑↓ | |
| #2 | rs7310079 | 2645738 | T | >0.05 | 0.027 | 0.044 | - | T | 0.034 | >0.05 | >0.05 | >0.05 | - | - | - | ↑↑- | |
| #3 | rs7306298 | 2647572 | A | - | 0.030 | 0.011 | - | - | - | - | - | - | A | >0.05 | 0.042 | ↑-↑ | |
| #3 | rs10744567 | 2651077 | G | >0.05 | 0.034 | 0.015 | - | - | - | - | - | - | G | >0.05 | 0.020 | ↑-↑ | |
| #3 | rs7136355 | 2653737 | C | - | 0.019 | 0.004 | - | - | - | - | - | - | C | >0.05 | 0.020 | ↑-↑ | |
| #3 | rs2302729 | 2654233 | C | - | 0.028 | 0.020 | >0.05 | - | - | - | - | - | C | >0.05 | 0.012 | ↑-↑ | |
Abbreviations: cerebellar cortex (CRBL), frontal cortex (FCTX), occipital cortex (OCTX), temporal cortex (TCTX), amygdale (AMYL), and anterior cingulate cortex (ACC). The italic bold lowercase risk allele is a major allele (f>0.5). #1, #2 and #3 are classified based on R2>0.85 within each class. ↑ and ↓, increase or decrease risk for disease, or mRNA expression level, referencing to the disease-risk alleles; “-”, missing.
3.2. The schizophrenia-risk alleles were significantly associated with the CACNA1C mRNA expression in the brain
All seventeen risk variants within the variant block showed significant associations with CACNA1C mRNA expression in different brain regions, including the cerebellar, frontal (including anterior cingulate (ACC)), occipital, temporal cortices, and amygdala (1.6×10−3≤p≤0.050). Specifically, the schizophrenia-risk allele of class #1 (including rs2238096) showed significant positive associations with CACNA1C mRNA expression in the frontal, occipital, and temporal cortices (1.6×10−3≤p≤0.050); the schizophrenia-risk alleles of class #2 showed a significant positive association with CACNA1C mRNA expression in the cerebellar cortex, but a negative association with expression in amygdala (0.020≤p≤0.050); and the schizophrenia-risk alleles of class #3 showed a significant positive association with CACNA1C mRNA expression in ACC (0.012≤p≤0.042). (Table 1).
3.3. The schizophrenia-risk alleles were significantly associated with subcortical GMVs
Seven of the seventeen risk variants within the variant block (41%) showed significant association with the GMVs of subcortical structures, including the nucleus accumbens, brainstem, and caudate (0.016≤p≤0.048). Specifically, the schizophrenia-risk allele of class #1 was positively associated with the GMVs of the caudate and brainstem (0.017≤p≤0.040); the schizophrenia-risk alleles of class #2 were negatively associated with the GMVs of the nucleus accumbens (0.034≤p≤0.048); and the schizophrenia-risk alleles of class #3 were positively associated with the GMVs of the nucleus accumbens and brainstem (0.016≤p≤0.028). (Table 2).
Table 2.
p values for SNP-gray matter volumes (GMV) associations in Europeans
| “SNP - GMV” associations | Effect direction | |||||
|---|---|---|---|---|---|---|
| Schizophrenia | Effect | Accumbens | Brainstem | Caudate | risk or effect | |
| SNP | -risk allele | allele | 28,697 | 28,809 | 30,153 | alleles |
| rs2238096 | a | a | >0.05 | 0.017 | 0.040 | ↑↑ |
| rs11062298 | A | C | 0.048 | >0.05 | >0.05 | ↑↓ |
| rs11062299 | T | C | 0.045 | >0.05 | >0.05 | ↑↓ |
| rs10848682 | C | G | 0.042 | >0.05 | >0.05 | ↑↓ |
| rs10774052 | C | T | 0.034 | >0.05 | >0.05 | ↑↓ |
| rs7315556 | G | A | 0.034 | >0.05 | >0.05 | ↑↓ |
| rs2302729 | C | C | 0.028 | 0.018 | >0.05 | ↑↑ |
The italic lowercase risk allele is a major allele (f>0.5). ↑ and ↓, increase or decrease risk for disease or GMV, respectively, referencing to the disease-risk alleles.
3.4. The schizophrenia-risk alleles were significantly associated with the cortical SA
All seventeen risk variants were significantly associated with the cortical SA of various brain regions, including frontal (pole, superior, and rostral middle), parietal (precuneus), temporal (pole), occipital (lateral, pericalcarine, lingual, and fusiform), and limbic [anterior (ACC) and posterior cingulate (PCC)] cortices (0.010≤p≤0.050). Specifically, the schizophrenia-risk allele of class #1 was positively (β>0) associated with the SA of ACC (p=0.043), but negatively (β<0) with the SA of parietal (precuneus) and temporal (pole) cortices, and PCC (0.015≤p≤0.037); the schizophrenia-risk alleles of class #2 were positively (β>0) associated with the SA of parietal (precuneus) cortex and PCC (0.010≤p≤0.050), but negatively (β<0) with the SA of occipital (pericalcarine and fusiform) cortices (0.020≤p≤0.050); and the schizophrenia-risk alleles of class #3 were positively (β>0) associated with the SA of occipital (lateral, pericalcarine, and lingual) cortices (0.017≤p≤0.047), but negatively (β<0) with the SA of frontal (pole, superior, and rostral middle) cortices (0.025≤p≤0.048). (Table 3).
Table 3.
p values for SNP-cortical surface area (SA) associations
| “SNP-cortical SA” associations | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| European | Mixed | Mixed | European | Mixed | Mixed | Mixed | Mixed | European | Mixed | Mixed | Effect | ||||
| Frontal | Parietal | Temporal | Limbic | Occipital | Limbic | direction | |||||||||
| rostral | posterior | peri- | anterior | of risk | |||||||||||
| Disease- | Effect | pole | superior | middle | precuneus | pole | cingulate | Effect | lateral | calcarine | lingual | fusiform | cingulate | or effect | |
| SNP | risk allele | allele | n=32742 | 35198 | 35198 | 32742 | 35198 | 35198 | allele | 35198 | 35198 | 32742 | 35198 | 35198 | alleles |
| rs2238096 | a | C | >0.05 | >0.05 | >0.05 | 0.032 | 0.015 | 0.037 | a | >0.05 | >0.05 | >0.05 | >0.05 | 0.043 | ↑↓↑ |
| rs10848680 | G | G | >0.05 | >0.05 | >0.05 | 0.018 | >0.05 | 0.028 | A | >0.05 | 0.034 | >0.05 | 0.036 | >0.05 | ↑↑↓ |
| rs216047 | T | T | >0.05 | >0.05 | >0.05 | 0.023 | >0.05 | 0.028 | C | >0.05 | 0.041 | >0.05 | 0.037 | >0.05 | ↑↑↓ |
| rs1894979 | G | G | >0.05 | >0.05 | >0.05 | 0.017 | >0.05 | 0.029 | A | >0.05 | 0.036 | >0.05 | 0.033 | >0.05 | ↑↑↓ |
| rs3794291 | C | C | >0.05 | >0.05 | >0.05 | 0.011 | >0.05 | 0.038 | T | >0.05 | 0.039 | >0.05 | 0.020 | >0.05 | ↑↑↓ |
| rs10161181 | A | A | >0.05 | >0.05 | >0.05 | 0.008 | >0.05 | 0.050 | C | >0.05 | 0.031 | >0.05 | 0.022 | >0.05 | ↑↑↓ |
| rs10161275 | T | T | >0.05 | >0.05 | >0.05 | 0.011 | >0.05 | 0.039 | C | >0.05 | 0.034 | >0.05 | 0.023 | >0.05 | ↑↑↓ |
| rs11062298 | A | A | >0.05 | >0.05 | >0.05 | 0.012 | >0.05 | 0.048 | C | >0.05 | 0.024 | >0.05 | 0.032 | >0.05 | ↑↑↓ |
| rs11062299 | T | T | >0.05 | >0.05 | >0.05 | 0.011 | >0.05 | 0.048 | C | >0.05 | 0.039 | >0.05 | 0.036 | >0.05 | ↑↑↓ |
| rs10848682 | C | C | >0.05 | >0.05 | >0.05 | 0.011 | >0.05 | >0.05 | - | - | - | - | - | - | ↑↑- |
| rs10774052 | C | C | >0.05 | >0.05 | >0.05 | 0.010 | >0.05 | >0.05 | T | >0.05 | 0.026 | >0.05 | 0.027 | >0.05 | ↑↑↓ |
| rs7315556 | G | G | >0.05 | >0.05 | >0.05 | 0.013 | >0.05 | 0.028 | A | >0.05 | 0.038 | >0.05 | 0.050 | >0.05 | ↑↑↓ |
| rs7310079 | T | T | >0.05 | >0.05 | >0.05 | 0.011 | >0.05 | >0.05 | C | >0.05 | 0.035 | >0.05 | 0.024 | >0.05 | ↑↑↓ |
| rs7306298 | A | G | >0.05 | >0.05 | 0.045 | >0.05 | >0.05 | >0.05 | A | 0.035 | 0.027 | >0.05 | >0.05 | >0.05 | ↑↓↑ |
| rs10744567 | G | A | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | G | 0.026 | 0.026 | >0.05 | >0.05 | >0.05 | ↑↓↑ |
| rs7136355 | C | T | >0.05 | 0.048 | >0.05 | >0.05 | >0.05 | >0.05 | C | 0.034 | 0.017 | 0.038 | >0.05 | >0.05 | ↑↓↑ |
| rs2302729 | C | T | 0.041 | 0.025 | >0.05 | >0.05 | >0.05 | >0.05 | C | 0.030 | 0.037 | 0.047 | >0.05 | >0.05 | ↑↓↑ |
The italic bold lowercase risk allele (class #1) is a major allele (f>0.5). ↑ and ↓, increase or decrease risk for disease, or effects on SA, respectively, referencing to the disease-risk alleles; “-”, missing.
3.5. The schizophrenia-risk alleles were significantly associated with the cortical TH
Sixteen of the seventeen risk variants were significantly associated with the cortical TH, which includes the frontal (pars opercularis), occipital (lateral, pericalcarine, and fusiform), temporal (middle and inferior), and limbic (entorhinal) cortices (0.004≤p≤0.050). Specifically, the schizophrenia-risk allele of class #1 was positively (β>0) associated with the TH of entorhinal cortex (p=0.039), but negatively (β<0) with the TH of frontal (pars opercularis) cortex (p=0.032); the schizophrenia-risk alleles of class #2 were positively (β>0) associated with the TH of occipital (fusiform), and temporal (middle and inferior) cortices (0.015≤p≤0.049); and the schizophrenia-risk alleles of class #3 were positively (β>0) associated with the TH of occipital (pericalcarine) cortex (0.004≤p≤0.009), but negatively (β<0) with the TH of occipital (lateral), and temporal (inferior) cortices (0.013≤p≤0.050). (Table 4).
Table 4.
p values for SNP-cortical thickness (TH) associations
| “SNP-cortical TH” associations in Europeans | Effect | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Frontal | Occipital | Temporal | Occipital | Limbic | direction | |||||||
| pars | peri- | of risk | ||||||||||
| Disease | Effect | opercularis | lateral | fusiform | middle | inferior | inferior | Effect | calcarine | entorhinal | or effect | |
| SNP | -risk allele | allele | n=9921 | 9921 | 23,909 | 23,909 | 23,909 | 9921 | allele | 23,909 | 9921 | alleles |
| rs2238096 | a | C | 0.032 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | a | >0.05 | 0.039 | ↑↓↑ |
| rs10848680 | G | G | >0.05 | >0.05 | 0.024 | 0.025 | 0.019 | >0.05 | G | >0.05 | >0.05 | ↑↑↑ |
| rs216047 | T | T | >0.05 | >0.05 | 0.031 | 0.029 | 0.019 | >0.05 | T | >0.05 | >0.05 | ↑↑↑ |
| rs1894979 | G | G | >0.05 | >0.05 | 0.033 | 0.031 | 0.023 | >0.05 | G | >0.05 | >0.05 | ↑↑↑ |
| rs3794291 | C | C | >0.05 | >0.05 | 0.050 | 0.044 | 0.018 | >0.05 | C | >0.05 | >0.05 | ↑↑↑ |
| rs10161181 | A | A | >0.05 | >0.05 | >0.05 | 0.049 | 0.015 | >0.05 | A | >0.05 | >0.05 | ↑↑↑ |
| rs10161275 | T | T | >0.05 | >0.05 | 0.049 | 0.042 | 0.015 | >0.05 | T | >0.05 | >0.05 | ↑↑↑ |
| rs11062298 | A | A | >0.05 | >0.05 | 0.038 | 0.037 | 0.030 | >0.05 | A | >0.05 | >0.05 | ↑↑↑ |
| rs11062299 | T | T | >0.05 | >0.05 | 0.038 | 0.037 | 0.030 | >0.05 | T | >0.05 | >0.05 | ↑↑↑ |
| rs10848682 | C | C | >0.05 | >0.05 | 0.040 | 0.042 | 0.032 | >0.05 | C | >0.05 | >0.05 | ↑↑↑ |
| rs10774052 | C | C | >0.05 | >0.05 | 0.036 | 0.048 | 0.047 | >0.05 | C | >0.05 | >0.05 | ↑↑↑ |
| rs7315556 | G | G | >0.05 | >0.05 | 0.037 | >0.05 | >0.05 | >0.05 | G | >0.05 | >0.05 | ↑↑↑ |
| rs7306298 | A | G | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | 0.038 | A | 0.005 | >0.05 | ↑↓↑ |
| rs10744567 | G | A | >0.05 | >0.05 | >0.05 | >0.05 | 0.046 | 0.040 | G | 0.004 | >0.05 | ↑↓↑ |
| rs7136355 | C | T | >0.05 | 0.050 | >0.05 | >0.05 | 0.036 | 0.033 | C | 0.009 | >0.05 | ↑↓↑ |
| rs2302729 | C | T | >0.05 | 0.033 | >0.05 | >0.05 | 0.033 | 0.013 | C | 0.005 | >0.05 | ↑↓↑ |
The italic bold lowercase risk allele (class #1) is a major allele (f>0.5). ↑ and ↓, increase or decrease risk for disease, or effects on TH, respectively, referencing to the disease-risk alleles; “-”, missing.
Discussion
We comprehensively examined a large set (n=847) of CACNA1C SNPs and identified two independent risk variant blocks associated with schizophrenia. The first block (n=20), located in intron 3, has been reported before13, and was significantly associated with schizophrenia across four Psychiatric Genomics Consortium cohorts (PGC) cohorts3,11,12,26 and the GMV of thalamus13. The second block (n=17), located in introns 36–45, is a novel finding and has distinct features from the first one, including that it was not associated with schizophrenia in those four PGC cohorts at all (data not shown), and was associated with the GMVs of multiple regions (see below) other than thalamus. These two blocks were associated with mRNA expression, cortical SA and TH in different brain regions too (see below)13.
We observed significant associations between the seventeen variants within the second risk block and schizophrenia in our samples, with one of the schizophrenia-risk alleles being a major allele (class #1) and all of the others being minor alleles (classes #2 and #3) with distinct functional effects. All risk alleles in three classes were positively associated with CACNA1C mRNA expression in the cerebellar, frontal (including ACC), occipital, and/or temporal cortices, but risk alleles in class #2 were negatively associated with it in amygdala. Risk alleles in classes #1 and #3 were positively associated with the GMVs of nucleus accumbens, brainstem, and/or caudate, but risk alleles in class #2 were negatively associated with the GMVs of nucleus accumbens. Additionally, risk alleles were significantly associated with the SA or TH in multiple regions. These findings suggest that CACNA1C might play a critical role in the pathogenesis of schizophrenia.
Most of the published studies reported reduction of cortical GMVs27–30 and enlargement of basal ganglia GMVs27,31–34 in schizophrenia. Our study found that the schizophrenia-risk alleles of classes #1 and #3 potentially increased the GMVs of caudate and nucleus accumbens, respectively, while potentially decreasing the SA of frontal (pole, superior, and rostral middle), parietal (precuneus), temporal (pole) cortices, and posterior cingulate cortex (PCC), as well as the TH of frontal (pars opercularis), occipital (lateral), and temporal (inferior) cortices. On the other hand, the schizophrenia-risk alleles of class #2 potentially decreased the SA of occipital (pericalcarine and fusiform) cortices. These findings are consistent with literatures, suggesting that CACNA1C might be implicated in schizophrenia via regulating these regional morphometrics.
The caudate is a critical component of the cognitive/associative pathways and one of the main input nuclei of the basal ganglia in the cortico-basal ganglia-thalamic loop. It receives inputs from various cortical structures35. The dorsolateral caudate receives projections from the dorsolateral prefrontal cortex, including the rostral middle frontal gyrus, and projects to the lateral dorsomedial globus pallidus (GPi) and rostral substantia nigra (SNr) where signals are relayed to the lateral ventral anterior nucleus pars magnocellularis (VAmc) and medial pars magnocellularis of the thalamus. This circuit is involved in executive functioning, emotion regulation, and working memory, which is known to be compromised in schizophrenic patients36. The ventromedial caudate, on the other hand, receives inputs from the lateral orbitofrontal cortex, the temporal gyrus, and ACC, and projects to the lateromedial GPi and rostrolateral SNr before looping into the cortex via the medial VAmc and medial magnocellularis. This circuit is central to decision-making and goal-directed action36; the ACC in particular is involved to behavioral adjustment to meet changing environmental challenges37. Dysfunction of this pathway may relate to the symptoms of schizophrenia, similar to those that result from damage to the prefrontal cortex38.
The pars opercularis of the inferior frontal cortex (Brodmann area 44) is part of Broca’s area and is associated with language processing39. The inferior temporal cortex is involved in complex visual processing, face perception, recognition of numbers and objects, social cognition, and memory40. The precuneus is involved in mental imagery concerning the self41, episodic memory42, and visuospatial imagery43, as well as mental effort44,45 and social cognition46. Along with the precuneus, the PCC has been implicated in awareness, arousal47, and episodic memory48. The occipital (pericalcarine and fusiform) cortices are involved with visual and sensory information-processing49. The entorhinal cortex plays an important role in declarative (autobiographical/episodic/semantic) memory formation and consolidation, and memory optimization in sleep50. Alteration of these regions might be also related to the symptoms, including hallucination, delusion, cognitive including memory impairment, and motivational dysfunction, as commonly observed in schizophrenia patients.
We observed that schizophrenia-risk alleles belonging to class #3 were positively correlated with the SA of the lateral occipital cortex but negatively correlated with its cortical TH. Similarly, risk alleles of class #2 were positively correlated with the TH of the fusiform but negatively correlated with its SA. While these findings may seem contradictory, they could potentially be explained by the radial unit hypothesis51, which proposes that distinct developmental mechanisms promote the expansion of SA and increases in TH. Nevertheless, further research is necessary to fully understand these associations.
Finally, in the current study, the genetic regulatory effects of risk alleles were examined only in individuals of European ancestry, which is a limitation. These effects should be examined in other major populations, such as Africans, when available in the future.
Overall, we have identified a novel risk variant block at CACNA1C for schizophrenia. These variants are biologically functional and robustly associated with schizophrenia, suggesting that they may play significant roles in the pathogenesis of this disorder.
Supplementary Material
{kind=link}
Acknowledgments:
We thank NIH GWAS Data Repository, the Contributing Investigator(s) who contributed the phenotype and genotype data from his/her original study (e.g., Drs. Lencz, Darvasi, Gejman, Levinson, Goldstein, Stefansson, Collier, Sklar, Collins, Cloninger, Owen, O’Donovan, Purcell, Fromer, Kirov, etc.), and the primary funding organization that supported the contributing study. Funding and other supports for phenotype and genotype data were provided through the National Institutes of Health (NIH) (RC2 MH089964, R01 MH084098, R01 MH67257, R01 MH59588, R01 MH59571, R01 MH59565, R01 MH59587, R01 MH60870, R01 MH59566, R01 MH59586, R01 MH61675, R01 MH60879, R01 MH81800, U01 MH46276, U01 MH46289 U01 MH46318, U01 MH79469, U01 MH79470, R01 HG005827 and R01 MH071681), Medical Research Council (MRC) Centre (G0800509) and Program Grants (G0801418), the European Community’s Seventh Framework Programme (HEALTH-F2-2010-241909 (Project EU-GEI)), the North Shore - LIJ Health System Foundation, the Hebrew University Genetic Resource, the Broad Institute, Cardiff University, Icahn School of Medicine at Mount Sinai, the Wellcome Trust Sanger Institute (WT089062 and WT098051), the Fidelity Foundations, the Sylvan Herman Foundation and philanthropic gifts from Kent and Liz Dauten, Ted and Vada Stanley, the Friedman Brain Institute, the Institute for Genomics and Multiscale Biology, and the Janssen Research Foundation.
Funding:
This study was funded by the National Natural Science Foundation of China (81201057 and 81371491), Natural Science Foundation of Shanghai (20ZR1448400), Shanghai Municipal Health Bureau Project (20124109), Chinese Medical Association, Psychiatry – Servier Youth Research Fund, Shanghai Mental Health Center international cooperation project (2013-) and Shanghai Municipal Center for Mental Health Clinical Research Program. This study was funded by Shanghai Pudong Municipal Health Commission (No. PWRI2020-11).
Footnotes
Ethics approval and consent to participate: All subjects signed written informed consents before participating in the study. All methods were carried out in accordance with relevant guidelines and regulations. All study procedures were reviewed and approved by the Human Investigation Committee of Yale University.
Consent for publication: All authors approved for publication.
Competing interests: The authors declared no competing interests.
Availability of data and materials:
The datasets used for the analyses described in this manuscript were obtained from dbGaP at http://www.ncbi.nlm.nih.gov/sites/entrez?Db=gap. The dbGaP accession numbers include phs000021.v3.p2, phs000448.v1.p1, phs000167.v1.p1 and phs000687.v1.p1.
References
- 1.O’Donovan MC, Craddock N, Norton N, et al. Identification of loci associated with schizophrenia by genome-wide association and follow-up. Nature genetics. 2008;40(9):1053–1055. [DOI] [PubMed] [Google Scholar]
- 2.Stefansson H, Ophoff RA, Steinberg S, et al. Common variants conferring risk of schizophrenia. Nature. 2009;460(7256):744–747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ripke S, O’Dushlaine C, Chambert K, et al. Genome-wide association analysis identifies 13 new risk loci for schizophrenia. Nature genetics. 2013;45(10):1150–1159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Nyegaard M, Demontis D, Foldager L, et al. CACNA1C (rs1006737) is associated with schizophrenia. Mol Psychiatry. 2010;15(2):119–121. [DOI] [PubMed] [Google Scholar]
- 5.Ivorra JL, Rivero O, Costas J, et al. Replication of previous genome-wide association studies of psychiatric diseases in a large schizophrenia case-control sample from Spain. Schizophrenia research. 2014;159(1):107–113. [DOI] [PubMed] [Google Scholar]
- 6.Green EK, Grozeva D, Jones I, et al. The bipolar disorder risk allele at CACNA1C also confers risk of recurrent major depression and of schizophrenia. Mol Psychiatry. 2010;15(10):1016–1022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bigos KL, Mattay VS, Callicott JH, et al. Genetic variation in CACNA1C affects brain circuitries related to mental illness. Archives of general psychiatry. 2010;67(9):939–945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zheng F, Zhang Y, Xie W, et al. Further evidence for genetic association of CACNA1C and schizophrenia: new risk loci in a Han Chinese population and a meta-analysis. Schizophrenia research. 2014;152(1):105–110. [DOI] [PubMed] [Google Scholar]
- 9.Guan F, Zhang B, Yan T, et al. MIR137 gene and target gene CACNA1C of miR-137 contribute to schizophrenia susceptibility in Han Chinese. Schizophrenia research. 2014;152(1):97–104. [DOI] [PubMed] [Google Scholar]
- 10.He K, An Z, Wang Q, et al. CACNA1C, schizophrenia and major depressive disorder in the Han Chinese population. Br J Psychiatry. 2014;204(1):36–39. [DOI] [PubMed] [Google Scholar]
- 11.Schizophrenia Psychiatric Genome-Wide Association Study Consortium. Genome-wide association study identifies five new schizophrenia loci. Nature genetics. 2011;43(10):969–976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Schizophrenia Working Group of the Psychiatric Genomics Consortium. Biological insights from 108 schizophrenia-associated genetic loci. Nature. 2014;511(7510):421–427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Wang Z, Chen W, Cao Y, et al. An independent, replicable, functional and significant risk variant block at intron 3 of CACNA1C for schizophrenia. Aust N Z J Psychiatry. 2022;56(4):385–397. [DOI] [PubMed] [Google Scholar]
- 14.American Psychiatric Association. Diagnostic and statistical manual of mental disorders. fourth edition ed. Washington, DC: American Psychiatric Press; 1994. [Google Scholar]
- 15.Gain Collaborative Research Group, Manolio TA, Rodriguez LL, et al. New models of collaboration in genome-wide association studies: the Genetic Association Information Network. Nature genetics. 2007;39(9):1045–1051. [DOI] [PubMed] [Google Scholar]
- 16.Lencz T, Guha S, Liu C, et al. Genome-wide association study implicates NDST3 in schizophrenia and bipolar disorder. Nature communications. 2013;4:2739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Sanders AR, Duan J, Levinson DF, et al. No significant association of 14 candidate genes with schizophrenia in a large European ancestry sample: implications for psychiatric genetics. The American journal of psychiatry. 2008;165(4):497–506. [DOI] [PubMed] [Google Scholar]
- 18.Fromer M, Pocklington AJ, Kavanagh DH, et al. De novo mutations in schizophrenia implicate synaptic networks. Nature. 2014;506(7487):179–184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Howie BN, Donnelly P, Marchini J. A flexible and accurate genotype imputation method for the next generation of genome-wide association studies. PLoS Genet. 2009;5(6):e1000529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Purcell S, Neale B, Todd-Brown K, et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. American journal of human genetics. 2007;81(3):559–575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ramasamy A, Trabzuni D, Guelfi S, et al. Genetic variability in the regulation of gene expression in ten regions of the human brain. Nature neuroscience. 2014;17(10):1418–1428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.GTEx Consortium. The Genotype-Tissue Expression (GTEx) project. Nature genetics. 2013;45(6):580–585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Satizabal CL, Adams HHH, Hibar DP, et al. Genetic architecture of subcortical brain structures in 38,851 individuals. Nature genetics. 2019;51(11):1624–1636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Grasby KL, Jahanshad N, Painter JN, et al. The genetic architecture of the human cerebral cortex. Science. 2020;367(6484):eaay6690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Fischl B FreeSurfer. NeuroImage. 2012;62(2):774–781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lam M, Chen CY, Li Z, et al. Comparative genetic architectures of schizophrenia in East Asian and European populations. Nature genetics. 2019;51(12):1670–1678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Breier A, Buchanan RW, Elkashef A, Munson RC, Kirkpatrick B, Gellad F. Brain morphology and schizophrenia. A magnetic resonance imaging study of limbic, prefrontal cortex, and caudate structures. Archives of general psychiatry. 1992;49(12):921–926. [DOI] [PubMed] [Google Scholar]
- 28.Hof PR, Haroutunian V, Friedrich VL Jr., et al. Loss and altered spatial distribution of oligodendrocytes in the superior frontal gyrus in schizophrenia. Biological psychiatry. 2003;53(12):1075–1085. [DOI] [PubMed] [Google Scholar]
- 29.Bonilha L, Molnar C, Horner MD, et al. Neurocognitive deficits and prefrontal cortical atrophy in patients with schizophrenia. Schizophrenia research. 2008;101(1–3):142–151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Tully LM, Lincoln SH, Liyanage-Don N, Hooker CI. Impaired cognitive control mediates the relationship between cortical thickness of the superior frontal gyrus and role functioning in schizophrenia. Schizophrenia research. 2014;152(2–3):358–364. [DOI] [PubMed] [Google Scholar]
- 31.Hokama H, Shenton ME, Nestor PG, et al. Caudate, putamen, and globus pallidus volume in schizophrenia: a quantitative MRI study. Psychiatry research. 1995;61(4):209–229. [DOI] [PubMed] [Google Scholar]
- 32.Bridle N, Pantelis C, Wood SJ, et al. Thalamic and caudate volumes in monozygotic twins discordantfor schizophrenia. Aust N Z J Psychiatry. 2002;36(3):347–354. [DOI] [PubMed] [Google Scholar]
- 33.Juuhl-Langseth M, Rimol LM, Rasmussen IA Jr., et al. Comprehensive segmentation of subcortical brain volumes in early onset schizophrenia reveals limited structural abnormalities. Psychiatry research. 2012;203(1):14–23. [DOI] [PubMed] [Google Scholar]
- 34.Juuhl-Langseth M, Hartberg CB, Holmen A, et al. Impaired Verbal Learning Is Associated with Larger Caudate Volumes in Early Onset Schizophrenia Spectrum Disorders. PLoS One. 2015;10(7):e0130435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Zhang S, Ide JS, Li CS. Resting-state functional connectivity of the medial superior frontal cortex. Cerebral cortex. 2012;22(1):99–111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Grahn JA, Parkinson JA, Owen AM. The role of the basal ganglia in learning and memory: neuropsychological studies. Behavioural brain research. 2009;199(1):53–60. [DOI] [PubMed] [Google Scholar]
- 37.Manza P, Hu S, Chao HH, Zhang S, Leung HC, Li CR. A dual but asymmetric role of the dorsal anterior cingulate cortex in response inhibition and switching from a non-salient to salient action. NeuroImage. 2016;134:466–474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Frank MJ, O’Reilly RC. A mechanistic account of striatal dopamine function in human cognition: psychopharmacological studies with cabergoline and haloperidol. Behav Neurosci. 2006;120(3):497–517. [DOI] [PubMed] [Google Scholar]
- 39.Schremm A, Noven M, Horne M, Soderstrom P, van Westen D, Roll M. Cortical thickness of planum temporale and pars opercularis in native language tone processing. Brain Lang. 2018;176:42–47. [DOI] [PubMed] [Google Scholar]
- 40.Conway BR. The Organization and Operation of Inferior Temporal Cortex. Annu Rev Vis Sci. 2018;4:381–402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lou HC, Luber B, Crupain M, et al. Parietal cortex and representation of the mental Self. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(17):6827–6832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Fletcher PC, Frith CD, Baker SC, Shallice T, Frackowiak RS, Dolan RJ. The mind’s eye--precuneus activation in memory-related imagery. NeuroImage. 1995;2(3):195–200. [DOI] [PubMed] [Google Scholar]
- 43.Oshio R, Tanaka S, Sadato N, Sokabe M, Hanakawa T, Honda M. Differential effect of double-pulse TMS applied to dorsal premotor cortex and precuneus during internal operation of visuospatial information. NeuroImage. 2010;49(1):1108–1115. [DOI] [PubMed] [Google Scholar]
- 44.Zhang S, Li CS. Task-related, low-frequency task-residual, and resting state activity in the default mode network brain regions. Front Psychol. 2012;3:172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Zhang S, Li CS. A neural measure of behavioral engagement: task-residual low-frequency blood oxygenation level-dependent activity in the precuneus. NeuroImage. 2010;49(2):1911–1918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Wang W, Zhornitsky S, Li CS, et al. Cerebral responses to self-initiated action during social interactions. Cogn Affect Behav Neurosci. 2019;19(6):1521–1535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Le TM, Zhornitsky S, Wang W, Ide J, Zhang S, Li CR. Posterior Cingulate Cortical Response to Active Avoidance Mediates the Relationship between Punishment Sensitivity and Problem Drinking. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2019;39(32):6354–6364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Nielsen FA, Balslev D, Hansen LK. Mining the posterior cingulate: segregation between memory and pain components. NeuroImage. 2005;27(3):520–532. [DOI] [PubMed] [Google Scholar]
- 49.Jalbrzikowski M, Jonas R, Senturk D, et al. Structural abnormalities in cortical volume, thickness, and surface area in 22q11.2 microdeletion syndrome: Relationship with psychotic symptoms. NeuroImage. Clinical 2013;3:405–415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Suthana N, Haneef Z, Stern J, et al. Memory enhancement and deep-brain stimulation of the entorhinal area. N Engl J Med. 2012;366(6):502–510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Rakic P Specification of cerebral cortical areas. Science. 1988;241(4862):170–176. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The datasets used for the analyses described in this manuscript were obtained from dbGaP at http://www.ncbi.nlm.nih.gov/sites/entrez?Db=gap. The dbGaP accession numbers include phs000021.v3.p2, phs000448.v1.p1, phs000167.v1.p1 and phs000687.v1.p1.