

Abstract
The halopyrimidine 5-bromo-2′-deoxyuridine (BrdU) is an exogenous marker of DNA synthesis. Since the introduction of monoclonal antibodies against BrdU, an increasing number of methodologies have been used for the immunodetection of this synthesized bromine-tagged base analogue into replicating DNA. BrdU labeling is widely used for identifying neuron precursors and following their fate during the embryonic, perinatal, and adult neurogenesis in a variety of vertebrate species including birds, reptiles, and mammals. Due to BrdU toxicity, its incorporation into replicating DNA presents adverse consequences on the generation, survival, and settled patterns of cells. This may lead to false results and misinterpretation in the identification of proliferative neuroblasts. In this review, I will indicate the detrimental effects of this nucleoside during the development of the central nervous system, as well as the reliability of BrdU labeling to detect proliferating neuroblasts. Moreover, it will show factors influencing BrdU immunodetection and the contribution of this nucleoside to the study of prenatal, perinatal, and adult neurogenesis. Human adult neurogenesis will also be discussed. It is my hope that this review serves as a reference for those researchers who focused on detecting cells that are in the synthetic phase of the cell cycle.
Keywords: 5-bromo-2′-deoxyuridine, adult neurogenesis, human adult neurogenesis, labeling, pitfalls, prenatal neurogenesis, proliferation, S-phase, suturing S-phase, toxicity
Introduction
Neuroblasts are cells able to generate progeny that undergo differentiation into functional neurons. The temporal sequence of neuron production throughout the nervous system is a precisely regulated event that can be linked to the patterns of anatomical connections in adulthood. During the embryonic and postnatal life, alterations in the proliferation of neuron precursors, genesis of neurons from their neuroblasts, migration to their final locations in the brain, and differentiation have important consequences on the normal development and cytoarchitecture of the central nervous system, which have been related to several pathological conditions such as schizophrenia, autism spectrum disorder, and deficit-hyperactivity disorder (Allin, 2016), Alzheimer’s disease (Kim et al., 2022), amyotrophic lateral sclerosis, Huntington’s disease, Parkinson’s disease, dementia with Lewy bodies, and frontotemporal dementia (Terreros-Roncal et al., 2021).
Administration and subsequent detection of 5-bromo-2′-deoxyuridine (BrdU) which incorporates into replicating DNA during the synthetic phase of the cell cycle are important approaches for inferring neuron generation and tracing the fate of proliferating neuroblasts and their progeny during the prenatal and postnatal development of the central nervous system. Despite this, BrdU labeling has important limitations, which may limit its usefulness in several studies. In this review, I will focus on (i) a short history of the BrdU and BrdU immunodetection, (ii) the reliability of BrdU labeling as a tool to detect neuroblasts undergoing S-phase, (iii) delivery and factors influencing the detection of BrdU into DNA, (iv) BrdU dosage and saturation of the S-phase, (v) BrdU toxicity, (vi) BrdU labeling and prenatal and perinatal neurogenesis, and (vii) BrdU labeling and adult neurogenesis. The effects of BrdU incorporation into DNA on human adult neurogenesis will be also discussed.
Search Strategy
All studies cited in this narrative review have been performed until March 2023. The manuscripts were searched electronically using the PubMed and Google Scholar databases. They represent the most relevant articles in the field. Only papers published in English were considered. A combination of the following keywords/terms were used: BrdU, BrdU labeling, S-phase, prenatal neurogenesis, adult neurogenesis, human adult neurogenesis.
5-Bromo-2’-Deoxyuridine Immunodetection
The original techniques utilized for the selective labeling of proliferating cells include the use of tritiated thymidine ([3H]TdR) and a detection step using either autoradiography or scintillation techniques (Leif et al., 2004; Cavanagh et al., 2011). [3H]TdR autoradiography has provided a way to measure the labeling index (fraction of S-phase cells), the percentage of mitotic figures labeled (Leif et al., 2004), as well as accurate information about the replication and repair of the DNA (Pederson, 2014). This methodology has also allowed the characterization of cell precursors under different experimental contexts, including cancer research (Clarkson et al., 1967), autoimmune diseases (Lawless et al., 2018), and genesis of neurons (Martí-Clúa, 2021a, 2022).
In the field of neurogenesis, an important goal in prenatal and adult neurogenesis research is to obtain, in tissue sections, a confident identification of those neuron precursors engaged in the DNA synthesis (Altman and Bayer, 1997; Duque et al., 2016). In this line, the possibility of experimentally labeling proliferating neuroblasts and dating neuronal birth started with the application of [3H]TdR, an exogenous marker of the DNA synthesis, and subsequent autoradiography in fixed tissue sections (Miale and Sidman, 1961). Despite this, [3H]TdR autoradiography is not cheap as it requires technical expertise in handling radiolabeled products, and the procedure takes several weeks in developing autoradiographs (Martí et al., 2002; Duque and Rakic, 2011). For decades, [3H]TdR and autoradiography have produced critical insights into the cellular mechanisms of central nervous system development, including developmental timetables of neurons, cell lineage, and also follow the migratory route of neurons from their neuroepithelial origin to their final locations (Nowakowski and Rakic, 1989; Altman and Bayer, 1997). Interestingly, [3H]TdR is not implicated in RNA synthesis.
The original process for marking the synthetic phase of the cell cycle with the use of [3H]TdR and high-resolution autoradiography was followed by the detection of BrdU incorporated into nascent DNA. This pyrimidine analogue is a chemically synthesized bromine-tagged base analogue that, during the S-phase of the cell cycle, is permanently incorporated in place of endogenous thymidine into newly synthesized DNA (Graztner, 1982; Nowakowski et al., 1989; Martí and Rodríguez-Vázquez, 2020; Table 1). BrdU resembles endogenous thymidine; therefore, BrdU is readily absorbed into the blood stream after being administered, spreading broadly in the body through the blood circulation system, and penetrating most organs, including the central nervous system (Solius et al., 2021). Once integrated into DNA, BrdU becomes stable, and it will remain in place and be passed down to daughter cells following mitosis (Lehner et al., 2011).
Table 1.
Examples of thymidine analogues used to detect de novo DNA synthesis and cell proliferation
| Agent | Antigen retrieval | Molar mass (g/mol) | Molecular formula | Antibody detection |
|---|---|---|---|---|
| BrdU | Yes | 307.10 | C9H11BrN2O5 | Yes |
| CldU | Yes | 262.65 | C9H11ClN2O5 | Yes(1) |
| IdU | Yes | 354.10 | C9H11IN2O5 | Yes(2) |
| EdU | No | 252.22 | C11H12N2O5 | Yes(3) |
| F-ara-EdU | No | 270.22 | C11H11FN2O5 | No(4) |
| AmdU | No | 283.24 | C10H13N5O5 | No(5) |
| VdU | No | 254.24 | C11H14N2O5 | No(6) |
AmdU: 5-Azidomethyl-2’-deoxyuridine; BrdU: 5-bromo-2’-deoxyuridine; CldU: 5-chloro-2’-deoxyuridine; EdU: 5-ethynyl-2’-deoxyuridine; F-ara-Edu: 2’-deoxy-2’-fluoro-5-ethynyluridine; IdU: 5-iodo-2’-deoxyuridine; VdU: 5-Vinyl-2’-deoxyuridine. (1): Rat monoclonal anti-BrdU (clone BU1/75) recognizes CldU; (2): rat monoclonal anti-BrdU (clone B/44) recognizes IdU; (3): most commercial anti-BrdU antibodies recognize EdU after single-stranded DNA production; (4–6): commercial antibodies are not available. A bioorthogonal chemical reaction was used for their detection.
It is important to indicate that, this chemically synthesized bromine-labeled base analogue, was introduced in the 1950s as a tool to detect and quantify DNA synthesis from proliferating cancer cells (Levkoff et al., 2008). Moreover, the application of antibodies against BrdU was the result of the necessity of overpassing the centrifugal cytology, a method that centrifuges cells onto a slice and then fixes the wet cells under the action of centrifugal force (Leif et al., 2004). In 1964, Erlanger and Beiser developed a polyclonal antibody to BrdU. Later, Gratzner et al. (1975) produced and purified a polyclonal antibody, which could be used for immunofluorescence analysis of DNA replication in single cells or to detect sister chromatid exchanges.
Since the introduction of the first monoclonal antibody against BrdU (Graztner, 1982), many antibodies for this analogue are commercially available that have been raised in a variety of host species such as mice, rats, rabbits, and sheep. It has facilitated cell cycle studies and provided an increasing number of immunocytochemical procedures for the detection of the BrdU that has been incorporated into replicating DNA. BrdU immunohistochemistry has provided new advances in the identification and characterization of neuroblasts during embryonic and adult neurogenesis in a variety of vertebrate species, including birds (Larson et al., 2019) and mammals (Miller and Nowakowski, 1988; Taupin, 2007).
Is 5-Bromo-2’-Deoxyuridine Labeling a Reliable Tool to Detect Neuroblasts in the S-phase of the Cell Cycle?
An important issue in the context of BrdU labeling is to ask the following question: is a BrdU-reactive nucleus a definitive sign of cell proliferation? The answer to this question is not simple. This is because the administration of this marker tags to a cohort of asynchronous cycling cells in the S-phase of the cell cycle and, therefore, the incorporation of BrdU into the nucleus should supply a realistic picture of the cell population fraction engaged in DNA synthesis during BrdU exposure. However, it is essential to accept that the detection of a BrdU-positive cell in a tissue section should not be a sufficient criterion to affirm that this cell was in the DNA synthetic phase of the cell cycle during marker administration. In this context, it has been demonstrated that BrdU can be incorporated into DNA without cell proliferation. This is because many other conditions, such as cell differentiation, abortive cell cycle, gene duplication, grafted cells, and DNA repair also promote BrdU labeling (Breunig et al. 2007; Taupin, 2007; Duque and Rakic, 2011; Zheng et al., 2011; Duque and Spector, 2019). In the context of the prenatal, perinatal, and adult neurogenesis, these findings have important implications for the interpretation of the results obtained from BrdU labeling. For example, a BrdU-stained nuclei may indicate either a cell is undergoing the S-phase of the cell cycle, or it is attempting to repair its DNA. Taken together, these studies indicate that in the absence of suitable controls, the possibility of false BrdU labeling or the erroneous interpretation of such labeling exists.
At the beginning of the central nervous system development, excessive numbers of neurons are produced, including many incorrect connections to their targets. Later, some of these die via apoptotic events (Svandova et al., 2023). On the other hand, the presence of neural stem cells and the generation of neurons in the adult brain are controlled at multiple levels, including programmed cell death (Ryu et al., 2016). Neurons are usually considered postmitotic cells (G0 phase of the cell cycle), being found in a quiescent state in the adult nervous system. Evidence has revealed that neurons can try to reactivate the cell cycle when they are exposed to insults (Frade and Ovejero-Benito, 2015). Under these circumstances, neurons can begin abortive DNA synthesis without cell division, and they die at the G1/S checkpoint (Pavulraj et al., 2023). In this context, it has been shown that apoptotic cornus ammonis 1 neurons can reenter the cell cycle, pass the G1/S phase checkpoint, resume DNA synthesis, and incorporate BrdU in their nuclei after cerebral hypoxia/ischemia (Kuan et al., 2004). Similar data have been observed when apoptotic events were induced in primary neuronal cultures of the prenatal rat cerebral cortex after treatment with homocysteine (Ye and Blain, 2010). Taken together, these results have indicated that, after an injury, the incorporation of BrdU alone cannot be considered as sufficient evidence of neurogenesis because it could lead to the misinterpretation of the results. The criteria for neuron production should include, in the same tissue section, at least three criteria: (i) proofs of DNA synthesis, (ii) identification of neuronal markers, and (iii) evidence showing the absence of apoptotic markers.
Neurons are highly differentiated cells and are usually considered to be in G0. Despite this, there are examples of neurons that, under normal physiological conditions, remain alive with 4C DNA content (Svandova et al., 2023). Retinal ganglion cells, and pyramidal neurons of the cerebrum and the neocortex are examples of tetraploid neurons (Nandakumar et al., 2021). On the other hand, it has also been reported that several aneuploid neurons have been found in the hippocampus of patients who have Alzheimer’s disease (Vincent et al., 1996) as well as in other pathological conditions including stroke, amyotrophic lateral sclerosis, encephalitis, and cognitive impairment (Herrup et al., 2004). The expression of proteins of the cell cycle in these regions of neurodegeneration suggests that these neurons have completed G1, S, and G2 but not the mitotic phase (Shepherd et al., 2018). This cellular event has important implications when studying neurogenesis. This is because BrdU is incorporated into the newly synthesized DNA of proliferating cells during the synthetic phase of the cell cycle. Therefore, BrdU labeling will not allow us to distinguish between the proliferation of neuroblasts and mature neurons duplicating their DNA without mitosis (Taupin, 2007; Lehner et al., 2011). Again, these results support the evidence that BrdU may not be a suitable tool to detect cell division.
BrdU has been used to discover the fate of transplanted cells in the central nervous system. In this scenario, cells are tagged in culture with this marker prior to grafting. Later, grafted cells are identified into the host by immunohistochemistry. Pitfalls and false positives have been reported because previous research has demonstrated that BrdU can be transferred from the graft to the host neuroblasts and glial cells (Burns et al., 2006), which may lead to a scenario where the integration and phenotype of the grafted cells into the host are incorrectly interpreted (Taupin, 2007; Lehner et al., 2011). The data from these investigations have supplied new evidence about the suitability of BrdU to study cell proliferation.
These issues have pivotal importance in the context of BrdU labeling, and therefore this artificial nucleoside should be considered as an indicator of DNA synthesis and not of cell division, as several times assumed (Breunig et al., 2007; Duque and Rakic, 2011).
Delivery and Factors Influencing the Detection of 5-Bromo-2’-Deoxyuridine into DNA
As compared with [3H]TdR autoradiography, BrdU labeling of neuroblasts is cheap, fast, easy to implement, and it requires neither the use of radiolabeled substances nor appropriate laboratories. Moreover, BrdU allows the detection of labeled cells throughout the relatively thick tissue sections required for stereological analyses of the brain (Taupin, 2007). Despite this, BrdU presents disadvantages. The first of these is that when detected by immunohistochemistry, BrdU incorporation into replicating DNA is not stoichiometric (Nowakowski and Hayes, 2000). Another source of potential technical problems is the way in which BrdU is delivered to the animals. BrdU can be supplied through intraperitoneal, intravenous, or intramuscular injections, implantation of osmotic pumps, and drinking water (Taupin, 2007; Solius et al., 2021).
These routes of administration are linked to alterations in BrdU uptake. Two examples have sustained evidence that the route of BrdU administration is related to variations in the number of BrdU-positive cells. In the first of them, BrdU is dissolved in drinking water and supplied to rats or mice. They are nocturnal animals, which means they are most active at night and during dawn and dusk. Due to their circadian dependence on water intake, treatment with this thymidine analogue dissolved in drinking water tags different numbers of proliferating cells during the light and dark phases of the day (Solius et al., 2021). Another interesting example, reporting differences in the number of BrdU-stained cells is that supplied by Zhao et al. (2003). In those experiments, BrdU was delivered by intraperitoneal injections or intracerebroventricular infusion into the lateral ventricle of adult mice. The results provide strong evidence that the intracerebroventricular administration of BrdU resulted in the labeling of a 5-fold higher number of BrdU-stained cells in the dentate gyrus and substantia nigra pars compacta compared with the intraperitoneal injections of this marker. These results have provided evidence that the route of BrdU administration is related to variations in the number of BrdU-positive cells.
When BrdU is administered, it is important to indicate that this nucleoside is metabolized through dehalogenation when integrated into the DNA. Once dehalogenated, the uracil residue would be excised from the DNA by the uracil glycosylase repair system. Several reports have indicated that BrdU is also metabolized through dehalogenation in the plasma. In humans, the half-life of BrdU in plasma is about 8–11 minutes (Hume and Saffhill, 1986; Taupin, 2007). These results have shown that the concentration of BrdU reaching the brain is only a fraction of the administered dose (Taupin, 2007), which may alter BrdU uptake by the neuroblasts. Despite this, BrdU is spread widely around the body through the blood circulation system, and crosses the brain blood barrier, and reaches germinal matrices or neurogenetic regions to label proliferating cells. Other tissues separated by barriers, including testis and placenta are also penetrated by the BrdU (Solius et al., 2021). An interesting approach to increase the concentration of BrdU in the nervous system is to supply this agent via intracerebroventricular injections (Zhao et al., 2003).
The specificity of BrdU immunodetection depends on several variables. This is because many factors have been described which influence the immunoreactivity of the BrdU epitope in paraffin-embedded tissue sections. One of these is the method of tissue fixation. Most procedures for the immunodetection of incorporated BrdU require tissue to be frozen or fixed in chemical agents, including methanol/acetic acid (Hervás et al., 2002), ethanol (Vanderlaan and Thomas, 1985), formalin (Rodríguez-Vázquez and Martí, 2021), or paraformaldehyde (Martí and Rodríguez-Vázquez, 2020). Results from my laboratory have shown no specific differences among the three fixative procedures (methanol/acetic acid, formalin, and paraformaldehyde), neither in BrdU-signal nor in the estimation of various cell cycle parameters (Hervás et al., 2002; Martí et al., 2016; Molina et al., 2017). However, it has been revealed that fixation with a methanol-Carnoy mixture, referred to as methacarn solution (60% methanol v/v, 30% chloroform v/v, and 10% glacial acetic acid v/v), provides better results than formalin-fixed tissue in the assessment of cell proliferation in rat mammary carcinomas (McGinley et al., 2000).
Besides the step of tissue fixation, attention should also be paid to the hydrolysis conditions leading to single-stranded DNA, the only form recognized by the anti-BrdU monoclonal antibodies (Gratzner, 1982). Usually, this methodological condition is obtained by incubating the tissue sections in hydrochloric acid (Dolbeare, 1995). In the same way, 5-chloro-2′-deoxyuridine (CldU) and 5-iodo-2′-deoxyuridine deoxyuridine (IdD) also need the production of single-stranded DNA because antibodies cannot gain access to native DNA (Solius et al., 2021; Table 1). The low pH of hydrochloric acid is considered to extract some histones from DNA and to partially cut the hydrogen bonds between DNA strands, thereby allowing the access of the anti-BrdU antibody (Wakayama et al., 2015). Pretreatment with hydrochloric acid prevents labeling of nuclei with propidium iodide and 4′6-diamidine-2′-phenylindole (Taupin, 2007), erodes cell and tissue components (Ffrench et al., 1994; Solius et al., 2021), and affects the immunoreactivity of a wide variety of proteins (Dinjens et al., 1992). To avoid these negative effects, several antigen retrieval techniques have been used, such as DNase I (Sekerkova et al., 2004a, b), heating in a sodium citrate buffer (Tang et al., 2007; Shimada et al., 2008) or the use of endonuclease Eco RI followed by exonuclease III (Dolbeare and Gray, 1988).
When these antigen retrieval procedures were systematically compared, it was found that the specificity of the staining pattern was affected by the antigen retrieval used (Dolbeare, 1995; Molina et al., 2017). In other words, these procedures give rise to considerable variability in the proportion of BrdU-stained cells. For example, protocols using 75 and 100 U/mL of DNase I, and 20 mM of heated sodium citrate buffer yielded similar numbers of BrdU-positive cells, although the values were smaller than those obtained with 3 N hydrochloric acid at 40°C during 15 minutes. 600 U/mL of endonuclease EcoRI combined with 600 U/mL of exonuclease III provided the smallest values. The reasons for these discrepancies are unknown, but they may derive from differences in the creation of single-stranded DNA. In this context, the disruption of the double-stranded DNA may be less accurate in those procedures using DNase I, heated citrate buffer or 600 U/mL of endonuclease EcoRI combined with 600 U/mL of exonuclease III, so that fewer antigenic sites can react with primary anti-BrdU antibody. Figure 1 depicts the variation in the number of labeled granule cell precursors in the cerebellar external granular layer among sections treated with several antigen retrieval solutions, such as hydrochloric acid (3 N at 40°C for 15 minutes) (Figure 1A), DNase I at 75 U/mL (Figure 1B), heated sodium citrate buffer at 20 mM (Figure 1C), and EcoRI (600 U/mL) combined with exonuclease III (600 U/mL) (Figure 1D). Taken together, these results have suggested that, depending on the antigen retrieval solution, the detection threshold for the BrdU incorporated into replicating DNA may be different.
Figure 1.
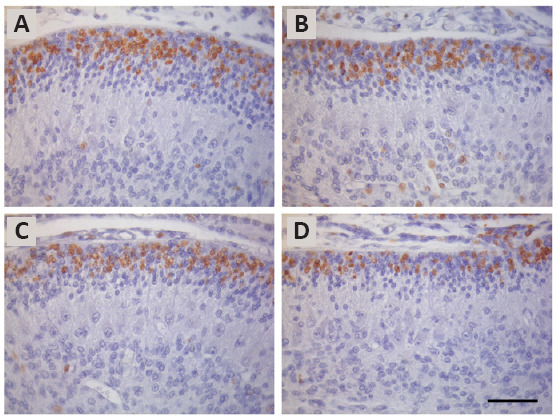
BrdU-immunolabeling in the cerebellar external granular of 10-day-old rats allowed to survive for 6 hours after a single injection of BrdU.
Sections representative of the vermis were treated with hydrochloric acid (3 N at 40°C for 15 minutes) (A), DNase I at 75 U/mL (B), heated sodium citrate buffer at 20 mM (C) or EcoRI (600 U/mL) combined with exonuclease III (600 U/mL) (D). BrdU-positive cells are those are those presenting a brown reaction product confined to nuclear material. Note that sections exposed to 3 N hydrochloric acid at 40°C for 15 minutes presents higher proportions of BrdU-reactive cells than those sections treated with DNase I at 75 U/mL or heated sodium citrate buffer at 20 mM. Sections incubated with EcoRI (600 U/mL) combined with exonuclease III (600 U/mL) showed the lowest proportion of labeled nuclei. Scale bar: 50 μm. Unpublished data.
Similar results have been observed when hydrochloric acid, sodium citrate buffer, and hydrochloric acid + formamide (Leuner et al., 2009) or several proteases (pepsin, pronase E, protease type XXIV, and trypsin type III) (Bak and Panos, 1997) were compared. These results have revealed that the interpretation of BrdU immunolabeling results should be carefully considered.
BrdU, CldU and IdD are analogues modified in the 5 position of the thymidine ring by halogen atoms (bromide, chloride, and iodine). Of all of them, BrdU is the most frequently used to infer the fraction of cells undergoing the S-phase of the cell cycle (Martí-Clúa, 2021b). CldU and IdD, on the other hand, allow us to perform studies involving double labeling for revealing the progression of cells through the cell cycle and the double and triple S-phase labeling schemes for distinguishing between proliferating and quiescent or senescent cells (Solius et al., 2021). In this context, the synthetic analogue, 5-ethynyl-2′-deoxyiridine (EdU) for thymidine substitution has been used for studying the times of neuron production (Fang et al., 2021; Table 1). Detection of this nucleoside does not need DNA denaturation for facilitating sterical access of antibodies. 5-Ethynyl-2′-deoxyiridine detection is based on the covalent coupling of a fluorescent azide to a terminal alkyne group of this agent through a Cu(I)-catalyzed cycloaddition reaction, which is named “click” (Solius et al., 2021). At the present, other nucleosides have emerged to detect de novo DNA synthesis, including (2′S)-2′-deoxy-2′-fluoro-5-ethynyluridine, 5-(azidomethyl)-2′-deoxyuridine, and 5-vinyl-2′-deoxyuridine (Solius et al., 2021; Table 1).
The use of BrdU, CldU and EdU has advantages and disadvantages. Advantages are that they incorporate into the nuclear DNA during the S-phase of the cell cycle, and their presence can rapidly be revealed by immunohistochemical procedures (Yamada et al., 2005; Sagga et al., 2018; Fang et al., 2021). In contrast, the disadvantages are that CldU and IdD present toxicity, but their noxious effects have been studied as an anticancer therapy and have not been yet analyzed in the context of labeling proliferating cells. When EdU is considered, there is evidence indicating that this marker has more severe toxic effects than an equimolar dose of BrdU (Solius et al., 2021).
At present, several pharmaceutical companies produce BrdU antibodies in various hosts (rat, mouse, rabbit, and sheep). These manufacturers indicate specifications (species reactivity, clone, clonality, and conjugation), scientific applications (ELISA, Western Blot, flow cytometry, and immunohistochemistry), protocols, as well as an important body of literature indicating references in which their primary BrdU antibody has been used. Despite this, neither the sensitivity nor specificity of these primary antibodies has been compared. Antibody selection is a key issue for detecting BrdU incorporation into replicating DNA. This is because it has been demonstrated that the number and the spatial distribution of BrdU-reactive cells in the dentate gyrus of adult rats (Leuner et al., 2009) and in their testes (Bak and Panos, 1997) are strongly dependent on the antibody used. Even more variability in BrdU labeling was observed after applying different primary anti-BrdU antibodies to HeLa cells incubated with BrdU (Liboska et al., 2012). These reports have revealed that primary BrdU antibodies originating from distinct commercial suppliers are not equally sensitive.
I suggest that before performing a study involving BrdU immunodetection, the specificity and sensitivity of some anti-BrdU antibodies should be carefully compared. In my opinion, this is an important issue to avoid inaccurate and conflicting results. In this context, using anti-BrdU antibodies from different companies and under similar laboratory conditions (histological procedures, strain and age of the animals, organ, dosage, and regimen of BrdU administration and DNA denaturation method), the number of BrdU-positive cells should be counted to determine if those BrdU antibodies label an equivalent number of cells.
5-Bromo-2’-Deoxyuridine Dosage and Saturation of the S-phase
Independently of the route of BrdU administration and variables affecting its immunodetection, fixation methods, antigen retrieval, and sensitivity of antibodies, a point deserving attention is the suturing dose of BrdU necessary to tag most of the neuroblasts undergoing the S-phase of the cell cycle. In other words, what dosage of BrdU is the most appropriate for labeling all S-phase cells? When using BrdU labeling, it is necessary to have a reliable picture of the neuroblast population engaged in DNA synthesis during the exposure to the marker. The available time of BrdU for dividing neuroblasts is one factor that should be interpreted with caution because, when studied on its own, minimal labeling time can be a misleading indicator of the BrdU acquisition rate. Identification of the S-phase saturation dose of BrdU is an important step in the evaluation of the mechanisms underlying regulation of neuroblasts division and will foster comparison of data among laboratories (Mandyam et al., 2007). This aspect should be addressed with caution since the saturating dose depends on the species and tissues (Solius et al., 2021), but the age of the animal also seems to be involved. For example, a single dose of 50 mg/kg BrdU is sufficient to label most of S-phase cells in the neuroepithelium of the embryonic cerebral wall of mice (Takahashi et al., 1995a), rat cerebellar neuroepithelium (Martí and Rodríguez-Vázquez, 2020) and rat cerebellar external granular layer (Hervás et al., 2002). On the other hand, when adult animals are considered, the bulk of the incorporation of 3H-BrdU tracer doses into DNA of mitotically active tissues of adult mice was completed within one hour of administration (Böswald et al., 1990). Moreover, the suturing dose of BrdU inferred by quantification of labeled neuroblast in the hippocampal dentate gyrus after a single delivery is 150 mg/kg in mice (Mandyam et al., 2007), and 200 mg/kg (Eadie et al., 2005) or 300 mg/kg in rats (Cameron and Mckay, 2001) but, in the ventricular zone of the adult songbirds, the saturation was achieved with a dose of 50 mg/kg of BrdU (Kubikova et al., 2020).
5-Bromo-2’-Deoxyuridine Toxicity
Before reviewing BrdU toxicity, it is necessary to provide a definition about what toxicity means. From a booklet developed in 1986 by the Hazard Evaluation and Information Service (HESIS) and the Labor Occupational Health Program (LOHP) at the University of California, toxicity can be defined as the ability of a substance to produce harmful health effects. These effects can strike on a cell type, a population of cells, a tissue, an organ system, or the entire body.
BrdU is usually considered a harmless substitute for the endogenous thymidine. Despite this, several studies have supplied evidence that the incorporation of this nucleoside into newly synthesized DNA may not be as innocuous as it is thought. The toxic effects of BrdU have been known since 1959, when Hakala (1959) reported that this agent affects the growth of mammalian cancer cells. Since then, several reports have been revealed that BrdU administration in the prenatal, perinatal, and postnatal life can be the source of unforeseen problems (Taupin, 2007; Duque and Rakic, 2011; Lehner et al., 2011; Martí-Clúa, 2021b).
In this context, it has been indicated that BrdU is phosphorylated and inserted into either newly replicating DNA or newly repaired DNA in place of the phosphorylated thymine (Morris et al., 1992). BrdU has a different chemical structure in comparison to endogenous thymidine, and this synthetic halogenated pyrimidine integrated as a totally foreign atom (Br) into replicating DNA when bromouracil is introduced instead of thymidine (Duque and Rakic, 2011). Moreover, the addition of exogenous BrdU modifies cellular nucleotide pool ratios. When the exogenous BrdU concentration is excessive or the ratio of deoxycytidine triphosphate to BrdU triphosphate decreases, the transformation of nucleotide triphosphates to deoxynucleotide triphosphates through the ribonucleotide reductases is inhibited. Under these conditions of pool imbalances, BrdU can be inserted into the genome opposite the nucleobase guanine (Morris et al., 1992). From these available data, it is plausible to assume that the genes that use bromosubstituted DNA are unlikely to transcribe appropriately into RNA and eventually the proper protein (Duque and Rakic, 2011).
The incorporation of BrdU into DNA produces important changes in the double helical structure of this nucleic acid, which are related to the toxicity of BrdU both in vitro and in vivo. Moreover, upon incorporation into DNA, BrdU produces destabilized nucleosome positioning which leads to alterations in heterochromatin organization and gene expression (En et al., 2023). In line with this, it has been indicated that, in combination with several stressors, including ionizing radiation, BrdU has adverse consequences for cancer cells (Levkoff et al., 2008). Moreover, in the absence of stressors, culture of same cancer cell lines, including human teratocarcinoma (Morris et al., 1992), human cutaneous T-cell lymphoma, human osteosarcoma, human thyroid tumor, and rat glioma (Levkoff et al., 2008), BrdU is an anticancer agent because a single exposure to this thymidine analogue produces a progressive and important impairment of cell cycle (accumulation in the G1 phase) as well as a lengthening of the time that the cells remain in S-phase (Morris et al., 1992; Ross et al., 2008). Incorporation of BrdU was also found to induce senescence-like processes on neurosphere cultures derived from neonatal mice (Ross et al., 2008) and adult rat brain (Leuner et al., 2009) as well as in tumoral (Mischishita et al., 1999; Levkoff et al., 2008) and non-tumoral cell lines (Ross et al., 2008). These results have suggested that BrdU activates several senescence pathways, including senescence-associated mRNAs and proteins present in both mortal and immortal mammalian cells (Mischishita et al., 1999; Suzuki et al., 2001). A recent study has revealed that a basic domain in the histone H2B N-terminal may be involved in the senescence produced by the BrdU (En et al., 2023). Moreover, it has been denoted that this halopyrimidine induces the differentiation of promyelocytic cell line (HL-60) to granulocytes (Keoffler et al., 1983) and promotes the differentiation of adult bone marrow-derived stem cells into neural and retinal cells (Qu et al., 2004). Further studies have shown that BrdU promotes the reprogramming of somatic cells into pluripotent stem cells (Long et al., 2015). The occurrence of sister-chromatid exchanges and double-strand breaks has also been observed in human teratocarcinoma cells cultured with BrdU (Morris et al., 1992).
From these previous studies, a question emerges. Why do BrdU-labeled cells stop proliferating? A possible link is the DNA methylation loss upon BrdU incorporation into the DNA. The mechanism by which this thymidine analogue affects DNA methylation is unclear, but these authors have proposed that in regions of DNA where a cytosine is followed by a guanine, the CpG sites, BrdU can be incorporated in place of cytidine, thus leading to the loss of CpG sites methylation. Interestingly, both CldU and IdD present comparable effects (Schneider and d’Adda di Fagagna, 2012). These results have supplied additional evidence indicating that the results obtained with BrdU labeling should be carefully interpreted.
In chicken embryos, BrdU administration produces delay in the development, growth retardation, mortality of embryos, and the appearance of defects in the ventral body wall (Barnigan et al., 1981; Gould et al., 1999a). In addition, when high doses of BrdU are administered into one of the embryo’s vitelline veins, isochronic clusters of neurons are produced in the chick dorsal telencephalon, which are artifacts. Interestingly, these structures were not observed when low doses of BrdU were administered (Rowell and Ragsdale, 2012).
In rodent embryos, on the other hand, it has been reported that BrdU alters the development of the heartbeat, visceral yolk sac circulation, erythrocyte and somite formation and closure of the otic vesicles (Nakashima et al., 1984). BrdU also interferes with the formation of Meckel’s cartilage, starting a chain of events leading through micrognathia and macroglossia to failure of palatal shelf reorientation and cleft palate (Bannigan et al., 1990). When administered in the prenatal life, BrdU also produces exencephaly (Bannigan et al., 1985), limbs, teeth, and tail deformities (Kolb et al., 1999), and behavioral abnormalities such as reduction of the male sexual behavior, alterations in the learning and memory in the Biel maze, increased ambulation and rearing in the open field (Kuwagata and Nagao, 1998). The most important data observed in this study is that rats injected with 50 mg/kg of BrdU on days 9 through 15 of gestation presented more locomotor activity and an impairment of learning and memory than animals administered with 100 mg/kg of this marker on days 16 through 20 of gestation. These results denote that the effects of the BrdU vary depending on the embryonic day of treatment. In the same line, Kolb et al. (1999) found that offspring of pregnant rats given two BrdU injections of 60 mg/kg spaced 6 hours apart on embryonic day 17 produced more severe deficits in spatial learning tasks than in animals administered with the marker on one of embryonic days 11, 12, 13 15 or 21. Taken together, these data have revealed that BrdU has detrimental effects on development and behavior of vertebrates. Therefore, it is proposed that, in terms of label proliferating cells, a low dose of BrdU should be used.
5-Bromo-2’-Deoxyuridine Labeling, and Prenatal and Perinatal Neurogenesis
BrdU has provided new opportunities for analyzing the fraction of S-phase cells during the prenatal and postnatal periods. BrdU labeling is a powerful tool to investigate cell proliferation and neurogenesis. It has generated valuable insights into the cellular mechanisms of the central nervous system development, including times of neuron origin (Martí et al., 2015, 2016), cell cycle studies, migration, and cell lineage (Nowakowski et al., 1989; Hervás et al., 2002; Molina et al., 2017; Martí and Rodríguez-Vázquez, 2020). Despite this, BrdU has unforeseen problems. The side effects of BrdU should not be considered negligible, neither in vitro nor in vivo. It has been reported that when cell lines derived from murine embryonic stem cells are exposed to BrdU, they lose the expression of stem cell markers like Nestin, Sox2 and Pax 6, and undergo glial differentiation, up-regulating the astrocytic marker GFAP. The latter was paralleled by a reduced expression of DNA methyltransferases and a rapid decrease of DNA methylation, suggesting that BrdU-labeled embryonic stem cells alter their DNA methylation status (Schneider and d’Adda di Fagagna, 2012). Other authors have reported that a single, low dose of BrdU has a severe antiproliferative effect in cultured neural stem cells, which is accompanied by altered cell differentiation, cell phenotype, and protein expression consistent with the induction of senescence (Ross et al., 2008). Biggers et al. (1987) have reported, during the organogenesis period, effects on neural tube differentiation of cultured embryos, as well as anomalies in the neurite outgrowth (reduction in length and number of these cytoplasmatic processes) from neuroblasts treated with BrdU. It has also been shown that BrdU incorporation in rat striatal precursors affects the differentiation of neurons, but no effects were reported when the differentiation of glial cells was considered. These results have revealed that neuronal precursors are more vulnerable than glial precursors to the toxic effects of the BrdU (Caldwell et al., 2005). In vivo studies, on the other hand, have shown that a single administration of BrdU at doses ranging from 100 to 300 mg/kg is able to alter the proliferative behavior of neuroblasts and leads to the activation of apoptotic cellular events in the rat cerebellar neuroepithelium (Rodríguez-Vázquez and Martí, 2021). Moreover, a single dose of BrdU (300 or 500 mg/kg) alters, in mice, the fusion of the neural folds due to cell necrosis in the neuroepithelium (Bannigan et al., 1985).
An important consideration about the BrdU detection into replicating DNA is that related to dilution of the marker across cell divisions. This is because after BrdU administration, BrdU-positive cells are expected to divide into BrdU-positive daughter cells. Repeated cell divisions may dilute the incorporated marker beyond the detection level. In this context, it is very important to choose the optimal dose of the BrdU to label cells in S-phase. This dose should be well tolerated and neither produces deleterious effects nor alters the proliferative dynamics of cells engaged in the synthetic phase of the cell cycle. A high dose of BrdU would have those effects. When administered in the embryonic period (Martí and Rodríguez-Vázquez, 2020; Rash et al., 2023) or in the perinatal life (Rodríguez-Vázquez and Martí, 2021), there is evidence showing that, in terms of label dividing neural progenitors, the dose of BrdU usually used in many laboratories (a single injection of 50–75 mg/kg) seems to be well tolerated and produces no striking effects, which suggest that this dosage is appropriate at least when the periods of time studied are short. Despite this, it cannot be excluded a more protracted effect of BrdU on neuroblasts’ cell biology, that is, neuron precursor differentiation and its final fate.
When the toxicity of repeated BrdU administrations on the proliferating neuroblasts are analyzed, the data are controversial. This is because previous reports have indicated that the administration of 50 mg/kg of BrdU in a cumulative labeling sequence during the prenatal life, is well tolerated and produces neither cytotoxic effects nor alters, at embryonic day 14, cell cycle dynamics in the dorsomedial region of the mouse cerebral wall (Takahashi et al., 1992, 1993, 1995a). No alterations have been reported when a succession of BrdU injections were administered on each of embryonic days 11 to 17 (Takahashi et al., 1995b). In line with these data, it has been shown that, in a cumulative BrdU (35 mg/kg) labeling sequence, no cytotoxic effects were observed in the rat cerebellar neuroepithelium (Martí and Rodríguez-Vázquez, 2020). In contrast, when several injections of BrdU were administered at 8-hour intervals over 2 successive days between days 11 and 21 of rat pregnancy and the offspring was analyzed in adulthood, defects in the proliferation, migration, and settling of the Purkinje cells were seen. Moreover, reductions in the sizes of the cerebellar cortex and deep nuclei, as well as foliation defects were found. Interestingly, the effects BrdU varied depending on the age of the animals (Sekerková et al., 2004b). In this line, results from my laboratory have revealed that when rats were administered with BrdU on embryonic days 13 and 14, following the abovementioned progressively delayed cumulative method, several granule cells were arrested in the molecular layer where they formed an ectopic zone. Examples of clusters of ectopic granule cells located in the bottom of the prima and secunda fissures are shown in Figure 2.
Figure 2.
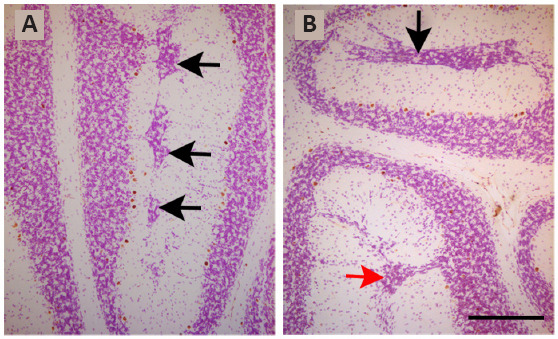
Light photomicrographs illustrating ectopic granule cells from rats administered with bromodeoxyuridine on embryonic days 13–14 and sacrificed on postnatal day 90.
Black arrows in “A” display ectopic neurons at the bottom of the prima fissure. Black and red arrows in “B” indicate ectopic granule cells at the bottom of the prima fissure and secunda fissures, respectively. Scale bar: 300 μm. Unpublished data.
On the other hand, when the timetables of neurogenesis and neurogenetic gradients of Purkinje cells and deep cerebellar nuclei were inferred in mice exposed to either BrdU or [3H]TdR as embryos are sacrificed in adulthood, systematic differences in the pattern of neurogenesis as well as in the spatial location of these macroneurons were observed (Martí et al., 2015). These results have indicated that both markers are not comparable, and discrepancies possibly arise from BrdU toxicity. In the same study, several cerebellar features of the cerebellum including length of the cerebellar cortex, the area of the molecular layer, the Purkinje cell number, the areas of the cerebellar nuclei, and the number of the deep cerebellar nuclei neurons were lower in animals exposed to BrdU than in those exposed to [3H]TdR. Again, data have revealed that the toxicity of BrdU alters the development of the central nervous system.
In line with these results, a significant study performed in macaque monkeys has provided strong evidence that the repeated administration of BrdU in the embryonic life compromised the number and survival of tagged neurons and disturbed neuroblasts’ migration as well as their final position in the adult cerebral cortex (Duque and Rakic, 2011).
5-Bromo-2’-Deoxyuridine Labeling and Adult Neurogenesis
The development of the central nervous system is a spatial and temporal regulated process. The vast majority of neurons in the adult mammalian brain arise during the embryonic and perinatal life. The adult mammalian brain and spinal cord present a limited capability to produce new neurons in adulthood. The neuroscientist Santiago Ramon y Cajal wrote: “once the development was ended, the fonts of growth and regeneration of the axons and dendrites dried up irrevocably. In the adult centers, the nerve paths are something fixed, ended and immutable. Everything may die, nothing may be regenerated. It is for the science of the future to change, if possible, this harsh decree” (Ramón y Cajal S, 1928). This dogma remained unchallenged until the early 1960s when Joseph Altman, using [3H]TdR autoradiography, described for first time the presence of newly generated neurons in the dentate gyrus of the adult rat hippocampus (Altman, 1962). Since then, several reports have indicated the addition of new neurons into the neocortex, the amygdala, the cornu Ammonis 1, the dorsal vagal complex of the brainstem, the spinal cord, and in the pars compacta of the substantia nigra adult mammals. Unfortunately, these reports have been unconfirmed, or they have been challenged with negative findings (Breunig et al., 2007; Duque and Spector, 2019).
There are two specific specialized brain regions which act as germinal centers. They have neural stem cells and sustain neurogenesis in their adult life. The first of these is the ventricular-subventricular area lining the lateral ventricles, and the second is the dentate gyrus in the hippocampus. In the first of these, new generated neurons migrate into the olfactory bulb via the rostral migratory stream. In the second, newly produced granule cells in the dentate gyrus migrate a short distance from the subgranular layer to the granular layer of the dentate gyrus and remain within this structure. Newly produced neurons in the ventricular-subventricular area and dentate gyrus establish synaptic contacts, and functional anatomical connections with neighboring neurons and glial cells (Lim and Alvarez-Buylla, 2016; Nogueira et al., 2022).
The existence of adult neurogenesis is accepted in several mammalian species. This phenomenon is considered to play an important role in cognitive ability, especially in learning and memory (Augusto-Oliveira et al., 2019; Petrik and Encinas, 2019). Production of neurons in adults is a well-established process in brain regions of lizards (González-Granero et al., 2023), birds, rodents, cows, dogs, and African elephants (Augusto-Oliveira et al., 2019). Furthermore, evidence of cell proliferation, neural stem cells and immature neurons has also been seen in marmosets, lemurs, macaques, and baboons (Petrik and Encinas, 2019). The absence or extremely low production rate of new hippocampal neurons during adulthood has been shown in chiropterans (Amrein et al., 2007), dolphins and whales (Patzle et al., 2015). Despite this adult neurogenesis comprises the production of new neurons from proliferating cells located in neurogenetic niches, the confirmation of BrdU-detected neurogenesis has not been accurately tested, and there are open questions such as how well this technology functions in adult animals. It is unknown whether, in adult animals from different species, different doses of BrdU have comparable labeling efficiencies for detecting S-phase cells in mitotically active areas of the nervous system. For example, in Gould et al. (Gould et al., 1999b), adult primates were administered one to five injections daily of 75 to 100 mg/kg of BrdU, and they were sacrificed from 2 hours to 2 weeks later. On the other hand, a single injection of 25–500 mg/kg was administered in mice (Mandyam et al., 2007), 50–600 mg/kg in rats (Cameron and Mckay, 2001). The issue dose is important because the toxicity of the BrdU as well as its detrimental effects on the cycle dynamic have been demonstrated (Taupin, 2007; Lehner et al., 2011; Rodríguez-Vázquez and Martí, 2021).
Adult human neurogenesis is currently a phenomenon under debate and intense controversy (Gould et al., 1999b; Duque and Spector, 2019; Nakafuku and del Águila, 2020; Moreno-Jiménez et al., 2021). From a historical perspective, in the late 1990s, Eriksson et al. (1990) in a pioneering study described the presence of adult neurogenesis in the human hippocampus after studying the post-mortem brain from five patients injected with 250 mg of intravenous BrdU for therapeutic purposes (tracking the proliferative activity of cancer cells). BrdU incorporation was found in hippocampal granule cells. Later, Curtis et al. (2007) showed, in adult humans administered with BrdU to assess the proliferative behavior of the tumor cells, the presence of the rostral migratory stream, which unsheathing the lateral olfactory ventricular extension. In the same study, it was revealed that this migratory pathway presents progenitor cells with migratory characteristics as well as cells that incorporated BrdU and become mature neurons in the olfactory bulb. These remarkable findings were complemented with the results from Ernst et al. (2014). They reported that the ventricular-subventricular zone is the source of neuroblasts and new neurons found in the human striatum. These investigations were further supported by research correlating atmospheric 14C released by nuclear bomb testing with the incorporation of 14C into DNA of proliferating cells (Ernst et al., 2014; Zhao and van Praag, 2020).
These studies were complemented by the discovery, in vitro, of BrdU-reactive neural precursor cells in surgical specimens from the adult human hippocampus (Kukekov et al., 1999; Roy et al., 2000; Arsenijevic et al., 2001; Nunes et al., 2003). In spite of the fact that these in vivo and in vitro experiments have shown several strong examples of BrdU-immunoreactive neurons, there are some limitations interpretating BrdU labeling that have not been considered. This is because, unfortunately, it has been ignoring the warning expressed in many reports published by the author and others about the possibility of false BrdU labeling or the incorrect interpretation of such tagging (Taupin 2007; Duque and Rakic, 2011; Lehner et al., 2011; Duque and Spector, 2019; Martí-Clúa, 2021b). The first of these limitations is related to the techniques used for producing a single-stranded DNA and subsequent BrdU immunodetection. Antigen retrieval procedures can produce BrdU-positive cells even in the absence of BrdU administration or even primary or secondary antibodies in human nervous tissue (Breuning et al., 2007). The second is due to the specificity of BrdU. This nucleoside is not a good marker of cell proliferation, but an indicator of DNA synthesis, which also takes place when the cell attempts to repair DNA itself or is undergoing an apoptotic cellular event (Breunig et al., 2007; Taupin, 2007; Duque and Rakic, 2011; Duque and Spector, 2019; Duque et al., 2022). These results emphasize the idea that even if BrdU is widely used and its immunodetection has been the basis for most studies on adult neurogenesis, its interpretation is not trivial.
BrdU is widely used to identify proliferating neuroblasts and their progenies. This synthetic halogenated pyrimidine is a powerful technology to investigate neuroblast proliferation and the genesis of neurons from the embryonic life to adulthood. Despite this, BrdU labeling presents methodological problems. They can be overcome via co-labeling, in the same tissue section, BrdU immunostaining with cell cycle markers (tritiated thymidine, Ki67, mini-chromosome maintenance protein-2, PCNA, phosphohistone-H3 and ribonucleotide reductase) and immature neuron markers (polysialylated neuronal cell adhesion molecule and doublecortin) (Zhao and van Praag, 2020; Sorrells et al., 2021; González-Granero et al., 2023). Furthermore, intracranial injection of retroviral vectors to label dividing progenitor cells allows to infer times of neuron origin and follow their fate. Another effective approach is the use of transgenic animals in which a fluorescent reporter is driven by promoters specific at different stages of new development, such as nestin, glial fibrillary acidic protein, sex determining region Y-box2, T-box brain protein 2, doublecortin and homeodomain-only protein (Zhao and van Praag, 2020).
Conclusions
BrdU immunohistochemistry is a common tool to investigate genesis, migration, and cell fate mapping. However, this current review has implications for the interpretation of results obtained by BrdU immunohistochemistry as an index of the proportion of S-phase cells. BrdU has pitfalls and methodological problems. The effect of BrdU incorporation into nascent DNA should not be underestimated by investigators, because it may lead to false results and misinterpretation in the identification of proliferative neuron precursors if: (i) the limitations of BrdU labeling are not accepted, and (ii) the data obtained are not prudently interpreted. The use of endogenous markers for cell cycle, markers of immature neurons retroviral vectors to tag proliferating progenitor cells and the use of transgenic animals will supply corroborating evidence of neurogenesis both in the prenatal and in the postnatal life.
Footnotes
Conflicts of interest: The authors declare no conflicts of interest.
Data availability statement: None declared.
C-Editors: Zhao M, Sun Y, Song LP; T-Editor: Jia Y
References
- 1.Allin MP. Novel insights from quantitative imaging of the developing cerebellum. Semin Fetal Neonatal Med. (2016);21:333–338. doi: 10.1016/j.siny.2016.06.003. [DOI] [PubMed] [Google Scholar]
- 2.Altman J. Are new neurons formed in the brains of adult mammals? Science. (1962);135:1127–1128. doi: 10.1126/science.135.3509.1127. [DOI] [PubMed] [Google Scholar]
- 3.Altman J, Bayer SA. CRC Press, Boca Raton; (1997). Development of the cerebellar system in relation to its evolution. structure, and functions. [Google Scholar]
- 4.Amrein I, Dechmann DK, Winter Y, Lipp HP. Absent or low rate of adult neurogenesis in the hippocampus of bats (Chiroptera) PLoS One. (2007);2:e455. doi: 10.1371/journal.pone.0000455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Arsenijevic Y, Villemure JG, Brunet JF, Bloch JJ, Déglon N, Kostic C, Zurn A, Aebischer P. Isolation of multipotent neural precursors residing in the cortex of the adult human brain. Exp Neurol. (2001);170:48–62. doi: 10.1006/exnr.2001.7691. [DOI] [PubMed] [Google Scholar]
- 6.Augusto-Oliveira M, Arrifano GPF, Malva JO, Crespo-Lopez ME. Adult hippocampal neurogenesis in different taxonomic groups:possible functional similarities and striking controversies. Cells. (2019);8:125. doi: 10.3390/cells8020125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bak PM, Panos RJ. Protease antigen recovery decreases the specificity of bromodeoxyuridine detection in formalin-fixed tissue. J Histochem Cytochem. (1997);45:1165–1170. doi: 10.1177/002215549704500814. [DOI] [PubMed] [Google Scholar]
- 8.Bannigan J, Langman J, van Breda A. The uptake of 5-bromodeoxyuridine by the chicken embryo and its effects upon growth. Anat Embryol (Berl) (1981);162:425–434. doi: 10.1007/BF00301868. [DOI] [PubMed] [Google Scholar]
- 9.Bannigan JG. The effects of 5-bromodeoxyuridine on fusion of the cranial neural folds in the mouse embryo. Teratology. (1985);32:229–239. doi: 10.1002/tera.1420320211. [DOI] [PubMed] [Google Scholar]
- 10.Bannigan JG, Cottell DC, Morris A. Study of the mechanisms of BUdR-induced cleft palate in the mouse. Teratology. (1990);42:79–89. doi: 10.1002/tera.1420420110. [DOI] [PubMed] [Google Scholar]
- 11.Biggers WJ, Barnea ER, Sanyal MK. Anomalous neural differentiation induced by 5-bromo-2'-deoxyuridine during organogenesis in the rat. Teratology. (1987);35:63–75. doi: 10.1002/tera.1420350110. [DOI] [PubMed] [Google Scholar]
- 12.Böswald M, Harasim S, Maurer-Schultze B. Tracer dose and availability time of thymidine and bromodeoxyuridine:application of bromodeoxyuridine in cell kinetic studies. Cell Tissue Kinet. (1990);23:169–181. doi: 10.1111/j.1365-2184.1990.tb01113.x. [DOI] [PubMed] [Google Scholar]
- 13.Breunig JJ, Arellano JI, Macklis JD, Rakic P. Everything that glitters isn't gold:a critical review of postnatal neural precursor analyses. Cell Stem Cell. (2007);1:612–627. doi: 10.1016/j.stem.2007.11.008. [DOI] [PubMed] [Google Scholar]
- 14.Burns TC, Ortiz-González XR, Gutiérrez-Pérez M, Keene CD, Sharda R, Demorest ZL, Jiang Y, Nelson-Holte M, Soriano M, Nakagawa Y, Luquin MR, Garcia-Verdugo JM, Caldwell MA, He X, Svendsen CN. 5-Bromo-2'-deoxyuridine is selectively toxic to neuronal precursors in vitro. Eur J Neurosci. (2005);22:2965–2970. doi: 10.1111/j.1460-9568.2005.04504.x. [DOI] [PubMed] [Google Scholar]
- 15.Cameron HA, McKay RD. Adult neurogenesis produces a large pool of new granule cells in the dentate gyrus. J Comp Neurol. (2001);435:406–417. doi: 10.1002/cne.1040. [DOI] [PubMed] [Google Scholar]
- 16.Cavanagh BL, Walker T, Norazit A, Meedeniya AC. Thymidine analogues for tracking DNA synthesis. Molecules. (2011);16:7980–7993. doi: 10.3390/molecules16097980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Clarkson B, Ohkita T, Ota K, Fried J. Studies of cellular proliferation in human leukemia. I. Estimation of growth rates of leukemic and normal hematopoietic cells in two adults with acute leukemia given single injections of tritiated thymidine. J Clin Invest. (1967);46:506–529. doi: 10.1172/JCI105553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Curtis MA, Kam M, Nannmark U, Anderson MF, Axell MZ, Wikkelso C, Holtås S, van Roon-Mom WM, Björk-Eriksson T, Nordborg C, Frisén J, Dragunow M, Faull RL, Eriksson PS. Human neuroblasts migrate to the olfactory bulb via a lateral ventricular extension. Science. (2007);315:1243–1249. doi: 10.1126/science.1136281. [DOI] [PubMed] [Google Scholar]
- 19.Dinjens WN, ten Kate J, Lenders MH, van der Linden EP, Bosman FT. Bromodeoxyuridine (BrdU) immunocytochemistry by exonuclease III (Exo III) digestion. Histochemistry. (1992);98:199–205. doi: 10.1007/BF00315878. [DOI] [PubMed] [Google Scholar]
- 20.Dolbeare F, Gray JW. Use of restriction endonucleases and exonuclease III to expose halogenated pyrimidines for immunochemical staining. Cytometry. (1988);9:631–635. doi: 10.1002/cyto.990090619. [DOI] [PubMed] [Google Scholar]
- 21.Dolbeare F. Bromodeoxyuridine:a diagnostic tool in biology and medicine, Part I:Historical perspectives, histochemical methods and cell kinetics. Histochem J. (1995);27:339–369. [PubMed] [Google Scholar]
- 22.Duque A, Rakic P. Different effects of bromodeoxyuridine and [3H]thymidine incorporation into DNA on cell proliferation, position , and fate. J Neurosci. (2011);31:15205–15217. doi: 10.1523/JNEUROSCI.3092-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Duque A, Krsnik Z, Kostović I, Rakic P. Secondary expansion of the transient subplate zone in the developing cerebrum of human and nonhuman primates. Proc Natl Acad Sci U S A. (2016);113:9892–9897. doi: 10.1073/pnas.1610078113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Duque A, Spector R. A balanced evaluation of the evidence for adult neurogenesis in humans:implication for neuropsychiatric disorders. Brain Struct Funct. (2019);224:2281–2295. doi: 10.1007/s00429-019-01917-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Eadie BD, Redila VA, Christie BR. Voluntary exercise alters the cytoarchitecture of the adult dentate gyrus by increasing cellular proliferation, dendritic complexity, and spine density. J Comp Neurol. (2005);486:39–47. doi: 10.1002/cne.20493. [DOI] [PubMed] [Google Scholar]
- 26.En A, Watanabe K, Ayusawa D, Fujii M. The key role of a basic domain of histone H2B N-terminal tail in the action of 5-bromodeoxyuridine to induce cellular senescence. FEBS J. (2023);290:692–711. doi: 10.1111/febs.16584. [DOI] [PubMed] [Google Scholar]
- 27.Eriksson PS, Perfilieva E, Björk-Eriksson T, Alborn AM, Nordborg C, Peterson DA, Gage FH. Neurogenesis in the adult human hippocampus. Nat Med. (1998);4:1313–1317. doi: 10.1038/3305. [DOI] [PubMed] [Google Scholar]
- 28.Erlanger BF, Beiser SM. Antibodies specific for ribonucleosides and ribonucleotides and their reaction with dna. Proc Natl Acad Sci U S A. (1964);52:68–74. doi: 10.1073/pnas.52.1.68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Ernst A, Alkass K, Bernard S, Salehpour M, Perl S, Tisdale J, Possnert G, Druid H, Frisén J. Neurogenesis in the striatum of the adult human brain. Cell. (2014);156:1072–1083. doi: 10.1016/j.cell.2014.01.044. [DOI] [PubMed] [Google Scholar]
- 30.Fang C, Wang H, Naumann RK. Developmental Patterning and Neurogenetic Gradients of Nurr1 Positive Neurons in the Rat Claustrum and Lateral Cortex. Front Neuroanat. (2021);15:786329. doi: 10.3389/fnana.2021.786329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ffrench M, Morel F, Souchier C, Benchaib M, Catallo R, Bryon PA. Choice of fixation and denaturation for the triple labelling of intra-cytoplasmic antigen, bromodeoxyuridine and DNA. Application to bone marrow plasma cells. Histochemistry. (1994);101:385–390. doi: 10.1007/BF00269001. [DOI] [PubMed] [Google Scholar]
- 32.Frade JM, Ovejero-Benito MC. Neuronal cell cycle:the neuron itself and its circumstances. Cell Cycle. (2015);14:712–720. doi: 10.1080/15384101.2015.1004937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.González-Granero S, Font E, Desfilis E, Herranz-Pérez V, García-Verdugo JM. Adult neurogenesis in the telencephalon of the lizard Podarcis liolepis. Front Neurosci. (2023);17:1125999. doi: 10.3389/fnins.2023.1125999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Gould TW, Burek MJ, Sosnowski JM, Prevette D, Oppenheim RW. The spatial-temporal gradient of naturally occurring motoneuron death reflects the time of prior exit from the cell cycle and position within the lateral motor column. Dev Biol. (1999a);216:611–621. doi: 10.1006/dbio.1999.9490. [DOI] [PubMed] [Google Scholar]
- 35.Gould E, Reeves AJ, Graziano MS, Gross CG. Neurogenesis in the neocortex of adult primates. Science. (1999b);286:548–552. doi: 10.1126/science.286.5439.548. [DOI] [PubMed] [Google Scholar]
- 36.Gratzner HG, Leif RC, Ingram DJ, Castro A. The use of antibody specific for bromodeoxyuridine for the immunofluorescent determination of DNA replication in single cells and chromosomes. Exp Cell Res. (1975);95:88–94. doi: 10.1016/0014-4827(75)90612-6. [DOI] [PubMed] [Google Scholar]
- 37.Gratzner HG, Pollack A, Ingram DJ, Leif RC. Deoxyribonucleic acid replication in single cells and chromosomes by immunologic techniques. J Histochem Cytochem. (1976);24:34–39. doi: 10.1177/24.1.815428. [DOI] [PubMed] [Google Scholar]
- 38.Gratzner HG. Monoclonal antibody to 5-bromo- and 5-iododeoxyuridine:a new reagent for detection of DNA replication. Science. (1982);218:474–475. doi: 10.1126/science.7123245. [DOI] [PubMed] [Google Scholar]
- 39.Hakala MT. Mode of action of 5-bromodeoxyuridine on mammalian cells in culture. J Biol Chem. (1959);234:3072–3076. [PubMed] [Google Scholar]
- 40.Herrup K, Neve R, Ackerman SL, Copani A. Divide and die:cell cycle events as triggers of nerve cell death. J Neurosci. (2004);24:9232–9239. doi: 10.1523/JNEUROSCI.3347-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Hervás JP, Martí-Clúa J, Muñoz-García A, Santa-Cruz MC. Proliferative activity in the cerebellar external granular layer evaluated by bromodeoxyuridine labeling. Biotech Histochem. (2002);77:27–35. [PubMed] [Google Scholar]
- 42.Hume WJ, Saffhill R. Iodo- and bromodeoxyuridine are excised at different rates from DNA of mouse tongue keratinocytes in vitro. Chem Biol Interact. (1986);60:227–232. doi: 10.1016/0009-2797(86)90031-1. [DOI] [PubMed] [Google Scholar]
- 43.Keoffler HP, Yen J, Carlson J. The study of human myeloid differentiation using bromodeoxyuridine (BrdU) J Cell Physiol. (1983);116:111–117. doi: 10.1002/jcp.1041160117. [DOI] [PubMed] [Google Scholar]
- 44.Kim HS, Shin SM, Kim S, Nam Y, Yoo A, Moon M. Relationship between adult subventricular neurogenesis and Alzheimer's disease:Pathologic roles and therapeutic implications. Front Aging Neurosci. (2022);14:1002281. doi: 10.3389/fnagi.2022.1002281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kolb B, Pedersen B, Ballermann M, Gibb R, Whishaw IQ. Embryonic and postnatal injections of bromodeoxyuridine produce age-dependent morphological and behavioral abnormalities. J Neurosci. (1999);19:2337–2346. doi: 10.1523/JNEUROSCI.19-06-02337.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kuan CY, Schloemer AJ, Lu A, Burns KA, Weng WL, Williams MT, Strauss KI, Vorhees CV, Flavell RA, Davis RJ, Sharp FR, Rakic P. Hypoxia-ischemia induces DNA synthesis without cell proliferation in dying neurons in adult rodent brain. J Neurosci. (2004);24:10763–10772. doi: 10.1523/JNEUROSCI.3883-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kubikova L, Polomova J, Mikulaskova V, Lukacova K. Effectivity of two cell proliferation markers in brain of a songbird zebra finch. Biology (Basel) (2020);9:356. doi: 10.3390/biology9110356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kukekov VG, Laywell ED, Suslov O, Davies K, Scheffler B, Thomas LB, O'Brien TF, Kusakabe M, Steindler DA. Multipotent stem/progenitor cells with similar properties arise from two neurogenic regions of adult human brain. Exp Neurol. (1999);156:333–344. doi: 10.1006/exnr.1999.7028. [DOI] [PubMed] [Google Scholar]
- 49.Kuwagata M, Nagao T. Behavior and reproductive function of rat male offspring treated prenatally with 5-bromo-2'-deoxyuridine. Reprod Toxicol. (1998);12:541–549. doi: 10.1016/s0890-6238(98)00037-9. [DOI] [PubMed] [Google Scholar]
- 50.Larson TA, Thatra NM, Hou D, Hu RA, Brenowitz EA. Seasonal changes in neuronal turnover in a forebrain nucleus in adult songbirds. J Comp Neurol. (2019);527:767–779. doi: 10.1002/cne.24552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lawless OJ, Bellanti JA, Brown ML, Sandberg K, Umans JG, Zhou L, Chen W, Wang J, Wang K, Zheng SG. In vitro induction of T regulatory cells by a methylated CpG DNA sequence in humans:Potential therapeutic applications in allergic and autoimmune diseases. Allergy Asthma Proc. (2018);39:143–152. doi: 10.2500/aap.2018.39.4113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Leif RC, Stein JH, Zucker RM. A short history of the initial application of anti-5-BrdU to the detection and measurement of S phase. Cytometry A. (2004);58:45–52. doi: 10.1002/cyto.a.20012. [DOI] [PubMed] [Google Scholar]
- 53.Lehner B, Sandner B, Marschallinger J, Lehner C, Furtner T, Couillard-Despres S, Rivera FJ, Brockhoff G, Bauer HC, Weidner N, Aigner L. The dark side of BrdU in neural stem cell biology:detrimental effects on cell cycle, differentiation and survival. Cell Tissue Res. (2011);345:313–328. doi: 10.1007/s00441-011-1213-7. [DOI] [PubMed] [Google Scholar]
- 54.Leuner B, Glasper ER, Gould E. Thymidine analog methods for studies of adult neurogenesis are not equally sensitive. J Comp Neurol. (2009);517:123–333. doi: 10.1002/cne.22107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Levkoff LH, Marshall GP, 2nd, Ross HH, Caldeira M, Reynolds BA, Cakiroglu M, Mariani CL, Streit WJ, Laywell ED. Bromodeoxyuridine inhibits cancer cell proliferation in vitro and in vivo. Neoplasia. (2008);10:804–816. doi: 10.1593/neo.08382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Liboska R, Ligasová A, Strunin D, Rosenberg I, Koberna K. Most anti-BrdU antibodies react with 2'-deoxy-5-ethynyluridine -- the method for the effective suppression of this cross-reactivity. PLoS One. (2012);7:e51679. doi: 10.1371/journal.pone.0051679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Lim DA, Alvarez-Buylla A. The adult ventricular-subventricular zone (V-SVZ) and olfactory bulb (OB) neurogenesis. Cold Spring Harb Perspect Biol. (2016);8:a018820. doi: 10.1101/cshperspect.a018820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Long Y, Wang M, Gu H, Xie X. Bromodeoxyuridine promotes full-chemical induction of mouse pluripotent stem cells. Cell Res. (2015);25:1171–1174. doi: 10.1038/cr.2015.96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Mandyam CD, Harburg GC, Eisch AJ. Determination of key aspects of precursor cell proliferation, cell cycle length and kinetics in the adult mouse subgranular zone. Neuroscience. (2007);146:108–122. doi: 10.1016/j.neuroscience.2006.12.064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Martí J, Wills KV, Ghetti B, Bayer SA. A combined immunohistochemical and autoradiographic method to detect midbrain dopaminergic neurons and determine their time of origin. Brain Res Brain Res Protoc. (2002);9:197–205. doi: 10.1016/s1385-299x(02)00145-9. [DOI] [PubMed] [Google Scholar]
- 61.Martí J, Santa-Cruz MC, Serra R, Hervás JP. Systematic differences in time of cerebellar-neuron origin derived from bromodeoxyuridine immunoperoxidase staining protocols and tritiated thymidine autoradiography:a comparative study. Int J Dev Neurosci. (2015);47:216–228. doi: 10.1016/j.ijdevneu.2015.09.001. [DOI] [PubMed] [Google Scholar]
- 62.Martí J, Santa-Cruz MC, Serra R, Hervás JP. Hydroxyurea treatment and development of the rat cerebellum:effects on the neurogenetic profiles and settled patterns of purkinje cells and deep cerebellar nuclei neurons. Neurotox Res. (2016);30:563–580. doi: 10.1007/s12640-016-9649-x. [DOI] [PubMed] [Google Scholar]
- 63.Martí J, Rodríguez-Vázquez L. An immunocytochemical approach to the analysis of the cell division cycle in the rat cerebellar neuroepithelium. Cell Cycle. (2020);19:2451–2459. doi: 10.1080/15384101.2020.1806425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Martí-Clúa J. Developmental timetables and gradients of neurogenesis in cerebellar Purkinje cells and deep glutamatergic neurons:a comparative study between the mouse and the rat. Anat Rec (Hoboken) (2021a);304:2856–2864. doi: 10.1002/ar.24607. [DOI] [PubMed] [Google Scholar]
- 65.Martí-Clúa J. Incorporation of 5-bromo-2'-deoxyuridine into DNA and proliferative behavior of cerebellar neuroblasts:all that glitters is not gold. Cells. (2021b);10:1453. doi: 10.3390/cells10061453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Martí-Clua J. Times of neuron origin and neurogenetic gradients in mice Purkinje cells and deep cerebellar nuclei neurons during the development of the cerebellum. A review. Tissue Cell. (2022);78:101897. doi: 10.1016/j.tice.2022.101897. [DOI] [PubMed] [Google Scholar]
- 67.McGinley JN, Knott KK, Thompson HJ. Effect of fixation and epitope retrieval on BrdU indices in mammary carcinomas. J Histochem Cytochem. (2000);48:355–362. doi: 10.1177/002215540004800305. [DOI] [PubMed] [Google Scholar]
- 68.Messier B, Leblond CP, Smart I. Presence of DNA synthesis and mitosis in the brain of young adult mice. Exp Cell Res. (1958);14:224–226. doi: 10.1016/0014-4827(58)90235-0. [DOI] [PubMed] [Google Scholar]
- 69.Miale IL, Sidman RL. An autoradiographic analysis of histogenesis in the mouse cerebellum. Exp Neurol. (1961);4:277–296. doi: 10.1016/0014-4886(61)90055-3. [DOI] [PubMed] [Google Scholar]
- 70.Michishita E, Nakabayashi K, Suzuki T, Kaul SC, Ogino H, Fujii M, Mitsui Y, Ayusawa D. 5-Bromodeoxyuridine induces senescence-like phenomena in mammalian cells regardless of cell type or species. J Biochem. (1999);126:1052–1059. doi: 10.1093/oxfordjournals.jbchem.a022549. [DOI] [PubMed] [Google Scholar]
- 71.Miller MW, Nowakowski RS. Use of bromodeoxyuridine-immunohistochemistry to examine the proliferation, migration and time of origin of cells in the central nervous system. Brain Res. (1988);457:44–52. doi: 10.1016/0006-8993(88)90055-8. [DOI] [PubMed] [Google Scholar]
- 72.Molina V, Rodríguez-Vázquez L, Owen D, Valero O, Martí J. Cell cycle analysis in the rat external granular layer evaluated by several bromodeoxyuridine immunoperoxidase staining protocols. Histochem Cell Biol. (2017);148:477–488. doi: 10.1007/s00418-017-1593-1. [DOI] [PubMed] [Google Scholar]
- 73.Moreno-Jiménez EP, Terreros-Roncal J, Flor-García M, Rábano A, Llorens-Martín M. Evidences for adult hippocampal neurogenesis in humans. J Neurosci. (2021);41:2541–2553. doi: 10.1523/JNEUROSCI.0675-20.2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Morris SM, Domon OE, McGarrity LJ, Kodell RL, Casciano DA. Effect of bromodeoxyuridine on the proliferation and growth of ethyl methanesulfonate-exposed P3 cells:relationship to the induction of sister-chromatid exchanges. Cell Biol Toxicol. (1992);8:75–87. doi: 10.1007/BF00119296. [DOI] [PubMed] [Google Scholar]
- 75.Nakafuku M, Del Águila Á. Developmental dynamics of neurogenesis and gliogenesis in the postnatal mammalian brain in health and disease:Historical and future perspectives. Wiley Interdiscip Rev Dev Biol. (2020);9:e369. doi: 10.1002/wdev.369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Nakashima K, Ninomiya H, Fujiki Y. The effect of 5-bromodeoxyuridine on mouse embryos during neurulation in vitro. Experientia. (1984);40:924–929. doi: 10.1007/BF01946441. [DOI] [PubMed] [Google Scholar]
- 77.Nandakumar S, Rozich E, Buttitta L. Cell cycle re-entry in the nervous system:from polyploidy to neurodegeneration. Front Cell Dev Biol. (2021);9:698661. doi: 10.3389/fcell.2021.698661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Nogueira AB, Hoshino HSR, Ortega NC, Dos Santos BGS, Teixeira MJ. Adult human neurogenesis:early studies clarify recent controversies and go further. Metab Brain Dis. (2022);37:153–172. doi: 10.1007/s11011-021-00864-8. [DOI] [PubMed] [Google Scholar]
- 79.Nowakowski RS, Lewin SB, Miller MW. Bromodeoxyuridine immunohistochemical determination of the lengths of the cell cycle and the DNA-synthetic phase for an anatomically defined population. J Neurocytol. (1989);18:311–318. doi: 10.1007/BF01190834. [DOI] [PubMed] [Google Scholar]
- 80.Nowakowski RS, Hayes NL. New neurons:extraordinary evidence or extraordinary conclusion? Science. (2000);288:771. doi: 10.1126/science.288.5467.771a. [DOI] [PubMed] [Google Scholar]
- 81.Nunes MC, Roy NS, Keyoung HM, Goodman RR, McKhann G, 2nd, Jiang L, Kang J, Nedergaard M, Goldman SA. Identification and isolation of multipotential neural progenitor cells from the subcortical white matter of the adult human brain. Nat Med. (2003);9:439–947. doi: 10.1038/nm837. [DOI] [PubMed] [Google Scholar]
- 82.Patzke N, Spocter MA, Karlsson KÆ, Bertelsen MF, Haagensen M, Chawana R, Streicher S, Kaswera C, Gilissen E, Alagaili AN, Mohammed OB, Reep RL, Bennett NC, Siegel JM, Ihunwo AO, Manger PR. In contrast to many other mammals, cetaceans have relatively small hippocampi that appear to lack adult neurogenesis. Brain Struct Funct. (2015);220:361–383. doi: 10.1007/s00429-013-0660-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Pavulraj S, Stout RW, Paulsen DB, Chowdhury SI. Live triple gene-deleted pseudorabies virus-vectored subunit PCV2b and CSFV vaccine undergoes an abortive replication cycle in the tg neurons following latency reactivation. Viruses. (2023);15:473. doi: 10.3390/v15020473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Pederson T. Tritiated thymidine:a breakthrough in DNA replication and repair. FASEB J. (2014);28:2739–2741. doi: 10.1096/fj.14-0701ufm. [DOI] [PubMed] [Google Scholar]
- 85.Petrik D, Encinas JM. Perspective:of mice and men - how widespread is adult neurogenesis? Front Neurosci. (2019);13:923. doi: 10.3389/fnins.2019.00923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Qu TY, Dong XJ, Sugaya I, Vaghani A, Pulido J, Sugaya K. Bromodeoxyuridine increases multipotency of human bone marrow-derived stem cells. Restor Neurol Neurosci. (2004);22:459–468. [PubMed] [Google Scholar]
- 87.Ramón, Cajal S. (May RM, ed) II. London: Oxford University Press; (1928). Degeneration and regeneration of the nervous system; p. 750. [Google Scholar]
- 88.Rash BG, Arellano JI, Duque A, Rakic P. Role of intracortical neuropil growth in the gyrification of the primate cerebral cortex. Proc Natl Acad Sci U S A. (2023);120:e2210967120. doi: 10.1073/pnas.2210967120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Rodríguez-Vázquez L, Martí J. Administration of 5-bromo-2'-deoxyuridine interferes with neuroblast proliferation and promotes apoptotic cell death in the rat cerebellar neuroepithelium. J Comp Neurol. (2021);529:1081–1096. doi: 10.1002/cne.25005. [DOI] [PubMed] [Google Scholar]
- 90.Ross HH, Levkoff LH, Marshall GP, 2nd, Caldeira M, Steindler DA, Reynolds BA, Laywell ED. Bromodeoxyuridine induces senescence in neural stem and progenitor cells. Stem Cells. (2008);26:3218–3227. doi: 10.1634/stemcells.2008-0299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Rowell JJ, Ragsdale CW. BrdU birth dating can produce errors in cell fate specification in chick brain development. J Histochem Cytochem. (2012);60:801–810. doi: 10.1369/0022155412458588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Roy NS, Wang S, Jiang L, Kang J, Benraiss A, Harrison-Restelli C, Fraser RA, Couldwell WT, Kawaguchi A, Okano H, Nedergaard M, Goldman SA. In vitro neurogenesis by progenitor cells isolated from the adult human hippocampus. Nat Med. (2000);6:271–277. doi: 10.1038/73119. [DOI] [PubMed] [Google Scholar]
- 93.Ryu JR, Hong CJ, Kim JY, Kim EK, Sun W, Yu SW. Control of adult neurogenesis by programmed cell death in the mammalian brain. Mol Brain. (2016);9:43. doi: 10.1186/s13041-016-0224-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Sagga N, Kuffová L, Vargesson N, Erskine L, Collinson JM. Limbal epithelial stem cell activity and corneal epithelial cell cycle parameters in adult and aging mice. Stem Cell Res. (2018);33:185–198. doi: 10.1016/j.scr.2018.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Schneider L, d'Adda di Fagagna F. Neural stem cells exposed to BrdU lose their global DNA methylation and undergo astrocytic differentiation. Nucleic Acids Res. (2012);40:5332–5342. doi: 10.1093/nar/gks207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Sekerková G, Ilijic E, Mugnaini E. Time of origin of unipolar brush cells in the rat cerebellum as observed by prenatal bromodeoxyuridine labeling. Neuroscience. (2004a);127:845–858. doi: 10.1016/j.neuroscience.2004.05.050. [DOI] [PubMed] [Google Scholar]
- 97.Sekerková G, Ilijic E, Mugnaini E. Bromodeoxyuridine administered during neurogenesis of the projection neurons causes cerebellar defects in rat. J Comp Neurol. (2004b);470:221–239. doi: 10.1002/cne.11016. [DOI] [PubMed] [Google Scholar]
- 98.Shepherd CE, Yang Y, Halliday GM. Region- and cell-specific aneuploidy in brain aging and neurodegeneration. Neuroscience. (2018);374:326–334. doi: 10.1016/j.neuroscience.2018.01.050. [DOI] [PubMed] [Google Scholar]
- 99.Shimada A, Shibata T, Komatsu K, Nifuji A. Improved methods for immunohistochemical detection of BrdU in hard tissue. J Immunol Methods. (2008);339:11–16. doi: 10.1016/j.jim.2008.07.013. [DOI] [PubMed] [Google Scholar]
- 100.Solius GM, Maltsev DI, Belousov VV, Podgorny OV. Recent advances in nucleotide analogue-based techniques for tracking dividing stem cells:an overview. J Biol Chem. (2021);297:101345. doi: 10.1016/j.jbc.2021.101345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Sorrells SF, Paredes MF, Zhang Z, Kang G, Pastor-Alonso O, Biagiotti S, Page CE, Sandoval K, Knox A, Connolly A, Huang EJ, Garcia-Verdugo JM, Oldham MC, Yang Z, Alvarez-Buylla A. Positive controls in adults and children support that very few, if any, new neurons are born in the adult human hippocampus. J Neurosci. (2021);41:2554–2565. doi: 10.1523/JNEUROSCI.0676-20.2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Suzuki T, Minagawa S, Michishita E, Ogino H, Fujii M, Mitsui Y, Ayusawa D. Induction of senescence-associated genes by 5-bromodeoxyuridine in HeLa cells. Exp Gerontol. (2001);36:465–474. doi: 10.1016/s0531-5565(00)00223-0. [DOI] [PubMed] [Google Scholar]
- 103.Svandova E, Lesot H, Sharpe P, Matalova E. Making the head:caspases in life and death. Front Cell Dev Biol. (2023);10:1075751. doi: 10.3389/fcell.2022.1075751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Takahashi T, Nowakowski RS, Caviness VS., Jr BUdR as an S-phase marker for quantitative studies of cytokinetic behaviour in the murine cerebral ventricular zone. J Neurocytol. (1992);21:185–197. doi: 10.1007/BF01194977. [DOI] [PubMed] [Google Scholar]
- 105.Takahashi T, Nowakowski RS, Caviness VS., Jr Cell cycle parameters and patterns of nuclear movement in the neocortical proliferative zone of the fetal mouse. J Neurosci. (1993);13:820–33. doi: 10.1523/JNEUROSCI.13-02-00820.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Takahashi T, Nowakowski RS, Caviness VS., Jr The cell cycle of the pseudostratified ventricular epithelium of the embryonic murine cerebral wall. J Neurosci. (1995a);15:6046–6057. doi: 10.1523/JNEUROSCI.15-09-06046.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Takahashi T, Nowakowski RS, Caviness VS., Jr Early ontogeny of the secondary proliferative population of the embryonic murine cerebral wall. J Neurosci. (1995b);15:6058–6068. doi: 10.1523/JNEUROSCI.15-09-06058.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Tang X, Falls DL, Li X, Lane T, Luskin MB. Antigen-retrieval procedure for bromodeoxyuridine immunolabeling with concurrent labeling of nuclear DNA and antigens damaged by HCl pretreatment. J Neurosci. (2007);27:5837–5844. doi: 10.1523/JNEUROSCI.5048-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Taupin P. BrdU immunohistochemistry for studying adult neurogenesis:paradigms, pitfalls , limitations, and validation. Brain Res Rev. (2007);53:198–214. doi: 10.1016/j.brainresrev.2006.08.002. [DOI] [PubMed] [Google Scholar]
- 110.Terreros-Roncal J, Moreno-Jiménez EP, Flor-García M, Rodríguez-Moreno CB, Trinchero MF, Cafini F, Rábano A, Llorens-Martín M. Impact of neurodegenerative diseases on human adult hippocampal neurogenesis. Science. (2021);374:1106–1113. doi: 10.1126/science.abl5163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Vanderlaan M, Thomas CB. Characterization of monoclonal antibodies to bromodeoxyuridine. Cytometry. (1985);6:501–505. doi: 10.1002/cyto.990060603. [DOI] [PubMed] [Google Scholar]
- 112.Vincent I, Rosado M, Davies P. Mitotic mechanisms in Alzheimer's disease? J Cell Biol. (1996);132:413–425. doi: 10.1083/jcb.132.3.413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Wakayama T, Nakata H, Kumchantuek T, Gewaily MS, Iseki S. Identification of 5-bromo-2'-deoxyuridine-labeled cells during mouse spermatogenesis by heat-induced antigen retrieval in lectin staining and immunohistochemistry. J Histochem Cytochem. (2015);63:190–205. doi: 10.1369/0022155414564870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Yamada K, Semba R, Ding X, Ma N, Nagahama M. Discrimination of cell nuclei in early S-phase, mid-to-late S-phase, and G(2)/M-phase by sequential administration of 5-bromo-2'-deoxyuridine and 5-chloro-2'-deoxyuridine. J Histochem Cytochem. (2005);53:1365–1370. doi: 10.1369/jhc.4A6601.2005. [DOI] [PubMed] [Google Scholar]
- 115.Ye W, Blain SW. S phase entry causes homocysteine-induced death while ataxia telangiectasia and Rad3 related protein functions anti-apoptotically to protect neurons. Brain. (2010);133:2295–2312. doi: 10.1093/brain/awq139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Zheng Y, Begum S, Zhang C, Fleming K, Masumura C, Zhang M, Smith P, Darlington C. Increased BrdU incorporation reflecting DNA repair, neuronal de-differentiation or possible neurogenesis in the adult cochlear nucleus following bilateral cochlear lesions in the rat. Exp Brain Res. (2011);210:477–487. doi: 10.1007/s00221-010-2491-0. [DOI] [PubMed] [Google Scholar]
- 117.Zhao M, Momma S, Delfani K, Carlen M, Cassidy RM, Johansson CB, Brismar H, Shupliakov O, Frisen J, Janson AM. Evidence for neurogenesis in the adult mammalian substantia nigra. Proc Natl Acad Sci U S A. (2003);100:7925–7930. doi: 10.1073/pnas.1131955100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Zhao X, van Praag H. Steps towards standardized quantification of adult neurogenesis. Nat Commun. (2020);11:4275. doi: 10.1038/s41467-020-18046-y. [DOI] [PMC free article] [PubMed] [Google Scholar]