

Abstract
The nucleocapsid (N) protein of the Toscana (TOS) virus was expressed in Escherichia coli by using a pET15b vector. The recombinant protein was purified by affinity chromatography and was characterized by sodium dodecyl sulfate-polyacrylamide gel electrophoresis, immunoblotting, and enzyme immunoassay (EIA). The recombinant antigen was reactive with positive human sera, and the reactivity correlated very well (r = 0.9) with that of a whole-virus antigen when tested by EIA with 30 TOS virus-positive and 30 TOS virus-negative serum samples. The results demonstrate that the recombinant N protein can be easily produced in a procaryotic system and used for diagnostic assays for TOS virus immunity.
Toscana (TOS) virus belongs to the Bunyaviridae family and possesses a tripartite negative-strand RNA genome, with the large segment coding for the virus polymerase, the medium segment coding for the G1 and G2 envelope glycoproteins, and the small segment coding for the nucleocapsid (N) protein and the nonstructural protein (1, 2, 6, 10). TOS virus has repeatedly been isolated from Phlebotomus perniciosus and Phlebotomus perfiliewi in different foci in central Italy (4, 13, 22). A high prevalence of antibodies against TOS virus was demonstrated in healthy individuals from the Tuscany region (3). The virus can cause headache, myalgia, fever, and aseptic meningitis or meningoencephalitis, which may be clinically more severe in adults (2, 18). This neuropathic infection is more frequent during summer (7, 8, 16, 17). At present, TOS virus infection is diagnosed directly by reverse transcription-PCR (RT-PCR) with cerebrospinal fluid (15, 20, 21) or by the serologic detection of specific antibodies by indirect immunofluorescence assay, complement fixation, hemagglutination-inhibition test (5), and enzyme immunoassay (EIA) with TOS virus antigens purified from infected cell cultures (15) or from previously infected mouse brain (3). Since the TOS virus N protein has been identified as the major immunodominant protein (14), we have produced this antigen by genetic expression in Escherichia coli to study whether the recombinant protein can be used in the diagnosis of TOS virus infection.
MATERIALS AND METHODS
Sera.
Sixty serum samples, 30 of which were drawn from patients hospitalized with acute meningitis or meningoencephalitis clinically and/or molecularly diagnosed as being infected with TOS virus (by indirect immunofluorescence assay and PCR) and 30 of which were drawn during the winter season from healthy children (1 to 5 years old) residing in Siena, Italy, were tested for the presence of TOS virus-specific immunoglobulin M (IgM) and IgG by EIA. Nine of 30 positive samples (samples 1 to 9) were collected at the convalescent stage of TOS virus infection, and 21 (samples 10 to 30) were collected at the acute phase of TOS virus infection.
Cells and virus.
Virus antigen was prepared from TOS virus (21)-infected Vero cells. Confluent monolayers of cells were infected with TOS virus at a multiplicity of infection of 1 50% tissue culture infective dose per cell and were cultivated for 4 days in Eagle’s minimal essential medium (Life Technologies, Milan, Italy) supplemented with 5% fetal calf serum (Life Technologies) and penicillin (200 U/ml)-streptomycin (200 μg/ml) (Sigma Co., Milan, Italy) until the appearance of a lytic cytopathic effect on cell culture. The broth cultures were thawed and frozen twice and centrifuged at 4,000 × g for 10 min to remove cell debris. The culture supernatant was then centrifuged at 100,000 × g for 90 min. The supernatant was discarded, and after sonication, the pellet was resuspended in TNE buffer (10 mM Tris-HCl, 0.15 M NaCl, 1 mM EDTA [pH 7.8]) and loaded onto a 20 to 60% sucrose gradient in TNE buffer and centrifuged at 75,000 × g for 3 h. The virus band was diluted in TNE buffer and sedimented at 100,000 × g for 90 min, resuspended in TNE buffer, and stored at −80°C until use. In order to purify the TOS virus N protein, the purified virus was resuspended in TNE buffer containing 1% Nonidet P-40 and the mixture was incubated for 30 min at room temperature. The virus was then stratified on a 20% sucrose cushion and centrifuged at 75,000 × g for 3 h. The pellet containing the N protein was resuspended in TNE buffer.
Expression of N protein.
TOS virus RNA was purified from infected cells by using a previously described method (15). RNA was then subjected to RT-PCR by using the sense primer 5′-GGATCCCATGTCAGACGAGAAT-3′ and the antisense primer 5′-GGATCCTCACTTGCCAACCTT-3′ containing the BamHI site as described elsewhere (22). Plasmid pET15b (Novagen, Milan, Italy) was used as a vector to express the TOS virus N protein. The target gene was cloned in BamHI (12) under control of a T7 promoter and was fused with a polyhistidine tag at the 5′ end (Fig. 1). Expression of the protein was directed by inducible T7 RNA polymerase engineered in E. coli BL21(λDE3). This strain contains a copy of the T7 RNA polymerase gene located in the chromosome under the control of an inducible lacUV5 promoter. Cultures were grown at 37°C in L broth containing ampicillin (100 μg/ml). T7 RNA polymerase was induced by the addition of isopropylthiogalactopyranoside (1 mM) when the culture reached an optical density of 0.6 at 600 nm. Induced cultures were allowed to grow for an additional 2 h at 37°C and were subsequently harvested by centrifugation.
FIG. 1.

Scheme for the genetic construct pSDTV-1. The N-protein gene (≃1,800 bp) of TOS virus is inserted at the BamHI site in pET15b (InVitrogen) under the control of the T7 promoter.
Purification of N protein.
Bacteria were pelleted and resuspended in lysis buffer (50 mM NAH2PO4, 300 mM NaCl [pH 8]) containing lysozyme (1 mg/ml) and antiprotease cocktail (Boehringer Mannheim, Milan, Italy) for 30 min on ice. The cells were then sonicated and centrifuged at 10,000 × g for 15 min. The supernatant was loaded on a His-Bind resin column (Novagen, Milan, Italy) previously equilibrated with 50 mM NaH2PO4 and 300 mM NaCl (pH 8.0). After washing with a buffer (50 mM NaH2PO4, 300 mM NaCl, 10% glycerol [pH 6.0]), the sample was eluted with a gradient of from 0 to 0.5 M imidazole. Samples were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis on 12% polyacrylamide gels. The expressed proteins were detected by immunoblotting.
Immunoblotting.
Purified TOS virus proteins (8 μg), the TOS virus N protein (8 μg), or the recombinant N protein (5 μg) were loaded onto a 12% polyacrylamide gel under reducing conditions, as described previously (11), transferred to a nitrocellulose sheet (19), and tested for the immunoreactivity with a positive and a negative human serum sample at a dilution of 1:200 (Fig. 2).
FIG. 2.
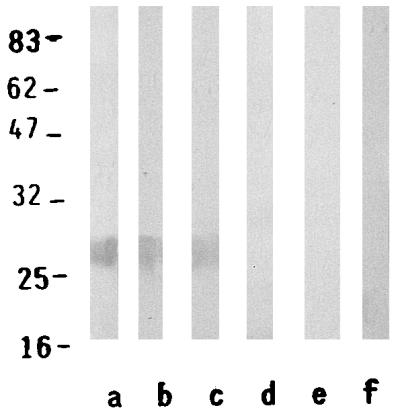
Immunoblot of a human positive serum (lanes a, b, and c) and a negative human serum (lanes e, f, and g) with TOS virus proteins, purified N protein, and recombinant N protein in the sets of lanes, respectively. Low-molecular-mass markers (in base pairs) are indicated on the left (New England Biolabs, Milan, Italy).
Recombinant TOS virus EIA.
Microtiter plates (Labsystem, Helsinki, Finland) were coated with the purified whole virus (1 μg/ml), purified N protein (1 μg/ml), or purified recombinant N protein (1 μg/ml) (100 μl/well) of TOS virus in 0.1 M carbonate buffer (pH 9.6) at room temperature overnight. After washing three times with buffer containing 0.05% Brij, 100 μl of 1:100 dilution of sera in phosphate-buffered saline (PBS) containing 0.05 Brij and 2% fetal calf serum was added to the wells in duplicate and the plates were incubated at 37°C for 45 min. After washing, 100 μl of peroxidase-conjugated anti-human IgG (diluted 1:20,000; Diesse, Siena, Italy) or IgM (diluted 1:6,000; Diesse) was added and the plates were incubated at 37°C for 1 h. Finally, the plates were washed three times and the enzymatic reaction was performed by adding 100 μl of 3,3′,5,5′-tetramethylbenzidine (Sigma) for 15 min. Then, the stop solution 1 N H2SO4 was added and the plates were read at 450 nm. A blank reagent and negative and positive controls were included in each test. For IgM detection sera were preliminarily screened for the presence of the rheumatoid factor, and positive sera were not included in the assay. Positive sera were considered those showing a value >2 standard deviations above the background value represented by negative controls.
RESULTS
Purification of recombinant N protein.
The recombinant N protein was purified by affinity chromatography. The protein was soluble and its yield was about 10 mg/liter of broth culture. The fractions were tested by immunoblot analysis by using human TOS virus-positive and -negative sera. The positive human serum reacted with the recombinant N antigen as well as the purified N protein, as shown in Fig. 2. The same band corresponding to the N protein was evident when the serum reacted with the whole-virus proteins. The control negative serum did not react with any of the TOS virus antigens.
Recombinant protein in EIA.
The antigenic properties of the recombinant N protein were finally evaluated by EIA. The results for all 60 serum samples tested with the purified virus or with the recombinant antigen were similar. The same result was obtained when the samples were tested with the purified TOS virus N protein. Figure 3A and B show the correlation of the reactivities of TOS virus IgM- and IgG-positive sera among the recombinant N protein, the whole-virus antigen, and the purified N protein. Among the positive sera, samples 1 to 9, which were drawn during the convalescent stage of the disease, had higher levels of IgG, while those collected during the acute phase had greater amounts of IgM (Fig. 3).
FIG. 3.

Comparison of the reactivities of the TOS virus IgM-positive sera (A) and IgG-positive sera (B) to the whole virus (□), the purified N protein ( ), and the recombinant N protein (■) by EIA. The results are expressed as the optical densities (OD) at 450 nm for three independent experiments. Samples 1 to 9 were from patients in the convalescent phase of TOS virus infection, and samples 10 to 30 were from patients in the acute phase of TOS virus infection.
DISCUSSION
It has previously been shown that the TOS virus N protein is the major virus antigen recognized by the humoral immune response to TOS virus infection. During acute TOS virus infection, specific IgM and IgG are produced against the N protein. It has been shown that the recombinant N proteins of Hantaan virus and the Puumala virus expressed in E. coli can be used as immunodiagnostic antigens (9, 23).
In the present study we expressed the TOS virus N protein in E. coli (10 mg/liter) and examined the immunological characteristics of this antigen when tested by EIA. The recombinant antigen has an apparent molecular mass of 28 kDa, which agrees with the molecular mass of the native TOS virus protein. The native protein is not glycosylated, and the recombinant expression in a procaryotic system is more likely to generate suitable antigenic conformations. To study the suitability of the recombinant antigen as a diagnostic reagent in a TOS virus antibody assay, 60 serum samples, including positive and negative serum samples, were tested by EIA. The assay appears to be sensitive in discriminating the acute from the convalescent phases of TOS virus infection. In fact, samples 10 to 30 had greater IgM response than samples 1 to 9, which had higher levels of IgG. The same results occurred when the purified N protein was used as antigen in the EIA, and the sera presented reactivities similar to those shown with the whole virus. We have also tested the reactivities of the recombinant N protein in an IgM antibody-capture EIA in which labeled anti-TOS virus antibodies were used. The results were similar (data not shown). Our results demonstrate that the antigenicities in the IgG and IgM EIAs correlate well with that of the whole virus or its nucleocapsid. Furthermore, this assay can be very useful for the diagnosis of TOS virus infection in patients who are negative by RT-PCR performed with cerebrospinal fluid but who have specific TOS virus IgM in their sera. With increased travel to areas of endemicity, particularly the Mediterranean region, imported cases of sandfly fever are increasing. This could cause diagnostic problems because a limited number of laboratories undertake suitable tests for the diagnosis of this infection. Thus, the recombinant protein has a good potential for use in the indirect diagnosis of TOS virus infection.
REFERENCES
- 1.Accardi L, Grò M C, Di Bonito P, Giorgi C. Toscana virus genomic L segment: molecular cloning, strategy and amino acid sequence in comparison with other negative strand RNA viruses. Virus Res. 1993;27:119–131. doi: 10.1016/0168-1702(93)90076-y. [DOI] [PubMed] [Google Scholar]
- 2.Bishop D H. Bunyaviridae and their replication. I. Bunyaviridae. In: Fields B N, Knipe D, Chanock R M, Melnick J L, Hirsh M S, Monath T P, Roizman B, editors. Virology. New York, N.Y: Raven Press; 1990. pp. 1155–1173. [Google Scholar]
- 3.Braito A, Corbisiero S, Marchi B, Sancasciani N, Fiorentini C, Ciufolini M G. Evidence of Toscana virus infections without central nervous system involvement: a serological study. Eur J Epidemiol. 1997;13:761–764. doi: 10.1023/a:1007422103992. [DOI] [PubMed] [Google Scholar]
- 4.Ciufolini, M. G., M. Maroli, and P. Verani. 1991. Laboratory reared sandflies (Diptera: psychodidae) and studies on phleboviruses. Parassitologia 33(Suppl. 1):137–142. [PubMed]
- 5.Clarke D H, Casals J. Techniques for hemagglutination and hemagglutination-inhibition with arthropod-born-viruses. Am J Trop Med Hyg. 1958;7:561–573. doi: 10.4269/ajtmh.1958.7.561. [DOI] [PubMed] [Google Scholar]
- 6.Di Bonito P, Monchi S, Grò M C, Giorgi C. Organization of the genomic segment of Toscana phlebovirus. J Gen Virol. 1997;78:77–81. doi: 10.1099/0022-1317-78-1-77. [DOI] [PubMed] [Google Scholar]
- 7.Ehrnst A, Peters C J, Niklasson B, Svedmyr A, Holmgren B. Neurovirulent Toscana virus (a sandfly fever virus) in a Swedish man after visit to Portugal. Lancet. 1985;i:212. doi: 10.1016/s0140-6736(85)92886-7. . (Letter.) [DOI] [PubMed] [Google Scholar]
- 8.Eitrem R, Niklasson B, Weiland O. Sandfly fever among Swedish tourists. Scand J Infect Dis. 1991;23:451–457. doi: 10.3109/00365549109075093. [DOI] [PubMed] [Google Scholar]
- 9.Gött P, Zöller L, Yang S, Stohwasser R, Bautz E K F, Darai G. Antigenicity of hantavirus nucleocapsid proteins expressed in E. coli. Virus Res. 1991;19:1–16. doi: 10.1016/0168-1702(91)90090-i. [DOI] [PubMed] [Google Scholar]
- 10.Grò M, Di Bonito P, Accardi L, Giorgi C. Analysis of 3′ and 5′ ends of N and NS messanger RNAs of Toscana phlebovirus. Virology. 1992;191:435–438. doi: 10.1016/0042-6822(92)90205-4. [DOI] [PubMed] [Google Scholar]
- 11.Laemmli U K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;270:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- 12.Maniatis T, Fritsch E F, Sambrook J. Molecular cloning: a laboratory manual. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory; 1982. [Google Scholar]
- 13.Nicoletti L, Verani P, Caciolli S, Ciufolini M G, Renzi A, Bartolozzi D, Paci P, Leoncini F, Padovani P, Traini E, Baldereschi M, Balducci M. Central nervous system involvement during infection by Phlebovirus toscanaof residents in natural foci in central Italy (1977–1988) Am J Trop Med Hyg. 1991;45:429–434. doi: 10.4269/ajtmh.1991.45.429. [DOI] [PubMed] [Google Scholar]
- 14.Schwarz T F, Gilch S, Pauli C, Jäger G. Immunoblot detection of antibodies to Toscana virus. J Med Virol. 1996;49:83–86. doi: 10.1002/(SICI)1096-9071(199606)49:2<83::AID-JMV2>3.0.CO;2-F. [DOI] [PubMed] [Google Scholar]
- 15.Schwarz T F, Jäger G, Gilch S, Nitschko H. Nested RT-PCR for detection of sandfly fever virus, serotype Toscana, in clinical specimens, with confirmation by nucleotide sequence analysis. Res Virol. 1995;146:355–362. doi: 10.1016/0923-2516(96)80598-x. [DOI] [PubMed] [Google Scholar]
- 16.Schwarz T F, Jäger G, Glich S, Pauli C. Serosurvey and laboratory diagnosis of imported sandfly fever virus serotype Toscana infection in Germany. Epidemiol Infect. 1995;114:501–510. doi: 10.1017/s0950268800052213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Schwarz T F, Glich S, Jäger G. Travel-related Toscana virus infection. Lancet. 1993;ii:803. doi: 10.1016/0140-6736(93)91568-7. . (Letter.) [DOI] [PubMed] [Google Scholar]
- 18.Tesh R B, Saidi S, Gajdamovic S J, Rodhain F, Vesenjak-Hirijan J. Serological studies on the epidemiology of sandfly fever in the Old World. Bull W H O. 1976;54:663–674. [PMC free article] [PubMed] [Google Scholar]
- 19.Towbin H, Stahelin T, Gordon J. Electrophoretic transfer of protein from polyacrylamide gels to nitrocellulose sheets: procedure and some application. Proc Natl Acad Sci USA. 1979;76:4350–4356. doi: 10.1073/pnas.76.9.4350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Valassina M, Cuppone A M, Bianchi S, Santini L, Cusi M G. Evidence of Toscana virus variants circulating in Tuscany, Italy, during the Summers of 1995 to 1997. J Clin Microbiol. 1998;36:2103–2104. doi: 10.1128/jcm.36.7.2103-2104.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Valassina M, Cusi M G, Valensin P E. Rapid identification of Toscana virus by nested PCR during an outbreak in the Siena area of Italy. J Clin Microbiol. 1996;34:2500–2502. doi: 10.1128/jcm.34.10.2500-2502.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Verani, P., L. Nicoletti, M. G. Ciufolini, and M. Balducci. 1991. Viruses transmitted by sandflies in Italy. Parassitologia 33(Suppl. 1):513–518. [PubMed]
- 23.Zöller L, Yang S, Gött P, Bautz E K F, Darai G. Use of recombinant nucleocapsid proteins of the Hantaan and nephropathia epidemica serotypes of hantaviruses as immunodiagnostic antigens. J Med Virol. 1993;39:200–207. doi: 10.1002/jmv.1890390305. [DOI] [PubMed] [Google Scholar]