

Abstract
Proteases (proteinases or peptidases) are a class of hydrolases that cleave peptide chains in proteins. Endopeptidases are a type of protease that hydrolyze the internal peptide bonds of proteins, forming shorter peptides; exopeptidases hydrolyze the terminal peptide bonds from the C-terminal or N-terminal, forming free amino acids. Microbial proteases are a popular instrument in many industrial applications. In this review, the classification, detection, identification, and sources of microbial proteases are systematically introduced, as well as their applications in food, detergents, waste treatment, and biotechnology processes in the industry fields. In addition, recent studies on techniques used to express heterologous microbial proteases are summarized to describe the process of studying proteases. Finally, future developmental trends for microbial proteases are discussed.
Keywords: protease, classification, detection, expression, application
1. Introduction
As recently highlighted by research and academic papers on enzymes, proteases constitute the largest product segment in the global industrial market for enzymes because they are extensively used in detergent and food industries (Acrofan, 2021; FOC Group, 2022). Additionally, with the development of science and technology, the use of protease enzymes in several bioremediation processes and leather treatments is increasing (Research and Markets, 2021). Moreover, protease enzymes are being extensively used in the production of medicines, as protease enzymes treat multiple diseases, such as lung, heart, eye, digestive tract, and skin ulcer diseases as well as soreness (Shrivastava et al., 2019). Thus, the demand for protease enzymes should continue to increase in the future.
The main sources of proteases are animals (e.g., calf stomach), plants (e.g., pineapple, fig, and papaya), microbes (e.g., Bacillus spp., Pseudomonas spp.; Jisha et al., 2013; Sun et al., 2016, 2019; Chitte and Chaphalkar, 2017). The production of enzymes from animal and plant sources, however, has been limited due to ethical issues, environmental reasons, and low-efficiency production processes. Commercially, microbial enzymes are popular due to their scientific and economic advantages as well as their broad biochemical diversity (Jisha et al., 2013).
In this paper, a detailed studies were reviewed on the classification, identification, testing, application and preparation of microbial protease due to their many advantages, including their rich variety (microbial proteases include acid, neutral, and alkaline proteases); ability to function under various industrial and even extreme conditions (such as high temperatures); and wide application potential and large market in various industry fields, including food, beverage, detergents, leather, animal feed, waste treatment, microbial fermentation and biotechnology industries. In addition, the number of potential proteases is very large (the main bioinformatics databases contains tens of millions of protease genes without functional verification).
2. Classification of microbial proteases
Microbial proteases can be categorized into the following categories: (1) proteases that can hydrolyze specific proteins (e.g., collagenase, elastase, and keratinase); (2) proteases that exhibit likeness to well-characterized proteolytic enzymes (e.g., chymotrypsin, trypsin, and pepsin); (3) proteases with an active pH range (e.g., alkaline, acid, or neutral); (4) proteases that exhibit mechanism of catalytic behavior (i.e., the amino acid residues are involved in the active site or center of the enzymes, such as aspartic proteases, cysteine proteases, metalloproteases, and serine proteases; Rao et al., 1998); and (5) proteases with hydrolysis sites specificity (endopeptidases and exopeptidases, which act internally in polypeptide chains and near the terminus of a polypeptide chain, respectively). The Enzyme Commission (EC) has denoted various endopeptidase and exopeptidase subtypes (see Table 1).
Table 1.
Classification and nomenclature of peptidases.
| Subclasses | EC code | Activity |
|---|---|---|
| Exopeptidases | 3.4.11–19 | Cleave near a terminus of peptides or proteins |
| Aminopeptidases | 3.4.11 | Remove a single amino acid from the free N-terminus |
| Dipeptidases | 3.4.13 | Exopeptidases specific for dipeptides |
| Dipeptidyl peptidases | 3.4.14 | Remove a dipeptide from the free N-terminus |
| Tripeptidyl peptidases | 3.4.14 | Remove a tripeptide from the free N-terminus |
| Peptidyldipeptidases | 3.4.15 | Release of free C-terminus liberates a dipeptide |
| Carboxypeptidases | 3.4.16–18 | Remove a single amino acid from the C-terminus |
| Serine proteases | 3.4.16 | Active sites contain serine |
| Metalloproteases | 3.4.17 | Active sites contain metal ions |
| Cysteine proteases | 3.4.18 | Active sites contain cysteine |
| Omega peptidases | 3.4.19 | Remove terminal residues that are substituted, cyclized or linked by isopeptide bonds |
| Endopeptidases | 3.4.21–24 | Cleave internally in peptides or proteins |
| Serine proteases | 3.4.21 | Active sites contain serine |
| Cysteine proteases | 3.4.22 | Active sites contain cysteine |
| Aspartic proteases | 3.4.23 | Active sites contain aspartate |
| Metalloproteases | 3.4.24 | Active sites contain metal ions |
| Threonine endopeptidases | 3.4.25 | Active sites contain threonine |
| Endopeptidases of unknown catalytic mechanism | 3.4.99 | Acting on peptide bonds |
Proteases are categorized in subgroup 4 of group 3 (hydrolase), per the Nomenclature Committee of the International Union of Biochemistry and Molecular Biology (International Union of Biochemistry, 1992).
A detailed system of classification has resulted from increased knowledge on the catalytic mechanism and structure. Depending on the evolutionary relationships and amino acid sequences of proteases, they are categorized into different clans and families (Rawlings et al., 2017). A clan (i.e., a group of families) does not exhibit significant similarities in sequence but does possess an evolutionary relationship. Clans can also include families from different catalytic classes because their catalytic-site residues follow an identical order and show similar tertiary folds. A family contains proteolytic enzymes that are homologous, which is revealed by a significant similarity in their amino acid sequence. They can be identified according to the family’s enzyme type or a homologous protein to the enzyme type, which thus is a family member. Based on this classification, the MEROPS database provides comprehensive details about different proteases. According to these phylogenetic relationships and mechanisms of action, all proteases in clans and families can be grouped into asparagine proteases, aspartic proteases, cysteine proteases, glutamic proteases, mixed proteases, metalloproteases, threonine proteases, serine proteases, and unknown proteases (Rawlings et al., 2017).
3. Detection of microbial proteases
3.1. Endopeptidase detection
3.1.1. Observation of halos
Protease production is indicated by the formation of clear halos around colonies that have grown on protein substrates in agar plates. This occurs when extracellular endopeptidases are produced by microorganisms in solid media. Growth media supplement the protein substrates, which were then poured into Petri plates. Commonly used substrates include skim milk agar (Masi et al., 2021; Shaikh et al., 2023), casein agar (Yokota et al., 1988; Rathod and Pathak, 2014), bovine serum albumin (BSA) agar (De Azeredo et al., 2001), gelatin agar (Mortezaei et al., 2021), keratin agar (Pereira et al., 2014; Nnolim et al., 2020), fibrin agar (Prabhu et al., 2021; Anis Ahamed et al., 2022), and elastin agar (Zins et al., 2001). When protease was produced in liquid media, the supernatant of fermentation broth (for extracellular proteases) or cell lysate (intracellular proteases) containing protease was collected. The same agar plates (containing protein substrates) as described for solid media were prepared, a well was created in the plate was made or an Oxford cup was placed on the plate for the enzyme liquid container to observe the halo (Wang et al., 2015; Yang et al., 2021).
The observation halo is the most intuitive and simple method used to identify proteases, but it is only suitable for endopeptidases and proteases that exhibit sufficiently strong activity to form clear halos. The activity of proteases is commonly detected by measuring the hydrolysate or the reduction in substrate caused by protease hydrolysis. There are many kinds of proteases that exhibit different activities, utilize different hydrolysis modes, and generate hydrolysis products with different characteristics; thus, different substrates and methods are needed to detect these proteases. To date, the substrates used to detect proteases are roughly divided into native substrates and modified substrates. The modified substrates are further mainly divided into chromogenic substrates and fluorescent substrates. Different substrates are detected with different methods.
3.1.2. Detection by natural protein substrates
Natural protein substrates are those that occur in nature (plant protein, animal protein, microbial protein, etc.). The most commonly used substrate for testing protease activity is casein. Protease hydrolyzes casein under certain temperature and pH conditions to produce peptides or amino acids that are soluble in an acidic solution. After undergoing acid deposition, the newly formed product dissolves in the upper acid solution, while the unhydrolyzed protein forms a precipitate (Yokota et al., 1988; Rathod and Pathak, 2014). The supernatant is collected by centrifugation, and the activity of the protease is determined by testing the resulting peptides or amino acids using Folin reagent, ninhydrin, TNBS or OPA, which each exhibit advantages and disadvantages (Table 2).
Table 2.
Detection of protease activity using a natural substrate.
| Test method/reagent | Detection principle | Advantages and disadvantages | References |
|---|---|---|---|
| Folin reagent | Proteases hydrolyze protein substrates to produces amino acids with phenolic groups (tyrosine, tryptophan) or peptides containing amino acids with phenolic groups, which can be reduced by the Folin reagent (Folin) under alkaline conditions to produce molybdenum blue and tungsten blue, the color of which is proportional to the content of amino acids with phenolic groups. The number of amino acids with phenolic groups produced by enzymatic digestion is obtained by detecting the absorbance at 680 nm, and thus calculating the protease activity | This method is easy to operate and the quantitative range is 5–100 μg amino acids; the color reaction of Folin reagent is caused by tyrosine, tryptophan and cysteine, so if the sample contains phenols, citric acid and sulfhydryl compounds, they will interfere with the detection; This method is affected by the type of protein substrate, and the color intensity of different proteins is slightly different due to the different content of tyrosine and tryptophan | Mcdonald and Chen (1965), Chen et al. (2022) |
| Ninhydrin | Amino acids and peptides with free α-amino and α-carboxyl groups react with ninhydrin to produce a blue–purple substance (proline and hydroxyproline react with ninhydrin to produce a (bright) yellow substance). The color shade of this compound is proportional to the amino acid content and the amino acid content is determined by measuring the absorbance at 570 nm (440 nm for proline and hydroxyproline) | A commonly used and sensitive method for the detection of proteases with a detection limit of 0.5 μg amino acids; however, the color developer has low stability and cannot be stored for a long time; different amino acids and ninhydrin develop color differently, resulting in partial deviation of the measurement results | Moore and Stein (1948), Zhang et al. (2013), Hamed et al. (2020) |
| Trinitrobenzene sulfonate (TNBS) | TNBS reacted with amino acids under alkaline conditions for 1 h at 37°C and cooled at room temperature for 30 min, followed by the detection of absorbance values at 420 nm, which were proportional to the amino acid content | The sensitivity of this method is reasonable, and the detection range is 0.05–0.4 μmol amino acids; the shortcomings are that the assay is time-consuming, the ε-amino group of leucine can also react with TNBS, which affects the accuracy of the determination results, and the lack of correlation between the proline and hydroxyproline contents and the absorbance values, which can easily produce bias | Adler-Nissen (1979), Spellman et al. (2003), Fathi et al. (2021), Han et al. (2021), Cermeño et al. (2022) |
| o-phthaldialdehyde (OPA) | Proteases release a free amino group for each peptide bond hydrolyzed. The free amino group reacts with OPA to form a yellow complex, the absorbance of which can be measured spectrophotometrically at 340 nm | The OPA method is a rapid and simple method to determine the hydrolysis of protein hydrolysates, and is 5–10 times more sensitive than the ninhydrin method; OPA determination of hydrolysis relies on a weak and unstable reaction between OPA and cysteine in the hydrolyzed substrate; this method is not suitable for cysteine-rich substrates, and proline and OPA do not react and cannot be detected; in addition, the detection process requires strict time control, because the detection value changes with time, and it is relatively difficult to obtain accurate measurement values | Church et al. (1983), Spellman et al. (2003), Hong et al. (2022), Alblooshi et al. (2023) |
3.1.3. Detection by modified protein substrates
3.1.3.1. Detection by chromogenic substrates
To increase substrate solubility and detection sensitivity, modified protein substrate is used in some methods to detect protease activity, and this substrate should generate a colored end product after proteolysis or a product that can be converted into a colored complex. One example is azocasein, a casein dyed with p-aminobenzenesulfonic acid, which produces a colored complex that is soluble in trichloroacetic acid and shows absorption at 440 nm after digestion by proteases (Cejudo-Bastante et al., 2022; de Matos et al., 2022; Marson et al., 2022). Succinyl casein, which possesses chemically succinlyated amino groups (Hatakeyama et al., 1992), easily dissolves at pH values greater than 4, unlike casein.
According to substrate specificity, the synthetic substrate can be identified by the type of protease screened, such as Tosyl-Gly-Pro-Arg-pNA for trypsin (Sandholt et al., 2018), Suc-Ala-Ala-Pro-Phe-pNA for chymotrypsin (Siigur et al., 2011; Németh et al., 2022) and Suc-Ala-Ala-Pro-Val-pNA for elastase (Ferreira et al., 2009). However, N-Cbz-Ala-Ala-Leu-pNA and N-Cbz-Gly-Gly-Leu-pNA are good substrates for subtilisins (Burchacka et al., 2022). The principle underlying the assay is that proteases hydrolyze the amide bond connecting p-nitroaniline (pNA) to the neighboring amino acid residue, and released pNA exhibits specific absorption at a 405 nm wavelength (enzyme activity is proportional to fluorescence intensity).
3.1.3.2. Detection by fluorogenic substrates
More sensitive methods are needed when the quantity or activity of protease enzymes are low, and sensitive fluorescent peptide substrates are available, through which the limit of detection reaches the ng level (Austin et al., 2022).
Fluorescent labeling applied to protease substrate modification can be divided into the following categories: 1, single fluorescence-based labeling, in which one kind of fluorescent dye labels the substrate protein after binding so that the substrate protein obtains fluorescent labeling. 2, Double fluorescence labeling, in which two different fluorescent dyes label the peptides. One dye is an energy acceptor and the other is an energy donor; the labeled peptide, which is activated by protease hydrolysis, does not show fluorescence. 3, Homotransfer fluorescence labeling, in which there is one kind of fluorescence labeling substrate protein, and fluorescence resonance energy transfer (FRET) occurs between the labeled fluorescent molecules, which do not show fluorescence but are hydrolyzed by proteases to activate fluorescence.
3.1.3.2.1. Single fluorescence-based labeling
Single fluorescence dye labeling involves introducing fluorophores attached to side chain amino acids, such as the N-terminus, C-terminus, Glu, Lys or Cys of a peptide. Nearly 30 types of fluorescence dyes have been developed thus far (Díaz-García and Badía-Laíño, 2018). The more widely used dyes are carboxyfluorescein (FAM; Feng et al., 2022), fluorescein isothiocyanate (FITC; Taylor et al., 2022), dansyl chloride (DNS-Cl; Yoo and Han, 2021), 2,4-dinitrophenylhydrazine (Dnp; Oliveira et al., 2001), 7-amino-4-methylcoumarin (AMC), 7-amino-4-trifluoromethyl coumarin (AFC; Breidenbach et al., 2020), carboxyrhodamine 110 (CR110; Lorey, 2002), Texas Red (Lorey, 2002), pentamethine cyanine (Cy5) and heptamethine cyanine (Cy7) dyes (Chin and Kim, 2018). Protease activity is measured as an enhanced emission generated after a peptide is cleaved by an enzyme and is released from the fluorophore. The detection limits of single fluorescence-based labeling for proteases can reach the ng level (Kasana et al., 2011). However, when detection is performed using a single fluorescently labeled protease substrate, the product and substrate must be separated, and the pH needs to be adjusted to enhance the detection signal. The detection steps remain relatively complex (Twining, 1984; Austin et al., 2022).
3.1.3.2.2. Fluorescence dye double labeling
In contrast to single fluorescence-based labeling, such as the commonly used FTC-casein assay, double fluorescence labeling provides a more convenient and precise method, which is based on the FRET concept (Clapp et al., 2004; Goulet et al., 2020). The kinetics of exo- and endopeptidases can be measured over a wide pH range using assay procedures that do not involve separation steps (Legare et al., 2022). The total substrate turnover can be measured at a fixed time after an enzyme is added (Elston et al., 2007). Decreased fluorescence quenching (i.e., increased total fluorescence), which occurs as peptides (labeled proteins) are digested into smaller fluorescein-labeled fragments, can be identified using FRET-based measurement. In classical FRET, electron energy transfer occurs between two fluorophores, an energy acceptor and energy donor. Table 3 lists common combinations of acceptors and donors.
Table 3.
Double fluorescence labeled donor-acceptor pair.
| Acceptors | Donors | Wavelength (nm) | |
|---|---|---|---|
| Excitation | Emission | ||
| Dnp (2,4-Dinitrophenyl) | Trp (Tryptophan) | 280 nm | 360 nm |
| 4-Nitro-Z (4-Nitro-benzyloxycarbonyl) | Trp (Tryptophan) | 280 nm | 360 nm |
| Dnp (2,4-Dinitrophenyl) | Mca (7-Methoxycoumarin-4-acetyl) | 325 nm | 392 nm |
| pNA (para-Nitroaniline) | Abz (2-Aminobenzoyl) | 320 nm | 420 nm |
| 3-Nitro-Tyr (3-Nitro-tyrosine) | Abz (2-Aminobenzoyl) | 320 nm | 420 nm |
| 4-Nitro-Phe (4-Nitro-phenylalanine) | Abz (2-Aminobenzoyl) | 320 nm | 420 nm |
| Dabcyl ((4-(4-Dimethylamino)phenyl)azo)benzoyl | EDANS (5-[(2-Aminoethyl)ami-no]-1-naphthalenesulfo-nic acid) | 340 nm | 490 nm |
| Dabsyl (4-(4-Diethylaminophenylazo)-benzenesulfonyl) | Lucifer Yellow | 430 nm | 520 nm |
| Dnp (2,4-Dinitrophenyl) | FITC (Fluorescein isothiocyanate) | 490 nm | 520 nm |
| 4-Nitro-Phe (4-Nitro-phenylalanine) | Dansyl (5-(Dimethylamino) naphthalene-1-sulfonyl) | 342 nm | 562 nm |
| QSY7 | 5-TAMRA (Carboxytetramethyl rhodamine) | 547 nm | 573 nm |
| QSY-7 | Eu (III) Chelate | 340 nm | 613 nm |
3.1.3.2.3. Homotransfer fluorescence labeling
As mentioned above, classical FRET involves electron energy transfer between two different fluorophores; however, FRET events can also occur as a result of fluorescence homotransfer in which fluorescein acts both as the energy “donor” and energy “acceptor” (Runnels and Scarlata, 1995; Chen et al., 2000; Thompson et al., 2000; Figure 1), which is called homotransfer fluorescence labeling. Compared to the FTC-casein assay, these assays are also easier to perform, and they are 100-fold more sensitive (Jones et al., 1997).
Figure 1.
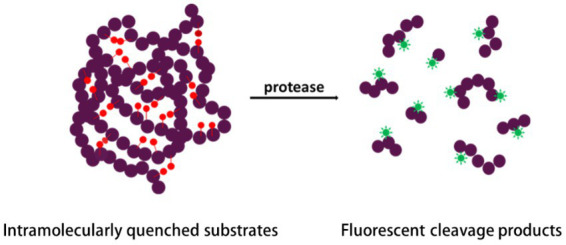
Principle of protease detection by fluorescence homotransfer.
A typical single-fluorescence dye used in FRET is the BODIPY dye (Jones et al., 1997): BODIPY dye molecules are attached to casein to prepare casein conjugates of BODIPY dyes. The dyes in these conjugates are labeled to achieve efficient quenching in the protein. This process yields nonfluorescent substrate molecules. These fluorogenic substrates release highly fluorescent BODIPY dye-labeled peptides during proteolysis and increase the fluorescence as it relates to enzymatic activity. Using standard fluorometers, filter fluorometers, or fluorescence microplate readers, this activity can be measured. Fluorescein excitation and emission wavelengths can be used to measure BODIPY casein hydrolysis. EnzChek™ Protease Assay kits from ThermoFisher Scientific contain a heavily labeled casein derivative. Green-fluorescent BODIPY FL dye and red-fluorescent BODIPY TR-X dye are commonly used for this application.
3.2. Exopeptidase detection
Endopeptidases hydrolyze proteins and mainly release peptides, and exopeptidases hydrolyze proteins and release free amino acids, so the methods used to detect endo−/exopeptidases must be different, and the method used to detect endopeptidases is not very sensitive to exopeptidases.
The assay used to measure exopeptidase activity usually involves synthetic peptide as the substrate; for the aminopeptidase assay, a peptide with two and three amino acid residues is synthesized to detect aminopeptidases (Mathew et al., 2000; Gu and Walling, 2002). For more sensitive detection, p-nitroaniline (pNA; Cahan et al., 2001; Schulze et al., 2018) or 7-methoxycoumarin-4-acetic acid (MCA; Chen et al., 2011, 2012; Schulze et al., 2018) are connected to the carboxyl terminus of peptides; after hydrolysis, a free pNA or MCA molecule is released in the reaction solution. This method can detect the specific absorbance value to determine the aminopeptidases activity.
For the carboxypeptidase assay, a peptide with an amino terminus blocked by benzyloxycarbonyl (CBZ; Fu et al., 2011; Song et al., 2021a) or benzoyl (BZ; Ramirez Zavala et al., 2004; Heylen et al., 2010) is most commonly used as a substrate. Only carboxypeptidase can release amino acids from the carboxyl terminus. After hydrolysis, free amino acids are released from the synthetic peptide and detected by ninhydrin or OPA reagent.
Fluorescent substrates can also be used to detect exopeptidases, including aminopeptidases (Chen, 2020; Liu S. Y. et al., 2022; Ma et al., 2023) and carboxypeptidases (Xiong et al., 2018; Yoo and Han, 2021), because of their extreme sensitivity.
Some protease detection methods, such as ELISAs or ultrasonic resolver technology assays, are also available. These methods are not widely used due to their limitations and are only used in special cases. For example, prior information on the structure of the enzyme is needed to perform ELISA-based assays (Blair and McDowell, 1995). For ultrasonic resolver technology, a different analytical method is needed and must be first correlated to the corresponding ultrasonic velocity signals in advance (Born et al., 2010). These methods will not be introduced in detail here. For details, please refer to related reviews (Kasana et al., 2011).
Among microbial resources, potential proteases are extremely abundant, and proteases detection methods are crucial for developing novel proteases. In the future, detection methods will be developed that are sensitive, fast, inexpensive, and suitable for high-throughput screening of proteases.
4. Application of microbial proteases
Microbial proteases have wide ranging applications in several fields, including baking, brewing, detergents, leather making, pharmaceuticals, meat tenderizing, cosmetics, medical diagnosis and so on (Christensen et al., 2022; Reddy et al., 2022; Akram et al., 2023; Mubeen et al., 2023). In addition, with the rapid development of new fields, applications of microbial proteases are expanding to new areas, such feed industries (Bernardeau et al., 2022; Cupi et al., 2022), hydrolysis applications to prepare active peptides (Christensen et al., 2022), and environmental protection applications, such as waste treatment and reuse (Ariaeenejad et al., 2022; Asitok et al., 2022; Zhai et al., 2022). These applications illustrate the diversity and importance of proteases. The applications of proteases and their respective microbial sources by examining acid protease, neutral protease and alkaline proteases and their classification were discussed and briefly summarized in Figure 2.
Figure 2.

The applications of proteases.
4.1. Alkaline proteases
Among the different proteases, alkaline proteases exhibit the highest activity in the pH range of 8 to 13. Alkaline proteases are commonly used in the following industries:
4.1.1. Detergent industry
Alkaline proteases represent the largest share of the enzyme market, are a commercially important group of enzymes and are used primarily as detergent additives (Sharma et al., 2017). By adding alkaline proteases to laundry detergents, proteinaceous material can be released from stains (Matkawala et al., 2019; Tanwar et al., 2022). Unlike traditional detergents, the addition of protease saves energy and improves washing efficiency. After soaking, shorter periods of agitation and lower wash temperatures can be used with the addition of proteases (Mubeen et al., 2023). Commercial alkaline proteases are effective at low levels (0.4–0.8%) and are compatible with various detergent components that contain oxidizing and sequestering agents. These proteases also exhibit high activity and stability over a broad range of pH values and temperatures as well as a long shelf life (Vojcic et al., 2015). Proteases are environmentally friendly, nonphosphate detergents, and washing powders containing proteases can be used in dry cleaning applications as stain and spot removers (Kumar et al., 2016).
4.1.2. Leather industry
Leathers are usually processed using an alkaline reagent. Because alkaline proteases exhibit keratinolytic and elastolytic activities, they can effectively biotreat leather, particularly the bating and dehairing of hides and skins (Tian et al., 2019; Srivastava et al., 2020). These methods are better choices than conventional methods, which use harsh chemicals, create disposal problems, exhibit increase safety risks, and cause chemical pollution (Hassan et al., 2020). Subsequent studies have successfully used alkaline proteases from Aspergillus, Streptomyces, and Bacillus in leather tanning (Ogino et al., 2008; Paul et al., 2016; El-Ghonemy and Ali, 2021; Hasan et al., 2022; Zhang et al., 2022).
4.1.3. Food industry
The most extensive application of alkaline protease is in the food industry.
4.1.3.1. Meat tenderization
Alkaline proteases can hydrolyze muscle fiber proteins and connective tissue proteins. Meat tenderization is achieved by immersing meat in a protease solution or sprinkling it with a powdered enzyme (Bureros et al., 2020). The vascular systems of animals are often injected with protease solutions 10–30 min before slaughter (Kalisz, 1988), including alkaline elastase (Qihe et al., 2006) and thermophilic alkaline protease (Wilson et al., 1992).
4.1.3.2. Meat solubilization
Soluble meat hydrolysates and meat-flavored hydrolysates are byproducts of the leather industry. These potential sources of protein are bone, offal (raw lung), and bone residues after mechanical deboning. The most beneficial enzyme in terms of solubilization, cost, and other factors is alcalase (Anzani et al., 2017), which can be used to produce fish protein hydrolysates (Noman et al., 2022).
4.1.3.3. Blood decolorization
Because of its intense color, blood is an underutilized source of food protein. Although the red cell fraction contains 75% of the protein in the blood, alcalase is preferred because it thoroughly and rapidly hydrolyzes red cells.
The red cell fraction contains 75% of the protein in the blood, of which more than 92% is hemoglobin. Hemoglobin is composed of heme and globin, and heme causes blood products to eventually appear black red and exhibits a strong bloody smell. Alcalase is the preferred blood decolorization protease because it thoroughly and rapidly hydrolyzes hemoglobin and releases polypeptides. After enzymatic cleavage, the remaining hydrophobic core formed by wrapping heme with hydrophobic peptide fragments forms precipitates under appropriate pH conditions. The supernatant is dried by spray to produce hemoglobin powder, which can remove the ugly black purple color and the bloody smell of blood products; furthermore, the powder can be used as feed additive, colorants in the food industry and pharmaceutical raw materials in the pharmaceutical industry (Pérez-Gálvez et al., 2011).
4.1.3.4. Soy products
In Asia, fungal proteases have long been used to prepare soy sauce and soy products (Devanthi and Gkatzionis, 2019). The alkaline and neutral proteases of Aspergillus are essential in the digestion of soybean protein and provide the rich flavor of true soy sauce (Zhao et al., 2020). They also play an important role in improving the quality of soy products during processing (Xu et al., 2013).
Members of the genus Bacillus have been screened for use in various industrial applications and have been identified as the predominant alkalophilic microorganism. They are a prolific source of alkaline proteases, including Bacillus amyloliquefaciens, Bacillus licheniformis, and some Bacillus sp. Many fungi produce extracellular alkaline proteases, most notably Aspergillus sp. (Table 4).
Table 4.
Representative alkaline proteases originated from microbial sources.
| Microbial sources | Proteases | References |
|---|---|---|
| Bacillus amyloliquefaciens | An extracellular alkaline protease | Chen W. et al. (2022) |
| An alkaline proteases AprM | Xie et al. (2022) | |
| An alkaline serine-protease APR68 | Cho (2019) | |
| Bacillus licheniformis | An alkaline protease AprE | Zhou et al. (2021) |
| A detergent stable thermophilic alkaline protease | Emran et al. (2020) | |
| Asubtilisin protease | Cupi et al. (2022) | |
| Bacillus sp. | An Alkaline protease with special reference to contact lens cleansing | Rejisha and Murugan (2021) |
| A novel alkaline serine protease | Ariyaei et al. (2019) | |
| A H2O2-tolerant alkaline protease | Yu et al. (2019) | |
| Deep-sea fungi | An alkaline and cold-tolerant proteases | Damare et al. (2006) |
| Aspergillus oryzae, Penicillium roquefortii and Aspergillus flavipes | Several alkaline proteases | Novelli et al. (2016) |
| Aspergillus sp. | An activator-dependent protease | Zhu X. et al. (2021) |
| An alkaline protease | Salihi et al. (2017) | |
| An alkaline serine protease | Yadav et al. (2015) | |
| An extracellular keratinolytic protease | Anitha and Palanivelu (2013) | |
| A solvent, salt and alkali-tolerant alkaline protease | Abdel Wahab and Ahmed (2018) |
4.2. Neutral proteases
Neutral proteases exhibit the highest activity at neutral, weakly alkaline or weakly acidic pH values. Neutral proteases are used in the following applications.
4.2.1. Bakers’ dough
To help in bread production, neutral proteases and amylases can be added to wheat or flour. Protease increases bread volume, improves dough elasticity, and improves crust texture (Zadeike et al., 2018; Gu et al., 2022; Xu et al., 2022; Li J. et al., 2023; Sun et al., 2023). In the process of making crackers, biscuits, and cookies, neutral proteases are used to improve the extensibility and strength of the dough and prevent dough from tearing when rolled thin. To prevent biscuits from bending and wrinkling in the oven, the dough must be soft (Borrelli et al., 2003; Sumantha et al., 2006; Mokashe et al., 2018; Nikinmaa et al., 2019). A soft and pliable dough is also necessary for the precise letters and decoration on biscuits. Bacterial neutral proteases are often used to achieve this (Ehren et al., 2009) because the enzymes’ highly specific endopeptidases are ideal for high protein flours.
4.2.2. Meat tenderization
Fresh meat pH is neutral, and therefore neutral proteases are best suited for hydrolysis; tenderization of meat is achieved by the action of endogenous proteases, especially neutral lysosomal cathepsins and neutral metalloprotease/cysteine endopeptidase (Prates et al., 2001; Thomas et al., 2004; Mikołajczak et al., 2019).
Neutral proteases are widely distributed among the Bacillus and Aspergillus species (Ward et al., 2009). Thermolysin [EC 3.4.24.27], which is produced by Bacillus thermoproteolyticus, is probably the best-known neutral protease (Inouye et al., 2007). Thermolysin was originally identified in the culture broth of Bacillus thermoproteolyticus Rokko and is an attractive target in protein engineering. Since its discovery in 1962, Thermolysin, which is a thermostable neutral zinc metalloprotease, has undergone extensive structural and mechanistic studies due to its halophilicity, catalytic mechanism, and thermostability. The Bacillus genera that produce neutral proteases include Bacillus subtilis, Bacillus licheniformis, Bacillus stearothermophilus, Bacillus nakamurai, and Bacillus tropicus, and the Aspergillus genera include Aspergillus oryzae, Aspergillus niger, Aspergillus sojae, Aspergillus nidulans, and Aspergillus tamarii (Table 5).
Table 5.
Representative neutral proteases originated from microbial sources.
| Microbial sources | Proteases | References |
|---|---|---|
| Bacillus subtilis | A neutral protease | Albillos et al. (2007) |
| Bacillus licheniformis | Neutral proteases | Reddy et al. (2022) |
| Bacillus stearothermophilus | A neutral protease | Mansfeld et al. (2005) |
| A neutral protease | Eijsink et al. (1991) | |
| Bacillus nakamurai | An Extracellular protease | Shaikh et al. (2023) |
| Bacillus tropicus | Keratinolytic proteases | Liya et al. (2023) |
| Aspergillus oryzae | A thermolysin-like protease, neutral protease I | Ma et al. (2016) |
| Neutral proteases | Belmessikh et al. (2013) | |
| Aspergillus niger | Neutral proteases | Reddy et al. (2022) |
| Aspergillus sojae | Neutral proteases | Li et al. (2023) |
| Aspergillus nidulans | Neutral proteases | Campbell and Peberdy (1983) |
| Aspergillus tamarii | Neutral proteases | Silva et al. (2018) |
4.3. Acid proteases
The proteases described here are active between pH 2 and 6. Acid proteases of microbial origin are mostly found in the food and beverage industries.
4.3.1. Food industry
Acid proteases are primarily used in the food industry for the clotting of milk during the manufacturing of cheese. When the milk proteins coagulate, they form solid masses or curds. Then, the whey is removed to generate cheese (Tsuchiya et al., 1993; Hellmuth, 2006; Theron and Divol, 2014). In addition to their application in the dairy industry, acid proteases are also used for baking. Similar to neutral proteases, acid proteases from Aspergillus oryzae can limit the proteolysis of wheat gluten and increase loaf volume. Fungal-derived acid proteases have also been extensively applied to create food seasonings and improve protein-rich foods (e.g., bread and related foodstuffs; Hamada et al., 2013; Purushothaman et al., 2019; Wu et al., 2022; Li X. et al., 2023; Niu et al., 2023).
4.3.2. Beverage industry
Acid proteases can degrade proteins in fruit juices and certain alcoholic beverages that cause turbidity (Espejo, 2021; Pati and Samantaray, 2022; Rasaq et al., 2023), including black currant (Landbo et al., 2006); cherry (Pinelo et al., 2010); pomegranate (Cerreti et al., 2017); and apple, orange, grape, and kiwi fruit juices (Guo et al., 2019). By adding acid proteases, the immediate turbidity is significantly reduced. Adding proline-specific proteases from Aspergillus niger (Lopez and Edens, 2005) or Aspergillus oryzae (Kang et al., 2014) when brewing beer can prevent chill-haze formation. This result indicates that proline-rich proteins perform hydrolysis due to a peptide fraction that cannot interact with polyphenols. Protein haze is also a problem that occurs during the production of white wine. Early research has found that by using acid proteases in wine, protein haze formation can be reduced without damaging wine quality (Marangon et al., 2012; Van Sluyter et al., 2013; Theron et al., 2018). Apart from preventing protein haze, acid proteases also increase the α-amino nitrogen concentration necessary for microbial growth and generate better flavor during beer brewing (Bell and Henschke, 2005; Lei et al., 2013; Wang et al., 2013; Serna-Saldivar and Rubio-Flores, 2017).
Acid proteases are mainly aspartic proteases and are distributed across all forms of life, including vertebrates, plants, fungi, bacteria and viruses (Theron and Divol, 2014). However, fungus-derived acid proteases, such as Aspergillopepsins I and II from Aspergillus niger are most commonly used in the food and beverage industries (Ichishima, 2004; Takahashi, 2004). They are the first and most commonly used acid proteases in the food industry. Recent reports on fungi-derived acid proteases have been used for various purposes, and the proteases mainly originate from Aspergillus oryzae, Aspergillus niger, Aspergillus foetidus, Aspergillus saitoi, Aspergillus clavatus, Rhizomucor miehei, Mucor miehei, and Rhizopus rhizopodiformis (Table 6).
Table 6.
Representative acid proteases originated from microbial sources.
| Microbial sources | Proteases | References |
|---|---|---|
| Aspergillus oryzae | An aspartate protease | Vishwanatha et al. (2009) |
| An acid protease as starter culture in doubanjiang fermentation | Niu et al. (2023) | |
| A valuable food acid protease | Murthy and Kusumoto (2015) | |
| An acid protease | de Castro and Sato (2014) | |
| An aspartate protease | Shu et al. (2020) | |
| A salt-tolerant acid protease | Lee et al. (2010) | |
| Aspergillus niger | Acid proteinase A | Takahashi et al. (1998) |
| Extracellular acid proteases | Aalbæk et al. (2002) | |
| Acid proteinases | Yang and Lin (1998) | |
| Aspartic proteases | Purushothaman et al. (2019) | |
| Acid proteinases | Li et al. (2020) | |
| Aspergillus foetidus | An aspartic protease | Souza et al. (2017) |
| A thermostable extracellular acid protease | Souza et al. (2015) | |
| Aspergillus saitoi | An acid carboxypeptidase | Chiba et al. (1993) |
| Aspergillus clavatus | An extracellular acid protease | Sampaio Silva et al. (2011) |
| Rhizomucor miehei | Acid proteinases | Aljammas et al. (2022a,b) |
| Rennet | Celebi et al. (2016) | |
| Rennet | Soltani et al. (2019) | |
| Mucor miehei | Acid proteases | Ayhan et al. (2001) |
| Acid proteases | Escobar and Barnett (1995) | |
| Rennet | Seker et al. (1999) | |
| Rhizopus rhizopodiformis | Extracellular acid proteases | Schindler et al. (1983), Sun et al. (2014) |
4.4. Expanding specific applications
Protease applications are still expanding as specific applications develop, and new areas of interest in recent years are described in the following section.
4.4.1. Protein hydrolysates that improve flavors and decrease bitterness
Due to their amino acid sequence and length, oligopeptides exhibit different flavors, including sweet, bitter, umami, sour, or salty taste. Twenty common amino acids also present different flavors, such as umami, sweetness and bitterness; glutamic acid presents an umami flavor; arginine, proline, leucine, isoleucine, phenylalanine, and tryptophan present a bitter taste for humans; and L-alanine and L-serine provide a sweet taste. Proteases (mainly endopeptidases) can hydrolyze proteins to produce oligopeptides, thus enhancing the flavor of protein-based food (Wang H. et al., 2022; Yan et al., 2022), and exopeptidases (mainly aminopeptidases and carboxypeptidases) can hydrolyze peptides to produce free amino acids, also enhancing (enriching) the flavor of protein-based food (Cheung et al., 2015; Fu et al., 2020; Ding et al., 2022). For example, Alcalase and Flavorzymes were used to prepare defatted flaxseed meal protein hydrolysates (Wei et al., 2018). After processing optimization, peptides with molecular weights above 1,000 Da enhanced the texture of food, while peptides with molecular weights ranging from 128 to 1,000 Da provided meat-like flavors and influenced other sensory features.
Aminopeptidases from Lactobacillus casei, Lactobacillus curvatus, and Lactobacillus sake were used to improve the sensory quality of dry fermented sausages (Nandan and Nampoothiri, 2020). Neutrase, which is a neutral bacterial protease, can modify flavor in dairy applications (Sumantha et al., 2006). During the fermenting of fish sauce, taste formation is affected by protease activity because it alters the content of Ala, Asp, Glu, Leu, Lys, TCA-soluble peptides, and succinic acid (Zhu W. et al., 2021).
Bitterness is inevitably produced when oligopeptides undergo protein hydrolysis, and the intensity of bitterness of hydrolysis products is mainly related to the content and position of hydrophobic amino acids (or more accurately, amino acid residues with Q values above 1,500 cal/mol, such as Leu, Ile, Phe, Tyr, Trp, Pro, Val, and Lys (Nishiwaki et al., 2002) in the peptide segment). Matoba and Hata (1972) described in detail that the bitterness of protein hydrolysate is great when hydrophobic amino acids are internal in the oligopeptides, the bitterness is comparatively weaker when the hydrophobic amino acid(s) are located at either the N- or C-terminus and the weakest occurs when the hydrophobic amino acids are in the free state. Therefore, specific endopeptidases and exopeptidases can reduce the bitterness of protein hydrolysate (FitzGerald and O'Cuinn, 2006; Soeryapranata et al., 2007). Endopeptidases can hydrolyze the hydrophobic amino acids forming bonds of oligopeptides and reduce bitterness (Capiralla et al., 2002; Edens et al., 2005; Zhang M. et al., 2021). Aminopeptidases (Lin et al., 2020; Nandan and Nampoothiri, 2020; Song et al., 2020a; Nakamura et al., 2023; Wang et al., 2023) and carboxypeptidase (Ding et al., 2022) from many different sources can continue to hydrolyze end hydrophobic amino acids and then reduce bitterness.
4.4.2. Protein modification
Microbial proteases are used to modify proteins. Protease-limited enzymatic hydrolysis of soybean protein can improve its solubility, emulsification, foaming and digestibility. Hydrolysis of peanut protein concentrates with Aspergillus oryzae crude protease extract resulted in their higher water- and oil-binding capacity as well as improved solubility, foam stability, and foaming capacity (Yadav et al., 2022). When soybean protein isolate (SPI) was treated with alkaline protease accompanied by high-speed shearing homogenization, it significantly improved the emulsion stability of the SPI hydrolysates. As a result, the foaming properties of SPI were improved significantly (Hao et al., 2022). Recent studies have examined methods to use microbial proteases from a variety of sources to improve the chemical and physical properties of animal (Ai et al., 2019; Du et al., 2022) and plant proteins (Zhang Q. et al., 2021; Lin et al., 2022; Liu Y. Q. et al., 2022; Ren and Li, 2022; Wang T. et al., 2022; Hariharan et al., 2023; Lv et al., 2023; Vogelsang-O’Dwyer et al., 2023).
4.4.3. Microbial fermentation
Proteases can hydrolyze the protein substrate in the fermentation medium into small peptides, making it easier for microorganisms to quickly absorb and utilize these substrates, improving fermentation efficiency. Other studies have found that during synergistic fermentation of bean dregs and soybean meal, adding multiple strains and protease promotes strain growth, organic acid secretion and amylase secretion and reduces sugar metabolism (Heng et al., 2022). Producing ethanol by microbial fermentation will cause hydrolysis by endogenous proteases and as a result will generate amino acids and peptides. Amino acids and peptides can support the growth of microorganisms, which subsequently increases ethanol production. To improve ethanol yield and reduce fermentation time, exogenous proteases can be used to hydrolyze protein sources available in the raw materials in feedstock used for ethanol production (Thomas and Ingledew, 1990). During high-gravity ethanol production from rice, proteases increased ethanol yield and decreased fermentation time during no-cook processes. Proteases have a significant impact on the size and growth of yeast and were found to enhance ethanol content by 2.4% v/v and shorten fermentation time by 48 h. External nitrogen addition was not needed for the SLSF-VHG process of rice (Tien et al., 2022).
4.4.4. Production of active peptides (or functional oligopeptides)
Active peptides are oligopeptides with specific compositions and sequences of amino acids. They are found in plant and animal proteins, and proteases can specifically hydrolyze proteins and release active peptides. Antioxidative, antidiabetic, antihypertensive, antimicrobial, antitumor, hypocholesterolemic, and many other biological properties may benefit from bioactive peptide structures (Karami and Akbari-adergani, 2019; Mada et al., 2019).
Microbial proteases have been used to produce high-value protein hydrolysates (Tacias-Pascacio et al., 2020; Mirzapour-Kouhdasht et al., 2021), especially antioxidant peptides (Mukhia et al., 2021; Noman et al., 2022; Pan et al., 2022). These proteases can be used in health food and cosmetic fields and show great application potential.
4.4.5. Animal feed
Processing feed ingredients and applying exogenous proteases are the primary uses of exogenous proteases in animal feed. These proteases can be used to maintain high performance and reduce dietary protein levels. Enzymatic hydrolysis is the best method when processing animal byproducts or plant-source feedstuffs. Interesting activities from peptides from plant or animal sources include antihypertensive, antimicrobial, antioxidant, and immunomodulatory activities. The environment also benefits from proteases by improving the utilization of protein materials and reducing nitrogen and ammonia excretions (Philipps-Wiemann, 2018; Hejdysz et al., 2020).
Proteases are used in the following applications:
4.4.5.1. Livestock feed
Adding Bacillus licheniformis to nursery diets that contain a low protein level can significantly improve nutrient digestibility, growth performance, and intestinal morphology of weaned pigs (Park et al., 2020). Keratinolytic proteases can also use low-energy consumption to convert poultry feathers to a nutritionally upgraded protein-rich feedstuff for livestock from a potent pollutant (Onifade et al., 1998).
4.4.5.2. Poultry feed
Protease supplementation can improve the growth performance of broilers. HuPro protease can be supplemented under low-protein conditions to achieve a breeding effect that is similar to a positive control (antibiotic). Proteases can alter the bacterial diversity in the cecum, which has a positive effect on broilers (Wang Y. et al., 2022).
4.4.5.3. Aquafeed
To improve the juiciness, flavor, tenderness, healthiness, and antioxidant capacity of grass carp meat, soy protein hydrolyzed by proteases has been added to a low-protein diet (Song et al., 2020b).
4.4.6. Waste recycling/environmental protection
To process various forms of protein-rich waste, proteases can be used for liquid, solid, and hazardous waste.
Tannery wastewater microbiota was screened for metagenome-derived PersiProtease1. The novel PersiProtease1 was extracted from the microbiota and was applied to biodegrade tannery wastewater protein, dehairing sheepskins, whey protein, chicken feathers, and waste X-ray films (Ariaeenejad et al., 2022).
Several studies have found that proteases exhibit excellent deproteinization for chitin processing of shrimp waste (Jellouli et al., 2011; Mhamdi et al., 2017a,b; Doan et al., 2019).
Another promising pathway for economic benefits and reduced carbon emissions in waste-activated sludge management is the recovery of short-chain fatty acids through anaerobic fermentation. Through alkaline protease–based pretreatment, waste-activated sludge flocs can be disintegrated following cell lysis, which releases biodegradable organic matter. This approach increased the α-glucosidase activities and endogenous protease, facilitated the biodegradation of dissolved organic matter, and encouraged short-chain fatty acid production. This is a promising method for disposing waste-activated sludge and recovering carbon. Short-chain fatty acids might meet 60% of the carbon gap in wastewater, making it a cost-effective and carbon-beneficial technology to manage (Pang et al., 2020, 2022).
Efficient waste-activated sludge dewatering can be achieved through neutral protease. Waste-activated sludge treatment and disposal in wastewater treatment plants require sludge dewaterability. After enzyme conditioning, the sludge supernatant of polysaccharides, proteins, and SCOD content increased, which demonstrated the excellent performance of neutral protease. The capillary suction time increased, and the sludge water content decreased (Kang et al., 2023).
Skatole, the main source of foul odor from feces, is released from the cecum and colon of pigs and is the main source of air pollution in the pig farming environment. A new protease from Lactobacillus brevis has been used to remove odor from pig manure (Meng et al., 2013).
Approximately 5–7% of the total weight of chicken originates from features, which are a major pollutant because of their recalcitrant nature. Feathers are composed of 90% keratin and thus are used as an organic fertilizer because they are good sources of amino acids, peptides, and minerals. Bacteria can degrade keratin through keratinase enzymes. These serine-type proteases have been used as alternatives to develop cost-effective, readily available, and eco-friendly nitrogen- and mineral-rich sources as organic fertilizers (Mazotto et al., 2010; Tamreihao et al., 2018).
Protease and protease-containing formulations can be used to clean hairs from clogged pipes and drains and can be used for depilation (Naveed et al., 2021).
Major contaminants in food bioprocessing sectors (e.g., milk and meat processing activities) result from protein-based residues. Alkaline protease has been used for waste management in different food-processing businesses as well as for activities at in residential areas (Majumder et al., 2015).
To minimize cleaning expenses, reduce environmental dangers, and increase equipment lifetime, various cleaning procedures have used protease alternatives. Because proteases are biodegradable, they will not cause environmental damage after they are used. Unlike other remediation approaches, biomass and chemicals cannot be removed to prevent accumulation. One disadvantage of using proteases for bioremediation is that the enzymes are expensive.
4.5. Special applications
4.5.1. Contact lens cleaning
Cleaning solutions for contact lenses are often prepared using animal (e.g., pancreatin, trypsin, and chymotrypsin) and plant (e.g., papain) proteases. Most of these solutions cause the cleansing bath to exhibit an unpleasant smell or develop an odor after use for a few hours (Liu et al., 2018). Reportedly, however, some microbial proteases can clean the debris off of contact lenses and tear film, making these cleaning compositions odorless and safe. For example, proteases from Bacillus species, Streptomyces sp., and Aspergillus sp. do not cause allergic reactions or eye irritation (Singh and Bajaj, 2017; Razzaq et al., 2019; Rejisha and Murugan, 2021).
4.5.2. Biotechnology processes
Some proteases are used for cleavage of various fusion tags after protein fusion expression in biotechnology protocols.
4.5.2.1. SUMO Protease
Small ubiquitin-related modifier (SUMO) is a kind of ubiquitin-related protein that can be fused with the target protein to promote its solubility and enhance its soluble expression. After expression, SUMO protease can specifically recognize and cut the SUMO sequence from the target protein.
4.5.2.2. Recombinant Kex2 protease
Kex2 protease, a yeast-derived precursor processing protease, is a calcium-dependent serine protease that specifically recognizes and cleaves the carboxy-terminal peptide bond of Arg-Arg, Lys-Arg, Pro-Arg and other bibasic amino acids. Kex2 protease was used for the cleavage of secreted peptides in yeast exogenous protein expression.
4.5.2.3. TEV Protease
TEV protease is a cysteine protease of tobacco etch virus that specifically recognizes the heptapeptide sequence Glu-Asn-Leu-Tyr-Phe-Gln-Gly/Ser and cleaves between Gln and Gly/Ser amino acid residues and is commonly used as a protease to remove GST, HIS or other tags from fusion proteins.
4.5.2.4. Proteinase K
This protease is used in genomic DNA extraction, enzyme digestion and removal in various common molecular biology experiments and cell biology experiments.
4.5.2.5. Recombinant trypsin
This protease is an endopeptidase that can be used for the hydrolysis of C-terminal peptide bonds of lysine and arginine to split macromolecular proteins into small peptides. Trypsin is widely used in various biotechnological processes, such as cell separation of various tissues in cell culture experiments, degradation of denatured protein, enzymatic hydrolysis and sequencing of proteins, stem cell therapy, and cell therapy of tumors.
The sources of microbial proteases are extensive and may originate from any type of microorganism. Fungal proteases have been used in the food industry due to their safety and enzymatic characteristics. Acid proteases, among other functions, may be used as a substitute for activities associated with renin, papain, and pepsin. Species of Aspergillus and Mucor are important acid protease sources. Acid proteases from Aspergillus flavus, Aspergillus oryzae, Aspergillus niger, Rhizomucor pusillus, Rhizomucor miehei, and Rhizopus species are all used to prepare oriental foods, such as tempeh and koji, and to produce cheese as a substitute for rennet. Important milk clotting enzymes are found in Rhizomucor pusillus, Rhizomucor miehei, Penicillium roqueforti, Penicillium camemberti, and Endothia parasitica (Mandujano-González et al., 2016; Aleksandrina, 2021; Ha et al., 2022). Mesophilic fungi have been used to release proteolytic enzymes. Thermophilic fungi with good protease activity include Achaetomium, Chaetomium, Humicola, Rhizomucor, Malbranchea, Penicillium, Rhizopus, Sporotrichum, Torula, and Talaromyces (Johri et al., 1985). Many of these species produce sufficient levels of acid, neutral, alkaline proteases and milk-clotting enzymes. Thermophilic fungi offer low fermenter contamination at high growth temperatures, which is a selective advantage. Thermophilic molds exhibit better enzymatic abilities because of their greater production and higher thermostability, resulting in their widespread commercial applications. Their enzymatic reactions have specificity, rapid speed, and efficiency even in small quantities. The possibility of commercial isolation of some of these thermophilic fungal species has received increased attention (Macchione et al., 2007). Proteases are ideal candidates for laundry detergent because of their thermostability and activity at high pH (Gupta et al., 2002). Proteases from the genus Bacillus meet this requirement most often, so proteases used in laundry detergent mostly originate from the genus Bacillus (David et al., 2018; Rekik et al., 2019; Akram et al., 2020; Emran et al., 2020). Some proteases derived from extreme environmental microorganisms also frequently appear in this field (Abdullah et al., 2022).
In the past few decades, the application field of microbial proteases has rapidly expanded and has played an indispensable role in the food and detergent industries. With the continuous discovery of new microbial proteases, the application fields of microbial proteases will continue to expand, and their application methods will develop toward green and energy-saving directions.
5. Heterologous expression of proteases
Almost all microorganisms can generate proteases, but the amount of protease naturally generated by microorganisms may be very low, and isolating proteases is difficult. To obtain a certain number of proteases for conducting further research or application, heterologous expression of proteases is an important method; furthermore, with the development of bioinformatics, tens of millions of genes have been predicted as proteases, which is a very large potential resource pool of proteases. To obtain valuable proteases, only heterologous expression method can be used to produce enzyme proteins and then perform functional verification. In addition, some characteristics of natural proteases may not meet the requirements of industrial application, so it is necessary to perform modifications, and performing modification on the original enzyme through the heterologous expression system is convenient.
Heterologous expression systems can be divided into prokaryotic expression systems and eukaryotic expression systems. The most representative prokaryotic expression systems are the Escherichia coli expression system and Bacillus subtilis expression system. Eukaryotic expression systems are representative of Pichia pastoris expression systems, and they are the most commonly selected expression systems for protease heterologous expression. The above heterologous expression systems have the characteristics of high expression of target proteins, low expression of background proteins, high expression efficiency and easy operation (Demain and Vaishnav, 2009).
A steady stream of protease-encoding genes has been cloned and expressed in new hosts, and the three major organisms of choice for cloning and overexpression are Escherichia coli, B. subtilis and the Pichia pastoris expression system (Table 7).
Table 7.
Heterologous expression of proteases.
| Host strains for cloning and overexpression | Proteases or encoding genes | Plasmid vectors | References |
|---|---|---|---|
| B. subtilis | |||
| B. subtilis | Glutamyl endopeptidase from Bacillus intermedius (gseBi) | pV | Shagimardanova et al. (2007) |
| B. subtilis DB104 | A thermophilic neutral protease from Bacillus stearothermophilus | Shuttle vector pHP13 | Zhang M. et al. (2008) |
| B. subtilis | An alkaline serine protease gene (GsProS8) from Geobacillus stearothermophilus | pWB980 | Chang et al. (2021) |
| B. subtilis WB800 | A gene coding for the nattokinase (Nk) from B. subtilis strain VTCC-DVN-12-01 | pAC7 | Nguyen et al. (2013) |
| B. subtilis WB800 | A subtilisin-like alkaline serine protease (ASP) from Bacillus halodurans C-125 | pMA0911 | Tekin et al. (2020) |
| B. subtilis WB600 | Keratinase (kerT) gene | pLY | Cao et al. (2019) |
| B. subtilis WB600 | Alkaline serine protease (BcaPRO) from B. clausii | pWBPRO1 constructed based on pWB980 | Liu et al. (2019) |
| B. subtilis SCK6 | A novel streptomyces trypsin | pWB980 | Wang et al. (2019) |
| A recombinant B. subtilis | Extracellular thermostable alkaline halophilic protease | pSaltExSePR5 | Promchai et al. (2018) |
| B. subtilis | Alanine aminopeptidase from Bacillus licheniformis E7 | pMA0911 | Chen Y. et al. (2022) |
| B. subtilis SCK6 | An extracellular keratinase | pMA0911 | Tian et al. (2019) |
| A recombinant B. subtilis | Serine protease from Bacillus intermedius | pCB22 | Sharipova et al. (2008) |
| B. subtilis | A fibrinolytic enzyme (subtilisin DFE) gene | pSUGV4 | Peng et al. (2004) |
| B. subtilis WB600 | Fibrinolytic protease of Bacillus licheniformis CH 3–17 | pHY300PLK | Jo et al. (2011) |
| E. coli | |||
| E. coli | An intracellular serine protease from isolated salt-tolerant Bacillus sp. LCB10 | pET-30a | Hou et al. (2019) |
| E. coli Transetta (DE3) | Subtilisin-like protease from a thermophilic Thermus thermophilus HB8 | pET-22b (+) | Xie et al. (2019) |
| E. coli strain BL21 | Serine alkaline protease | pET-15b | Suberu et al. (2019) |
| E. coli BL21-Gold (Stratagene), E. coli ORIGAMI B and E. coli Rosetta2 | Extracellular serine proteases from Stenotrophomonas maltophilia | pMS470Δ8 | Ribitsch et al. (2012) |
| E. coli BL21(DE3) pLysS and E. coli BL21-AI™ | Serine alkaline protease (SAPN) from Melghiribacillus genus. | pUT57 and pTrc99A, Gateway™ pDEST™ 17 | Mechri et al. (2021) |
| E. coli BL21 | A novel alkaline serine protease gene from native Iranian Bacillus sp. | pET-28 a (+) | Ariyaei et al. (2019) |
| E. coli | Serine protease from Nocardiopsis sp. | pET-39b (+) and pET-22b (+) | Rohamare et al. (2015) |
| E. coli BL21 (DE3) | A thermo- and surfactant-stable protease from Thermomonospora curvata | pET-25b (+) | Sittipol et al. (2019) |
| E. coli BL21 (DE3) | Serine proteases from Oceanobacillus iheyensis O.M.A18 and Haloalkaliphilic bacterium O.M.E12 | pET-21a (+) | Purohit and Singh (2014) |
| E. coli BL21 (DE3) | Subtilisin-like Protease Myroicolsin | pET-22b (+) | Ran et al. (2014) |
| E. coli BL21 | An alkaline protease from Bacillus licheniformis | pET–28b (+) | Lin et al. (2015) |
| E. coli BL21(DE3) | A fibrinolytic protease gene from the polychaeta, Periserrula leucophryna | pT7-7 | Joo et al. (2013) |
| E. coli BL21(DE3) | A novel fibrinolytic serine protease from Arenicola cristata | pET-21a (+) | Zhao and Ju (2014) |
| E. coli BL21(DE3) | A novel aspartic protease gene from marine-derived Metschnikowia reukaufii | pET-24a (+) | Li et al. (2008) |
| E. coli BL21(DE3) | A novel extracellular subtilisin-like protease from the hyperthermophile Aeropyrum pernix K1 | pGEX | Catara et al. (2003) |
| E. coli BL21(DE3) | A new thiol-dependent, alkaline serine protease | pET-28a (+) | Masilamani and Natarajan (2015) |
| E. coli BL21(DE3) pLysS | A serine protease-like protein from silkworm (Bombyx mori) | pGEX-5X-1 | Kim et al. (2009) |
| E. coli BL21 | Lon protease from rice (Oryza sativa) | pET-32a | Su et al. (2006) |
| E. coli Rosetta-gami (DE3) | Serine protease aprv2 from virulent isolate Dichelobacter nodosus | pET-22b (+) | Wani et al. (2016) |
| E. coli BL21 (DE3) | A novel extracellular cold-adapted alkaline protease gene of the marine bacterium strain YS-80-122 | pET-28a | Wang et al. (2010) |
| E. coli Shuffle®T7. | Three main serine carboxypeptidases (SCP3, SCP20, and SCP47) from Nepenthes mirabilis | pET-28a-SUMO | Porfírio et al. (2022) |
| E. coli BL21(DE3) | Carboxypeptidases genes (dacA, dacB, dacC, and dacF) in Bacillus subtilis CW14 | pET-28a (+) | Xu et al. (2021) |
| E. coli | A collagenolytic aspartic protease from Thermomucor indicae-seudaticae | pET-28a (+) | Pereira et al. (2020) |
| E. coli BL21(DE3) pLysS | Novel protease from Bacillus licheniformis strain K7A | pGEM-T Easy | Hadjidj et al. (2018) |
| E. coli BL21 (DE3) | A low salt-adapted extracellular protease from the extremely halophilic archaeon Halococcus salifodinae | pET-28a | Hou et al. (2021) |
| E. coli BL21 | Avihepatovirus 3C protease | pET-32a | Sun et al. (2019) |
| E. coli BL21 (DE3) pLysS | Site-2 protease | pHEN6 | Schacherl et al. (2017) |
| E. coli BL21 (DE3) | A dual-functional aminopeptidase from Streptomyces canus T20 | pET-24a (+) | Qin et al. (2021) |
| P. pastoris | |||
| P. pastoris SMD1168 and X33 | Serine alkaline protease from Melghiribacillus genus | pPICZαC | Mechri et al. (2021) |
| P. pastoris GS115 | Serine protease from thermophilic fungus Thermoascus aurantiacus var. levisporus | pPIC9K | Li et al. (2011) |
| Saccharomyces cerevisiae W3124 | An aspartic protease | pYES 2.0 | Bang et al. (1999) |
| P. pastoris | Proteinase K | pPink-HC | Skowron et al. (2020) |
| P. pastoris KM71 | Protease from Aspergillus niger | pPIC9K | Ke et al. (2019) |
| P. pastoris | A plant aspartic protease (preprogaline B) | pGAPZα A | Feijoo-Siota et al. (2018) |
| P. pastoris | A novel serine protease | A self-construct plasmid | Shu et al. (2016) |
| P. pastoris X-33 and GS115 (his4) | Streptokinase | pPICZαA and pGAPZαA | Adivitiya Dagar et al. (2016) |
| P. pastoris | Bacillus pumilus 3–19 protease | pPINK-HC | Pudova et al. (2021) |
| P. pastoris | A zinc-dependent proteases of the metzincin superfamily of metalloproteases | pPIC9K | Schlenzig and Schilling (2017) |
| P. pastoris GS115 | Recombinant protease MarP from Mycobacterium tuberculosis | pPICZα | García-González et al. (2021) |
| P. pastoris | A collagenolytic aspartic protease from Thermomucor indicae-seudaticae | pPICZαA | Pereira et al. (2020) |
| P. pastoris | A novel aminopeptidase B from Aspergillus niger | pPIC9K | Song and Feng (2021) |
| P. pastoris X-33 and SMD1168 strains | A Mucor circinelloides aspartic protease | pGAM1 | Kangwa et al. (2018) |
| P. pastoris | A new serine protease from Crotalus durissus collilineatus venom | PICZαA | Boldrini-França et al. (2015) |
| P. pastoris | Keratinolytic serine protease gene sfp2 from Streptomyces fradiae var. k11 | pPIC9K | Li et al. (2007) |
| P. pastoris KM71 | Subtilisin Pr1A gene from a strain of locust specific fungus, Metarhizium anisopliae | pPIC9K | Zhang W. et al. (2008) |
| P. pastoris GS115 | Subtilisin QK | pPICZα | Zhou et al. (2017) |
| P. pastoris SMD1168 and X33 | A serine alkaline protease from Melghiribacillus thermohalophilus | pPICZαC | Mechri et al. (2021) |
| P. pastoris KM71 | Aspergillus sojae alkaline protease | pPIC9K | Ke et al. (2018) |
| P. pastoris GS115 | Recombinant Aspergillus oryzae alkaline protease | pPIC9K | Guo and Ma (2008) |
| P. pastoris | Keratinase (kerA) gene from Bacillus licheniformis | pPICZαA | Porres et al. (2002) |
| P. pastoris | A thermostable serine protease (TfpA) from Thermomonospora fusca YX | pPICZαA | Kim and Lei (2005) |
| P. pastoris GS115 | The Bacillus subtilis subsp. subtilis str. BSP1 YwaD aminopeptidase | pHBM905A | Tang et al. (2016) |
| P. pastoris GS115 | Carboxypeptidase Y from Saccharomyces cerevisiae | pHBM905A | Yu et al. (2014) |
| P. pastoris X-33 | Prolyl aminopeptidase | pPIC9K | Yang et al. (2016) |
| P. pastoris X-33 | X-Prolyl-dipeptidyl aminopeptidase from Basidiomycete ustilago maydis | pPICZαB | Juárez-Montiel et al. (2014) |
| P. pastoris X33 or SMD1168H | A metalloprotease | pPICZαC | Schlenzig et al. (2015) |
| P. pastoris GS 115 | A new dipeptidyl-peptidase isolated from Aspergillus fumigatus | pHIL-S1 | Beauvais et al. (1997) |
| P. pastoris GS115 | A metalloprotease | pPIC9K | Song et al. (2021b) |
| P. pastoris GS115 | A new carboxypeptidase from Aspergillus niger | pPIC9K | Song et al. (2021a) |
| P. pastoris GS115 | Two new Aspergillus niger aspartic proteases | pPIC9K | Song Y. et al. (2020) |
| Pichia pastoris KM71H | Serine protease from Bothrops pauloensis snake venom | pPICZαA | Isabel et al. (2016) |
| P. pastoris GS115 | A thermolysin-like protease, neutral protease I, from Aspergillus oryzae | pHBM905BDM | Ma et al. (2016) |
| P. pastoris GS115 | First fibrinolytic enzyme from mushroom (Cordyceps militaris) | pPIC9K | Katrolia et al. (2019) |
| P. pastoris GS115 | Thrombolytic enzyme (lumbrokinase) from earthworm | pPICZα-B | Ge et al. (2005) |
From the above table, we can find that proteases from animals, plants, microorganisms or viruses can be successfully expressed in the three heterologous expression systems (E. coli, B. subtilis and P. pastoris). Another important feature of proteases is that a significant part is found in nature as a protein precursor (or zymogen). These proteases can be synthesized as inactive or less active precursor molecules, which have developed after evolution. These principal mechanisms can control the activity of proteases. The propeptide sequence of a protein precursor is connected to the C- or N-terminus of the material protein. The propeptides within protease precursors likely perform the following physiological functions: (1) help fold the mature enzyme; (2) provide the protease interaction with the bacterial cell surveillance mechanisms, including protease translocation through the cell wall; and (3) inhibit the proteases to protect the host cells from proteolytic damage (Baker et al., 1992; Baardsnes et al., 1998; Serkina et al., 2001; Varón et al., 2006).
For a protease to function, the propeptide must be removed, and the zymogen must be activated to produce a functional mature protease, so activation of the zymogen is important for proteases. Regulation of proteolytic enzyme activity is necessary for cells and tissues because proteolysis at the wrong time and location may be lethal.
The mechanisms by which zymogens activate proteolytic enzymes are diverse and naturally occurring. They are activated, in some cases, upon enzymatic or nonenzymatic cofactor triggering, an appropriate signal such as acidification, Ca++-binding or, in other cases, by limited intra- or intermolecular proteolysis cleaving off an inhibitory peptide (Khan and James, 1998; Marie-Claire et al., 1998; Takagi et al., 2001; Wiederanders et al., 2003).
However, regarding the heterologous expression of some proteases, their activation mechanism or whether the activation mechanism of zymogens occurs in the heterologous host are unable to predict in advance; whether the propeptide is retained is unknown? Some relevant reports were summarized and the result showed that most of the successfully expressed proteases were expressed with the retention of the propeptide, but some of them were successful in removing the leading peptide and expressing the mature peptide directly (Table 8).
Table 8.
Heterologously expressed proteases with (without) the retention of the propeptide.
| Proteases | Protease gene sources | Expression host | Expression with propreptide (Y/N) | Remarks | Reference |
|---|---|---|---|---|---|
| A keratinolytic serine protease | Streptomyces fradiae var.k11 | E. coli | Y | Recombinants expressing the proenzyme exhibited markedly higher activity than that recombinant expressing mature enzyme | Meng et al. (2007) |
| Keratinase | Bacillus licheniformis | Pichia pastoris | Y | Expressing the pro-mature structure was reported to increase the production of the keratinases gene (kerA) | Carlsson et al. (2001) |
| A thermolysin-like neutral protease | Bacillus stearothermophilus | E. coli | N | A much more effective access to active mature protease was found when TLP-ste (devoid of its prosequence) was expressed this confirming that the propeptide is not essential for proper folding of the enzyme or its stabilization during the folding process | Mansfeld et al. (2005) |
| A novel extracellular serine protease | Aeropyrum pernix K1 | E. coli | N | / | Catara et al. (2003) |
| Subtilisin | Thermus thermophilus HB8 | E. coli | Y | Expression of the mature-subtilisin gene was found to produce inactive inclusion bodies, expression of the pro-subtilisin gene resulted in active mature-subtilisin | Xie et al. (2019) |
| Subtilisin E | Bacillus subtilis 168 | E. coli | Y | When the entire coding region for pre-pro-subtilisin E was cloned into an Escherichia coli expression vector, active mature subtilisin E was secreted into the periplasmic space; When the propeptide was absence to the mature subtilisin sequence, no protease activity was detected | Ikemura et al. (1987) |
| Candida secreted aspartic proteases | Candida albicans | Pichia patoris | N | Expression of the C. albicans SAP1 gene lacking the propeptide-coding region in the Pichia pastoris does not lead to the secretion of the enzyme into the culture supernatant, but results in an accumulation of recombinant protein in the cell. Co-expression in this system of the unattached propeptide from Sap1p, as well as from other Saps, restored Sap1p secretion | Beggah et al. (2000) |
| Keratinase | Bacillus licheniformis BBE11-1 | E. coli | Y | Optimizing the C-terminus of propeptide will affect the cleavage efficiency of propeptide. The primary structure of C-terminus propeptide is crucial for the mature keratinase production. | Liu et al. (2014) |
| Leucyl aminopeptidase A | Aspergillus oryzae RIB40 | E. coli and Pichia pastoris | Y | / | Baltulionis et al. (2021) |
| Subtilisin-like serine proteases | Tomato | E. coli | Y | / | Meyer et al. (2016) |
| Nattokinase | Bacillus subtilis VTCC-DVN-12-01 | Bacillus subtilis WB800 | Y | / | Nguyen et al. (2013) |
For the heterologous expression of a novel protease, the precursor sequence should be cloned, and if there is no functional enzyme expression, then in turn clone the mature peptide sequence. The identification of the propeptide sequence of a protease can be performed by referring to the literature or using prediction software of the peptide clearance sites: https://services.healthtech.dtu.dk/service.php?ProP-1.0. Notably, the propeptide sequence of a protease may be at the N-terminal end or the C-terminal end of the protein sequence.
Heterologous expression is an important method for detecting novel microbial proteases and will continue to play a crucial role. Moreover, as an optimization of this approach, some new technologies, such as CRISPR and directed evolution, will continue to be applied to optimize this approach and improve the method’s efficiency. As representatives of prokaryotic expression systems, E. coli expression systems and B. subtilis expression systems, as well as eukaryotic expression systems, P. pastoris expression systems can now meet the heterologous expression of proteases from prokaryotic and eukaryotic microorganisms. Expression levels could be increased and more functional protease expression could be obtained to further improve heterologous expression.
6. Conclusion and future prospects
Because enzymes are environmentally friendly chemicals, they could completely replace or reduce the use of hazardous chemicals in industrial processes. As a result, enzymes show promising applications for sustainable manufacturing and production. Proteases are superior to many industrial enzymes because of their varied application in many different bioindustries, such as detergent, leather, textiles, and food, as well as pharmaceutical, biotechnology, and waste treatment processes. Among proteases from diverse sources, microbial proteases have been the preferred source for applications owing to their fast growth, efficient production, wide diversity, longer shelf life, and potential for genetic manipulation of microorganisms compared to plant or animal sources.
It is certain that microbial protease, as a green, efficient tool, will be continuously applied in various industry applications with the development of biological technology, and it will lead the development of the abovementioned fields or promote the development of each field. To increase our fundamental knowledge on microbial ecology (e.g., enzymes, their evolution, and their relevance in industrial sectors), “omics” and biological technologies should continue to be used for molecular characterization, crystallography, and enzymatic modulation by applying algorithms, bioinformatics tools, and genetic engineering. Genetic engineering and immobilization techniques should be further developed to discover new proteases, enzyme systems that are more effective and efficient should be developed, the functions of existing proteases should be optimized, and fewer resources and energy should be consumed while achieving maximum product yields.
In the next decade or few decades, research should be conducted on proteases regarding enzyme preparation methods and usage methods to improve efficiency, such as developing immobilized enzyme technology, enzyme modification technology, and protease fusion application with chemical approaches and developing faster and more efficient methods for detecting and analyzing proteases to facilitate the development of new proteases. De novo design of new proteases using artificial intelligence and various algorithms should also be applied. It is also necessary to develop general methods for long-term preservation of proteases to mitigate inactivation caused by self-hydrolysis; greater accuracy and control during the production process is critical in terms of improving product value or expanded substrate extensiveness of proteases with a goal of obtaining better hydrolysis efficiency. Biochemical attributes of microbial proteases, such as thermostable, cold-active, and halophilic extreme environmental properties, should be further studied to determine significant applications in bioprocesses, as proteases and even enzymes are always of research interest.
Author contributions
PS and FWe: conceptualization. PS, XZ, SW, WX, and FWa: literature search. PS, XZ, and FWa: writing–original draft preparation. PS, SW, FWe, and RF: writing–review and editing. PS and FWe: funding acquisition. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the Natural Science Foundation of Shandong Province (grant number ZR2022MC159).
Conflict of interest
PS and SW were employed by Shandong Aobo Biotech Co. Ltd.; PS and RF were employed by Jiangxi Zymerck Biotechnology Co., Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We would like to thank AJE (https://www.aje.cn) for English language editing.
References
- Aalbæk T., Reeslev M., Jensen B., Eriksen S. H. (2002). Acid protease and formation of multiple forms of glucoamylase in batch and continuous cultures of Aspergillus niger. Enzym. Microb. Technol. 30, 410–415. doi: 10.1016/s0141-0229(02)00006-6 [DOI] [Google Scholar]
- Abdel Wahab W. A., Ahmed S. A. (2018). Response surface methodology for production, characterization and application of solvent, salt and alkali-tolerant alkaline protease from isolated fungal strain Aspergillus niger WA 2017. Int. J. Biol. Macromol. 115, 447–458. doi: 10.1016/j.ijbiomac.2018.04.041, PMID: [DOI] [PubMed] [Google Scholar]
- Abdullah A. A., Babu J., Mohammed S. A., Pramod W. R. (2022). “Chapter 17-current applications and future trends of extremozymes in detergent industries”, in Microbial extremozymes. ed. Kuddus Mohammed. (Salt Lake City, UT: Academic Press; ), 223–230. [Google Scholar]
- Acrofan (2021). Global industrial enzymes market (2020 to 2027)-featuring BASF, Novozymes and DSM among others–research and markets. Available at: http://us.acrofan.com/detail.php?number=511126 [Accessed February 18, 2023]
- Adivitiya Dagar V. K., Devi N., Khasa Y. P. (2016). High level production of active streptokinase in Pichia pastoris fed-batch culture. Int. J. Biol. Macromol. 83, 50–60. doi: 10.1016/j.ijbiomac.2015.11.062, PMID: [DOI] [PubMed] [Google Scholar]
- Adler-Nissen J. (1979). Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzene sulfonic acid. J. Agric. Food Chem. 27, 1256–1262. doi: 10.1021/jf60226a042, PMID: [DOI] [PubMed] [Google Scholar]
- Ai M., Tang T., Zhou L., Ling Z., Guo S., Jiang A. (2019). Effects of different proteases on the emulsifying capacity, rheological and structure characteristics of preserved egg white hydrolysates. Food Hydrocoll. 87, 933–942. doi: 10.1016/j.foodhyd.2018.09.023 [DOI] [Google Scholar]
- Akram Z., Asgher M., Ahmad S. Q. (2023). “Chapter 18 - microbial proteases—robust biocatalytic tools for greener biotechnology” in Developments in applied microbiology and biotechnology, microbial biomolecules. eds. Kumar A., Bilal M., Ferreira L. F. R., Kumari M. (Salt Lake City, UT: Academic Press; ), 405–427. [Google Scholar]
- Akram F., Haq I., Jabbar Z. (2020). Production and characterization of a novel thermo- and detergent stable keratinase from Bacillus sp. NKSP-7 with perceptible applications in leather processing and laundry industries. Int. J. Biol. Macromol. 164, 371–383. doi: 10.1016/j.ijbiomac.2020.07.146, PMID: [DOI] [PubMed] [Google Scholar]
- Albillos S. M., Busto M. D., Perez-Mateos M., Ortega N. (2007). Analysis by capillary electrophoresis of the proteolytic activity of a Bacillus subtilis neutral protease on bovine caseins. Int. Dairy J. 17, 1195–1200. doi: 10.1016/j.idairyj.2007.02.003 [DOI] [Google Scholar]
- Alblooshi M., Devarajan A. R., Singh B. P., Ramakrishnan P., Mostafa H., Kamal H., et al. (2023). Multifunctional bioactive properties of hydrolysates from colocynth (Citrullus colocynthis) seeds derived proteins: characterization and biological properties. Plant Physiol. Biochem. 194, 326–334. doi: 10.1016/j.plaphy.2022.11.026, PMID: [DOI] [PubMed] [Google Scholar]
- Aleksandrina P. (2021). Fungal proteases: current and potential industrial applications. Encyclo. Mycol. 2, 348–357. doi: 10.1016/B978-0-12-819990-9.00025-1 [DOI] [Google Scholar]
- Aljammas H. A., Yazji S., Azizieh A. (2022a). Enhancement of protease production from Rhizomucor miehei by mutagenesis with ethyl methanesulfonate, ultraviolet, and microwaves- a preliminary study. Biores. Tech. Rep. 20:101287. doi: 10.1016/j.biteb.2022.101287 [DOI] [Google Scholar]
- Aljammas H. A., Yazji S., Azizieh A. (2022b). Optimization of protease production from Rhizomucor miehei Rm4 isolate under solid-state fermentation. J. Genet. Eng Biotech. 20, 82–13. doi: 10.1186/s43141-022-00358-9, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anis Ahamed N., Arif I. A., Al-Rashed S., Panneerselvam A., Ambikapathy V. (2022). In vitro thrombolytic potential of fibrinolytic enzyme from Brevibacterium sp. isolated from the root of the plant, aloe castellorum. J. King Saud. Univ. Sci. 34:101868. doi: 10.1016/j.jksus.2022.101868 [DOI] [Google Scholar]
- Anitha T. S., Palanivelu P. (2013). Purification and characterization of an extracellular keratinolytic protease from a new isolate of Aspergillus parasiticus. Protein Expr. Purif. 88, 214–220. doi: 10.1016/j.pep.2013.01.007, PMID: [DOI] [PubMed] [Google Scholar]
- Anzani C., Prandi B., Tedeschi T., Baldinelli C., Sorlini G., Wierenga P. A., et al. (2017). Degradation of collagen increases nitrogen solubilization during enzymatic hydrolysis of fleshing meat. Waste. Biomass. Valori. 9, 1113–1119. doi: 10.1007/s12649-017-9866-4 [DOI] [Google Scholar]
- Ariaeenejad S., Kavousi K., Mamaghani A. S. A., Ghasemitabesh R., Hosseini Salekdeh G. (2022). Simultaneous hydrolysis of various protein-rich industrial wastes by a naturally evolved protease from tannery wastewater microbiota. Sci. Total Environ. 815:152796. doi: 10.1016/j.scitotenv.2021.152796, PMID: [DOI] [PubMed] [Google Scholar]
- Ariyaei A., Farhadi A., Moradian F., Rahimi Mianji G. (2019). Cloning, expression and characterization of a novel alkaline serine protease gene from native Iranian Bacillus sp.; a producer of protease for use in livestock. Gene 693, 10–15. doi: 10.1016/j.gene.2019.01.020, PMID: [DOI] [PubMed] [Google Scholar]
- Asitok A., Ekpenyong M., Takon I., Antai S., Ogarekpe N., Antigha R., et al. (2022). Overproduction of a thermo-stable halo-alkaline protease on agro-waste-based optimized medium through alternate combinatorial random mutagenesis of Stenotrophomonas acidaminiphila. Biotechnol. Rep. 35:e00746. doi: 10.1016/j.btre.2022.e00746, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Austin M. J., Schunk H., Watkins C., Ling N., Chauvin J., Morton L., et al. (2022). Fluorescent peptomer substrates for differential degradation by metalloproteases. Biomacromolecules 23, 4909–4923. doi: 10.1021/acs.biomac.2c01077, PMID: [DOI] [PubMed] [Google Scholar]
- Ayhan F., Çelebi S. S., Tanyolaç A. (2001). The effect of fermentation parameters on the production of Mucor miehei acid protease in a chemically defined medium. J. Chem. Technol. Biotechnol. 76, 153–160. doi: 10.1002/jctb.364 [DOI] [Google Scholar]
- Baardsnes J., Sidhu S., MacLeod A., Elliott J., Morden D., Watson J., et al. (1998). Streptomyces griseus protease B: secretion correlates with the length of the propeptide. J. Bacteriol. 180, 3241–3244. doi: 10.1128/JB.180.12.3241-3244.1998, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baker D., Silen J. L., Agard D. A. (1992). Protease pro region required for folding is a potent inhibitor of the mature enzyme. Prot. Struct. Funct. Bioinform. 12, 339–344. doi: 10.1002/prot.340120406, PMID: [DOI] [PubMed] [Google Scholar]
- Baltulionis G., Blight M., Robin A., Charalampopoulos D., Watson K. A. (2021). The role of propeptide-mediated autoinhibition and intermolecular chaperone in the maturation of cognate catalytic domain in leucine aminopeptidase. J. Struct. Biol. 213:107741. doi: 10.1016/j.jsb.2021.107741, PMID: [DOI] [PubMed] [Google Scholar]
- Bang M. L., Villadsen I., Sandal T. (1999). Cloning and characterization of an endo-β-1,3(4)glucanase and an aspartic protease from Phaffia rhodozyma CBS 6938. Appl. Microbiol. Biotechnol. 51, 215–222. doi: 10.1007/s002530051384, PMID: [DOI] [PubMed] [Google Scholar]
- Beauvais A., Monod M., Debeaupuis J. P., Diaquin M., Kobayashi H., Latgé J. P. (1997). Biochemical and antigenic characterization of a new dipeptidyl-peptidase isolated from Aspergillus fumigatus. J. Biol. Chem. 272, 6238–6244. doi: 10.1074/jbc.272.10.6238, PMID: [DOI] [PubMed] [Google Scholar]
- Beggah S., Léchenne B., Reichard U., Foundling S., Monod M. (2000). Intra- and intermolecular events direct the propeptide-mediated maturation of the candida albicans secreted aspartic proteinase Sap1p. Microbiology 146 (Pt 11), 2765–2773. doi: 10.1099/00221287-146-11-2765, PMID: [DOI] [PubMed] [Google Scholar]
- Bell S. J., Henschke P. A. (2005). Implications of nitrogen nutrition for grapes, fermentation and wine. Aust. J. Grape Wine R. 11, 242–295. doi: 10.1111/j.1755-0238.2005.tb00028.x [DOI] [Google Scholar]
- Belmessikh A., Boukhalfa H., Mechakra-Maza A., Gheribi-Aoulmi Z., Amrane A. (2013). Statistical optimization of culture medium for neutral protease production by Aspergillus oryzae. Comparative study between solid and submerged fermentations on tomato pomace. J. Taiwan. Inst. Chem. E 44, 377–385. doi: 10.1016/j.jtice.2012.12.011 [DOI] [Google Scholar]
- Bernardeau M., Hibberd A. A., Saxer G., Velayudhan D. E., Marchal L., Vinyeta E. (2022). O122 intrinsic properties of 3 Bacillus spp. strains from animal origin constituent of a direct fed Microbials/protease blend having growth performance in pigs fed high fiber diet. An. Sci. Proc. 13, 394–395. doi: 10.1016/j.anscip.2022.07.132 [DOI] [Google Scholar]
- Blair I. S., McDowell D. A. (1995). Detection of extracellular proteinase of Pseudomonas fragi by enzyme-linked immunosorbent assay. Curr. Microbiol. 31, 180–185. doi: 10.1007/BF00293551 [DOI] [Google Scholar]
- Boldrini-França J., Rodrigues R. S., Santos-Silva L. K., de Souza D. L. N., Gomes M. S. R., Cologna C. T., et al. (2015). Expression of a new serine protease from Crotalus durissus collilineatus venom in Pichia pastoris and functional comparison with the native enzyme. Appl. Microbiol. Biotechnol. 99, 9971–9986. doi: 10.1007/s00253-015-6836-2, PMID: [DOI] [PubMed] [Google Scholar]
- Born K., Manns A., Dzeyk K., Lutz-Wahl S., Gau D., Fischer L. (2010). Evaluation of ultrasound velocity measurements for estimating protease activities using casein as substrate. Biotechnol. Lett. 32, 249–253. doi: 10.1007/s10529-009-0135-x, PMID: [DOI] [PubMed] [Google Scholar]
- Borrelli R. C., Mennella C., Barba F., Russo M., Russo G. L., Krome K., et al. (2003). Characterization of colored compounds obtained by enzymatic extraction of bakery products. Food Chem. Toxicol. 41, 1367–1374. doi: 10.1016/S0278-6915(03)00140-6, PMID: [DOI] [PubMed] [Google Scholar]
- Breidenbach J., Bartz U., Gütschow M. (2020). Coumarin as a structural component of substrates and probes for serine and cysteine proteases. BBA-Biomemb. Prot. Prot. 1868:140445. doi: 10.1016/j.bbapap.2020.140445, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burchacka E., Pięta P., Łupicka-Słowik A. (2022). Recent advances in fungal serine protease inhibitors. Biomed. Pharmacother. 146:112523. doi: 10.1016/j.biopha.2021.112523, PMID: [DOI] [PubMed] [Google Scholar]
- Bureros K. J. C., Dizon E. I., Israel K. A. C., Abanto O. D., Tambalo F. Z. (2020). Physicochemical and sensory properties of carabeef treated with Bacillus subtilis (Ehrenberg) Cohn protease as meat tenderizer. J. Food. Sci. Tech. Mys. 57, 310–318. doi: 10.1007/s13197-019-04062-4, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cahan R., Axelrad I., Safrin M., Ohman D. E., Kessler E. (2001). A secreted aminopeptidase of Pseudomonas aeruginosa. J. Biol. Chem. 276, 43645–43652. doi: 10.1074/jbc.m106950200 [DOI] [PubMed] [Google Scholar]
- Campbell J. M., Peberdy J. F. (1983). Properties of Aspergillus nidulans neutral protease. FEMS Microbiol. Lett. 16, 49–53. doi: 10.1111/j.1574-6968.1983.tb00257.x [DOI] [Google Scholar]
- Cao S., Li D., Ma X., Xin Q., Song J., Lu F., et al. (2019). A novel unhairing enzyme produced by heterologous expression of keratinase gene (kerT) in Bacillus subtilis. World J. Microbiol. Biotechnol. 35, 122–132. doi: 10.1007/s11274-019-2701-2, PMID: [DOI] [PubMed] [Google Scholar]
- Capiralla H., Hiroi T., Hirokawa T., Maeda S. (2002). Purification and characterization of a hydrophobic amino acid—specific endopeptidase from Halobacterium halobium S9 with potential application in debittering of protein hydrolysates. Process Biochem. 38, 571–579. doi: 10.1016/S0032-9592(02)00180-2 [DOI] [Google Scholar]
- Carlsson P., Thormählen P., Skoglundh M., Persson H., Fridell E., Jobson E., et al. (2001). Periodic control for improved low-temperature catalytic activity. Top. Catal. 16-17:343. doi: 10.1023/A:1015083007746 [DOI] [Google Scholar]
- Catara G., Ruggiero G., La Cara F., Digilio F. A., Capasso A., Rossi M. (2003). A novel extracellular subtilisin-like protease from the hyperthermophile Aeropyrum pernix K1: biochemical properties, cloning, and expression. Extremop. Life Under Ext. Cond. 7, 391–399. doi: 10.1007/s00792-003-0337-4, PMID: [DOI] [PubMed] [Google Scholar]
- Cejudo-Bastante M. J., Oliva-Sobrado M., González-Miret M. L., Heredia F. J. (2022). Optimization of the methodology for obtaining enzymatic protein hydrolysates from an industrial grape seed meal residue. Food Chem. 370:131078. doi: 10.1016/j.foodchem.2021.131078, PMID: [DOI] [PubMed] [Google Scholar]
- Celebi M., Topuzogullari M., Kuzu H. (2016). Thermal destabilization of Rhizomucor miehei rennet with aldehyde dextran sulfate: purification, bioconjugation and milk-clotting activities. Appl. Biochem. Biotechnol. 180, 261–273. doi: 10.1007/s12010-016-2097-5, PMID: [DOI] [PubMed] [Google Scholar]
- Cermeño M., Bascón C., Amigo-Benavent M., Felix M., Fitz Gerald R. J. (2022). Identification of peptides from edible silkworm pupae (Bombyx mori) protein hydrolysates with antioxidant activity. J. Funct. Foods 92:105052. doi: 10.1016/j.jff.2022.105052 [DOI] [Google Scholar]
- Cerreti M., Liburdi K., Benucci I., Emiliani Spinelli S., Lombardelli C., Esti M. (2017). Optimization of pectinase and protease clarification treatment of pomegranate juice. LWT-Food Sci. Technol. 82, 58–65. doi: 10.1016/j.lwt.2017.04.022 [DOI] [Google Scholar]
- Chang C., Gong S., Liu Z., Yan Q., Jiang Z. (2021). High level expression and biochemical characterization of an alkaline serine protease from Geobacillus stearothermophilus to prepare antihypertensive whey protein hydrolysate. BMC Biotechnol. 21:21. doi: 10.1186/s12896-021-00678-7, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Y. (2020). Fluorescent probes for detection and bioimaging of leucine aminopeptidase. Mat. Today Chem. 15:100216. doi: 10.1016/j.mtchem.2019.100216 [DOI] [Google Scholar]
- Chen S., Cao M., Su W., Wu G. (2011). Purification and characterization of a novel leucine aminopeptidase from the earthworm Eisenia foetida. Process Biochem. 46, 1641–1648. doi: 10.1016/j.procbio.2011.05.008 [DOI] [Google Scholar]
- Chen C. S., Chen W. N. U., Zhou M., Arttamangkul S., Haugland R. P. (2000). Probing the cathepsin D using a BODIPY FL–pepstatin a: applications in fluorescence polarization and microscopy. J. Biochem. Biophys. Methods 42, 137–151. doi: 10.1016/s0165-022x(00)00048-8, PMID: [DOI] [PubMed] [Google Scholar]
- Chen W., Li L., Ye C., Zhao Z., Huang K., Zou D., et al. (2022). Efficient production of extracellular alkaline protease in Bacillus amyloliquefaciens by host strain construction. Food Sci. Tech. Brazil. 163:113620. doi: 10.1016/j.lwt.2022.113620 [DOI] [Google Scholar]
- Chen X., Wu G., Cai Q., Liu G., Osatomi K., Su W., et al. (2012). Biochemical characterization of an aminopeptidase with highest preference for lysine from Japanese flounder skeletal muscle. Food Chem. 130, 679–686. doi: 10.1016/j.foodchem.2011.07.109 [DOI] [Google Scholar]
- Chen Y., Zhang R., Zhang W., Xu Y. (2022). Alanine aminopeptidase from Bacillus licheniformis E7 expressed in bacillus subtilis efficiently hydrolyzes soy protein to small peptides and free amino acids. Food Sci. Technol. 165:113642. doi: 10.1016/j.lwt.2022.113642 [DOI] [Google Scholar]
- Cheung L. K. Y., Aluko R. E., Cliff M. A., Li-Chan E. C. Y. (2015). Effects of exopeptidase treatment on antihypertensive activity and taste attributes of enzymatic whey protein hydrolysates. J. Funct. Foods 13, 262–275. doi: 10.1016/j.jff.2014.12.036 [DOI] [Google Scholar]
- Chiba Y., Yamagata Y., Iijima S., Nakajima T., Ichishima E. (1993). The carbohydrate moiety of the acid carboxypeptidase from Aspergillus saitoi. Curr. Microbiol. 27, 281–288. doi: 10.1007/bf01575993, PMID: [DOI] [PubMed] [Google Scholar]
- Chin J., Kim H. J. (2018). Near-infrared fluorescent probes for peptidases. Coord. Chem. Rev. 354, 169–181. doi: 10.1016/j.ccr.2017.07.009 [DOI] [Google Scholar]
- Chitte R., Chaphalkar S. (2017). “The world of proteases across microbes, insects, and medicinal trees” in The proteases in physiology and pathology. eds. Chakraborti S., Dhalla N. (Singapore: Springer; ) [Google Scholar]
- Cho S. (2019). Primary structure and characterization of a protease from Bacillus amyloliquefaciens isolated from meju, a traditional Korean soybean fermentation starter. Proc. Biochem. 80, 52–57. doi: 10.1016/j.procbio.2019.02.011 [DOI] [Google Scholar]
- Christensen L. F., García-Béjar B., Bang-Berthelsen C. H., Hansen E. B. (2022). Extracellular microbial proteases with specificity for plant proteins in food fermentation. Int. J. Food Microbiol. 381:109889. doi: 10.1016/j.ijfoodmicro.2022.109889, PMID: [DOI] [PubMed] [Google Scholar]
- Church F. C., Swaisgood H. E., Porter D. H., Catignani G. L. (1983). Spectrophotometric assay using o-phthaldialdehyde for determination of proteolysis in milk and isolated milk Proteins1. J. Dairy Sci. 66, 1219–1227. doi: 10.3168/jds.S0022-0302(83)81926-2 [DOI] [Google Scholar]
- Clapp A. R., Medintz I. L., Mauro J. M., Fisher B. R., Bawendi M. G., Mattoussi H. (2004). Fluorescence resonance energy transfer between quantum dot donors and dye-labeled protein acceptors. J. Am. Chem. Soc. 126, 301–310. doi: 10.1021/ja037088b, PMID: [DOI] [PubMed] [Google Scholar]
- Cupi D., Thorsen M., Elvig-Jørgensen S. G., Wulf-Andersen L., Berti-Sorbara J., Cowieson A. J., et al. (2022). Efficacy and safety profile of a subtilisin protease produced by fermentation in Bacillus licheniformis to be used as a feed additive. Heliyon 8:e10030. doi: 10.1016/j.heliyon.2022.e10030, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Damare S., Raghukumar C., Muraleedharan U. D., Raghukumar S. (2006). Deep-sea fungi as a source of alkaline and cold-tolerant proteases. Enzym. Microb. Technol. 39, 172–181. doi: 10.1016/j.enzmictec.2006.03.032 [DOI] [Google Scholar]
- David A., Singh Chauhan P., Kumar A., Angural S., Kumar D., Puri N., et al. (2018). Coproduction of protease and mannanase from Bacillus nealsonii PN-11 in solid state fermentation and their combined application as detergent additives. Int. J. Biol. Macromol. 108, 1176–1184. doi: 10.1016/j.ijbiomac.2017.09.037, PMID: [DOI] [PubMed] [Google Scholar]
- De Azeredo L. A., Leite S. G., Freire D. M., Benchetrit L., Coelho R. R. (2001). Proteases from actinomycetes interfere in solid media plate assays of hyaluronidase activity. J. Microbiol. Methods 45, 207–212. doi: 10.1016/s0167-7012(01)00251-2, PMID: [DOI] [PubMed] [Google Scholar]
- de Castro R. J. S., Sato H. H. (2014). Advantages of an acid protease from Aspergillus oryzae over commercial preparations for production of whey protein hydrolysates with antioxidant activities. Biocatal. Agric. Biotechnol. 3, 58–65. doi: 10.1016/j.bcab.2013.11.012 [DOI] [Google Scholar]
- de Matos F. M., de Lacerda J., Gomes T. J., Zanetti G., de Castro R., Soares J. (2022). Production of black cricket protein hydrolysates with α-amylase, α-glucosidase and angiotensin I-converting enzyme inhibitory activities using a mixture of proteases. Biocatal. Agric. Biotechnol. 39:102276. doi: 10.1016/j.bcab.2022.102276 [DOI] [Google Scholar]
- Demain A. L., Vaishnav P. (2009). Production of recombinant proteins by microbes and higher organisms. Biotechnol. Adv. 27, 297–306. doi: 10.1016/j.biotechadv.2009.01.008, PMID: [DOI] [PubMed] [Google Scholar]
- Devanthi P. V. P., Gkatzionis K. (2019). Soy sauce fermentation: microorganisms, aroma formation, and process modification. Food Res. Int. 120, 364–374. doi: 10.1016/j.foodres.2019.03.010, PMID: [DOI] [PubMed] [Google Scholar]
- Díaz-García M. E., Badía-Laíño R. (2018). Fluorescence/Fluorescence Labeling. Reference Module in Chemistry, Molecular Sciences and Chemical Engineering.
- Ding S., Mao B., Lu X., Zhuge B., Zong H. (2022). Efficient production and biochemical characterization of a thermostable carboxypeptidase from Bacillus megaterium and its application on flavor improvement of soy isolate protein hydrolysates. Eur. Food Res. Technol. 248, 2135–2143. doi: 10.1007/s00217-022-04036-5 [DOI] [Google Scholar]
- Doan C. T., Tran T. N., Nguyen V. B., Vo T. P. K., Nguyen A. D., Wang S. L. (2019). Chitin extraction from shrimp waste by liquid fermentation using an alkaline protease-producing strain, Brevibacillus parabrevis. Int. J. Biol. Macromol. 131, 706–715. doi: 10.1016/j.ijbiomac.2019.03.117, PMID: [DOI] [PubMed] [Google Scholar]
- Du X., Jing H., Wang L., Huang X., Wang X., Wang H. (2022). Characterization of structure, physicochemical properties, and hypoglycemic activity of goat milk whey protein hydrolysate processed with different proteases. Food. Sci. Technol. 159:113257. doi: 10.1016/j.lwt.2022.113257 [DOI] [Google Scholar]
- Edens L., Dekker P., van der Hoeven R., Deen F., de Roos A., Floris R. (2005). Extracellular prolyl endoprotease from Aspergillus niger and its use in the debittering of protein hydrolysates. J. Agric. Food Chem. 53, 7950–7957. doi: 10.1021/jf050652c, PMID: [DOI] [PubMed] [Google Scholar]
- Ehren J., Morón B., Martin E., Bethune M. T., Gray G. M., Khosla C. (2009). A food-grade enzyme preparation with modest gluten detoxification properties. PLoS One 4:e6313. doi: 10.1371/journal.pone.0006313, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eijsink V. G. H., van der Zee R., van den Burg B., Vriend G., Venema G. (1991). Improving the thermostability of the neutral protease of Bacillus stearothermophilus by replacing a buried asparagine by leucine. FEBS Lett. 282, 13–16. doi: 10.1016/0014-5793(91)80434-5, PMID: [DOI] [PubMed] [Google Scholar]
- El-Ghonemy D. H., Ali T. H. (2021). Effective bioconversion of feather-waste keratin by thermo-surfactant stable alkaline keratinase produced from Aspergillus sp. DHE7 with promising biotechnological application in detergent formulations. Biocatal. Agric. Biotechnol. 35:102052. doi: 10.1016/j.bcab.2021.102052 [DOI] [Google Scholar]
- Elston C., Wallach J., Saulnier J. (2007). New continuous and specific fluorometric assays for Pseudomonas aeruginosa elastase and LasA protease. Anal. Biochem. 368, 87–94. doi: 10.1016/j.ab.2007.04.041, PMID: [DOI] [PubMed] [Google Scholar]
- Emran M. A., Ismail S. A., Hashem A. M. (2020). Production of detergent stable thermophilic alkaline protease by Bacillus licheniformis ALW1. Biocatal. Agric. Biotechnol. 26:101631. doi: 10.1016/j.bcab.2020.101631 [DOI] [Google Scholar]
- Escobar J., Barnett S. (1995). Synthesis of acid protease from Mucor miehei: integration of production and recovery. Process Biochem. 30, 695–700. doi: 10.1016/0032-9592(95)00005-4 [DOI] [Google Scholar]
- Espejo F. (2021). Role of commercial enzymes in wine production: a critical review of recent research. J. Food Sci. Technol. 58, 9–21. doi: 10.1007/s13197-020-04489-0, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fathi P., Moosavi-Nasab M., Mirzapour-Kouhdasht A., Khalesi M. (2021). Generation of hydrolysates from rice bran proteins using a combined ultrasonication-Alcalase hydrolysis treatment. Food Biosci. 42:101110. doi: 10.1016/j.fbio.2021.101110 [DOI] [Google Scholar]
- Feijoo-Siota L., Rama J. L. R., Sánchez-Pérez A., Villa T. G. (2018). Expression, activation and processing of a novel plant milk-clotting aspartic protease in Pichia pastoris. J. Biotechnol. 268, 28–39. doi: 10.1016/j.jbiotec.2018.01.006, PMID: [DOI] [PubMed] [Google Scholar]
- Feng T., Yan S., Hou S., Fan X. (2022). Novel fluorescence biosensor custom-made for protein tyrosine phosphatase 1B detection based on titanium dioxide-decorated single-walled carbon nanohorn nanocomposite. Spectrochim. Acta A Mol. Biomol. Spectrosc. 280:121548. doi: 10.1016/j.saa.2022.121548, PMID: [DOI] [PubMed] [Google Scholar]
- Ferreira G. A., Magliano A. C. M., Pral E. M. F., Alfieri S. C. (2009). Elastase secretion in Acanthamoeba polyphaga. Acta Trop. 112, 156–163. doi: 10.1016/j.actatropica.2009.07.015, PMID: [DOI] [PubMed] [Google Scholar]
- FitzGerald R. J., O'Cuinn G. (2006). Enzymatic debittering of food protein hydrolysates. Biotechnol. Adv. 24, 234–237. doi: 10.1016/j.biotechadv.2005.11.002, PMID: [DOI] [PubMed] [Google Scholar]
- FOC Group (2022). Industrial enzymes market worth $11.05 bn by 2029. Focus. Catal. 5:2. doi: 10.1016/J.FOCAT.2022.04.006 [DOI] [Google Scholar]
- Fu J., Li L., Yang X. Q. (2011). Specificity of carboxypeptidases from Actinomucor elegans and their debittering effect on soybean protein hydrolysates. Appl. Biochem. Biotechnol. 165, 1201–1210. doi: 10.1007/s12010-011-9338-4, PMID: [DOI] [PubMed] [Google Scholar]
- Fu Y., Liu J., Zhang W., Wæhrens S. S., Tøstesen M., Hansen E. T., et al. (2020). Exopeptidase treatment combined with maillard reaction modification of protein hydrolysates derived from porcine muscle and plasma: structure–taste relationship. Food Chem. 306:125613. doi: 10.1016/j.foodchem.2019.125613, PMID: [DOI] [PubMed] [Google Scholar]
- García-González G., Ascacio-Martínez J. Á., Hernández-Bello R., González G. M., Palma-Nicolás J. P. (2021). Expression of recombinant protease MarP from Mycobacterium tuberculosis in Pichia pastoris and its effect on human monocytes. Biotechnol. Lett. 43, 1787–1798. doi: 10.1007/s10529-021-03149-3, PMID: [DOI] [PubMed] [Google Scholar]
- Ge T., Sun Z. J., Fu S. H., Liang G. D. (2005). Cloning of thrombolytic enzyme (lumbrokinase) from earthworm and its expression in the yeast Pichia pastoris. Protein Expr. Purif. 42, 20–28. doi: 10.1016/j.pep.2005.04.005, PMID: [DOI] [PubMed] [Google Scholar]
- Goulet D. L., Fraaz U., Zulich C. J., Pilkington T. J., Siemann S. (2020). Specificity-directed design of a FRET-quenched heptapeptide for assaying thermolysin-like proteases. Anal. Biochem. 604:113826. doi: 10.1016/j.ab.2020.113826, PMID: [DOI] [PubMed] [Google Scholar]
- Gu M., Hong T., Ma Y., Xi J., Zhao Q., Xu D., et al. (2022). Effects of a commercial peptidase on rheology, microstructure, gluten properties of wheat dough and bread quality. Food Sci. Tech. 160:113266. doi: 10.1016/j.lwt.2022.113266 [DOI] [Google Scholar]
- Gu Y. Q., Walling L. L. (2002). Identification of residues critical for activity of the wound-induced leucine aminopeptidase (LAP-A) of tomato. Eur. J. Biochem. 269, 1630–1640. doi: 10.1046/j.1432-1327.2002.02795.x, PMID: [DOI] [PubMed] [Google Scholar]
- Guo J. P., Ma Y. (2008). High-level expression, purification and characterization of recombinant Aspergillus oryzae alkaline protease in Pichia pastoris. Protein Expr. Purif. 58, 301–308. doi: 10.1016/j.pep.2007.12.005, PMID: [DOI] [PubMed] [Google Scholar]
- Guo Y., Tu T., Yuan P., Wang Y., Ren Y., Yao B., et al. (2019). High-level expression and characterization of a novel aspartic protease from Talaromyces leycettanus JCM12802 and its potential application in juice clarification. Food Chem. 281, 197–203. doi: 10.1016/j.foodchem.2018.12.096, PMID: [DOI] [PubMed] [Google Scholar]
- Gupta R., Beg Q. K., Lorenz P. (2002). Bacterial alkaline proteases: molecular approaches and industrial applications. Appl. Microbiol. Biotechnol. 59, 15–32. doi: 10.1007/s00253-002-0975-y, PMID: [DOI] [PubMed] [Google Scholar]
- Ha M., El-Din Bekhit A., McConnell M., Carne A. (2022). A simple method for enrichment of [beta]-lactoglobulin from bovine milk whey involving selective hydrolysis by two fungal protease preparations. Food Chem. 368:130820. doi: 10.1016/j.foodchem.2021.130820, PMID: [DOI] [PubMed] [Google Scholar]
- Hadjidj R., Badis A., Mechri S., Eddouaouda K., Khelouia L., Annane R., et al. (2018). Purification, biochemical, and molecular characterization of novel protease from Bacillus licheniformis strain K7A. Int. J. Biol. Macromol. 114, 1033–1048. doi: 10.1016/j.ijbiomac.2018.03.167, PMID: [DOI] [PubMed] [Google Scholar]
- Hamada S., Suzuki K., Aoki N., Suzuki Y. (2013). Improvements in the qualities of gluten-free bread after using a protease obtained from Aspergillus oryzae. J. Cereal Sci. 57, 91–97. doi: 10.1016/j.jcs.2012.10.008 [DOI] [Google Scholar]
- Hamed M. B., El-Badry M. O., Kandil E. I., Borai I. H., Fahmy A. S. (2020). A contradictory action of procoagulant ficin by a fibrinolytic serine protease from Egyptian Ficus carica latex. Biotechnol. Rep. 27:e00492. doi: 10.1016/j.btre.2020.e00492, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han R., Hernández Álvarez A. J., Maycock J., Murray B. S., Boesch C. (2021). Comparison of alcalase- and pepsin-treated oilseed protein hydrolysates – experimental validation of predicted antioxidant, antihypertensive and antidiabetic properties. Curr. Res. Food Sci. 4, 141–149. doi: 10.1016/j.crfs.2021.03.001, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hao J., Zhang Z., Yang M., Zhang Y., Wu T., Liu R., et al. (2022). Micronization using combined alkaline protease hydrolysis and high-speed shearing homogenization for improving the functional properties of soy protein isolates. Biores. Bioproc. 9, 1–12. doi: 10.1186/s40643-022-00565-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hariharan S., Patti A., Arora A. (2023). Functional proteins from biovalorization of peanut meal: advances in process technology and applications. Plant Food Hum. Nutr. 78, 13–24. doi: 10.1007/s11130-022-01040-8, PMID: [DOI] [PubMed] [Google Scholar]
- Hasan M. J., Haque P., Rahman M. M. (2022). Protease enzyme based cleaner leather processing: a review. J. Clean. Prod. 365:132826. doi: 10.1016/j.jclepro.2022.132826 [DOI] [Google Scholar]
- Hassan M. A., Abol-Fotouh D., Omer A. M., Tamer T. M., Abbas E. (2020). Comprehensive insights into microbial keratinases and their implication in various biotechnological and industrial sectors: a review. Int. J. Biol. Macromol. 154, 567–583. doi: 10.1016/j.ijbiomac.2020.03.116, PMID: [DOI] [PubMed] [Google Scholar]
- Hatakeyama T., Kohzaki H., Yamasaki N. (1992). A microassay for proteases using succinylcasein as a substrate. Anal. Biochem. 204, 181–184. doi: 10.1016/0003-2697(92)90158-4, PMID: [DOI] [PubMed] [Google Scholar]
- Hejdysz M., Kaczmarek S. A., Kubiś M., Wiśniewska Z., Peris S., Budnik S., et al. (2020). The effect of protease and Bacillus licheniformis on nutritional value of pea, faba bean, yellow lupin and narrow-leaved lupin in broiler chicken diets. Br. Poult. Sci. 61, 287–293. doi: 10.1080/00071668.2020.1716303, PMID: [DOI] [PubMed] [Google Scholar]
- Hellmuth K. (2006). Industrial scale production of chymosin with Aspergillus niger. Microb. Cell Factories 5, 1–2. doi: 10.1186/1475-2859-5-S1-S31 [DOI] [Google Scholar]
- Heng X., Chen H., Lu C., Feng T., Li K., Gao E. (2022). Study on synergistic fermentation of bean dregs and soybean meal by multiple strains and proteases. Food Sci. Technol. 154:112626. doi: 10.1016/j.lwt.2021.112626 [DOI] [Google Scholar]
- Heylen E., Van Goethem S., Augustyns K., Hendriks D. (2010). Measurement of carboxypeptidase U (active thrombin-activatable fibrinolysis inhibitor) in plasma: challenges overcome by a novel selective assay. Anal. Biochem. 403, 114–116. doi: 10.1016/j.ab.2010.03.045, PMID: [DOI] [PubMed] [Google Scholar]
- Hong S., Lin Y., Dia V. P. (2022). Anti-inflammatory and antioxidant properties of hempseed protein enzymatic hydrolysates. Food. Hydrocolloids for Health. 2:100082. doi: 10.1016/j.fhfh.2022.100082 [DOI] [Google Scholar]
- Hou Y., Lu F., Tian J., Tian Y. (2019). Cloning, heterologous expression and characterization of an intracellular serine protease from Bacillus sp. LCB10. Appl. Biochem. Microbiol. 55, 482–488. doi: 10.1134/s0003683819050168 [DOI] [Google Scholar]
- Hou J., Yin X. M., Li Y., Han D., Lü B., Zhang J. Y., et al. (2021). Biochemical characterization of a low salt-adapted extracellular protease from the extremely halophilic archaeon Halococcus salifodinae. Int. J. Biol. Macromol. 176, 253–259. doi: 10.1016/j.ijbiomac.2021.02.081, PMID: [DOI] [PubMed] [Google Scholar]
- Ichishima E. (2004). “Aspergillopepsin I” in The handbook of proteolytic enzymes. eds. Barrett A. J., Rawlings N. D., Woessner J. F. (Salt Lake City, UT: Academic Press; ), 92–99. [Google Scholar]
- Ikemura H., Takagi H., Inouye M. (1987). Requirement of pro-sequence for the production of active subtilisin E in Escherichia coli. J. Biol. Chem. 262, 7859–7864. doi: 10.1016/S0021-9258(18)47646-6, PMID: [DOI] [PubMed] [Google Scholar]
- Inouye K., Kusano M., Hashida Y., Minoda M., Yasukawa K. (2007). Engineering, expression, purification, and production of recombinant thermolysin. Biotechnol. Annu. Rev. 13, 43–64. doi: 10.1016/s1387-2656(07)13003-9, PMID: [DOI] [PubMed] [Google Scholar]
- International Union of Biochemistry (1992). Enzyme nomenclature. New York: Academic Press. [Google Scholar]
- Isabel T. F., Costa G. N. M., Pacheco I. B., Barbosa L. G., Santos-Junior C. D., Fonseca F. P. P., et al. (2016). Expression and partial biochemical characterization of a recombinant serine protease from Bothrops pauloensis snake venom. Toxicon 115, 49–54. doi: 10.1016/j.toxicon.2016.03.002, PMID: [DOI] [PubMed] [Google Scholar]
- Jellouli K., Ghorbel-Bellaaj O., Ayed H. B., Manni L., Agrebi R., Nasri M. (2011). Alkaline-protease from Bacillus licheniformis MP1: purification, characterization and potential application as a detergent additive and for shrimp waste deproteinization. Process Biochem. 46, 1248–1256. doi: 10.1016/j.procbio.2011.02.012 [DOI] [Google Scholar]
- Jisha V. N., Smitha R. B., Pradeep S., Sreedevi S., Unni K. N., Sajith S., et al. (2013). Versatility of microbial proteases. Adv. Enzy. Res. 1, 39–51. doi: 10.4236/aer.2013.13005 [DOI] [Google Scholar]
- Jo H. D., Kwon G. H., Park J. Y., Cha J., Song Y. S., Kim J. H. (2011). Cloning and overexpression of aprE3-17 encoding the major fibrinolytic protease of Bacillus licheniformis CH 3-17. Biotechnol. Biopro. E 16, 352–359. doi: 10.1007/s12257-010-0328-0 [DOI] [Google Scholar]
- Johri B. N., Jain S., Chouhan S. (1985). Enzymes from Thermophilic fungi: proteases and lipases. Proc. Ind. Acad. Sci. 94, 175–196. doi: 10.1007/bf03053136 [DOI] [Google Scholar]
- Jones L. J., Upson R. H., Haugland R. P., Panchuk-Voloshina N., Zhou M., Haugland R. P. (1997). Quenched BODIPY dye-labeled casein substrates for the assay of protease activity by direct fluorescence measurement. Anal. Biochem. 251, 144–152. doi: 10.1006/abio.1997.2259, PMID: [DOI] [PubMed] [Google Scholar]
- Joo H. S., Ra K. S., Park H. S., Choi J. W. (2013). Molecular cloning and functional expression of a fibrinolytic protease gene from the polychaeta, Periserrula leucophryna. Biotechnol. Biopro. E 18, 209–217. doi: 10.1007/s12257-012-0800-0 [DOI] [Google Scholar]
- Juárez-Montiel M., Ibarra J. A., Chávez-Camarillo G., Hernández-Rodríguez C., Villa-Tanaca L. (2014). Molecular cloning and heterologous expression in Pichia pastoris of X-prolyl-dipeptidyl aminopeptidase from Basidiomycete ustilago maydis. Appl. Biochem. Biotechnol. 172, 2530–2539. doi: 10.1007/s12010-013-0682-4, PMID: [DOI] [PubMed] [Google Scholar]
- Kalisz H. M. (1988). “Microbial proteinases” in Enzyme studies. Advances in biochemical engineering/biotechnology. ed. Fiechter A. (New York: Springer, Berlin, Heidelberg; ), 1–65. [DOI] [PubMed] [Google Scholar]
- Kang X., Li C., Ding W., Ma Y., Gao S., Zhou X., et al. (2023). Optimization of operating conditions in the biological enzymes for efficient waste activated sludge dewatering. Proc. Saf. Environ. 170, 545–552. doi: 10.1016/j.psep.2022.12.046 [DOI] [Google Scholar]
- Kang C., Yu X. W., Xu Y. (2014). Cloning and expression of a novel prolyl endopeptidase from Aspergillus oryzae and its application in beer stabilization. J. Ind. Microbiol. Biotechnol. 42, 263–272. doi: 10.1007/s10295-014-1571-8, PMID: [DOI] [PubMed] [Google Scholar]
- Kangwa M., Salgado J. A. G., Fernandez-Lahore H. M. (2018). Identification and characterization of N-glycosylation site on a Mucor circinelloides aspartic protease expressed in Pichia pastoris: effect on secretion, activity and thermo-stability. AMB Express 8:157. doi: 10.1186/s13568-018-0691-3, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karami Z., Akbari-adergani B. (2019). Bioactive food derived peptides: a review on correlation between structure of bioactive peptides and their functional properties. J. Food Sci. Technol. 56, 535–547. doi: 10.1007/s13197-018-3549-4, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kasana R. C., Salwan R., Yadav S. K. (2011). Microbial proteases: detection, production, and genetic improvement. Crit. Rev. Microbiol. 37, 262–276. doi: 10.3109/1040841X.2011.577029, PMID: [DOI] [PubMed] [Google Scholar]
- Katrolia P., Liu X., Zhao Y., Kumar Kopparapu N., Zheng X. (2019). Gene cloning, expression and homology modeling of first fibrinolytic enzyme from mushroom (Cordyceps militaris). Int. J. Biol. Macromol. 146, 897–906. doi: 10.1016/j.ijbiomac.2019.09.212, PMID: [DOI] [PubMed] [Google Scholar]
- Ke Y., Wei M., Fu Y., Zhu Y., Zhan X. (2019). Enzymatic characteristics of a recombinant protease (rPepD) from Aspergillus niger expressed in Pichia pastoris. Protein Expr. Purif. 162, 67–71. doi: 10.1016/j.pep.2019.06.002, PMID: [DOI] [PubMed] [Google Scholar]
- Ke Y., Yuan X., Li J., Zhou W., Huang X., Wang T. (2018). High-level expression, purification, and enzymatic characterization of a recombinant Aspergillus sojae alkaline protease in Pichia pastoris. Protein Expr. Purif. 148, 24–29. doi: 10.1016/j.pep.2018.03.009, PMID: [DOI] [PubMed] [Google Scholar]
- Khan A. R., James M. N. G. (1998). Molecular mechanisms for the conversion of zymogens to active proteolytic enzymes. Protein Sci. 7, 815–836. doi: 10.1002/pro.5560070401, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim S. Y., Jeong E. J., Song K. J., Park K. S. (2009). Molecular cloning and characterization of a serine protease-like protein from silkworm (Bombyx mori). Gen. Genom. 31, 387–395. doi: 10.1007/bf03191257 [DOI] [Google Scholar]
- Kim T., Lei X. G. (2005). Expression and characterization of a thermostable serine protease (TfpA) from Thermomonospora fusca YX in Pichia pastoris. Appl. Microbiol. Biotechnol. 68, 355–359. doi: 10.1007/s00253-005-1911-8, PMID: [DOI] [PubMed] [Google Scholar]
- Kumar N. S., Devi P. S. S., Nair A. S. (2016). A review on microbial proteases. Int. J. Adv. Res. 4, 2048–2053. doi: 10.21474/IJAR01/1091 [DOI] [Google Scholar]
- Landbo A. K. R., Pinelo M., Vikbjerg A. F., Let M. B., Meyer A. S. (2006). Protease-assisted clarification of black currant juice: synergy with other clarifying agents and effects on the phenol content. J. Agric. Food Chem. 54, 6554–6563. doi: 10.1021/jf060008d, PMID: [DOI] [PubMed] [Google Scholar]
- Lee S. K., Hwang J. Y., Choi S. H., Kim S. M. (2010). Purification and characterization of Aspergillus oryzae LK-101 salt-tolerant acid protease isolated from soybean paste. Food Sci. Biotechnol. 19, 327–334. doi: 10.1007/s10068-010-0047-5 [DOI] [Google Scholar]
- Legare S., Heide F., Bailey-Elkin B. A., Stetefeld J. (2022). Improved SARS-CoV-2 main protease high-throughput screening assay using a 5-carboxyfluorescein substrate. J. Biol. Chem. 298:101739. doi: 10.1016/j.jbc.2022.101739, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lei H., Zhao H., Zhao M. (2013). Proteases supplementation to high gravity worts enhances fermentation performance of brewer’s yeast. Biochem. Eng. J. 77, 1–6. doi: 10.1016/j.bej.2013.04.016 [DOI] [Google Scholar]
- Li J., Chi Z., Liu Z., Yue L., Peng Y., Wang L. (2008). Cloning and characterization of a novel aspartic protease gene from marine-derived Metschnikowia reukaufii and its expression in E. coli. Appl. Biochem. Biotechnol. 159, 119–132. doi: 10.1007/s12010-008-8400-3, PMID: [DOI] [PubMed] [Google Scholar]
- Li J., Liu B., Feng X., Zhang M., Ding T., Zhao Y., et al. (2023). Comparative proteome and volatile metabolome analysis of Aspergillus oryzae 3.042 and Aspergillus sojae 3.495 during koji fermentation. Food Res. Int. 165:112527. doi: 10.1016/j.foodres.2023.112527, PMID: [DOI] [PubMed] [Google Scholar]
- Li J., Shi P., Han X., Meng K., Yang P., Wang Y., et al. (2007). Functional expression of the keratinolytic serine protease gene sfp2 from streptomyces fradiae var. k11 in Pichia pastoris. Protein Expr. Purif. 54, 79–86. doi: 10.1016/j.pep.2007.02.012, PMID: [DOI] [PubMed] [Google Scholar]
- Li X., Wang L., Jiang P., Zhu Y., Zhang W., Li R., et al. (2023). The effect of wheat bran dietary fibre and raw wheat bran on the flour and dough properties: a comparative study. Food Sci. Tech. 173:114304. doi: 10.1016/j.lwt.2022.114304 [DOI] [Google Scholar]
- Li A. N., Xie C., Zhang J., Zhang J., Li D. C. (2011). Cloning, expression, and characterization of serine protease from thermophilic fungus Thermoascus aurantiacus var. levisporus. J. Microbiol. 49, 121–129. doi: 10.1007/s12275-011-9355-6, PMID: [DOI] [PubMed] [Google Scholar]
- Li C., Zhou J., Du G., Chen J., Takahashi S., Liu S. (2020). Developing Aspergillus niger as a cell factory for food enzyme production. Biotechnol. Adv. 44:107630. doi: 10.1016/j.biotechadv.2020.107630, PMID: [DOI] [PubMed] [Google Scholar]
- Lin X., Dong L., Yu D., Wang B., Pan L. (2020). High-level expression and characterization of the thermostable leucine aminopeptidase Thelap from the thermophilic fungus Thermomyces lanuginosus in Aspergillus niger and its application in soy protein hydrolysis. Protein Expr. Purif. 167:105544. doi: 10.1016/j.pep.2019.105544, PMID: [DOI] [PubMed] [Google Scholar]
- Lin Z., Wei H., Zhang Y., Liu P., Liu Y., Huang Z., et al. (2022). Improving emulsification properties of alkaline protein extract from green tea residue by enzymatic methods. Curr. Res. Food Sci. 5, 1235–1242. doi: 10.1016/j.crfs.2022.07.016, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin S., Zhang M., Liu J., Jones G. S. (2015). Construction and application of recombinant strain for the production of an alkaline protease from Bacillus licheniformis. J. Biosci. Bioeng. 119, 284–288. doi: 10.1016/j.jbiosc.2014.08.002, PMID: [DOI] [PubMed] [Google Scholar]
- Liu Y. Q., Huang Y. Y., Deng Y. Q., Li Z. M., Lian W. T., Zhang G., et al. (2022). Effect of enzymatic hydrolysis followed after extrusion pretreatment on the structure and emulsibility of soybean protein. Process Biochem. 116, 173–184. doi: 10.1016/j.procbio.2022.03.012 [DOI] [Google Scholar]
- Liu J., Sharma A., Niewiara M. J., Singh R., Ming R., Yu Q. (2018). Papain-like cysteine proteases in Carica papaya: lineage-specific gene duplication and expansion. BMC Genomics 19:26. doi: 10.1186/s12864-017-4394-y, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Y., Shi C., Li D., Chen X., Li J., Zhang Y., et al. (2019). Engineering a highly efficient expression system to produce BcaPRO protease in Bacillus subtilis by an optimized promoter and signal peptide. Int. J. Biol. Macromol. 138, 903–911. doi: 10.1016/j.ijbiomac.2019.07.175, PMID: [DOI] [PubMed] [Google Scholar]
- Liu S. Y., Wang H. L., Zou X. T., Nie G. (2022). De novo design of an ultrasensitive fluorogenic probe for aminopeptidase N sensing in living system. Sensor. Actuat. B-Chem. 363:131828. doi: 10.1016/j.snb.2022.131828 [DOI] [Google Scholar]
- Liu B., Zhang J., Fang Z., Du G., Chen J., Liao X. (2014). Functional analysis of the C-terminal propeptide of keratinase from Bacillus licheniformis BBE11-1 and its effect on the production of keratinase in Bacillus subtilis. Process Biochem. 49, 1538–1542. doi: 10.1016/j.procbio.2014.04.021 [DOI] [Google Scholar]
- Liya S. M., Umesh M., Nag A., Chinnathambi A., Alharbi S. A., Jhanani G. K., et al. (2023). Optimized production of keratinolytic proteases from Bacillus tropicus LS27 and its application as a sustainable alternative for dehairing, destaining and metal recovery. Environ. Res. 221:115283. doi: 10.1016/j.envres.2023.115283, PMID: [DOI] [PubMed] [Google Scholar]
- Lopez M., Edens L. (2005). Effective prevention of chill-haze in beer using an acid proline-specific endoprotease from Aspergillus niger. J. Agric. Food Chem. 53, 7944–7949. doi: 10.1021/jf0506535, PMID: [DOI] [PubMed] [Google Scholar]
- Lorey S. (2002). Transcellular proteolysis demonstrated by novel cell surface-associated substrates of dipeptidyl peptidase IV (CD26). J. Biol. Chem. 277, 33170–33177. doi: 10.1074/jbc.m200798200, PMID: [DOI] [PubMed] [Google Scholar]
- Lv Y., Chen L., Liu F., Xu F., Zhong F. (2023). Improvement of the encapsulation capacity and emulsifying properties of soy protein isolate through controlled enzymatic hydrolysis. Food Hydrocoll. 138:108444. doi: 10.1016/j.foodhyd.2022.108444 [DOI] [Google Scholar]
- Ma M., Dai D., Ma P., Wu Q., Gao D., Song D. (2023). Biocompatible ratiometric fluorescent chemosensor for ultrasensitive detection of endogenous aminopeptidase N in vitro and in vivo. Sensor. Actuat. B-Chem. 379:133228. doi: 10.1016/j.snb.2022.133228 [DOI] [Google Scholar]
- Ma X., Liu Y., Li Q., Liu L., Yi L., Ma L., et al. (2016). Expression, purification and identification of a thermolysin-like protease, neutral protease I, from Aspergillus oryzae with the Pichia pastoris expression system. Protein Expr. Purif. 128, 52–59. doi: 10.1016/j.pep.2016.08.008 [DOI] [PubMed] [Google Scholar]
- Macchione M. M., Merheb C. W., Gomes E., da Silva R. (2007). Protease production by different Thermophilic fungi. Appl. Biochem. Biotechnol. 146, 223–230. doi: 10.1007/s12010-007-8034-x, PMID: [DOI] [PubMed] [Google Scholar]
- Mada S. B., Ugwu C. P., Abarshi M. M. (2019). Health promoting effects of food-derived bioactive peptides: a review. Int. J. Pept. Res. Ther. 26, 831–848. doi: 10.1007/s10989-019-09890-8 [DOI] [Google Scholar]
- Majumder R., Banik S. P., Ramrakhiani L., Khowala S. (2015). Bioremediation by alkaline protease (AkP) from edible mushroom Termitomyces clypeatus: optimization approach based on statistical design and characterization for diverse applications. J. Chem. Technol. Biotechnol. 90, 1886–1896. doi: 10.1002/jctb.4500 [DOI] [Google Scholar]
- Mandujano-González V., Villa-Tanaca L., Anducho-Reyes M. A., Mercado-Flores Y. (2016). Secreted fungal aspartic proteases: a review. Rev. Iberoam. Micol. 33, 76–82. doi: 10.1016/j.riam.2015.10.003 [DOI] [PubMed] [Google Scholar]
- Mansfeld J., Petermann E., Dürrschmidt P., Ulbrich-Hofmann R. (2005). The propeptide is not required to produce catalytically active neutral protease from Bacillus stearothermophilus. Protein Expr. Purif. 39, 219–228. doi: 10.1016/j.pep.2004.10.008 [DOI] [PubMed] [Google Scholar]
- Marangon M., Van Sluyter S. C., Robinson E. M. C., Muhlack R. A., Holt H. E., Haynes P. A., et al. (2012). Degradation of white wine haze proteins by aspergillopepsin I and II during juice flash pasteurization. Food Chem. 135, 1157–1165. doi: 10.1016/j.foodchem.2012.05.042, PMID: [DOI] [PubMed] [Google Scholar]
- Marie-Claire C., Roques B. P., Beaumont A. (1998). Intramolecular processing of prothermolysin. J. Biol. Chem. 273, 5697–5701. doi: 10.1074/jbc.273.10.5697, PMID: [DOI] [PubMed] [Google Scholar]
- Marson G. V., Lacour S., Hubinger M. D., Belleville M. P. (2022). Serial fractionation of spent brewer’s yeast protein hydrolysate by ultrafiltration: a peptide-rich product with low RNA content. J. Food Eng. 312:110737. doi: 10.1016/j.jfoodeng.2021.110737 [DOI] [Google Scholar]
- Masi C., Gemechu G., Tafesse M. (2021). Isolation, screening, characterization, and identification of alkaline protease-producing bacteria from leather industry effluent. Ann. Microbiol. 71, 1–11. doi: 10.1186/s13213-021-01631-x [DOI] [Google Scholar]
- Masilamani R., Natarajan S. (2015). Molecular cloning, overexpression and characterization of a new thiol-dependent, alkaline serine protease with destaining function and fibrinolytic potential from Marinobacter aquaeolei MS2-1. Biologia 70, 1143–1149. doi: 10.1515/biolog-2015-0144 [DOI] [Google Scholar]
- Mathew Z., Knox T. M., Miller C. G. (2000). Salmonella enterica serovar typhimurium peptidase B is a leucyl aminopeptidase with specificity for acidic amino acids. J. Bacteriol. 182, 3383–3393. doi: 10.1128/JB.182.12.3383-3393.2000, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matkawala F., Nighojkar S., Kumar A., Nighojkar A. (2019). Enhanced production of alkaline protease by Neocosmospora sp. N1 using custard apple seed powder as inducer and its application for stain removal and dehairing. Biocat. Agric. Biotech. 21:101310. doi: 10.1016/j.bcab.2019.101310 [DOI] [Google Scholar]
- Matoba T., Hata T. (1972). Relationship between bitterness of peptides and their chemical structures. Agric. Biol. Chem. 36, 1423–1431. doi: 10.1271/bbb1961.36.1423 [DOI] [Google Scholar]
- Mazotto A. M., de Melo A. C. N., Macrae A., Rosado A. S., Peixoto R., Cedrola S. M. L., et al. (2010). Biodegradation of feather waste by extracellular keratinases and gelatinases from Bacillus spp. World J. Microbiol. Biotechnol. 27, 1355–1365. doi: 10.1007/s11274-010-0586-1 [DOI] [PubMed] [Google Scholar]
- Mcdonald C. E., Chen L. L. (1965). The Lowry modification of the folin reagent for determination of proteinase activity. Anal. Biochem. 10, 175–177. doi: 10.1016/0003-2697(65)90255-1, PMID: [DOI] [PubMed] [Google Scholar]
- Mechri S., Zaraî Jaouadi N., Bouacem K., Allala F., Bouraoui A., Ferard C., et al. (2021). Cloning and heterologous expression of subtilisin SAPN, a serine alkaline protease from Melghiribacillus thermohalophilus Nari2AT in Escherichia coli and Pichia pastoris. Process Biochem. 105, 27–41. doi: 10.1016/j.procbio.2021.03.020 [DOI] [Google Scholar]
- Meng X., He Z. F., Li H. J. (2013). Purification and characterization of a novel skatole-degrading protease from Lactobacillus brevis 1.12. Food Sci. Biotechnol. 22, 1–7. doi: 10.1007/s10068-013-0224-4 [DOI] [Google Scholar]
- Meng K., Li J., Cao Y., Shi P., Wu B., Han X., et al. (2007). Gene cloning and heterologous expression of a serine protease from Streptomyces fradiae var.k11. Can. J. Microbiol. 53, 186–195. doi: 10.1139/w06-122, PMID: [DOI] [PubMed] [Google Scholar]
- Meyer M., Leptihn S., Welz M., Schaller A. (2016). Functional characterization of propeptides in plant subtilases as intramolecular chaperones and inhibitors of the mature protease. J. Biol. Chem. 291, 19449–19461. doi: 10.1074/jbc.M116.744151, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mhamdi S., Bkhairia I., Nasri R., Mechichi T., Nasri M., Kamoun A. S. (2017a). Evaluation of the biotechnological potential of a novel purified protease BS1 from Bacillus safensis S406 on the chitin extraction and detergent formulation. Int. J. Biol. Macromol. 104, 739–747. doi: 10.1016/j.ijbiomac.2017.06.062, PMID: [DOI] [PubMed] [Google Scholar]
- Mhamdi S., Ktari N., Hajji S., Nasri M., Sellami Kamoun A. (2017b). Alkaline proteases from a newly isolated Micromonospora chaiyaphumensis S103: characterization and application as a detergent additive and for chitin extraction from shrimp shell waste. Int. J. Biol. Macromol. 94, 415–422. doi: 10.1016/j.ijbiomac.2016.10.036, PMID: [DOI] [PubMed] [Google Scholar]
- Mikołajczak B., Iwańska E., Spychaj A., Danyluk B., Montowska M., Grześ B., et al. (2019). An analysis of the influence of various tenderizing treatments on the tenderness of meat from Polish Holstein-Friesian bulls and the course of changes in collagen. Meat Sci. 158:107906. doi: 10.1016/j.meatsci.2019.107906, PMID: [DOI] [PubMed] [Google Scholar]
- Mirzapour-Kouhdasht A., Moosavi-Nasab M., Yousefi R., Eun J. (2021). Bio/multi-functional peptides derived from fish gelatin hydrolysates: technological and functional properties. Biocatal. Agric. Biotechnol. 36:102152. doi: 10.1016/j.bcab.2021.102152 [DOI] [Google Scholar]
- Mokashe N., Chaudhari B., Patil U. (2018). Operative utility of salt-stable proteases of halophilic and halotolerant bacteria in the biotechnology sector. Int. J. Biol. Macromol. 117, 493–522. doi: 10.1016/j.ijbiomac.2018.05.217, PMID: [DOI] [PubMed] [Google Scholar]
- Moore S., Stein W. H. (1948). Photometric ninhydrin method for use in the chromatography of amino acids. J. Biol. Chem. 176, 367–388. doi: 10.1016/S0021-9258(18)51034-6, PMID: [DOI] [PubMed] [Google Scholar]
- Mortezaei M., Dadmehr M., Korouzhdehi B., Hakimi M., Ramshini H. (2021). Colorimetric and label free detection of gelatinase positive bacteria and gelatinase activity based on aggregation and dissolution of gold nanoparticles. J. Microbiol. Methods 191:106349. doi: 10.1016/j.mimet.2021.106349, PMID: [DOI] [PubMed] [Google Scholar]
- Mubeen A., Nazim H., Zulqarnain B., Muhammad B., Ajay K., Luiz F. R. F., et al. (2023). “Bioprospecting microbial proteases in various industries/sectors,” in Microbial biomolecules. eds. Kumar Ajay, Bilal Muhammad, Kumari Madhuree, et al. (Salt Lake City, UT: Academic Press; ), 301–324. [Google Scholar]
- Mukhia S., Kumar A., Kumar R. (2021). Generation of antioxidant peptides from soy protein isolate through psychrotrophic Chryseobacterium sp. derived alkaline broad temperature active protease. Food Sci. Technol. 143:111152. doi: 10.1016/j.lwt.2021.111152 [DOI] [Google Scholar]
- Murthy P. S., Kusumoto K. (2015). Acid protease production by Aspergillus oryzae on potato pulp powder with emphasis on glycine releasing activity: a benefit to the food industry. Food Bioprod. Process. 96, 180–188. doi: 10.1016/j.fbp.2015.07.013 [DOI] [Google Scholar]
- Nakamura R., Saito M., Maruyama M., Yamanaka S., Tanemura T., Watanabe M., et al. (2023). Reduction in the bitterness of protein hydrolysates by an aminopeptidase from Aspergillus oryzae. Food Sci. Technol. Res. 29, 71–77. doi: 10.3136/fstr.FSTR-D-22-00110 [DOI] [Google Scholar]
- Nandan A., Nampoothiri K. M. (2020). Therapeutic and biotechnological applications of substrate specific microbial aminopeptidases. Appl. Microbiol. Biotechnol. 104, 5243–5257. doi: 10.1007/s00253-020-10641-9, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naveed M., Nadeem F., Mehmood T., Bilal M., Anwar Z., Amjad F. (2021). Protease-a versatile and ecofriendly biocatalyst with multi-industrial applications: an updated review. Catal. Lett. 151, 307–323. doi: 10.1007/s10562-020-03316-7 [DOI] [Google Scholar]
- Németh B. Z., Demcsák A., Micsonai A., Kiss B., Schlosser G., Geisz A., et al. (2022). Arg236 in human chymotrypsin B2 (CTRB2) is a key determinant of high enzyme activity, trypsinogen degradation capacity, and protection against pancreatitis. BBA-Biomemb. Prot. Prot. 1870:140831. doi: 10.1016/j.bbapap.2022.140831 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nguyen T. T., Quyen T. D., Le H. T. (2013). Cloning and enhancing production of a detergent- and organic-solvent-resistant nattokinase from Bacillus subtilis VTCC-DVN-12-01 by using an eight-protease-gene-deficient Bacillus subtilis WB800. Microb. Cell Factories 12:79. doi: 10.1186/1475-2859-12-79 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nikinmaa M., Mattila O., Holopainen-Mantila U., Heiniö R., Nordlund E. (2019). Impact of lactic acid bacteria starter cultures and hydrolytic enzymes on the characteristics of wholegrain crackers. J. Cereal Sci. 88, 1–8. doi: 10.1016/j.jcs.2019.04.016 [DOI] [Google Scholar]
- Nishiwaki T., Yoshimizu S., Furuta M., Hayashi K. (2002). Debittering of enzymatic hydrolysates using an aminopeptidase from the edible basidiomycete Grifola frondosa. J. Biosci. Bioeng. 93, 60–63. doi: 10.1016/S1389-1723(02)80055-X, PMID: [DOI] [PubMed] [Google Scholar]
- Niu C., Xing X., Yang X., Zheng F., Liu C., Wang J., et al. (2023). Isolation, identification and application of Aspergillus oryzae BL18 with high protease activity as starter culture in doubanjiang (broad bean paste) fermentation. Food Biosci. 51:102225. doi: 10.1016/j.fbio.2022.102225 [DOI] [Google Scholar]
- Nnolim N. E., Okoh A. I., Nwodo U. U. (2020). Proteolytic bacteria isolated from agro-waste dumpsites produced keratinolytic enzymes. Biotechnol. Rep. 27:e00483. doi: 10.1016/j.btre.2020.e00483, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noman A., Wang Y., Zhang C., Yin L., Abed S. M. (2022). Fractionation and purification of antioxidant peptides from chinese sturgeon (Acipenser sinensis) protein hydrolysates prepared using papain and alcalase 2.4L. Arab. J. Chem. 15:104368. doi: 10.1016/j.arabjc.2022.104368 [DOI] [Google Scholar]
- Novelli P. K., Barros M. M., Fleuri L. F. (2016). Novel inexpensive fungi proteases: production by solid state fermentation and characterization. Food Chem. 198, 119–124. doi: 10.1016/j.foodchem.2015.11.089, PMID: [DOI] [PubMed] [Google Scholar]
- Ogino H., Otsubo T., Ishikawa H. (2008). Screening, purification, and characterization of a leather-degrading protease. Biochem. Eng. J. 38, 234–240. doi: 10.1016/j.bej.2007.07.008, PMID: 37284689 [DOI] [Google Scholar]
- Oliveira V., Campos M., Melo R. L., Ferro E. S., Camargo A. C. M., Juliano M. A., et al. (2001). Substrate specificity characterization of recombinant metallo oligo-peptidases thimet oligopeptidase and neurolysin. Biochemistry 40, 4417–4425. doi: 10.1021/bi002715k [DOI] [PubMed] [Google Scholar]
- Onifade A. A., Al-Sane N. A., Al-Musallam A. A., Al-Zarban S. (1998). A review: potentials for biotechnological applications of keratin-degrading microorganisms and their enzymes for nutritional improvement of feathers and other keratins as livestock feed resources. Bioresour. Technol. 66, 1–11. doi: 10.1016/S0960-8524(98)00033-9 [DOI] [Google Scholar]
- Pan L., Peng Q., Li W., Yan C., Li Z., You S., et al. (2022). Antioxidant peptides derived from mulberry seed protein by ionic liquid-enhanced microfluidic hydrolysis with immobilized protease. Biomass Conver. Bior. 12, 4435–4447. doi: 10.1007/s13399-022-02410-7 [DOI] [Google Scholar]
- Pang H., Jiao Q., He J., Zhang Z., Wang L., Yan Z., et al. (2022). Enhanced short-chain fatty acids production through a short-term anaerobic fermentation of waste activated sludge: synergistic pretreatment of alkali and alkaline hydrolase blend. J. Clean. Prod. 342:130954. doi: 10.1016/j.jclepro.2022.130954 [DOI] [Google Scholar]
- Pang H., Pan X., Li L., He J., Zheng Y., Qu F., et al. (2020). An innovative alkaline protease-based pretreatment approach for enhanced short-chain fatty acids production via a short-term anaerobic fermentation of waste activated sludge. Bioresour. Technol. 312:123397. doi: 10.1016/j.biortech.2020.123397 [DOI] [PubMed] [Google Scholar]
- Park S., Lee J. J., Yang B. M., Cho J. H., Kim S., Kang J., et al. (2020). Dietary protease improves growth performance, nutrient digestibility, and intestinal morphology of weaned pigs. J. Anim. Sci Technol. 62, 21–30. doi: 10.5187/jast.2020.62.1.21, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pati S., Samantaray D. P. (2022). “Enzymes in brewing and wine industries,” in Novel food grade enzymes. eds. Dutt Tripathi A., Darani K. K., Srivastava S. K. (Singapore: Springer; ), 165–181. [Google Scholar]
- Paul T., Jana A., Mandal A. K., Mandal A., Das Mohpatra P. K., Mondal K. C. (2016). Bacterial keratinolytic protease, imminent starter for NextGen leather and detergent industries. Sustain. Chem. Pharm. 3, 8–22. doi: 10.1016/j.scp.2016.01.001 [DOI] [Google Scholar]
- Peng Y., Yang X. J., Xiao L., Zhang Y. Z. (2004). Cloning and expression of a fibrinolytic enzyme (subtilisin DFE) gene from Bacillus amyloliquefaciens DC-4 in Bacillus subtilis. Res. Microbiol. 155, 167–173. doi: 10.1016/j.resmic.2003.10.004 [DOI] [PubMed] [Google Scholar]
- Pereira W. E. S., da Silva R. R., de Amo G. S., Ruller R., Kishi L. T., Boscolo M., et al. (2020). A collagenolytic aspartic protease from Thermomucor indicae-seudaticae expressed in Escherichia coli and Pichia pastoris. Appl. Biochem. Biotechnol. 191, 1258–1270. doi: 10.1007/s12010-020-03292-z, PMID: [DOI] [PubMed] [Google Scholar]
- Pereira J. Q., Lopes F. C., Petry M. V., Medina L. F. D. C., Brandelli A. (2014). Isolation of three novel Antarctic psychrotolerant feather-degrading bacteria and partial purification of keratinolytic enzyme from Lysobacter sp. A03. Int. Biodeterior. Biodegradation 88, 1–7. doi: 10.1016/j.ibiod.2013.11.012 [DOI] [Google Scholar]
- Pérez-Gálvez R., Almécija M. C., Espejo F. J., Guadix E. M., Guadix A. (2011). Bi-objective optimization of the enzymatic hydrolysis of porcine blood protein. Biochem. Eng. J. 53, 305–310. doi: 10.1016/j.bej.2010.12.004 [DOI] [Google Scholar]
- Philipps-Wiemann P. (2018). “Proteases—animal feed,” in Enzymes in human and animal nutrition. eds. Simões Nunes Carlos, Kumar Vikas. (Salt Lake City, UT: Academic Press; ), 279–297. [Google Scholar]
- Pinelo M., Zeuner B., Meyer A. S. (2010). Juice clarification by protease and pectinase treatments indicates new roles of pectin and protein in cherry juice turbidity. Food Bioprod. Process. 88, 259–265. doi: 10.1016/j.fbp.2009.03.005 [DOI] [Google Scholar]
- Porfírio C. T. M. N., Souza P. F. N., Ramos M. V., Campos F. A. P., Freitas S. F., Oliveira J. P. B., et al. (2022). Serine carboxypeptidases from the carnivorous plant nepenthes mirabilis: partial characterization and heterologous expression. Int. J. Biol. Macromol. 198, 77–86. doi: 10.1016/j.ijbiomac.2021.12.104, PMID: [DOI] [PubMed] [Google Scholar]
- Porres J. M., Benito M. J., Lei X. G. (2002). Functional expression of keratinase(kera) gene from Bacillus licheniformis in Pichia pastoris. Biotechnol. Lett. 24, 631–636. doi: 10.1023/A:1015083007746, PMID: 23472192 [DOI] [Google Scholar]
- Prabhu N., Gajendran T., Karthikadevi S., Archana A., Arthe R. (2021). Utilization of sugarcane bagasse for enhancement production of fibrinolytic enzyme using statistical approach. Clean. Eng. Tech. 5:100269. doi: 10.1016/j.clet.2021.100269 [DOI] [Google Scholar]
- Prates J. A. M., Ribeiro A. M. R., Correia A. A. D. (2001). Role of cysteine endopeptidases (EC 3.4.22) in rabbit meat tenderisation and some related changes. Meat Sci. 57, 283–290. doi: 10.1016/S0309-1740(00)00103-0, PMID: [DOI] [PubMed] [Google Scholar]
- Promchai R., Boonchalearn A., Visessanguan W., Luxananil P. (2018). Rapid production of extracellular thermostable alkaline halophilic protease originating from an extreme haloarchaeon, Halobacterium salinarum by recombinant Bacillus subtilis. Biocatal. Agric. Biotechnol. 15, 192–198. doi: 10.1016/j.bcab.2018.06.017 [DOI] [Google Scholar]
- Pudova D. S., Vasilyeva Y. A., Sharipova M. R. (2021). Heterologous expression of Bacillus pumilus 3–19 protease in Pichia pastoris and its potential use as a feed additive in poultry farming. Bionanoscience 11, 989–997. doi: 10.1007/s12668-021-00899-2 [DOI] [Google Scholar]
- Purohit M. K., Singh S. P. (2014). Cloning, over expression and functional attributes of serine proteases from Oceanobacillus iheyensis O.M.A18 and haloalkaliphilic bacterium O.M.E12. Process Biochem. 49, 61–68. doi: 10.1016/j.procbio.2013.07.009 [DOI] [Google Scholar]
- Purushothaman K., Bhat S. K., Singh S. A., Marathe G. K., Appu Rao A. R. G. (2019). Aspartic protease from Aspergillus niger: molecular characterization and interaction with pepstatin A. Int. J. Biol. Macromol. 139, 199–212. doi: 10.1016/j.ijbiomac.2019.07.133, PMID: [DOI] [PubMed] [Google Scholar]
- Qihe C., Guoqing H., Yingchun J., Hui N. (2006). Effects of elastase from a Bacillus strain on the tenderization of beef meat. Food Chem. 98, 624–629. doi: 10.1016/j.foodchem.2005.06.043 [DOI] [Google Scholar]
- Qin Q., Tang C., Wu J., Chen S., Yan Z. (2021). A dual-functional aminopeptidase from Streptomyces canus T20 and its application in the preparation of small rice peptides. Int. J. Biol. Macromol. 167, 214–222. doi: 10.1016/j.ijbiomac.2020.11.175, PMID: [DOI] [PubMed] [Google Scholar]
- Ramirez Zavala B., Mercado Flores Y., Hernandez Rodriguez C., Villa Tanaca L. (2004). Purification and characterization of a serine carboxypeptidase from Kluyveromyces marxianus. Int. J. Food Microbiol. 91, 245–252. doi: 10.1016/S0168-1605(03)00409-4, PMID: [DOI] [PubMed] [Google Scholar]
- Ran L. Y., Su H. N., Zhou M. Y., Wang L., Chen X. L., Xie B. B., et al. (2014). Characterization of a novel subtilisin-like protease myroicolsin from deep sea bacterium Myroides profundi D25 and molecular insight into its collagenolytic mechanism. J. Biol. Chem. 289, 6041–6053. doi: 10.1074/jbc.M113.513861, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rao M. B., Tanksale A. M., Ghatge M. S., Deshpande V. V. (1998). Molecular and biotechnological aspects of microbial proteases. Microbiol. Mol. Biol R 62, 597–635. doi: 10.1128/MMBR.62.3.597-635.1998, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rasaq S. A., Bahiru T., Swarna J., Amit K. J. (2023). “An overview of industrial enzymes in beverage production and processing,” in Value-addition in beverages through enzyme technology. eds. Kuddus Mohammed, Hossain Mohammad. (Salt Lake City, UT: Academic Press; ), 1–26. [Google Scholar]
- Rathod M. G., Pathak A. P. (2014). Wealth from waste: optimized alkaline protease production from agro-industrial residues by Bacillus alcalophilus LW8 and its biotechnological applications. J. Taibah. Univ. Sci. 8, 307–314. doi: 10.1016/j.jtusci.2014.04.002 [DOI] [Google Scholar]
- Rawlings N. D., Barrett A. J., Thomas P. D., Huang X., Bateman A., Finn R. D. (2017). The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucleic Acids Res. 46, D624–D632. doi: 10.1093/nar/gkx1134 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Razzaq A., Shamsi S., Ali A., Ali Q., Sajjad M., Malik A., et al. (2019). Microbial proteases applications. Front. Bioeng. Biotech. 7:110. doi: 10.3389/fbioe.2019.00110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reddy N., Deekonda V., Seshagiri S., Reddy R., Gangula A. K. (2022). Production, characterization and applications of proteases produced by Bacillus licheniformis, Acinetobacter pittii and Aspergillus niger using neem seed oil cake as the substrate. Ind. Crop. Prod. 187:115403. doi: 10.1016/j.indcrop.2022.115403 [DOI] [Google Scholar]
- Rejisha R. P., Murugan M. (2021). Alkaline protease production by halophilic Bacillus sp. strain SP II-4 and characterization with special reference to contact lens cleansing. Mat. Today Proc. 45, 1757–1760. doi: 10.1016/j.matpr.2020.08.624 [DOI] [Google Scholar]
- Rekik H., Zaraî Jaouadi N., Gargouri F., Bejar W., Frikha F., Jmal N., et al. (2019). Production, purification and biochemical characterization of a novel detergent-stable serine alkaline protease from Bacillus safensis strain RH12. Int. J. Biol. Macromol. 121, 1227–1239. doi: 10.1016/j.ijbiomac.2018.10.139, PMID: [DOI] [PubMed] [Google Scholar]
- Ren Y., Li L. (2022). The influence of protease hydrolysis of lactic acid bacteria on the fermentation induced soybean protein gel: protein molecule, peptides and amino acids. Food Res. Int. 156:111284. doi: 10.1016/j.foodres.2022.111284, PMID: [DOI] [PubMed] [Google Scholar]
- Research and Markets (2021). https://www.globenewswire.com/news-release/2021/08/05/2275265/0/en/The-Worldwide-Industrial-Enzymes-Industry-is-Expected-to-Reach-17-4-Billion-by-2027.html [Accessed February 18, 2023]
- Ribitsch D., Heumann S., Karl W., Gerlach J., Leber R., Birner-Gruenberger R., et al. (2012). Extracellular serine proteases from Stenotrophomonas maltophilia: screening, isolation and heterologous expression in E. coli. J. Biotechnol. 157, 140–147. doi: 10.1016/j.jbiotec.2011.09.025, PMID: [DOI] [PubMed] [Google Scholar]
- Rohamare S., Gaikwad S., Jones D., Bhavnani V., Pal J., Sharma R., et al. (2015). Cloning, expression and in silico studies of a serine protease from a marine actinomycete (Nocardiopsis sp. NCIM 5124). Process Biochem. 50, 378–387. doi: 10.1016/j.procbio.2014.12.025 [DOI] [Google Scholar]
- Runnels L. W., Scarlata S. F. (1995). Theory and application of fluorescence homotransfer to melittin oligomerization. Biophys. J. 69, 1569–1583. doi: 10.1016/S0006-3495(95)80030-5, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salihi A., Asoodeh A., Aliabadian M. (2017). Production and biochemical characterization of an alkaline protease from Aspergillus oryzae CH93. Int. J. Biol. Macromol. 94, 827–835. doi: 10.1016/j.ijbiomac.2016.06.023, PMID: [DOI] [PubMed] [Google Scholar]
- Sampaio Silva T. A. E., Knob A., Tremacoldi C. R., Brochetto-Braga M. R., Carmona E. C. (2011). Purification and some properties of an extracellular acid protease from Aspergillus clavatus. World J. Microbiol. Biotechnol. 27, 2491–2497. doi: 10.1007/s11274-011-0717-3 [DOI] [Google Scholar]
- Sandholt G. B., Stefansson B., Scheving R., Gudmundsdottir Á. (2018). Biochemical characterization of a native group III trypsin ZT from Atlantic cod (Gadus morhua). Int. J. Biol. Macromol. 125, 847–855. doi: 10.1016/j.ijbiomac.2018.12.099 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schacherl M., Gompert M., Pardon E., Lamkemeyer T., Steyaert J., Baumann U. (2017). Crystallographic and biochemical characterization of the dimeric architecture of site-2 protease. BBA-Biomem. 1859, 1859–1871. doi: 10.1016/j.bbamem.2017.05.006, PMID: [DOI] [PubMed] [Google Scholar]
- Schindler J., Lehmann R., Pfeiffer H., Schmid R. (1983). “Extracellular Acid Protease of Rhizopus rhizopodiformis,” in: Enzyme technology. eds. Lafferty R. M. (Heidelberg, Berlin: Springer; ), 69–77. [Google Scholar]
- Schlenzig D., Schilling S. (2017). Heterologous expression of the astacin protease meprin β in Pichia pastoris. Matrix Metal. 1579, 35–45. doi: 10.1007/978-1-4939-6863-3_3 [DOI] [PubMed] [Google Scholar]
- Schlenzig D., Wermann M., Ramsbeck D., Moenke-Wedler T., Schilling S. (2015). Expression, purification and initial characterization of human meprin β from Pichia pastoris. Protein Expr. Purif. 116, 75–81. doi: 10.1016/j.pep.2015.08.001, PMID: [DOI] [PubMed] [Google Scholar]
- Schulze A., Wermann M., Demuth H. U., Yoshimoto T., Ramsbeck D., Schlenzig D., et al. (2018). Continuous assays for meprin alpha and beta using prolyl tripeptidyl aminopeptidase (PtP) from Porphyromonas gingivalis. Anal. Biochem. 559, 11–16. doi: 10.1016/j.ab.2018.08.005, PMID: [DOI] [PubMed] [Google Scholar]
- Seker S., Beyenal H., Tanyolac A. (1999). Modeling milk clotting activity in the continuous production of microbial rennet from Mucor miehei. J. Food Sci. 64, 525–529. doi: 10.1111/j.1365-2621.1999.tb15076.x [DOI] [Google Scholar]
- Serkina A. V., Shevelev A. B., Chestukhina G. G. (2001). Structures and functions of precursors of bacterial proteases. Russ. J. Bioorg Chem. 27, 285–305. doi: 10.1023/A:1012322813107 [DOI] [Google Scholar]
- Serna-Saldivar S. O., Rubio-Flores M. (2017). “Role of intrinsic and supplemented enzymes in brewing and beer” in In the microbial enzyme technology in food applications. eds. Ray R. C., Rosell C. M. (Boca Raton, FL: CRC Press; ), 271–294. [Google Scholar]
- Shagimardanova E. I., Chastukhina I. B., Shamsutdinov T. R., Balaban N. P., Mardanova A. M., Kostrov S. V., et al. (2007). Heterologous expression of Bacillus intermedius gene of glutamyl endopeptidase in Bacillus subtilis strains defective in regulatory proteins. Microbiology 76, 569–574. doi: 10.1134/S0026261707050098, PMID: [DOI] [PubMed] [Google Scholar]
- Shaikh I. A., Turakani B., Malpani J., Goudar S. V., Mahnashi M. H., Hamed Al-Serwi R., et al. (2023). Extracellular protease production, optimization, and partial purification from Bacillus nakamurai PL4 and its applications. J. King Saud. Univ. Sci. 35:102429. doi: 10.1016/j.jksus.2022.102429 [DOI] [Google Scholar]
- Sharipova M., Balaban N., Kayumov A., Kirillova Y., Mardanova A., Gabdrakhmanova L., et al. (2008). The expression of the serine proteinase gene of Bacillus intermedius in Bacillus subtilis. Microbiol. Res. 163, 39–50. doi: 10.1016/j.micres.2006.03.003, PMID: [DOI] [PubMed] [Google Scholar]
- Sharma K. M., Kumar R., Panwar S., Kumar A. (2017). Microbial alkaline proteases: optimization of production parameters and their properties. J. Genet. Eng. Biotech. 15, 115–126. doi: 10.1016/j.jgeb.2017.02.001, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shrivastava A., Shrivastava N., Singh P. K. (2019). Chapter 34-enzymes in pharmaceutical industry. Enzy. Food Biotech. 34, 591–602. doi: 10.1016/b978-0-12-813280-7.00034-7 [DOI] [Google Scholar]
- Shu M., Shen W., Yang S., Wang X., Wang F., Wang Y., et al. (2016). High-level expression and characterization of a novel serine protease in Pichia pastoris by multi-copy integration. Enzym. Microb. Technol. 92, 56–66. doi: 10.1016/j.enzmictec.2016.06.007, PMID: [DOI] [PubMed] [Google Scholar]
- Shu L., Si X., Yang X., Ma W., Sun J., Zhang J., et al. (2020). Enhancement of acid protease activity of Aspergillus oryzae using atmospheric and room temperature plasma. Front. Microbiol. 11:1418. doi: 10.3389/fmicb.2020.01418 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siigur E., Tõnismägi K., Trummal K., Samel M., Vija H., Aaspõllu A., et al. (2011). A new tyrosine-specific chymotrypsin-like and angiotensin-degrading serine proteinase from Vipera lebetina snake venom. Biochimie 93, 321–330. doi: 10.1016/j.biochi.2010.10.004, PMID: [DOI] [PubMed] [Google Scholar]
- Silva O. S. D., Alves R. O., Porto T. S. (2018). PEG-sodium citrate aqueous two-phase systems to in situ recovery of protease from Aspergillus tamarii URM4634 by extractive fermentation. Biocatal. Agric. Biotechnol. 16, 209–216. doi: 10.1016/j.bcab.2018.08.001 [DOI] [Google Scholar]
- Singh S., Bajaj B. K. (2017). Potential application spectrum of microbial proteases for clean and green industrial production. Energy Ecol. Environ. 2, 370–386. doi: 10.1007/s40974-017-0076-5 [DOI] [Google Scholar]
- Sittipol D., Saelao P., Lohnoo T., Lerksuthirat T., Kumsang Y., Yingyong W., et al. (2019). Cloning, expression, purification and characterization of a thermo- and surfactant-stable protease from Thermomonospora curvata. Biocatal. Agric. Biotechnol. 19:101111. doi: 10.1016/j.bcab.2019.101111 [DOI] [Google Scholar]
- Skowron P. M., Krefft D., Brodzik R., Kasperkiewicz P., Drag M., Koller K. P. (2020). An alternative for proteinase K-heat-sensitive protease from Fungus Onygena corvina for biotechnology: cloning, engineering, expression, characterization and special application for protein sequencing. Microb. Cell Factories 19:135. doi: 10.1186/s12934-020-01392-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soeryapranata E., Powers J. R., Ünlü G. (2007). Cloning and characterization of debittering peptidases, PepE, PepO, PepO2, PepO3, and PepN, of Lactobacillus helveticus WSU19. Int. Dairy J. 17, 1096–1106. doi: 10.1016/j.idairyj.2007.02.002 [DOI] [Google Scholar]
- Soltani M., Sahingil D., Gokce Y., Hayaloglu A. A. (2019). Effect of blends of camel chymosin and microbial rennet (Rhizomucor miehei) on chemical composition, proteolysis and residual coagulant activity in iranian ultrafiltered white cheese. J. Food Sci. Technol. 56, 589–598. doi: 10.1007/s13197-018-3513-3, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song P., Cheng L., Tian K., Zhang M., Mchunu N. P., Niu D., et al. (2020a). Biochemical characterization of two new Aspergillus niger aspartic proteases. 3 Biotech. 10:303. doi: 10.1007/s13205-020-02292-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song P., Cheng L., Tian K., Zhang M., Singh S., Niu D., et al. (2020b). A novel aminopeptidase with potential debittering properties in casein and soybean protein hydrolysates. Food Sci. Biotechnol. 29, 1491–1499. doi: 10.1007/s10068-020-00813-8, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song P., Feng W. (2021). Functional expression and characterization of a novel aminopeptidase B from Aspergillus niger in Pichia pastoris. 3 Biotech. 11:366. doi: 10.1007/s13205-021-02915-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song P., Xu W., Wang K., Zhang Y., Wang F., Zhou X., et al. (2021a). Cloning, expression and characterization of metalloproteinase HypZn from Aspergillus niger. PLoS One 16:e0259809. doi: 10.1371/journal.pone.0259809, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song P., Xu W., Zhang Y., Wang F., Zhou X., Shi H., et al. (2021b). A new carboxypeptidase from Aspergillus niger with good thermostability, pH stability and broad substrate specificity. Sci. Rep. UK 11:18745. doi: 10.1038/s41598-021-98003-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song Y., Yan L., Jiang W., Xiao W., Feng L., Wu P., et al. (2020). Enzyme-treated soy protein supplementation in low protein diet improved flesh tenderness, juiciness, flavor, healthiness, and antioxidant capacity in on-growing grass carp (Ctenopharyngodon idella). Fish Physiol. Biochem. 46, 213–230. doi: 10.1007/s10695-019-00710-w, PMID: [DOI] [PubMed] [Google Scholar]
- Souza P. M., Aliakbarian B., Filho E. X. F., Magalhães P. O., Junior A. P., Converti A., et al. (2015). Kinetic and thermodynamic studies of a novel acid protease from aspergillus foetidus. Int. J. Biol. Macromol. 81, 17–21. doi: 10.1016/j.ijbiomac.2015.07.043, PMID: [DOI] [PubMed] [Google Scholar]
- Souza P. M., Werneck G., Aliakbarian B., Siqueira F., Ferreira Filho E. X., Perego P., et al. (2017). Production, purification and characterization of an aspartic protease from Aspergillus foetidus. Food Chem. Toxicol. 109, 1103–1110. doi: 10.1016/j.fct.2017.03.055, PMID: [DOI] [PubMed] [Google Scholar]
- Spellman D., McEvoy E., O’Cuinn G., FitzGerald R. J. (2003). Proteinase and exopeptidase hydrolysis of whey protein: comparison of the TNBS, OPA and pH stat methods for quantification of degree of hydrolysis. Int. Dairy J. 13, 447–453. doi: 10.1016/S0958-6946(03)00053-0 [DOI] [Google Scholar]
- Srivastava B., Khatri M., Singh G., Arya S. K. (2020). Microbial keratinases: an overview of biochemical characterization and its eco-friendly approach for industrial applications. J. Clean. Prod. 252:119847. doi: 10.1016/j.jclepro.2019.119847 [DOI] [Google Scholar]
- Su W., Lin C., Wu J., Li K., He G., Qian X., et al. (2006). Molecular cloning and expression of a cDNA encoding lon protease from rice (Oryza sativa). Biotechnol. Lett. 28, 923–927. doi: 10.1007/s10529-006-9022-x [DOI] [PubMed] [Google Scholar]
- Suberu Y., Akande I., Samuel T., Lawal A., Olaniran A. (2019). Cloning, expression, purification and characterization of serine alkaline protease from Bacillus subtilis RD7. Biocatal. Agric. Biotechnol. 20:101264. doi: 10.1016/j.bcab.2019.101264 [DOI] [Google Scholar]
- Sumantha A., Larroche C., Pandey A. (2006). Microbiology and industrial biotechnology of food-grade proteases: a perspective. Food Technol. Biotechnol. 44, 211–220. doi: 10.1177/1082013206063838 [DOI] [Google Scholar]
- Sun D., Wang M., Wen X., Mao S., Cheng A., Jia R., et al. (2019). Biochemical characterization of recombinant Avihepatovirus 3C protease and its localization. Virol. J. 16:54. doi: 10.1186/s12985-019-1155-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun Q., Wang X. P., Yan Q. J., Chen W., Jiang Z. Q. (2014). Purification and characterization of a chymosin from Rhizopus microsporus var. rhizopodiformis. Appl. Biochem. Biotechnol. 174, 174–185. doi: 10.1007/s12010-014-1044-6, PMID: [DOI] [PubMed] [Google Scholar]
- Sun X., Wu S., Li W., Koksel F., Du Y., Sun L., et al. (2023). The effects of cooperative fermentation by yeast and lactic acid bacteria on the dough rheology, retention and stabilization of gas cells in a whole wheat flour dough system - a review. Food Hydrocoll. 135:108212. doi: 10.1016/j.foodhyd.2022.108212 [DOI] [Google Scholar]
- Sun Q., Zhang B., Yan Q. J., Jiang Z. Q. (2016). Comparative analysis on the distribution of protease activities among fruits and vegetable resources. Food Chem. 213, 708–713. doi: 10.1016/j.foodchem.2016.07.029, PMID: [DOI] [PubMed] [Google Scholar]
- Tacias-Pascacio V. G., Morellon-Sterling R., Siar E., Tavano O., Berenguer-Murcia Á., Fernandez-Lafuente R. (2020). Use of alcalase in the production of bioactive peptides: a review. Int. J. Biol. Macromol. 165, 2143–2196. doi: 10.1016/j.ijbiomac.2020.10.060, PMID: [DOI] [PubMed] [Google Scholar]
- Takagi H., Koga M., Katsurada S., Yabuta Y., Shinde U., Inouye M., et al. (2001). Functional analysis of the propeptides of subtilisin E and aqualysin I as intramolecular chaperones. FEBS Lett. 508, 210–214. doi: 10.1016/S0014-5793(01)03053-8 [DOI] [PubMed] [Google Scholar]
- Takahashi K. (2004). “Aspergillopepsin II,” in Handbook of proteolytic enzymes (second edition). eds. Barrett Alan J., Rawlings Neil D., Fred Woessner J. (Salt Lake City, UT: Academic Press; ), 221–224. [Google Scholar]
- Takahashi K., Kagami N., Huang X. P., Kojima M., Inoue H. (1998). “Aspergillus niger acid proteinase A” in In the advances in experimental medicine and biology. ed. James M. N. G. (Boston, MA: Springer; ), 4615–5373. [DOI] [PubMed] [Google Scholar]
- Tamreihao K., Mukherjee S., Khunjamayum R., Devi L. J., Asem R. S., Ningthoujam D. S. (2018). Feather degradation by keratinolytic bacteria and biofertilizing potential for sustainable agricultural production. J. Basic Microbiol. 59, 4–13. doi: 10.1002/jobm.201800434 [DOI] [PubMed] [Google Scholar]
- Tang W., Li Z., Li C., Yu X., Wang F., Wan X., et al. (2016). High-level expression and characterization of the Bacillus subtilis subsp. subtilis str. BSP1 YwaD aminopeptidase in Pichia pastoris. Protein Expr. Purif. 122, 23–30. doi: 10.1016/j.pep.2016.02.009, PMID: [DOI] [PubMed] [Google Scholar]
- Tanwar M., Debnath M., Debnath S., Sharma P., Mukhopadhay A., Kakar N., et al. (2022). Exploring the utility of nanoprotease as environmentally friendly benign laundry detergent fabric cleaner. J. Clean. Prod. 334:130243. doi: 10.1016/j.jclepro.2021.130243 [DOI] [Google Scholar]
- Taylor G., Frommherz Y., Katikaridis P., Layer D., Sinning I., Carroni M., et al. (2022). Antibacterial peptide Cyclomarin a creates toxicity by deregulating the mycobacterium tuberculosis ClpC1–ClpP1P2 protease. J. Biol. Chem. 298:102202. doi: 10.1016/j.jbc.2022.102202, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tekin A., Uzuner U., Sezen K. (2020). Homology modeling and heterologous expression of highly alkaline subtilisin-like serine protease from Bacillus halodurans C-125. Biotechnol. Lett. 43, 479–494. doi: 10.1007/s10529-020-03025-6 [DOI] [PubMed] [Google Scholar]
- Theron L. W., Bely M., Divol B. (2018). Monitoring the impact of an aspartic protease (MpAPr1) on grape proteins and wine properties. Appl. Microbiol. Biotechnol. 102, 5173–5183. doi: 10.1007/s00253-018-8980-y, PMID: [DOI] [PubMed] [Google Scholar]
- Theron L. W., Divol B. (2014). Microbial aspartic proteases: current and potential applications in industry. Appl. Microbiol. Biotechnol. 98, 8853–8868. doi: 10.1007/s00253-014-6035-6, PMID: [DOI] [PubMed] [Google Scholar]
- Thomas A. R., Gondoza H., Hoffman L. C., Oosthuizen V., Naudé R. J. (2004). The roles of the proteasome, and cathepsins B, L, H and D, in ostrich meat tenderization. Meat Sci. 67, 113–120. doi: 10.1016/j.meatsci.2003.10.001, PMID: [DOI] [PubMed] [Google Scholar]
- Thomas K. C., Ingledew W. M. (1990). Fuel alcohol production: effects of free amino nitrogen on fermentation of very-high-gravity wheat mashes. Appl. Environ. Microbiol. 56, 2046–2050. doi: 10.1128/aem.56.7.2046-2050.1990, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson V. F., Saldaña S., Cong J., Goll D. E. (2000). A BODIPY fluorescent microplate assay for measuring activity of calpains and other proteases. Anal. Biochem. 279, 170–178. doi: 10.1006/abio.1999.4475, PMID: [DOI] [PubMed] [Google Scholar]
- Tian J., Xu Z., Long X., Tian Y., Shi B. (2019). High-expression keratinase by Bacillus subtilis SCK6 for enzymatic dehairing of goatskins. Int. J. Biol. Macromol. 135, 119–126. doi: 10.1016/j.ijbiomac.2019.05.131, PMID: [DOI] [PubMed] [Google Scholar]
- Tien T. N., Nguyen T. C., Nguyen C. N., Nguyen T. T., Pham T. A., Pham N. H., et al. (2022). Protease increases ethanol yield and decreases fermentation time in no-cook process during very-high-gravity ethanol production from rice. Process Biochem. 117, 10–18. doi: 10.1016/j.procbio.2022.03.005 [DOI] [Google Scholar]
- Tsuchiya K., Gomi K., Kitamoto K., Kumagai C., Tamura G. (1993). Secretion of calf chymosin from the filamentous fungus Aspergillus oryzae. Appl. Microbiol. Biotechnol. 40, 327–332. doi: 10.1007/BF00170388 [DOI] [PubMed] [Google Scholar]
- Twining S. S. (1984). Fluorescein isothiocyanate-labeled casein assay for proteolytic enzymes. Anal. Biochem. 143:30. doi: 10.1016/0003-2697(84)90553-0, PMID: [DOI] [PubMed] [Google Scholar]
- Van Sluyter S. C., Warnock N. I., Schmidt S., Anderson P., van Kan J. A. L., Bacic A., et al. (2013). Aspartic acid protease from Botrytis cinerea removes haze-forming proteins during white winemaking. J. Agric. Food Chem. 61, 9705–9711. doi: 10.1021/jf402762k, PMID: [DOI] [PubMed] [Google Scholar]
- Varón R., García-Moreno M., Valera-Ruipérez D., García-Molina F., García-Cánovas F., Ladrón-de Guevara R. G., et al. (2006). Kinetic analysis of a general model of activation of aspartic proteinase zymogens. J. Theor. Biol. 242, 743–754. doi: 10.1016/j.jtbi.2006.04.013, PMID: [DOI] [PubMed] [Google Scholar]
- Vishwanatha K., Appurao A., Singh S. (2009). Characterization of acid protease expressed from Aspergillus oryzae MTCC 5341. Food Chem. 114, 402–407. doi: 10.1016/j.foodchem.2008.09.070 [DOI] [Google Scholar]
- Vogelsang-O’Dwyer M., Sahin A. W., Bot F., O’Mahony J. A., Bez J., Arendt E. K., et al. (2023). Enzymatic hydrolysis of lentil protein concentrates for modification of physicochemical and techno-functional properties. Eur. Food Res. Technol. 249, 573–586. doi: 10.1007/s00217-022-04152-2 [DOI] [Google Scholar]
- Vojcic L., Pitzler C., Körfer G., Jakob F., Martinez R., Maurer K., et al. (2015). Advances in protease engineering for laundry detergents. New Biotechnol. 32, 629–634. doi: 10.1016/j.nbt.2014.12.010, PMID: [DOI] [PubMed] [Google Scholar]
- Wang P., Guo Q., Ma Y., Li S., Lu X., Zhang X., et al. (2015). DegQ regulates the production of fengycins and biofilm formation of the biocontrol agent Bacillus subtilis NCD-2. Microbiol. Res. 178, 42–50. doi: 10.1016/j.micres.2015.06.006, PMID: [DOI] [PubMed] [Google Scholar]
- Wang F., Hao J., Yang C., Sun M. (2010). Cloning, expression, and identification of a novel extracellular cold-adapted alkaline protease gene of the marine bacterium strain YS-80-122. Appl. Biochem. Biotechnol. 162, 1497–1505. doi: 10.1007/s12010-010-8927-y, PMID: [DOI] [PubMed] [Google Scholar]
- Wang Y., Li Z., Li H., Selomulya C. (2022). Effect of hydrolysis on the emulsification and antioxidant properties of plant-sourced proteins. Curr. Opin. Food Sci. 48:100949. doi: 10.1016/j.cofs.2022.100949 [DOI] [Google Scholar]
- Wang Z., Li X., Tian J., Chu Y., Tian Y. (2019). Cloning, heterologous expression and characterization of a novel streptomyces trypsin in Bacillus subtilis SCK6. Int. J. Biol. Macromol. 147, 890–897. doi: 10.1016/j.ijbiomac.2019.09.248 [DOI] [PubMed] [Google Scholar]
- Wang T., Ling H., Zhang W., Zhou Y., Li Y., Hu Y., et al. (2022). Protease or clostridium butyricum addition to a low-protein diet improves broiler growth performance. Appl. Microbiol. Biotechnol. 106, 7917–7931. doi: 10.1007/s00253-022-12264-8, PMID: [DOI] [PubMed] [Google Scholar]
- Wang H., Xu J., Liu Q., Xia X., Sun F., Kong B. (2022). Effect of the protease from Staphylococcus carnosus on the proteolysis, quality characteristics, and flavor development of Harbin dry sausage. Meat Sci. 189:108827. doi: 10.1016/j.meatsci.2022.108827, PMID: [DOI] [PubMed] [Google Scholar]
- Wang J., Xu A., Wan Y., Li Q. (2013). Purification and characterization of a new metallo-neutral protease for beer brewing from Bacillus amyloliquefaciens SYB-001. Appl. Biochem. Biotechnol. 170, 2021–2033. doi: 10.1007/s12010-013-0350-8, PMID: [DOI] [PubMed] [Google Scholar]
- Wang Y., Zhao P., Zhou Y., Hu X., Xiong H. (2023). From bitter to delicious: properties and uses of microbial aminopeptidases. World J. Microbiol. Biotechnol. 39:72. doi: 10.1007/s11274-022-03501-3 [DOI] [PubMed] [Google Scholar]
- Wani A. H., Sharma M., Salwan R., Singh G., Chahota R., Verma S. (2016). Cloning, expression, and functional characterization of serine protease Aprv2 from virulent isolate Dichelobacter nodosus of indian origin. Appl. Biochem. Biotechnol. 180, 576–587. doi: 10.1007/s12010-016-2117-5, PMID: [DOI] [PubMed] [Google Scholar]
- Ward O. P., Rao M. B., Kulkarni A. (2009). “Proteases, production”, in Encyclopedia of microbiology (third edition). ed. Schaechter Moselio. (Salt Lake City, UT: Academic Press; ), 495–511. [Google Scholar]
- Wei C., Thakur K., Liu D., Zhang J., Wei Z. (2018). Enzymatic hydrolysis of flaxseed (Linum usitatissimum L.) protein and sensory characterization of maillard reaction products. Food Chem. 263, 186–193. doi: 10.1016/j.foodchem.2018.04.120 [DOI] [PubMed] [Google Scholar]
- Wiederanders B., Kaulmann G., Schilling K. (2003). Functions of propeptide parts in cysteine proteases. Curr. Protein. Pept Sc. 4, 309–326. doi: 10.2174/1389203033487081, PMID: [DOI] [PubMed] [Google Scholar]
- Wilson S. A., Young O. A., Coolbear T., Daniel R. M. (1992). The use of proteases from extreme thermophiles for meat tenderisation. Meat Sci. 32, 93–103. doi: 10.1016/0309-1740(92)90019-Z, PMID: [DOI] [PubMed] [Google Scholar]
- Wu J., Ren L., Zhao N., Wu T., Liu R., Sui W., et al. (2022). Solid-state fermentation by Rhizopus oryzae improves flavor of wheat bran for application in food. J. Cereal Sci. 107:103536. doi: 10.1016/j.jcs.2022.103536 [DOI] [Google Scholar]
- Xie F., Feng F., Liu D., Quan S., Liu L., Zhang X., et al. (2022). Bacillus amyloliquefaciens 35 M can exclusively produce and secrete proteases when cultured in soybean-meal-based medium. Colloid Surf. B 209:112188. doi: 10.1016/j.colsurfb.2021.112188 [DOI] [PubMed] [Google Scholar]
- Xie G., Shao Z., Zong L., Li X., Cong D., Huo R. (2019). Heterologous expression and characterization of a novel subtilisin-like protease from a thermophilic Thermus thermophilus HB8. Int. J. Biol. Macromol. 138, 528–535. doi: 10.1016/j.ijbiomac.2019.07.101, PMID: [DOI] [PubMed] [Google Scholar]
- Xiong H., Yang J. Q., Kang W. M., Wu F. X., Zuo J. L., Zhou Z. Y., et al. (2018). Fluorogenic and chromogenic detection of carboxypeptidase Y with a nonpeptide-based small-molecule probe. Sensor. Actuat. B-Chem. 269, 127–134. doi: 10.1016/j.snb.2018.04.102 [DOI] [Google Scholar]
- Xu D., Guan W., Wu F., Jin Y., Yang N., Jin Z., et al. (2022). Improvement of baked wheat chips quality by protease-mediated enzymatic hydrolysis of wheat flour. Food Sci. Tech. 157:113043. doi: 10.1016/j.lwt.2021.113043 [DOI] [Google Scholar]
- Xu D., Li C., Zhao M., Fengc Y., Sun L., Wang Y. (2013). Assessment on the improvement of soy sauce fermentation by Aspergillus oryzae HG76. Biocatal. Agric. Biotechnol. 2, 344–351. doi: 10.1016/j.bcab.2013.07.002, PMID: 31807347 [DOI] [Google Scholar]
- Xu X., Pang M., Liu J., Wang Y., Wu X., Huang K., et al. (2021). Genome mining reveals the genes of carboxypeptidase for OTA-detoxification in Bacillus subtilis CW14. Int. J. Biol. Macromol. 186, 800–810. doi: 10.1016/j.ijbiomac.2021.07.085, PMID: [DOI] [PubMed] [Google Scholar]
- Yadav S. K., Bisht D., Tiwari S., Darmwal N. (2015). Purification, biochemical characterization and performance evaluation of an alkaline serine protease from Aspergillus flavus MTCC 9952 mutant. Biocatal. Agric. Biotechnol. 4, 667–677. doi: 10.1016/j.bcab.2015.08.007 [DOI] [Google Scholar]
- Yadav D. N., Mir N. A., Wadhwa R., Tushir S., Sethi S., Anurag R. K., et al. (2022). Hydrolysis of peanut (Arachis hypogea L) protein concentrate by fungal crude protease extract: effect on structural, functional and in-vitro protein digestibility. J. Food Sci. Technol. 59, 2141–2149. doi: 10.1007/s13197-021-05225-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan Z., Yuan S., Qin Q., Wu J. (2022). Enhancement of rice protein hydrolysate quality using a novel dual enzyme system. Food. Sci. Technol. 158:113110. doi: 10.1016/j.lwt.2022.113110 [DOI] [Google Scholar]
- Yang F. C., Lin I. H. (1998). Production of acid protease using thin stillage from a rice-spirit distillery by Aspergillus niger. Enzym. Microb. Technol. 23, 397–402. doi: 10.1016/S0141-0229(98)00070-2 [DOI] [Google Scholar]
- Yang X., Wang Z., Zhang C., Wang L., Pang L., Zhang D., et al. (2021). Assessment of the production of Bacillus cereus protease and its effect on the quality of ultra-high temperature-sterilized whole milk. J. Dairy Sci. 104, 6577–6587. doi: 10.3168/jds.2020-19818, PMID: [DOI] [PubMed] [Google Scholar]
- Yang H., Zhu Q., Zhou N., Tian Y. (2016). Optimized expression of prolyl aminopeptidase in Pichia pastoris and its characteristics after glycosylation. World J. Microbiol. Biotechnol. 32:176. doi: 10.1007/s11274-016-2135-z [DOI] [PubMed] [Google Scholar]
- Yokota K., Furusawa N., Abe T., Takenaka S. (1988). Application of casein agar plate method for the determination of protease activity. Eisei. Kagaku. 34, 241–247. doi: 10.1248/jhs1956.34.241, PMID: 36125759 [DOI] [Google Scholar]
- Yoo S., Han M. S. (2021). New strategy to design fluorescent substrates of carboxypeptidases using a combination of dansylated peptides and albumin. Dyes Pigments 196:109804. doi: 10.1016/j.dyepig.2021.109804 [DOI] [Google Scholar]
- Yu P., Huang X., Ren Q., Wang X. (2019). Purification and characterization of a H2O2-tolerant alkaline protease from Bacillus sp. ZJ1502, a newly isolated strain from fermented bean curd. Food Chem. 274, 510–517. doi: 10.1016/j.foodchem.2018.09.013, PMID: [DOI] [PubMed] [Google Scholar]
- Yu X., Zhai C., Zhong X., Tang W., Wang X., Yang H., et al. (2014). High-level expression and characterization of carboxypeptidase Y from Saccharomyces cerevisiae in Pichia pastoris GS115. Biotechnol. Lett. 37, 161–167. doi: 10.1007/s10529-014-1667-2 [DOI] [PubMed] [Google Scholar]
- Zadeike D., Jukonyte R., Juodeikiene G., Bartkiene E., Valatkeviciene Z. (2018). Comparative study of ciabatta crust crispness through acoustic and mechanical methods: effects of wheat malt and protease on dough rheology and crust crispness retention during storage. Food Sci. Tech. 89, 110–116. doi: 10.1016/j.lwt.2017.10.034 [DOI] [Google Scholar]
- Zhai W., Li X., Duan X., Gou C., Wang L., Gao Y. (2022). Development of a microbial protease for composting swine carcasses, optimization of its production and elucidation of its catalytic hydrolysis mechanism. BMC Biotechnol. 22:36. doi: 10.1186/s12896-022-00768-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Q., Cheng Z., Wang Y., Zheng S., Wang Y., Fu L. (2021). Combining alcalase hydrolysis and transglutaminase-cross-linking improved bitterness and techno-functional properties of hypoallergenic soybean protein hydrolysates through structural modifications. Food. Sci. Technol. 151:112096. doi: 10.1016/j.lwt.2021.112096 [DOI] [Google Scholar]
- Zhang Y., Fu Y., Zhou S., Kang L., Li C. (2013). A straightforward ninhydrin-based method for collagenase activity and inhibitor screening of collagenase using spectrophotometry. Anal. Biochem. 437, 46–48. doi: 10.1016/j.ab.2013.02.030, PMID: [DOI] [PubMed] [Google Scholar]
- Zhang N., Huang P. H., Wang P., Yu Y. Y., Zhou M., Wang Q. (2022). Combined cutinase and keratinolytic enzyme to endow improved shrink-resistance to wool fabric. Fiber Polym. 23, 985–992. doi: 10.1007/s12221-022-4445-0 [DOI] [Google Scholar]
- Zhang M., Xin X., Wu H., Zhang H. (2021). Debittering effect of partially purified proteases from soybean seedlings on soybean protein isolate hydrolysate produced by alcalase. Food Chem. 362:130190. doi: 10.1016/j.foodchem.2021.130190, PMID: [DOI] [PubMed] [Google Scholar]
- Zhang W., Yueqing C., Yuxian X. (2008). Cloning of the subtilisin Pr1A gene from a strain of locust specific fungus, Metarhizium anisopliae, and functional expression of the protein in Pichia pastoris. World J. Microbiol. Biotechnol. 24, 2481–2488. doi: 10.1007/s11274-008-9771-x [DOI] [Google Scholar]
- Zhang M., Zhao C., Du L., Lu F., Gao C. (2008). Expression, purification, and characterization of a thermophilic neutral protease from Bacillus stearothermophilus in Bacillus subtilis. Sci. China Ser. C Life Sci. 51, 52–59. doi: 10.1007/s11427-008-0009-9, PMID: [DOI] [PubMed] [Google Scholar]
- Zhao C., Ju J. (2014). Cloning, expression and activity analysis of a novel fibrinolytic serine protease from Arenicola cristata. J. Ocean U. China. 14, 533–540. doi: 10.1007/s11802-015-2488-1 [DOI] [Google Scholar]
- Zhao Y., Zhao X., Sun-Waterhouse D., Ivan Neil Waterhouse G., Zhao M., Zhang J., et al. (2020). Two-stage selective enzymatic hydrolysis generates protein hydrolysates rich in Asn-pro and ala-his for enhancing taste attributes of soy sauce. Food Chem. 345:128803. doi: 10.1016/j.foodchem.2020.128803 [DOI] [PubMed] [Google Scholar]
- Zhou K., Dong Y., Zheng H., Chen B., Mao R., Zhou L., et al. (2017). Expression, fermentation, purification and lyophilisation of recombinant Subtilisin QK in Pichia pastoris. Process Biochem. 54, 1–8. doi: 10.1016/j.procbio.2016.12.028 [DOI] [Google Scholar]
- Zhou C., Yang G., Zhang L., Zhang H., Zhou H., Lu F. (2021). Construction of an alkaline protease overproducer strain based on Bacillus licheniformis 2709 using an integrative approach. Int. J. Biol. Macromol. 193, 1449–1456. doi: 10.1016/j.ijbiomac.2021.10.208, PMID: [DOI] [PubMed] [Google Scholar]
- Zhu X., Hua Y., Li X., Kong X., Zhang C., Chen Y. (2021). Isolation and characterization of an activator-dependent protease from Aspergillus ochraceus screened from low denatured defatted soybean meal and the proteolysis of soy proteins. Food Sci. Tech. 150:112026. doi: 10.1016/j.lwt.2021.112026 [DOI] [Google Scholar]
- Zhu W., Luan H., Bu Y., Li J., Li X., Zhang Y. (2021). Changes in taste substances during fermentation of fish sauce and the correlation with protease activity. Food Res. Int. 144:110349. doi: 10.1016/j.foodres.2021.110349, PMID: [DOI] [PubMed] [Google Scholar]
- Zins M. M., Zimprich C. A., Petermann S. R., Rust L. (2001). Expression and partial characterization of an elastase from Chromobacterium violaceum. Vet. Microbiol. 80, 63–74. doi: 10.1016/s0378-1135(00)00370-9 [DOI] [PubMed] [Google Scholar]