

Abstract
Background and Aims
Plants can respond to drought by changing their relative investments in the biomass and morphology of each organ. The aims of this study were to quantify the relative contribution of changes in morphology vs. allocation and determine how they affect each other. These results should help us understand the mechanisms that plants use to respond to drought events.
Methods
In a glasshouse experiment, we applied a drought treatment (well-watered vs. drought) at early and late stages of plant growth, leading to four treatment combinations (well-watered in both early and late periods, WW; drought in the early period and well-watered in the late period, DW; well-watered in the early period and drought in the late period, WD; drought in both early and late periods, DD). We used the variance partitioning method to compare the contribution of organ (leaf and root) biomass allocation and morphology to the leaf area ratio, root length ratio and root area ratio, for the rhizomatous grass Leymus chinensis (Trin.) Tzvelev.
Key Results
Compared with the continuously well-watered treatment, the leaf area ratio, root length ratio and root area ratio showed increasing trends under various drought treatments. The contribution of leaf mass allocation to leaf area ratio differed among the drought treatments and was 2.1- to 5.3-fold greater than leaf morphology, and the contribution of root mass allocation to root length ratio was ~2-fold greater than that of root morphology. In contrast, root morphology contributed more to the root area ratio than biomass allocation under drought in both the early and late periods. There was a negative correlation between the ratio of leaf mass fraction to root mass fraction and the ratio of specific leaf area to specific root length (or specific root area).
Conclusions
This study suggested that organ biomass allocation drove a larger proportion of variation than morphological traits for the absorption of resources in this rhizomatous grass. These findings should help us understand the adaptive mechanisms of plants when they are confronted with drought stress.
Keywords: Rhizomatous grass, biomass allocation, plant functional trait, drought stress, drought timing
INTRODUCTION
The responses of plant above- and below-ground organs to changing environments can reflect the adaptive strategies of resource utilization and plant performance under environmental change (Laughlin et al., 2021; Luo et al., 2021). A shift in resource availability can trigger phenotypic changes in functional traits with the aim of mitigating stress levels and increasing uptake of limiting resources (Volaire, 2018; Zhang et al., 2021a, b).
Biomass allocation in plants is thought to reflect the distribution of ‘resources’, allowing plants to survive under changing climatic conditions (Müller et al., 2000; Weiner, 2004). Optimal partitioning theory states that plants invest more biomass into the organ that can improve access to the current limiting factor (Acuña-Míguez et al., 2022). Plants can modify their biomass allocation to different organs in response to habitat characteristics (e.g. abiotic factors), and they can even use resources efficiently when they are scarce (Sun and Wang, 2016). For example, plants can increase biomass allocation to roots or decrease allocation to leaves to maximize acquisition of water and minimize water loss under rainfall-deficient scenarios (Zhang et al., 2017; Zhang et al., 2021a). Besides, plants have been observed to grow rapidly when sufficient water is resupplied after dehydration (Wang et al., 2016). This means that plants can sometimes undergo over-compensatory growth that offsets the losses caused by the previous drought stress (Spieß et al., 2012).
Variation in plant morphological traits also plays an important role in adaptation to environment change (March‐Salas et al., 2022). For example, leaf traits such as specific leaf area (SLA) are known to increase with decreasing light availability in tree species (Bebre et al., 2022). Changes in leaf structure are also a fundamental response to drought stress, and changes to leaf area (Poorter et al., 2012) can prevent heat accumulation in leaves and lower leaf surface temperature (Dudley et al., 2014; Wright et al., 2017; Zhang et al., 2021a). Furthermore, leaf cells are generally smaller and more compact under dry conditions which can decrease overall SLA (Poorter et al., 2009; Luong et al., 2021). Reductions in SLA have been associated with slower plant growth rate, and enhanced water-use efficiency and osmotic potential due to accumulation of soluble compounds facilitating turgor maintenance (Wellstein et al., 2017). Meanwhile, plants can also lower their mean leaf size to reduce evapotranspiration through the surface of the leaves (Wyka et al., 2019). Alterations to leaf area can decrease the overall area to biomass ratio (leaf area ratio, LAR; Freschet et al., 2015a, b).
Similarly, for root traits, variation in water supply directly affects root length and branching (March-Salas et al., 2022). Constant precipitation leads to higher rooting depth and lower root branching, while water deficit generally drives increases in the length and area of roots for a given unit of plant mass [i.e. the root length ratio (RLR) and root area ratio (RAR); Ryser and Lambers, 1995] to expand root depth in order to increase water absorption (Bardgett et al., 2014; Siddiqui et al., 2021). Moreover, plants can increase the numbers of lateral roots and crown roots for water uptake from shallow soil layers (Comas et al., 2013; Li et al., 2021). This may result from an increase in the root length and area per unit root mass [i.e. the specific root length (SRL) and specific root area (SRA); Geng et al., 2014; Lozano et al., 2020] and/or the proportion of total plant mass allocated to the root [i.e. root mass fraction (RMF); Du et al., 2019].
There is a trade-off between leaf biomass allocation and morphological characteristics for resource absorption (Migliavacca et al., 2021). Plasticity in SLA has been shown to be clearly greater than in leaf mass fraction (LMF) (Poorter et al., 2012; Freschet et al., 2015a, b). For example, plants increase LAR mainly by adjusting leaf morphology (SLA) rather than leaf biomass allocation (LMF) under shading treatments (Poorter et al., 2010). Increasing attention has recently been paid to the impact of environmental change on root traits (Poorter et al., 2012; Fry et al., 2018; Lozano et al., 2020; Freschet et al., 2021; March‐Salas et al., 2022). Plant responses to resource availability, in terms of changes in morphology or/and biomass allocation, have been studied extensively (Geng et al., 2014; Freschet et al., 2015a, b; Rehling et al., 2021). However, the contribution of morphology vs. biomass allocation to resource absorption capacity (leaf area ratio, root length ratio and root area ratio) in response to environmental change was comparatively insufficient.
Despite some degree of variation in plant resource requirements under environmental changes, plants need to consume energy and substrates produced by metabolism during the process of absorbing and transporting nutrients (Elser et al., 2010; Freschet et al., 2015b). This means that acquiring resources such as CO2, water and nutrients in a way that is consistent with these basic requirements can maximize plant benefits while minimizing resource acquisition costs (Maire et al., 2013). Under drought stress, plants may employ multiple adaptation strategies that influence their resource acquisition capacities, such as adjusting their size and architecture (Zhang et al., 2021a) and changing the type and intensity of mycorrhizal associations and nutrient resorption processes (Aerts and Chapin, 1999; Remke et al., 2021). Nevertheless, a particularly critical and widespread way for plants to modulate water acquisition and transpiration is to adjust the ratio of the total surface area of leaves for transpiration to the total length of the roots exploring the soil (Waisel et al., 2002). The ratio of leaf area to root length depends not only on changes in leaf and root biomass but also on morphological characteristics such as SLA and SRL. Therefore, differences in the ratio of SLA to SRL can counteract differences in leaf to root mass allocation and limit variation in the ratio of leaf area to root length (Freschet et al., 2015a, b). The relationship between the ratio of SLA to SRL and the ratio of leaf to root mass should be negative, which means that plants adopt strategies ranging from maintaining more biomass in leaves but with a lower SLA and less biomass in roots with a higher SRL, to maintaining more biomass in roots but with a lower SRL and less biomass in leaves but with a higher SLA (Aerts and Chapin, 1999). Changes in the environment will alter the observed regression patterns among these traits. In a study of several northwest European herbaceous species, no differences in regression slopes between SLA:SRL vs. LMF:RMF were observed due to similar direction and intensity of the trait responses to contrasting amounts of nutrient and light supply, but the y-intercepts of each regression significantly differed due to interspecific differences in the traits (Freschet et al., 2015b). However, it remains poorly understood how plants balance these two trends under drought treatments at different stages of the growing season.
The occurrence of plants and the extent of their growth are controlled to a greater degree by water supply than by any other factor, and losses to crop and forage yields due to drought exceed the losses from all other abiotic and/or biotic factors (Boyer, 1982; Volaire, 2018). Through analysis of observed precipitation in China over the last 50 years, the amount and distribution of growing season precipitation has shown significant changes (Lu et al., 2014). In addition, the demand for water by plants varies seasonally. Thus, it is important to understand the impact of drought occurring at different stages of the growing season on plant organ morphology and biomass allocation, while also testing the relative contributions of these plant traits to resource absorption capacity. Estimating these contributions would tell us how important these processes are in the overall response of plants to drought at different times and the subsequent effects of these changes on ecosystem properties, and would provide a complementary approach for clarifying the various adaptive strategies of plants under a changing climate (Poorter et al., 2012).
First, we hypothesized that plants would allocate less biomass to leaves and decrease SLA under drought treatment, increasing the root biomass allocation, SRL and SRA. Second, we hypothesized that plants would respond more strongly under drought occurring earlier rather than later in the season, as the peak of growth occurs in the early growing season. Third, we hypothesized that below-ground variation in RLR and RAR under different drought stresses is driven predominantly by plastic adjustments in root morphology rather than root biomass allocation. Fourth, we hypothesized that drought stress occurring at different times would cause changes in the regression model of SLA:SRL vs. LMF:RMF (and SLA:SRA vs. LMF:RMF). We tested these hypotheses by measuring plant leaf and root biomass allocation and morphology under four drought treatments.
MATERIALS AND METHODS
Plant culture
Grassland mono-dominated by Leymus chinensis (Trin.) Tzvel., a perennial rhizomatous grass with good palatability and high forage value, is widespread in the semi-arid and saline soil area on the eastern edge of the Euro-Asia steppe (Xu et al., 2009; Wang et al., 2019; Shi et al., 2021). Under the dual pressure of overgrazing and sharp climate changes, this steppe has suffered substantial degradation in recent decades (Yu et al., 2018; Zhang et al., 2021c). When this is combined with drought and global warming, soil moisture becomes more critical as daily temperature increases during the growing season. Thus, drought has become a major limiting factor for L. chinensis growth and survival.
On 10 May, 2018, samples of evenly growing L. chinensis were collected from the natural grassland and transplanted to the Jilin Songnen Grassland Ecosystem National Observation and Research Station (44°45ʹN, 123°45ʹE). Mean annual precipitation and temperature at the research station are 445 mm and 5.6 °C, respectively. The soil substrate was collected from the 0–30-cm soil layer in the natural grassland. Soil bulk density was measured by the cutting-ring method (McNabb et al., 2001). Soil field water-holding capacity was measured gravimetrically (soil samples were saturated with water and then weighed). Subsequently, the soil samples were dried at 50 °C until they reached a constant weight (Li et al., 2016). Soil bulk density was 1.45 ± 0.04 g cm−3 and field water-holding capacity was 15.93 ± 0.9 %.
In the grassland study area, 400 sampling points were selected to collect plants. Sampling points were separated by at least 50 m to avoid genetic similarities. Samples were transplanted to plastic pots (the top of the pot is 24 cm in diameter, the bottom is 18 cm, and the height is 26 cm; the volume of the pot is 9 × 103 cm3). Each plastic pot was filled with 8 kg of the collected dry soil (saline meadow soil with pH = 8.05, soil total carbon content of 4.9 g kg−1, soil total nitrogen content of 0.7 g kg−1, soil total phosphorus content of 0.68 g kg−1), and 20 individual plants were spaced evenly in each pot. For the establishment period, all pots received the same amount of water. After 10 d (20 May, 2018), plants with similar heights (average of 13.2 ± 1.4 cm) were retained at a density of 15 individuals per pot after thinning (based on the population size of natural L. chinensis grassland, Zhu, 2004), and drought treatments were started.
Experimental design
All pots were placed in a naturally illuminated glasshouse to exclude precipitation. Based on the length of the growing season of L. chinensis at the local site (May–October) and its phenological stages, we divided the experiment into early (20 May–20 July) and late (21 July–20 September) periods during the 2018 growing season (Fig. 1). Two soil relative water content (SRWC) intensities were applied: well-watered by maintaining a 75–80 % water-holding capacity, and drought by maintaining a 30–35 % water-holding capacity (Xu et al., 2009). We applied a fully crossed factorial design as follows: early treatment with two levels (drought vs. well-watered) and late treatment with two levels (drought vs. well-watered) plus their interactions. This led to four treatment combinations: (1) plants were well-watered in both the early and late periods (WW); (2) plants experienced drought in the early period and were well-watered in the late period (DW); (3) plants were well-watered in the early period and experienced drought in the late period (WD); and (4) plants were subjected to drought in both the early and late periods (DD). Five replicates for each treatment were implemented, for a total of 20 (4 × 5) pots and 300 (20 × 15) individual plants (Fig. 2). These pots were spaced 50 cm from each other. Water was added to return the soil moisture to the upper limit by weighing the soil with the pot at 17:00–19:00 h every 2 d. SRWC was calculated as follows:
Fig. 1.

General morphology of Leymus chinensis (Zhang et al., 2009) and its phenological stages (Shi et al., 2017).
Fig. 2.
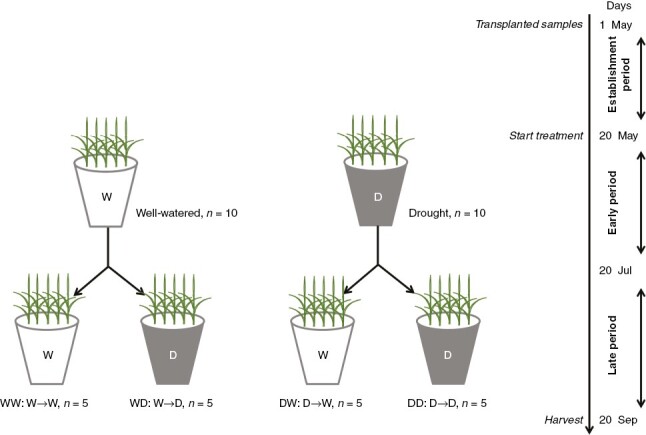
Schematic illustration of the overall experimental design, including the timing of treatment and harvest throughout the whole experiment (W, well-watered; D, drought). The four treatments included: WW, plants were well-watered in both the early and late periods; DW, plants experienced drought in the early period and were well-watered in the late period; WD, plants were well-watered in the early period and experienced drought in the late period; and DD, plants were drought affected in both the early and late periods.
where Wsoil is the current soil weight, Wpot is the weight of the empty pot, DWsoil is the dry soil weight, and WFC is the soil weight at field capacity (Xu et al., 2009). When combining these parameters, (Wsoil + Wpot) is pot weight with current soil, (DWsoil + Wpot) is pot weight with dry soil, and (WFC + Wpot) is pot weight with field capacity soil. The increases in pot weight due to plant biomass growth (i.e. 5.3–8.3 g dry weight at the end of the experiment) had a minor effect on the SRWC calculation. During the experiment, we randomly changed the position of the pots every week. At the same time, we calibrated the soil moisture content using a TRIME Pico64 moisture probe (IMKO GmbH, Ettlingen, Germany) three times (10 May, 20 July and 20 September).
Plant trait measurements
At the end of the experiment (20 September 2018), ten evenly growing individuals that had not been affected by insects or pathogens were selected and marked with tags. We then collected the third or fourth leaf from the top of each plant (healthy and fully expanded) and placed it in a polythene bag with a piece of moist filter paper. The bags were stored in a 4 °C refrigerator for a maximum of 24 h before being scanned to determine leaf area. Collected leaves were surface-dried with filter paper and scanned for single leaf area with a portable leaf area meter (AM-350, ADC Bioscientific Ltd, UK). We then placed the leaves into envelopes, dried them at 65 °C for 48 h and weighed them. SLA (cm2 g−1) was estimated as the ratio of single leaf area to single leaf dry mass.
Next, individual plants were cleaned with water and sorted into leaf, stem, rhizome and root components. Roots were also placed into polythene bags with moist filter paper and stored in a 4 °C refrigerator, for up to 24 h prior to scanning and analysed using analysis software (WinRHIZO Pro 2016b, Regent Instruments, Canada) for root length (m) and root surface area (cm2). All plant parts were subsequently dried at 65 °C for 48 h and weighed. SRL (m g−1) and SRA (cm2 g−1) were calculated as the ratio of root length to root dry mass and the ratio of root area to root dry mass, respectively. Total leaf area was estimated as total leaf mass per plant multiplied by SLA (Konôpka et al., 2021). For each plant, LMF and RMF were calculated as proportions of the total plant dry mass (g g−1). Individual values for plant LAR, RLR and RAR are the total leaf area, root length and root surface area per total plant dry mass (cm2 g−1, m g−1 and cm2 g−1), respectively. The abbreviations, explanations and measurement methods for plant traits in this article are shown in Supplementary Data Table S1.
Data analysis
In this study, the linear mixed-effect model (LMM), the emmeans function and standardized major axis were performed using R version 3.6.0 (R Core Team, 2020). Variance partitioning was calculated with a macro in Excel. The specific steps for data analysis were as follows. First, we tested normality and homogeneity across all the data, and the results showed that LMF and SRA needed sqrt-transformation to meet normality and homoscedasticity of residuals. Then we analysed the treatment effects on plant organ biomass, biomass allocation and morphology using the LMM implemented in the nlme package and the lme function (Pinheiro et al., 2021). The analysis was based on 50 individual replicates for each treatment (5 pots × 10 individual plants). During the LMM analysis, early and late treatment (two levels of soil moisture conditions: well-watered vs. drought) and their interactions were included as fixed factors, and the pots as a random factor, and plant height measured before drought (original plant height) was treated as covariate. We used the emmeans package to undertake post-hoc contrast analysis. The emmeans package is intended for use with a wide variety of ANOVA models, including mixed-effect models (Bruce, 2021).
The calculation method for LMF was leaf dry mass divided by total plant dry mass, SLA was leaf area divided by leaf dry mass, and LAR was leaf area divided by total plant dry mass. Thus, LMF × SLA can decompose into the following formula:
The resulting cancellation of leaf dry biomass produces the formula:
We used the variance partitioning method to compare the contribution of LMF and SLA to LAR via a macro in Excel (Rees et al., 2010; Freschet et al., 2015a). The equations LAR = LMF × SLA, RLR = RMF × SRL, and RAR = RMF × SRA, can be further expressed as: lar = lmf + sla, rlr = rmf + srl, and rar = rmf + sra, where lowercase characters are used to indicate loge-transformed variables. Consequently, the variance decomposition of lar, for example, can be expressed as: Var (lar) = Var (lmf) + Var (sla) + Cov (lmf, sla), where Var is the variance and Cov is the covariance. Following this, the contribution of variation in lmf to the variation in lar can be written as: Cont (lmf) = (Var (lmf) + Cov (lmf, sla))/Var (lar) (Freschet et al., 2015a, b). Similarly, we partitioned the variance in LAR, RLR and RAR across all four treatments.
We used the standardized major axis (SMA) regression (SMART version 2.0; Fellmann, 2012) to test the potential counteracting effects of SLA:SRL and LMF:RMF on the ratio of LAR to RLR and SLA:SRA and LMF:RMF on the ratio of LAR to RAR. For each drought treatment, SMA regression was performed between LMF:RMF and SLA:SRL and between LMF:RMF and SLA:SRA. The regression equation is given as y=αx + β, where α is the slope and β the y-intercept. Heterogeneity among SMA regression equations under the four drought treatments was tested. If the slopes were significantly different, we only compared the heterogeneity of slopes; the y-intercept homogeneity comparisons were performed only when slopes were homogeneous (Cui et al., 2020).
RESULTS
Variable responses of plant biomass allocation and morphology to drought stress
The results of the LMM showed that the original plant height of L. chinensis had no significant effects on biomass, biomass allocation and organ morphology (Supplementary Data Table S2; Table 1). Early treatment had significant effects on either biomass allocation (LMF and RMF) or organ morphology (SLA, SRL, SRA, LAR, RLR and RAR) (Table 1). The results indicated that all the plant traits (biomass allocation and organ morphology) were significantly different between WW, WD and DW, DD (Fig. S1). Late treatment had significant effects only on SLA and two of the root morphology traits (RLR and RAR) (Table 1). SLA, RLR and RAR were significantly different between WW, DW and WD, DD (Fig. S1). The early and late treatment had significant interactive effects on biomass allocation and organ morphology (Table 1). LAR was greater under the DW treatment than under the WW treatment, through a slight increase in SLA and a significant increase in LMF (Fig. 3A, D, F). WD treatment had only a small positive effect on plant LAR, and this result stemmed from the counteracting effects of a significantly decreased SLA and increased LMF. DD treatment also had a slightly positive effect on plant LAR, owing to a significant decrease in SLA, which was counterbalanced by a significant increase in LMF (Fig. 3A, D, F).
Table 1.
The effect of experimental drought in different periods on plant biomass allocation and organ morphology. Response variables were specific leaf area (SLA), specific root length (SRL), specific root area (SRA), leaf area ratio (LAR), root length ratio (RLR), root area ratio (RAR), leaf mass fraction (LMF) and root mass fraction (RMF). Significant treatments (‘E’ and ‘L’ refer to early and late treatment), the interactive effects and the covariates from original plant height are shown
| Response variable | Parameter | d.f. | Sample size | F | P |
|---|---|---|---|---|---|
| SLA | E | 1 | 100 | 9.32 | 0.003 |
| L | 1 | 100 | 52.6 | <0.001 | |
| E × L | 1 | 100 | 10.1 | 0.002 | |
| Covariate | 1 | 100 | 0.125 | 0.724 | |
| SRL | E | 1 | 100 | 83.6 | <0.001 |
| L | 1 | 100 | 1.17 | 0.281 | |
| E × L | 1 | 100 | 66.6 | <0.001 | |
| Covariate | 1 | 100 | 1.492 | 0.223 | |
| SRA | E | 1 | 100 | 38.8 | <0.001 |
| L | 1 | 100 | 1.418 | 0.235 | |
| E × L | 1 | 100 | 33.9 | <0.001 | |
| Covariate | 1 | 100 | 2.073 | 0.152 | |
| LAR | E | 1 | 100 | 8.72 | 0.004 |
| L | 1 | 100 | 2.419 | 0.122 | |
| E × L | 1 | 100 | 10.8 | 0.001 | |
| Covariate | 1 | 100 | 0.692 | 0.407 | |
| RLR | E | 1 | 100 | 108.6 | <0.001 |
| L | 1 | 100 | 6.044 | 0.015 | |
| E × L | 1 | 100 | 88.3 | <0.001 | |
| Covariate | 1 | 100 | 3.48 | 0.064 | |
| RAR | E | 1 | 100 | 107.1 | <0.001 |
| L | 1 | 100 | 10.9 | 0.001 | |
| E × L | 1 | 100 | 75.9 | <0.001 | |
| Covariate | 1 | 100 | 2.399 | 0.123 | |
| LMF | E | 1 | 100 | 24.1 | <0.001 |
| L | 1 | 100 | 3.107 | 0.08 | |
| E × L | 1 | 100 | 4.338 | 0.039 | |
| Covariate | 1 | 100 | 0.965 | 0.327 | |
| RMF | E | 1 | 100 | 23.3 | <0.001 |
| L | 1 | 100 | 3.718 | 0.055 | |
| E × L | 1 | 100 | 16.9 | <0.001 | |
| Covariate | 1 | 100 | 0.333 | 0.564 |
P-values of significant parameters are shown in bold.
Fig. 3.

Violin plots of leaf and root biomass allocation and morphology measured under drought treatment. (A) Specific leaf area, SLA; (B) specific root length, SRL; (C) specific root area, SRA; (D) leaf mass fraction, LMF; (E) root mass fraction, RMF; (F) leaf area ratio, LAR; (G) root length ratio, RLR; (H) root area ratio, RAR. Different capital letters denote a P < 0.05 difference based on a Tukey’s post-hoc test among treatments (WW: plants were well-watered in both the early and late periods, DW: plants experienced drought in the early period and were well-watered in the late period, WD: plants were well-watered in the early period and experienced drought in the late period, and DD: plants experienced drought in both the early and late periods).
RLR was greater under drought treatment (including DW, WD and DD) than under WW treatment through an increase in both SRL and RMF (Fig. 3B, E, G). The changes in RAR and RLR were consistent, and there were similarities to the trends in both SRA and SRL (Fig. 3C, E, H). Interestingly, the maximum values in LAR, RLR and RAR appeared in the DW treatment (Fig. 3F–H).
Relative contributions of morphological changes and biomass allocation to plant above- and below-ground parts
Above ground, LAR responded significantly to changes in water supply, and 68–84 % of the plastic variation in LAR was the result of changes in LMF (Fig. 4). In the WW treatment, the contribution of LMF to the total LAR variation was 74 %, and for SLA it was 26 % (Fig. 4). Similar results were observed for the DW and WD treatments (for DW, LMF contributed 68 % and SLA contributed 32 %; for WD, LMF contributed 79 % and SLA contributed 21 %). However, the contribution of LMF increased to 84 % for the LAR variation under the DD treatment, which was 5.3-fold larger than SLA (Fig. 4).
Fig. 4.

Relative contribution of leaf and root biomass allocation (leaf mass fraction: LMF, or root mass fraction: RMF, light grey bars) and morphology (specific leaf area: SLA, specific root length: SRL, or specific root area: SRA, dark grey bars) to the total variation in leaf area ratio (LAR), root length ratio (RLR) and root area ratio (RAR).
Below ground, RLR and RAR responded significantly to changes in water supply (Fig. 3). The contributions of RMF to total the RLR variation were 75, 65, 66 and 67 % under WW, DW, WD and DD conditions, respectively (Fig. 4). Regarding RAR variation, the contribution of RMF was ~1.6-fold larger than SRA under the WW, DW and WD treatments (Fig. 4). Conversely, the contribution of SRA to RAR variation was 69 %, which was 2.2-fold larger than RMF under the DD treatment (Fig. 4).
Counteracting effects of leaf and root biomass allocation on leaf-root morphology
Significant negative relationships between the ratios of SLA:SRL and LMF:RMF were found in all four treatments (Fig. 5A; Supplementary Data Table S3). Meanwhile, significant differences were observed between regression slopes of WW and the three drought treatments (Fig. 5A, Table 2). The reason for this result was that the increase in SLA and decrease in SRL increased SLA:SRL under WW treatment (Fig. 3), and this ultimately changed the regression slope. Although there were no significant differences among the DW, WD and DD slopes, the y-intercepts were markedly different (Table 2), which indicated that the plants had a higher LMF:RMF for a given SLA:SRL under the WD treatment, followed by the DW and DD treatments. The regression was also significantly negative between SLA:SRA and LMF:RMF for DW, WD and DD, but not for the WW treatment (Fig. 5B; Table S3). Significant differences were observed in slopes or y-intercepts between every pair of regression lines, except for WD vs. DD (Table 2).
Fig. 5.

Standardized major axis (SMA) regressions between (A) the leaf to root mass ratio and the specific leaf area to specific root length (SLA:SRL) ratio under the WW (blue), DW (black), WD (purple) and DD (red) treatments. SMA regressions between (B) the leaf to root mass ratio and the specific leaf area to specific root area (SLA:SRA) ratio. Significant relationships are indicated by solid lines and non-significant relationships are indicated by dashed lines.
Table 2.
Results of the standardized major axis regression (SMA) analyses of the specific leaf area to specific root length (SLA:SRL) ratio and the leaf to root mass ratio, and the specific leaf area to specific root area (SLA:SRA) ratio and the leaf to root mass ratio among different drought treatments
| Comparison object | Slope | y-intercept | Comparison object | Slope | y-intercept | ||
|---|---|---|---|---|---|---|---|
| LMF:RMF–SLA:SRL | WW vs. DW | 0.027 | LMF:RMF–SLA:SRA | WW vs. DW | <0.001 | ||
| WW vs. WD | <0.001 | WW vs. WD | 0.02 | ||||
| WW vs. DD | <0.001 | WW vs. DD | 0.182 | 0.03 | |||
| DW vs. WD | 0.127 | <0.001 | DW vs. WD | 0.091 | <0.001 | ||
| DW vs. DD | 0.111 | <0.001 | DW vs. DD | 0.01 | |||
| WD vs. DD | 0.925 | 0.007 | WD vs. DD | 0.333 | 0.341 |
Significant differences in slope and y-intercept (P < 0.05) are shown in bold.
DISCUSSION
Understanding the mechanisms of plant responses to various drought conditions is a crucial issue for plants to adapt to future rainfall changes (Kumordzi et al., 2019). Our study showed that drought occurring in the early growing season had a greater impact on plant biomass allocation and organ morphology than drought in the later period. The contribution of LMF to LAR was greater than SLA under different drought treatments, and the contribution of RMF to RLR was greater than SRL. However, the contribution of SRA to RAR was greater than RMF under persistent drought treatment (drought in both the early and the late periods).
Leaf and root systems adopted water conservation and water absorption strategies, respectively, to cope with drought stress
Based on optimal partitioning theory, plants will allocate a proportionally greater amount of resources to the structure, with the limiting resources captured so that plant performance is optimized (Bloom et al., 1985; Eziz et al., 2017). For example, plants invest more biomass in roots and less in leaves when experiencing moisture deficit (Poorter et al., 2012). In this study, L. chinensis allocated more biomass to roots under the early drought treatments (DW and DD) than the early well-watered treatments (WW and WD) to promote water absorption (Fig. 3; Supplementary Data Fig. S1e), which was consistent with optimal partitioning theory (Freschet et al., 2018; Enquist and Niklas, 2002; Rehling et al., 2021). Specifically, the maximum value of RMF occurred in DW, followed by DD, WD and WW (Fig. 3E). This was generated by greater root biomass production under the DW treatment (Fig. S2). In accordance with the compensatory growth hypothesis, rapid plant growth can be observed when sufficient water is resupplied after dehydration (Spieß et al., 2012; Wang et al., 2016; Wang et al., 2017). The reason for the rapid development of the root system may be that it offsets the losses caused by previous drought stress so that the possibility of future drought can be managed. For LMF, the value under early drought (DW and DD) was obviously higher than the early well-watered treatments (WW and DW) (Fig. 3D; Fig. S1). The minimum LMF value appeared in the WW treatment, and was significantly lower than in the DW, WD and DD treatments (Fig. 3D). This result differs from a global worldwide meta-analysis that reported a significant decrease in the fraction of leaf mass under drought (Eziz et al., 2017). The reason for this discrepancy is that wetter conditions promoted L. chinensis biomass accumulation in leaves, stems, roots and especially rhizomes (Fig. S2, Table S4), and rhizome formation caused the increase in total plant biomass to exceed total leaf biomass, finally resulting in a reduction in LMF. In contrast to other plants, rhizomatous grasses not only grow upward with above-ground tillers, but also expand laterally via rhizomes (Huang et al., 2019; Wang et al., 2019; Fig. 1). This also led to a decrease in LAR in L. chinensis (Fig. 3F). Compared with previous studies on other species (Eziz et al., 2017; Puglielli et al., 2021), the current work on L. chinensis has indicated the potential for species-specific biomass allocation patterns under future rainfall changes.
In addition to the changes in biomass allocation to plant organs, plants can also alter the morphology of each of their organs to respond to various environments (Poorter and Ryser, 2015; Poorter et al., 2019). In the current experiment, SRL and SRA increased significantly with drought treatment (Fig. 3B, C). This result is consistent with previous research on the meadow grass Microlaena stipoides under changing precipitation (Ryalls et al., 2018). This is because low water availability induced high SRL and SRA values, allowing better exploration and exploitation of deeper soil horizons (Alvarez-Flores et al., 2014; Freschet et al., 2015b). However, Coffea arabica has been shown to exhibit the opposite response (Isaac et al., 2017). Therefore, the mechanisms underlying the diverse response patterns of different plant functional types warrant further exploration. Furthermore, the maximum values of SRL and SRA appeared in the DW treatment (Fig. 3B, C), so this means that the early drought and then well-watered treatment can extend the total length and area of root achieved per unit root mass, thus facilitating water absorption. For leaf morphology, SLA decreased significantly in response to water deficit except in the DW treatment. SLA reduction is often interpreted as a mechanism of dehydration avoidance, and this effect could be the composite outcome of decreased leaf area and increased leaf thickness (Markesteijn et al., 2011), enabling leaves to maintain turgor and minimize cell damage under drought, and this has been related to an increase in survival under drought conditions (Bongers et al., 2017; Lozano et al., 2020). In this study, the value of SLA in the DW treatment was significantly higher than in the other drought treatments. This intriguing difference indicated that the soil moisture status during later periods determined the state of SLA.
Based on the discussion above, our study exploring the intraspecific responses of organ biomass allocation and other traits to changing water availability has revealed that both optimal partitioning theory and resource economics theory are tenable. The data therefore support our first hypothesis that both variations in organ mass allocation and morphology contributed to reducing transpiration and strengthening moisture absorption. However, this result is not consistent with the second hypothesis because root biomass, length and surface area under DW were greater than under WD (Supplementary Data Figs S2 and S3). In response, resource limitations would induce plant functional adjustment consistent with an improvement in the plants’ capacity to acquire the limiting resource (Freschet et al., 2018). In our study, low water availability induced a high RLR and RMF, allowing better exploration and exploitation of deeper soil horizons (Freschet et al., 2015b), which is consistent with the root foraging theory (Alvarez-Flores et al., 2014). At the same time, SLA decreased with drought stress, indicating a smaller leaf area per unit of blade biomass (de Smedt et al., 2012). Moreover, a small leaf area under drought stress (Fig. S3) could decrease water loss by reducing the amount of transpiring tissues and then retain water to facilitate survival, growth and necessary physiological activities (Markesteijn and Poorter, 2009).
Drought stress alters the relative contributions of plant biomass allocation and morphology to resource acquisition
Previous studies have demonstrated that changes in SLA play a more prominent role than LMF in LAR variation, but the magnitude of the contribution has varied according to geographical location. For example, a 9-fold higher contribution of SLA compared with LMF was found in a subalpine plant community on New Zealand’s South Island (Freschet et al., 2015a), while it was only 2.5-fold higher for northwestern European herbaceous species in the Sheffield region of the UK (Freschet et al., 2015b), and a meta-analysis from a worldwide array of experimental data estimated a 7-fold higher contribution of SLA than of LMF (Poorter et al., 2012). In the current study, we observed the opposite results: LMF variation contributed more than SLA to plastic differences in LAR, and soil moisture altered the magnitude of the contribution (Fig. 4). These findings suggested that the degree of contribution was dependent on geographical location and species differences, and this was even the case for major contributors to above-ground plant resources such as leaf biomass allocation and morphology.
For the root system in our study, the variation in RMF made an ~2.3 times greater contribution towards modulating RLR (relative length of roots exploring soil) than the contribution by SRL. This result is in line with the work of Freschet et al. (2015a, b) where RMF was 2.5 times stronger than SRL in contributing to differences in RLR in alpine species in New Zealand and in herbaceous species in the UK. Furthermore, we also calculated the contribution of RMF and SRA to RAR (relative area of roots touching soil). Similar to the result for RLR, changes in root biomass allocation were a 1.6-fold stronger contribution to RAR than SRA under WW, DW and WD. However, the opposite result was found under DD treatment (2.2-fold higher contribution of SRA than RMF to differences in RAR). In general, plasticity in root morphology may not be very important for plant resource acquisition (Fig. 4; Freschet et al., 2015a, b), while variation in SRA may increase the contribution as the drought period becomes longer. Thus, root traits involved in resource acquisition other than SRL also deserve attention, such as SRA, root architecture, root hair length and density, and the type and intensity of mycorrhizal associations.
The asynchronous development of plant organs altered the plant absorption balance between above- and below-ground parts in response to drought
Generally, the plants can achieve balanced resource acquisition by trading off the biomass vs. morphology of above- and below-ground organs (Freschet et al., 2015a, b). This was particularly true in our study, and the differences in the ratio of SLA to SRL (or the ratio of SLA to SRA) caused by changes in soil moisture were counteracted by differences in leaf to fine root mass allocation, similar to that described by Freschet et al. (2015a). Although the direction of relationships between LMF:RMF and SLA:SRL (or SLA:SRA) were stable and negative, the correlation indexes (slope or y-intercept) were significantly different among the soil moisture treatments (Fig. 5, Table 2). For example, the slope increased with water supply, but the y-intercept decreased (Fig. 5, Table 2). This is in contrast to the findings of Freschet et al. (2015b) who observed no significant difference between regression slopes but a significant decrease in y-intercept occurred in the low soil nutrient treatment. Our results partially support the initial part of the fourth hypothesis that the regression between LMF:RMF and SLA:SRL was indeed negative but oppose the later part of the hypothesis that soil fertility and moisture improvement can increase the regression intercepts. The reason for this phenomenon is that a favourable soil environment may promote the lateral expansion of rhizomatous grass through rhizomes (Fig. 1; Supplementary Data Fig. S2). Under the WW treatment, the roots and buds (mainly axillary rhizome buds and tiller buds) formed on the rhizome, but many buds had not yet developed into above-ground parts (leaves and stems), which resulted in a higher rhizome biomass under WW than the DW, WD and DD treatments (3.5, 2.4 and 3.4 times greater, respectively), while the biomass of leaves under WW was 1.3, 1.2 and 1.5 times greater than the other three treatments, respectively (Fig. S2). This allometric growth between organs led to a decrease in LMF in the WW treatment and then reduced the y-intercept of the regression between LMF:RMF and SLA:SRL (or SLA:SRA). Our study thus emphasizes the influence of the environment on the absorption balance in above- and below-ground resources by affecting the development of plant organs.
CONCLUSION
For L. chinensis, we revealed that biomass allocation drove a larger proportion of variation than morphological changes in response to differences in water supply. Our results demonstrated that leaf morphology displayed a water conservation strategy (lower SLA), while leaf biomass allocation did not support this strategy (higher LMF). Root biomass allocation and morphology displayed a water absorption strategy. Moreover, disproportionate growth among leaves, roots and rhizomes due to different water conditions regulated the absorption balance of above- and below-ground resources. These and previous results help us to better understand the range of variation in plant phenotypic plasticity in response to rainfall changes in the future.
SUPPLEMENTARY DATA
Supplementary data are available online at https://academic.oup.com/aob and consist of the following. Table S1: Abbreviations used in this article, together with explanation, measuring method and the units applied. Table S2: The effects of original plant height as a covariate on L. chinensis organ biomass, biomass allocation and morphology traits according to the linear mixed-effect model. Table S3: Standardized major axis regression analyses of specific leaf area. Table S4: Experimental drought effects on plant organ biomass and organ morphology. Fig. S1: The effects of early treatment and late treatment on plant biomass allocation and organ morphology. Fig. S2: Above- and below-ground biomass. Fig. S3: Leaf area, root length and root area.
ACKNOWLEDGEMENTS
We are grateful for the help of Dr Freschet, Dr Xuefeng Wu and Dr Xue Yang for their advice on data analysis questions, Mr Jingtian Chen and Mr Chao Li who assisted with field work, and Ms Xiuquan Yue and Ms Ya’nan Li who contributed to the laboratory analyses. Dr Rowan Sage and Karolina Heyduk and two reviewers made insightful remarks on a previous version of the manuscript.
Contributor Information
Yuheng Yang, Institute of Grassland Science, Key Laboratory of Vegetation Ecology of the Ministry of Education, Jilin Songnen Grassland Ecosystem National Observation and Research Station, Northeast Normal University, Changchun 130024, China.
Yujie Shi, Institute of Grassland Science, Key Laboratory of Vegetation Ecology of the Ministry of Education, Jilin Songnen Grassland Ecosystem National Observation and Research Station, Northeast Normal University, Changchun 130024, China.
Xiaowei Wei, Institute of Grassland Science, Key Laboratory of Vegetation Ecology of the Ministry of Education, Jilin Songnen Grassland Ecosystem National Observation and Research Station, Northeast Normal University, Changchun 130024, China.
Jiayu Han, Institute of Grassland Science, Key Laboratory of Vegetation Ecology of the Ministry of Education, Jilin Songnen Grassland Ecosystem National Observation and Research Station, Northeast Normal University, Changchun 130024, China.
Junfeng Wang, Institute of Grassland Science, Key Laboratory of Vegetation Ecology of the Ministry of Education, Jilin Songnen Grassland Ecosystem National Observation and Research Station, Northeast Normal University, Changchun 130024, China.
Chunsheng Mu, Institute of Grassland Science, Key Laboratory of Vegetation Ecology of the Ministry of Education, Jilin Songnen Grassland Ecosystem National Observation and Research Station, Northeast Normal University, Changchun 130024, China.
Jinwei Zhang, Institute of Grassland Science, Key Laboratory of Vegetation Ecology of the Ministry of Education, Jilin Songnen Grassland Ecosystem National Observation and Research Station, Northeast Normal University, Changchun 130024, China.
FUNDING
This work was supported by the National Natural Science Foundation of China (31971505), the Natural Science Foundation of Jilin Province (YDZJ202101ZYTS081), the Youth Talent Support Project of Jilin Province (QT202007), the Science and Technology Project of the Jilin Provincial Education Department (JJKH20221169KJ) and the China Postdoctoral Science Foundation (2021M690030).
LITERATURE CITED
- Acuña-Míguez B, Olano JM, Valladares F, García-Hidalgo M, Bravo-Oviedo A.. 2022. Increased recovery in coarse‐root secondary growth improves resilience to drought in transition forests. Journal of Ecology 111: 170–181. doi: 10.1111/1365-2745.14024. [DOI] [Google Scholar]
- Aerts R, Chapin FS.. 1999. The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns. Advances in Ecological Research 30: 1–67. doi: 10.1016/S0065-2504(08)60016-1. [DOI] [Google Scholar]
- Alvarez-Flores R, Winkel T, Nguyen-Thi-Truc A, Joffre R.. 2014. Root foraging capacity depends on root system architecture and ontogeny in seedlings of three Andean Chenopodium species. Plant and Soil 380: 415–428. doi: 10.1007/s11104-014-2105-x. [DOI] [Google Scholar]
- Bardgett RD, Mommer L, de Vries FT.. 2014. Going underground: root traits as drivers of ecosystem processes. Trends in Ecology & Evolution 29: 692–699. doi: 10.1016/j.tree.2014.10.006. [DOI] [PubMed] [Google Scholar]
- Bebre I, Marques I, Annighöfer P.. 2022. Biomass allocation and leaf morphology of saplings grown under various conditions of light availability and competition types. Plants 11: 305. doi: 10.3390/plants11030305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bloom AJ, Chapin III FS, Mooney HA.. 1985. Resource limitation in plants – an economic analogy. Annual Review of Ecology and Systematics 16: 363–392. doi: 10.1146/annurev.ecolsys.16.1.363. [DOI] [Google Scholar]
- Bongers FJ, Olmo M, Lopez-Iglesias B, Anten NPR, Villar R.. 2017. Drought responses, phenotypic plasticity and survival of Mediterranean species in two different microclimatic sites. Plant Biology 19: 386–395. doi: 10.1111/plb.12544. [DOI] [PubMed] [Google Scholar]
- Boyer JS. 1982. Plant productivity and environment. Science 218: 443–448. doi: 10.1126/science.218.4571.443. [DOI] [PubMed] [Google Scholar]
- Bruce D. 2021. One way ANOVA with R, chapter 6: Beginning to Explore the emmeans package for post hoc tests and contrasts. Albany: University at Albany Press. [Google Scholar]
- Comas LH, Becker SR, Cruz VMV, Byrne PF, Dierig DA.. 2013. Root traits contributing to plant productivity under drought. Frontiers in Plant Science 4: 442. doi: 10.3389/fpls.2013.00442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cui EQ, Weng ES, Yan ER, Xia JY.. 2020. Robust leaf trait relationships across species under global environmental changes. Nature Communications 11: 2999. doi: 10.1038/s41467-020-16839-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du L, Liu HY, Guan WB, Li JM, Li JS.. 2019. Drought affects the coordination of belowground and aboveground resource-related traits in Solidago canadensis in China. Ecology and Evolution 9: 9948–9960. doi: 10.1002/ece3.5536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dudley BD, Hughes RF, Ostertag R.. 2014. Groundwater availability mediates the ecosystem effects of an invasion of Prosopis pallida. Ecological Applications 24: 1954–1971. doi: 10.1890/13-1262.1. [DOI] [PubMed] [Google Scholar]
- Elser JJ, Fagan WF, Kerkhoff AJ, Swenson NG, Enquist BJ.. 2010. Biological stoichiometry of plant production: metabolism, scaling and ecological response to global change. The New Phytologist 186: 593–608. doi: 10.1111/j.1469-8137.2010.03214.x. [DOI] [PubMed] [Google Scholar]
- Enquist BJ, Niklas KJ.. 2002. Global allocation rules for patterns of biomass partitioning in seed plants. Science 295: 1517–1520. doi: 10.1126/science.1066360. [DOI] [PubMed] [Google Scholar]
- Eziz A, Yan ZB, Tian D, Han WX, Tang ZY, Fang JY.. 2017. Drought effect on plant biomass allocation: A meta-analysis. Ecology and Evolution 7: 11002–11010. doi: 10.1002/ece3.3630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fellmann CD. 2012. Ontogeny of anatomical mechanical advantage of the biceps brachii muscle in Macaques. In: Wang Q, ed. Bones, genetics, and behavior of rhesus macaques. New York: Springer, 159–176. [Google Scholar]
- Freschet GT, Kichenin E, Wardle DA.. 2015a. Explaining within-community variation in plant biomass allocation: a balance between organ biomass and morphology above vs below ground? Journal of Vegetation Science 26: 431–440. doi: 10.1111/jvs.12259. [DOI] [Google Scholar]
- Freschet GT, Roumet C, Comas LH, et al. 2021. Root traits as drivers of plant and ecosystem functioning: current understanding, pitfalls and future research needs. The New Phytologist 232: 1123–1158. doi: 10.1111/nph.17072. [DOI] [PubMed] [Google Scholar]
- Freschet GT, Swart EM, Cornelissen JHC.. 2015b. Integrated plant phenotypic responses to contrasting above- and below-ground resources: key roles of specific leaf area and root mass fraction. The New Phytologist 206: 1247–1260. doi: 10.1111/nph.13352. [DOI] [PubMed] [Google Scholar]
- Freschet GT, Violle C, Bourget MY, Scherer-Lorenzen M, Fort F.. 2018. Allocation, morphology, physiology, architecture: the multiple facets of plant above- and below-ground responses to resource stress. The New Phytologist 219: 1338–1352. doi: 10.1111/nph.15225. [DOI] [PubMed] [Google Scholar]
- Fry EL, Evans AL, Sturrock CJ, Bullock JM, Bardgett RD.. 2018. Root architecture governs plasticity in response to drought. Plant and Soil 433: 189–200. doi: 10.1007/s11104-018-3824-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geng Y, Wang L, Jin DM, Liu HY, He JS.. 2014. Alpine climate alters the relationships between leaf and root morphological traits but not chemical traits. Oecologia 175: 445–455. doi: 10.1007/s00442-014-2919-5. [DOI] [PubMed] [Google Scholar]
- Huang LS, Flavell R, Donnison IS, et al. 2019. Collecting wild Miscanthus germplasm in Asia for crop improvement and conservation in Europe whilst adhering to the guidelines of the United Nations’ Convention on Biological Diversity. Annals of Botany 124: 591–604. doi: 10.1093/aob/mcy231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Isaac ME, Martin AR, de Melo Virginio Filho E, Rapidel B, Roupsard O, van den Meersche K.. 2017. Intraspecific trait variation and coordination: root and leaf economics spectra in coffee across environmental gradients. Frontiers in Plant Science 8: 1196. doi: 10.3389/fpls.2017.01196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Konôpka B, Pajtík J, Šebeň V, Surový P, Merganičová K.. 2021. Woody and foliage biomass, foliage traits and growth efficiency in young trees of four broadleaved tree species in a temperate forest. Plants 10: 2155. doi: 10.3390/plants10102155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumordzi BB, Aubin I, Cardou F, et al. 2019. Geographic scale and disturbance influence intraspecific trait variability in leaves and roots of North American understorey plants. Functional Ecology 33: 1771–1784. doi: 10.1111/1365-2435.13402. [DOI] [Google Scholar]
- Laughlin DC, Mommer L, Sabatini FM, et al. 2021. Root traits explain plant species distributions along climatic gradients yet challenge the nature of ecological trade-offs. Nature Ecology & Evolution 5: 1123–1134. doi: 10.1038/s41559-021-01471-7. [DOI] [PubMed] [Google Scholar]
- Li CN, Li L, Reynolds MP, et al. 2021. Recognizing the hidden half in wheat: root system attributes associated with drought tolerance. Journal of Experimental Botany 72: 5117–5133. doi: 10.1093/jxb/erab124. [DOI] [PubMed] [Google Scholar]
- Li JM, Du LS, Guan WB, Yu FH, van Kleunen M.. 2016. Latitudinal and longitudinal clines of phenotypic plasticity in the invasive herb Solidago canadensis in China. Oecologia 182: 755–764. doi: 10.1007/s00442-016-3699-x. [DOI] [PubMed] [Google Scholar]
- Lozano YM, Aguilar-Trigueros CA, Flaig IC, Rillig MC.. 2020. Root trait responses to drought are more heterogeneous than leaf trait responses. Functional Ecology 34: 2224–2235. doi: 10.1111/1365-2435.13656. [DOI] [Google Scholar]
- Lu E, Zeng YT, Luo YL, et al. 2014. Changes of summer precipitation in China: The dominance of frequency and intensity and linkage with changes in moisture and air temperature. Journal of Geophysical Research Atmospheres 119: 12575–12587. doi: 10.1002/2014JD022456. [DOI] [Google Scholar]
- Luo WT, Griffin-Nolan RJ, Ma W, et al. 2021. Plant traits and soil fertility mediate productivity losses under extreme drought in C3 grasslands. Ecology 102: e03465. doi: 10.1002/ecy.3465. [DOI] [PubMed] [Google Scholar]
- Luong JC, Holl KD, Loik ME.. 2021. Leaf traits and phylogeny explain plant survival and community dynamics in response to extreme drought in a restored coastal grassland. Journal of Applied Ecology 58: 1670–1680. doi: 10.1111/1365-2664.13909. [DOI] [Google Scholar]
- Maire V, Gross N, Hill D, et al. 2013. Disentangling coordination among functional traits using an individual-centred model: impact on plant performance at intra- and inter-specific levels. PLoS One 8: e77372. doi: 10.1371/journal.pone.0077372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- March-Salas M, van Kleunen M, Fitze PS.. 2022. Effects of intrinsic precipitation-predictability on root traits, allocation strategies and the selective regimes acting on them. Oikos 1: e07970. doi: 10.1111/oik.07970. [DOI] [Google Scholar]
- Markesteijn L, Poorter L.. 2009. Seedling root morphology and biomass allocation of 62 tropical tree species in relation to drought- and shade-tolerance. Journal of Ecology 97: 311–325. doi: 10.1111/j.1365-2745.2008.01466.x. [DOI] [Google Scholar]
- Markesteijn L, Poorter L, Paz H, Sack L, Bongers F.. 2011. Ecological differentiation in xylem cavitation resistance is associated with stem and leaf structural traits. Plant, Cell & Environment 34: 137–148. doi: 10.1111/j.1365-3040.2010.02231.x. [DOI] [PubMed] [Google Scholar]
- McNabb DH, Startsev AD, Nguyen H.. 2001. Soil wetness and traffic level effects on bulk density and air-filled porosity of compacted boreal forest soils. Soil Science Society of America Journal 65: 1238–1247. doi: 10.2136/sssaj2001.6541238x. [DOI] [Google Scholar]
- Migliavacca M, Musavi T, Mahecha MD, et al. 2021. The three major axes of terrestrial ecosystem function. Nature 598: 468–472. doi: 10.1038/s41586-021-03939-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Müller I, Schmid B, Weiner J.. 2000. The effect of nutrient availability on biomass allocation patterns in 27 species of herbaceous plants. Perspectives in Plant Ecology, Evolution and Systematics 3: 115–127. doi: 10.1078/1433-8319-00007. [DOI] [Google Scholar]
- Pinheiro J, Bates D, DebRoy S, Sarkar D; R Core Team. 2021. nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-152. https://svn.r-project.org/R-packages/trunk/nlme/
- Poorter H, Niinemets U, Ntagkas N, et al. 2019. A meta-analysis of plant responses to light intensity for 70 traits ranging from molecules to whole plant performance. The New Phytologist 223: 1073–1105. doi: 10.1111/nph.15754. [DOI] [PubMed] [Google Scholar]
- Poorter H, Niinemets U, Poorter L, Wright IJ, Villar R.. 2009. Causes and consequences of variation in leaf mass per area (LMA): a meta-analysis. The New Phytologist 182: 565–588. doi: 10.1111/j.1469-8137.2009.02830.x. [DOI] [PubMed] [Google Scholar]
- Poorter H, Niklas KJ, Reich PB, Oleksyn J, Poot P, Mommer L.. 2012. Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control. The New Phytologist 193: 30–50. doi: 10.1111/j.1469-8137.2011.03952.x. [DOI] [PubMed] [Google Scholar]
- Poorter H, Niinemets U, Walter A, Fiorani F, Schurr U.. 2010. A method to construct dose–response curves for a wide range of environmental factors and plant traits by means of a meta-analysis of phenotypic data. Journal of Experimental Botany 61: 2043–2055. doi: 10.1093/jxb/erp358. [DOI] [PubMed] [Google Scholar]
- Poorter H, Ryser P.. 2015. The limits to leaf and root plasticity: what is so special about specific root length? The New Phytologist 206: 1188–1190. doi: 10.1111/nph.13438. [DOI] [PubMed] [Google Scholar]
- Puglielli G, Laanisto L, Poorter H, Niinemets U.. 2021. Global patterns of biomass allocation in woody species with different tolerances of shade and drought: evidence for multiple strategies. The New Phytologist 229: 308–322. doi: 10.1111/nph.16879. [DOI] [PubMed] [Google Scholar]
- R Core Team. 2020. R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available online at: https://www.R-project.org/ [Google Scholar]
- Rees M, Osborne CP, Woodward FI, Hulme SP, Turnbull LA, Taylor SH.. 2010. Partitioning the components of relative growth rate: how important is plant size variation? The American Naturalist 176: E152–E161. doi: 10.1086/657037. [DOI] [PubMed] [Google Scholar]
- Rehling F, Sandner TM, Matthies D.. 2021. Biomass partitioning in response to intraspecific competition depends on nutrients and species characteristics: A study of 43 plant species. Journal of Ecology 109: 2219–2233. doi: 10.1111/1365-2745.13635. [DOI] [Google Scholar]
- Remke MJ, Johnson NC, Wright J, Williamson M, Bowker MA.. 2021. Sympatric pairings of dryland grass populations, mycorrhizal fungi and associated soil biota enhance mutualism and ameliorate drought stress. Journal of Ecology 109: 1210–1223. doi: 10.1111/1365-2745.13546. [DOI] [Google Scholar]
- Ryalls JMW, Moore BD, Johnson SN, Connor M, Hiltpold I.. 2018. Root responses to domestication, precipitation and silicification: weeping meadow grass simplifies and alters toughness. Plant and Soil 427: 291–304. doi: 10.1007/s11104-018-3650-5. [DOI] [Google Scholar]
- Ryser P, Lambers H.. 1995. Root and leaf attributes accounting for the performance of fast- and slow-growing grasses at different nutrient supply. Plant and Soil 170: 251–265. doi: 10.1007/bf00010478. [DOI] [Google Scholar]
- Sun J, Wang HM.. 2016. Soil nitrogen and carbon determine the trade-off of the above- and below-ground biomass across alpine grasslands, Tibetan Plateau. Ecological Indicators 60: 1070–1076. doi: 10.1016/j.ecolind.2015.08.038. [DOI] [Google Scholar]
- Shi YJ, Gao S, Zhou DH, et al. 2017. Fall nitrogen application increases seed yield, forage yield and nitrogen use efficiency more than spring nitrogen application in Leymus chinensis a perennial grass. Field Crops Research 214: 66–72. doi: 10.1016/j.fcr.2017.08.022. [DOI] [Google Scholar]
- Shi YJ, Wang JF, Ao YN, et al. 2021. Responses of soil N2O emissions and their abiotic and biotic drivers to altered rainfall regimes and co-occurring wet N deposition in a semi-arid grassland. Global Change Biology 27: 4894–4908. doi: 10.1111/gcb.15792. [DOI] [PubMed] [Google Scholar]
- Siddiqui MN, Léon J, Naz AA, Ballvora A.. 2021. Genetics and genomics of root system variation in adaptation to drought stress in cereal crops. Journal of Experimental Botany 72: 1007–1019. doi: 10.1093/jxb/eraa487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Smedt S, Cuní Sanchez A, van den Bilcke N, Simbo D, Potters G, Samson R.. 2012. Functional responses of baobab (Adansonia digitata L.) seedlings to drought conditions: Differences between western and south-eastern Africa. Environmental and Experimental Botany 75: 181–187. doi: 10.1016/j.envexpbot.2011.09.011. [DOI] [Google Scholar]
- Spieß N, Oufir M, Matušíková I, et al. 2012. Ecophysiological and transcriptomic responses of oak (Quercus robur) to long-term drought exposure and rewatering. Environmental and Experimental Botany 77: 117–126. doi: 10.1016/j.envexpbot.2011.11.010. [DOI] [Google Scholar]
- Volaire F. 2018. A unified framework of plant adaptive strategies to drought: Crossing scales and disciplines. Global Change Biology 24: 2929–2938. doi: 10.1111/gcb.14062. [DOI] [PubMed] [Google Scholar]
- Waisel Y, Eshel A, Kafkafi U. (eds) 2002. Plant roots: the hidden half, 3rd edn. New York: Marcel Dekker. [Google Scholar]
- Wang JF, Shi YJ, Ao YN, et al. 2019. Summer drought decreases Leymus chinensis productivity through constraining the bud, tiller and shoot production. Journal of Agronomy and Crop Science 205: 554–561. doi: 10.1111/jac.12354. [DOI] [Google Scholar]
- Wang N, Gao J, Zhang SQ.. 2017. Overcompensation or limitation to photosynthesis and root hydraulic conductance altered by rehydration in seedlings of sorghum and maize. The Crop Journal 5: 337–344. doi: 10.1016/j.cj.2017.01.005. [DOI] [Google Scholar]
- Wang XL, Wang JJ, Sun RH, et al. 2016. Correlation of the corn compensatory growth mechanism after post-drought rewatering with cytokinin induced by root nitrate absorption. Agricultural Water Management 166: 77–85. doi: 10.1016/j.agwat.2015.12.007. [DOI] [Google Scholar]
- Weiner J. 2004. Allocation, plasticity and allometry in plants. Perspectives in Plant Ecology, Evolution and Systematics 6: 207–215. doi: 10.1078/1433-8319-00083. [DOI] [Google Scholar]
- Wellstein C, Poschlod P, Gohlke A, et al. 2017. Effects of extreme drought on specific leaf area of grassland species: A meta-analysis of experimental studies in temperate and sub-Mediterranean systems. Global Change Biology 23: 2473–2481. doi: 10.1111/gcb.13662. [DOI] [PubMed] [Google Scholar]
- Wright IJ, Dong N, Maire V, et al. 2017. Global climatic drivers of leaf size. Science 357: 917–921. doi: 10.1126/science.aal4760. [DOI] [PubMed] [Google Scholar]
- Wyka TP, Bagniewska-Zadworna A, Kuczyńska A, et al. 2019. Drought-induced anatomical modifications of barley (Hordeum vulgare L.) leaves: An allometric perspective. Environmental and Experimental Botany 166: 103798. doi: 10.1016/j.envexpbot.2019.103798. [DOI] [Google Scholar]
- Xu ZZ, Zhou GS, Shimizu H.. 2009. Are plant growth and photosynthesis limited by pre-drought following rewatering in grass? Journal of Experimental Botany 60: 3737–3749. doi: 10.1093/jxb/erp216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu PJ, Liu SW, Zhang L, Li Q, Zhou DW.. 2018. Selecting the minimum data set and quantitative soil quality indexing of alkaline soils under different land uses in northeastern China. The Science of the Total Environment 61: 564–571. doi: 10.1016/j.scitotenv.2017.10.301. [DOI] [PubMed] [Google Scholar]
- Zhang B, Zhu JJ, Pan QM, et al. 2017. Grassland species respond differently to altered precipitation amount and pattern. Environmental and Experimental Botany 137: 166–176. doi: 10.1016/j.envexpbot.2017.02.006. [DOI] [Google Scholar]
- Zhang JW, Gao X, Zheng X, et al. 2021a. A high stem to leaf ratio reduced rainfall use efficiency under altered rainfall patterns in a semi-arid grassland in Northeast China. Plant Biology 23: 760–769. doi: 10.1111/plb.13278. [DOI] [PubMed] [Google Scholar]
- Zhang JW, Shen XJ, Mu BF, et al. 2021b. Moderately prolonged dry intervals between precipitation events promote production in Leymus chinensis in a semi-arid grassland of Northeast China. BMC Plant Biology 21: 147. doi: 10.1186/s12870-021-02920-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang JW, Wu XF, Shi YJ, et al. 2021c. A slight increase in soil pH benefits soil organic carbon and nitrogen storage in a semi-arid grassland. Ecological Indicators 130: 108037. doi: 10.1016/j.ecolind.2021.108037. [DOI] [Google Scholar]
- Zhang JT, Mu CS, Wang DL, Wang JF, Chen GX.. 2009. Shoot population recruitment from a bud bank over two seasons of undisturbed growth of Leymus chinensis. Botany 87: 1242–1249. doi: 10.1139/B09-080. [DOI] [Google Scholar]
- Zhu T. 2004. Biological ecology of Leymus chinensis. Jilin: Jilin Science and Technology Press. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.