

Abstract
Aims
Molecular hydrogen has been exhibited a protective function in heart diseases. Our previous study demonstrated that hydrogen‐rich saline (HRS) could scavenge free radicals selectively and alleviate the inflammatory response in the myocardial ischaemia/reperfusion (I/R) injury, but the underlying mechanism has not been fully clarified.
Methods and results
Adult (10 weeks) C57BL/6 male mice and neonatal rat cardiomyocytes were used to establish I/R and hypoxia/reoxygenation (H/R) injury models. I/R and H/R models were treated with HRS to classify the mechanisms of cardioproctective function. In this study, we found that miR‐124‐3p was significantly decreased in both I/R and H/R models, while it was partially ameliorated by HRS pretreatment. HRS treatment also alleviated ischaemia‐induced apoptotic cell death and increased cell viability during I/R process, whereas silencing expression of miR‐124‐3p abolished this protective effect. In addition, we identified calpain1 as a direct target of miR‐124‐3p, and up‐regulation of miR‐124‐3 produced both activity and expression of calpain1. It was also found that compared with the HRS group, overexpression of calpain1 increased caspase‐3 activities, promoted cleaved‐caspase3 and Bax protein expressions, and correspondingly decreased Bcl‐2, further reducing cell viability. These results illustrated that calpain1 overexpression attenuated protective effect of HRS on cardiomyocytes in H/R model.
Conclusions
The present study showed a protective effect of HRS on I/R injury, which may be associated with miR‐124‐3p–calpain1 signalling pathway.
Keywords: Hydrogen‐rich saline, Cardiomyocyte apoptosis, Myocardial ischaemia/reperfusion injury, miR‐124‐3p
Introduction
Ischaemic heart disease including myocardial infarction (MI) and myocardial injury is one of the leading causes of morbidity and mortality in China, with age‐standardized mortality increased by 20.6% from 1990. 1 Unrelieved ischaemia causes permanent damage to the myocardium during acute MI. Reperfusion therapy is the most common and effective treatment against ischaemic heart disease; however, the process of reperfusion after ischaemia causes ischaemia/reperfusion injury (I/R injury), which may induce sustained myocardial damage. 2 Hydrogen (H2) has been proved to be protective in ischaemia reperfusion injury of several organs, including brain, liver, transplanted ischaemic kidney and myocardium, 3 , 4 , 5 , 6 and even in experimental settings in humans, 7 but the accurate mechanism is not fully understood. Recently, it was reported that hydrogen‐rich saline (HRS) exhibits the protective effects through interfering miRNA expressions, and the research suggested that HRS could regulate miR‐21 and miR‐210 in brain I/R process, further offering a neuroprotective effect. 8 , 9
MicroRNAs (miRNAs) are endogenous RNAs that play regulatory roles in multicellular organism by targeting mRNAs for degradation or translational repression. 10 , 11 It has been found that miRNA participated in the process of myocardial I/R injury by regulating apoptosis‐related genes. Some miRNAs inhibited myocardial apoptosis; for example, overexpression of miR‐494 reduced I/R injury and exerted myocardial protection by regulating pro‐apoptotic gene PTEN, 12 whereas many miRNAs increased cell apoptosis, as reported that miR‐15b promoted cardiomyocyte apoptosis through inhibiting the expression of Bcl‐2 protein, promoted the release of mitochondrial cytochrome c, and increased the depolarization of mitochondrial membrane. 13
MiR‐124 was firstly reported being highly expressed in the central nervous system. 14 However, the critical roles of miR‐124 in the pathophysiology of various diseases are being progressively recognized, especially in regulating process of cell apoptosis. Several studies have suggested that miR‐124 possesses anti‐apoptosis effects, as proved that miR‐124 inhibits spinal neuronal apoptosis through binding to GCH1 15 and that miR‐124‐3p attenuates hyperphosphorylation of Tau protein‐induced apoptosis. 16 Conversely, inhibition of miR‐124 reduces cardiomyocyte apoptosis following MI via targeting signal transducer and activator of transcription 3 (STAT3). 17 However, it remains unclear whether miR‐124 was downstream regulated by HRS and which genes it targets during myocardial I/R injury.
Calpains belong to a family within the unique property of using protein cleavage to modify the activity or function of their substrate proteins, which was shown to be critical in numerous cellular functions, including cytoskeletal remodelling, cellular signalling, and apoptosis. 18 , 19 Previous studies uncovered the effects of calpain1 in heart diseases such as heart failure, pathological cardiac remodelling, and cardiac hypertrophy. 20 , 21 , 22 However, the function of calpain1 in I/R injury and its significance has not been fully demonstrated. It was found in the prediction using TargetScan software that calpain1 might be a potential target for miR‐124. Here, we hypothesized that miR‐124–calpain1 signalling exerts a vital regulatory effect on the protective function of HRS during I/R injury.
Methods
Animals
Adult C57BL/6 mice weighing 20–25 g were obtained from the Shanghai SLAC Laboratory Animal Co., Ltd and housed with free access to food and water under 12 h light–dark cycle and acclimated for 7 days prior to any experimental procedure. A total of 50 adult male mice were divided into five groups; among the 10 mice per group, heart tissues from five mice were used for the TTC/Evans staining and the others were collected and stored at −80°C until detection. All experimental procedures were approved by the Institutional Animal Care and Use Committee of the Shanghai Changzheng Hospital and were conducted in strict accordance with the National Institutes of Health guidelines for the use of experimental animals. The investigation conforms to the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH Publication No. 85‐23, revised 1985).
Hydrogen‐rich saline production
According to the procedure described in our previous studies, 23 , 24 hydrogen was dissolved in 0.9% saline for 6 h under high pressure (0.4 MPa) to a supersaturated level by using a hydrogen producing apparatus. HRS was stored under an atmospheric pressure at 4°C in an aluminium bag with no dead volume, sterilized by gamma radiation. To confirm the concentration of hydrogen, gas chromatography was performed. HRS was freshly prepared every week to ensure that the concentration was maintained at 0.6 mmol/L.
Isolation of neonatal rat cardiomyocytes
Neonatal rat cardiomyocytes (NRCMs) were isolated according to previous research. 25 , 26 Briefly, neonatal rats (1‐day‐old) were anaesthetized with carbon dioxide and sacrificed by cervical dislocation. The hearts were quickly taken and cut into pieces of ~1 mm3 in the pre‐cooled Hank's Balanced Salt Solution, and the blood clot around the heart tissue was removed as much as possible. Then cardiac tissues were transferred to a glass bottle and digested in the enzyme solution containing 0.125% trypsin and 0.1% type II collagenase. Next, the bottle was put into a preheated cell vibrator for 30 min with 0.22 g at 37°C. After digestion, the supernatant was collected and added into Dulbecco's modified Eagle's medium (DMEM)/F12 medium containing 10% foetal bovine serum (FBS). Fresh enzyme solution buffer was added into the remaining tissue pieces for another 20 min. The digestion procedure was repeated four times. After the last collection of all cell suspensions, the cells were filtered through a 70 μm filter and centrifuged at 112 g for 10 min, and the supernatant was discarded. The total cells were resuspended in DMEM/F12 medium containing 10% FBS and seeded in a cell culture dish. After 2 h of incubation, cardiac fibroblasts adhered to the culture dish while NRCM cells remained to suspend in medium. At this time, the cell suspension was collected and seeded into a new culture dish at a concentration of 5 × 105/mL, and BrdU was added to make the final concentration of 0.1 mmol/L. Finally, NRCM cells were cultured overnight for subsequent experiments.
In vitro hypoxia/reoxygenation injury
Hydrogen was diluted in normal culture medium to prepare 0.6 mmol/L of HRS culture medium as reported previously. 23 In order to maintain a supersaturated concentration, HRS culture medium was freshly prepared every week. The procedure of in vitro hypoxia/reoxygenation (H/R) injury was performed as previously published. 27 NRCM cells were cultured in both serum‐ and glucose‐free DMEM in a humidified environment with 95% N2 and 5% CO2 for hypoxic conditions. After 4 h of incubation, cells of the control group were transferred with regular culture medium while cells of the HRS group were cultured in HRS medium for 12 h of reoxygenation.
In vivo mice ischaemia/reperfusion model
Ten‐week‐old male C57BL/6 mice were used in in vivo experiments. The mice I/R injury model was established according to previous research. 28 , 29 Briefly, the mice were anaesthetized with sodium pentobarbital (50 mg/kg i.p.) and thoracotomy was performed between the second and third ribs. After the pericardium was incised, a 7–0 ligature was passed underneath the left anterior descending coronary artery (LAD) and was tied to produce an occlusion for 30 min of ischaemia. The HRS solution (10 mL/kg) or normal saline (10 mL/kg) was administered via intraperitoneal injection at 5 min prior to reperfusion. Then the ligature was removed and the chest was closed for 24 h of reperfusion. The mice of the sham operation group included all procedures except ligation of the LAD.
In vivo delivery of miR‐124‐3p inhibitor
After mice received sodium pentobarbital and thoracotomy was performed, miR‐124‐3p inhibitor was prepared for injection. The administered dosage for each mouse was 3 nmol, and every mouse received three injection points in left ventricular anterior wall. After 24 h of injection, mice received thoracotomy again, and then I/R model was established. After LAD was ligated for 30 min and reperfusion for 24 h, the TTC/Evans staining was performed and then the heart was harvested.
Assessment of lactate dehydrogenase and caspase‐3 activity
The release of lactate dehydrogenase (LDH) in cell culture medium was measured to detect the degree of myocardial injury. LDH activities were detected using LDH detection kits (Beyotime, China) according to the manufacturer's instructions. Caspase‐3 activity was determined using a commercial caspase‐3 activity assay kit (Promega Biotech Co., Ltd). Briefly, NRCMs were plated in 96‐well plates and exposed to H/R injury, and the solution was replaced with either regular medium or HRS medium before the reoxygenation process. Ninety‐six‐well plates containing cells were removed from the incubator, and 100 μL of Caspase‐Glo® 3 Reagent was added to each well. After being mixed softly for 1 min, the 96‐well plates were incubated at 37°C for another 1 h. The luminescence was then measured by luminometer.
Luciferase reporter assay and cell transfection
The full length of 3′‐untranslated region (3′‐UTR) calpain1 containing the predicted miR‐124‐3p binding sites were amplified by PCR and cloned into the downstream of the dual luciferase reporter gene in psiCHECK2 (Promega, USA) vectors. Site‐directed Gene Mutagenesis Kit (Beyotime) was used to generate recombinant calpain1‐mutation vector. 293A cells were cotransfected with wild‐type calpain1 (calpain1‐wt) or mutant‐type calpain1 (calpain1‐mut). The cells were also cotransfected with miR‐124‐3p mimics or miR‐negative control using the same methods. After 36 h of transfection, the luciferase activity was determined using the Dual Luciferase Reporter Gene Assay Kit (Beyotime). The Renilla luciferase activity was regarded as the normalization. Transfections were performed using Lipofectamine 3000 (Invitrogen) according to the manufacturer's instructions. miR‐124‐3p mimics and inhibitors (Guangzhou RiboBio Co., Ltd) were also transfected into NRCM cells using Lipo 3000 as a transfection reagent.
Quantitative real‐time RT–PCR of miRNA
RNA (including miRNA) was extracted using a TRIzol kit (Life Technologies) according to the product instructions. miRcute Plus miRNA Kit (Tiangen Biotech) was used for reverse transcription into cDNA. The housekeeping genes (U6) was used for normalization. The primer sequences used were as follows: miR‐124‐3p‐forward: 5′‐GCTTAAGGCACGCGG‐3′, miR‐124‐3p‐reverse: 5′‐GTGCAGGGTCCGAGG‐3′; U6‐forward: 5′‐CTCGCTTCGGCAGCACATATACT‐3′, U6‐reverse: 5′‐ACGCTTCACGAATTTGCGTGT‐3′.
Western blot
After different kinds of treatment, including miRNA transfection, adenovirus infection, and HRS pretreatment, NRCMs were exposed to H/R injury. After reoxygenation, cells were harvested in phosphate‐buffered saline and lysates were prepared by tissue lysis buffer. Protein concentration was determined by bicinchoninic acid protein assay. Equal amounts were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis gels and then transferred to a polyvinylidene difluoride (PVDF) membrane. The band density of specific proteins was quantified after normalization with the density of glyceraldehyde 3‐phosphate dehydrogenase.
Heart function evaluation
The heart function variables of C57BL/6 mice including ejection fraction (EF%), heart rate, left ventricular end‐diastolic volume (LVEDV), left ventricular end‐systolic volume (LVESV), and left ventricular posterior wall were measured by ultrasound (Vevo‐2100). EF was calculated as EF = [(LVEDV − LVESV)/LVEDV] × 100%.
TTC/Evans staining
At the end of the myocardial I/R protocol, the heart was removed immediately, and then ~0.5 mL of 4% Evans blue dye (Sigma) was injected into the aorta to show the non‐ischaemic area. Myocardial areas not stained with Evans blue were defined as risk areas (AARS). The heart was wrapped in polyethylene and frozen for 10 min to harden. After that, the heart was cut into seven transverse slices and incubated at room temperature in 1% TTC in phosphate‐buffered saline for 15 min to distinguish between viable (brick red) and infarcted (pale) myocardium. Then, the heart slices were fixed overnight in 10% neutral buffered formaldehyde for better colour contrast and digital imaging. Infarct size, AARS, and non‐risk area were quantified by the percentage of total myocardial area using the Image‐Pro Plus 5.0 software.
Assessment of creatine kinase‐MB and cardiac troponin I
Mouse blood samples were collected from the heart, and then blood were centrifuged at 1000 g for 5 min and stored at −80°C until detection. The levels of serum creatine kinase‐MB and cardiac troponin I were measured with ELISA Kit (Sigma and Abcam, USA) in accordance with the manufacturer's instructions. Briefly, 100 μL standard or samples were added to each well for 1.5 h at 37°C. Then, the liquid was removed and added with conjugate antibody working solution for 1 h at 37°C. Thereafter, the wells were washed three times before adding 100 μL of horseradish peroxidase‐avidin (1×) to each well for 1 h at 37°C. After washing for three times, substrate reagent was added into each well for 15 min at 37°C in the dark. At last, the reaction was stopped by adding top solution to each well and read at 450 nm within 5 min.
Cell viability assessment
A Cell Counting Kit‐8 (CCK‐8) assay (Beijing Solarbio Science & Technology Co., Ltd) was used to assess cell viability according to the protocol of the kit. Briefly, NRCMs were seeded into 96‐well plates and grown to 75% confluence. Following transfection, CCK‐8 reagent was added to each well and incubated for 1 h at 37°C. The absorbance of each well was measured at a wavelength of 450 nm on a microplate reader (Thermo Fisher Scientific, Inc.). For clonogenic assays, NRCMs were plated into six‐well plates at a density of 4000–8000 cells per well. After 24 h, cells were cultured in a hypoxia chamber (Billups‐Rothenberg, Inc.) at 37°C for the indicated time points. Cells were fixed with 10% formalin at room temperature for 10 min, followed by staining with 0.02% crystal violet at room temperature for 10 min. Then, cell numbers were counted under a dissection microscope.
Bioinformatics and statistical analysis
The miRNA targets predicted by computer‐aided algorithms were obtained from TargetScanMouse 7.1 (http://www.targetscan.org). Statistical analysis was performed using GraphPad Prism 5.01 software. All the data were presented as means ± standard error of the mean. Comparisons between the two groups were analysed using a Student's t‐test. A one‐way ANOVA followed by post hoc Tukey's test was used to analyse differences between multiple groups. P < 0.05 was considered as statistical significance.
Results
Hydrogen‐rich saline improves heart function after ischaemia/reperfusion and increases miRNA‐124‐3p expression
To determine the in vivo and in vitro protective effects of HRS, we first established mice I/R model and NRCM H/R model as described in the Methods section. As shown in Figure 1 A,B,E,G , HRS treatment significantly improved mice heart function after I/R injury. Furthermore, treatment of HRS solution alleviated the damage of hypoxia to cardiomyocytes. CCK‐8 assay revealed that cell viability was markedly increased while the expression of LDH in culture medium and cell caspase‐3 activity was both decreased after being exposed to HRS solution (Figure 2 A–C ). To clarify which miRNAs were involved in protective effect of HRS in myocardial I/R injury, real‐time quantitative PCR analysis was conducted and the results showed that the expression of miR‐124‐3p in cardiomyocyte was decreased both in I/R injury and in H/R injury, which was partially rescued by HRS treatment, indicating that miR‐124‐3p may exert potential role in HRS treatment.
Figure 1.
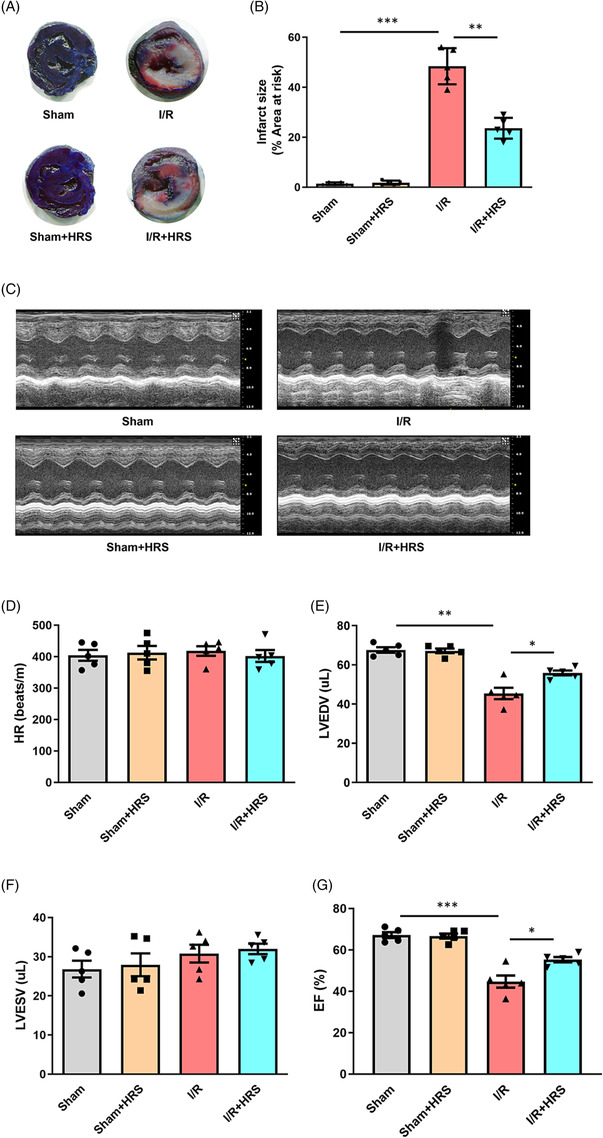
Hydrogen‐rich saline (HRS) improves heart function after ischaemia/reperfusion (I/R). (A) TTC/Evans staining of mice heart after I/R injury. (B) Infarct size of mice heart after I/R injury. (C) Representative images of ultrasound for mice heart function. (D–G) Heart rate (HR), left ventricular end‐diastolic volume (LVEDV), left ventricular end‐systolic volume (LVESV), and ejection fraction (EF) in different groups. *P < 0.05, **P < 0.01, and ***P < 0.001.
Figure 2.
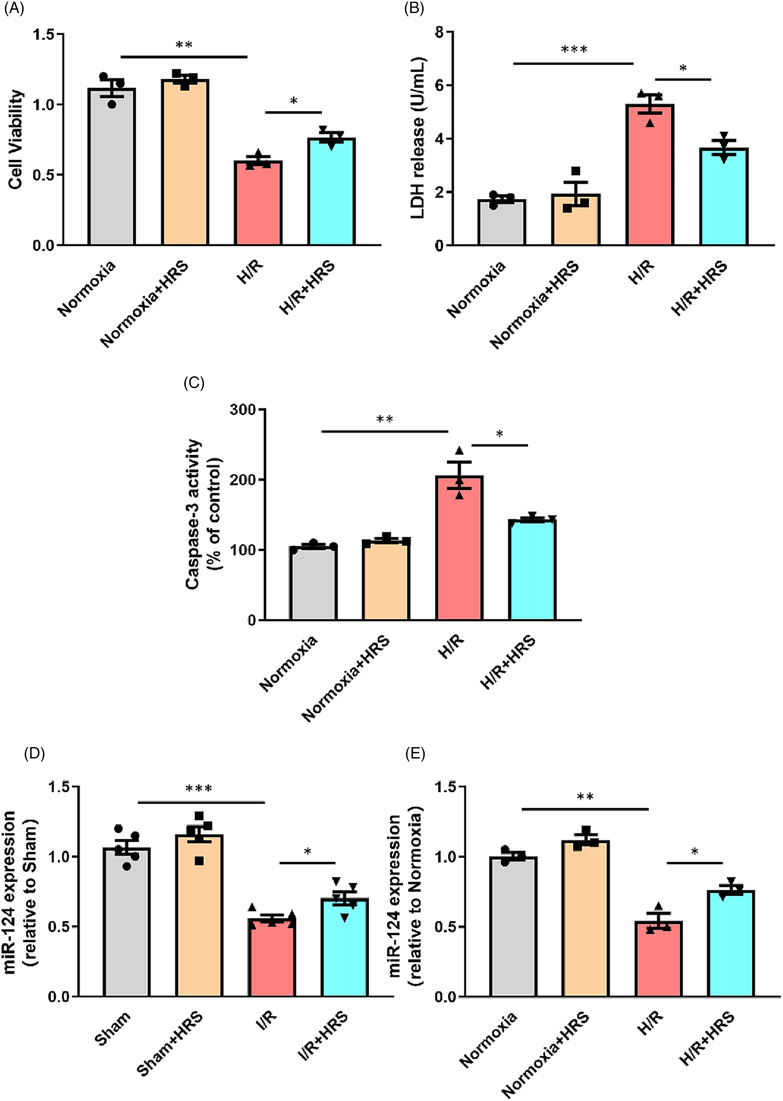
Hydrogen‐rich saline (HRS) decreases neonatal rat cardiomyocyte (NRCM) apoptosis in response to hypoxia/reoxygenation (H/R) injury and increases miRNA‐124‐3p expression. (A) Cell viability of NRCMs after H/R injury was measured. (B) Lactate dehydrogenase (LDH) activity of NRCM culture medium after H/R injury was measured. (C) Caspase‐3 activity of NRCMs after H/R injury was measured. (D, E) miRNA‐124‐3p expression was measured after ischaemia/reperfusion (I/R) and H/R injury. *P < 0.05, **P < 0.01, and ***P < 0.001.
Overexpression of miRNA‐124‐3p decreases neonatal rat cardiomyocyte apoptosis induced by hypoxia/reoxygenation injury
At present, many researchers have reported the function of miR‐124‐3p in regulating apoptosis. However, there are few studies focused on the role of miR‐124‐3p in the cardiovascular system. We found that miR‐124‐3p was decreased after I/R in vivo and H/R injury in vitro (Figure 2 D,E ). In that case, we then examined the effect of miR‐124‐3p on H/R‐induced myocardial apoptosis. As shown in Figure 3 A , the expression level of miR‐124‐3p was effectively elevated by miR‐124‐3p mimic. Caspase‐3 activities and LDH were reduced and cell viability was increased in the miRNA‐124‐3p overexpression group compared with the H/R group. Then, we tested the effect of miR‐124‐3p mimic on the apoptosis proteins, such as cleaved‐caspase3, Bax, and Bcl‐2 via western blot assay. As shown in Figure 3 , the protein expressions of cleaved‐caspase3 and Bax significantly declined after being transfected with miR‐124‐3p mimic, while Bcl‐2 protein was increased.
Figure 3.
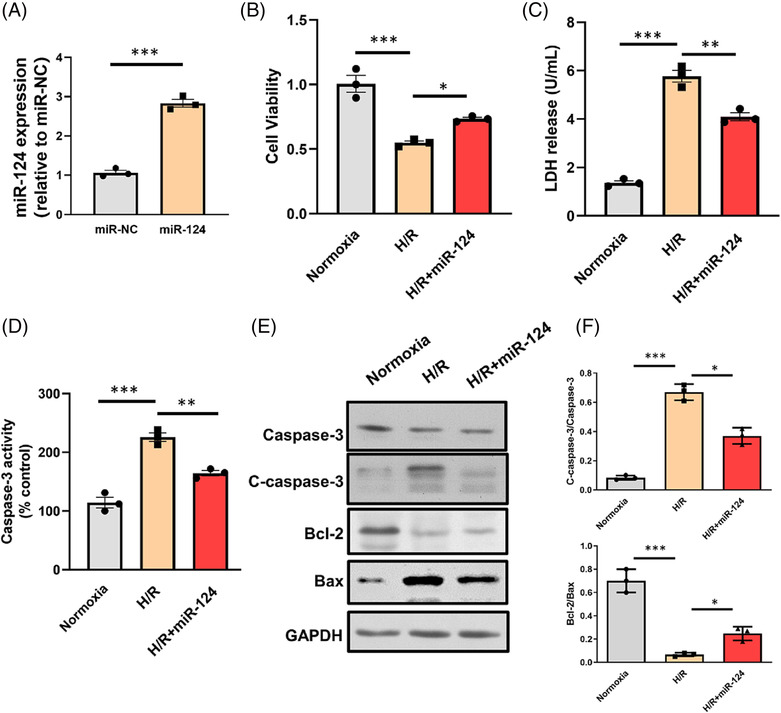
Overexpression of miRNA‐124‐3p decreases myocardial apoptosis induced by hypoxia/reoxygenation (H/R) injury. (A) Expression of miR‐124‐3p increased after transfection of miR‐124‐3p mimic in neonatal rat cardiomyocytes (NRCMs). (B) Cell viability of NRCMs after H/R injury was measured. (C) Lactate dehydrogenase (LDH) activity of NRCM culture medium after H/R injury was measured. (D) Caspase‐3 activity of NRCMs after H/R injury was measured. (E, F) Protein expressions of caspase‐3, Bcl‐2, and Bax were detected by western blot. *P < 0.05, **P < 0.01, and ***P < 0.001.
Down‐regulation of miR‐124‐3p abolishes heart protective effect of hydrogen‐rich saline
To explore the role of miR‐124‐3p in HRS‐induced heart protective effect, the expression of miR‐124‐3p was repressed by miR‐124 inhibitor. As shown in Figure 4 A , the expression level of miR‐124‐3p was effectively reduced by miR‐124‐3p inhibitor both in vivo and in vitro. The results of CCK‐8, LDH, and caspase‐3 activity assays showed that after H/R injury, LDH from culture medium and caspase‐3 activity in the HRS + miR‐124‐3p inhibitor group were remarkably increased, while cell viability was lower, than those in the HRS + miR‐negative‐control inhibitor group, and the differences were statistically significant. In vivo experiments showed that significant decreased myocardial infarct size was observed in the HRS treatment group relative to the I/R group. However, we found that in vivo delivery of miR‐124‐3p inhibitor into the myocardium markedly abolished HRS‐induced cardiac protection Figure 5 . In addition, the values of EF in HRS + miR‐124‐3p inhibitor transfected hearts were significantly lower than the HRS group after I/R.
Figure 4.
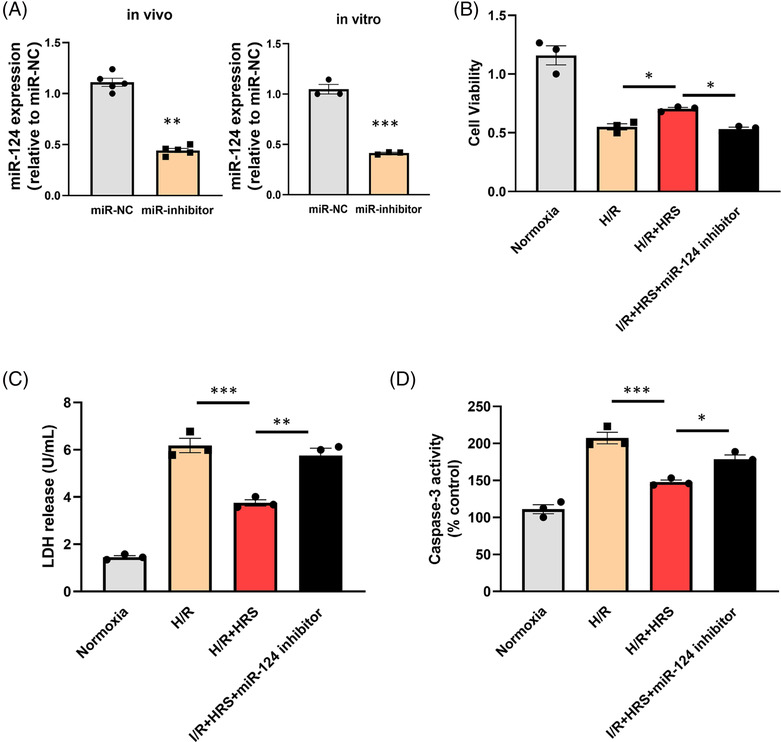
Down‐regulation of miR‐124‐3p abolishes heart protective effect of hydrogen‐rich saline (HRS) induced by hypoxia/reoxygenation (H/R) injury. (A) Expression of miR‐124‐3p decreased both in vivo and in vitro after being treated with miR‐124‐3p inhibitor. (B) Cell viability of neonatal rat cardiomyocytes (NRCMs) after H/R injury was measured. (C) Lactate dehydrogenase (LDH) activity of NRCM culture medium after H/R injury was measured. (D) Caspase‐3 activity of NRCMs after H/R injury was measured. *P < 0.05, **P < 0.01, and ***P < 0.001.
Figure 5.
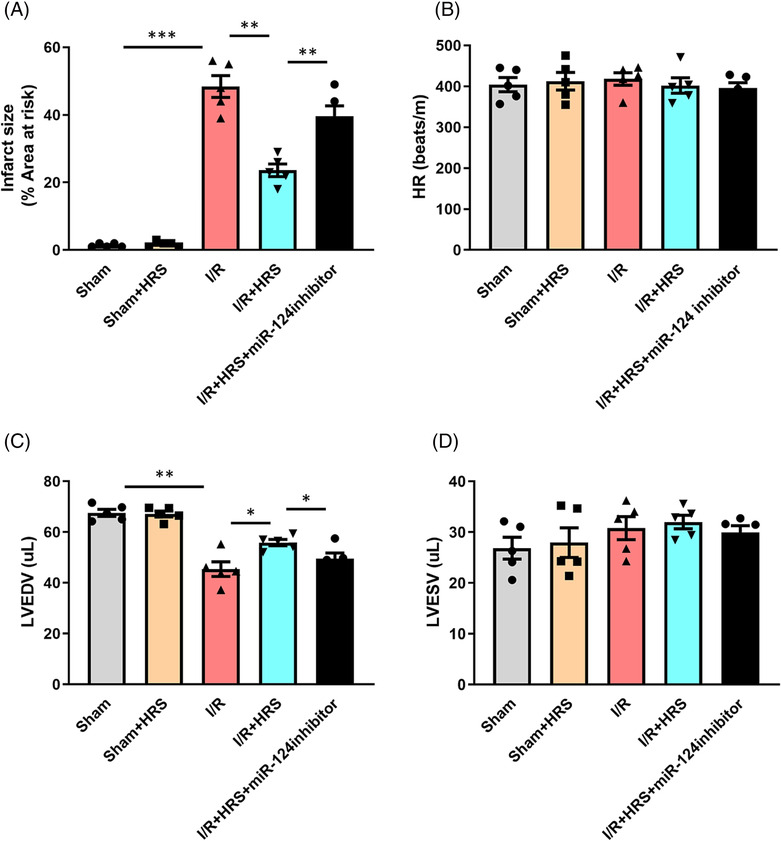
Down‐regulation of miR‐124‐3p abolishes heart protective effect of hydrogen‐rich saline (HRS). (A) Infarct size of mice heart after ischaemia/reperfusion (I/R) injury. (B) Heart rate (HR), (C) left ventricular end‐diastolic volume (LVEDV), (D) left ventricular end‐systolic volume (LVESV). *P < 0.05, **P < 0.01, and ***P < 0.001.
Calpain1 is a direct target of miR‐124‐3p
As a key cellular survival factor, calpain1 played crucial roles in regulating apoptosis not only in cancer cells 19 , 30 , 31 but also in cardiac pathophysiology. Previous studies demonstrated that calpain1 is important in promoting cardiomyocyte apoptosis under various pathological conditions. 32 , 33 , 34 In response to hypoxia, calpain1 is activated and contributes to I/R injury. However, the upstream regulatory molecules of calpain1 and the relationship between calpain1 and miRNAs had not been fully clarified. In this study, it was found that calpain1 might be a potential target gene for miR‐124‐3p analysed by TargetScan 7.1 software, and one miR‐124‐3p binding site was identified within the 3′‐UTR of calpain1 mRNA. In order to further confirm the interaction between miR‐124‐3p and calpain1 in cardiomyocyte, relevant assays were applied. As shown in Figure 6 C–E , transfection of miR‐124‐3p mimic decreased both expression of calpain1 and calpain1 activity in NRCMs. In order to further confirm the interaction between miR‐124‐3p and calpain1, the responses of the calpain1‐wt and calpain1‐mut to miR‐124‐3p and miR‐NC were examined by dual luciferase assay. The results showed that the luciferase activity was significantly decreased in the miR‐124‐3p mimic + calpain1‐wt group compared with the miR‐NC + calpain1‐wt group, while it had no significant changes in the miR‐124‐3p mimic + calpain1‐mut group (Figure 6 A,B ). These results demonstrated that miR‐124‐3p mimic could decrease the level of calpain1 through targeting the predicted sequence. All the data illustrated that miR‐124‐3p inhibited the expression of calpain1 in cardiomyocyte.
Figure 6.
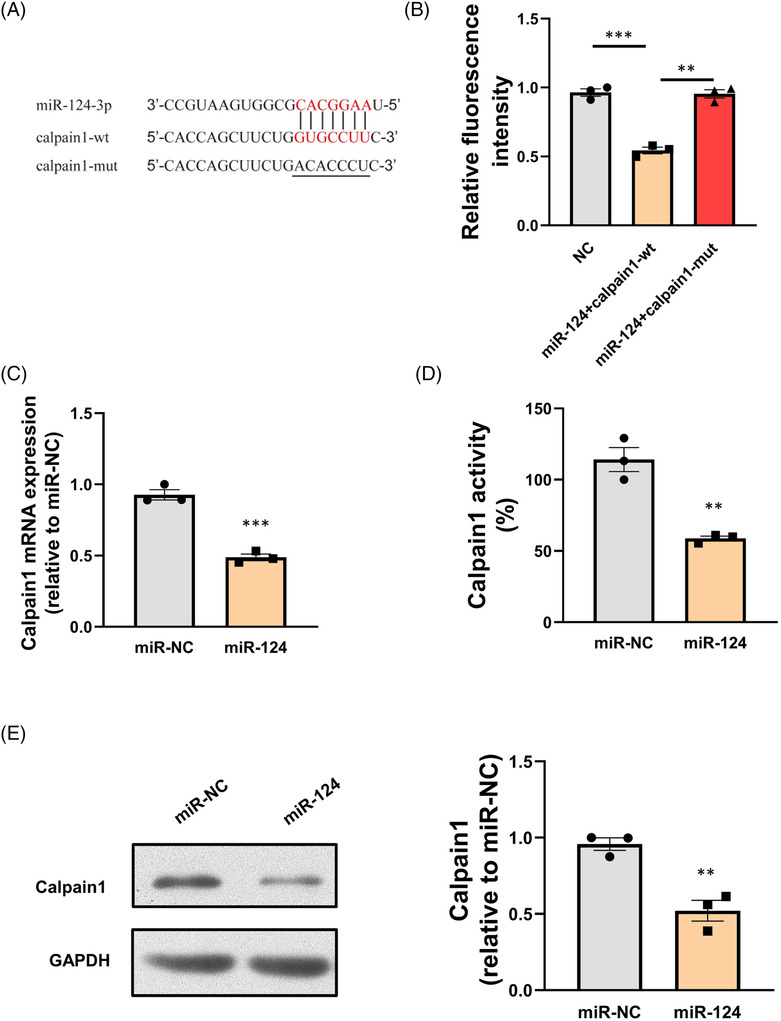
Calpain1 is a direct target of miR‐124‐3p. (A) The predicted binding sites for miR‐124‐3p in calpain1 mRNA. (B) MiR‐124‐3p significantly decreased the luciferase activity of wild‐type calpain1 (calpain1‐wt) but not mutant‐type calpain1 (calpain1‐mut) in 293A cells. (C) Expression of calpain1 mRNA decreased after overexpression of miR‐124‐3p. (D) Calpain1 activity decreased after overexpression of miR‐124‐3p. (E) Expression of calpain1 protein decreased after overexpression of miR‐124‐3p.
Calpain1 overexpression attenuates protective effect of hydrogen‐rich saline on cardiomyocytes
To further confirm the functional involvement of calpain1 in the regulation of apoptosis by HRS solution, both adenovirus of calpain1 and HRS were used in the following experiments. We found that ad‐calpain1 significantly increased the calpain1 expression in NRCMs. Furthermore, as shown in Figure 7 , caspase‐3 activities and LDH were increased and cell viability was reduced in the calpain1 overexpression group compared with the HRS group. Next, apoptosis‐related proteins including cleaved‐caspase3, Bax, and Bcl‐2 were examined via western blot assay. We found that compared with HRS‐treated NRCMs, the protein expressions of cleaved‐caspase3 and Bax significantly increased after calpain1 overexpression, while bcl‐2 protein was decreased. These results illustrated that calpain1 overexpression abolished protective effect of HRS on cardiomyocytes in H/R model.
Figure 7.
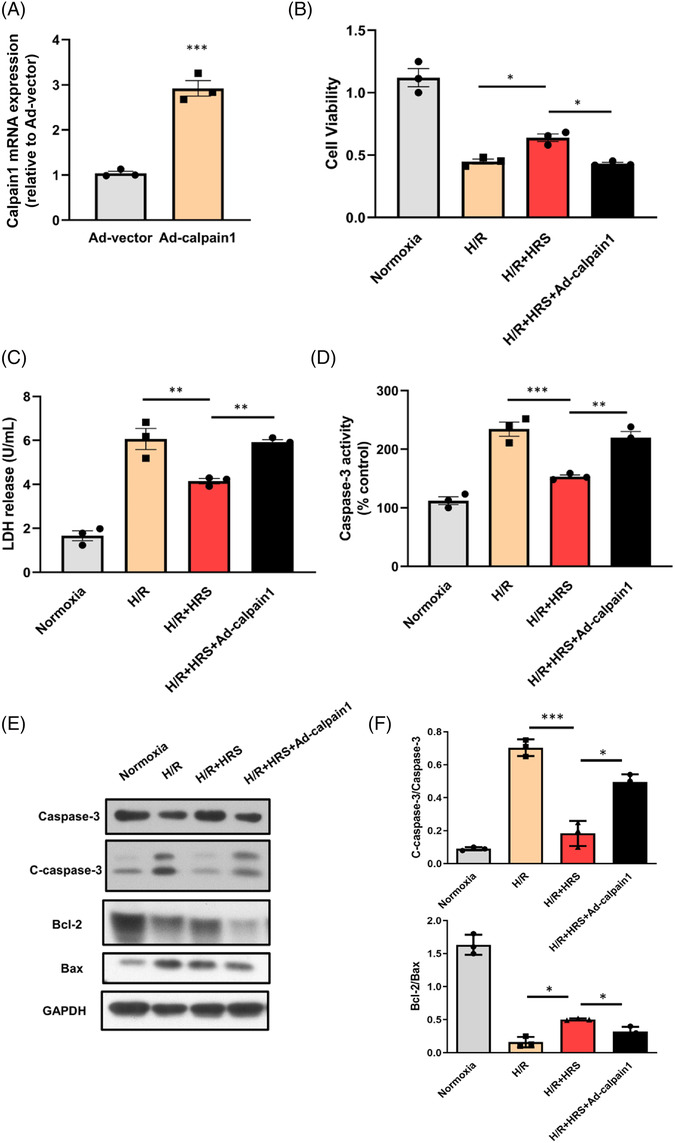
Calpain1 overexpression attenuates protective effect of hydrogen‐rich saline (HRS) on cardiomyocytes. (A) Expression of calpain1 increased after being treated with Ad‐calpain1 in neonatal rat cardiomyocytes (NRCMs). (B) Cell viability of NRCMs after hypoxia/reoxygenation (H/R) injury was measured. (C) Lactate dehydrogenase (LDH) activity of NRCM culture medium after H/R injury was measured. (D) Caspase‐3 activity of NRCMs after H/R injury was measured. (E, F) Protein expressions of caspase‐3, Bcl‐2, and Bax were detected by western blot. *P < 0.05, **P < 0.01, and ***P < 0.001.
Discussion
HRS has been shown to possess anti‐inflammatory, anti‐apoptosis, and anti‐oxidative stress effects, further exhibiting a protective function in heart diseases, 6 , 35 , 36 but the accurate mechanism underlying this protection is not very clear. Recently, it was reported that the protective effect of HRS on acute cerebral I/R was partially involved in regulating the expression of miR‐21 and miR‐210. Several studies have investigated that HRS treatment up‐regulated the expression of miR‐21 and down‐regulated the expression of miR‐1 in the myocardium. We also reported the miR‐1 in the protective effect of hydrogen sulfide against cardiomyocyte apoptosis induced by I/R. 37 Additionally, HRS pretreatment altered levels of a total of 64 intestinal I/R‐responsive miRNAs, including miR‐199a‐3p, miR‐296‐5p, miR‐5126, and miR‐6538. 8 , 9 However, the miRNAs regulated by HRS in myocardial I/R injury remain unknown. In the present study, we focused on the expression changes of miRNAs and its potential function during HRS treatment in mice I/R model. Our data illustrated that miR‐124 was decreased after I/R injury, while HRS treatment partly reversed this effect, indicating that miR‐124 may take part in the HRS‐induced protective effect.
MiR‐124 is highly expressed in normal brain of human. 38 Previous studies mainly focused on the role of miR‐124 in neuronal functions and injuries. 39 , 40 , 41 Studies have shown that miR‐124 plays an important role in the neuroprotection following I/R injury, suggesting that it may be involved in the pathophysiology of heart I/R injury. Recently, many researchers have illustrated that miR‐124 is also involved in the regulation of heart diseases. For example, it was found that cardiac expression of miR‐124 was increased in patients and mice with heart failure, and miR‐124 is an important negative regulator of cardiac angiogenesis and function, likely by suppressing the expression of CD151 in heart cells. 42 Moreover, doxorubicin has been shown to promote the generation of reactive oxygen species and cardiomyocyte apoptosis, while miR‐124 could attenuate this oxidative stress injury by inhibiting p66Shc signalling pathway. 43 In addition, miR‐124 has been shown to possess regulatory functions in MI. By using the bioinformatics network analysis of microarray data, miR‐124 was identified as a critical modulator in MI. 44 It was found that miR‐124 was increased in a mouse MI model. 17 Furthermore, miR‐124 overexpression aggravated hypoxia injury by decreasing viability, migration, and invasion, as well as promoting apoptosis. 45 Nevertheless, the function of miR‐124‐3p in the pathogenesis of I/R remains unclear. A recent study 46 showed that the expression of miR‐124 in the infarct zone was increased relative to the remote zone and miR‐124 promoted cell apoptosis in a rat MI model. In this study, however, we found that miR‐124 decreased in mice I/R model with 30 min of ischaemia and reperfusion for 24 h compared with the sham group, indicating that the miR‐124 level might fluctuate with different reperfusion durations. However, further research is needed to fully understand the effects of HRS treatment on the expression profile of miRNAs in the myocardium.
It was proved that miR‐124 played an important role in modulating the process of cell apoptosis and related to several target genes, such as GCH1, EDNRB, and PDCD6. 47 , 48 , 49 In this study, it was found that, for the first time, calpain1 might be another direct target gene for miR‐124 analysed by TargetScan software, and the interaction between miR‐124 and calpain1 in cardiomyocyte was confirmed through relevant assays. Previous studies showed that calpain1 not only participated in the progress of apoptosis but also possessed both anti‐apoptosis and pro‐apoptosis effects. 32 , 50 , 51 In the present study, we found that both the activity and protein expression of calpain1 increased after H/R injury, and it could be reversed by HRS treatment. In addition, it was showed that ad‐calpain1 significantly increased the calpain1 expression and caspase‐3 activities were increased in the calpain1 overexpression group compared with the HRS group. Furthermore, we found that compared with HRS‐treated NRCMs, the apoptosis‐related protein expressions of cleaved‐caspase3 and Bax significantly increased after calpain1 overexpression, while Bcl‐2 protein was decreased. These results illustrated that calpain1 overexpression abolished protective effect of HRS on cardiomyocytes in H/R injury, further supporting the role of calpain1 as a pro‐apoptotic factor in myocardial H/R injury. In this study, we focused on the effects of miR‐124 on the expression of calpain1. Excluding calpain1, miR‐124 has several putative target molecules including tumour necrosis factor‐α, interleukin‐6, hypoxia‐inducible factor‐1α, and STAT3 according to the papers published previously, but these additional target molecules were not investigated in this study. It is possible that these target molecules could contribute to the protective effect of HRS. However, few studies focused on the relationship between HRS–miR‐124 and target molecules, and further investigations are needed to explore the effects of miR‐124 modulation on these potential targets and to determine their role in the protective effect of HRS.
Several study limitations should be considered in this study. First, the present study did not include the long‐term effects of HRS treatment on cardiac function and overall survival after I/R injury, which needs further investigation. Second, the effect of miR‐124 cardiac‐specific overexpression on the I/R mouse model was not investigated in this study. Third, the study mainly focused on the miR‐124‐3p and calpain1 signalling pathway, but other potential signalling contributing to the cardioprotective effects of HRS might not have been considered.
In conclusion, our study demonstrated that HRS treatment could protect cardiomyocytes from I/R and H/R injury by regulating the miR‐124‐3p/calpain1 pathway. These findings provide new insights into the underlying mechanisms of HRS‐mediated cardioprotection and suggest that HRS, miR‐124‐3p, and calpain1 may be potential therapeutic targets for the clinical treatment of myocardial I/R injury in human.
Conflict of interest
None declared.
Funding
This work was supported by the National Natural Science Foundation of China (81300102 and 81770244), the University Basic Medical Research Project (2022QN084), the Cultivation of Teaching Achievement Project (JXPY2021C14), the Sailing Talent Program (2019‐QH‐29), and the Shanghai Education Development Foundation and Shanghai Municipal Education Commission (Shuguang Program, 21SG37).
Xue, X. , Xi, W. , Li, W. , Xiao, J. , Wang, Z. , and Zhang, Y. (2023) Hydrogen‐rich saline alleviates cardiomyocyte apoptosis by reducing expression of calpain1 via miR‐124‐3p. ESC Heart Failure, 10: 3077–3090. 10.1002/ehf2.14492.
Xiaofei Xue, Wang Xi, and Wei Li contributed equally to this study.
Contributor Information
Wang Xi, Email: anjiechen@yeah.net.
Zhinong Wang, Email: wangzn007@smmu.edu.cn.
Yufeng Zhang, Email: zyflwj@smmu.edu.cn.
References
- 1. Zhou M, Wang H, Zeng X, Yin P, Zhu J, Chen W, et al. Mortality, morbidity, and risk factors in China and its provinces, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2019;394:1145‐1158. doi: 10.1016/S0140-6736(19)30427-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Heusch G. Myocardial ischaemia–reperfusion injury and cardioprotection in perspective. Nat Rev Cardiol 2020;17:773‐789. doi: 10.1038/s41569-020-0403-y [DOI] [PubMed] [Google Scholar]
- 3. Chu X, Cao L, Yu Z, Xin D, Li T, Ma W, et al. Hydrogen‐rich saline promotes microglia M2 polarization and complement‐mediated synapse loss to restore behavioral deficits following hypoxia‐ischemic in neonatal mice via AMPK activation. J Neuroinflammation 2019;16:104. doi: 10.1186/s12974-019-1488-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Li S, Fujino M, Ichimaru N, Kurokawa R, Hirano S, Mou L, et al. Molecular hydrogen protects against ischemia‐reperfusion injury in a mouse fatty liver model via regulating HO‐1 and Sirt1 expression. Sci Rep 2018;8:14019. doi: 10.1038/s41598-018-32411-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Liu Y, Yang L, Tao K, Vizcaychipi MP, Lloyd DM, Sun X, et al. Protective effects of hydrogen enriched saline on liver ischemia reperfusion injury by reducing oxidative stress and HMGB1 release. BMC Gastroenterol 2014;14:12. doi: 10.1186/1471-230X-14-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Yao L, Chen H, Wu Q, Xie K. Hydrogen‐rich saline alleviates inflammation and apoptosis in myocardial I/R injury via PINK‐mediated autophagy. Int J Mol Med 2019;44:1048‐1062. doi: 10.3892/ijmm.2019.4264 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Fontanari P, Badier M, Guillot C, Tomei C, Burnet H, Gardette B, et al. Changes in maximal performance of inspiratory and skeletal muscles during and after the 7.1‐MPa Hydra 10 record human dive. Eur J Appl Physiol 2000;81:325‐328. doi: 10.1007/s004210050050 [DOI] [PubMed] [Google Scholar]
- 8. Li Q, Yu P, Zeng Q, Luo B, Cai S, Hui K, et al. Neuroprotective effect of hydrogen‐rich saline in global cerebral ischemia/reperfusion rats: Up‐regulated Tregs and down‐regulated miR‐21, miR‐210 and NF‐κB expression. Neurochem Res 2016;41:2655‐2665. doi: 10.1007/s11064-016-1978-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Yao W, Lin X, Han X, Zeng L, Guo A, Guan Y, et al. MicroRNA files in the prevention of intestinal ischemia/reperfusion injury by hydrogen rich saline. Biosci Rep 2020;40: doi: 10.1042/BSR20191043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Bartel DP. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004;116:281‐297. doi: 10.1016/S0092-8674(04)00045-5 [DOI] [PubMed] [Google Scholar]
- 11. Farh KK, Grimson A, Jan C, Lewis BP, Johnston WK, Lim LP, et al. The widespread impact of mammalian MicroRNAs on mRNA repression and evolution. Science 2005;310:1817‐1821. doi: 10.1126/science.1121158 [DOI] [PubMed] [Google Scholar]
- 12. Wang X, Zhang X, Ren XP, Chen J, Liu H, Yang J, et al. MicroRNA‐494 targeting both proapoptotic and antiapoptotic proteins protects against ischemia/reperfusion‐induced cardiac injury. Circulation 2010;122:1308‐1318. doi: 10.1161/CIRCULATIONAHA.110.964684 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Liu L, Zhang G, Liang Z, Liu X, Li T, Fan J, et al. MicroRNA‐15b enhances hypoxia/reoxygenation‐induced apoptosis of cardiomyocytes via a mitochondrial apoptotic pathway. Apoptosis 2014;19:19‐29. doi: 10.1007/s10495-013-0899-2 [DOI] [PubMed] [Google Scholar]
- 14. Li X, Yu Z, Li Y, Liu S, Gao C, Hou X, et al. The tumor suppressor miR‐124 inhibits cell proliferation by targeting STAT3 and functions as a prognostic marker for postoperative NSCLC patients. Int J Oncol 2015;46:798‐808. doi: 10.3892/ijo.2014.2786 [DOI] [PubMed] [Google Scholar]
- 15. Yuan S, Wang YX, Gong PH, Meng CY. MiR‐124 inhibits spinal neuronal apoptosis through binding to GCH1. Eur Rev Med Pharmacol Sci 2019;23:4564‐4574. doi: 10.26355/eurrev_201906_18032 [DOI] [PubMed] [Google Scholar]
- 16. Kang Q, Xiang Y, Li D, Liang J, Zhang X, Zhou F, et al. MiR‐124‐3p attenuates hyperphosphorylation of Tau protein‐induced apoptosis via caveolin‐1‐PI3K/Akt/GSK3β pathway in N2a/APP695swe cells. Oncotarget 2017;8:24314‐24326. doi: 10.18632/oncotarget.15149 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. He F, Liu H, Guo J, Yang D, Yu Y, Yu J, et al. Inhibition of microRNA‐124 reduces cardiomyocyte apoptosis following myocardial infarction via targeting STAT3. Cell Physiol Biochem 2018;51:186‐200. doi: 10.1159/000495173 [DOI] [PubMed] [Google Scholar]
- 18. Baudry M, Bi X. Calpain‐1 and calpain‐2: The yin and yang of synaptic plasticity and neurodegeneration. Trends Neurosci 2016;39:235‐245. doi: 10.1016/j.tins.2016.01.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Ma D, Fang J, Liu Y, Song JJ, Wang YQ, Xia J, et al. High level of calpain1 promotes cancer cell invasion and migration in oral squamous cell carcinoma. Oncol Lett 2017;13:4017‐4026. doi: 10.3892/ol.2017.5970 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Wang Y, Chen B, Huang CK, Guo A, Wu J, Zhang X, et al. Targeting calpain for heart failure therapy: Implications from multiple murine models. JACC Basic Transl Sci 2018;3:503‐517. doi: 10.1016/j.jacbts.2018.05.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Kunkel GH, Kunkel CJ, Ozuna H, Miralda I, Tyagi SC. TFAM overexpression reduces pathological cardiac remodeling. Mol Cell Biochem 2019;454:139‐152. doi: 10.1007/s11010-018-3459-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Han Q, Liu Q, Zhang H, Lu M, Wang H, Tang F, et al. Simvastatin improves cardiac hypertrophy in diabetic rats by attenuation of oxidative stress and inflammation induced by calpain‐1‐mediated activation of nuclear factor‐κB (NF‐κB). Med Sci Monit 2019;25:1232‐1241. doi: 10.12659/MSM.913244 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Zhang Y, Sun Q, He B, Xiao J, Wang Z, Sun X. Anti‐inflammatory effect of hydrogen‐rich saline in a rat model of regional myocardial ischemia and reperfusion. Int J Cardiol 2011;148:91‐95. doi: 10.1016/j.ijcard.2010.08.058 [DOI] [PubMed] [Google Scholar]
- 24. He B, Zhang Y, Kang B, Xiao J, Xie B, Wang Z. Protection of oral hydrogen water as an antioxidant on pulmonary hypertension. Mol Biol Rep 2013;40:5513‐5521. doi: 10.1007/s11033-013-2653-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. du Pre BC, Dierickx P, Crnko S, Doevendans PA, Vos MA, Geijsen N, et al. Neonatal rat cardiomyocytes as an in vitro model for circadian rhythms in the heart. J Mol Cell Cardiol 2017;112:58‐63. doi: 10.1016/j.yjmcc.2017.08.009 [DOI] [PubMed] [Google Scholar]
- 26. Quan D, Huang H. In vitro study of the effects of reprogramming neonatal rat fibroblasts transfected with TBX18 on spontaneous beating in neonatal rat cardiomyocytes. Mol Med Rep 2018;18:5520‐5526. doi: 10.3892/mmr.2018.9566 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Fan Z, Gao Y, Huang Z, Xue F, Wu S, Yang J, et al. Protective effect of hydrogen‐rich saline on pressure overload‐induced cardiac hypertrophyin rats: Possible role of JAK‐STAT signaling. BMC Cardiovasc Disord 2018;18:32. doi: 10.1186/s12872-018-0773-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Zhu L, Xu C, Huo X, Hao H, Wan Q, Chen H, et al. The cyclooxygenase‐1/mPGES‐1/endothelial prostaglandin EP4 receptor pathway constrains myocardial ischemia‐reperfusion injury. Nat Commun 2019;10:1888. doi: 10.1038/s41467-019-09492-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Huang ZQ, Xu W, Wu JL, Lu X, Chen XM. MicroRNA‐374a protects against myocardial ischemia‐reperfusion injury in mice by targeting the MAPK6 pathway. Life Sci 2019;232:116619. doi: 10.1016/j.lfs.2019.116619 [DOI] [PubMed] [Google Scholar]
- 30. Huang Q, Chen H, Wang F, Brost BC, Li J, Gao Y, et al. Reduced syncytin‐1 expression in choriocarcinoma BeWo cells activates the calpain1‐AIF‐mediated apoptosis, implication for preeclampsia. Cell Mol Life Sci 2014;71:3151‐3164. doi: 10.1007/s00018-013-1533-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Wang Y, Hersheson J, Lopez D, Hammer M, Liu Y, Lee KH, et al. Defects in the CAPN1 gene result in alterations in cerebellar development and cerebellar ataxia in mice and humans. Cell Rep 2016;16:79‐91. doi: 10.1016/j.celrep.2016.05.044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Yan M, Chen K, He L, Li S, Huang D, Li J. Uric acid induces cardiomyocyte apoptosis via activation of calpain‐1 and endoplasmic reticulum stress. Cell Physiol Biochem 2018;45:2122‐2135. doi: 10.1159/000488048 [DOI] [PubMed] [Google Scholar]
- 33. Zheng D, Wang G, Li S, Fan GC, Peng T. Calpain‐1 induces endoplasmic reticulum stress in promoting cardiomyocyte apoptosis following hypoxia/reoxygenation. Biochim Biophys Acta 2015;1852:882‐892. doi: 10.1016/j.bbadis.2015.01.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Blunt BC, Creek AT, Henderson DC, Hofmann PA. H2O2 activation of HSP25/27 protects desmin from calpain proteolysis in rat ventricular myocytes. Am J Physiol Heart Circ Physiol 2007;293:H1518‐H1525. doi: 10.1152/ajpheart.00269.2006 [DOI] [PubMed] [Google Scholar]
- 35. LeBaron TW, Kura B, Kalocayova B, Tribulova N, Slezak J. A new approach for the prevention and treatment of cardiovascular disorders. Molecular hydrogen significantly reduces the effects of oxidative stress. Molecules 2019;24:2076. doi: 10.3390/molecules24112076 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Gao Y, Yang H, Fan Y, Li L, Fang J, Yang W. Hydrogen‐rich saline attenuates cardiac and hepatic injury in doxorubicin rat model by inhibiting inflammation and apoptosis. Mediators Inflamm 2016;2016:1320365. doi: 10.1155/2016/1320365 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Kang B, Hong J, Xiao J, Zhu X, Ni X, Zhang Y, et al. Involvement of miR‐1 in the protective effect of hydrogen sulfide against cardiomyocyte apoptosis induced by ischemia/reperfusion. Mol Biol Rep 2014;41:6845‐6853. doi: 10.1007/s11033-014-3570-2 [DOI] [PubMed] [Google Scholar]
- 38. Landgraf P, Rusu M, Sheridan R, Sewer A, Iovino N, Aravin A, et al. A mammalian microRNA expression atlas based on small RNA library sequencing. Cell 2007;129:1401‐1414. doi: 10.1016/j.cell.2007.04.040 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Veremeyko T, Kuznetsova IS, Dukhinova M, W. Y. Yung A, Kopeikina E, Barteneva NS, et al. Neuronal extracellular microRNAs miR‐124 and miR‐9 mediate cell‐cell communication between neurons and microglia. J Neurosci Res 2019;97:162‐184. doi: 10.1002/jnr.24344 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Geng L, Liu W, Chen Y. miR‐124‐3p attenuates MPP+‐induced neuronal injury by targeting STAT3 in SH‐SY5Y cells. Exp Biol Med (Maywood) 2017;242:1757‐1764. doi: 10.1177/1535370217734492 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Cheng LC, Pastrana E, Tavazoie M, Doetsch F. miR‐124 regulates adult neurogenesis in the subventricular zone stem cell niche. Nat Neurosci 2009;12:399‐408. doi: 10.1038/nn.2294 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Zhao Y, Yan M, Chen C, Gong W, Yin Z, Li H, et al. MiR‐124 aggravates failing hearts by suppressing CD151‐facilitated angiogenesis in heart. Oncotarget 2018;9:14382‐14396. doi: 10.18632/oncotarget.24205 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Liu Y, Li Y, Ni J, Shu Y, Wang H, Hu T. MiR‐124 attenuates doxorubicin‐induced cardiac injury via inhibiting p66Shc‐mediated oxidative stress. Biochem Biophys Res Commun 2020;521:420‐426. doi: 10.1016/j.bbrc.2019.10.157 [DOI] [PubMed] [Google Scholar]
- 44. Li Y, He XN, Li C, Gong L, Liu M. Identification of candidate genes and microRNAs for acute myocardial infarction by weighted gene coexpression network analysis. Biomed Res Int 2019;2019:5742608. doi: 10.1155/2019/5742608 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Ghafouri‐Fard S, Shoorei H, Bahroudi Z, Abak A, Majidpoor J, Taheri M. An update on the role of miR‐124 in the pathogenesis of human disorders. Biomed Pharmacother 2021;135:111198. doi: 10.1016/j.biopha.2020.111198 [DOI] [PubMed] [Google Scholar]
- 46. Liu BF, Chen Q, Zhang M, Zhu YK. MiR‐124 promotes ischemia‐reperfusion induced cardiomyocyte apoptosis by targeting sphingosine kinase 1. Eur Rev Med Pharmacol Sci 2019;23:7049‐7058. doi: 10.26355/eurrev_201908_18747 [DOI] [PubMed] [Google Scholar]
- 47. Fu W, Wu X, Yang Z, Mi H. The effect of miR‐124‐3p on cell proliferation and apoptosis in bladder cancer by targeting EDNRB. Arch Med Sci 2019;15:1154‐1162. doi: 10.5114/aoms.2018.78743 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Wang S, Wu G, Han Y, Song P, Chen J, Wu Y, et al. miR‐124 regulates STAT3‐mediated cell proliferation, migration and apoptosis in bladder cancer. Oncol Lett 2018;16:5875‐5881. doi: 10.3892/ol.2018.9341 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Yuan L, Li S, Zhou Q, Wang D, Zou D, Shu J, et al. MiR‐124 inhibits invasion and induces apoptosis of ovarian cancer cells by targeting programmed cell death 6. Oncol Lett 2017;14:7311‐7317. doi: 10.3892/ol.2017.7157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Liu ZF, Zheng D, Fan GC, Peng T, Su L. Heat stress prevents lipopolysaccharide‐induced apoptosis in pulmonary microvascular endothelial cells by blocking calpain/p38 MAPK signalling. Apoptosis 2016;21:896‐904. doi: 10.1007/s10495-016-1263-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Li H, Chen LP, Wang T, Wang SG, Liu JH. Calpain inhibition improves erectile function in diabetic mice via upregulating endothelial nitric oxide synthase expression and reducing apoptosis. Asian J Androl 2018;20:342‐348. doi: 10.4103/aja.aja_63_17 [DOI] [PMC free article] [PubMed] [Google Scholar]