

Abstract
Overlapping genes are widely prevalent; however, their expression and consequences are poorly understood. Here, we describe and functionally characterize a novel zyx-1 overlapping gene, azyx-1, with distinct regulatory functions in Caenorhabditis elegans. We observed conservation of alternative open reading frames (ORFs) overlapping the 5′ region of zyxin family members in several animal species, and find shared sites of azyx-1 and zyxin proteoform expression in C. elegans. In line with a standard ribosome scanning model, our results support cis regulation of zyx-1 long isoform(s) by upstream initiating azyx-1a. Moreover, we report on a rare observation of trans regulation of zyx-1 by azyx-1, with evidence of increased ZYX-1 upon azyx-1 overexpression. Our results suggest a dual role for azyx-1 in influencing zyx-1 proteoform heterogeneity and highlight its impact on C. elegans muscular integrity and locomotion.
Overlapping genes are widely prevalent but their expression and consequences are poorly understood. This study shows that the C. elegans zyxin locus, zyx-1, contains a small ORF, azyx-1, which has a dual cis- and trans-acting role in regulating zyx-1 isoform diversity.
Introduction
Zyxins belong to a subfamily of conserved LIM domain-containing proteins found across eukaryotes and characterized for their role in cell-ECM (extra-cellular matrix) adhesion and cytoskeleton organization [1–4]. Characterized by a proline-rich N-terminus and 3 consecutive LIM domains in their C-terminal region [1,3,5], zyxins regulate actin assembly and remodeling, as well as cell motility [6–9]. In line with this, human zyxin is implicated in stretch-induced gene expression changes via active nuclear translocation [10]. Moreover, it also promotes apoptosis in response to DNA damage [11].
While multiple distinct zyxin proteins are present in vertebrates (for example, Lpp, Trip6, and Zyx), Caenorhabditis elegans contains a unique zyxin gene, zyx-1, with 5 annotated protein isoforms, of which isoforms a and b are predominantly expressed [5,12,13]. There are anatomical differences in isoform expression, with ZYX-1b observed in body wall muscle, pharynx, vulva, spermathecae, and multiple neurons, whereas ZYX-1a mainly localizes to tail phasmid neurons and uterine muscle, and weakly so, body wall muscle [13]. In C. elegans, zyx-1 has been postulated to have a minor role in reproduction, however, the mechanism(s) and isoform(s) involved remain elusive [14,15]. Beyond this, C. elegans zyx-1 is hypothesized to be functionally analogous to vertebrate zyxin, with LIM domains acting as mechanical stabilizers at focal adhesions, and the proline-rich N-terminus involved in sensing muscle cell damage [12].
Previous studies revealed that only the ZYX-1b isoform regulates synapse maintenance and development, while in the context of a dystrophic mutant background, the longer ZYX-1a isoform partially rescues muscle degeneration in an ATN-1-dependent manner, highlighting how not only expression patterns, but also molecular functions are isoform-dependent for C. elegans zyxin [12,13]. At a gene-regulatory level, this use of alternative splicing products and functional diversification of proteoforms for C. elegans zyxin is in line with observations made for several LIM domain proteins across eukaryotes [16].
In general, alternative and overlapping open reading frames (ORFs) arising out of polycistronic mRNA can contribute to posttranscriptional regulation [17–19]. A recent community-wide effort for annotation of such genomic loci categorized overlapping genes based on their initiation and termination codon with respect to the main coding sequence of a given transcript [20]. Of these, ORFs with an upstream start site (upstream or upstream overlapping; uORF and uoORF) often influence the translation of the main coding sequence, based on the evidence of the ones that have been detected and investigated in detail [21–25]. From a more human-centered future perspective, uORFs are a rather unexplored niche for translational research: with a predicted prevalence in over 50% of human genes and first examples regulating translation of disease-associated genes already emerging [22,26], the field is bound to not only lead to more fundamental, but also application-oriented insights. Keeping this broader context in mind, we here focus on more fundamental principles of uORFs in a model organism context.
We previously provided mass spectrometric evidence for 467 splice variants and 85 noncanonical gene products, including from polycistronic and ncRNA translation, in C. elegans [27]. Of these, 1 newly discovered gene, azyx-1 (alternative N-terminal ORF of zyx-1), was identified as an 166 amino acid-long protein, translated from the 5′ UTR of zyx-1. In this study, we provide evidence for 2 protein isoforms of azyx-1, one initiating upstream and another downstream of zyx-1 AUG, and both overlapping the proline-rich N-terminus of ZYX-1 long isoforms. To understand whether zyxin proteins could also be regulated by upstream/overlapping ORFs, we here explore functional relevance of azyx-1 and its relation to zyxin in the model organism C. elegans.
Results
A novel gene, azyx-1, reveals putative syntenic conservation of overlapping genes on eukaryotic zyxin
The C. elegans genome contains 2,468 predicted ORFs initiating in 5′ UTRs; however, only 8 of those are supported by mass spectrometric evidence [27,28]. One of these is azyx-1, a noncanonical gene overlapping the gene encoding the only C. elegans zyxin, zyx-1 [27]. azyx-1 is translated from a different reading frame than zyx-1, starting from an AUG 184 bp upstream of the zyx-1 initiation site and delivering a polypeptide product with a sequence length of 166 amino acids (AZYX-1a, Fig 1A). The same locus also embeds a putative shorter AZYX-1 isoform (AZYX-1b), initiating within the zyx-1 coding sequence and containing only the 106 C-terminal amino acids of AZYX-1a (Fig 1A). Six different transcripts have been described for the zyx-1 locus (F24G4.3, WormBase version WS283): 2 are associated with isoform a, and then a single transcript for isoforms b-e each (Fig 1A). Based on these, AZYX-1a can be translated from the F42G4.3a.1 transcript that contains a longer 5′ UTR, whereas AZYX-1b could be translated from all 3 transcripts of the longer zyxin isoforms a and e: F42G4.3a.1, F42G4.3a.2, and F42G4.3e.1. We previously mapped unique peptides to azyx-1, covering >60% of the 166 amino acids-long protein sequence at 1% FDR [27], showing that azyx-1 indeed is translated. Since LIM-domain proteins, to which zyxins belong, often result from alternative splicing and display strong domain conservation across eukaryotes [16], we asked whether azyx-1 or the phenomenon of overlapping genes at zyxin loci is also conserved across eukaryotes. To that end, we searched for predicted ORFs initiating in 5′ UTRs (uORF and uoORF) or within the coding sequence (oORF) of zyxin orthologs (lpp, trip6, and zyx) in 9 animals (OpenProt release 1.6, S4 Table). We identified 14 ORFs initiating upstream of zyx-1 orthologs in 7 species including C. elegans (Fig 1B and S4 Table). Moreover, 6 of these 5′ UTR initiating ORFs were conserved in vertebrates as alternative oORFs within the coding region of zyxin orthologs (S4 Table). Together, our observations suggest a syntenic conservation of overlapping genes towards the 5′ end of zyxin orthologs, in combination with the widely prevalent alternative splicing observed in zyxins.
Fig 1. Schematic representation of zyxin loci including overlapping uoORFs.

Exons (pink: canonical, blue: uoORF/oORF), introns (line), and UTRs (purple) indicated. All loci are shown according to the direction of transcription (arrowhead). (A) Six zyx-1 transcripts are recognized in C. elegans, corresponding to 5 isoforms. The coding sequence for the uoORF/oORF (blue) azyx-1 as identified previously by Parmar and colleagues is contained in the long transcripts. (B) Examples of uoORFs (blue) predicted by OpenProt [28] with respect to zyxin orthologs of B. taurus, D. melanogaster, and M. musculus. The gene names (left), transcript ID (right), and OpenProt IDs are provided next to each schematic representation.
azyx-1 and zyx-1a reporters are observed in partially overlapping anatomical locations
To understand which cells or tissues of C. elegans may express azyx-1, we generated an extrachromosomal azyx-1 reporter strain (LSC1959). While the zyx-1a start codon is contained within this sequence, it does not share the azyx-1 reading frame. Hence, the azyx-1::mNeonGreen fusion construct cannot lead to a fluorescent signal should translation initiate at the downstream zyx-1a start codon. We observed a strong fluorescent signal in body wall muscle, vulval muscle, and very faintly in unidentified structures in the head and tail (Fig 2A–2D). This fluorescence pattern was consistent, as observed in L4 and adult worms (Figs 2 and S1A–S1D). Next, we looked for expression of zyx-1 isoforms as reported via extrachromosomal arrays by Luo and colleagues. In line with their report, we also observe the zyx-1 long isoform predominantly in tail neurons, and faintly in body wall muscle, and additionally also faintly in the pharynx, and more brightly in an unidentified structure in the head (Fig 4A, middle panel of control). Interestingly, the short zyx-1b isoform is also strongly expressed in body wall and vulval muscle [13] (Luo and colleagues), where we observe azyx-1 expression (Fig 2A). Thus, beyond alternative splicing in zyxins, our observations suggest that in those locations, proteoform diversification and localization fine-tuning may putatively also occur via polycistronic transcripts, with the observed expression patterns suggesting the possibility of mutually exclusive translation from long zyx transcripts.
Fig 2. azyx-1 is prominently expressed in body wall and vulval muscle and faintly in the head and tail region.
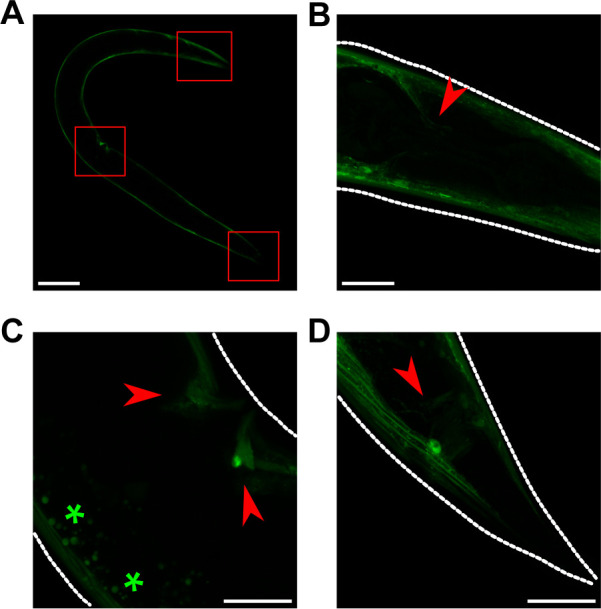
Anterior always to the left. (A–D) For azyx-1 localization, azyx-1p::azyx-1::mNeonGreen:: azyx-1 3′ UTR was expressed extra-chromosomally in a wild-type background. The reporter protein is clearly visible in (A) body wall muscle (scale bar, 100 μm), with red boxes indicating (B) 2 unidentified neurites in the head, (C) vulval muscle, and (D) in the tail region (scale bar, 20 μm, autofluorescence *). For L4 life stage, see S1A–S1C Fig.
Fig 4. azyx-1 overexpression increases zyxin reporter signal and leads to zyxin accumulation in motor neurons.

(A) The integrated reporter control (LSC1870) (upper) contains ZYX-1::GFP (representing all isoforms) and mCherry::ZYX-1 (representing long isoforms), which is compared against a strain with the same genetic reporter background (lower), extrachromosomally overexpressing azyx-1 (azyx-1OE, LSC1960). Anterior to the right; scale bar, 100 μm. (B) GFP (p = 4.85 × 10−8) and (C) mCherry (p = 4.82 × 10−5) quantification showing increase in signal upon azyx-1 overexpression, with * indicating significance (ANOVA) for n ≥ 14 per condition. (D) Ratio of mCherry/GFP. (E) Increased accumulation of GFP along the ventral nerve cord as observed in azyx-1OE (LSC1960) compared to control reporter (LSC1870), *intestinal autofluorescence and (F) colocalization (merged mCherry and GFP, red arrows) of zyx-1::GFP in motor neurons upon azyx-1 overexpression as imaged in adult head, mid-body, and tail. Upper panels: control worms (LSC1998) only expressing the neuronal reporter construct (unc-47p::mCherry) and *co-injection marker (myo-2p::mCherry), lower panels: azyx-1 OE (LSC1999) in the control background. Scale bars: 20 μm. (G) GFP quantification in unc-47p::mCherry-positive cells showing increase in signal upon azyx-1 overexpression (Welch’s t test p = 2.25 × 10−11, n ≥ 35 neurons per condition). Data used to generate figures can be found in S1 Data.
AZYX-1 and ZYX-1 levels vary with age
To understand which protein products are generated by these overlapping genes, we quantified AZYX-1 and ZYX-1 at 3 different ages: L4 larvae, and young (day 1) and post-reproductive (day 8) adults. We selected these life stages based on azyx-1 and zyx-1 expression in body wall muscle (Fig 2), combined with the knowledge that C. elegans musculature deteriorates with age [29]. We observed a relative increase of about 4-fold for AZYX-1 (p = 8.3 × 10−5) and 6-fold for ZYX-1 (p = 1.9 × 10−5) in day 1 adult versus L4 stages (Figs 3A and S2). While AZYX-1 at day 8 of adulthood remained high (p = 0.77, Figs 3A and S2A), ZYX-1 of these post-reproductive animals was intermediary between L4 and day 1 adult levels (Fig 3A). By relying on quantitative data of the specific peptides making up these proteins, we can further deduce that changes in adult zyxin levels differ for different proteoforms: at day 8 versus day 1 of adulthood, 2 out of 3 quantified N-terminal peptides (ZYX-1.1 and 1.3) were comparable to day 1 levels, while the quantified C-terminal peptides (ZYX-1.4 and ZYX-1.5) significantly reduced (p = <0.02, Figs 3B and S2B). For AZYX-1, 5 out of 7 peptides remain unchanged while 2 (shared by both AZYX-1a and AZYX-1b) show a decline (Fig 3C). Together, these data suggest that AZYX-1 levels remain stable while ZYX-1 levels decline between day 1 and day 8 of adulthood. This is corroborated by similar transcript level fold change of zyx-1 for day 1 and day 8 adulthood [30]. Furthermore, it appears that shorter isoforms of zyx-1 may reduce with age, whereas the longer isoforms might be less susceptible to such a post-reproductive decline.
Fig 3. Targeted quantitation in aging worms and in an azyx-1a deletion mutant reveals age-dependent zyx-1 fold change and cis regulation by azyx-1.

Graph axes indicate log2 fold change (left) and absolute fold change (right). Data normalized to a spike-in peptide (1 fmol/worm) for aging wt worms and GPD-3 for mutant vs. wt comparison, with 4 biological replicates (diamonds) for each time point, mean (circle), and standard error bars. (A) Fold change at day 1 (black) and day 8 (gray) of adulthood in comparison to L4 larval stage for all quantified peptides of AZYX-1 (7 peptides) and ZYX-1 (6 peptides) combined. (B) Fold change at day 8 in comparison to day 1 of adulthood for individual ZYX-1 peptides (1.1 to 1.6). zyx-1 gene model indicates the location of peptides unique to long isoform a and e (1.1–1.3) and peptides shared by longer and shorter isoforms (1.4–1.6). (C) Fold change at day 8 in comparison to day 1 of adulthood for individual AZYX-1 peptides (1.1 to 1.7). AZYX-1.1 and 1.2 are unique to AZYX-1a while the rest are shared by AZYX-1a and AZYX-1b. Significantly decreased peptides are colored as per their position relative to the overlapping zyxin gene model (red: long, green: shared; fold change cut-off = <0.66, p-value = <0.02). (D) Peptides unique to ZYX-1 long isoforms (ZYX-1.1–1.3) show a consistent increase in the azyx-1 deletion mutant, while peptides shared between all ZYX-1 (1.4–1.6) isoforms remain similar to wild-type levels (at day 1 of adulthood); p-value: * <0.05, ** <0.01, *** <0.001, ns = not significant, n = 4 biological replicates, with data normalized to GPD-3. For spike-in and HIS-24 normalization, see S3A and S3B Fig. Data used to generate figures can be found in S1 Data.
azyx-1 likely exhibits cis control over zyx-1
Since azyx-1 initiation lies 184 bp upstream of zyx-1, we hypothesized a cis regulation of downstream zyx-1 major ORF by azyx-1. To test whether this could be the case, we generated an azyx-1a deletion mutant (LSC1898, ∆27bp of azyx-1a) and quantified ZYX-1 peptides. We observed a substantial increase in N-terminal ZYX-1 peptides (ZYX-1.1–1.3) for azyx-1 mutants versus wild type (Fig 3D). Peptides that were not specific to the long zyxin proteoforms (ZYX-1.4–1.6) showed a much more modest change—or even none at all—in mutant versus control. The N-terminal peptides, specific to zyx-1 long isoforms (ZYX-1a/e), showed a 2- to 3-fold range increase upon azyx-1a mutation. To confirm that the observations for the deletion mutant most likely result from loss of ribosomal initiation at the AZYX-1a start site, we generated a second mutant through precise nucleotide exchange to mutate it from ATG to TAC (PHX7030). Indeed, both mutants similarly increase ZYX-1long peptides while leaving the others unaffected (S4A Fig). Our observations also hold true for all 3 independent means of data normalization that were available (GDP-3, spike-in, HIS-24; Figs 3D, S3A, S3B, and S3D), supporting the solidity of the claim that removal of the azyx-1a start codon increases zyxin levels in a proteoform-biased way. AZYX-1a mutants also suppressed all downstream peptides shared by AZYX-1b, with several remaining below detection limit in mutants. This suggests that AZYX-1a might be the prominently translated isoform (S3C and S4B Figs). Together, our results fit the hypothesis that azyx-1 may inhibit translation of downstream zyx-1 isoforms, likely affecting the longer isoforms more due to a shared transcript. This does not rule out the possibility of trans regulation (see below) or of an internal ribosome entry site (IRES) downstream of the AZYX-1a start codon for ZYX-1a initiation; however, based on in silico analysis, there is no evidence of IRES within the region downstream of the AZYX-1a start site.
azyx-1 overexpression increases ZYX-1 levels and accumulation in GABAergic motor neurons
While we suspect that uoORF azyx-1a may exhibit cis control over translation of zyx-1 long isoforms, measurable quantities of AZYX-1 peptides are being produced in vivo (Figs 2 and 3, Parmar et al. 2021 [27]). Given the detection of two isoform-specific peptides (S2 Table), these certainly are translation products of azyx-1a. A putative shorter isoform, azyx-1b, with a predicted start codon downstream of that of the zyx-1 long isoforms could also contribute to the remainder of the measured signal (Figs 1A, 2, and 3; Parmar and colleagues [27]). Therefore, we asked whether azyx-1 might also influence C. elegans zyxin in trans. We made use of a reporter system that fuses all zyxin isoforms to GFP and only the long ones (a/e) additionally to mCherry [13] and extrachromosomally expressed azyx-1 (Fig 4A). We measured a significant increase in all (zyx-1::GFP), as well as in the long (mCherry::zyx-1) proteoform reporter levels specifically, upon azyx-1 overexpression in the reporter strain (p < 0.001; Fig 4B and 4C). This observation suggests that production of ZYX-1 may be stimulated by the presence of AZYX-1. In addition, the ratios for mCherry/GFP show no significant differences, suggesting an overall ZYX-1 increase under forced overexpression of azyx-1, irrespective of proteoform (Fig 4D). So far, our results support the presence of both cis and trans control of zyx-1 by azyx-1 in vivo.
Upon azyx-1 overexpression, we also observed an increase in GFP signal aggregation along the ventral nerve cord in what appeared to be motor neurons (Fig 4E). Colocalization with unc-47p::mCherry-positive cells identifies GABAergic neurons along the ventral nerve cord as sites of this zyx-1 accumulation under azyx-1 overexpression with an observed increase in GFP signal (Fig 4F and 4G). zyx-1 has previously been reported to be expressed in GABAergic neurons [31], and its accumulation in these motor neurons upon increased AZYX-1 levels made us hypothesize that azyx-1 overexpressors may suffer locomotion-related impairments.
azyx-1 overexpression affects muscle integrity and neuromuscular behavior in C. elegans
Our expression analysis revealed that azyx-1 and zyx-1 are abundantly expressed in body wall muscle (Figs 2A and S1). In line with this, using blinded and randomized manual scoring (Fig 5A), we observed a quantifiable loss of muscle fiber integrity in azyx-1 overexpression conditions but not in deletion mutants, as compared to control animals (Fig 5B; Fisher’s exact test: wild type versus OE line 1 p = 4.13 × 10−4, versus OE line 2 p = 0.0287, versus mutant p = 1). To test whether manipulating AZYX-1 levels could, as is already known for zyxin deletion [12], affect muscle performance, we performed burrowing assays. These evaluate the ability of worms crawling through Pluronic F-127, a transparent and biocompatible gel, stimulated by a chemoattractant, giving insight into the neuromuscular capabilities of the animals [32]. Here, the genetic overexpression of azyx-1 resulted in defective burrowing as compared to wild type (Fig 5C, two-way ANOVA, Tukey HSD wild type versus azyx-1 OE line 1 p = 3.14 × 10−6, versus azyx-1 OE line 2 p = 4.35 × 10−10; S5 Fig, versus azyx-1 OE line 3 p = 0.046), while azyx-1 mutants displayed contrasting results. The ∆27bp mutant displayed borderline-significant defective burrowing (S5 Fig, p = 0.047) that could be rescued by extrachromosomal resupplementation of azyx-1 (S5 Fig, p = 0.401), but mutating the azyx-1a start site did not affect burrowing capabilities (Fig 5C, p = 1). Together, these data suggest that abnormally high doses of AZYX-1 affect muscular integrity and neuromuscular behavior.
Fig 5. azyx-1 overexpression affects muscle integrity and burrowing behavior.

A fluorescent myo-3 reporter (RW1596) was either crossed with an azyx-1 deletion mutant (LSC2001) or injected to create azyx-1 overexpressor strains (LSC2000, LSC2055) to score muscular quality of day 1 adults via blinded image analysis (n ≥ 40 per condition). (A) Images were scored as disorganized when differing from controls at least as severely as the example image shown here, deteriorated region indicated with arrow. (B) Distribution of observed muscular phenotypes in azyx-1 mutant and overexpressor (OE) lines as compared to controls; Fisher’s exact test p = 1.483 × 10−5, pairwise wild type vs. OE line 1 p = 4.13 × 10−4, vs. OE line 2 p = 0.0287. (C) Chemotaxis index of burrowing assay in day 1 adults, as cumulatively observed over 180 min for the azyx-1 ATG-to-TAC mutant (PHX7030) and for overexpressor lines (line 1: LSC2052, line 2: LSC2053) in comparison to positive (zyx-1(gk190)) and negative (wild type) controls, n ≥ 30 per condition in 4 biologically independent replicates (≥30 × 4); p-value two-way ANOVA 2 × 10−16 for strain and time. Tukey HSD wild type vs. azyx-1 OE line 1 p = 3.14 × 10−6, vs. azyx-1 OE line 2 p = 4.35 × 10−10, vs. zyx-1 p = 4.51 × 10−8. Data used to generate figures can be found in S1 Data.
Discussion
Although noncanonical ORFs are widely prevalent in eukaryotic genomes [33–35], they often tend to be species specific [18,36,37]. These ghost genes often are not susceptible to BLAST analysis due to their small size and inherent genomic variation over species. Upon manually examining the genomic loci of zyxin orthologs, we found evidence of putative syntenic conservation of azyx-1 across 7 species (Fig 2B and S4 Table). This exemplifies how noncanonical ORFs can escape functional annotation, as the existing automated means for orthology mapping are restricted to (large) sequence similarity. Naturally, overlapping genes are more likely to be conserved across species, as a consequence of parent gene conservation. It is interesting however, that overlapping genes could also be harbored within specific gene isoforms to potentially regulate gene expression at a proteoform level, which we propose is the case for azyx-1a and zyx-1a.
Since azyx-1a and zyx-1a likely share a transcript, we asked whether there might be a preference for translation initiation from either of the 2 start codons. Upon azyx-1a start codon mutation, we observed a substantial increase in ZYX-1 long isoform abundance, while the downstream short isoforms largely remain unaffected (Figs 3D, S3A and S3B). Given the global linearity of eukaryotic ribosomal scanning and translation [38,39], our results suggest that azyx-1a exhibits cis control over zyx-1 long isoform(s), by occupying the scanning ribosome in +2 reading frame. While we cannot fully exclude a direct effect on zyx-1a transcription due to a possibly nested effect of any azyx-1 mutation with changes to a putatively unknown regulatory element of the zyxin gene, our observations in 2 independent mutants are in coherence with the widespread cis repression of downstream major ORF translation by upstream ORFs, predicted across vertebrates and observed in cells and animal models [40–42].
Interestingly, azyx-1 also contains a downstream oORF isoform, prompting a putative trans function. One option would be for AZYX-1 to drag the remaining transcript to nonsense-mediated decay (NMD), as has been observed for other uoORFs in plants, yeast, and mammalian cells [43–47]. However, zyx-1 is not present in the available C. elegans resource for NMD targets [48] and contrary to such expectations, we observe a positive trans effect of azyx-1 on zyx-1 reporter constructs (Fig 4). We therefore propose there may be a feed-forward loop that regulates zyx-1 (long and short) translation via azyx-1 (Fig 6). This is in any case a rare observation of reinforcing trans control by an upstream (overlapping) gene, which based on our observations is opposed to its proposed uoORF-mediated cis effect (Fig 3D). A similar observation of a combined cis and trans regulation has been made before for human hepatitis B virus [49], and it is possible that the regulatory context could be proteoform specific. While the cis regulation can be explained by standard ribosomal scanning, the molecular interplay involved in sensing AZYX-1 to then regulate zyx-1 in trans remains to be investigated. Since azyx-1 extends 2,475 bp into the zyx-1 long ORF, it is unlikely for ribosomes to reinitiate at zyx-1 long start codons after being released from the azyx-1 termination site. We hypothesize that the zyx-1-overlapping C-terminus of azyx-1 contains a functional trans domain responsible for the feed-forward loop. Previous studies have shown physical interaction between overlapping genes [50,51], but the mechanisms by which this occurs often remain undiscovered. Our results suggest that azyx-1/zyx-1 is an interesting candidate to elucidate such interactions and intragenic trans regulation.
Fig 6. Proposed model of cis and trans regulation of zyx-1 by azyx-1.

Ribosomes upon entry onto zyx-1 (long) mRNA initiate translation (green) at uoORF AZYX-1a AUG in +2 reading frame, thus repressing translation of downstream ZYX-1 in cis. Leaky scanning through upstream AZYX-1a initiation leads to ZYX-1 translation (pink). Accumulation of translated AZYX-1 causes increased translation of zyx-1 (long and short) mRNA in a feed-forward loop via an unknown signaling cascade.
Phenotypically, increased localization of zyx-1 in motor neurons (Fig 4F and 4G), defective muscle spindles, and burrowing chemotaxis upon azyx-1 overexpression (Fig 5) suggest azyx-1 as a key player in zyx-1-mediated muscular integrity and locomotion. Since zyx-1 shorter isoforms (b and d) are mainly expressed in body wall muscle and motor neurons [12,13], we anticipate that the observed increase upon azyx-1 overexpression corresponds to shorter and not zyx-1 long isoforms (Fig 5B). Because long zyxin transcripts (which also contain azyx-1) can be detected in different anatomical sites (Figs 2 and S1A–S1C; Lecroisey and colleagues and Luo and colleagues [12,13]), it will be interesting to, in future experiments, resolve how all observed effects may depend on cell- or tissue-dictated differences in (a)zyx-1 transcription and translation.
Previous studies observed substantial decrease in worm muscle degeneration upon RNAi knockdown of 5′ region of zyx-1a and 3′ of zyx-1 in dystrophic background (dyc-1; hlh-1), with a distinct isoform involvement [12]. Our results show that azyx-1 overexpression leads to zyx-1 increase and muscle deterioration, while a azyx-1 mutants have significantly increased zyx-1 long isoforms and do not suffer measurable muscular defects (Fig 5B). We hypothesize that the observed effect stems from 2 distinct pathways, which may or may not involve direct modulation by azyx-1. The zyx-1 long isoform, with its characteristic N-terminal proline rich region, has been hypothesized to translocate to the nucleus under cytoplasmic stress and activate transcription of genes involved in repair of damage [12]. This would explain the muscular integrity of azyx-1 mutants, possibly driven by their zyx-1 long isoform increase. On the other hand, azyx-1 overexpression could be causing an imbalance in long/short zyx-1 ratio via excess LIM domain expression and consequent disturbance of endogenous zyx-1 homeostasis, similar to that observed previously upon LIM domain overexpression in Vero cells [52]. Conversely, azyx-1 could harbor a hitherto unknown functional domain, which upon overexpression causes loss of muscular integrity. Whether AZYX-1 trans effects involve other players than ZYX-1, and how these all causally fit together in regulating neuromuscular physiology and function, remains to be detailed. One observation that we cannot yet explain, is the difference in burrowing readout for the 2 azyx-1 deletion alleles, despite their identical results for quantification of AZYX-1 loss and ZYX-1 long increase (S4 Fig). This suggests not everything about this locus is yet known. Given the conservation of LIM domain proteins across eukaryotes, and the evidence of putative syntenic conservation for azyx-1 (Fig 1), we believe this locus provides an interesting model context for future research on LIM domain proteins and prevalence of similar overlapping genes in eukaryotic systems.
Materials and methods
Reagents and tools table in Table 1
Table 1. Reagents and tools table.
| Reagent/resource | Reference or source | Identifier or catalog number |
|---|---|---|
| Experimental models | ||
| N2 Bristol | Caenorhabditis Genetics Center, University of Minnesota, USA | N/A |
| VC299 | Caenorhabditis Genetics Center, University of Minnesota, USA | N/A |
| NM4325 | Gift of Professor Michael Nonet, Washington University Medical School, USA | N/A |
| NM4031 | Gift of Professor Michael Nonet, Washington University Medical School, USA | N/A |
| RW1596 | Gift of Professor Bart Braeckman, Ghent University, Belgium | N/A |
| PHX7030 | SunyBiotech, China | N/A |
| LSC1898 | This study | N/A |
| LSC1950 | N/A | |
| LSC1951 | N/A | |
| LSC1959 | N/A | |
| LSC1870 | N/A | |
| LSC1960 | N/A | |
| LSC1998 | N/A | |
| LSC1999 | N/A | |
| LSC2000 | N/A | |
| LSC2001 | N/A | |
| LSC2052 | N/A | |
| LSC2053 | N/A | |
| LSC2055 | N/A | |
| Recombinant DNA | ||
| azyx-1p::azyx-1::mNeonGreen::UTR | This study | N/A |
| azyx-1p::azyx-1(gBlock)::UTR | This study | N/A |
| unc47p::mCherry | This study | N/A |
| unc133p::mCherry | This study | N/A |
| myo2p::mCherry | This study | N/A |
| Oligonucleotides and sequence-based reagents | This study | S2 Table |
| Chemicals, enzymes and other reagents | ||
| Spike-in peptide EAVSEILETSR | GL Biochem | N/A |
| Modified Trypsin | Promega | V5111 |
| Formic Acid | Pierce | 85178 |
| Acetonitrile | Sigma/Merck | 100029 |
| Dimethyl sulfoxide | Sigma | 276855 |
| Pluronic F-127 | Sigma | P2443 |
| SspI Fast Digest | Thermo Scientific | FD0774 |
| NEBuilder HiFi DNA assembly | New England Biolabs | E2621 |
| Fluorodeoxyuridine | Sigma | F0503 |
| Dithiothreitol | Serva | 20710 |
| Iodoacetamide | Serva | 26710 |
| Urea | Sigma | U5378 |
| Thiourea | Sigma | 88810 |
| HEPES | Sigma | H3375 |
| Software | ||
| MaxQuant (v1.6.17.0) with Andromeda | [53] | N/A |
| Skyline (version 64-bit 21.1.0.278, 22.2.0.527) | [54] | N/A |
| Prosit | [55] | N/A |
| MSStats | [56] | N/A |
| Fiji | https://imagej.net/software/fiji | N/A |
| Blinder | [57] | N/A |
| Rstudio | https://www.rstudio.com/ | N/A |
| Inkscape | https://inkscape.org | N/A |
| Affinity Designer | https://affinity.serif.com | N/A |
| Other | ||
| iRT Kit | Biognosys | N/A |
| Wizard Genomic DNA purification kit | Promega | A1120 |
| COPAS | Union Biometrica | |
| Pierce C18 spin columns | Thermo Scientific | 89870 |
| UltiMate 3000 RSLCnano liquid chromatography system | Thermo Fisher Scientific | N/A |
| Q-Exactive HF-X Hybrid Quadrupole-Orbitrap Mass Spectrometer | Thermo Fisher Scientific | N/A |
| sssReproSil Gold C18-AQ, 3 μm, 450 mm × 75 μm | Dr. Maisch Gmbh | r13.aq.0003 |
| ReproSil-pur C18-AQ, 5 μm, 20 mm × 75 μm | Dr. Maisch Gmbh | r15.aq.0001 |
Worm culture
All strains used in this study (S1 Table) were cultured at 20°C on nematode growth medium (NGM) plates seeded with E. coli OP50 [58,59].
Molecular biology
For azyx-1 localization, 3,474 bp upstream of the azyx-1 stop codon were amplified by PCR from wild-type genomic DNA, along with 558 bp of azyx-1 3′ UTR and fused to 5′ and 3′ ends of mNeonGreen (minus its start-AUG) using HiFi DNA assembly (NEBuilder). The resultant linear transgene was purified (Wizard Genomic DNA purification kit, Promega) and confirmed by sequencing (oligos p001-p006, S3 Table), and injected into wild-type N2 to generate C. elegans strain LSC1959 (see “Transgenesis” and S1 Table). For overexpression and rescue strains (LSC1950, LSC1951, LSC1960, LSC1997, LSC1999, LSC2001, LSC2052, LSC2053, LSC2055; S1 Table), a 757 bp promoter region upstream of azyx-1 was amplified, as were 535 bp of its 3′ UTR. Next, these were fused to 5′ and 3′ ends of a 587 bp synthesized azyx-1 gBlock (Integrated DNA Technologies (IDT); containing all azyx-1 exons and its first intron, with the ATG at the zyx-1a start mutated to CTG) using HiFi DNA assembly (NEBuilder) and confirmed by sequencing (oligos p007-p010, S3 Table). To build the neuronal marker transgene, mCherry was fused to 1,800 bp of the unc-47 promoter region and 497 bp of the unc-47 3′ UTR using HiFi DNA assembly (NEBuilder) and confirmed by sequencing (oligos p011-p016, S3 Table).
CRISPR/Cas9-mediated knockout of azyx-1a
For the generation of the lst1687 allele, which contains a 27 bp deletion at the beginning of the azyx-1 ORF (positions -182 to -155 upstream of the zyx-1a start site), the dpy-10 co-CRISPR strategy was used with homology-directed repair (HDR) according to Paix and colleagues [60]. The injection mix comprised 2.5 μl of recombinant codon-optimized Cas9 enzyme, 2.5 μl tracrRNA (0.17 mol/l, IDT), 1 μl dpy-10 crRNA (0.6 nmol/μl, IDT), 1 μl of azyx-1 crRNA (0.6 nmol/μl, IDT, S3 Table), 1 μl dpy-10 repair template (0.5 mg/ml, Merck), and 1 μl repair template for azyx-1 containing a 27 bp deletion that encompasses azyx-1a start codon (1 mg/ml, IDT, S3 Table). The mix was micro-injected in the gonads of young adult N2 Bristol wild types (Zeiss Axio Observer A1 with Eppendorf Femtojet and Eppendorf Injectman NI2) [61]. Offspring were screened for the desired CRISPR/Cas9-mediated deletion by SspI (FastDigest Thermo Fisher) cleavage of PCR products of the azyx-1 locus, which is only possible after HDR (oligos p007 and p017, S3 Table).
The syb7030 allele was acquired commercially (SunyBiotech) and corresponds to a precise nucleotide exchange of the azyx-1a start codon, wherein the -184th to -182nd basepairs upstream of zyx-1a were mutated from ATG to TAC. The presence of homozygous lst1687 and syb7030 alleles in the azyx-1a mutant strains LSC1898 and PHX7030 (S1 Table) was confirmed via sequencing.
Sample collection and preparation for proteomics
Adult worms were synchronized by standard hypochlorite treatment [62]. After overnight incubation in S-basal (5.85 g NaCl, 1 g K2 HPO4, 6 g KH2PO4 in 1 L milliQ) on a rotor at 20°C, the L1 arrested animals were grown on NGM plates seeded with E. coli OP50. For wild-type sampling at different ages, we collected worms at larval (L4, 48h post L1 refeeding), day 1 adult (20 h post L4 harvest) and post-reproductive, day 8 of adulthood stages. For day 8 samples, offspring were avoided by supplementing the worm cultures with 50 μl of a 50 μm fluorodeoxyuridine (FUDR) solution every 48 h, as of the L4 larval stage (i.e., L4, and days 2, 4, and 6 of adulthood) [63]. For comparisons of wild types with azyx-1 deletion mutants, both strains were synchronized and then grown until larval stage L4 or day 1 adult stage. For sampling, worms were washed off NGM plates with S-basal and allowed to settle in conical tubes for 10 min. Following that, the supernatant was removed and worm pellets were diluted to 15 ml in S-basal for sorting. Worms were sorted using a Complex Object Parametric Analyzer and Sorter (COPAS) platform (Union Biometrica, Holliston, Massachusetts, United States of America) for each sample individually. Four independently grown populations of worms were used per condition. We collected 200 animals per sample for day 8 or 1,000 animals per sample for all other conditions into a 1.5 ml Eppendorf LoBind tube. The worms were pelleted by spinning at 1,500 g for 1 min, S-basal was removed, and 200 μl of 50 mM HEPES were added to the worm pellet, spun at 1,500 g for 1 min and the supernatant was carefully discarded ensuring no worms were lost in pipetting. Finally, the pellet was supplemented with 1 fmol/worm of synthetic spike-in peptide (EAVSEILETSRVSGWRLFKKIS), comprising a proteotypic peptide for quantitation (EAVSEILETSR) (Vandemoortele and colleagues [64]) fused to a HiBit Tag (VSGWRLFKKIS) via a tryptic cleavage site, from a stock solution in water at a concentration of 100 fmol/μl. The pellet was snap frozen in liquid nitrogen and stored at -80°C until further processing. The duration from initial worm collection off NGM until snap freezing was approximately 20 min and carried out at 20°C.
For protein extraction, worm pellets were thawed on ice with 100 μl of lysis buffer (8 M Urea, 2 M Thiourea in 10 mM HEPES) and lysed by sonication using a probe sonicator (40% amplitude, 5 s ON, 10 s OFF × 10). The lysate was spun at 15,000 g for 10 min and the supernatant was transferred to a fresh 1.5 ml Eppendorf LoBind tube. Protein concentration was estimated using a Bradford assay and sample aliquots corresponding to 50 μg of total protein were processed further for LC-MS/MS. For this, each sample was reduced with 5 mM dithiothreitol at 56°C for 30 min and alkylated with 25 mM of iodoacetamide for 20 min at room temperature. The lysate was diluted to 1M urea and digested overnight at 37°C with 2 μg of sequencing-grade trypsin (Promega), after which the sample was acidified to 0.1% formic acid, cleaned using Pierce C18 spin columns as per the manufacturer’s protocol and dried in a Savant SpeedVac. The dried peptides were dissolved to 0.1 μg/μl in 2% acetonitrile/98% H2O/0.1% formic acid (FA)/0.1X Biognosys iRT peptides (for retention time calibration).
Peptide ion selection for targeted quantification
Peptide ions useful for quantification of proteins of interest (ZYX-1 and AZYX-1), and of proteins used for data normalization (GPD-3, HIS-24, spike-in) were selected based on an unscheduled parallel reaction monitoring (PRM) experiment. To accurately normalize data across age and conditions, we chose 3 normalization options: 2 relying on endogenous C. elegans proteins—viz. GPD-3 (GAPDH homolog—4 peptides), HIS-24 (Histone homolog—4 peptides)—and one relying on the externally added synthetic spike-in peptide (1 peptide). Skyline-daily was used to build an initial library [54]. For all proteins of interest, all theoretically predicted tryptic peptides with a length between 7 and 26 amino acids were added to the initial spectral library. In total, 98 peptide precursor ions were selected and measured in an unscheduled PRM experiment that was run on a pooled sample consisting of all peptide samples used in this study and analyzed with Skyline. Subsequently, 23 measured peptide precursors representing 22 peptides and 5 target proteins (ZYX-1, AZYX-1, GPD-3, HIS-24, Spike-in) were selected for the final PRM measurements. Additionally, 11 MS1 ions of the Biognosys iRT reference peptides were included in the precursor list. Details of all peptides and corresponding protein(s), including their uniqueness in the proteome database, can be found in S2 Table.
Targeted LC-MS/MS measurements
Targeted measurements using scheduled PRM were performed with a 50-min linear gradient on a Dionex UltiMate 3000 RSLCnano system coupled to a Q-Exactive HF-X mass spectrometer (Thermo Fisher Scientific). The spectrometer was operated in PRM and positive ionization mode. MS1 spectra (360 to 1,300 m/z) were recorded at a resolution of 60,000 using an AGC target value of 3 × 106 and a MaxIT of 100 ms. Targeted MS2 spectra were acquired at 60,000 resolution with a fixed first mass of 100 m/z, after HCD with a normalized collision energy of 26%, and using an AGC target value of 1 × 106, a MaxIT of 118 ms and an isolation window of 0.9 m/z. In a single PRM measurement, 23 + 11 MS1 peptide ions (see above) were targeted with a 5-min scheduled retention time window. The cycle time was approximately 2.1 s, which leads to about 10 data points per chromatographic peak.
Targeted mass spectrometric data analysis
PRM data were analyzed using Skyline (version 64-bit 21.1.0.278 and 22.2.0.527) [54]. Peak integration, transition interferences, and integration boundaries were reviewed manually, considering 4 to 6 transitions per peptide. To discriminate true from false positive peptide detection, filtering according to correlation of PRM fragment ion intensities was carried out. For this purpose, an experimental spectral library was built from the PRM data itself, by searching these with MaxQuant and then loading the generated search results back into Skyline. For confident peptide identification, a “Library Dot Product” ≥0.85, as well as a mass accuracy ≤10 ppm (“Average Mass Error PPM”) were required. We also manually verified the correlation between PRM fragment ion intensties and spectra predicted with the artifical intellegence algorithm Prosit [55]. For peptide and protein quantification, chromatographic peak areas were exported from Skyline in MSStats format, and further processing, quantification, statistical analysis, and visualization were performed in RStudio with the MSStats package [56]. For HIS-24, 3 most consistent peptides out of 4 were considered for downstream analysis and peptide FISQNYK was omitted. The data were log2 transformed, processed as per default MSStats parameters, and visualized using the ggplot2 package of R. For age analysis, data were normalized to spike-in peptide (1 fmol/worm) and L4 samples were used as the reference. For azyx-1a mutant and wild-type comparison, all 3 normalizations were considered (i.e., GPD-3, HIS-24, and spike-in). To evaluate the internal control stability the pair-wise ratios of each combination were calculated based on Vandesompele and colleagues [65] and the equality of variance was evaluated using Levene’s test. The mass spectrometric raw files acquired in PRM mode and the Skyline analysis files have been deposited to Panorama Public (Sharma and colleagues [66]) and can be accessed via https://panoramaweb.org/azyx1.url.
Transgenesis
For in vivo localization of azyx-1, the lstEx1065 extra-chromosomal array was generated by mixing 25 ng/μl purified linear transgene [azyx-1p::azyx-1+mNeonGreen::azyx-1 3′ UTR] (see “Molecular biology”) with 25 ng/μl coelomocyte-restricted co-injection marker [unc-122p::DsRed] and 50 ng/μl 1-kb ladder (Thermo Scientific) as carrier DNA. This was injected into Bristol wild type (N2) to generate LSC1959 (S1 Table).
All genetic overexpressions and rescues of azyx-1 strains were created via injection of 25 ng/μl of linear transgene [azyx-1p::azyx-1(gBlock)::azyx-1 3′ UTR] with 12.5 ng/μl pharyngeal co-injection marker [myo-2p::mCherry] and 50 ng/μl of 1-kb ladder (Thermo Scientific) as carrier DNA unless otherwise noted (S1 Table). For strains transgenically expressing the unc-47p::mCherry neuronal marker (LSC1998 and LSC1999; S1 Table), 10 ng/μl of this linear construct was injected along with 5 ng/μl pharyngeal co-injection marker [myo-2p::mCherry] and 50 ng/μl of 1-kb ladder (Thermo Scientific), with the addition of 10 ng/μl overexpression transgene [azyx-1p::azyx-1(gBlock):: azyx-1 3′ UTR] for LSC1999.
Transgenic strains were always confirmed by observation of co-injection marker presence via fluoresence microscopy, followed by PCR and sequencing of the added target sequences. For overexpression of azyx-1 in the zyx-1 reporter background (LSC1870, which expresses mCherry::zyx-1;zyx-1::GFP, S1 Table), the extrachromosomal array (NM3425) was integrated with UV irradiation as per [67] and outcrossed twice with wild type (N2 Bristol). All injections using a Zeiss Axio Observer A1 with Eppendorf Femtojet and Eppendorf Injectman NI2 were performed targeting syncytial gonads of young adults.
Confocal imaging
Confocal microscopy was performed using either an Olympus FluoView 1000 (IX81) or a Zeiss LSM900 confocal microscope. To obtain synchronized L4 larvae, a timed egg-laying was performed 48 h before imaging, whereas day 1 adults were synchronized by picking L4 larvae approximately 16 h before imaging. Worms were anaesthetized using of 500 mM sodium azide and mounted on 2% agarose pads. Using Fiji [68], resulting z-stacks were analyzed by performing a sum of slices projection and selecting the region of interest (worm or neuron) with the polygon selection tool. In this region of interest, the mean pixel intensity was measured.
Manual scoring of muscular filaments
Day 1 adults were imaged with confocal microscopy to investigate muscle fiber integrity in control (RW1595, n = 75), overexpressor (LSC2000, n = 52; LSC2055 n = 40) and mutant strains (LSC2001, n = 57). The resulting image files were randomized in Blinder freeware (Cothren and colleagues), visually assessed and scored between 2 qualitative classes of muscle filaments: (1) normal well-organized or (2) mildly damaged or disorganized (Fig 5A), based on previously reported manual scoring parameters [69]. Briefly: muscle fibers displaying tightly organized filaments aligned in a parallel manner were classified as normal. The increased presence of breakage, thinning, or disorganization of individual muscle filaments were classified as disorganized. Fisher’s exact test was used to determine association between strains and muscle integrity. Bonferroni corrections were used for multiple comparisons and adjusted p-values were reported.
Burrowing assay
Burrowing assays were performed with synchronized day 1 adults and executed essentially as described by Lesanpezeshki and colleagues [32], with minor adjustments. Briefly, 20 gravid adults for each replicate were allowed to lay eggs on a seeded NGM plate for 3 h and subsequently removed while allowing the eggs to hatch and grow at 20°C. After 70 h, worms were washed off the NGM plates with S-basal and transferred onto unseeded NGM plates to induce a starvation response. After 1 h, 30 adult worms were dropped in 50 μl 26% w/v Pluronic F-127 in a well of a Corning Costar 12-well plate and covered with 2.5 ml of 26% w/v Pluronic F-127 (Sigma-Aldrich) at 14°C. After 15 min at 20°C, the Pluronic F-127 had gelated and a droplet of 20 μl 100 mg/ml E. coli HB101 was added on top as a chemoattractant (time = 0). The bacterial droplet was monitored every 30 min for 3 h to calculate a chemotaxis index as the percentage of worms that had cumulatively reached the bacteria (out of the 100% corresponding to n ≥ 30). At each time point of observation, worms that had reached the bacterial pellet were removed to avoid crowding and reburrowing. Statistical significance was determined by two-way ANOVA.
Supporting information
Localization of azyx-1p::azyx-1+mNeonGreen::UTR in (A) body wall (scale bar, 100 μm), (B) head, (C) vulval muscle, and (D) tail (scale bar, 20 μm) of L4 stage worms.
(TIF)
Normalized intensity of transition ions for peptides corresponding to (A) AZYX-1 (7 peptides) and (B) ZYX-1 (6 peptides) measured at L4 (orange), day 1 (green) and day 8 (blue) of adulthood (n = 4) with mean intensity across replicates (horizontal black line). Data normalized to spike-in (1 fmol/worm). Formal statistical analysis of data was performed and indicated in main text corresponding to Fig 3A–3D. Data used to generate figures can be found in S1 Data.
(TIF)
Fold change and standard error of individual ZYX-1 peptides in azyx-1 mutant vs. wt with data normalized to (A) spike-in peptide or (B) HIS-24. (C) Fold change and standard error of the only detectable AZYX-1 peptide in azyx-1 mutant strain (LSC1898), likely corresponding to AZYX-1b isoform, upon AZYX-1a start codon deletion with a significant down-regulation in comparison to wt at day 1 of adulthood (normalized to GPD-3, p = 0.0017). (D) Distribution of raw intensity of transition ions with mean and standard error for spike-in (1 peptide), GPD-3 (4 peptides), and HIS-24 (3 peptides) across 4 (colored) biological replicates. None of the normalization methods differed significantly from the others (Levene’s test p = 0.991 for normalized ratios of raw intensities, with pairwise p-values vs. spike-in 0.99, vs. GPD-3 0.72, and vs. HIS-24 0.72). Data used to generate figures can be found in S1 Data.
(TIF)
Fold change and standard error of peptides in azyx-1 mutant strains (Δ27bp and ATG -> TAC) corresponding to (A) ZYX-1, where ZYX-1.1–1.3 peptides are specific to long zyxin isoforms and ZYX-1.5 being the only detectable peptide shared between all isoforms, and (B) the only detectable AZYX-1 peptides likely corresponding to AZYX-1b isoform; p-value: *** <0.001, n = 5 biological replicates, L4 larval stage, with data normalized to GPD-3. Data used to generate figures can be found in S1 Data.
(TIF)
Chemotaxis index of burrowing assay in day 1 adults, as cumulatively observed over 180 min for the azyx-1 (Δ27bp) mutant (LSC1898), rescue (LSC1951), and overexpressor (LSC1950) strains in comparison to positive (zyx-1(gk190)) and negative (wild type) controls, n = 30 per condition in 5 biologically independent replicates (30 × 5); two-way ANOVA p = 0.0025 for strain and time. Tukey HSD wild type vs. azyx-1 OE p = 0.047, vs. azyx-1 (Δ27bp) p = 0.018, vs. zyx-1 p = 0.002. Data used to generate figure can be found in S1 Data.
(TIF)
(XLSX)
elegans proteome.
(XLSX)
(XLSX)
For each, OpenProt ID, species, protein length (# amino acids), molecular weight (kDa), isoelectric point, transcript ID, (localization vs.) associated LIM gene, sequence, and ORF type are provided.
(XLSX)
(XLSX)
Acknowledgments
We would like to thank Prof. Kathrin Gieseler (Université Claude Bernard Lyon 1, France), Prof. Michael Nonet (Washington University in St. Louis, USA) and Prof. Bart Braeckman (UGent, Belgium) for providing strains, and Dr. Marlies Peeters and Dr. Gerben Menschaert (UGent, Belgium) for valuable discussions.
Abbreviations
- ECM
extra-cellular matrix
- FA
formic acid
- HDR
homology-directed repair
- IRES
internal ribosome entry site
- NGM
nematode growth medium
- NMD
nonsense-mediated decay
- ORF
open reading frame
Data Availability
Mass spectrometry data are available via Panorama Public via https://panoramaweb.org/azyx1.url. All other relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by the Fonds Wetenschappelijk Onderzoek Flanders (FWO Flanders, grant G052217N to LT; www.fwo.be), the Katholieke Universiteit Leuven (KU Leuven, grant C16/19/003 to LT; www.kuleuven.be), and by EPIC-XS support to BSP, CL, LT through grant 823839 of the Horizon 2020 programme of the European Union (epic-xs.eu). Some strains were provided by the CGC, which is funded by NIH Office of Research Infrastructure Programs (P40 OD010440). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Beckerle MC. Zyxin: Zinc fingers at sites of cell adhesion. Bioessays. 1997. Nov 1;19(11):949–57. doi: 10.1002/bies.950191104 [DOI] [PubMed] [Google Scholar]
- 2.Zheng Q, Zhao Y. The diverse biofunctions of LIM domain proteins: determined by subcellular localization and protein—protein interaction. Biol Cell. 2007. Sep 1;99(9):489–502. doi: 10.1042/BC20060126 [DOI] [PubMed] [Google Scholar]
- 3.Sadler I, Crawford AW, Michelsen JW, Beckerle MC. Zyxin and cCRP: two interactive LIM domain proteins associated with the cytoskeleton. J Cell Biol. 1992. Dec 15;119(6):1573–87. doi: 10.1083/jcb.119.6.1573 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Freyd G, Kim SK, Horvitz HR. Novel cysteine-rich motif and homeodomain in the product of the Caenorhabditis elegans cell lineage gene lin-11. Nature. 1990. Apr 12;344(6269):876–9. doi: 10.1038/344876a0 [DOI] [PubMed] [Google Scholar]
- 5.Kadrmas JL, Beckerle MC. The LIM domain: from the cytoskeleton to the nucleus. Nat Rev Mol Cell Biol. 2004. Nov 1;5(11):920–31. doi: 10.1038/nrm1499 [DOI] [PubMed] [Google Scholar]
- 6.Fraley SI, Feng Y, Giri A, Longmore GD, Wirtz D. Dimensional and temporal controls of three-dimensional cell migration by zyxin and binding partners. Nat Commun. 2012. Mar 6;3(1):1–13. doi: 10.1038/ncomms1711 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hoffman LM, Jensen CC, Kloeker S, Wang CLA, Yoshigi M, Beckerle MC. Genetic ablation of zyxin causes Mena/VASP mislocalization, increased motility, and deficits in actin remodeling. J Cell Biol. 2006. Feb 27;172(5):771. doi: 10.1083/jcb.200512115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Nix DA, Beckerle MC. Nuclear–Cytoplasmic Shuttling of the Focal Contact Protein, Zyxin: A Potential Mechanism for Communication between Sites of Cell Adhesion and the Nucleus. J Cell Biol. 1997. Sep 8;138(5):1139–47. doi: 10.1083/jcb.138.5.1139 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Winkelman JD, Anderson CA, Suarez C, Kovar DR, Gardel ML. Evolutionarily diverse LIM domain-containing proteins bind stressed actin filaments through a conserved mechanism. Proc Natl Acad Sci U S A. 2020. Oct 13;117(41):25532–42. doi: 10.1073/pnas.2004656117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Wójtowicz A, Babu SS, Li L, Gretz N, Hecker M, Cattaruzza M. Zyxin mediation of stretch-induced gene expression in human endothelial cells. Circ Res. 2010. Oct 1;107(7):898–902. doi: 10.1161/CIRCRESAHA.110.227850 [DOI] [PubMed] [Google Scholar]
- 11.Crone J, Glas C, Schultheiss K, Moehlenbrink J, Krieghoff-Henning E, Hofmann TG. Zyxin Is a Critical Regulator of the Apoptotic HIPK2-p53 Signaling Axis. Cancer Res. 2011. Mar 15;71(6):2350–9. doi: 10.1158/0008-5472.CAN-10-3486 [DOI] [PubMed] [Google Scholar]
- 12.Lecroisey C, Brouilly N, Qadota H, Mariol MC, Rochette NC, Martin E, et al. ZYX-1, the unique zyxin protein of Caenorhabditis elegans, is involved in dystrophin-dependent muscle degeneration. Mol Biol Cell. 2013. Apr 15;24(8):1232–49. doi: 10.1091/mbc.E12-09-0679 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Luo S, Schaefer AM, Dour S, Nonet ML. The conserved LIM domain-containing focal adhesion protein ZYX-1 regulates synapse maintenance in Caenorhabditis elegans. Development. 2014. Oct 15;141(20):3922–33. doi: 10.1242/dev.108217 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Castaneda PG, Wu N, Qiu Z, Lee M, Cram EJ. ZYX-1/Zyxin plays a minor role in oocyte transit through the spermatheca in C. elegans. MicroPubl Biol. 2021. Oct 21;2021: doi: 10.17912/micropub.biology.000489 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Smith P, Leung-Chiu WM, Montgomery R, Orsborn A, Kuznicki K, Gressman-Coberly E, et al. The GLH proteins, Caenorhabditis elegans P granule components, associate with CSN-5 and KGB-1, proteins necessary for fertility, and with ZYX-1, a predicted cytoskeletal protein. Dev Biol. 2002. Nov 15;251(2):333–47. doi: 10.1006/dbio.2002.0832 [DOI] [PubMed] [Google Scholar]
- 16.Zheng QH, Zhang AH, Zhao Y. Structure and function of LIM domain proteins are regulated by alternative splicing. Vol. 9, Int J Clin Exp Med. 2016. Jul 9;13281–13290. [Google Scholar]
- 17.Brunet MA, Leblanc S, Roucou X. Reconsidering proteomic diversity with functional investigation of small ORFs and alternative ORFs. Exp Cell Res. 2020. Aug 1;393(1):112057. doi: 10.1016/j.yexcr.2020.112057 [DOI] [PubMed] [Google Scholar]
- 18.Makalowska I, Lin CF, Makalowski W. Overlapping genes in vertebrate genomes. Comput Biol Chem. 2005. Feb 1;29(1):1–12. doi: 10.1016/j.compbiolchem.2004.12.006 [DOI] [PubMed] [Google Scholar]
- 19.Wright BW, Molloy MP, Jaschke PR. Overlapping genes in natural and engineered genomes. Nat Rev Genet. 2021. 2021. Oct 5;1–15. doi: 10.1038/s41576-021-00417-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Mudge JM, Ruiz-Orera J, Prensner JR, Brunet MA, Calvet F, Jungreis I, et al. Standardized annotation of translated open reading frames. Nat Biotechnol. 2022. Jul 13;40(7):994–9. doi: 10.1038/s41587-022-01369-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Chew GL, Pauli A, Schier AF. Conservation of uORF repressiveness and sequence features in mouse, human and zebrafish. Nat Commun. 2016. May 24;7. doi: 10.1038/ncomms11663 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Schulz J, Mah N, Neuenschwander M, Kischka T, Ratei R, Schlag PM, et al. Loss-of-function uORF mutations in human malignancies. Sci Rep. 2018. Feb 5;8(1):1–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zhang H, Wang Y, Lu J. Function and Evolution of Upstream ORFs in Eukaryotes. Trends Biochem Sci. 2019. Sep 1;44(9):782–94. doi: 10.1016/j.tibs.2019.03.002 [DOI] [PubMed] [Google Scholar]
- 24.Young SK, Wek RC. Upstream open reading frames differentially regulate genespecific translation in the integrated stress response. J Biol Chem. 2016. Aug 12;291(33):16927–35. doi: 10.1074/jbc.R116.733899 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Calvo SE, Pagliarini DJ, Mootha VK. Upstream open reading frames cause widespread reduction of protein expression and are polymorphic among humans. Proc Natl Acad Sci U S A. 2009. May 5;106(18):7507–12. doi: 10.1073/pnas.0810916106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lee DSM, Park J, Kromer A, Baras A, Rader DJ, Ritchie MD, et al. Disrupting upstream translation in mRNAs is associated with human disease. Nat Commun. 2021. Mar 9;12(1):1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Parmar BS, Peeters MKR, Boonen K, Clark EC, Baggerman G, Menschaert G, et al. Identification of Non-Canonical Translation Products in C. elegans Using Tandem Mass Spectrometry. Front Genet. 2021. Oct 25;0:1910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Brunet MA, Ene Brunelle M, Lucier JFO, Delcourt V, Levesque M, Fréd F, et al. OpenProt: a more comprehensive guide to explore eukaryotic coding potential and proteomes. Nucleic Acids Res. 2018. Jan 8;47:403–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Herndon LA, Schmeissner PJ, Dudaronek JM, Brown PA, Listner KM, Sakano Y, et al. Stochastic and genetic factors influence tissue-specific decline in ageing C. elegans. Nature. 2002. Oct 24;419(6909):808–14. doi: 10.1038/nature01135 [DOI] [PubMed] [Google Scholar]
- 30.Roux AE, Yuan H, Podshivalova K, Hendrickson D, Kerr R, Kenyon C, et al. The complete cell atlas of an aging multicellular organism. bioRxiv. 2022. Jun 16;2022.06.15.496201. [Google Scholar]
- 31.Spencer WC, Zeller G, Watson JD, Henz SR, Watkins KL, McWhirter RD, et al. A spatial and temporal map of C. elegans gene expression. Genome Res. 2011. Feb 1;21(2):325–41. doi: 10.1101/gr.114595.110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lesanpezeshki L, Hewitt JE, Laranjeiro R, Antebi A, Driscoll M, Szewczyk NJ, et al. Pluronic gel-based burrowing assay for rapid assessment of neuromuscular health in C. elegans. Sci Rep. 2019. Oct 23;9(1):1–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Chen J, Brunner AD, Cogan JZ, Nuñez JK, Fields AP, Adamson B, et al. Pervasive functional translation of noncanonical human open reading frames. Science (80-). 2020. Mar 6;367(6482):1140 LP–1146. doi: 10.1126/science.aay0262 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ingolia NT, Brar GA, Stern-ginossar N, Harris MS, Talhouarne GJ, Jackson SE, et al. Ribosome Profiling Reveals Pervasive Translation Outside of Annotated Protein-Coding Genes. Cell Rep. 2014. Sep 11;8(5):1365–79. doi: 10.1016/j.celrep.2014.07.045 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Bazzini AA, Johnstone TG, Christiano R, Mackowiak SD, Obermayer B, Fleming ES, et al. Identification of small ORFs in vertebrates using ribosome footprinting and evolutionary conservation. EMBO J. 2014. May 2;33(9):981–93. doi: 10.1002/embj.201488411 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Veeramachaneni V, Makałowski W, Galdzicki M, Sood R, Makałowska I. Mammalian Overlapping Genes: The Comparative Perspective. Genome Res. 2004. Feb;14(2):280. doi: 10.1101/gr.1590904 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Soldà G, Suyama M, Pelucchi P, Boi S, Guffanti A, Rizzi E, et al. Non-random retention of protein-coding overlapping genes in Metazoa. BMC Genomics. 2008. Apr 16;9. doi: 10.1186/1471-2164-9-174 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Vassilenko KS, Alekhina OM, Dmitriev SE, Shatsky IN, Spirin AS. Unidirectional constant rate motion of the ribosomal scanning particle during eukaryotic translation initiation. Nucleic Acids Res. 2011. Jul 1;39(13):5555–67. doi: 10.1093/nar/gkr147 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Hinnebusch AG. Molecular Mechanism of Scanning and Start Codon Selection in Eukaryotes. Microbiol Mol Biol Rev. 2011. Sep 1;75(3):434. doi: 10.1128/MMBR.00008-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Ribone PA, Capella M, Arce AL, Chan RL. A uORF Represses the Transcription Factor AtHB1 in Aerial Tissues to Avoid a Deleterious Phenotype. Plant Physiol. 2017. Nov 1;175(3):1238–53. doi: 10.1104/pp.17.01060 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Vattem KM, Wek RC. Reinitiation involving upstream ORFs regulates ATF4 mRNA translation in mammalian cells. Proc Natl Acad Sci U S A. 2004. Aug 3;101(31):11269–74. doi: 10.1073/pnas.0400541101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Johnstone TG, Bazzini AA, Giraldez AJ. Upstream ORFs are prevalent translational repressors in vertebrates. EMBO J. 2016. Apr 1;35(7):706–23. doi: 10.15252/embj.201592759 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Nyikó T, Sonkoly B, Mérai Z, Benkovics AH, Silhavy D. Plant upstream ORFs can trigger nonsense-mediated mRNA decay in a size-dependent manner. Plant Mol Biol. 2009. Aug 4;71(4–5):367–78. doi: 10.1007/s11103-009-9528-4 [DOI] [PubMed] [Google Scholar]
- 44.Hurt JA, Robertson AD, Burge CB. Global analyses of UPF1 binding and function reveal expanded scope of nonsense-mediated mRNA decay. Genome Res. 2013. Oct 1;23(10):1636–50. doi: 10.1101/gr.157354.113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Mendell JT, Sharifi NA, Meyers JL, Martinez-Murillo F, Dietz HC. Nonsense surveillance regulates expression of diverse classes of mammalian transcripts and mutes genomic noise. Nat Genet. 2004. Sep 26;36(10):1073–8. doi: 10.1038/ng1429 [DOI] [PubMed] [Google Scholar]
- 46.Gaba A, Jacobson A, Sachs MS. Ribosome occupancy of the yeast CPA1 upstream open reading frame termination codon modulates nonsense-mediated mRNA decay. Mol Cell. 2005. Nov 11;20(3):449–60. doi: 10.1016/j.molcel.2005.09.019 [DOI] [PubMed] [Google Scholar]
- 47.Johansson MJO, Jacobson A. Nonsense-mediated mRNA decay maintains translational fidelity by limiting magnesium uptake. Genes Dev. 2010. Jul 15;24(14):1491–5. doi: 10.1101/gad.1930710 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Muir VS, Gasch AP, Anderson P. The substrates of nonsense-mediated mRNA decay in Caenorhabditis elegans. G3. 2018. Jan 1;8(1):195–205. doi: 10.1534/g3.117.300254 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Chen A, Kao YF, Brown CM. Translation of the first upstream ORF in the hepatitis B virus pregenomic RNA modulates translation at the core and polymerase initiation codons. Nucleic Acids Res. 2005. Mar 1;33(4):1169–81. doi: 10.1093/nar/gki251 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Bergeron D, Lapointe C, Bissonnette C, Tremblay G, Motard J, Roucou X. An out-of-frame overlapping reading frame in the ataxin-1 coding sequence encodes a novel ataxin-1 interacting protein. J Biol Chem. 2013. Jul 26;288(30):21824–35. doi: 10.1074/jbc.M113.472654 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Klemke M, Kehlenbach RH, Huttner WB. Two overlapping reading frames in a single exon encode interacting proteins—A novel way of gene usage. EMBO J. 2001. Jul 16;20(14):3849–60. doi: 10.1093/emboj/20.14.3849 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Nix DA, Fradelizi J, Bockholt S, Menichi B, Louvard D, Friederich E, et al. Targeting of Zyxin to Sites of Actin Membrane Interaction and to the Nucleus *. J Biol Chem. 2001. Sep 14;276(37):34759–67. doi: 10.1074/jbc.M102820200 [DOI] [PubMed] [Google Scholar]
- 53.Tyanova S, Temu T, Cox J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nat Protoc. 2016. Oct 27;11(12):2301–19. doi: 10.1038/nprot.2016.136 [DOI] [PubMed] [Google Scholar]
- 54.Pino LK, Searle BC, Bollinger JG, Nunn B, MacLean B, MacCoss MJ. The Skyline ecosystem: Informatics for quantitative mass spectrometry proteomics. Mass Spectrom Rev. 2020. May 1;39(3):229–44. doi: 10.1002/mas.21540 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Gessulat S, Schmidt T, Zolg DP, Samaras P, Schnatbaum K, Zerweck J, et al. Prosit: proteome-wide prediction of peptide tandem mass spectra by deep learning. Nat Methods. 2019. May 27;16(6):509–18. doi: 10.1038/s41592-019-0426-7 [DOI] [PubMed] [Google Scholar]
- 56.Choi M, Chang CY, Clough T, Broudy D, Killeen T, MacLean B, et al. MSstats: an R package for statistical analysis of quantitative mass spectrometry-based proteomic experiments. Bioinformatics. 2014. Sep 1;30(17):2524–6. doi: 10.1093/bioinformatics/btu305 [DOI] [PubMed] [Google Scholar]
- 57.Cothren SD, Meyer JN, Hartman JH. Blinded Visual Scoring of Images Using the Freely-available Software Blinder. Bioanalysis. 2018. Dec 5;8(23). doi: 10.21769/BioProtoc.3103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Lewis JA, Fleming JT. Chapter 1: Basic Culture Methods. Methods Cell Biol. 1995. Jan 1;48(C):3–29. [PubMed] [Google Scholar]
- 59.Brenner S. The genetics of Caenorhabditis elegans. Genetics. 1974. May;77(1):71–94. doi: 10.1093/genetics/77.1.71 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Paix A, Folkmann A, Seydoux G. Precision genome editing using CRISPR-Cas9 and linear repair templates in C. elegans. Methods. 2017. May 15;121–122:86–93. doi: 10.1016/j.ymeth.2017.03.023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Mello CC, Kramer JM, Stinchcomb D, Ambros V. Efficient gene transfer in C.elegans: extrachromosomal maintenance and integration of transforming sequences. EMBO J. 1991. Dec 1;10(12):3959–70. doi: 10.1002/j.1460-2075.1991.tb04966.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Porta-De-La-Riva M, Fontrodona L, Villanueva A, Cerón J. Basic Caenorhabditis elegans Methods: Synchronization and Observation. J Vis Exp. 2012. Jun 10;(64). doi: 10.3791/4019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Mitchell DH, Stiles JW, Santelli J, Rao Sanadi D. Synchronous Growth and Aging of Caenorhabditis elegans in the Presence of Fluorodeoxyuridine. J Gerontol. 1979. Jan 1;34(1):28–36. doi: 10.1093/geronj/34.1.28 [DOI] [PubMed] [Google Scholar]
- 64.Vandemoortele G, Staes A, Gonnelli G, Samyn N, De Sutter D, Vandermarliere E, et al. An extra dimension in protein tagging by quantifying universal proteotypic peptides using targeted proteomics. Sci Rep. 2016. Jun 6;6:27220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002. Apr 10;3(7):research0034.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Sharma V, Eckels J, Schilling B, Ludwig C, Jaffe JD, MacCoss MJ, MacLean B. Panorama Public: A Public Repository for Quantitative Data Sets Processed in Skyline. Mol Cell Proteomics. 2018. Jun;17(6):1239–1244. doi: 10.1074/mcp.RA117.000543 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Mariol MC, Walter L, Bellemin S, Gieseler K. A rapid protocol for integrating extrachromosomal arrays with high transmission rate into the C. elegans genome. J Vis Exp. 2013. Sep 12;(82). doi: 10.3791/50773 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T, et al. Fiji: an open-source platform for biological-image analysis. Nat Methods. 2012. Jun 28;9(7):676–82. doi: 10.1038/nmeth.2019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Dhondt I, Verschuuren C, Zečić A, Loier T, Braeckman BP, de Vos WH. Prediction of biological age by morphological staging of sarcopenia in caenorhabditis elegans. Dis Model Mech. 2021. Nov 1;14(11). doi: 10.1242/dmm.049169 [DOI] [PMC free article] [PubMed] [Google Scholar]