

Abstract
Following a request from the European Commission, the EFSA Panel on Plant Health performed a quantitative pest risk assessment to assess whether the import of cut roses provides a pathway for the introduction of Thaumatotibia leucotreta (Lepidoptera: Tortricidae) into the EU. The assessment was limited to the entry and establishment steps. A pathway model was used to assess how many T. leucotreta individuals would survive and emerge as adults from commercial or household wastes in an EU NUTS2 region climatically suitable in a specific season. This pathway model for entry consisted of three components: a cut roses distribution model, a T. leucotreta developmental model and a waste model. Four scenarios of timing from initial disposal of the cut roses until waste treatment (3, 7, 14 and 28 days) were considered. The estimated median number of adults escaping per year from imported cut roses in all the climatically suitable NUTS2 regions of the EU varied from 49,867 (90% uncertainty between 5,298 and 234,393) up to 143,689 (90% uncertainty between 21,126 and 401,458) for the 3‐ and 28‐day scenarios. Assuming that, on average, a successful mating will happen for every 435 escaping moths, the estimated median number of T. leucotreta mated females per year from imported cut roses in all the climatically suitable NUTS2 regions of the EU would vary from 115 (90% uncertainty between 12 and 538) up to 330 (90% uncertainty between 49 and 923) for the 3‐ and 28‐day scenarios. Due to the extreme polyphagia of T. leucotreta, host availability will not be a limiting factor for establishment. Climatic suitability assessment, using a physiologically based demographic modelling approach, identified the coastline extending from the northwest of the Iberian Peninsula through the Mediterranean as area suitable for establishment of T. leucotreta. This assessment indicates that cut roses provide a pathway for the introduction of T. leucotreta into the EU.
Keywords: Africa, Israel, false codling moth, climate suitability, pathway model, quantitative assessment, waste management
Summary
Following a request from the European Commission on whether the importation of cut flowers of roses (Rosa sp.) into the EU could constitute a potential pathway for the introduction of Thaumatotibia leucotreta (Meyrick) (Lepidoptera: Tortricidae), the EFSA Panel on Plant Health performed a quantitative pest risk assessment limited to the entry and the establishment of T. leucotreta, the false codling moth. The assessment focused on the pathway of import of cut roses from the areas where T. leucotreta is known to occur and the likelihood of introduction (i.e. entry, including transfer, and establishment) in the EU.
This polyphagous insect pest (affecting more than 100 genera of host plants in more than 50 botanical families) occurs in sub‐Saharan Africa and has spread to Israel. The pest is regularly intercepted on cut roses and other fresh produce imported into the EU from its areas of occurrence.
The area potentially suitable for the establishment of this pest in the EU was assessed using a climate matching approach according to Köppen–Geiger categories and a physiologically based demographic model (PBDM). The predictions of the PBDM were validated with the occurrence data from the areas of origin of the pest and from invaded areas.
T. leucotreta larvae on roses imported into the EU will be primarily affected by cold stress exacerbated by a lack of dormancy/diapause. The area of potential establishment includes the coastline extending from the northwest of the Iberian Peninsula through the Mediterranean. The estimated densities of populations in the EU do not reach the high population densities projected for East Africa. Other published models are in broad agreement with these predictions regarding the potential major areas at risk of establishment (which is related to the common use of data on temperature requirements of this pest).
Additional areas in the EU indicated as suitable with low population numbers are most likely associated with transient populations only. The main uncertainties about possible establishment in these areas are caused by the lack of developmental data at different temperatures. Besides outdoors establishment in regions climatically suitable, as stated by EPPO (2013), T. leucotreta could overwinter in greenhouses in other areas with horticultural production.
Considering entry, the pest has been frequently intercepted on cut roses and observational records exist of flying adults of the pest in a few locations in the EU. An entry pathway model was used to assess the probability of entry of T. leucotreta, considering different steps of the pathway, and to identify the uncertainties of the assessment. The model consisted of three components to determine the number of adults which would escape from cut roses imported from countries with reported occurrence of T. leucotreta (African countries and Israel):
a cut roses distribution model which describes the proportion of the imported infested roses distributed to the NUTS2 regions in the EU with suitable climate;
a developmental model that describes the proportion of T. leucotreta adults that would emerge from infested cut roses depending on the number of days after import into the EU;
a waste model which describes the proportion of T. leucotreta adults that would survive and escape prior to different types of waste treatments.
The model estimates how many T. leucotreta individuals would survive and emerge as adults from commercial or household disposal of infested cut roses, in the EU NUTS2 regions where the establishment is estimated possible based on physiologically based demographic modelling. The number of T. leucotreta adults escaping from disposed cut roses per year and per season are calculated using trade and temperature data.
Four scenarios were considered, for the timespan from the initial disposal of the cut roses at the household until the waste treatment: 3, 7, 14 and 28 days.
According to model results, the median number of T. leucotreta adults escaping from imported cut roses in all the climatically suitable NUTS2 regions of the EU was estimated as 49,867 per year (90% uncertainty between 5,298 and 234,393) for the 3‐day scenario of time from initial disposal of cut roses at the household until waste treatment, and as 143,689 per year (90% uncertainty between 21,126 and 401,458) for the 28‐day scenario. The differences across the scenarios are due to the escapes from the regional waste management processes, whereas the escapes from the private compost remain constant across all scenarios.
Assuming as a realistic scenario that on average one of every 435 escaping T. leucotreta moths results in a successful mating, the estimated median number of T. leucotreta mated females per year from imported cut roses in all the climatically suitable NUTS2 regions of the EU would vary from 115 (90% uncertainty between 12 and 539) up to 330 (90% uncertainty between 49 and 923) for the 3‐ and 28‐day scenarios, respectively. When analysing the results for the EU by seasons, the highest number of expected mated females is predicted in summer compared to the other seasons in the 14‐ and 28‐day scenarios. In particular, for the 28‐day scenario, the number of mated females in summer would be 185 (90% uncertainty between 28 and 480), contributing more than 50% of the total annual mated females. Factors like clustering of infestations in cut roses or spatial or temporal clustering of cut roses consumption in a particular residential area or during favourable times of the year would increase the probability of mating and transfer to suitable host.
With regard to host plants availability, the Panel agreed with EPPO (2013) on the wide availability of suitable hosts in the coastal areas of Southern Europe. A female of T. leucotreta, having an extremely wide range of plants suitable for oviposition and further larval development, will likely find suitable hosts for oviposition even during the winter in the areas climatically suitable. Due to the extreme polyphagia of T. leucotreta immature stages, host availability should not be a limiting factor for establishment in climatically suitable areas.
Overall, regular escape of pest insects on the territory of the EU is predicted but so far it has not led to outbreaks (other than few incursions) in the EU, possibly because of the relatively recent shift of pest pressure in Africa towards cut roses and the fact that much of the consumption of cut roses in the EU occurs in regions with lower climate suitability. However, observations of flying adults have been reported in the EU.
The outputs of this quantitative pest risk assessment indicate that cut roses provide a pathway for the introduction of T. leucotreta to the EU
The number of escaped adults of T. leucotreta with a possible mating partner, from the imported cut roses in the realistic clustering scenario, is predicted to be higher during summer than in the other seasons, particularly when the 14‐ and 28‐day scenarios until waste treatment are considered. This seasonality is explained by the faster development of T. leucotreta in the warmer season.
Sensitivity analysis of the pathway model showed that the main uncertainties remain regarding: the infestation rate in the imported cut roses; and main parameters of the waste model, especially the proportion of waste privately composted and the timing between initial disposal of the cut roses in the household and the waste treatment in the public facilities.
To reduce the uncertainties, data collection and research are recommended on the following key topics: the ecology and biology of T. leucotreta in its natural environment and in cut rose production in Eastern Africa; the level of infestation and clustering of T. leucotreta in the cut roses consignments; the level of effectiveness of the export and import border inspections in detecting the different life stages of T. leucotreta in cut roses; the actual waste management processes at NUTS2 level in the EU, including the proportion of private composting and the timing between the initial waste disposal and the waste treatment.
1. Introduction
1.1. Background and terms of reference as provided by the requestor
1.1.1. Background
False codling moth (Thaumatotibia leucotreta [Meyrick]) is a moth species belonging to the family Tortricidae under the order of Lepidoptera. Larvae of the moth feed on a wide range of fruit, vegetable and other crops. It is not known to occur in the EU and it is regulated as a Union Quarantine Pest i.e., it is included in the annexes of Commission Implementing Regulation (EU) 2019/2072. It is also listed as a priority pest under Regulation (EU) 2019/1702. The pest is polyphagous and has a strong dispersal potential. The eggs and larvae of the pest are regularly intercepted on cut flowers of roses (Rosa sp.) imported into the EU from non‐EU countries.
Cut flowers of roses (Rosa sp.) are included in Annex XI.A of Regulation (EU) 2019/2072 as a commodity that is subject to phytosanitary certificates. They are, however, not included among commodities for which special import requirements for T. leucotreta are required under Annex VII. of the aforesaid Regulation.
The reason for which EFSA is requested to prepare its scientific opinion is related to the inclusion of cut roses 1 in the system of reduced frequencies of physical checks during import plant health inspection (Regulation (EC) No 1756/2004). The inclusion in the system of cut roses imported from African countries (Kenya in particular), is put in question due to the number of interceptions of the pest.
Over the last few years, the number of interceptions of eggs and larvae of T. leucotreta in imported cut flowers of roses, especially from Kenya, has increased. It would be therefore helpful, if the risk of cut roses as a potential pathway for the introduction of the pest in the Union was clarified. This would complement the non‐conclusive guidance on this potential pathway provided by the pest risk analysis carried out by EPPO in 2013 (modified in 2021), 2 and would guide the Commission and the Member States on further regulatory approach on the pest in the commodity.
1.1.2. Terms of reference (ToR)
EFSA is requested, pursuant to Article 29(1) of Regulation (EC) No 178/2002, to provide a scientific opinion in the field of plant health.
EFSA is requested to deliver an opinion whether the importation of cut flowers of roses (Rosa sp.) into the EU constitutes a potential pathway for the introduction of Thaumatotibia leucotreta (Meyrick). In order to reach that conclusion, EFSA shall take into account all relevant scientific and technical information, including data collected by Member States on interceptions of the pest in cut roses.
1.2. Interpretation of the terms of reference
The terms of reference specify that the requested opinion should address the probability of introduction, which is defined by the International Standard on Phytosanitary Measures ISPM No 5 (IPPC Secretariat, 2022) as the entry of a pest resulting in its establishment, thus including entry (including transfer) and establishment.
The assessment of spread and impact is therefore outside the scope of this mandate and not included in this opinion. The Panel therefore undertook a partial quantitative pest risk assessment, according to the principles laid down in its guidance on quantitative pest risk assessment (EFSA PLH Panel, 2018), as it is limited to the steps of entry (including transfer) and establishment.
As the mandate focuses on the assessment of the probability of introduction with cut roses under current conditions, no additional scenario of risk reduction options is included.
This opinion only deals with the introduction of T. leucotreta (the false codling moth (FCM)) via the pathway of imported cut roses from countries where the pest occurs, i.e. African countries and Israel (as specified in the mandate background).
2. Data and methodologies
For this opinion, the following data were searched:
Data on the EU import of cut roses from Africa and Israel;
Data on the import volume and destinations of cut roses into the EU;
Data on the waste treatment procedures in the EU;
Data on T. leucotreta developmental biology in relation to temperature.
The assessment was based on a combination of literature review, interviews with hearing experts and Expert Knowledge Elicitation (EKE) with experts or Panel members and EFSA staff to assess quantities that could not be well identified from literature or databases alone (EFSA, 2014). To link pest entry with establishment potential, the distribution of infested plant material entering the EU was assessed using the NUTS 2 statistical regions of the EU as the spatial resolution.
Hearings with experts from the National Plant Protection Organisations (NPPO) of Kenya and Uganda took place on 10 November 2022. Experts presented the pest status of T. leucotreta on cut roses in Kenya and Uganda and the phytosanitary measures that are in place against T. leucotreta including their limitations. The NPPOs of Kenya and Ethiopia submitted a report about T. leucotreta pest status on cut roses in their countries and the respective phytosanitary measures along with their limitations. Upon request of the EFSA, the NPPO of Kenya (KEPHIS) sent additional information, on anonymised trapping data of T. leucotreta populations in and around six rose farms (KEPHIS, personal communication, 11 January 2023).
Information on the pest distribution was retrieved from the European and Mediterranean Plant Protection Organization (EPPO) Global Database (EPPO, online) and relevant literature.
Data on interceptions and outbreaks of the pest within the risk assessment area were searched in the EUROPHYT and TRACES databases.
Data on cut roses import sampling and inspection procedures, infestation rates and the development stage of intercepted T. leucotreta specimens were provided by the Netherlands Food and Consumer Product Safety Authority (NVWA) (NVWA, 2022. Development of T. leucotreta on cut roses, personal communication, 23 September 2022; NVWA, 2022. Development stage of intercepted T. leucotreta specimens, personal communication, 12 October 2022; NVWA, 2022. Infestation rates of consignments of cut‐roses infested by T. leucotreta, personal communication, 12 October 2022).
Data on interception and development stage of intercepted T. leucotreta specimens in cut roses import were provided by the Belgian Federal Agency for the Safety of the Food Chain (FAVV‐AFSCA) (FAVV‐AFSCA, 2022. Reply to EFSA request of info on the FCM cut roses interceptions and checks in Belgium, personal communication, 25 January 2023).
2.1. Temporal and spatial scales
The cut roses pathway model calculates the flow per year, on average, over a period of 5 years (2022–2026).
The Köppen–Geiger climate classification used 30 years of climate data, 1981–2010. The Köppen–Geiger climate classification uses a 0.08 × 0.08° world grid.
The physiologically based demographic model (PBDM) (Gutierrez, 1996) uses 2000–2010 gridded temperature data with the scale of about 0.25° × 0.25°. The projected distribution and relative abundance map of T. leucotreta used the same resolution (Ruane et al., 2015; AgMERRA data, https://data.giss.nasa.gov/impacts/agmipcf/agmerra).
2.2. Data on T. leucotreta occurrence and interceptions
2.2.1. Occurrences of T. leucotreta
An extensive literature search for T. leucotreta global distribution was conducted in Web of Science (all databases, excluding Data Citation Index and Zoological Record) and Scopus on 12 May 2022 (Rossi et al., 2023; see Appendix D). The search string was based only on the scientific and English common name of the pest. No other keywords were used in order not to limit the retrieval of distribution data, often reported as secondary information. The review followed a two‐step approach. The first step was based on the title and the abstract, while the second one was based on the full text. The search yielded 240 documents including information on pest distribution. From these documents, 751 records of the presence of T. leucotreta were extracted, out of which 516 were specific locations reporting geographic coordinates, or very small administrative units (e.g. small provinces) for which coordinates from Google Earth were used, and 235 were related to larger administrative units (Figure 1). The full description of the literature search methodology and the results is available in Rossi et al. (2023) (see Appendix D).
Figure 1.
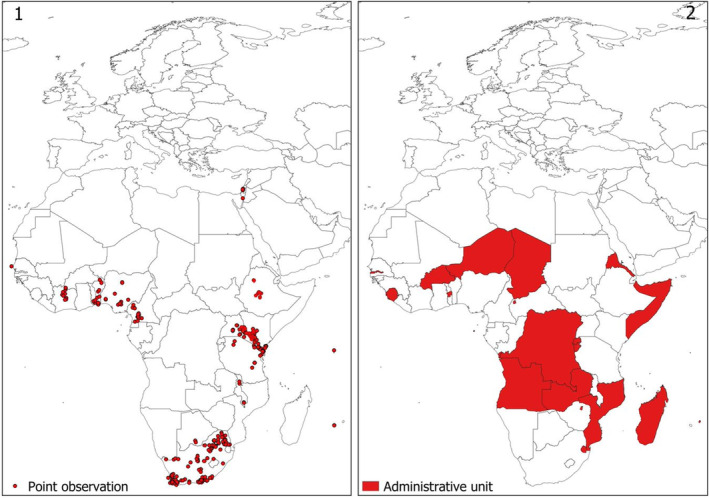
Observed distribution of T. leucotreta. The map on the left (1) shows point observations. The map on the right (2) shows observations at the administrative unit level, for the areas where point observations were not found (Rossi et al., 2023; Appendix D)
2.2.2. Interceptions of T. leucotreta in border inspections
The Panel searched for interceptions of T. leucotreta on any commodity from 1994 until 2022, in EUROPHYT (last accessed on 10 February 2023) and TRACES (accessed on 10 February 2023).
From 2014 to 2022, a total of 517 interceptions of T. leucotreta were found in the EU (Table 1). Most of the infested consignments were intercepted in the Netherlands, Belgium, France, Germany and Spain. In the same period, 261 interceptions were found in cut roses, with the majority of interceptions on cut roses in the Netherlands and Belgium and few interceptions from France and Germany (Table 2). No interceptions in cut roses were reported in any other EU Member State (MS) for the period 2014–2022; however, this can be explained by their limited trade in cut roses (see Figure 4).
Table 1.
Annual interceptions of T. leucotreta on all commodities per EU MS, from 2014 until November 2022
| Total interceptions | Year | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Country | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | Total |
| Belgium | 11 | 6 | 9 | 13 | 20 | 7 | 2 | 2 | 70 | |
| Bulgaria | 2 | 2 | ||||||||
| Cyprus | 1 | 1 | ||||||||
| France | 7 | 6 | 7 | 7 | 4 | 3 | 1 | 4 | 39 | |
| Germany | 3 | 5 | 1 | 2 | 2 | 1 | 14 | |||
| Ireland | 1 | 1 | ||||||||
| Italy | 2 | 2 | ||||||||
| Lithuania | 2 | 2 | ||||||||
| The Netherlands | 23 | 16 | 21 | 13 | 48 | 43 | 48 | 79 | 69 | 360 |
| Portugal | 1 | 1 | 2 | 4 | ||||||
| Spain | 6 | 1 | 2 | 3 | 12 | |||||
| Sweden | 1 | 3 | 2 | 1 | 2 | 1 | 10 | |||
| Total | 40 | 41 | 41 | 37 | 68 | 77 | 57 | 81 | 76 | 517 |
Table 2.
Annual interceptions of T. leucotreta on cut roses per EU MS, from 2014 until November 2022
| Rosa sp. | Year | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Import country | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | Total |
| Belgium | 2 | 7 | 6 | 2 | 17 | |||||
| France | 3 | 3 | ||||||||
| Germany | 2 | 1 | 3 | |||||||
| The Netherlands | 1 | 29 | 25 | 43 | 79 | 61 | 238 | |||
| Total | 1 | 0 | 0 | 0 | 31 | 34 | 50 | 81 | 61 | 261 |
Figure 4.
Distribution of cut roses imported by the EU (2011–2020)
From 2014 until January 2023, T. leucotreta was intercepted in 148 shipments of cut roses originating from Kenya, 84 shipments of cut roses from Uganda, 46 from Tanzania, 36 from Ethiopia, 20 from Zambia, 14 from Zimbabwe and 3 from Rwanda (Table 3). A peak in interceptions both in cut roses as well as in other consignments was recorded in 2021 (Table 4).
Table 3.
Annual interceptions of T. leucotreta on cut roses per exporting Country, from 2014 until January 2023
| Rosa sp. | Year | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Exporting country | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | 2023 (January) | Total |
| Ethiopia | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 10 | 23 | 1 | 36 |
| Kenya | 0 | 0 | 0 | 0 | 37 | 39 | 9 | 44 | 18 | 1 | 148 |
| Rwanda | 0 | 0 | 0 | 0 | 0 | 1 | 2 | 0 | 0 | 0 | 3 |
| Tanzania | 0 | 0 | 0 | 0 | 33 | 12 | 1 | 0 | 0 | 0 | 46 |
| Uganda | 1 | 0 | 1 | 0 | 9 | 19 | 13 | 19 | 19 | 3 | 84 |
| Zambia | 0 | 0 | 0 | 0 | 4 | 4 | 0 | 9 | 2 | 1 | 20 |
| Zimbabwe | 0 | 0 | 0 | 0 | 13 | 0 | 0 | 0 | 1 | 0 | 14 |
| Total | 1 | 0 | 1 | 0 | 96 | 76 | 26 | 82 | 63 | 6 | 351 |
Table 4.
Annual interceptions of T. leucotreta in the EU from 2014 until January 2023, in cut roses versus in all other commodities
| Year | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Commodities | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | Jan‐23 | Total |
| Rosa sp. | 1 | 0 | 1 | 0 | 96 | 76 | 26 | 82 | 63 | 6 | 351 |
| Other commodities | 169 | 260 | 146 | 141 | 112 | 116 | 34 | 56 | 14 | 3 | 1,051 |
| Total | 170 | 260 | 147 | 141 | 208 | 192 | 60 | 138 | 77 | 9 | 1,402 |
Live samples of egg and larval stages of T. leucotreta were repeatedly found during border inspections of cut roses from African countries conducted by the phytosanitary inspection services of the Netherlands (NVWA, 2022: Development stage of intercepted Thaumatotibia leucotreta specimens; personal communication, 12 October 2022; see Appendix A section A.7.1 ) and Belgium (FAVV‐AFSCA, 2022: Reply to EFSA request of info on the FCM cut roses interceptions and checks in Belgium; personal communication, 25 January 2023; data not shown). In experiments conducted by the Dutch NPPO, T. leucotreta adults were bred from some of these larvae under simulated waste‐bin conditions (NVWA 2022: Development of Thaumatotibia leucotreta on cut roses; personal communication, 23 September 2022).
2.2.3. Clustering of T. leucotreta in intercepted consignments
In the interception records of the Dutch NPPO some indications on clustered infestations of T. leucotreta in cut roses were given (NVWA, NVWA, 2022: Infestation rates of consignments of cut roses infested by T. leucotreta; personal communication, 12 October 2022). The Dutch NPPO reported, from all its imports of cut roses between January 2019 and September 2022, that 217 interceptions occurred, from which 77% of the samples had only one specimen, while 15% had two, and 8% had three specimens of T. leucotreta.
Neither the size of the consignment nor the sample size per consignment were recorded in the laboratory's database. As an approximation the Dutch NPPO assumed a median consignment size based on the import data for 2020 of 25,400 roses, which corresponds to a sample size of 400 roses according to the Dutch inspection rules (NVWA, 2022: Infestation rates of consignments of cut roses infested by T. leucotreta, personal communication, 12 October 2022, see also Appendix A Section A.7.2) (Table 5).
Table 5.
Number of T. leucotreta specimens per sample in consignments of cut roses intercepted by the Dutch NPPO in the period January 2019–September 2022 (NVWA, 2022: Infestation rates of consignments of cut roses infested by T. leucotreta; personal communication, 12 October 2022)
| Number of specimens per sample in infested consignments | Number of infested consignments | Total number of T. leucotreta specimens | ||
|---|---|---|---|---|
| Absolut | Relative (% consignments) | Absolut | Relative (% specimens) | |
| 1 | 167 | 77% | 167 | 59% |
| 2 | 32 | 15% | 64 | 22% |
| 3 | 18 | 8% | 54 | 19% |
| 4 or more | 0 | 0% | 0 | 0% |
| Total | 217 | 100% | 285 | 100% |
Changing the view from consignments to specimens: 59% of the specimens were single individuals in the sample of the consignment, while 41% of the specimens were present as two or more insects per sample (= 22% + 19% + 0%; see Table 5 column on ‘Relative [% specimens]’). Using the latter value, two scenarios on the clustering of two or more specimens of T. leucotreta in the imported consignments of cut roses were constructed:
Cluster scenario 1 (worst case): Looking at the 41% of the specimens having two or more insects per sample, it is assumed that at least two specimens:
are on the same cut rose;
have similar life stages (e.g. egg, early larva), develop in parallel and escape at a similar time in the same location.
The likelihood that one insect would have a possible mating partner in the same rose is at least:
Where:
Likelihood of multiple infestation per sample (41%) is derived from Table 5 and described above;
Likelihood of having a female (2/3) and the likelihood of having a male (1/3) are derived from the sex ratio 2:1 of T. leucotreta, meaning two females per one male (Mkiga et al., 2019).
Hence, under this scenario, in total 9% of the insects will be females and will have a partner for mating in the same rose (i.e. at least one in every 11 T. leucotreta).
The worst‐case scenario is not further elaborated, because the Panel considers it as unlikely referring to:
-
–
the egg laying behaviour of the insect (according to COLEACP et al., 2020, the T. leucotreta female moth lays over 100 eggs at night, usually singly on flower petals or other parts of the rose bush);
-
–
the lack of records from border inspections on multiple T. leucotreta specimens on a single cut rose. Although, according to COLEACP et al. (2020), generally only one to three larvae survive in each rose flower, there is no record so far of multiple T. leucotreta specimens on a single cut rose upon EU import border inspection.
Cluster scenario 2 (realistic case): Looking at the 41% of the specimens having two or more insects per sample, it is assumed that:
Infested roses have no more than one T. leucotreta specimen per cut rose;
Distribution of the cut roses is done in bunches of 10 cut roses;
The specimens of T. leucotreta are uniformly distributed among the bunches of cut roses;
The specimens of T. leucotreta in a bunch of cut roses have similar life stages (e.g. egg, early larvae), develop in parallel and escape at similar time in the same location from the bunch.
The likelihood that one insect would have a possible mating partner in the same bunch of 10 roses is at least:
Where:
Likelihood of multiple infestation per sample (41%) is derived from Table 5 and described above;
Likelihood of multiple infestation per bunch (2.5%). A sample of 400 cut roses (as described above) has 40 bunches of 10 rose stems each; assuming that one bunch is infested with one insect, there is one out 40 possibilities for a second insect to infest the same bunch;
Likelihood of having a female (2/3) and the likelihood of having a male (1/3) are derived from the sex ratio 2:1 of T. leucotreta, meaning two females per one male (Mkiga et al., 2019).
Hence, under this scenario, in total 0.23% of the insect will be females and will have a partner for mating in the same bunch of 10 roses (i.e. at least one in every 435 T. leucotreta).
This realistic scenario is further elaborated in the assessment, because it is representing typical market conditions and egg laying behaviour of the insect.
Clustered scenario 3 (best case): in Appendix A, Section A.1.4, the Panel has also estimated the average number of escaped T. leucotreta adults per 1 km radius in the residential areas of the NUTS2 regions within a 10‐day period. This scenario could be interpreted as ‘best case’, when no temporal or spatial clustering of the cut roses consumption occurs, and all escaping adults are homogeneously distributed within the residential area of a NUTS2 region throughout the year.
2.2.4. Records of T. leucotreta in the EU, the UK and USA
Since 1965, more than 30 adult specimens of T. leucotreta have been reported in the EU. Occasional records have been made in Finland (Karvonen, 1983), the UK (Knill‐Jones, 1994; Langmaid, 1996; NBN atlas, 1997), Denmark and the Netherlands (Huisman and Koster, 2000), Sweden (Svensson, 2002), France (Rogard, 2015), Czechia (Šumpich et al., 2021, 2022) and Belgium and Germany (Rennwald, 2022). Recordings consisted of a single adult specimen either found in private homes, likely imported as larvae with produce from Africa (oranges; FI: Karvonen, 1983; NL: Huisman and Koster, 2000; SE: Svensson, 2002; DE: Rennwald, 2022) or captured in light traps outdoors (the UK: Langmaid, 1996; NBN atlas, 1997; FR: Rogard, 2015; BE and DE: Rennwald, 2022). However, these were all isolated findings without any evidence of established populations.
Since 2009, three incursions 3 of T. leucotreta have been reported in Europe: In the Netherlands, one larva was found in a glasshouse on habanero peppers (Capsicum chinense) in 2009 (EPPO, 2010; Potting and van der Straten, 2011), and one larva and three adults on sweet peppers (Capsicum annuum) in 2013 (EPPO, 2014); and in Germany, one male was found in 2018 on a pheromone trap in a glasshouse producing sweet peppers (C. annuum) (EPPO, 2018). These incursions have been found near packaging facilities of imported fruits (the Netherlands) and near a supermarket waste container (Germany). No populations were established.
A single adult male was detected in a trap in California (USA) in 2008 (Gilligan et al., 2011), but, in spite of extensive surveys conducted for T. leucotreta throughout the state, no further detections of the pest occurred.
2.3. Entry
The process of pest introduction is defined by the IPPC (International Plant Protection Convention) as the entry of a pest resulting in its establishment 4 (IPPC Secretariat, 2022 ). Introduction can therefore be divided into the assessment of pest entry and the assessment of pest establishment, with the process of pest transfer to a host being the step that links entry to establishment.
Estimation of the number of T. leucotreta adults escaping from the cut roses imported from African countries into the EU was performed for the NUTS2 regions identified as suitable for establishment as described in Section 2.4.
To estimate the number of T. leucotreta adults that will escape from infested cut roses imported from countries with reported occurrence of T. leucotreta, a pathway model was used that consisted of three submodels: (i) a rose distribution model (Section 2.3.1) describing how the imported and infested roses are distributed into the NUTS2 regions in the EU area suitable for establishment, (ii) a developmental model (Section 2.3.2) describing the proportion of T. leucotreta adults emerging depending on the number of days after import into the EU and (iii) a waste model (Section 2.3.3) estimating which proportion of T. leucotreta will survive and escape different waste treatments. The last component was included because the life cycle of imported cut roses was assumed to end with cut roses being disposed. For the assessment of entry as estimated by the overall entry model (Section 2.3.4), and of potential establishment of T. leucotreta, it is therefore important to estimate the proportion of T. leucotreta that survives and escapes all possible waste treatments.
2.3.1. The cut roses distribution model
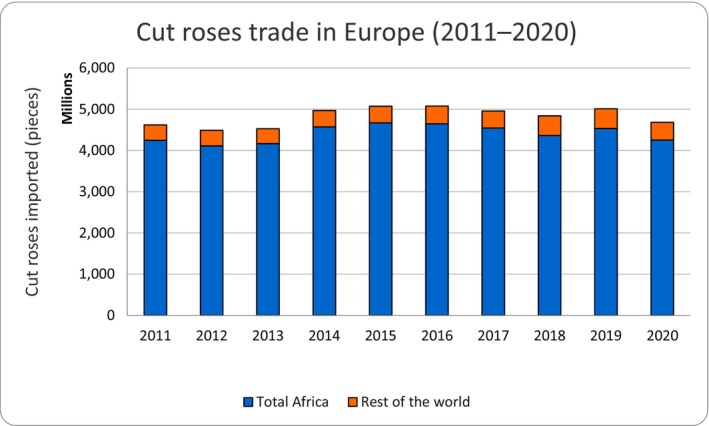
The average annual volume of cut roses imported into EU MS over the years 2011–2020 was 4,825 million stems. Most of the imported roses (91%) came from Africa and Israel (4,411 million stems). The remaining 9% (414 million stems) entered from the rest of the world, mainly Ecuador and Colombia, followed by the UK (Figure 2). The remaining countries contributed with only 1.5%. The EU countries on average exchanged 3,039 million stems among themselves. This includes trade of their own production and internal EU trade of imported cut roses from third countries.
Figure 2.
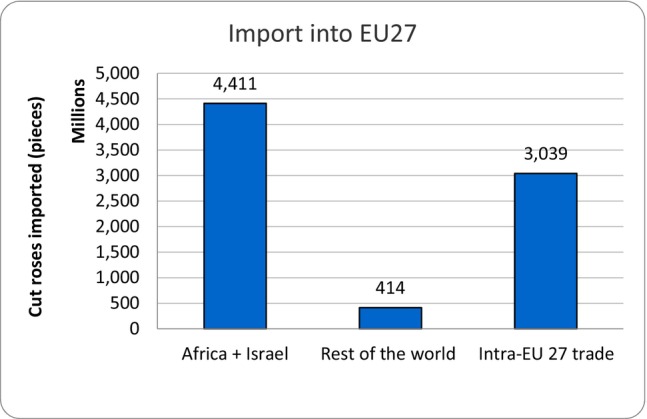
Trade in cut roses in the EU (average of 2011–2020)
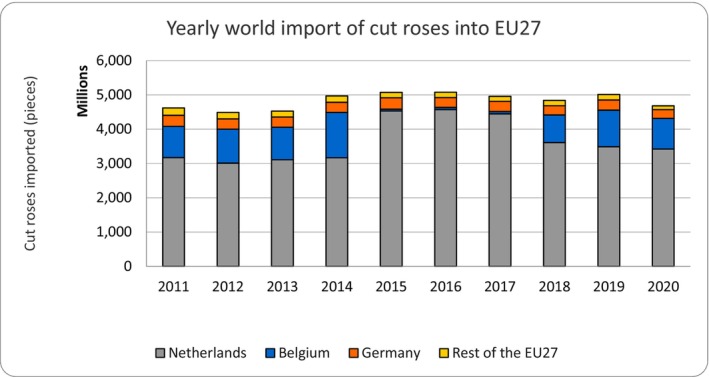
The volume of cut roses imported into the EU has been relatively stable in period 2011–2020 (Figure 3), reaching a peak in 2016. The Covid‐19 pandemic had only a marginal effect on the trade as indicated by the slight reduction in 2020.
Figure 3.
Trends in the EU of import of cut roses (2011–2020)
The imported roses are not distributed evenly through the EU. Most of the cut roses (on average 75.5%) enter the Netherlands, 15.0% enter Belgium, 6.1% enter Germany and only 3.4% enter the remaining 24 EU countries (Figure 4). However, the intra‐EU trade complicates the calculations of the pathway model for the EU 27 MS, e.g. between 2015 and 2017 the import into Belgium was reduced, while that of the Netherlands was increased by a similar amount, indicating intra‐EU trade relations between neighbouring countries. Due to the observed intra‐EU trade relations, to limit the pathway model to the main intra‐EU trade relations, some EU countries were therefore clustered for manageability of the pathway model calculations: ‘The Netherlands/Belgium’, ‘Germany/Luxembourg’, ‘Spain/Portugal’, ‘Italy/Malta’ and ‘Greece/Cyprus’.
The pathway of cut roses in the EU starts with entries mainly via airplane followed by repacking, auction, wholesalers and retail sales to the consumers (Figure 5).
Figure 5.
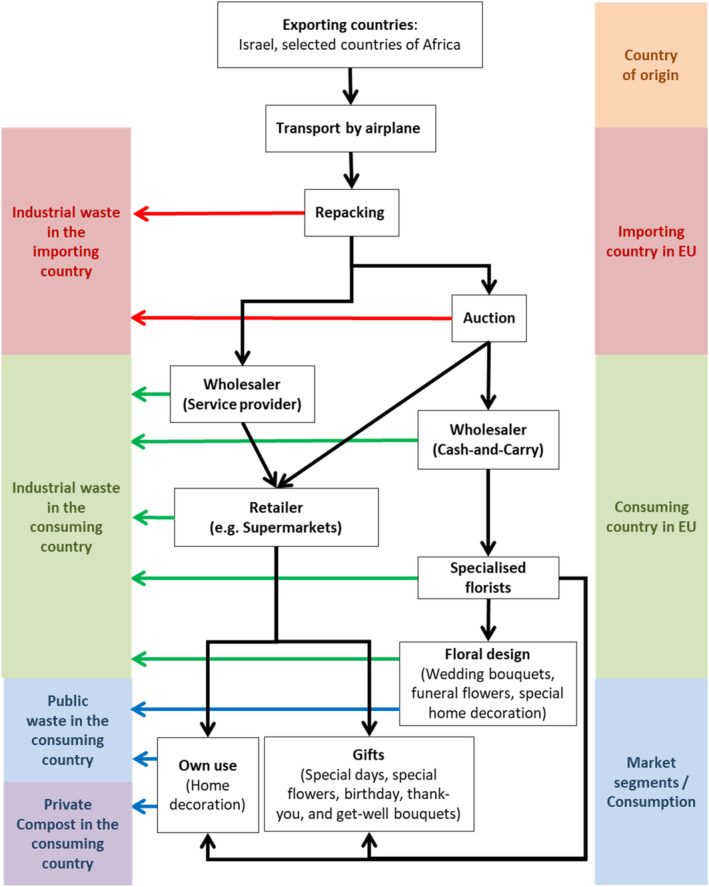
Pathway of cut roses from import to waste (adapted from CBI, 2017)
All cut roses will finally end in the waste, differentiated by commercial waste in different countries, public waste and private compost.
According to CBI (2017), the European cut flower market consists of two market channels: one focusing on specialised florists and the other on unspecialised retail (e.g. supermarkets). The main differences between the channels are the role of the flower auction and wholesalers, and the characteristics of the products.
Some of the imported cut roses are sold directly to high‐end customers, but most are reserved for Dutch auctions, a major hub in the redistribution of cut flowers in the EU and worldwide. In 2020, in total 2,912 million stems were imported from African countries (CBS, 2022), 2,814 million stems were traded through Dutch auctions (Royal Flora Auctions, 2021). A minor part of cut roses consumed in the EU is produced in European greenhouses (the Netherlands 20.0%, Germany 0.4% and Belgium 0.2%). A large part (78.0%) of the cut rose stems auctioned in the Netherlands are imported from rose farms in Africa (Ethiopia 44.0%, Kenya 31.0%, Uganda 1.5% and others from Rwanda, Tanzania, Zimbabwe, Zambia and South Africa) (see Appendix A). Germany and France are the major intra‐EU trade markets for auctioned roses (Rabobank, 2022). From the auction houses, cut roses are redistributed to various points of sale at retailers, any delay affects their vase life. Direct sales can drastically reduce the farm‐to‐vase time.
Due to missing precise information on the proportion of trade via specialised florists and supermarkets, the pathway model only distinguishes direct import into the consuming country/NUTS2 region, and re‐exporting via European countries (indirect trade, see Figure 6).
Figure 6.
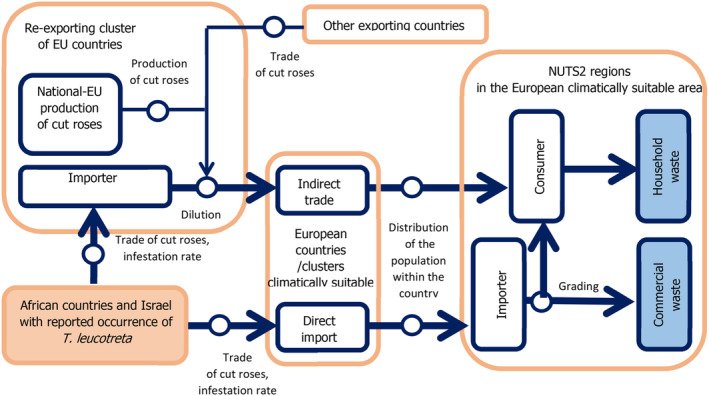
Pathway of cut roses from import to waste as conceptualised for the rose distribution model
The waste resulting from rejected roses by grading/repacking will end in the commercial waste of the consuming country or stays in the importing country. A diagram presenting the pathway model for the rose distribution model is outlined in Figure 6.
The rose distribution model comprises two main pathways: ‘direct’ and ‘indirect’ import into the EU area climatically suitable. The latter one is further divided into two main clusters for intra‐EU trade: ‘The Netherlands/Belgium’ and ‘Germany/Luxembourg’.
-
Direct import of cut roses from African countries and Israel, with reported occurrence of T. leucotreta by a country with areas climatically suitable.
For this pathway, the trade volume reported by Eurostat on national and monthly basis was considered in the model. To cover most of the trade between the countries within the areas climatically suitable, the following country clusters were established: (i) Spain and Portugal, (ii) Italy and Malta and (iii) Greece and Cyprus. In the model, it is assumed that the distribution within a cluster will follow the distribution of potential consumers, thus the distribution of the population within the countries/clusters. Different regional consumption habits are not considered.
In this pathway, importers, including when applicable wholesalers and distribution centres, are in the NUTS2 region of consumption and it is assumed that they perform the necessary grading and repacking.
-
The cut roses are imported by the Netherlands and Belgium and further traded ‘intra‐EU’ to the EU countries with areas climatically suitable.
Because only the total trade volume between the Netherlands/Belgium and other EU flower consuming countries is reported to Eurostat, it is assumed that the imported cut roses (from Africa and Israel) will be equally diluted by other imports of cut roses, and the production in the Netherlands and Belgium. Preferences for the intra‐EU re‐trade of African cut roses are not considered.
In this pathway, grading and repacking is assumed to be done in the Netherlands and Belgium, and then, the cut roses will go directly to retailers or specialised florist shops in the consuming countries.
It is assumed that the national EU distribution will follow again the distribution of potential consumers, thus the distribution of the population within the cluster. Different regional consumption habits are not considered.
-
The cut roses are imported by Germany and Luxembourg and further traded intra‐EU the countries with areas climatically suitable.
For this pathway, the same assumptions made for re‐exporting for the Netherlands and Belgium are valid (see Section 2 for details).
No other European cluster for the re‐export of African roses is considered due to minor trade volumes. Due to the low infestation, the model further assumes that an infested rose contains one insect, and this could be in any life stage.
In all scenarios, the annual trade is further stratified by seasons: December–February, March–May, June–August, September–November; to reflect the different climatic condition per season (see developmental model).
The overall number of T. leucotreta individuals per season ending in commercial or household waste in a NUTS2 region climatically suitable in a specific season was therefore calculated as below. A description of the parameters used in the rose distribution model is presented in Table 6.
Table 6.
Description and source of evidence for the parameters used in the rose distribution model
| Parameter | Description | Source | Distribution |
|---|---|---|---|
| FCMcommercial, NUTS2, Season FCMhousehold, NUTS2, Season | Number of T. leucotreta individuals ending in commercial or household waste in a NUTS2 region climatically suitable during a specific season | Calculated | |
| AF | African countries and Israel with reported occurrence of T. leucotreta | EPPO Global Database (Appendix A, Section A.2) | Strata |
| EURA | European countries/clusters with areas climatically suitable/under risk | Countries with at least one NUTS2 region with a climate suitability classification by the physiologically based demographic model above 0 (Appendix A, Section A.3) | Strata |
| NUTS2 | NUTS2 regions climatically suitable | NUTS2 regions with at least one grid cell with a climate suitability classification by the physiologically based demographic model above 0 (Appendix A, Section A.3) | Strata |
| NLBE | Cluster of re‐exporting countries: the Netherlands/Belgium | Annual trade of cut flowers from 2011 to 2020 (Appendix A, Section A.5) | Strata |
| DELU | Cluster of re‐exporting countries: Germany/Luxembourg | Annual trade of cut flowers from 2011 to 2020 (Appendix A, Section A.6) | Strata |
| Season | Stratification by season: winter, spring, summer, autumn; or total | Strata | |
| TradeAF‐EURA, Season | Direct trade of cut roses from AF to European climatically suitable countries/clusters (Unit: [pcs]) | Eurostat monthly trade of cut roses (CN 03061100) in 2011–2020 (Appendix A, Section A.4) | NORMAL |
| PopulationNUTS2/EURA | Proportion of the human population in the NUTS2 region climatically suitable in relation to the whole country or cluster (Unit: [−]) | Eurostat population 1 January 2020 (Appendix A, Section A.4.2) | CONSTANT |
| Grading | Proportion of direct imported cut roses, which on average is discarded due to quality issues (Unit: [−]) | Default value (Appendix A, Section A.4.4) | UNIFORM |
| TradeAF➔NLBE, Season TradeAF➔DELU, Season | Trade of cut roses from AF to the European re‐exporting clusters in a specific season (Unit: [pcs]) | Eurostat monthly trade of cut roses (CN 03061100) in 2011–2020 (Appendix A, Sections A.5, A.6) | NORMAL |
| TradeOthers➔NLBE, Season TradeOthers➔DELU, Season | Trade of cut roses from other countries to the European re‐exporting clusters in a specific season (Unit: [pcs]) | Eurostat monthly trade of cut roses (CN 03061100) in 2011–2020 (Appendix A, Sections A.5, A.6) | NORMAL |
| ProductionNLBE, Season ProductionDELU, Season | Average production of cut roses in European re‐exporting clusters in a specific season (Unit: [pcs]) | Calculated | |
| AreaNL AreaDE | Average, annual area for the production of cut roses in specific MS (Unit: [ha]) | National statistics on cut roses production surface | CONSTANT |
| ExtrapolationNL➔NLBE ExtrapolationDE➔DELU | Extrapolation from national production area to the area in a re‐exporting cluster (Unit: [−]) | Eurostat farm structure | CONSTANT |
| Conversionha➔pcs | Productivity of cut roses (Unit: [pcs/ha]) | Productivity of different roses | CONSTANT |
| ProportionSeason/Total | Proportion of cut roses produced in the re‐exporting clusters in a specific season compared to the annual production | Default value: equally distributed | CONSTANT |
| DilutionNLBE,Season DilutionDELU,Season | Dilution of T. leucotreta‐infested cut roses, imported from Africa and Israel, by other imports and own production in a specific re‐exporting cluster and season (Unit: [−]) | Calculated for scenarios without/with Intra‐EU trade | UNIFORM |
| InfestationAF | Average infestation rate of cut roses from AF (Unit: [−]) | EKE question 2 (Appendix A, Section A.7) | GENERALBETA |
It is assumed that commercial waste only appears in the grading step at import or auction. Thus, for indirect import (intra‐trade) via EU countries outside the EU climatically suitable areas (the EU climatically suitable area is indicated in the equations as the EU risk area, abbreviated as EURA), no commercial waste has to be considered. For the direct import, the number of T. leucotreta (FCM) entering a NUTS2 region in the EU climatically suitable area in a specific season (FCMcommercial, NUTS2 Season) is calculated as:
where TradeAF‐EURA, Season, InfestationAF, PopulationNUTS2/EURA and Grading are the volume of direct import from Africa into a country cluster in a specific season, the infestation rate of African roses, the proportion of the population within a NUTS2 region climatically suitable (compared to the total population of the country cluster) and the proportion of roses deselected by grading due to quality problems or similar, respectively.
Because all roses will finally be wasted, the household waste consists in the EU climatically suitable areas of the marketed (not deselected during grading) roses, and the imported roses from the ‘Dutch/Belgian’ and ‘German/Luxembourg’ cluster.
Roses re‐traded intra‐EU are diluted by national production and by other imports into these clusters (‘Dutch/Belgian’ and ‘German/Luxembourg’ clusters). The dilution factor is calculated as proportion of African and Israelian roses in relation to the total amount of roses within the corresponding country cluster. The total amount of roses in a country cluster consists of import from Africa, other imports and own production. This assumes that all roses within an intra‐EU trade cluster are mixed before intra‐EU re‐trade.
Missing information on the cut rose production of Belgium and Luxembourg was extrapolated from the Dutch and German production and converted from the production area.
2.3.2. The developmental model
The developmental model of T. leucotreta after entry into the EU (start at the border) estimates the number of T. leucotreta adults emerging in the importing country considering three sequential compartments:
-
Transport and storage under cold conditions. This compartment includes the handling after entry, cold transportation to the region for consumption, and cold storage there. After harvest and sorting on site in Africa, cut roses are stored in water at a temperature of 2–6°C. As a result, the moisture content is maintained. During transport by truck/trailer, a constant temperature of 4–6°C and humidity level (60–80%) is maintained to deliver the flowers to destinations in optimal condition (at 4°C ± 1°C, Flower Watch, 2011; Carrier Transicold, 2022). Temperatures, however, may fluctuate for short periods during handling (up to 8°C) and reloading (short periods up to 16°C) from storage into transport and from trucks into the aircraft cabin (Flower Watch, 2011; Carrier Transicold, 2022) Cut roses are mainly transported by air freight. A total turnover time from the grower up to and including the auction takes between 1 and 2 days by air transport (Flower Watch, 2011; Carrier Transicold, 2022). Upon arrival in Europe (mostly the Netherlands), the time from purchasing trade to the retailer takes a maximum of ~ 4 days at a temperature of 8°C ± 1°C, 75% RH.
During the whole chain from grower to retailer, a constant temperature of 8°C is assumed as a conservative (i.e. worst) scenario.
The duration of cut roses in this model compartment assumes a triangular distribution with parameters minimum, most likely and maximum equal to 1.0, 4.5 and 8.0 days, respectively, that adequately describe the uncertainty in the duration of transport. These values were also set considering the interest in keeping the transport time as short as possible by the flower industry. Additionally, the cold storage at the final place of sales to consumers is part of this compartment. It should be noted that even considering the extreme values for temperature and duration, transportation and cold storage do not support full development of T. leucotreta.
-
Retail and private home under climatisation. This compartment includes the provision to and the use of the flowers by the consumer.
Vase life at the consumer is aimed at a period of 7.0–10.0 days for imported cut roses (VBN, 2017; Harkema et al., 2017) and 2 weeks for locally produced cut roses (FloraNews, 2015), but can vary between 4.0 and 14.0 days (Yakimova et al., 1996; Ichimura et al., 2006) Hence, the duration of cut roses in this compartment was modelled assuming a triangular distribution with parameters minimum, most likely and maximum equal to 7.0, 10.5 and 14.0 days, respectively. Temperature in this compartment is assumed to be set to the ideal temperature values for human living and therefore modelled assuming it could adequately be described by a triangular distribution with parameters minimum, most likely and maximum equal to 18.0, 21.0 and 24.0°C, respectively.
Organic waste at ambient temperature. This compartment comprises the situation after waste disposal by the consumer. It is assumed that the temperatures are ambient according to the regional and seasonal situation. Due to missing information on the duration within this compartment, four scenarios for the time between waste disposal and waste treatment are constructed: 3, 7, 14 and 28 days (see the waste model in Section 2.3.3).
A diagram presenting the compartment model (time on the horizontal axis) for the developmental model is outlined in Figure 7.
Figure 7.
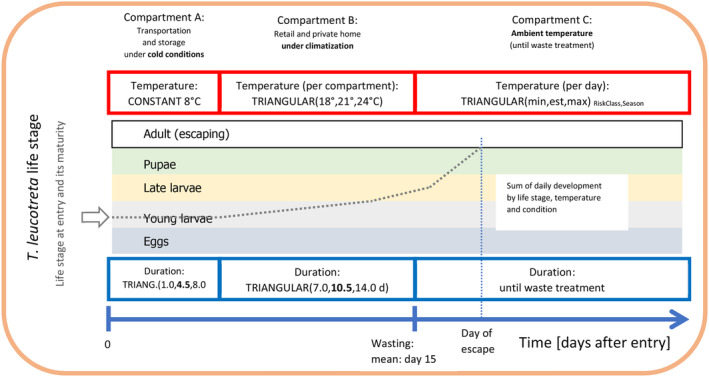
Visualisation of three phases of development after entry of cut roses. The blue axis at the bottom represents the time axis and three phases: (1) transport, (2) vase life, (3) post‐waste disposal. The red boxes on top represent the assumed temperature during each phase, while the blue boxes at the bottom represent the duration. The dotted line in the middle of the graph indicates how larvae on the roses progress through their life stages during the three stages of the cut roses pathway, from production to waste.
The following simulation process was done using @RISK software version 7.6 (see Appendix C): An individual of T. leucotreta will enter the EU at a specific life stage. According to the temperature, the individual will develop each day a bit, additionally depending on a random component related to the insect and host conditions (see @RISK file data Appendices). Individuals will develop through successive life stages accordingly. When emerging from the pupae, the adult moth will escape the cut rose or waste. The day of escape was simulated for 10,000 T. leucotreta arriving in the EU. The simulation results in a profile of life stages for each day after entry, or the proportion of adults for a fixed day depending on the ambient temperatures for specific climate suitability classes and seasons. A description of the parameters used to inform the developmental model is presented in Table 7.
Table 7.
Description and source of evidence for the parameters used in the developmental model
| Parameter | Description | Source | Distribution |
|---|---|---|---|
| ESCtime, xd, CSClass, Season | Proportion of T. leucotreta which will escape the cut roses until x days (x = 3, 7, 14 or 28 days depending on the scenario) after initial disposal (at day 15) in a region of a specific climate suitability class and season | Calculated | |
| PropEggs PropYoung/all larvae | Proportion of T. leucotreta life stages, when arriving at the border | EKE question 1a and 1b (Appendix A Section A.8) | GENERALBETA |
| Maturity | Level of maturity within a specific life stage, when arriving at the border | Uninformative on age | UNIFORM |
| DevelopLifeStage,Temp Condition | Daily development of a specific life stage according to the temperature and a random component related to the insect and host conditions. Reciprocal of the duration of the specific life stage at a specific temperature. The condition is the position within a range of values | Scientific literature | TRIANGULAR with correlation of the life stages |
| TemperatureCompA TemperatureCompB TemperatureCompC | Temperature during the stay in a compartment. For compartments A and B, the temperature is assumed as constant, while for compartment C, the temperature may change every day | Default values for compartment A and B, seasonal weather data for 2001–2010 (at NUTS2 level) | TRIANGULAR |
| DurationCompA DurationCompB | Duration of the stay in compartments A and B | Default values according to the objectives of the flower industry | TRIANGULAR |
2.3.3. The waste model
The waste model estimates the proportion of T. leucotreta that survive and escape different waste treatments: private compost and the communal treatments (landfill, composting and incineration/anaerobic digestion). The proportion of each treatment is estimated for household and commercial (sorted vegetal) waste.
Because no data on the duration from initial disposal to waste treatment for normal household waste were available, and a dependence on local conditions is assumed, the calculations were done for four different times: treatment 3.0, 7.0, 14.0 and 28.0 days after initial disposal at the household.
To model the escape of T. leucotreta moths before the treatment of the waste, four scenarios for the collection of household waste are calculated:
Scenario 1: Fast collection and treatment of household waste. In this scenario, treatment occurs 3 days after initial disposal (day 18 after entry). This is a ‘best case’ scenario where consumers would take their waste for collection on the same day that the cut roses were disposed of, and waste is also collected within 24 h and treated without delays.
Scenario 2: Treatment occurs 7 days after initial waste disposal (day 22 after entry). This is for example the case where roses are kept for a few days in the waste bin at private consumers before being taken out for collection. The rest of the waste management chain runs smoothly with a possible 1‐ or 2‐day delay between collection and treatment.
Scenario 3: Treatment occurs after 14 days after initial disposal (day 29 after entry). This is a scenario assuming more severe delays, e.g. when roses are kept for a few days in the waste bin at the private consumer's before being taken out for collection. At the same time, there is a delay in either the waste collection or treatment, e.g. by intermediate storage.
Scenario 4: Slow collection and treatment of household waste. In this scenario, treatment occurs 28 days after initial disposal (day 43 after entry). This should be regarded as the ‘worst‐case’ scenario in which long delays occur and there is maximum opportunity for T. leucotreta adults to escape from waste. For example, the waste is not taken out for collection every week and/or there is a longer storage in the waste management chain before the treatment.
For the commercial waste, the Scenario 1 (3 days after initial disposal) is assumed. The pathway of the waste model is presented in Figure 8.
Figure 8.
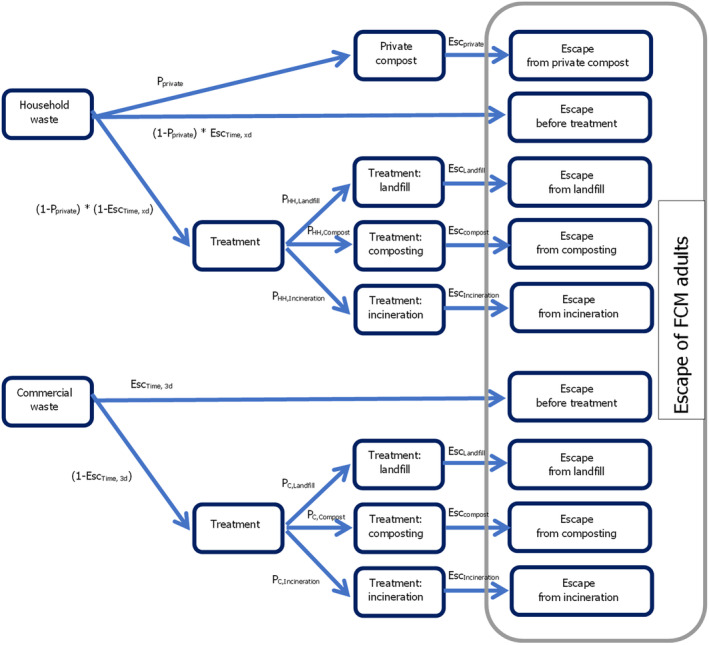
Pathway model for the waste management component (see text below, Table 8 and Appendix A for details)
The overall number of T. leucotreta individuals/year escaping from the household or commercial waste were therefore calculated as the escape from private compost, escape from household waste before the treatment and escape during the three types of treatment: landfill, composting or incineration.
Additionally, T. leucotreta may escape the commercial waste. This may happen before the treatment (within 3 days after wasting), or during the different types of treatment.
A description of the parameters used to inform the waste model is presented in Table 8.
Table 8.
Description and source of evidence for the parameters used in the waste model
| Parameter | Description | Source | Distribution |
|---|---|---|---|
| ESCHousehold, NUTS2, Season | Proportion of T. leucotreta, which will escape the cut roses used at consumer level or household waste | Calculated | |
| ESCCommercial, NUTS2, Season | Proportion of T. leucotreta, which will escape the commercial waste of cut roses | Calculated | |
| Pprivate | Proportion of household waste, which goes to private composting (not subjected to regional waste treatment) | EKE question 3 (Appendix A.9) | GENERALBETA |
| ESCtime, xd, CSClass, Season | Proportion of T. leucotreta, which will escape the cut roses until x days after initial disposal in a region of a specific climate suitability class (CSClass) and season | Calculated by the developmental model | |
| PHH,Landfill,country PC,Landfill,country | Proportion of household (HH) or commercial (C) waste treated by ‘landfill’ | Eurostat waste treatment for household or vegetal waste (Appendix A.9.2 and A.9.3) | CONSTANT |
| PHH,Compostl,country PC,Compostl,country | Proportion of household (HH) or commercial (C) waste treated by ‘composting’ | Eurostat waste treatment for household or vegetal waste (Appendix A.9.2 and A.9.3) | CONSTANT |
| PHH,Incineration,country PC,Incineration,country | Proportion of household (HH) or commercial (C) waste treated by ‘incineration or anaerobic digestion’ | Eurostat waste treatment for household or vegetal waste (Appendix A.9.2 and A.9.3) | CONSTANT |
| ESCprivate | Proportion of T. leucotreta, which will escape private compost | Conservative assumption | CONSTANT = 100% |
| ESCLandfill | Proportion of T. leucotreta, which will escape waste treated by ‘landfill’ | EKE question 4a (Appendix A.9.4) | UNIFORM |
| ESCCompost | Proportion of T. leucotreta, which will escape waste treated by ‘composting’ | EKE question 4b (Appendix A.9.4) | UNIFORM |
| ESCIncineration | Proportion of T. leucotreta, which will escape waste treated by ‘incineration or anaerobic digestion’ | EKE question 4c (Appendix A.9.4) | CONSTANT = 0% |
Waste treatments for both commercial and household waste include landfill, composting (and home composting), incineration or anaerobic digestion. A high variability in the proportions of waste treated by the different methods occurs across MSs, particularly in relation to the household waste (Figures 9 and 10).
Figure 9.
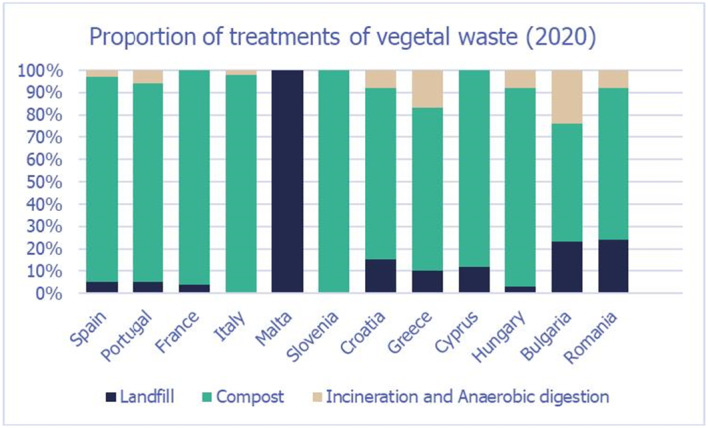
Proportion vegetal waste 5 treated as landfill, compost and incineration/anaerobic digestion in the European countries considered in the assessment
Figure 10.
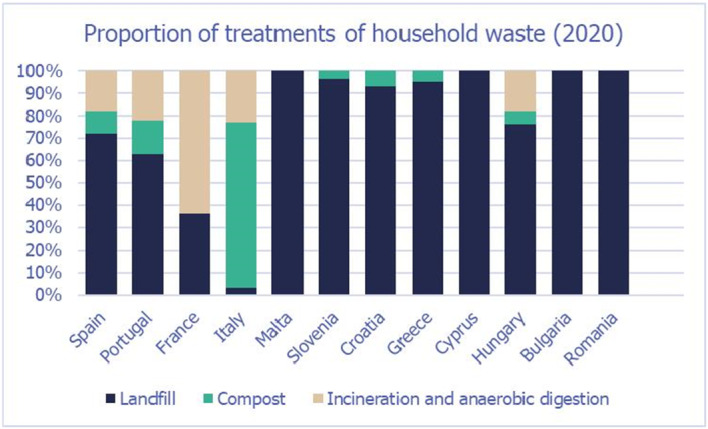
Proportion household waste treated as landfill, compost and incineration/anaerobic digestion in the European countries considered in the assessment
5 For each type of treatment, EKE was used to determine the proportion of T. leucotreta that survives and escapes the treatment. The main characteristics of the different treatments are described in Appendix A.9. This information was used as part of the evidence for the expert judgement on: (i) proportion of organic waste going to private composting (Appendix A section A.9), (ii) survival rate of T. leucotreta at landfill, during composting and incineration/anaerobic digestion (Appendix A Sections A.9.4, A.9.5 and A.9.6).
For the scope of the opinion, the following assumptions were made: (i) Public waste management is also applicable for cut roses wasted by private consumers and (ii) commercial waste of cut flowers will be handled as treatment of vegetal waste.
2.3.4. The overall entry model
The overall model for entry combines the three submodels and calculates the average number of T. leucotreta adults escaping from cut roses over all seasons according to the equation below.
To adjust for the natural mortality occurring during the development of the insect along the pathway, an overall mortality factor of 44–60% was introduced that reduced the total number of escaping adults.
Neither data nor information were found in the scientific and grey literature on the mortality of T. leucotreta in cut roses or in organic waste; however, in other commodities (sweet orange and sweet pepper, see Section 3.1.2), a larval mortality range was reported between 25% and 37%. Data or information on pupal mortality rate for T. leucotreta are also lacking; however, the Panel assumes that for T. leucotreta the pupal mortality and the larval mortality are comparable, as it is the case for other Tortricidae species (Milonas and Savopoulou‐Soultani, 2000, Gutierrez et al., 2012). Therefore, as there are no data on mortality of T. leucotreta in cut roses and in organic waste and considering also that the temperature‐dependent mortality remains low in the range of temperatures experienced by cut roses from entry to disposal (see Figure 16), a mortality rate between 25% (such as in sweet oranges) and 37% (sweet pepper) for developing larvae at 25.0 ± 2.0°C was used as a lower bound of the overall mortality for larvae in the cut roses pathway. Further assuming that the mortality range (25–37%) would be similar also for pupae, an overall mortality range of 44–60% from larvae to pupae was estimated and applied overall to the entry model results on the numbers of adult moths predicted to escape per year.
Figure 16.
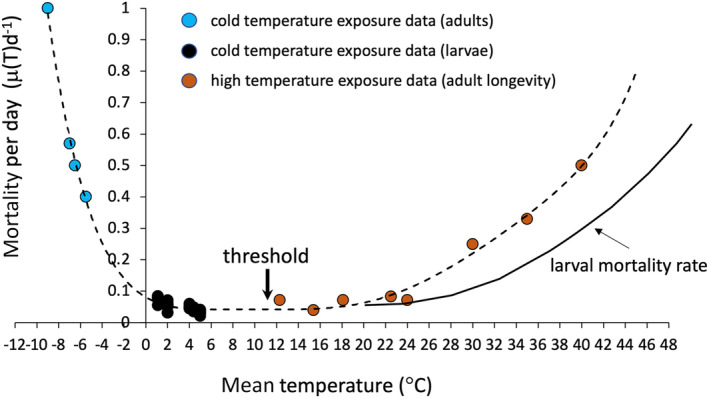
The biodemographic functions for larval and adult temperature‐dependent mortality (source: Myburgh, 1965; Daiber, 1980; Boardman et al., 2012, 2013; Moore et al., 2016, 2022) with the 4°C displacement of larval mortality rates (see Terblanche et al., 2017) indicated as a solid line. Same mortality function was assumed for eggs and pupae (see, e.g. for other Tortricidae species: Milonas and Savopoulou‐Soultani, 2000, Gutierrez et al., 2012)
Descriptions of the parameters used in the overall model are reported in Table 9.
Table 9.
Description and source of evidence for the parameters used in the final model
| Parameter | Description | Source | Distribution |
|---|---|---|---|
| FCMEscape, NUTS2 | Annual average number of T. leucotreta adults escaping from cut roses imported from AF (African countries with FCM occurrence and Israel) in a specific NUTS2 region | ||
| Mortnatural | Natural developmental mortality |
Observed larval mortality on other crops Extrapolation to pupae |
UNIFORM |
| FCMCommercial, NUTS2, Season FCMHousehold, NUTS2, Season | Number of FCM T. leucotreta individuals ending in commercial or household waste in a NUTS2 region climatically suitable in a specific season | Calculated in the distribution model | |
| ESCHousehold, NUTS2, Season | Proportion of T. leucotreta which will escape the cut roses used at consumer level or household waste | Calculated in the waste model | |
| ESCCommercial, NUTS2, Season | Proportion of T. leucotreta which will escape the commercial waste of cut roses | Calculated in the waste model |
Results of the overall model are expressed as average number of adult escapes per NUTS2 region with suitable climatic conditions for establishment.
To provide an interpretation of the results in terms of possible mating under different level of clustering, the results of the final model are also transformed to represent two scenarios:
Cluster Scenario 2: Clustering in bunches of 10 roses as defined in Section 2.2.3. Under this scenario, a mating will happen for every 435 escaping T. leucotreta (No. mated femalesNUTS2/year = FCMEscape, NUTS2/435) (see results in Sections 3.4.1 and 3.4.2 and in Appendix A Sections A.1.2 and A.1.3).
Cluster scenario 3: No temporal or spatial clustering and all escaping adults are homogeneously distributed within the residential area of a NUTS2 area and throughout a year. To this end, the average number of adult escapees is standardised to a circle of a 1‐km radius (flying radius of T. leucotreta males), considering only the residential area of each NUTS2 regions during a time of 10 days (because the majority of eggs are laid within the first 10 days of female adult stage) (see results in Section 3.4.3 and in Appendix A Section A.1.4).
Results transformed as the number of mated females NUTS2/year are presented for the EU and some selected NUTS2 regions as disaggregated values by season for all scenarios of time between waste disposal and waste treatment (i.e. 3, 7, 14 and 28 days).
To combine the uncertainties in the model parameter (expressed as distributions), the model is implemented in a Monte‐Carlo simulation 10,000 times (using @RISK version 7.6) (see Appendix D).
2.3.5. Uncertainties of entry
Uncertainties on entry are generally quantitatively assessed in the model and in the EKE. Aspects not quantified and assumptions are described in Section 3.3. To identify the parameters of the pathway model driving most of the uncertainty on the output estimates, a sensitivity analysis was conducted on each scenario (3, 7, 14 and 28 days) by calculating the relative decomposition of R2 from the calculated correlation coefficients between inputs and outputs.
2.4. Establishment
2.4.1. Climate suitability methodology
In the EPPO PRA for T. leucotreta (EPPO, 2013), a CLIMEX compare locations model was explored but then abandoned because of lack of knowledge on factors influencing winter survival and the climatic limits of its distribution. A simple rule based on diurnal temperatures (based on the difference between weekly maxima and minima) was adopted although recognising its uncertainty (as it was based on very few locations and there was uncertainty of the characteristics of the coldest winter that T. leucotreta could survive).
Other quantitative approaches to assess T. leucotreta establishment were found in literature but not all addressing the EU region (Table 10 below). The Panel used two approaches for assessing the area of potential establishment of T. leucotreta. The first is the Köppen–Geiger climate classification (MacLeod and Korycinska, 2019) matching climate categories in the EU with those in locations where T. leucotreta is known to occur in Africa and Israel (see Section 2.2.1 and Figure 1). The second is a physiologically based demographic model (hereinafter referred to as PBDM; Gutierrez, 1996).
Table 10.
Methodological approaches to model climate and niche suitability of T. leucotreta
| Model basis | Source | EU data |
|---|---|---|
| Host presence + pest occurrence + climate characteristics | EPPO (2013) | Yes, map |
| Pest occurrence + climate categories | (Köppen–Geiger) in this EFSA Scientific Opinion; Rossi et al. (2023) | Yes, map (Figure 23) |
| Pest occurrence + host presence | Venette et al. (2003) | None |
| Pest occurrence + degree‐days + host presence | Li et al. (2022) | Yes, map (pers.comm, Figure 27) |
| Pest biology+Degree‐day + cold stress (CLIMEX) | Barker and Coop (2019) | Yes, map (pers.comm) |
| Physiologically based demography | Gutierrez and Ponti (2013, 2023a,b) | Yes, map (Figure 25 and Figure 26) |
In this document, the establishment is measured in terms of interannual average abundance of life stages of the pest, given entry. Establishment is interpreted as the similarity of predicted pest abundance (output of the PBD Model) for EU regions in comparison with after invasion locations that report establishment of the pest in newly invaded areas in South Africa and Israel (Giliomee and Riedl, 1998; Hofmeyr et al., 2015).
2.4.2. EPPO PRA
The EPPO PRA led to the following conclusions.
Based on the assumption that the capacity to survive cold stresses during the winter is the key climatic factor influencing the establishment in the EPPO region and the finding that, in the South African locations where T. leucotreta is known to occur with the minimum lowest winter temperature, maximum temperatures in the winter are up to 15–17°C, a simple rule was applied to estimate EPPO regions where the climate was suitable for T. leucotreta. As a result, according to the conclusions of EPPO (2013), not only the known distribution in South Africa and the Israeli coastal plain, but also parts of the Mediterranean coast in Europe (Spain, Italy [Sicily], Malta and Cyprus) and North Africa (Morocco, Algeria and Tunisia) together with Portugal, the Canary Islands, Azores and Jordan were shown to have temperatures above the threshold. It was deemed possible that T. leucotreta can establish in a wider area in the EU because of limited knowledge of T. leucotreta cold tolerance and the fact that recent climatic data suggest that the threshold is also likely to be exceeded in southern France, e.g. Corsica, and larger areas of southern Portugal, Greece (Crete), Spain and Italy. Based on this rule, in areas further north in Europe, conditions are too cold (low minimum temperatures below 0°C, or absolute/mean minimum temperatures in January, as low as 1–3°C) and not coupled by maximum temperatures within the 15–18°C range or warmer. However, up to about 55° of latitude north, sufficient degree days above the minimum development threshold of 12°C may accrue during warmer periods for T. leucotreta to complete at least one transient generation (EPPO, 2013).
2.4.3. Previous climate suitability assessments
Although the potential establishment range of T. leucotreta was assessed with different methodologies, the exploited data were similar and limited to development + fecundity (Daiber, 1979a,b,c) and cold stress data for certain life stages (Stotter and Terblanche, 2009). The methodologies differ from each other regarding whether the suitability was estimated based on observed presence or, by the predictive interpretation of the set of experimental data as drivers of the population development over generations (see Table 10).
2.4.4. Climate matching based on the Köppen–Geiger climate classification
The climate matching approach based on the Köppen–Geiger climate classification maps areas including climate types that fulfil two conditions: (i) the organism has been found to occur in them in its endemic range, (ii) the climate type occurs in EU. Thus, if the organism occurs in a climate type that does not occur in the EU, this climate is not mapped as a relevant climate for the assessment. The Panel used the implementation of Köppen–Geiger climate classification currently available in SCANClim (EFSA and Maiorano, 2022, published by the Institute for Veterinary Public Health of the University of Vienna (Kottek et al., 2006), for the period 1986–2010, rescaled after Rubel et al. (2017) (https://koeppen-geiger.vu-wien.ac.at/present.htm).
2.4.5. Physiologically based demographic model
The climate matching with Köppen–Geiger classification was difficult to interpret because one climate zone intersected with the occurrence data on the African continent to a small extent, which would add all middle and northern EU MS to the climatically suitable area for establishment. Therefore, the EU territory was assessed using a physiologically based demographic model (PBDM; see CASAS Global https://www.casasglobal.org/). This approach models the potential establishment of the pest from the physiological response of its developmental stages to daily weather, using daily climatic variables as input. The approach does not rely on occurrence records. The technique is taken from the literature with several applications to other pests (see e.g. Gutierrez and Ponti, 2013, 2023a,b).
PBDMs are based on the notion that analogous weather‐driven submodels for resource acquisition and birth–death dynamics can be used to predict explicitly the biology and dynamics of heterotherm species across trophic levels (Gutierrez & Baumgärtner, 1984; Gutierrez, 1992, 1996; Gutierrez and Ponti, 2023b). When driven by site specific daily weather, PBDMs predict the phenology, age structure and abundance dynamics of the target species (e.g. an invasive insect herbivore) and as appropriate of interacting species in its food web (e.g. its host plant and natural enemies) across wide geographic areas (see Gutierrez et al., 2008, 2010).
The technical model documentation can be found in Appendix B of this scientific opinion. The model characteristics are summarised according to the POE protocol (Purpose, Overview, Evidence) (Grimm et al., 2020). Purpose of the PBD Model: a temperature‐driven version of the PBDM (see Gutierrez and Ponti, 2013) was implemented to exploit local differences in climate on the scale of about 0.25° × 0.25° for predicting the average annual level of the life stages of T. leucotreta in the EU, the Mediterranean Basin and Africa, using 10 consecutive years of daily maximum and minimum temperature values (years 2001–2010) as driver of daily T. leucotreta biology. Overview: The model describes the growth and survival of local pest populations (on a 0.25° × 0.25° latitude/longitude grid resulting in horizontal resolution of about 25 km) based on interpreted temperature‐dependent development, fecundity and mortality data available from the literature across all life stages of the pest (see Section 3.1). With daily resolution, the thermal biology data of the four developmental stages of T. leucotreta were summarised using biodemographic functions for development, fecundity and mortality (see Box 1 below and Appendix B). Biodemographic functions of T. leucotreta with available data are temperature‐dependent developmental rates, per capita lifetime reproductive profiles and temperature‐dependent mortality rates (see Section 3.1).
Box 1. Biodemographic functions used in the PBD model of T. leucotreta.
Oviposition profile on age x = days at 24°C times a scalar for the effects of temperature.
Stage developmental rates on temperature (delstage):
Temperature (T) dependent mortality rate:
Polynomial mortality function for all stages (μstage):
Modification for larval temperature tolerance (τ; displacement of mortality curve):
Density‐dependent predation mortality in eggs/pupae/adults (Lx):
Erlang parameters:
kegg = klarvae = kpupae = 25 (age classes of each stage, see Appendix B)
kadult = 15 (age classes of the adult stage, see Appendix B)
Mean developmental times (degree days > threshold):
eggs (86.1dd > 8.75°C)
larvae (148,8dd > 10.7°C)
pupae (185.2dd > 10.3°C)
adult females (191.8dd > 10.3°C)
Adult immigration rates (adultimmigration)
Sex ratio = 0.5
The growth and abundance of the pest at a geographic location emerge from the daily minimum and maximum temperature values (AgMERRA daily weather data; see Ruane et al., 2015) and a background density‐dependent mortality term on all stages but larvae which are inside the rose bud. Local population may go extinct during extended frost/cold periods. In the model, new introduction of adults at very low levels is immediately assumed when daily temperature, T, is above 11°C. The model is analysed in each spatial unit in discrete daily steps evaluating the life stage cohorts independently. The model output per 25 × 25 km spatial unit refers to the (average) number of pupae over 10 consecutive years of temperature data and pest population dynamics. For selected locations across the EU, the physiologically based pest abundance is shown by multi‐annual time‐series plots to understand qualitative differences on the prospective local population dynamics of the pest.
Evidence: The PBDM of T. leucotreta is adapted from an established modelling framework in support of QPRA of different pests and the modelling approach has repeatedly been validated with the occurrence or occurrence‐based models of other pests (see e.g. Gutierrez and Ponti, 2013, 2023a,b). Likewise, the model output of the T. leucotreta version was assessed with the occurrence reports of indigenous and invasive T. leucotreta in Africa and Israel. The approach could predict positive densities of T. leucotreta for the regions with pest observations, in both indigenous and invaded locations (see Appendix B).
The model was modified to explore the impact of higher thermal tolerance of the larval stage when compared to the adult stage (see Box 1, modification for larval temperature tolerance). Literature provides strong indication of higher thermal tolerance of larvae (Uys, 2014; Terblanche et al., 2017), but data are not sufficiently detailed. Therefore, model scenarios were considered using different larval thermal tolerance. The displacement of larval temperature‐dependent mortality curve by 4°C resp. 6°C facilitated the explanation of T. leucotreta occurrence records in western parts of Africa (Table 11 and Figure 11). However, the physiological aspect is of marginal relevance to predictions for EU climates, as the main constraint to T. leucotreta distribution in Europe is cold weather and the absence of dormancy in the pest (Terblanche et al., 2014).
Table 11.
Comparison of average number of pupae per year in the period 2001–2010, as projected by the PBDM model, under three scenarios of larval heat‐stress mortality being displaced by 0°C, +4°C and 6°C. Percentages in brackets compare model predictions to observations
| Continent/Country | Number of T. leucotreta observations with geographical coordinates | No of T. leucotreta observations with predicted no pupae > 1 in scenario + 0 | No of T. leucotreta observations with predicted no pupae > 1 in scenario +4°C | No of T. leucotreta observations with predicted no pupae > 1 in scenario +6°C |
|---|---|---|---|---|
| African continent | 468 | 412 (88%) | 446 (95%) | 448 (96%) |
| Southern Africa | 265 | 264 (99.6%) | 265 (100%) | 265 (100%) |
| Eastern Africa | 123 | 112 (91%) | 115 (93%) | 115 (93%) |
| Western Africa | 46 | 9 (19.6%) | 41 (89%) | 43 (93%) |
Figure 11.
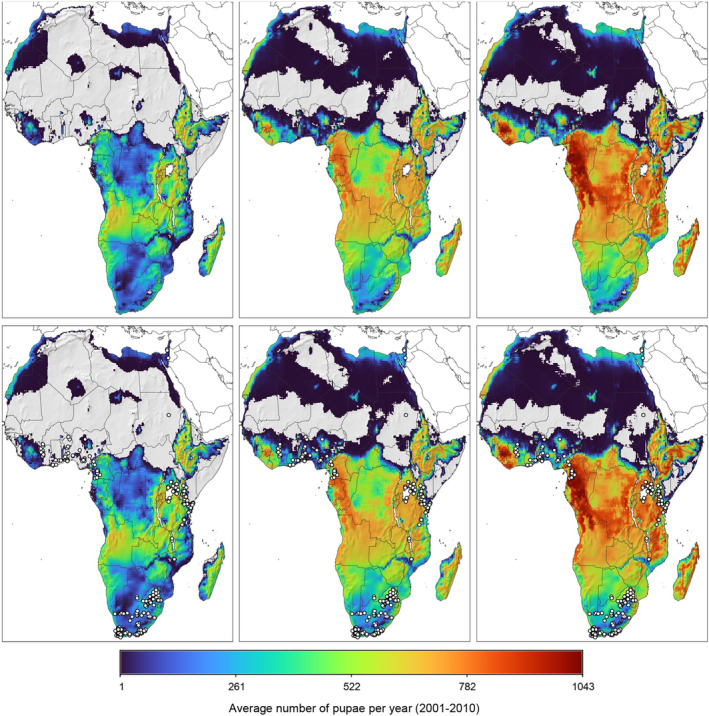
Maps of Africa with geographic distribution and relative abundance of T. leucotreta (average number of pupae during 2001–2010) as projected by the PBDM model shown for three scenarios of larval temperature‐dependent mortality displacement by 0°C (left), 4°C (middle) and 6°C (right). The maps in the bottom row additionally show T. leucotreta occurrence records (white circles with black outline). In scenario 0°C, the predicted abundance is in the range of [1, 696] (left), with 4°C according to Terblanche et al. (2017) the range is [1, 828] (middle), and with 6°C according to Uys (2014) a range of [1, 1,043] average pupae per year (right) was predicted for the whole continent. The colour legend uses the turbo colour palette (Mikhailov, 2020) to maximise graphical inference (Reda and Szafir, 2021). The absolute minimum and maximum values of the colour scaling are equal for the three maps (i.e. 1–1043) to ease comparisons. PBDM simulations are mapped and analysed using GRASS (GRASS Development Team, 2022), a multipurpose open‐source GIS (Neteler et al., 2012)
Interestingly, the predicted population at 4°C and 6°C displacement of larval mortality keeps the original predictions for Kenya and Ethiopia nearly unchanged while the prediction for the southwest coastal areas of Africa increased. In summary, the T. leucotreta model including displacement of larval temperature‐dependent mortality captures the distribution of T. leucotreta in Eastern parts of Africa (Kenya above the midpoint of the density range) and hotter regions such as Nigeria, Benin, Togo and Ghana (0.25–0.5 of the density range). In particular (and independent of the displacement scenario), the occurrences from South Africa, an invaded area of T. leucotreta, are captured in every detail of the ecological structure (Table 11 and Figure 11).
Limitations and assumptions: The biological data summarised from the literature and included in the PBDM for assessing establishment of T. leucotreta, enabled modelling temperature dependence of development, fecundity and mortality. These data, however, were generated from experiments designed for other purposes, such as developmental or quarantine studies, but not specifically addressing temperature‐dependent population dynamics as would be most suitable for PBDM modelling.
Population dynamics in the model is driven by local daily temperature data intrinsically capturing altitude structure. Other dependencies of the pest on microclimatic parameters are not yet studied and quantitative data are unavailable. Nutritional and seasonal energy budget aspects are not included due to unavailable physiologically based data.
The model assumes that hosts for T. leucotreta are available year‐round and would not limit the development of the pest. The assumption was motivated by the polyphagia of the pest and the wide distribution of hosts like tomato, maize, citrus, etc. in the areas climatically suitable for its establishment in the EU.
The predictions of the PBDM do not result from observational or studies in the field and do not include shelter from low temperature the pest may receive when inside a host subunit (e.g. fruit or pupae in soil). To average temperature‐dependent population dynamics, local populations are maintained above a low minimum computational threshold by randomly adding a fraction of adults each day to each location (0.00001 adults/day per cell = per 625 km2, equivalent to about one individual every 27 years). Hence, if local conditions favour population growth over the seasons/years, then the minimal density of stabilising (i.e. background) invasion does not have a quantitative impact on the output. True local extinction, however, cannot be inferred from the model results.
3. Assessment
The assessment of introduction of FCM into the EU via the pathway of imported cut roses has four chapters.
Pest biology, including thermal characteristics, is summarised in Section 3.1.
In Section 3.3, aspects not quantified for this QPRA are presented together with the main assumptions.
In Section 3.4, the Panel covers the entry and transfer of T. leucotreta, and discusses relevant aspects of the production, trade and use of the commodity (cut roses) that can act as a pathway.
In Section 3.5, the Panel covers the potential establishment of the pest in the EU and discusses climatic factors and host distribution.
3.1. Review of pest biology
A description of the biology of T. leucotreta is provided in the EFSA pest survey card (EFSA, 2020) with additional details in Adom and Fening (2021) and Mkiga et al. (2019). Here, we provide a summary overview with a focus on temperature‐dependent developmental biology.
Immature stages of T. leucotreta include the egg, the larva (with five instars) and the pupa. After hatching, the first larval instar seeks a suitable host fruit for development, which may last a few hours before the larva penetrates the fruit (Kelly, 1914). Penetration of the host fruit is easier and faster through lesions or scars on the fruit surface (Daiber, 1979b). Each life stage exhibits different requirements in temperature for development (Boersma, 2018). The duration of larval development shows great variability and can last from 25 days in mature sweet oranges to 173 days in immature/maturing ones. However, in most cases, the larval developmental time of T. leucotreta recorded in sweet oranges, guavas and plums lasted from 50 to 70 days (reviewed in [Daiber, 1979b]). There are no detailed published data of T. leucotreta larvae on cut roses. Therefore, developmental data on artificial diet and other host fruits are reported.
3.1.1. Larval development in artificial media
In a set of laboratory experiments, the duration of larval development in artificial media was estimated and it was found to decrease as temperature increased from 12.1°C to 30.0°C (Table 12) (Daiber, 1979b, 1989). The lower temperature threshold for development was calculated at 11.6°C and a thermal constant of 156 day‐degrees above the lower threshold is required for larval development. Shorter development times are observed in the first four larval instars, while the fifth instar before entering the prepupal stage takes significantly longer. Larval survival and development appear to be affected by the quality of the rearing medium (Daiber, 1979b).
Table 12.
Developmental duration (in days) of T. leucotreta immatures developmental stages in artificial media under different temperatures (Daiber, 1979a,b,c; Daiber, 1989)
| Temperature in °C | Egg | Larval | Pupae |
|---|---|---|---|
| 10.9 | 22.0 | ||
| 12.1 | 61.0 | 68.0 | |
| 15.0 | 45.6 | 45.0 | |
| 15.3 | 14.0 | 39.0 | |
| 17.9 | 11.0 | 21.0 | 22.0 |
| 20.0 | 7.0 | 18.8 | |
| 21.2 | 14.0 | 15.0 | |
| 23.8 | 6.0 | 12.0 | 15.0 |
| 25.0 | 11.6 | ||
| 30.0 | 4.0 | 7.0 | 10.0 |
| 35.0 | 3.0 | – | |
| 40.0 | – |
(−) = no development was observed; blank = not tested.
Unfortunately, there are limited detailed data on larval development for each larval instar across various temperatures. Using the data from Table 12, the total development time is estimated at different constant temperatures for egg to adult and for first‐instar larva to adult. Using the composition of immature population of T. leucotreta (larval instars, prepupae and pupae) in artificial medium, under three different constant temperatures (15, 20 and 25°C), as reported by Daiber (1979a,b,c), the development time for the remaining larval instars to adult was estimated at 15, 20 and 25°C by deducting the number of days from oviposition until first appearance of the relevant instar larvae (Figures 12 and 13).
Figure 12.
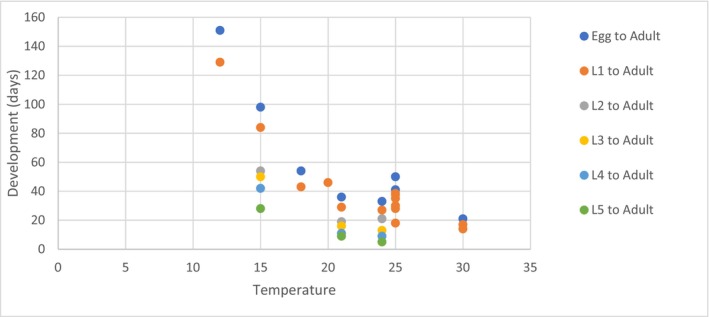
Total development time from egg to adult, L1 to adult and L2 to adult at different constant temperatures
Figure 13.
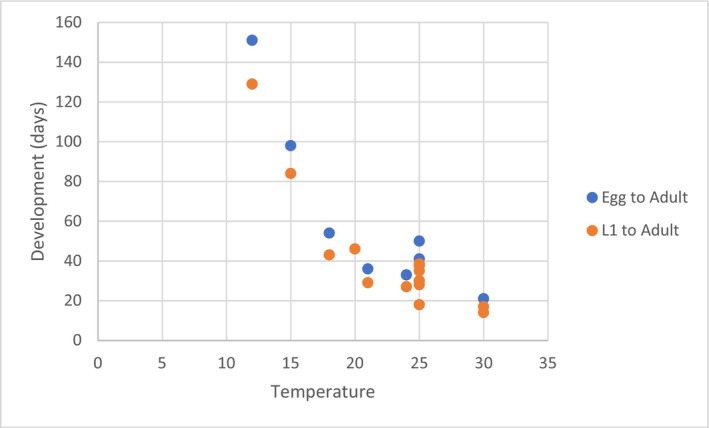
Total development time from egg to adult and from L1 to adult at different constant temperatures
3.1.2. Larval development in artificially infested fruits and vegetables
Larval development is affected by host fruit and rearing substrate. At 25.0 ± 2.0°C, larval development time was ~ 19 days on sweet oranges and 24 days on sweet pepper. Besides development time, survival is also affected by host fruit. Survival of developing larvae was estimated at 75% on sweet oranges and approximately at 63% on sweet pepper.
Larval survival and development time were also assessed in apples, pears, grapes and sweet oranges (De Jager, 2013). Total development time until adulthood was estimated at 50 days on pear and 41 days on sweet oranges (25°C). Apples proved to be an unviable host for T. leucotreta, with no fruit being penetrated, leading to no offspring.
In South Africa, natural infestation of peaches by T. leucotreta was investigated in commercial orchards and backyards of several areas (Pretoria, Rietondale, Silvertone, Hartbeespoortdam, Waterkloof). Low natural infestation (4%) of the late peach cultivar Keimoes was recorded in green and immature fruits. However, 20–30% of mature fruits, sampled during the week of harvesting, were found infested (Daiber, 1976).
Orange infestation was assessed in low, mid‐ and high‐altitude areas of Kenya and Tanzania (25 orchards in each altitude, 300 fruit per orchard). In addition, okra, sweet peppers, chilli pepper and African eggplant (300 mature vegetables per species) were sampled from vegetable fields situated within 100 m from orange orchards. Fruit loss of up to 46% for sweet orange and 12% for solanaceous vegetables was reported in Kenya and Tanzania (Mkiga et al., 2019). Odanga et al. (2018) reported losses of more than 25% for avocado fruit produced in Taita hills and Mount Kilimanjaro of Kenya and Tanzania, respectively. Also, higher percentages of infestation were recorded in high and mid altitudes compared to low ones. African eggplant, okra, sweet and chilli pepper are grown throughout the year in irrigated fields which provide a continuous availability of host plants for this pest. In Kenya, maize, which is a favourable host for T. leucotreta, is widely cultivated in areas where greenhouses with cut roses occur. According to the NPPO of Ethiopia, T. leucotreta presence in greenhouses with roses is characterised as none to medium without further details. In Kenya, KEPHIS reports T. leucotreta prevalence from low to medium‐high. T. leucotreta infestation is reported to be closely associated with crown galls affected by Agrobacterium spp. Farmers keep windows and doors open to reduce relative humidity to avoid grey mould, Botrytis, infections. However, windows often lack insect proof net, which allows entry of T. leucotreta adults in the greenhouse.
KEPHIS experts characterise T. leucotreta prevalence as high when adults trapped are > 20 without further details. KEPHIS provided trap capture data from a few farms without specifying their exact location. According to these scattered monitoring data provided by KEPHIS, T. leucotreta is persistent throughout the year in several locations where cut roses are grown both outside as well as inside the greenhouses. In these locations, outside catches were higher or comparable to those inside the greenhouse.
3.1.3. Thermal biology and population dynamics
Population dynamics of T. leucotreta life stages were predicted with the PBDM physiologically based population model (Appendix B; see e.g. Gutierrez and Ponti, 2013) that includes temperature‐dependent development (Figure 14), fecundity (Figure 15) and mortality functions (Figure 16). These biodemographic functions are fitted to experimental data (Daiber, 1979a,b,c; Terblanche et al., 2014, 2017; Sections 3.1.1 and 3.1.2 of this opinion). Experimental data for T. leucotreta from South Africa report low‐ and high‐temperature susceptibility and absence of diapause. T. leucotreta has adapted to a range of temperatures between 10°C and 35°C with the optimum near 24–25°C akin to that of a temperate species.
Figure 14.
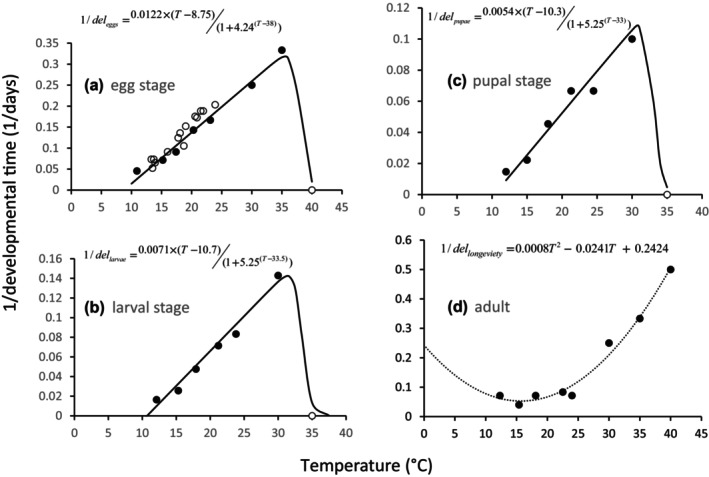
Development data applied in the model: (a) egg, (b) larval, (c) pupal and (d) adult stage. Experimental data on temperature dependence of development of the four life stages of T. leucotreta. Note that the data for adults are for adult longevity (i.e. the rate to final death) (Daiber 1979a,b,c, 1980)
Figure 15.
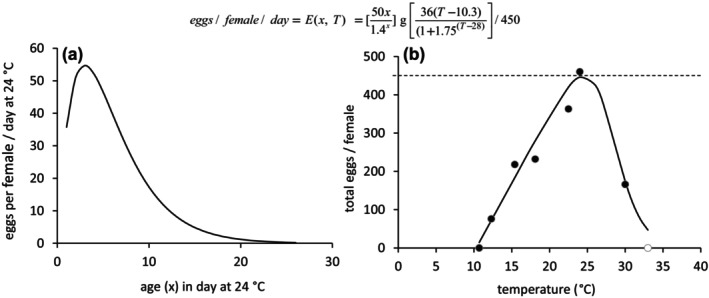
Fecundity functions used in the model. The left panel is the age specific per capita oviposition profile at 24°C. The effects of temperature on oviposition (right panel) showing a thermal threshold of 10.3°C with a maximum of 24°C and rapidly declining at higher temperatures with threshold about 33°C (Daiber (1980), see also Section 3.1 on biology)
The biodemographic functions for the development of T. leucotreta are based on data of the four life stages (egg, larva, pupa and adult) (Figure 14).
The three‐dimensional biodemographic function for fecundity used in the model combines data of the temperature‐dependent number of eggs with the age‐dependent oviposition rate profile. The product of the two functions is used to predict the oviposition rate [third dimension] at age (a) (second dimension) and temperature (T) [first dimension]. The left panel in Figure 15 is the age (a) specific oviposition profile at 24°C, while the right skewed function for total fecundity with a maximum of about 24°C in the right panel is the effect of temperature on oviposition. When normalised by dividing by maximum fecundity (i.e. 450), the latter function is used as a scalar of the age‐specific profile (the rightmost half of the function in Figure 15).
Experimental high‐temperature treatments had no significant effect on subsequent low‐temperature survival of the insect Stotter and Terblanche (2009); therefore, individual data were combined over the temperature range of the model, assuming no effect if temperature changes between high and low or vice versa over short daily time scales. The biodemographic function for temperature‐dependent daily mortality applied in the model for the T. leucotreta life stages is shown in Figure 16. It combines adult cold stress data (blue data points in Figure 16, Stotter and Terblanche, 2009), adult longevity data (brown data points in Figure 16; Daiber 1979a,b,c. see Figure 14) with larval data from Boardman et al. (2012, 2013) and Moore et al. (2022) (black data points in Figure 16).
The temperature‐related mortality is assumed the same for the egg, pupal and adult stages, but non‐systematic, survivorship data indicate the larval stage is more tolerant to high temperatures than the adult stage (Daiber, 1979a,b,c; Uys, 2014; Terblanche et al., 2017; and others). Although the thermal tolerance data by Terblanche et al. (2017) are insufficient to explicitly model the different patterns of larval mortality across temperatures, the data still suggest a 4°C displacement of the onset of increasing larval mortality versus those of adults above 24°C. With such displacement, the high‐temperature mortality of larvae starts to increase above 28°C compared to 24°C for adults. A 6°C displacement of larval mortality rates was also implemented to account for even wider differences in high‐temperature tolerance of larvae versus adults, as hinted in studies conducted by Uys (2014). In Figure 16, the 4°C displacement of larval mortality rates is indicated by the solid line. The prospective population dynamics of T. leucotreta were predicted for selected geographic locations in Europe, the Middle East and Africa (Figure 17, Table 13).
Figure 17.
Map of selected geographic locations for which T. leucotreta stage‐structured population dynamics are presented
Table 13.
List of selected geographic locations for which T. leucotreta stage‐structured population dynamics are presented in Figure 17
| Label | Longitude (degrees E) | Latitude (degrees N) | Elevation (meters) | Köppen–Geiger classification |
|---|---|---|---|---|
| The Netherlands | 4.869 | 52.229 | −5.0 | Cfb |
| Hungary 1 | 21.980 | 47.838 | 142.0 | Cfb |
| Hungary 2 | 17.358 | 47.664 | 115.0 | Cfb |
| Hungary 3 | 19.505 | 47.001 | 127.0 | Cfb |
| France | −0.561 | 44.215 | 114.0 | Cfb |
| Spain 1 | −4.242 | 43.250 | 208.0 | Cfb |
| Spain 2 | −8.833 | 42.975 | 323.0 | Csb |
| Italy | 12.454 | 41.774 | 72.0 | Csa |
| Spain 3 | −2.725 | 36.722 | 46.0 | BSh |
| Israel | 34.880 | 32.164 | 67.0 | Csa |
| Kenya 1 | 34.803 | 1.035 | 2017.0 | Cfb |
| Kenya 2 | 35.341 | 0.398 | 2186.0 | Cfb |
| Kenya 3 | 37.650 | 0.050 | 1596.0 | Csb |
| Kenya 4 | 35.549 | −0.236 | 2162.0 | Cfb |
| Kenya 5 | 36.761 | −1.238 | 1747.0 | Cfb |
The resulting time series show qualitative differences in the stage‐structured population profile over time depending on the temperature characteristic of the location (Figure 18) using the same low initial values in the model. Note that the populations in Kenya grow to high levels but oscillate with changes in seasonal temperatures (Figure 18k–o). In highly unfavourable areas (Figure 18a–d), populations remain low due to random inputs in the model, and in intermediately favourable areas (coastal Israel, Figure 18j), the effects of seasonal cooler weather cause deep oscillations in the dynamics.
Figure 18.
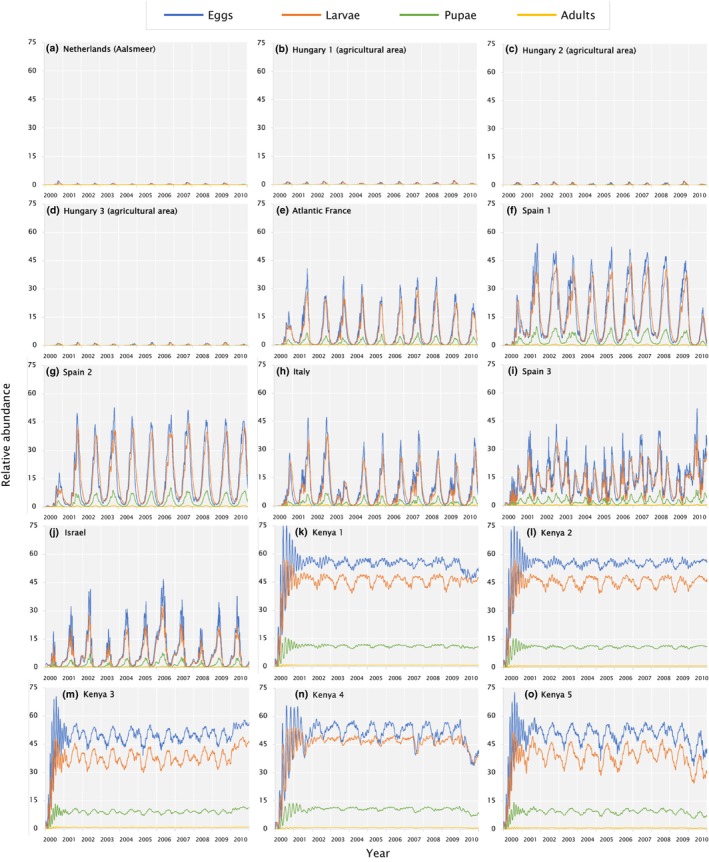
Population dynamics of T. leucotreta eggs (blue), larvae (orange), pupae (green) and adults (yellow) at selected locations for years 2000–2010. Plots in subfigures (a) to (o) are ordered so that the associated locations fall on a North to South gradient (see map of locations in Figure 17)
The predicted maps of establishment across EU MS, south‐eastern Asia and Africa (see Section 3.5.2) are generated by averaging the population time series (Figure 18) over 10 years (2001–2010) as the mean abundance per cell per life stage of the insect. Only the pupal stage is shown in those figures (Section 3.5.2). Population dynamics for the first year of PBDM simulation (2000) are included in each plot of Figure 18 to show how the model equilibrates to local weather conditions but are not used for computing summary statistics (i.e. average pupae) shown on maps in Section 3.5.2. PBDM simulations are mapped and analysed using GRASS (GRASS Development Team, 2022), a multi‐purpose open‐source GIS (Neteler et al., 2012).
3.2. Pest introduction
Pest introduction is the combination of entry (which includes transfer to hosts) and establishment.
In Section 3.3, the aspects not quantified in this opinion and the main assumptions are presented.
In Section 3.4, the Panel covers the entry and transfer of T. leucotreta, and discusses relevant aspects of the production, trade and use of the commodity (cut roses) that can act as a pathway.
Climatic factors and host distribution in the EU which can affect potential for establishment across are covered in Section 3.5.
3.3. Aspects not quantified and assumptions
Although a small but consistent percentage of cut flowers including cut roses is used for funerals and in cemeteries, the percentage varies among countries (5–13%, Rabobank, 2022) (see further below). This use is not specifically assessed in this scientific opinion, but it is covered by the different scenarios of the pathway model.
In line with EPPO (2013), it is assumed that T. leucotreta, being extremely polyphagous, will likely find suitable hosts for establishment outdoors in the climatically suitable areas.
Scenarios of possible changes in trade patterns or climate are not considered in this assessment.
Most common rose varieties are ‘standard roses’ developing one bloom per stem. There are, however, ‘spray rose’ varieties which develop multiple blooms per stem (typically three to seven flowers per stem). The pathway model assumes that one infested stem corresponds to one T. leucotreta insect. This is reasoned by the low level of infestation and the egg laying behaviour of T. leucotreta; however, spray rose varieties with multiple blooms per stem may not meet this assumption.
3.4. Entry into the EU where establishment is possible (NUTS2 resolution)
The Panel modelled the entry of T. leucotreta estimating the infestation level in cut roses imported into the EU, the trade flow and distribution into EU NUTS2 regions and the escape of T. leucotreta adults from the waste before and after waste treatment in the EU NUTS2 regions presenting areas suitable for the establishment of T. leucotreta (for methodology, see Section 2.3). NUTS2 regions were considered in the model when having at least one grid cell with at least 1 pupa per year (see Table 14 for the definition of the climate suitability classes and Figure 19).
Table 14.
Definition of climate suitability classes in the pathway model
| No. pupae/year per NUTS2 area | Climate suitability class | Interpretation | Colour |
|---|---|---|---|
| 0 | 0 | Not suitable | |
| 1 ≤ × < 128 | 1 |
Low climate suitability
High climate suitability |
|
| 128 ≤ × < 255 | 2 | ||
| 255 ≤ × < 382 | 3 | ||
| 382 ≤ × < 509 | 4 |

Figure 19.

NUTS 2 regions in EU and some neighbouring countries, and their climate suitability classes according to the potential establishment of T. leucotreta based on the average number of pupae/year as estimated by the PBDM model. 0 = less than 1 pupae/year, 1 = between 1 and 128 pupae/year, 2 = between 128 and 255 pupae/year, 3 = between 255 and 382 pupae/year and 4 = between 382 and 509 pupae/year. Note: Kosovo and Bosnia and Herzegovina are not covered in the layer of NUTS 2 boundaries by Eurostat (year 2021; https://ec.europa.eu/eurostat/web/gisco/geodata/reference-data/administrative-units-statistical-units/nuts)
Considering the data included in the assessment, the regular escape of T. leucotreta adult insects in the territory of the EU is predicted as:
Average number of escaped adults per year per each NUTS2 (presented in Section 3.4.1);
Average number of escaped adults per year/(3.14 Km2 × 10 days) in residential areas of each NUTS2 region (presented in Section 3.4.3 and in Appendix A Section A.1.4).
3.4.1. Results of the entry pathway model at NUTS2 level
Based on the outputs of the entry model, the median number of adults escaping from cut roses in all the climatically suitable areas of the EU is estimated to be 49,867 per year (90% uncertainty between 5,298 and 234,393) for the scenario of timing until waste treatment of 3 days and up to 143,689 per year (90% uncertainty between 21,126 and 401,458) for 28 days timing until waste treatment under the worst‐case scenario (see Table A.1 in Appendix A). Results of the median number of escapes per year for the different climatically suitable NUTS2 regions are shown in log‐scale in Figure 20 for the four considered scenarios of timing until waste treatment.
Table A.1.
Number of Thaumatotibia leucotreta adults escaping within a year from cut roses imported from Africa (including Israel) calculated by a pathway model for NUTS2 regions in the climatically suitable area of Europe. Results are stratified by four scenarios of timing from disposal at the consumer until the waste treatment, and given as median and 90% certainty interval (from P5 to P95)
| Country | NUTS code | Label | Climate suitability class | Annual escape (3 days) | Annual escape (7 days) | Annual escape (14 days) | Annual escape (28 days) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (Adults/NUTS2) | (Adults/NUTS2) | (Adults/NUTS2) | (Adults/NUTS2) | ||||||||||||
| P5 | Median | P95 | P5 | Median | P95 | P5 | Median | P95 | P5 | Median | P95 | ||||
| Total | Countries in the climatically suitable area | 5,298 | 49,867 | 234,393 | 6,059 | 52,805 | 238,832 | 9,431 | 69,501 | 264,975 | 21,126 | 143,689 | 401,458 | ||
| Spain & Portugal | ESPT | Spain and Portugal | 261 | 2,794 | 16,149 | 313 | 2,994 | 16,444 | 569 | 4,408 | 18,442 | 1,284 | 8,883 | 27,212 | |
| ES11 | Galicia | 1 | 12 | 137 | 794 | 14 | 142 | 801 | 21 | 173 | 844 | 47 | 333 | 1,107 | |
| ES12 | Principado de Asturias | 1 | 5 | 52 | 299 | 5 | 54 | 302 | 8 | 65 | 318 | 18 | 126 | 418 | |
| ES13 | Cantabria | 1 | 3 | 29 | 171 | 3 | 31 | 173 | 4 | 37 | 182 | 10 | 72 | 239 | |
| ES21 | País Vasco | 1 | 10 | 111 | 643 | 11 | 115 | 649 | 17 | 140 | 683 | 38 | 270 | 897 | |
| ES22 | Comunidad Foral de Navarra | 1 | 3 | 33 | 193 | 3 | 35 | 195 | 5 | 42 | 205 | 11 | 81 | 269 | |
| ES23 | La Rioja | 1 | 1 | 16 | 93 | 2 | 17 | 94 | 2 | 20 | 99 | 5 | 39 | 129 | |
| ES24 | Aragón | 1 | 6 | 67 | 391 | 7 | 70 | 394 | 10 | 85 | 415 | 23 | 164 | 545 | |
| ES30 | Comunidad de Madrid | 1 | 31 | 342 | 1,982 | 35 | 355 | 2,000 | 52 | 432 | 2,106 | 117 | 832 | 2,765 | |
| ES41 | Castilla y León | 1 | 11 | 122 | 705 | 12 | 126 | 712 | 18 | 154 | 750 | 42 | 296 | 984 | |
| ES42 | Castilla‐la Mancha | 2 | 10 | 104 | 602 | 11 | 110 | 610 | 19 | 154 | 669 | 43 | 300 | 937 | |
| ES43 | Extremadura | 2 | 5 | 54 | 312 | 6 | 57 | 317 | 10 | 80 | 347 | 22 | 156 | 487 | |
| ES51 | Cataluña | 2 | 36 | 389 | 2,250 | 43 | 412 | 2,283 | 72 | 575 | 2,502 | 162 | 1,121 | 3,506 | |
| ES52 | Comunitat Valenciana | 3 | 24 | 257 | 1,481 | 31 | 283 | 1,517 | 65 | 481 | 1,819 | 143 | 989 | 2,877 | |
| ES53 | Illes Balears | 2 | 6 | 62 | 356 | 7 | 65 | 361 | 11 | 91 | 396 | 26 | 177 | 555 | |
| ES61 | Andalucía | 4 | 42 | 435 | 2,498 | 54 | 485 | 2,574 | 116 | 845 | 3,131 | 269 | 1,848 | 5,440 | |
| ES62 | Región de Murcia | 3 | 7 | 77 | 443 | 9 | 85 | 454 | 19 | 144 | 544 | 43 | 296 | 861 | |
| ES63 | Ciudad de Ceuta | 3 | 0 | 4 | 25 | 1 | 5 | 25 | 1 | 8 | 30 | 2 | 17 | 48 | |
| ES64 | Ciudad de Melilla | 2 | 0 | 4 | 25 | 0 | 5 | 25 | 1 | 6 | 28 | 2 | 12 | 39 | |
| PT11 | Norte | 2 | 17 | 182 | 1,051 | 20 | 193 | 1,067 | 34 | 269 | 1,169 | 76 | 524 | 1,638 | |
| PT15 | Algarve | 3 | 2 | 22 | 129 | 3 | 25 | 132 | 6 | 42 | 158 | 12 | 86 | 251 | |
| PT16 | Centro (PT) | 3 | 11 | 113 | 653 | 14 | 125 | 669 | 29 | 212 | 802 | 63 | 436 | 1,268 | |
| PT17 | Área Metropolitana de Lisboa | 3 | 14 | 146 | 843 | 18 | 161 | 863 | 37 | 273 | 1,035 | 81 | 563 | 1,637 | |
| PT18 | Alentejo | 3 | 3 | 36 | 207 | 4 | 40 | 212 | 9 | 67 | 255 | 20 | 139 | 403 | |
| France | FR | France | 2,302 | 21,884 | 104,347 | 2,581 | 23,070 | 105,876 | 3,865 | 29,054 | 114,867 | 8,831 | 60,342 | 171,723 | |
| FRG0 | Pays‐de‐la‐Loire | 1 | 288 | 2,736 | 13,048 | 323 | 2,885 | 13,239 | 483 | 3,633 | 14,363 | 1,104 | 7,545 | 21,473 | |
| FRH0 | Bretagne | 1 | 253 | 2,407 | 11,476 | 284 | 2,537 | 11,645 | 425 | 3,195 | 12,633 | 971 | 6,637 | 18,887 | |
| FRI1 | Aquitaine | 1 | 262 | 2,493 | 11,887 | 294 | 2,628 | 12,061 | 440 | 3,310 | 13,085 | 1,006 | 6,874 | 19,562 | |
| FRI3 | Poitou‐Charentes | 1 | 137 | 1,300 | 6,197 | 153 | 1,370 | 6,288 | 230 | 1,726 | 6,822 | 525 | 3,584 | 10,199 | |
| FRJ1 | Languedoc‐Roussillon | 1 | 216 | 2,053 | 9,789 | 242 | 2,164 | 9,933 | 363 | 2,726 | 10,776 | 829 | 5,661 | 16,110 | |
| FRJ2 | Midi‐Pyrénées | 1 | 233 | 2,212 | 10,549 | 261 | 2,332 | 10,703 | 391 | 2,937 | 11,612 | 893 | 6,100 | 17,360 | |
| FRK2 | Rhône‐Alpes | 1 | 505 | 4,796 | 22,868 | 566 | 5,056 | 23,203 | 847 | 6,367 | 25,174 | 1,935 | 13,224 | 37,634 | |
| FRL0 | Provence‐Alpes‐Côte d'Azur | 1 | 383 | 3,639 | 17,351 | 429 | 3,836 | 17,605 | 643 | 4,831 | 19,100 | 1,468 | 10,033 | 28,554 | |
| FRM0 | Corse | 1 | 26 | 248 | 1,182 | 29 | 261 | 1,199 | 44 | 329 | 1,301 | 100 | 683 | 1,945 | |
| Italy & Malta | ITMT | Italy and Malta | 1,854 | 18,232 | 86,462 | 2,164 | 19,409 | 88,284 | 3,603 | 26,401 | 98,639 | 7,823 | 54,231 | 150,632 | |
| ITC1 | Piemonte | 1 | 132 | 1,316 | 6,258 | 150 | 1,380 | 6,360 | 223 | 1,706 | 6,838 | 494 | 3,463 | 9,918 | |
| ITC3 | Liguria | 1 | 47 | 465 | 2,213 | 53 | 488 | 2,249 | 79 | 603 | 2,418 | 175 | 1,225 | 3,508 | |
| ITC4 | Lombardia | 1 | 308 | 3,060 | 14,556 | 349 | 3,209 | 14,793 | 519 | 3,968 | 15,904 | 1,150 | 8,055 | 23,070 | |
| ITF1 | Abruzzo | 1 | 40 | 395 | 1,878 | 45 | 414 | 1,909 | 67 | 512 | 2,052 | 148 | 1,039 | 2,977 | |
| ITF2 | Molise | 1 | 9 | 92 | 436 | 10 | 96 | 443 | 16 | 119 | 477 | 34 | 241 | 691 | |
| ITF3 | Campania | 2 | 180 | 1,754 | 8,308 | 213 | 1,883 | 8,502 | 369 | 2,697 | 9,716 | 806 | 5,539 | 15,180 | |
| ITF4 | Puglia | 2 | 125 | 1,214 | 5,750 | 148 | 1,303 | 5,884 | 256 | 1,866 | 6,724 | 558 | 3,833 | 10,506 | |
| ITF5 | Basilicata | 1 | 17 | 169 | 803 | 19 | 177 | 816 | 29 | 219 | 877 | 63 | 444 | 1,273 | |
| ITF6 | Calabria | 2 | 60 | 582 | 2,755 | 71 | 624 | 2,819 | 122 | 894 | 3,222 | 267 | 1,837 | 5,034 | |
| ITG1 | Sicilia | 3 | 160 | 1,510 | 7,108 | 203 | 1,686 | 7,352 | 429 | 2,981 | 9,431 | 952 | 6,393 | 16,924 | |
| ITG2 | Sardegna | 2 | 51 | 495 | 2,344 | 60 | 531 | 2,399 | 104 | 761 | 2,741 | 227 | 1,563 | 4,283 | |
| ITH2 | Provincia Autonoma di Trento | 1 | 17 | 166 | 792 | 19 | 175 | 805 | 28 | 216 | 865 | 63 | 438 | 1,255 | |
| ITH3 | Veneto | 1 | 150 | 1,489 | 7,082 | 170 | 1,561 | 7,198 | 253 | 1,931 | 7,738 | 559 | 3,919 | 11,225 | |
| ITH4 | Friuli‐Venezia Giulia | 1 | 37 | 368 | 1,751 | 42 | 386 | 1,779 | 62 | 477 | 1,913 | 138 | 969 | 2,775 | |
| ITH5 | Emilia‐Romagna | 1 | 137 | 1,362 | 6,480 | 155 | 1,428 | 6,585 | 231 | 1,766 | 7,080 | 512 | 3,586 | 10,270 | |
| ITI1 | Toscana | 2 | 117 | 1,134 | 5,371 | 138 | 1,217 | 5,496 | 239 | 1,743 | 6,281 | 521 | 3,581 | 9,813 | |
| ITI2 | Umbria | 1 | 27 | 266 | 1,263 | 30 | 278 | 1,284 | 45 | 344 | 1,380 | 100 | 699 | 2,002 | |
| ITI3 | Marche | 1 | 46 | 462 | 2,196 | 53 | 484 | 2,231 | 78 | 599 | 2,399 | 173 | 1,215 | 3,480 | |
| ITI4 | Lazio | 2 | 182 | 1,768 | 8,371 | 215 | 1,897 | 8,567 | 372 | 2,717 | 9,790 | 812 | 5,581 | 15,296 | |
| MT00 | Malta | 2 | 17 | 159 | 750 | 20 | 170 | 767 | 34 | 244 | 876 | 73 | 500 | 1,368 |
| Slovenia | SI | Slovenia | 43 | 437 | 2,081 | 50 | 460 | 2,116 | 75 | 584 | 2,305 | 175 | 1,214 | 3,456 | |
| SI03 | Vzhodna Slovenija | 1 | 23 | 230 | 1,092 | 26 | 242 | 1,110 | 39 | 307 | 1,210 | 92 | 637 | 1,814 | |
| SI04 | Zahodna Slovenija | 1 | 21 | 208 | 989 | 24 | 219 | 1,005 | 36 | 278 | 1,095 | 83 | 577 | 1,642 | |
| Croatia | HR | Croatia | 80 | 857 | 4,351 | 91 | 907 | 4,424 | 141 | 1,171 | 4,871 | 315 | 2,414 | 7,855 | |
| HR02 | Pannonian Croatia | 1 | 21 | 226 | 1,148 | 24 | 239 | 1,167 | 37 | 309 | 1,285 | 83 | 637 | 2,072 | |
| HR03 | Adriatic Croatia | 1 | 27 | 287 | 1,459 | 31 | 304 | 1,484 | 47 | 393 | 1,634 | 106 | 810 | 2,634 | |
| HR05 | City of Zagreb | 1 | 16 | 170 | 862 | 18 | 180 | 876 | 28 | 232 | 965 | 62 | 478 | 1,555 | |
| HR06 | Northern Croatia | 1 | 16 | 174 | 883 | 19 | 184 | 898 | 29 | 238 | 989 | 64 | 490 | 1,594 | |
| Greece & Cyprus | Greece and Cyprus | 280 | 1,517 | 6,686 | 307 | 1,625 | 6,873 | 439 | 2,382 | 8,165 | 794 | 4,673 | 13,210 | ||
| EL30 | Attiki | 3 | 104 | 506 | 2,175 | 116 | 555 | 2,257 | 176 | 916 | 2,891 | 319 | 1,831 | 5,035 | |
| EL41 | Voreio Aigaio | 2 | 5 | 30 | 132 | 6 | 32 | 135 | 8 | 44 | 155 | 14 | 84 | 242 | |
| EL42 | Notio Aigaio | 2 | 8 | 45 | 200 | 9 | 48 | 204 | 12 | 66 | 235 | 21 | 128 | 367 | |
| EL43 | Kriti | 3 | 18 | 86 | 370 | 20 | 94 | 384 | 30 | 156 | 492 | 54 | 312 | 857 | |
| EL51 | Anatoliki Makedonia, Thraki | 1 | 12 | 75 | 342 | 13 | 78 | 347 | 16 | 96 | 377 | 29 | 184 | 558 | |
| EL52 | Kentriki Makedonia | 1 | 36 | 234 | 1,070 | 39 | 245 | 1,084 | 50 | 300 | 1,178 | 92 | 575 | 1,744 | |
| EL53 | Dytiki Makedonia | 1 | 5 | 33 | 151 | 6 | 35 | 153 | 7 | 42 | 166 | 13 | 81 | 247 | |
| EL54 | Ipeiros | 2 | 8 | 43 | 192 | 8 | 46 | 196 | 11 | 63 | 226 | 21 | 122 | 352 | |
| EL61 | Thessalia | 1 | 14 | 89 | 409 | 15 | 93 | 414 | 19 | 115 | 450 | 35 | 220 | 666 | |
| EL62 | Ionia Nisia | 2 | 5 | 26 | 117 | 5 | 28 | 119 | 7 | 39 | 138 | 13 | 75 | 214 | |
| EL63 | Dytiki Ellada | 2 | 15 | 84 | 374 | 16 | 89 | 383 | 22 | 124 | 441 | 40 | 239 | 687 | |
| EL64 | Sterea Ellada | 2 | 13 | 72 | 320 | 14 | 76 | 327 | 19 | 106 | 376 | 34 | 204 | 587 | |
| EL65 | Peloponnisos | 3 | 16 | 77 | 333 | 18 | 85 | 345 | 27 | 140 | 442 | 49 | 280 | 770 | |
| CY00 | Kypros | 2 | 20 | 115 | 511 | 22 | 122 | 522 | 31 | 169 | 601 | 55 | 326 | 937 | |
| Hungary | HU | Hungary | 88 | 893 | 4,286 | 100 | 938 | 4,361 | 156 | 1,202 | 4,746 | 368 | 2,542 | 7,298 | |
| HU12 | Pest | 1 | 19 | 194 | 932 | 22 | 204 | 948 | 34 | 261 | 1,032 | 80 | 553 | 1,587 | |
| HU23 | Dél‐Dunántúl | 1 | 13 | 131 | 629 | 15 | 138 | 640 | 23 | 176 | 696 | 54 | 373 | 1,070 | |
| HU31 | Észak‐Magyarország | 1 | 16 | 167 | 804 | 19 | 176 | 818 | 29 | 225 | 890 | 69 | 477 | 1,369 | |
| HU32 | Észak‐Alföld | 1 | 21 | 216 | 1,037 | 24 | 227 | 1,055 | 38 | 291 | 1,148 | 89 | 615 | 1,765 | |
| HU33 | Dél‐Alföld | 1 | 18 | 184 | 885 | 21 | 194 | 900 | 32 | 248 | 980 | 76 | 525 | 1,506 |
| Bulgaria | BG | Bulgaria | 55 | 421 | 2,020 | 59 | 442 | 2,058 | 83 | 559 | 2,263 | 165 | 1,123 | 3,619 | |
| BG31 | Severozapaden | 1 | 6 | 44 | 212 | 6 | 46 | 216 | 9 | 59 | 237 | 17 | 118 | 379 | |
| BG32 | Severen tsentralen | 1 | 6 | 47 | 225 | 7 | 49 | 229 | 9 | 62 | 252 | 18 | 125 | 403 | |
| BG33 | Severoiztochen | 1 | 7 | 56 | 269 | 8 | 59 | 274 | 11 | 74 | 301 | 22 | 149 | 481 | |
| BG34 | Yugoiztochen | 1 | 8 | 62 | 298 | 9 | 65 | 303 | 12 | 82 | 333 | 24 | 165 | 533 | |
| BG41 | Yugozapaden | 1 | 16 | 127 | 609 | 18 | 133 | 620 | 25 | 168 | 682 | 50 | 338 | 1,090 | |
| BG42 | Yuzhen tsentralen | 1 | 11 | 85 | 409 | 12 | 89 | 416 | 17 | 113 | 458 | 33 | 227 | 732 | |
| Romania | RO | Romania | 217 | 2,212 | 11,673 | 248 | 2,350 | 11,938 | 366 | 3,071 | 13,232 | 766 | 6,265 | 22,162 | |
| RO11 | Nord‐Vest | 1 | 40 | 408 | 2,150 | 46 | 433 | 2,199 | 67 | 566 | 2,437 | 141 | 1,154 | 4,082 | |
| RO22 | Sud‐Est | 1 | 37 | 380 | 2,006 | 43 | 404 | 2,052 | 63 | 528 | 2,274 | 132 | 1,077 | 3,809 | |
| RO31 | Sud ‐ Muntenia | 1 | 46 | 464 | 2,449 | 52 | 493 | 2,504 | 77 | 644 | 2,776 | 161 | 1,314 | 4,649 | |
| RO32 | Bucuresti ‐ Ilfov | 1 | 37 | 371 | 1,960 | 42 | 395 | 2,004 | 61 | 516 | 2,222 | 129 | 1,052 | 3,721 | |
| RO41 | Sud‐Vest Oltenia | 1 | 30 | 306 | 1,613 | 34 | 325 | 1,649 | 50 | 424 | 1,828 | 106 | 865 | 3,061 | |
| RO42 | Vest | 1 | 28 | 283 | 1,495 | 32 | 301 | 1,529 | 47 | 393 | 1,695 | 98 | 802 | 2,839 |
Figure 20.
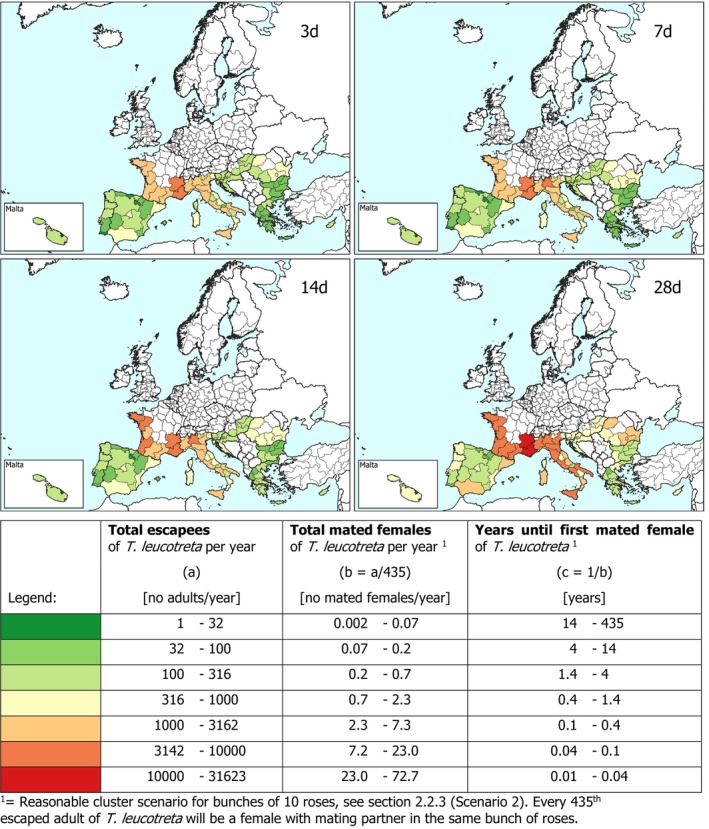
Maps of the European NUTS2 regions that are climatically suitable for T. leucotreta establishment showing the results of the pathway model for T. leucotreta in cut roses expressed in total number (log‐scale) of T. leucotreta adults predicted to escape from the cut roses disposed per year and each NUTS2 region, under four different scenarios of timing from the initial disposal until the waste treatment of 3, 7, 14 and 28 days
It has to be noted, however, that the differences across the scenarios are entirely due to the escapees from the regional waste management processes. In fact, considering that private composting is assumed not having a time gap between the initial disposal at household and the treatment, the escapees from the private compost remain constant across all scenarios. The detailed results (median) and 90% certainty intervals (P5 to P95) for each NUTS2 region in the climatically suitable NUTS2 regions of the EU are presented in Appendix A Section A.1 for the four considered scenarios of timing until waste treatment.
In NUTS2 regions with higher climate suitability (i.e. within the climate suitability classes 3 and 4 of Table 14), and for a timing until waste treatment of 3 days, the median number of adults of T. leucotreta per year escaping from the cut roses varied from 1,510 for Sicily (Italy) to 506 for Attica (Greece), to 435 in Andalusia (southern Spain) and 257 in the Region of Valencia (eastern Spain) (see Table A.1 in Appendix A).
In some of the NUTS2 regions with lower climate suitability class (i.e. within the climate suitability class 1, see e.g. the blue areas in France in Figure 19), the estimated median number of adults escaping from cut roses was larger than in other more climatically suitable areas (e.g. Spain) (Figure 20), due to the large number of cut roses from Africa consumed in the region. This was the case for the Rhône‐Alpes region (France) where the estimated median number of adults escaping from cut roses was 4,796 per year (Figure 20 and Table A.1 in Appendix A).
When the period until waste treatment becomes longer, the number of escapees increases, and this could become relevant in NUTS2 regions with higher climate suitability class. For example, for Andalusia (Spain), the median number of escapees per year increases from 435 (90% uncertainty between 42 and 2,498) to 1,848 (90% uncertainty between 269 and 5,440) when considering a time from initial disposal until waste treatment of 3 and 28 days, respectively. Similarly, in Sicily (Italy), the estimated median number of adults escaping from cut roses per year increased from 1,510 (90% uncertainty between 160 and 7,108) to 6,393 (90% uncertainty between 952 and 16,924) from scenario 3 to 28 days. Again, for Attica region (Greece), the estimated median number of escapees raised from 506 (90% uncertainty between 104 and 2,175) to 1,831 (90% uncertainty between 319 and 5,035) from scenario 3 to 28 days, respectively (Figure 20 and Table A.1 in Appendix A).
It should be noted that the period from the initial disposal at household level until the waste treatment remains a substantial source of uncertainty both within and between NUTS2 regions. For this reason, the Panel decided to follow a scenarios approach, ranging from the best‐case scenario of 3 days timespan between disposal and treatment to the worst‐case scenario of 28 days. The relative frequency of occurrence of the four scenarios in the real settings is, however, not known. The scenarios are expected to apply to individual communities or waste collection systems in a particular NUTS2, but the Panel does not expect that such scenarios will apply simultaneously in all NUTS2 in the same year.
Figure 20 also shows the predicted median number of mated females per year escaping from cut roses in the climatically suitable areas of the EU, under the realistic scenario that, on average, for every 435 escaping T. leucotreta adults a mating will occur (see Section 2.2.3). Under such scenario, the total estimated median number of T. leucotreta mated females per year from imported cut roses in all the climatically suitable NUTS2 regions of the EU would vary from 115 (90% uncertainty between 12 and 538) up to 330 (90% uncertainty between 49 and 923) for the 3‐ and 28‐day scenarios, respectively (see Appendix A Table A.1.2 and Appendix D).
The number of years until the occurrence of the first mated female of T. leucotreta from imported infested cut roses in a climatically suitable NUTS2 region, varies from less than one up to few hundreds, depending on the NUTS2 region and on the scenario of timing from disposal until treatment (Figure 20). As observed above, when commenting the median number of T. leucotreta adults escape per NUTS2, many of these mated females will occur in NUTS2 regions only marginally suitable for the insect's establishment. However, under all the four scenarios of 3, 7, 14 and 28 days of timing until treatments, there are NUTS2 regions in the coastal areas of the Mediterranean and of the Iberian Peninsula where a mated female is predicted to result in less than 1 year.
Due to the polyphagous nature of T. leucotreta, host availability will not be a limiting factor (see also Section 3.4.5), while the likelihood for establishment may be limited by the climatic conditions (esp. winter cold) and other factors (e.g. natural enemies). It also has to be considered that the likelihood of mating will decrease with decreasing size of the flower bunch, but it will increase when several infested bunches would simultaneously occur during a suitable period (e.g. in the first 10 days of the female adult stage when the majority of eggs are laid [Daiber, 1980; Mkiga et al., 2019; EFSA, 2020]) within a suitable area (i.e. a radius of 1 km, flying radius of T. leucotreta males responding to females [EFSA, 2020]) (see also Sections 3.4.3 and 3.4.4).
3.4.2. Seasonality of predicted T. leucotreta escapes and matings from infested cut roses
The escape of adults of T. leucotreta is more likely, when the time between the initial disposal at the consumer household and the waste treatment is prolonged, especially 14 or 28 days. However, the effect is more pronounced when the temperatures induce faster development, especially in summer. In the 28‐day scenario, the longer duration is sufficient to increase the number of escapes also in spring and autumn. To support the interpretation, a reasonable clustering scenario is assumed (see Section 2.3.3), namely the escape of females T. leucotreta with possible mating partner in a bunch of 10 roses. This is shown at EU level for all the climatically suitable NUTS2 regions in Table 15 and in Figure 21.
Table 15.
Escape of T. leucotreta females with possible mating partner (median with 90% uncertainty range [P5–P95]) in a bunch of 10 cut roses (realistic scenario) in the EU climatically suitable areas during an average season over 10 years, for four scenarios on the timing between the initial disposal and waste treatment: 3, 7, 14 or 28 days
| Duration between initial disposal and waste treatment | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 days | 7 days | 14 days | 28 days | |||||||||
| Season | Med | P5 | P95 | Med | P5 | P95 | Med | P5 | P95 | Med | P5 | P95 |
| Winter | 31 | 3 | 148 | 31 | 3 | 148 | 31 | 3 | 148 | 31 | 3 | 148 |
| Spring | 32 | 3 | 153 | 32 | 3 | 153 | 34 | 4 | 155 | 43 | 6 | 170 |
| Summer | 24 | 3 | 112 | 30 | 4 | 121 | 61 | 9 | 177 | 185 | 28 | 480 |
| Autumn | 27 | 3 | 129 | 28 | 3 | 130 | 32 | 4 | 136 | 56 | 8 | 177 |
Figure 21.
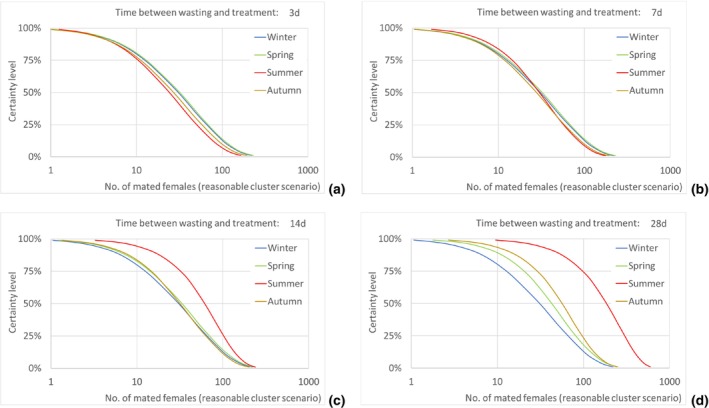
Escape of T. leucotreta females with possible mating partner in a bunch of 10 cut roses (realistic clustering scenario) during an average season over 10 years (blue line = winter, green = spring, red = summer and yellow = autumn) in the climatically suitable NUTS2 regions in the EU with certainty curves for different scenarios between initial wasting and waste treatment: (a) 3 days, (b) 7 days, (c) 14 days and (d) 28 days
Considering the results at the EU level, the highest estimated number of adults escapes and of mated females is predicted during the summer (Figure 21, Table 15). This is generally observed also in individual NUTS2 regions (see Appendix A: e.g. Rhone Alpes (FR), Andalusia (ES) and Sicily (IT)).
However, when considering individual NUTS2 regions, in specific regions, higher escape values may occur also in autumn, especially in the 28 days scenario (e.g. in Sicily (Figure 22) or in Andalusia [Appendix A Figure A.3]).
Figure 22.
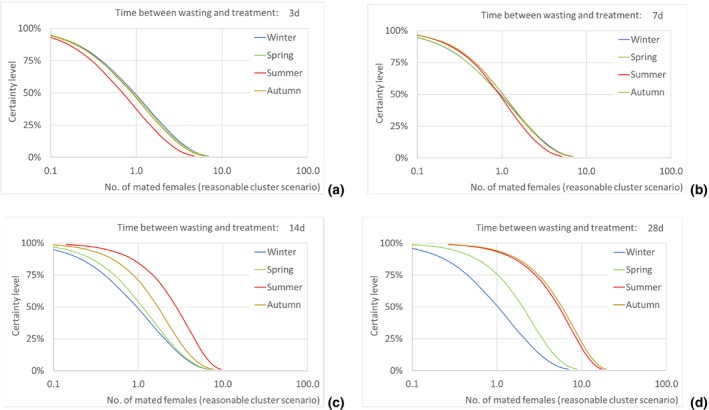
Escape of T. leucotreta females with possible mating partner in a bunch of 10 cut roses (realistic clustering scenario) during an average season over 10 years (blue line = winter, green = spring, red = summer and yellow = autumn) in Sicily (Italy) with certainty curves for different scenarios between initial wasting and waste treatment: (a) 3 days, (b) 7 days, (c) 14 days and (d) 28 days
3.4.3. Escape per circle of 1 km radius in the residential areas of the NUTS2 regions within 10 days (scenario 3)
The scenario 3 (see Section 2.2.3 and Appendix A Section A.1.4) assumes the complete absence of clustering of the infestation in the cut roses and the uniform distribution of disposed cut roses in the residential areas of each NUTS2 region. Under this scenario, on average more than one mated female (for the 95th percentile) is expected in summer under the 28 days waste treatment scenario in few NUTS 2 regions such as Corsica (France), Liguria and Sicily (Italy) and Malta. It should, however, be noted that these standardised values represent a uniform spatial and temporal distribution of insect escapes, but that such uniform distributions would be extremely unlikely to occur in reality. Hence, the details are reported in the Appendix A (Section A.1.4).
3.4.4. Spatial and temporal clustering of cut roses consumption
The Panel notes that an increase of the likelihood of T. leucotreta adults escaping and mating would also be expected in case of spatial or temporal clustering of cut roses consumption in a particular residential area or during a particular time of the year. Peaks of cut roses consumption are expected during the year, e.g. with peaks in Valentine's Day, Mother's Day and Easter. Spatial and temporal clustering may also occur due to ceremonies or cultural events (e.g. marriages, funerals, celebrations or festivals). However, data on such spatial and temporal clustering of cut roses are not available at NUTS2 level, and no adjustments to the calculations were made.
3.4.5. Host plants availability
With regard to host plants availability, in line with EPPO (2013), it is assumed that a female of T. leucotreta, having an extremely wide range of plants suitable for oviposition, will likely find suitable plants for oviposition even during the winter in the areas climatically suitable (see also Section 3.5). This is particularly true for the coastal areas of Southern Europe, where areas of intense horticulture occur close to highly urbanised regions and to natural Mediterranean evergreen vegetation. Citrus host plants are also widely used as ornamentals in urban areas and hence provide host fruits for oviposition.
3.4.6. Uncertainties affecting the assessment of entry
The key uncertainties on the entry assessment for T. leucotreta via the cut roses pathway regard:
data on the prevalence in the environment and the level of infestation of T. leucotreta in the cut roses farms in the exporting countries;
data on the abundance and clustering of T. leucotreta in single consignments of cut roses. Clustering could potentially increase the probability of entry by: (1) limiting the detection efficiency at the border; (2) increasing the likelihood of mating (and therefore the probability of transfer) when highly infested bouquets or consignments of cut roses are distributed in the same location and time;
data on the effectiveness of the border inspections at export in the countries of origin in detecting the different life stages of T. leucotreta in cut roses, especially with regard to the details of the national procedures in place;
data on the effectiveness of the border inspections at import in the EU MS in detecting the different life stages of T. leucotreta in cut roses;
information on the practical implementation and timelines of the waste management processes in the EU Member States at NUTS2 level, particularly about: the private composting; the actual time between the initial waste disposal and the waste treatment in the EU MSs at NUTS2 level;
lack of knowledge about development to adulthood of T. leucotreta in cut roses and in waste. These stages (e.g. during composting) might pose additional mortality (in addition to the overall mortality range already included in the pathway model). The Panel noted, however, that preliminary trials suggested that the complete development of T. leucotreta on cut roses in a simulated waste bin environment is possible (NVWA 2022: Development of Thaumatotibia leucotreta on cut roses; personal communication, 23 September 2022);
there is a lack of information about spatial and temporal clustering of cut roses and their eventual disposal. For instance, one might infer that there is an increase of disposal of cut roses in the period after Valentine's Day, mother's day, Easter or other festivities.
Results of the sensitivity analysis of the pathway model using the contribution of each factor to total R2 are shown in Table 16 below.
Table 16.
Sensitivity analysis of the pathway model showing the contribution of each factor to total R2 = 1
| Rel. decomposition of R2 | ||||
|---|---|---|---|---|
| Uncertainty decomposition | Total FCM (3 days) | Total FCM (7 days) | Total FCM (14 days) | Total FCM (28 days) |
| Infestation rate | 50.7% | 52.9% | 63.4% | 88.0% |
| Proportion private compost | 47.8% | 45.7% | 34.8% | 9.5% |
| Natural survival | 1.1% | 1.1% | 1.4% | 1.9% |
From the relative decomposition of the R2 (Table 16 and Appendix D), it emerges that the overall uncertainty in the output is mostly driven by the infestation rate and the relative proportion of waste treated as private compost; it has to be noted that the uncertainty of the proportion of private composting decreases as the time to treatment increases.
3.4.7. Conclusion on the assessment of entry
The Panel modelled the entry of T. leucotreta estimating the infestation level in cut roses imported into the EU, the trade flow and distribution into EU NUTS2 regions and the escape of T. leucotreta adults from the cut roses disposal before and after the waste treatment in the EU NUTS2 regions with areas climatically suitable for the establishment of T. leucotreta. Four scenarios, for the timespan from the initial disposal of the cut roses at the household until the waste treatment, were considered: 3, 7, 14 and 28 days.
According to model results, the median number of T. leucotreta adults escaping from imported cut roses in all the climatically suitable NUTS2 regions of the EU was estimated in 49,867 per year (90% uncertainty between 5,298 and 234,393) for the scenario of timing from initial disposal until waste treatment of 3 days and up to 143,689 per year (90% uncertainty between 21,126 and 401,458) for 28 days timing until waste treatment. The major differences across the scenarios are due to the escapes of adults across the regional waste management processes, whereas the escapes from the private compost remain constant across all scenarios.
In EU NUTS2 regions with higher climate suitability, for the scenario of 3 days until waste, the median number of T. leucotreta adults per year escaping from the cut roses varied from 1510 for Sicily (Italy) to 506 for Attica (Greece), to 435 in Andalusia (southern Spain) and 257 in the Region of Valencia (eastern Spain). In some EU NUTS2 regions with lower climate suitability, the estimated median number of adults escaping from cut roses was larger (e.g. in the Rhône‐Alpes region in France) than in other more climatically suitable areas (e.g. in Spain) (Figure 20), due to the large number of cut roses from Africa consumed in the region.
When the period until waste treatment becomes longer, the number of escapes increases. For example, for Andalusia (Spain), the median number of escapes per year increased to 1,848 (90% uncertainty between 269 and 5,440) when the time from initial disposal until waste treatment was 28 days.
As evidenced by the sensitivity analysis in Table 16 and Appendix D, the uncertainties in the model outputs are mostly driven by the input parameters: (i) infestation rate and (ii) the relative proportion of waste treated as private compost while the contribution of the uncertainty in trade data and natural survival shown only marginal contribution.
Assuming as a realistic scenario that on average one in every 435 escaping T. leucotreta results in a mating, the estimated median number of T. leucotreta mated females per year from imported cut roses in all the climatically suitable NUTS2 regions of the EU would vary from 115 (90% uncertainty between 12 and 538) up to 330 (90% uncertainty between 49 and 923) for the 3‐ and 28‐day scenarios, respectively. From the results disaggregated into seasons, in the EU a higher number of mated females are predicted in summer compared to the other seasons in the longer scenarios of 14 and 28 days until treatment. In particular, for the 28 days scenario, the number of mated females in summer would be 185 (90% uncertainty between 28 and 480), contributing more than 50% of the total annual mated females.
Factors like the clustering of infestation in the cut roses or the spatial or temporal clustering of cut roses consumption in a particular residential area or during particular times of the year would increase the number of mated females.
With regard to host plants availability, the Panel agreed with EPPO (2013) on the wide availability of suitable hosts in the coastal areas of Southern Europe. A female of T. leucotreta, being extremely polyphagous, would likely find suitable hosts for oviposition even during the winter in the climatically suitable areas.
3.5. Assessment of climatic factors and host distribution affecting establishment
Climatic mapping is the principal method for identifying areas that could provide suitable conditions for the establishment of a pest taking key abiotic factors into account (Baker, 2002). Climatic factors and availability of hosts are considered in the following sections.
3.5.1. Köppen–Geiger climate classification approach
Figure 23 shows the results of the Köppen–Geiger climate classification approach. It shows the climate types that are present in the EU and have also been associated with occurrences of FCM. Most of the observations occur under the semi‐arid (BSk, BSk) and temperate dry (Csa, Csb) climates, while fewer observations, mainly clustered around the east side of Lake Victoria and on the coast of South Africa, fall under temperate wet climates (Cfa, Cfb, Cfc).
Figure 23.
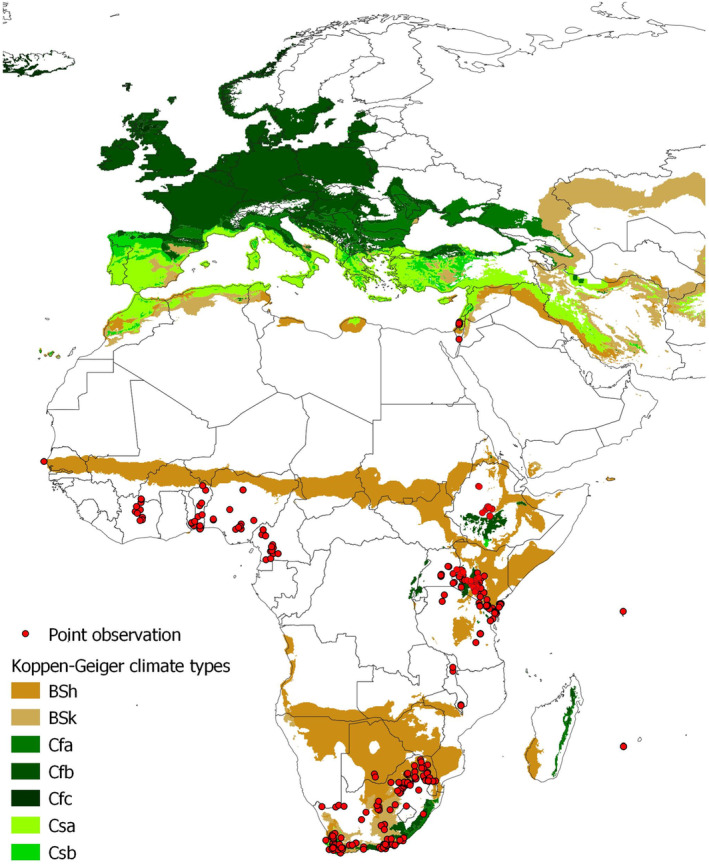
Climate suitability map of T. leucotreta for Europe and Africa, based on the Köppen–Geiger climate classification. Red dots indicate point observations of the pest (coordinates). Climates that are not present in the EU are not mapped. The legend shows the list of Köppen–Geiger climates occurring in the locations of observation
Figure 24 focuses on the location of observations in Southern Africa and Eastern Africa.
Figure 24.
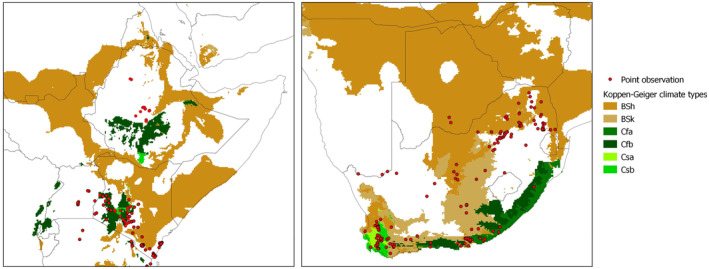
Climate suitability map of T. leucotreta for Eastern Africa, and South Africa, based on the Köppen–Geiger climate classification. Red dots indicate point observations of the pest (with coordinates). Climates not present in EU are not mapped. The legend shows the list of Köppen–Geiger climates occurring in the locations of observation
3.5.2. Physiologically based demographic model
The PBDM of T. leucotreta was applied to predict the average number of pupae for all 25 × 25 km cells in EU MS (see Figures 25 and 26) based on climate data of 2000–2010 (1990–2000 climate was additionally applied to balance the model dynamics). The outcome is presented as a colour map with colours referring to the average annually number of pupae in the cell. PBDM model predictions were mapped using the Turbo colour map (Mikhailov, 2020) to maximise graphical inference (see Reda and Szafir, 2021).
Figure 25.
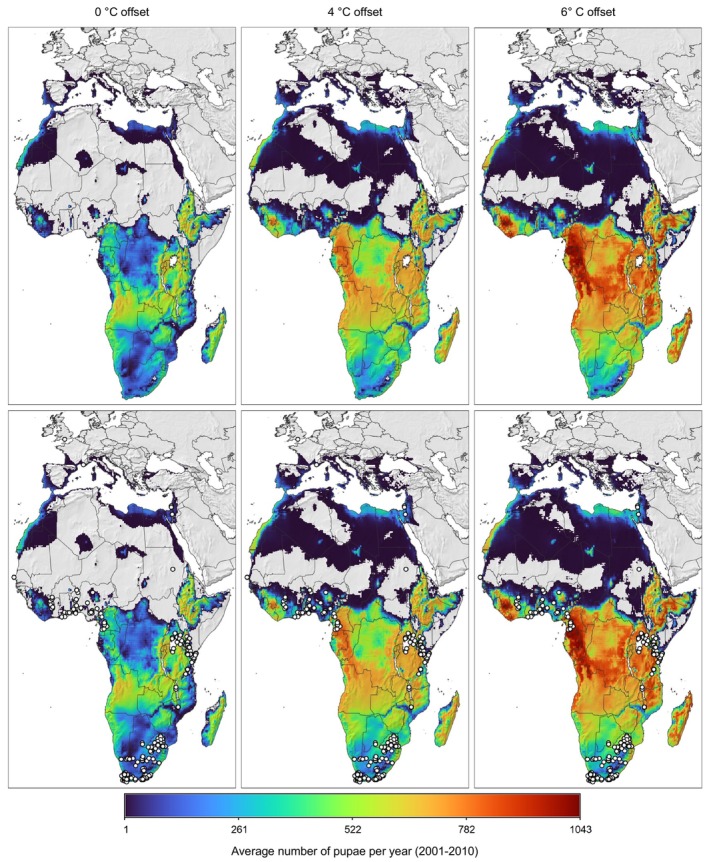
Maps showing average T. leucotreta pupal density in Europe, the Middle East and Africa for the period 2000–2010 (average number of pupae per year, as projected by the PBDM). Columns of maps show the effect of larval heat tolerance assuming displaced heat‐stress mortality: left column without [data range (1, 696]), middle column with 4°C (Terblanche et al., 2017) [data range (1, 828)] and right column with 6°C (Uys, 2014) [data range (1, 1043)] displacement of larvae mortality relative to adults (heat tolerance of larvae increases left to right). The top versus bottom rows of maps differ only in that the bottom maps have T. leucotreta occurrence records superimposed (white circles with black outline). Note the model predicts infested areas from Gabon south to Angola with no records of T. leucotreta. The colour legend uses the turbo colour palette (Mikhailov, 2020) to maximise graphical inference (Reda and Szafir, 2021) and extends to the absolute minimum and maximum values across the three maps (i.e. 1–1043) to facilitate comparison between maps.
Figure 26.
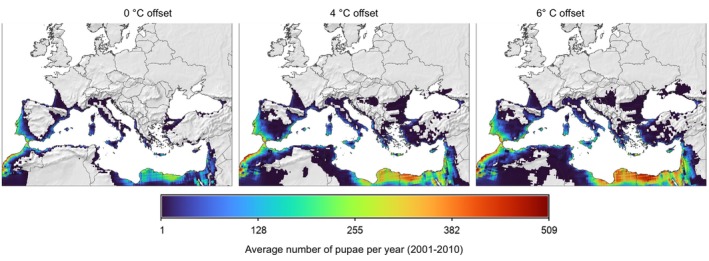
The predicted level of T. leucotreta establishment following entry into EU MS and the Mediterranean Basin (average number of pupae per year in the period 2001–2010, as projected by the PBDM model). The three columns of maps show the effect of different levels of heat tolerance. Left without [data range (1, 362)], middle with 4°C [data range (1, 467)] and right with 6°C [data range (1, 509)] displacement of larvae mortality relative to adults (cf. Figure 16). The colour legend uses the turbo colour rule (Mikhailov, 2020) to maximise graphical inference (Reda and Szafir, 2021) and extends to the absolute minimum and maximum values for Europe and the Mediterranean Basin across the three maps (i.e. 1–509)
Predictions for EU MS are first displayed in maps encompassing Africa, south‐west Asia and Europe to show the potential geographic distribution and relative abundance of T. leucotreta in the context of its wider native (sub‐Saharan) and current invasive (South African and Israeli) distribution. Then, the focus moves to maps including Europe and the Mediterranean Basin for a more detailed assessment of T. leucotreta establishment risk in EU MS and neighbouring countries.
Using 4°C and 6°C temperature offset scenarios of increased larval tolerance to high temperatures (see Sections 2.4.5 and 3.1), T. leucotreta predicted population densities remain nearly unchanged for Kenya and Ethiopia, whereas increasing T. leucotreta densities are predicted in southwest Africa due to the increased larval tolerance to heat (Figure 25). The T. leucotreta model with larval heat tolerance displacement captures the distribution of T. leucotreta in eastern parts of Africa (note that Kenya remains above the midpoint of the density range) but marginally the infestations in South Africa (invaded areas) and hotter regions such as Nigeria, Benin, Togo and Ghana (0.25–0.50 of the pupal density range).
Restricting the attention to Europe, maps with 0, 4 and 6°C displacements for Europe and the Mediterranean Basin indicate little change in the underlying geographic distribution of T. leucotreta, but increased population densities in hotter areas such as Morocco, Egypt and Israel/Palestine. No areas show predicted T. leucotreta population densities as high as those of the occurrence data points in Kenya and Ethiopia, with maximum densities being ~ 40–50% lower. However, there are spots in Europe which have shown predicted levels comparable with those in the Western Cape of South Africa (Figure 26) where the moth is considered naturalised (established although not indigenous) (Giliomee and Riedl, 1998; Hofmeyr et al., 2015).
Due to the cold limitations in large parts of the EU MS, the building of greater numbers of pupae over the 10 years of model analysis is prevented. The areas that have average pupae density like Israel (established invasion; Wysoki, 1986) are limited to the southern parts of the Mediterranean and Atlantic coastline of the EU (in particular in Portugal, Spain, Italy, Greece (i.e. Crete) and Cyprus).
For comparison, the abundance projection of the PBD is shown together for Africa and Europe (i.e. combining Figure 11 with predictions for Europe in Figure 25) but using the same colour code across the whole map. Indeed, in the southern part of the Iberian Peninsula, the model projects equal pupal abundance as shown in parts of South Africa where the moth has presence reports while being naturalised there. However, population densities predicted in native cooler subtropical areas of Africa (e.g. Kenya) are not predicted anywhere in the EU.
3.5.3. Uncertainties affecting the assessment of climatic suitability and presence of hosts in the risk assessment area
The most important uncertainty when assessing habitat suitability for establishment was that most data are about temperature dependence and all experimental data were produced for a different purpose than parameterising a quantitative pest risk assessment.
The literature on T. leucotreta is primarily from South Africa, where the pest is considered non‐native. The data on its thermal biology were developed to inform quarantine procedures for the export of agricultural products and are based on stock colonies maintained at optimal non‐fluctuating temperatures in the laboratory. This research focused on short period exposures of T. leucotreta life stages to cold and high temperatures for varying short periods of time (hours) to assess survival. While the data are useful for quarantine purposes, such data do not enable accurate evaluation of the potential of T. leucotreta natural populations to survive and establish in novel areas worldwide (e.g. Europe).
T. leucotreta developmental, mortality and reproductive rates are affected by temperature, moisture and host quality that vary in time and geographic space. Available data on the thermal biology for T. leucotreta development and survival were generally adequate in the favourable range (10–30°C) but were inadequate at low (< 10°C) and high (> 30°C) temperatures. This led to the development of a composite function for mortality that incorporated data from different life stages.
The niche model of T. leucotreta for Australia (Li et al., 2022) suggests a potentially inverse dependence of habitat suitability on precipitation in the driest quarter. There are no experimental data on this parameter (or what it means), and hence, it could not be included in the PBDM projections. Interestingly, Kenyan hearing experts did report ‘T. leucotreta is more prevalent at the onset of the rainy season’. Although the aspect of the T. leucotreta niche might be less relevant under precipitation regimes in EU territories, it likely could improve the validity of the PBDM model. It is therefore assumed important to consider precipitation with upcoming exposure experiments on the pest.
To refine the predictions of invasive risk of T. leucotreta following data should be assembled:
experimental data on the effects of temperature on developmental and survival rates of all life stages, and the effects of temperature on oviposition profiles and life tables especially on cut roses;
the effects of relative humidity (or vapour pressure deficit) on the survival of the life stages exposed to ambient conditions are required to refine temperature‐dependent stage‐specific survival rates, and reproduction by adult females;
the genetic background of populations in areas of Africa with higher temperatures such as West Africa where T. leucotreta is also reported to occur.
There is generally a convincing agreement between model predictions and observation data. This argues for the validity of PBDM predictions that do not rely on occurrence data. Interestingly, another related approach produced a map comparable to that shown in Figure 26 (Barker and Coop, pers. Comm) using degree day patterns to predict cold stress‐related exclusion areas and else predicting the multiplicity of generations given entry. The data of a third model projecting T. leucotreta habitat quality to the EU territory using MAXENT niche modelling plus degree day approach (see Figure 27, by courtesy of Li and McKirdy, personal communication, 13 January 2023) is in general agreement with Panel predictions. The predicted average pupal density above one (Figure 26) overlaps with the part of the inclusion area where Barker (personal communication, data not shown) predicts at least three generations per annum, while Li and McKirdy (Li and McKirdy, 2023, personal communication, 13 January 2023) project similar habitats along the Atlantic coast of the Iberian Peninsula and the Mediterranean coast of the EU (Figure 27).
Figure 27.

Climatic suitability map (with continued data) for the potential establishment of T. leucotreta in Europe, according to the species distribution model developed for Australia by Li et al. (2022). Personal communication by courtesy of Xingyu Li and Simon McKirdy, received by email on 13 January 2023
All three models are based on climate data with Li et al. (2022) additionally exploiting occurrence data. Hence, the PBD Model used here is confirmed by two independent model outcomes when extrapolating temperature‐dependent T. leucotreta biology to local suitability for establishment and/or population growth.
The apparent difference between the PBDM and the two other approaches is regions north of the Pyrenees, Ireland and the UK. These areas were not excluded by the degree‐day approach due to the absence of mortality during cold periods and yet predicted sufficient degree days for the development of up to two generations of T. leucotreta during favourable periods but tell little about the dynamics or the magnitude of the population. But the temporal cohesion of the degree‐days is not considered and strongly influences the continuous development time in the PBD Model during cold periods. If the development days are not in a consecutive chain, fewer life cycles can be completed, resulting in lower average numbers of pupae. If the output of the PBD model was plotted for values below one, the apparent difference between models factually disappeared (data not shown).
It has to be noted that also the preliminary climate suitability assessment by EPPO (2013) projects similar climate suitability.
The uncertainty always remains as to whether other, yet unidentified climatic or ecological features might impact the temperature‐dependent physiology and expand/contract the region of possible establishment. Hence, it is possible that the inclusion of other physiologically based determinants in the PBDM, such as those related to precipitation and humidity, for which no information is available, could further affect the estimated risk of establishment of the pest and the size of the risk area of establishment.
3.5.4. Conclusions on establishment
In summary, T. leucotreta larvae on African roses imported into the EU will be primarily affected by cold stress exacerbated by a lack of dormancy/diapause in the pest.
All model‐based projections identify coastal areas with moderate temperatures (Mediterranean coasts, Iberian Atlantic coast, Pacific coast in USA and Australian coasts). These areas are characterised by limited cold periods of cool temperatures and by warm but not excessively hot summers.
The Mediterranean coastline and the western coast of the Iberian Peninsula are the main areas for potential establishment of T. leucotreta in the EU. Moreover, additional areas have the potential to host transient populations but not establishment due to limited population growth and discontinued winter survival outdoors preventing multiple generations.
As stated by EPPO (2013), T. leucotreta could overwinter in greenhouse in areas in cooler areas with intense horticultural production.
3.6. Overall uncertainty
The major sources of uncertainty identified in this assessment and for which data collection is recommended are:
data on T. leucotreta infestation rate of cut roses in the country of origin and in consignments at export;
data on the actual timing between the waste disposal at household and the waste treatments in the EU MSs at NUTS2 level;
data on biology and ecology of T. leucotreta in natural environment and in cut roses in East Africa. In particular, there is a need to collect data used to estimate relevant parameters of the thermal biology of T. leucotreta originate from experiments carried out at constant temperatures using laboratory‐reared specimens.
4. Conclusions
Following a request from the European Commission, the EFSA Panel on Plant Health performed a partial quantitative pest risk assessment of T. leucotreta for the EU, via the cut roses import pathway.
The area potentially suitable for establishment of the pest was assessed using the climate matching according to Köppen–Geiger categories and the physiologically based demographic model. The predictions of the PBDM were validated with the occurrence data from the areas of origin of the pest and recent invasion areas.
T. leucotreta larvae entering into the EU will suffer by cold stress exacerbated by a lack of dormancy/diapause in the pest, thus limiting the NUTS2 regions climatically suitable for establishment.
The area of potential establishment includes the coastline extending from the northwest of the Iberian Peninsula through the Mediterranean. However, the estimated population densities in the EU are not reaching the high population densities projected for T. leucotreta‐native East Africa.
Additional areas in the EU, indicated as suitable but with low population numbers, are most likely associated with transient populations. Besides outdoor establishment in regions climatically suitable, as stated by EPPO (2013), T. leucotreta could overwinter in greenhouses in other areas with horticultural production.
Other published models are in broad agreement regarding the major areas at risk of establishment in the EU, which is related to the common use of data of temperature requirements for this pest.
The main uncertainties about possible establishment in the EU territory are caused by the lack of demographic studies at different temperatures and population studies in the cultivated and natural environment in the areas of pest distribution.
Considering entry, the pest has been frequently intercepted in cut roses and there are observational records of flying adults of the pest in a few locations in the EU.
The Panel modelled the entry of T. leucotreta estimating the infestation level in cut roses imported into the EU, the trade flow and distribution into EU NUTS2 regions, and the escape of T. leucotreta adults from the cut roses disposal before and after the waste treatment in the EU NUTS2 regions presenting areas climatically suitable for the establishment of T. leucotreta. Four scenarios, for the timespan from the initial disposal of the cut roses at the household until the waste treatment, were considered: 3, 7, 14 and 28 days.
According to model results, the median number of T. leucotreta adults escaping from imported cut roses in all the climatically suitable NUTS2 regions of the EU was estimated in 49,867 per year (90% uncertainty between 5,298 and 234,393) for the scenario of timing from initial disposal until waste treatment of 3 days and up to 143,689 per year (90% uncertainty between 21,126 and 401,458) for 28 days timing until waste treatment. The differences across the scenarios are due to the escapes across the regional waste management processes, whereas the escapes from the private compost remain constant across all scenarios. Assuming as a realistic scenario that, on average, for every 435 escaping T. leucotreta a mating would happen, the estimated median number of T. leucotreta mated females per year from imported cut roses in all the climatically suitable NUTS2 regions of the EU would vary from 115 (90% uncertainty between 12 and 538) up to 330 (90% uncertainty between 49 and 923) for the 3‐ and 28‐day scenarios, respectively. From the results disaggregated into seasons, summer showed a higher number of expected mated females respect to the other seasons for the EU in the longer scenarios of 14 and 28 days until treatment. In particular, for the 28 days scenario, the mated females in summer would be 185 (90% uncertainty between 28 and 480), contributing to more than 50% of the total annual mated females. Factors like the clustering of the infestation in the cut roses or the spatial or temporal clustering of cut roses consumption in a particular residential area or in a particular time of the year would increase the probability of mating and transfer to suitable host.
With regard to host plants availability, the Panel agreed with EPPO (2013) on the wide availability of suitable hosts in the coastal areas of Southern Europe. A female of T. leucotreta, having an extremely wide range of suitable plants for oviposition, will likely find suitable plants for oviposition even during the winter in the areas climatically suitable. Due to the extreme polyphagia of T. leucotreta immature stages, host availability should not be a limiting factor for establishment in climatically suitable areas.
Overall, regular escape of pest insects on the territory of the EU is predicted through the cut roses but so far it has not led to outbreaks (other than few incursions) in the EU, possibly because of the relatively recent shift of pest pressure in Africa towards cut roses and of the fact that much of the cut roses consumption in the EU occurs in regions with less climate suitability. However, observations of flying adults have been reported in the EU.
The outputs of this quantitative pest risk assessment indicate that cut roses provide a pathway for the introduction of T. leucotreta to the EU.
Regarding the seasonality, the number of escaped adults of T. leucotreta with a possible mating partner from the imported cut roses in the realistic clustering scenario is predicted to be higher during summer, particularly when the 14‐ and 28‐day scenarios until waste treatment are considered, due to the predicted faster development of T. leucotreta in the warmer season.
Sensitivity analysis of the pathway model showed that the main uncertainties remain regarding the infestation rate in the imported cut roses and main parameters of the waste model, especially the proportion of waste privately composted and the timing between initial disposal of the cut roses in the household and the waste treatment in the public facilities.
To reduce the uncertainties, data collection and research are recommended on the following key topics: the ecology and biology of T. leucotreta in its natural environment and in cut roses in Eastern Africa; the level of infestation and clustering of T. leucotreta in the cut roses consignments; the level of effectiveness of the export and import border inspections in detecting the different life stages of T. leucotreta in cut roses; the actual waste management processes at NUTS2 level in the EU, including the proportion of private composting and the timing between the initial waste disposal and the waste treatment.
Abbreviations
- CN
Combined Nomenclature (8‐digit code building on HS codes to provide greater resolution)
- CR
Certainty Range
- DD
Degree Days
- EI
Ecoclimatic index (an index of climatic suitability used by CLIMEX)
- EKE
Expert Knowledge Elicitation
- ENM
Ecological niche model
- EPPO
European and Mediterranean Plant Protection Organisation
- FAVV‐AFSCA
Federaal Agentschap voor de Veiligheid van de Voedselketen‐ Agence Fédérale pour la Sécurité de la Chaîne Alimentaire (Federal Agency for the Safety of the Food Chain)
- FCM
False Codling Moth
- HS
Harmonised System (6‐digit World Customs Organisation system to categorise goods)
- IPM
Integrated Pest Management
- IPPC
International Plant Protection Convention
- KEPHIS
Kenya Plant Health Inspectorate Service
- MS
Member State (of the EU)
- NPPO
National Plant Protection Organisation
- NUTS
Nomenclature Units for Territorial Statistics
- NVWA
Nederlandse Voedsel‐ en Warenautoriteit (Netherlands Food and Consumer Product Safety Authority)
- PBDM
Physiologically Based Demographic Model
- PRA
Pest Risk Analysis
- ToR
Terms of Reference
Appendix A – Pathway model on the entry of Thaumatotibia leucotreta via cut roses from Africa
A.1. Result table
The following table compares the annual escape of Thaumatotibia leucotreta by different waste collection scenarios: The timing from disposal by the consumer to waste treatment varies from 3, 7, 14 to 28 days.
A.1, A.2, A.3, A.4, A.5, A.6, A.7, A.8, A.9, A.10, A.11, A.12, A.13, A.14, A.15
Figure A.1.
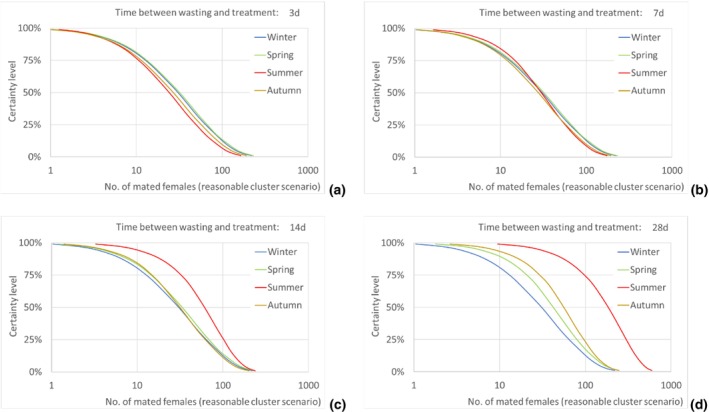
Average seasonal number of female T. leucotreta with possible mating partner in a bunch of 10 roses (Reasonable clustering scenario) per season (blue line = winter, green = spring, red = summer and yellow = autumn) in the EU climatically suitable area with certainty curves for different scenarios between initial wasting and waste treatment: (a) 3 days, (b) 7 days, (c) 14 days and (d) 28 days
Figure A.4.
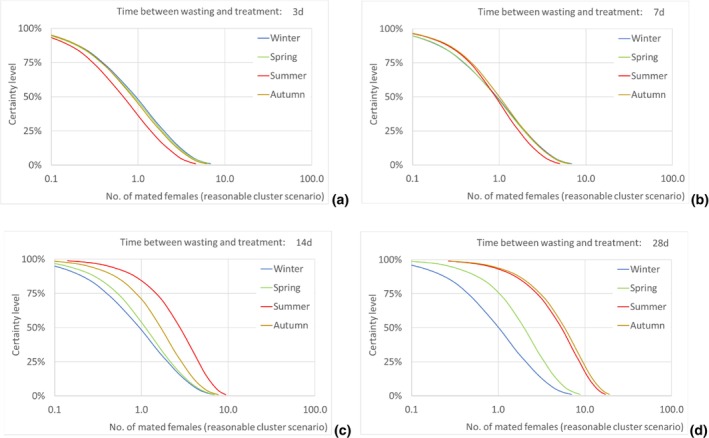
Average seasonal number of female T. leucotreta with possible mating partner in a bunch of 10 roses (Reasonable clustering scenario) per season (blue line = winter, green = spring, red = summer and yellow = autumn) in Sicily (Italy) with certainty curves for different scenarios between initial wasting and waste treatment: (a) 3 days, (b) 7 days, (c) 14 days and (d) 28 days
Figure A.5.
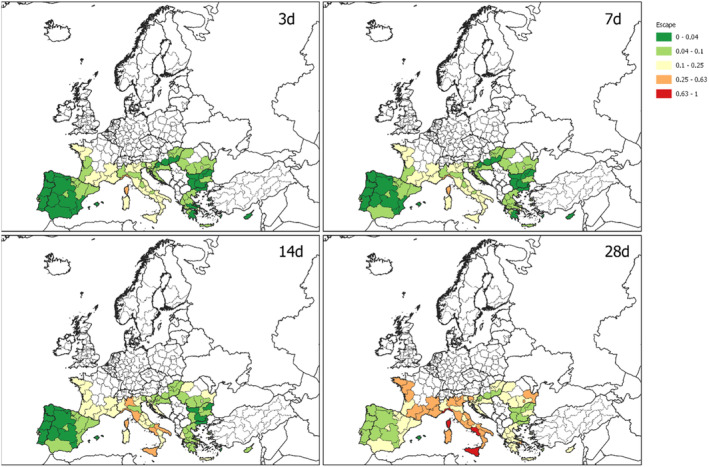
Maps of the European NUTS2 regions that are climatically suitable for T. leucotreta establishment showing the estimated average numbers of T. leucotreta adult escapes standardised to an area of 3.14 km2 over 10 days in the residential area of each NUTS2 region (log‐scale). Results are presented for the four different scenarios of timing from the initial disposal until the waste treatment of 3, 7, 14 and 28 days
Figure A.6.
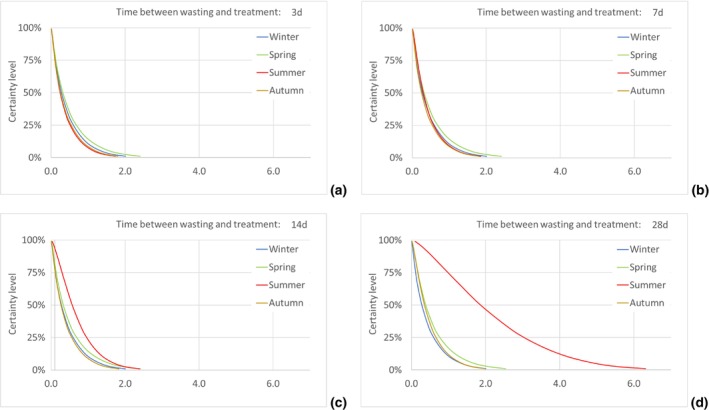
Estimated number of T. leucotreta adults escaping from disposed infested cut roses in a 3.14‐km2 circle per 10 days in the residential areas of Corsica (France) with distribution functions for different scenarios between initial wasting and waste treatment: (a) 3 days, (b) 7 days, (c) 14 days and (d) 28 days
Figure A.7.
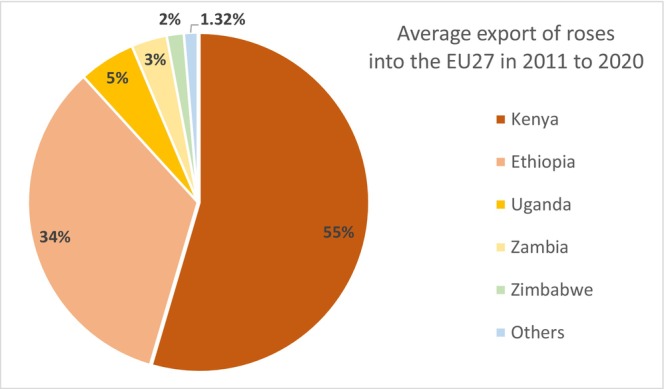
Average relative export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to EU (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8 [DS‐645593], CN 06031100, online, accessed on 4 October 2022)
Figure A.8.
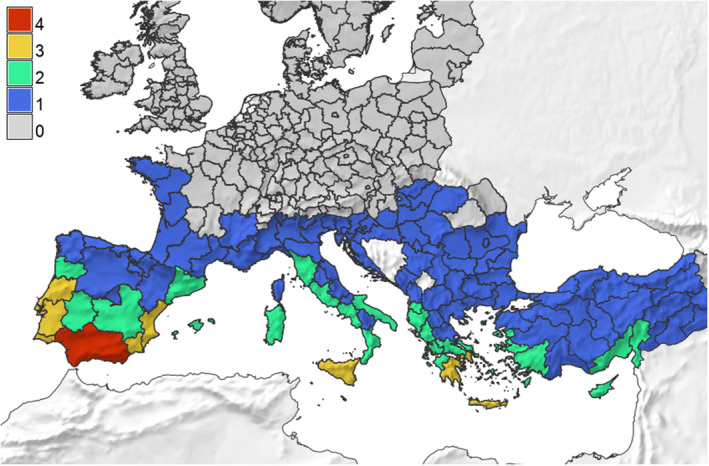
NUTS 2 regions in EU and some neighbouring countries, and their climate suitability classes according to the potential establishment of T. leucotreta based on the average number of pupae/year as estimated by the physiologically based demographic model (PBDM, see Appendix B). 0 = less than 1 pupae/year, 1 = between 1 and 128 pupae/year, 2 = between 128 and 255 pupae/year, 3 = between 255 and 382 pupae/year and 4 = between 382 and 509 pupae/year. Note: Kosovo and Bosnia and Herzegovina are not covered in the layer of NUTS 2 boundaries by Eurostat (year 2021; https://ec.europa.eu/eurostat/web/gisco/geodata/reference-data/administrative-units-statistical-units/nuts)
Figure A.9.
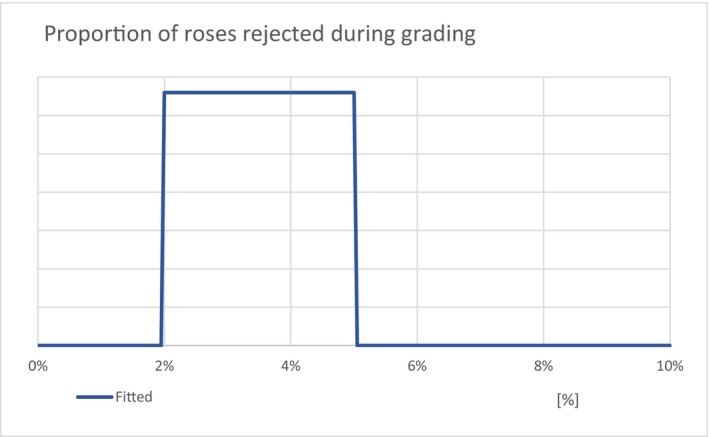
Density function showing the uncertainty distribution of the proportion of cut roses rejected in grading step directly after import into the EU
Figure A.10.
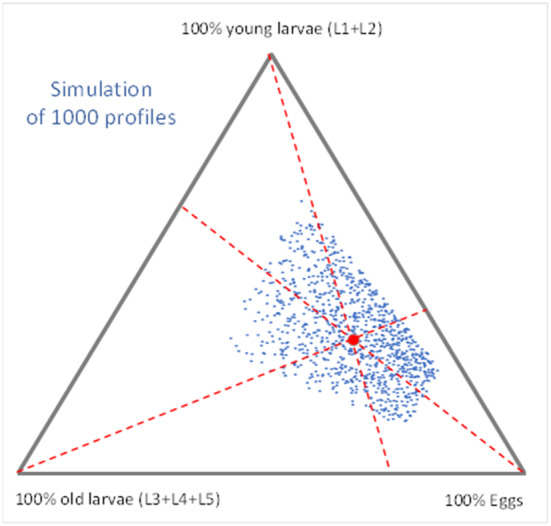
Simulation of the profile of infestation of cut roses at the EU border with immature life stages of FCM (in a triangular diagram: Each corner represents a pure infestation with only one life stage)
Figure A.11.
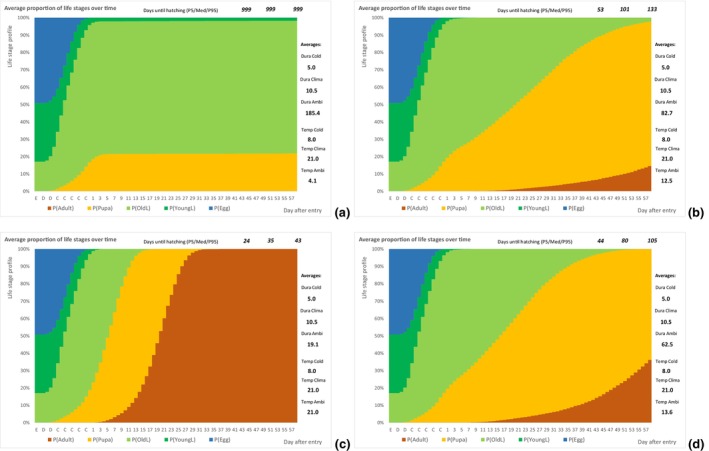
Average proportion of different life stages (blue = eggs, dark green = L1–L2, light green = L3–L5, yellow = pupae, red = adults) of Thaumatotibia leucotreta by day (horizontal axis) starting with entering the EU (E), distribution in cooled conditions (D), consumption in climatised environment (C) and days after wasting by the consumer. Results by the developmental model are given for NUTS2 regions in climate suitability class 1 for winter (a), spring (b), summer (c) and autumn (d)
Figure A.12.
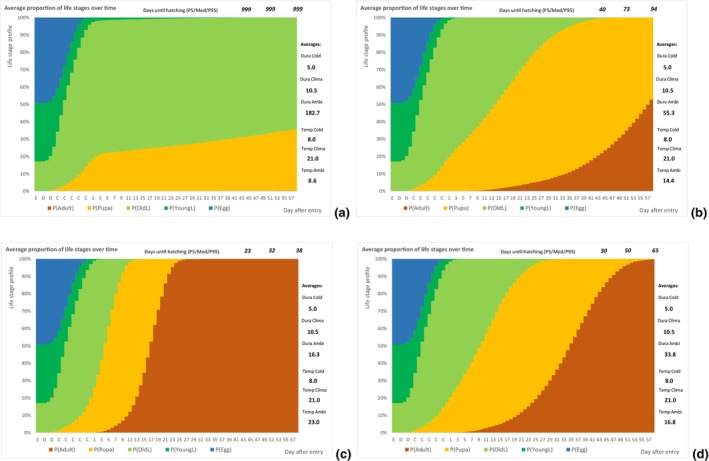
Average proportion of different life stages (blue = eggs, dark green = L1–L2, light green = L3–L5, yellow = pupae, red = adults) of Thaumatotibia leucotreta by day (horizontal axis) starting with entering the EU (E), distribution in cooled conditions (D), consumption in climatised environment (C) and days after wasting by the consumer. Results by the developmental model are given for NUTS2 regions in climate suitability class 2 for winter (a), spring (b), summer (c) and autumn (d)
Figure A.13.
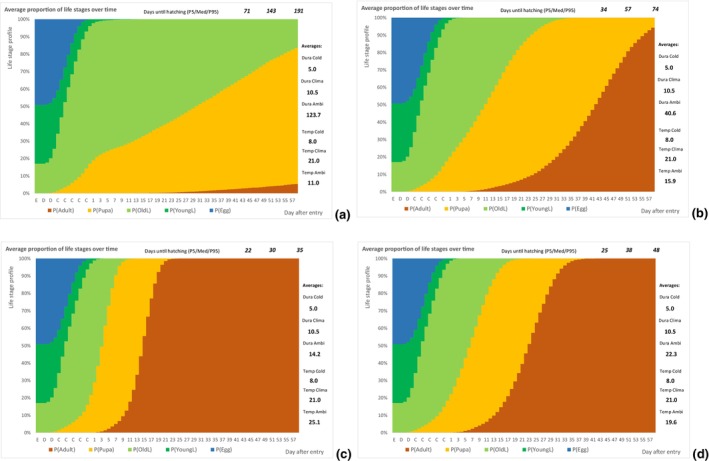
Average proportion of different life stages (blue = eggs, dark green = L1–L2, light green = L3–L5, yellow = pupae, red = adults) of Thaumatotibia leucotreta by day (horizontal axis) starting with entering the EU (E), distribution in cooled conditions (D), consumption in climatised environment (C) and days after wasting by the consumer. Results by the developmental model are given for NUTS2 regions in climate suitability class 3 for winter (a), spring (b), summer (c) and autumn (d)
Figure A.14.
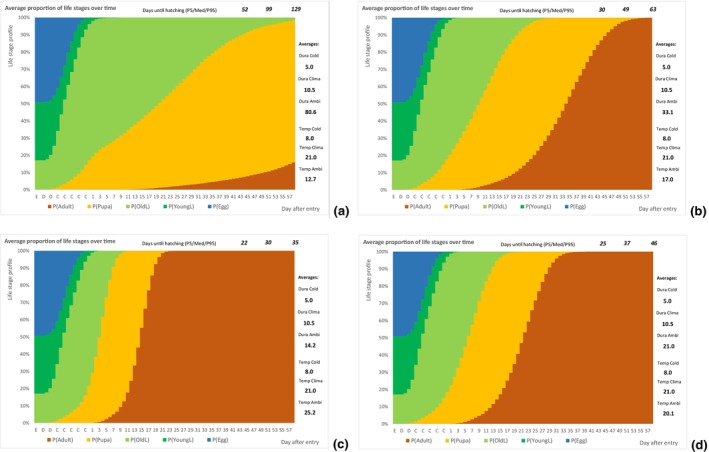
Average proportion of different life stages (blue = eggs, dark green = L1–L2, light green = L3–L5, yellow = pupae, red = adults) of Thaumatotibia leucotreta by day (horizontal axis) starting with entering the EU (E), distribution in cooled conditions (D), consumption in climatised environment (C) and days after wasting by the consumer. Results by the developmental model are given for NUTS2 regions in climate suitability class 4 for winter (a), spring (b), summer (c) and autumn (d)
Figure A.15.
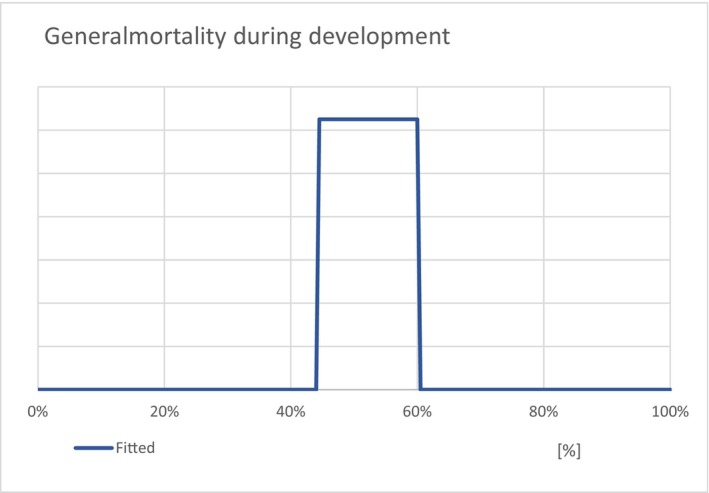
Density function showing the uncertainty distribution of the mortality during natural development of Thaumatotibia leucotreta
A.1, A.2, A.3, A.4, A.5, A.6, A.7, A.8, A.9, A.10, A.11, A.12, A.13, A.14, A.15, A.16, A.17, A.18, A.19, A.20, A.21, A.22, A.23, A.24, A.25, A.26, A.27, A.28, A.29, A.30, A.31, A.32, A.33, A.34, A.35, A.36, A.37, A.38, A.39, A.40, A.41, A.42, A.43, A.44, A.45, A.46, A.47, A.48, A.49, A.50, A.51, A.52, A.53, A.54, A.55, A.56, A.57, A.58, A.59, A.60, A.61, A.62, A.63, A.64, A.65, A.66, A.67, A.68, A.69, A.70, A.71, A.72, A.73, A.74, A.75, A.76, A.77, A.78, A.79, A.80, A.81, A.82, A.83, A.84, A.85, A.86, A87, A.88, A.89, A.90, A.91, A.92
Table A.2.
Number of Thaumatotibia leucotreta adults escaping within a year from cut roses imported from Africa (including Israel) calculated by a pathway model for NUTS2 regions in the climatically suitable area of Europe. Results are stratified by waste treated by private compost and waste treated after communal collection with four scenarios of timing from disposal at the consumer until the waste treatment. Values are given as median and 90% certainty interval (from P5 to P95)
| NUTS code | Label | Climate suitability class | Annual escape from private compost | Annual escape from collected waste (3 days) | Annual escape from collected waste (7 days) | Annual escape from collected waste (14 days) | Annual escape from collected waste (28 days) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (Adults/year) | (Adults/year) | (Adults/year) | (Adults/year) | (Adults/year) | ||||||||||||||
| P5 | Median | P95 | P5 | Median | P95 | P5 | Median | P95 | P5 | Median | P95 | P5 | Median | P95 | ||||
| Spain & Portugal | ESPT | Spain and Portugal | ||||||||||||||||
| ES11 | Galicia | 1 | 12 | 135 | 791 | 0 | 2 | 5 | 1 | 6 | 17 | 5 | 30 | 84 | 24 | 160 | 451 | |
| ES12 | Principado de Asturias | 1 | 4 | 51 | 298 | 0 | 1 | 2 | 0 | 2 | 6 | 2 | 11 | 32 | 9 | 60 | 170 | |
| ES13 | Cantabria | 1 | 3 | 29 | 171 | 0 | 0 | 1 | 0 | 1 | 4 | 1 | 6 | 18 | 5 | 35 | 97 | |
| ES21 | País Vasco | 1 | 10 | 109 | 641 | 0 | 1 | 4 | 1 | 5 | 14 | 4 | 24 | 68 | 20 | 130 | 365 | |
| ES22 | Comunidad Foral de Navarra | 1 | 3 | 33 | 192 | 0 | 0 | 1 | 0 | 2 | 4 | 1 | 7 | 20 | 6 | 39 | 110 | |
| ES23 | La Rioja | 1 | 1 | 16 | 93 | 0 | 0 | 1 | 0 | 1 | 2 | 1 | 4 | 10 | 3 | 19 | 53 | |
| ES24 | Aragón | 1 | 6 | 66 | 390 | 0 | 1 | 2 | 1 | 3 | 8 | 2 | 15 | 41 | 12 | 79 | 222 | |
| ES30 | Comunidad de Madrid | 1 | 29 | 336 | 1,976 | 1 | 4 | 11 | 3 | 16 | 43 | 12 | 75 | 210 | 60 | 400 | 1,126 | |
| ES41 | Castilla y León | 1 | 10 | 120 | 703 | 0 | 1 | 4 | 1 | 6 | 15 | 4 | 27 | 75 | 22 | 142 | 401 | |
| ES42 | Castilla‐la Mancha | 2 | 9 | 102 | 599 | 0 | 2 | 4 | 1 | 7 | 19 | 6 | 40 | 112 | 25 | 165 | 459 | |
| ES43 | Extremadura | 2 | 5 | 53 | 311 | 0 | 1 | 2 | 1 | 4 | 10 | 3 | 21 | 58 | 13 | 86 | 238 | |
| ES51 | Cataluña | 2 | 33 | 381 | 2,241 | 2 | 6 | 17 | 5 | 26 | 71 | 23 | 151 | 418 | 93 | 618 | 1,717 | |
| ES52 | Comunitat Valenciana | 3 | 22 | 251 | 1,473 | 1 | 5 | 14 | 5 | 27 | 72 | 28 | 181 | 501 | 96 | 639 | 1,764 | |
| ES53 | Illes Balears | 2 | 5 | 60 | 355 | 0 | 1 | 3 | 1 | 4 | 11 | 4 | 24 | 66 | 15 | 98 | 272 | |
| ES61 | Andalucía | 4 | 37 | 423 | 2,483 | 3 | 10 | 27 | 9 | 51 | 138 | 52 | 336 | 931 | 187 | 1,259 | 3,530 | |
| ES62 | Región de Murcia | 3 | 7 | 75 | 441 | 0 | 2 | 4 | 1 | 8 | 21 | 8 | 54 | 150 | 29 | 191 | 528 | |
| ES63 | Ciudad de Ceuta | 3 | 0 | 4 | 25 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 3 | 8 | 2 | 11 | 29 | |
| ES64 | Ciudad de Melilla | 2 | 0 | 4 | 25 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 2 | 5 | 1 | 7 | 19 | |
| PT11 | Norte | 2 | 16 | 178 | 1,047 | 1 | 3 | 8 | 2 | 12 | 33 | 11 | 70 | 195 | 44 | 289 | 802 | |
| PT15 | Algarve | 3 | 2 | 22 | 128 | 0 | 0 | 1 | 0 | 2 | 6 | 2 | 16 | 44 | 8 | 56 | 154 | |
| PT16 | Centro (PT) | 3 | 10 | 111 | 649 | 1 | 2 | 6 | 2 | 12 | 32 | 12 | 80 | 221 | 43 | 282 | 778 | |
| PT17 | Área Metropolitana de Lisboa | 3 | 12 | 143 | 838 | 1 | 3 | 8 | 3 | 15 | 41 | 16 | 103 | 285 | 55 | 364 | 1,004 | |
| PT18 | Alentejo | 3 | 3 | 35 | 206 | 0 | 1 | 2 | 1 | 4 | 10 | 4 | 25 | 70 | 14 | 90 | 247 | |
| France | FR | France | ||||||||||||||||
| FRG0 | Pays‐de‐la‐Loire | 1 | 261 | 2,689 | 12,975 | 17 | 46 | 95 | 38 | 164 | 419 | 131 | 783 | 2,108 | 626 | 4,161 | 11,324 | |
| FRH0 | Bretagne | 1 | 230 | 2,365 | 11,412 | 15 | 40 | 84 | 34 | 144 | 369 | 115 | 689 | 1,854 | 551 | 3,660 | 9,960 | |
| FRI1 | Aquitaine | 1 | 238 | 2,449 | 11,820 | 15 | 42 | 87 | 35 | 150 | 382 | 119 | 713 | 1,920 | 571 | 3,791 | 10,316 | |
| FRI3 | Poitou‐Charentes | 1 | 124 | 1,277 | 6,163 | 8 | 22 | 45 | 18 | 78 | 199 | 62 | 372 | 1,001 | 297 | 1,976 | 5,379 | |
| FRJ1 | Languedoc‐Roussillon | 1 | 196 | 2,017 | 9,735 | 13 | 34 | 71 | 29 | 123 | 315 | 98 | 588 | 1,581 | 470 | 3,122 | 8,496 | |
| FRJ2 | Midi‐Pyrénées | 1 | 211 | 2,174 | 10,490 | 14 | 37 | 77 | 31 | 133 | 339 | 106 | 633 | 1,704 | 506 | 3,364 | 9,155 | |
| FRK2 | Rhône‐Alpes | 1 | 458 | 4,712 | 22,741 | 30 | 80 | 167 | 67 | 288 | 735 | 229 | 1,373 | 3,694 | 1,098 | 7,293 | 19,847 | |
| FRL0 | Provence‐Alpes‐Côte d'Azur | 1 | 347 | 3,575 | 17,254 | 23 | 61 | 127 | 51 | 218 | 558 | 174 | 1,041 | 2,803 | 833 | 5,534 | 15,058 | |
| FRM0 | Corse | 1 | 24 | 244 | 1,175 | 2 | 4 | 9 | 3 | 15 | 38 | 12 | 71 | 191 | 57 | 377 | 1,026 | |
| Italy & Malta | ITMT | Italy and Malta | ||||||||||||||||
| ITC1 | Piemonte | 1 | 126 | 1,298 | 6,235 | 5 | 15 | 34 | 13 | 68 | 176 | 54 | 341 | 910 | 277 | 1,833 | 4,912 | |
| ITC3 | Liguria | 1 | 44 | 459 | 2,205 | 2 | 5 | 12 | 5 | 24 | 62 | 19 | 121 | 322 | 98 | 648 | 1,737 | |
| ITC4 | Lombardia | 1 | 292 | 3,019 | 14,503 | 11 | 35 | 80 | 30 | 158 | 409 | 125 | 793 | 2,117 | 643 | 4,263 | 11,426 | |
| ITF1 | Abruzzo | 1 | 38 | 390 | 1,871 | 1 | 5 | 10 | 4 | 20 | 53 | 16 | 102 | 273 | 83 | 550 | 1,474 | |
| ITF2 | Molise | 1 | 9 | 90 | 435 | 0 | 1 | 2 | 1 | 5 | 12 | 4 | 24 | 63 | 19 | 128 | 342 | |
| ITF3 | Campania | 2 | 166 | 1,720 | 8,262 | 9 | 29 | 66 | 26 | 135 | 348 | 125 | 802 | 2,133 | 500 | 3,310 | 8,778 | |
| ITF4 | Puglia | 2 | 115 | 1,190 | 5,718 | 7 | 20 | 45 | 18 | 93 | 241 | 87 | 555 | 1,476 | 346 | 2,291 | 6,075 | |
| ITF5 | Basilicata | 1 | 16 | 167 | 800 | 1 | 2 | 4 | 2 | 9 | 23 | 7 | 44 | 117 | 35 | 235 | 630 | |
| ITF6 | Calabria | 2 | 55 | 570 | 2,740 | 3 | 10 | 22 | 9 | 45 | 115 | 42 | 266 | 707 | 166 | 1,097 | 2,911 | |
| ITG1 | Sicilia | 3 | 142 | 1,468 | 7,051 | 12 | 36 | 82 | 35 | 182 | 470 | 195 | 1,257 | 3,322 | 669 | 4,432 | 11,788 | |
| ITG2 | Sardegna | 2 | 47 | 485 | 2,331 | 3 | 8 | 19 | 7 | 38 | 98 | 35 | 226 | 602 | 141 | 934 | 2,477 | |
| ITH2 | Provincia Autonoma di Trento | 1 | 16 | 164 | 789 | 1 | 2 | 4 | 2 | 9 | 22 | 7 | 43 | 115 | 35 | 232 | 621 | |
| ITH3 | Veneto | 1 | 142 | 1,469 | 7,057 | 5 | 17 | 39 | 15 | 77 | 199 | 61 | 386 | 1,030 | 313 | 2,074 | 5,559 | |
| ITH4 | Friuli‐Venezia Giulia | 1 | 35 | 363 | 1,745 | 1 | 4 | 10 | 4 | 19 | 49 | 15 | 95 | 255 | 77 | 513 | 1,374 | |
| ITH5 | Emilia‐Romagna | 1 | 130 | 1,344 | 6,457 | 5 | 16 | 36 | 14 | 70 | 182 | 56 | 353 | 942 | 286 | 1,898 | 5,087 | |
| ITI1 | Toscana | 2 | 108 | 1,112 | 5,341 | 6 | 19 | 42 | 17 | 87 | 225 | 81 | 518 | 1,379 | 323 | 2,140 | 5,674 | |
| ITI2 | Umbria | 1 | 25 | 262 | 1,259 | 1 | 3 | 7 | 3 | 14 | 35 | 11 | 69 | 184 | 56 | 370 | 991 | |
| ITI3 | Marche | 1 | 44 | 455 | 2,188 | 2 | 5 | 12 | 5 | 24 | 62 | 19 | 120 | 319 | 97 | 643 | 1,724 | |
| ITI4 | Lazio | 2 | 168 | 1,733 | 8,325 | 10 | 29 | 66 | 26 | 136 | 351 | 126 | 808 | 2,149 | 503 | 3,335 | 8,845 | |
| MT00 | Malta | 2 | 15 | 155 | 744 | 1 | 3 | 8 | 3 | 13 | 33 | 12 | 73 | 193 | 45 | 299 | 792 | |
| Slovenia | SI | Slovenia | ||||||||||||||||
| SI03 | Vzhodna Slovenija | 1 | 22 | 226 | 1,085 | 0 | 3 | 8 | 2 | 13 | 35 | 10 | 65 | 175 | 52 | 352 | 943 | |
| SI04 | Zahodna Slovenija | 1 | 20 | 205 | 983 | 0 | 2 | 7 | 2 | 12 | 31 | 9 | 59 | 159 | 48 | 319 | 854 | |
| Croatia | HR | Croatia | ||||||||||||||||
| HR02 | Pannonian Croatia | 1 | 20 | 222 | 1,142 | 0 | 3 | 9 | 1 | 13 | 43 | 7 | 64 | 223 | 36 | 344 | 1,203 | |
| HR03 | Adriatic Croatia | 1 | 26 | 283 | 1,452 | 0 | 3 | 11 | 2 | 16 | 55 | 9 | 81 | 284 | 46 | 438 | 1,530 | |
| HR05 | City of Zagreb | 1 | 15 | 167 | 857 | 0 | 2 | 7 | 1 | 10 | 32 | 5 | 48 | 168 | 27 | 258 | 903 | |
| HR06 | Northern Croatia | 1 | 16 | 171 | 879 | 0 | 2 | 7 | 1 | 10 | 33 | 5 | 49 | 172 | 28 | 265 | 926 | |
| Greece & Cyprus | Greece and Cyprus | |||||||||||||||||
| EL30 | Attiki | 3 | 41 | 431 | 2,096 | 31 | 71 | 132 | 48 | 113 | 226 | 104 | 408 | 1,093 | 235 | 1,264 | 3,493 | |
| EL41 | Voreio Aigaio | 2 | 3 | 26 | 129 | 1 | 3 | 6 | 2 | 5 | 9 | 4 | 14 | 38 | 9 | 50 | 144 | |
| EL42 | Notio Aigaio | 2 | 4 | 40 | 195 | 2 | 5 | 9 | 3 | 7 | 14 | 6 | 22 | 58 | 14 | 76 | 217 | |
| EL43 | Kriti | 3 | 7 | 73 | 357 | 5 | 12 | 22 | 8 | 19 | 38 | 18 | 69 | 186 | 40 | 215 | 595 | |
| EL51 | Anatoliki Makedonia, Thraki | 1 | 7 | 69 | 336 | 2 | 6 | 11 | 3 | 9 | 18 | 6 | 22 | 60 | 17 | 97 | 293 | |
| EL52 | Kentriki Makedonia | 1 | 21 | 216 | 1,049 | 7 | 18 | 34 | 10 | 27 | 55 | 19 | 69 | 187 | 53 | 304 | 916 | |
| EL53 | Dytiki Makedonia | 1 | 3 | 30 | 148 | 1 | 2 | 5 | 1 | 4 | 8 | 3 | 10 | 26 | 7 | 43 | 130 | |
| EL54 | Ipeiros | 2 | 4 | 38 | 187 | 2 | 4 | 8 | 3 | 7 | 14 | 6 | 21 | 55 | 13 | 73 | 208 | |
| EL61 | Thessalia | 1 | 8 | 82 | 401 | 3 | 7 | 13 | 4 | 10 | 21 | 7 | 27 | 71 | 20 | 116 | 350 | |
| EL62 | Ionia Nisia | 2 | 2 | 23 | 114 | 1 | 3 | 5 | 2 | 4 | 8 | 3 | 13 | 34 | 8 | 45 | 127 | |
| EL63 | Dytiki Ellada | 2 | 7 | 75 | 365 | 4 | 9 | 16 | 5 | 13 | 27 | 11 | 40 | 108 | 26 | 143 | 407 | |
| EL64 | Sterea Ellada | 2 | 6 | 64 | 312 | 3 | 7 | 14 | 5 | 11 | 23 | 9 | 35 | 92 | 22 | 122 | 348 | |
| EL65 | Peloponnisos | 3 | 6 | 66 | 321 | 5 | 11 | 20 | 7 | 17 | 35 | 16 | 62 | 167 | 36 | 193 | 535 | |
| CY00 | Kypros | 2 | 10 | 102 | 498 | 5 | 12 | 22 | 7 | 18 | 37 | 15 | 55 | 148 | 36 | 195 | 555 | |
| Hungary | HU | Hungary | ||||||||||||||||
| HU12 | Pest | 1 | 18 | 191 | 928 | 0 | 2 | 6 | 2 | 11 | 31 | 9 | 57 | 157 | 46 | 309 | 849 | |
| HU23 | Dél‐Dunántúl | 1 | 12 | 129 | 626 | 0 | 2 | 4 | 1 | 8 | 21 | 6 | 39 | 106 | 31 | 209 | 572 | |
| HU31 | Észak‐Magyarország | 1 | 16 | 165 | 801 | 0 | 2 | 5 | 1 | 10 | 26 | 7 | 50 | 136 | 40 | 267 | 732 | |
| HU32 | Észak‐Alföld | 1 | 21 | 213 | 1,033 | 0 | 3 | 7 | 2 | 12 | 34 | 9 | 64 | 175 | 51 | 344 | 944 | |
| HU33 | Dél‐Alföld | 1 | 17 | 182 | 881 | 0 | 2 | 6 | 2 | 11 | 29 | 8 | 55 | 149 | 44 | 294 | 806 | |
| Bulgaria | BG | Bulgaria | ||||||||||||||||
| BG31 | Severozapaden | 1 | 4 | 42 | 209 | 1 | 2 | 4 | 1 | 4 | 10 | 3 | 13 | 42 | 9 | 64 | 217 | |
| BG32 | Severen tsentralen | 1 | 4 | 44 | 222 | 1 | 2 | 4 | 2 | 4 | 10 | 3 | 14 | 44 | 10 | 68 | 230 | |
| BG33 | Severoiztochen | 1 | 5 | 53 | 265 | 1 | 3 | 5 | 2 | 5 | 12 | 4 | 17 | 53 | 11 | 81 | 275 | |
| BG34 | Yugoiztochen | 1 | 6 | 59 | 294 | 1 | 3 | 6 | 2 | 6 | 13 | 4 | 19 | 59 | 13 | 89 | 305 | |
| BG41 | Yugozapaden | 1 | 12 | 120 | 601 | 3 | 6 | 11 | 4 | 11 | 27 | 8 | 38 | 120 | 26 | 183 | 623 | |
| BG42 | Yuzhen tsentralen | 1 | 8 | 81 | 404 | 2 | 4 | 8 | 3 | 8 | 18 | 6 | 25 | 80 | 17 | 123 | 418 | |
| Romania | RO | Romania | ||||||||||||||||
| RO11 | Nord‐Vest | 1 | 36 | 400 | 2,135 | 2 | 7 | 20 | 4 | 25 | 91 | 12 | 118 | 463 | 55 | 624 | 2,475 | |
| RO22 | Sud‐Est | 1 | 34 | 373 | 1,992 | 2 | 7 | 18 | 4 | 23 | 85 | 11 | 110 | 432 | 51 | 582 | 2,309 | |
| RO31 | Sud ‐ Muntenia | 1 | 42 | 455 | 2,432 | 2 | 8 | 22 | 5 | 28 | 103 | 14 | 134 | 527 | 63 | 710 | 2,819 | |
| RO32 | Bucuresti ‐ Ilfov | 1 | 33 | 364 | 1,946 | 2 | 7 | 18 | 4 | 23 | 83 | 11 | 107 | 422 | 50 | 568 | 2,256 | |
| RO41 | Sud‐Vest Oltenia | 1 | 27 | 300 | 1,601 | 2 | 5 | 15 | 3 | 19 | 68 | 9 | 88 | 347 | 41 | 468 | 1,856 | |
| RO42 | Vest | 1 | 25 | 278 | 1,485 | 2 | 5 | 14 | 3 | 17 | 63 | 8 | 82 | 322 | 38 | 434 | 1,721 | |
Table A.3.
Average seasonal escape of females T. leucotreta with possible mating partner in a bunch of 10 roses (reasonable scenario) in the climatically suitable EU areas given for four scenarios on the timing between initial disposal and waste treatment: 3, 7, 14 or 28 days
| Average escape | Duration between initial disposal and waste treatment | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 days | 7 days | 14 days | 28 days | |||||||||
| Season | Med | P5 | P95 | Med | P5 | P95 | Med | P5 | P95 | Med | P5 | P95 |
| Winter | 31 | 3 | 148 | 31 | 3 | 148 | 31 | 3 | 148 | 31 | 3 | 148 |
| Spring | 32 | 3 | 153 | 32 | 3 | 153 | 34 | 4 | 155 | 43 | 6 | 170 |
| Summer | 24 | 3 | 112 | 30 | 4 | 121 | 61 | 9 | 177 | 185 | 28 | 480 |
| Autumn | 27 | 3 | 129 | 28 | 3 | 130 | 32 | 4 | 136 | 56 | 8 | 177 |
Table A.4.
Average density of Thaumatotibia leucotreta adults escaping within 10 days from cut roses imported from Africa (including Israel) per circle with a radius of 1 km (3.14 km2) in the residential area. Area and duration allow females to find a mating partner. Results are stratified by four scenarios of timing from disposal at the consumer until the waste treatment. Values are given as median and 90% certainty interval (from P5 to P95)
| NUTS code | Label | Climate suitability class | Total escape of FCM per 3.14 km2 and 10 days (3 days) | Total escape of FCM per 3.14 km2 and 10 days (7 days) | Total escape of FCM per 3.14 km2 and 10 days (14 days) | Total escape of FCM per 3.14 km2 and 10 days (28 days) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (adults/[3.14 m2 × 10 days]) | (adults/[3.14 m2 × 10 days]) | (adults/[3.14 m2 × 10 days]) | (adults/[3.14 m2 × 10 days]) | ||||||||||||||||
| P5 | Median | P95 | P(X ≥ 2) | P5 | Median | P95 | P(X ≥ 2) | P5 | Median | P95 | P(X ≥ 2) | P5 | Median | P95 | P(X ≥ 2) | ||||
| Spain & Portugal | ESPT | Spain and Portugal | |||||||||||||||||
| ES11 | Galicia | 1 | 0.002 | 0.018 | 0.102 | 0.000% | 0.002 | 0.018 | 0.103 | 0.000% | 0.003 | 0.022 | 0.108 | 0.000% | 0.006 | 0.043 | 0.142 | 0.000% | |
| ES12 | Principado de Asturias | 1 | 0.003 | 0.031 | 0.178 | 0.000% | 0.003 | 0.032 | 0.179 | 0.000% | 0.005 | 0.039 | 0.189 | 0.000% | 0.010 | 0.075 | 0.248 | 0.000% | |
| ES13 | Cantabria | 1 | 0.002 | 0.026 | 0.149 | 0.000% | 0.003 | 0.027 | 0.151 | 0.000% | 0.004 | 0.033 | 0.159 | 0.000% | 0.009 | 0.063 | 0.208 | 0.000% | |
| ES21 | País Vasco | 1 | 0.006 | 0.063 | 0.364 | 0.000% | 0.006 | 0.065 | 0.368 | 0.000% | 0.010 | 0.079 | 0.387 | 0.000% | 0.022 | 0.153 | 0.508 | 0.000% | |
| ES22 | Comunidad Foral de Navarra | 1 | 0.006 | 0.062 | 0.358 | 0.000% | 0.006 | 0.064 | 0.361 | 0.000% | 0.009 | 0.078 | 0.380 | 0.000% | 0.021 | 0.150 | 0.499 | 0.000% | |
| ES23 | La Rioja | 1 | 0.005 | 0.052 | 0.300 | 0.000% | 0.005 | 0.054 | 0.303 | 0.000% | 0.008 | 0.065 | 0.319 | 0.000% | 0.018 | 0.126 | 0.418 | 0.000% | |
| ES24 | Aragón | 1 | 0.005 | 0.055 | 0.322 | 0.000% | 0.006 | 0.058 | 0.324 | 0.000% | 0.008 | 0.070 | 0.342 | 0.000% | 0.019 | 0.135 | 0.449 | 0.000% | |
| ES30 | Comunidad de Madrid | 1 | 0.006 | 0.068 | 0.395 | 0.000% | 0.007 | 0.071 | 0.398 | 0.000% | 0.010 | 0.086 | 0.419 | 0.000% | 0.023 | 0.166 | 0.551 | 0.000% | |
| ES41 | Castilla y León | 1 | 0.002 | 0.024 | 0.138 | 0.000% | 0.002 | 0.025 | 0.139 | 0.000% | 0.004 | 0.030 | 0.147 | 0.000% | 0.008 | 0.058 | 0.192 | 0.000% | |
| ES42 | Castilla‐la Mancha | 2 | 0.002 | 0.019 | 0.110 | 0.000% | 0.002 | 0.020 | 0.112 | 0.000% | 0.004 | 0.028 | 0.123 | 0.000% | 0.008 | 0.055 | 0.172 | 0.000% | |
| ES43 | Extremadura | 2 | 0.002 | 0.017 | 0.097 | 0.000% | 0.002 | 0.018 | 0.099 | 0.000% | 0.003 | 0.025 | 0.108 | 0.000% | 0.007 | 0.048 | 0.152 | 0.000% | |
| ES51 | Cataluña | 2 | 0.005 | 0.056 | 0.324 | 0.000% | 0.006 | 0.059 | 0.329 | 0.000% | 0.010 | 0.083 | 0.360 | 0.000% | 0.023 | 0.161 | 0.505 | 0.000% | |
| ES52 | Comunitat Valenciana | 3 | 0.003 | 0.032 | 0.184 | 0.000% | 0.004 | 0.035 | 0.188 | 0.000% | 0.008 | 0.060 | 0.226 | 0.000% | 0.018 | 0.123 | 0.357 | 0.000% | |
| ES53 | Illes Balears | 2 | 0.001 | 0.013 | 0.073 | 0.000% | 0.001 | 0.013 | 0.074 | 0.000% | 0.002 | 0.019 | 0.081 | 0.000% | 0.005 | 0.036 | 0.113 | 0.000% | |
| ES61 | Andalucía | 4 | 0.004 | 0.040 | 0.229 | 0.000% | 0.005 | 0.044 | 0.236 | 0.000% | 0.011 | 0.077 | 0.287 | 0.000% | 0.025 | 0.169 | 0.498 | 0.000% | |
| ES62 | Región de Murcia | 3 | 0.002 | 0.021 | 0.123 | 0.000% | 0.003 | 0.023 | 0.126 | 0.000% | 0.005 | 0.040 | 0.151 | 0.000% | 0.012 | 0.082 | 0.238 | 0.000% | |
| ES63 | Ciudad de Ceuta | 3 | 0.002 | 0.019 | 0.108 | 0.000% | 0.002 | 0.021 | 0.111 | 0.000% | 0.005 | 0.035 | 0.133 | 0.000% | 0.010 | 0.072 | 0.210 | 0.000% | |
| ES64 | Ciudad de Melilla | 2 | 0.002 | 0.027 | 0.155 | 0.000% | 0.003 | 0.028 | 0.157 | 0.000% | 0.005 | 0.040 | 0.172 | 0.000% | 0.011 | 0.077 | 0.241 | 0.000% | |
| PT11 | Norte | 2 | 0.002 | 0.024 | 0.140 | 0.000% | 0.003 | 0.026 | 0.142 | 0.000% | 0.005 | 0.036 | 0.156 | 0.000% | 0.010 | 0.070 | 0.218 | 0.000% | |
| PT15 | Algarve | 3 | 0.002 | 0.017 | 0.098 | 0.000% | 0.002 | 0.019 | 0.100 | 0.000% | 0.004 | 0.032 | 0.120 | 0.000% | 0.009 | 0.065 | 0.190 | 0.000% | |
| PT16 | Centro (PT) | 3 | 0.001 | 0.015 | 0.084 | 0.000% | 0.002 | 0.016 | 0.086 | 0.000% | 0.004 | 0.027 | 0.103 | 0.000% | 0.008 | 0.056 | 0.163 | 0.000% | |
| PT17 | Área Metropolitana de Lisboa | 3 | 0.005 | 0.057 | 0.325 | 0.000% | 0.007 | 0.062 | 0.333 | 0.000% | 0.014 | 0.106 | 0.399 | 0.000% | 0.031 | 0.217 | 0.632 | 0.000% | |
| PT18 | Alentejo | 3 | 0.002 | 0.016 | 0.095 | 0.000% | 0.002 | 0.018 | 0.097 | 0.000% | 0.004 | 0.031 | 0.116 | 0.000% | 0.009 | 0.063 | 0.184 | 0.000% | |
| France | FR | France | |||||||||||||||||
| FRG0 | Pays‐de‐la‐Loire | 1 | 0.013 | 0.119 | 0.570 | 0.000% | 0.014 | 0.126 | 0.578 | 0.000% | 0.021 | 0.159 | 0.627 | 0.000% | 0.048 | 0.329 | 0.937 | 0.004% | |
| FRH0 | Bretagne | 1 | 0.011 | 0.107 | 0.508 | 0.000% | 0.013 | 0.112 | 0.515 | 0.000% | 0.019 | 0.141 | 0.559 | 0.000% | 0.043 | 0.294 | 0.836 | 0.000% | |
| FRI1 | Aquitaine | 1 | 0.008 | 0.077 | 0.368 | 0.000% | 0.009 | 0.081 | 0.374 | 0.000% | 0.014 | 0.103 | 0.406 | 0.000% | 0.031 | 0.213 | 0.606 | 0.000% | |
| FRI3 | Poitou‐Charentes | 1 | 0.010 | 0.098 | 0.467 | 0.000% | 0.012 | 0.103 | 0.474 | 0.000% | 0.017 | 0.130 | 0.514 | 0.000% | 0.040 | 0.270 | 0.768 | 0.000% | |
| FRJ1 | Languedoc‐Roussillon | 1 | 0.017 | 0.161 | 0.768 | 0.003% | 0.019 | 0.170 | 0.779 | 0.004% | 0.028 | 0.214 | 0.845 | 0.005% | 0.065 | 0.444 | 1.264 | 0.280% | |
| FRJ2 | Midi‐Pyrénées | 1 | 0.011 | 0.107 | 0.510 | 0.000% | 0.013 | 0.113 | 0.518 | 0.000% | 0.019 | 0.142 | 0.562 | 0.000% | 0.043 | 0.295 | 0.840 | 0.000% | |
| FRK2 | Rhône‐Alpes | 1 | 0.018 | 0.175 | 0.832 | 0.006% | 0.021 | 0.184 | 0.845 | 0.007% | 0.031 | 0.232 | 0.916 | 0.017% | 0.070 | 0.481 | 1.370 | 0.570% | |
| FRL0 | Provence‐Alpes‐Côte d'Azur | 1 | 0.016 | 0.151 | 0.722 | 0.002% | 0.018 | 0.160 | 0.732 | 0.002% | 0.027 | 0.201 | 0.794 | 0.003% | 0.061 | 0.417 | 1.188 | 0.146% | |
| FRM0 | Corse | 1 | 0.029 | 0.277 | 1.322 | 0.859% | 0.033 | 0.292 | 1.341 | 0.929% | 0.049 | 0.368 | 1.455 | 1.371% | 0.112 | 0.764 | 2.175 | 7.192% | |
| Italy & Malta | ITMT | Italy and Malta | |||||||||||||||||
| ITC1 | Piemonte | 1 | 0.009 | 0.088 | 0.417 | 0.000% | 0.010 | 0.092 | 0.424 | 0.000% | 0.015 | 0.114 | 0.456 | 0.000% | 0.033 | 0.231 | 0.661 | 0.000% | |
| ITC3 | Liguria | 1 | 0.025 | 0.251 | 1.192 | 0.482% | 0.029 | 0.263 | 1.211 | 0.548% | 0.042 | 0.325 | 1.302 | 0.755% | 0.094 | 0.659 | 1.889 | 3.889% | |
| ITC4 | Lombardia | 1 | 0.019 | 0.193 | 0.918 | 0.014% | 0.022 | 0.202 | 0.933 | 0.027% | 0.033 | 0.250 | 1.003 | 0.066% | 0.073 | 0.508 | 1.455 | 0.890% | |
| ITF1 | Abruzzo | 1 | 0.014 | 0.135 | 0.642 | 0.000% | 0.015 | 0.142 | 0.653 | 0.000% | 0.023 | 0.175 | 0.702 | 0.000% | 0.051 | 0.356 | 1.018 | 0.008% | |
| ITF2 | Molise | 1 | 0.007 | 0.072 | 0.343 | 0.000% | 0.008 | 0.076 | 0.348 | 0.000% | 0.012 | 0.093 | 0.375 | 0.000% | 0.027 | 0.190 | 0.543 | 0.000% | |
| ITF3 | Campania | 2 | 0.023 | 0.225 | 1.067 | 0.209% | 0.027 | 0.242 | 1.092 | 0.255% | 0.047 | 0.346 | 1.248 | 0.531% | 0.104 | 0.712 | 1.950 | 4.395% | |
| ITF4 | Puglia | 2 | 0.020 | 0.194 | 0.919 | 0.014% | 0.024 | 0.208 | 0.940 | 0.029% | 0.041 | 0.298 | 1.074 | 0.131% | 0.089 | 0.612 | 1.678 | 1.914% | |
| ITF5 | Basilicata | 1 | 0.011 | 0.107 | 0.508 | 0.000% | 0.012 | 0.112 | 0.516 | 0.000% | 0.018 | 0.138 | 0.555 | 0.000% | 0.040 | 0.281 | 0.804 | 0.000% | |
| ITF6 | Calabria | 2 | 0.013 | 0.126 | 0.598 | 0.000% | 0.015 | 0.135 | 0.612 | 0.000% | 0.027 | 0.194 | 0.699 | 0.000% | 0.058 | 0.398 | 1.092 | 0.018% | |
| ITG1 | Sicilia | 3 | 0.022 | 0.206 | 0.970 | 0.061% | 0.028 | 0.230 | 1.004 | 0.086% | 0.058 | 0.407 | 1.287 | 0.512% | 0.130 | 0.873 | 2.310 | 9.502% | |
| ITG2 | Sardegna | 2 | 0.014 | 0.141 | 0.668 | 0.000% | 0.017 | 0.151 | 0.684 | 0.000% | 0.030 | 0.217 | 0.781 | 0.001% | 0.065 | 0.445 | 1.221 | 0.103% | |
| ITH2 | Provincia Autonoma di Trento | 1 | 0.016 | 0.161 | 0.767 | 0.002% | 0.018 | 0.169 | 0.780 | 0.002% | 0.027 | 0.209 | 0.838 | 0.003% | 0.061 | 0.425 | 1.216 | 0.172% | |
| ITH3 | Veneto | 1 | 0.012 | 0.120 | 0.572 | 0.000% | 0.014 | 0.126 | 0.581 | 0.000% | 0.020 | 0.156 | 0.625 | 0.000% | 0.045 | 0.317 | 0.907 | 0.002% | |
| ITH4 | Friuli‐Venezia Giulia | 1 | 0.007 | 0.069 | 0.331 | 0.000% | 0.008 | 0.073 | 0.336 | 0.000% | 0.012 | 0.090 | 0.361 | 0.000% | 0.026 | 0.183 | 0.524 | 0.000% | |
| ITH5 | Emilia‐Romagna | 1 | 0.007 | 0.069 | 0.326 | 0.000% | 0.008 | 0.072 | 0.331 | 0.000% | 0.012 | 0.089 | 0.356 | 0.000% | 0.026 | 0.180 | 0.517 | 0.000% | |
| ITI1 | Toscana | 2 | 0.017 | 0.163 | 0.772 | 0.002% | 0.020 | 0.175 | 0.790 | 0.002% | 0.034 | 0.250 | 0.902 | 0.004% | 0.075 | 0.515 | 1.410 | 0.537% | |
| ITI2 | Umbria | 1 | 0.008 | 0.078 | 0.370 | 0.000% | 0.009 | 0.081 | 0.376 | 0.000% | 0.013 | 0.101 | 0.404 | 0.000% | 0.029 | 0.205 | 0.586 | 0.000% | |
| ITI3 | Marche | 1 | 0.013 | 0.126 | 0.598 | 0.000% | 0.014 | 0.132 | 0.608 | 0.000% | 0.021 | 0.163 | 0.654 | 0.000% | 0.047 | 0.331 | 0.949 | 0.004% | |
| ITI4 | Lazio | 2 | 0.016 | 0.152 | 0.721 | 0.001% | 0.019 | 0.163 | 0.738 | 0.001% | 0.032 | 0.234 | 0.843 | 0.003% | 0.070 | 0.481 | 1.317 | 0.294% | |
| MT00 | Malta | 2 | 0.026 | 0.247 | 1.168 | 0.403% | 0.031 | 0.265 | 1.194 | 0.467% | 0.053 | 0.380 | 1.364 | 0.906% | 0.114 | 0.778 | 2.131 | 6.571% | |
| Slovenia | SI | Slovenia | |||||||||||||||||
| SI03 | Vzhodna Slovenija | 1 | 0.007 | 0.073 | 0.346 | 0.000% | 0.008 | 0.077 | 0.352 | 0.000% | 0.012 | 0.097 | 0.384 | 0.000% | 0.029 | 0.202 | 0.575 | 0.000% | |
| SI04 | Zahodna Slovenija | 1 | 0.012 | 0.124 | 0.591 | 0.000% | 0.014 | 0.131 | 0.601 | 0.000% | 0.021 | 0.166 | 0.654 | 0.000% | 0.050 | 0.345 | 0.981 | 0.002% | |
| Croatia | HR | Croatia | |||||||||||||||||
| HR02 | Pannonian Croatia | 1 | 0.006 | 0.066 | 0.337 | 0.000% | 0.007 | 0.070 | 0.343 | 0.000% | 0.011 | 0.091 | 0.377 | 0.000% | 0.024 | 0.187 | 0.608 | 0.000% | |
| HR03 | Adriatic Croatia | 1 | 0.007 | 0.079 | 0.403 | 0.000% | 0.008 | 0.084 | 0.410 | 0.000% | 0.013 | 0.108 | 0.451 | 0.000% | 0.029 | 0.223 | 0.727 | 0.000% | |
| HR05 | City of Zagreb | 1 | 0.008 | 0.089 | 0.450 | 0.000% | 0.009 | 0.094 | 0.457 | 0.000% | 0.015 | 0.121 | 0.503 | 0.000% | 0.033 | 0.249 | 0.812 | 0.001% | |
| HR06 | Northern Croatia | 1 | 0.003 | 0.033 | 0.167 | 0.000% | 0.004 | 0.035 | 0.170 | 0.000% | 0.005 | 0.045 | 0.187 | 0.000% | 0.012 | 0.093 | 0.302 | 0.000% | |
| Greece & Cyprus | Greece and Cyprus | ||||||||||||||||||
| EL30 | Attiki | 3 | 0.022 | 0.108 | 0.465 | 0.000% | 0.025 | 0.119 | 0.483 | 0.000% | 0.038 | 0.196 | 0.618 | 0.000% | 0.068 | 0.392 | 1.076 | 0.010% | |
| EL41 | Voreio Aigaio | 2 | 0.007 | 0.039 | 0.174 | 0.000% | 0.008 | 0.042 | 0.178 | 0.000% | 0.010 | 0.058 | 0.205 | 0.000% | 0.019 | 0.111 | 0.320 | 0.000% | |
| EL42 | Notio Aigaio | 2 | 0.003 | 0.017 | 0.077 | 0.000% | 0.003 | 0.019 | 0.079 | 0.000% | 0.005 | 0.026 | 0.091 | 0.000% | 0.008 | 0.049 | 0.142 | 0.000% | |
| EL43 | Kriti | 3 | 0.013 | 0.065 | 0.281 | 0.000% | 0.015 | 0.072 | 0.291 | 0.000% | 0.023 | 0.118 | 0.373 | 0.000% | 0.041 | 0.236 | 0.650 | 0.000% | |
| EL51 | Anatoliki Makedonia, Thraki | 1 | 0.004 | 0.029 | 0.132 | 0.000% | 0.005 | 0.030 | 0.134 | 0.000% | 0.006 | 0.037 | 0.145 | 0.000% | 0.011 | 0.071 | 0.215 | 0.000% | |
| EL52 | Kentriki Makedonia | 1 | 0.011 | 0.072 | 0.331 | 0.000% | 0.012 | 0.076 | 0.335 | 0.000% | 0.015 | 0.093 | 0.364 | 0.000% | 0.028 | 0.178 | 0.540 | 0.000% | |
| EL53 | Dytiki Makedonia | 1 | 0.007 | 0.046 | 0.210 | 0.000% | 0.008 | 0.048 | 0.212 | 0.000% | 0.010 | 0.059 | 0.230 | 0.000% | 0.018 | 0.113 | 0.341 | 0.000% | |
| EL54 | Ipeiros | 2 | 0.013 | 0.072 | 0.321 | 0.000% | 0.014 | 0.077 | 0.329 | 0.000% | 0.019 | 0.106 | 0.378 | 0.000% | 0.034 | 0.205 | 0.590 | 0.000% | |
| EL61 | Thessalia | 1 | 0.007 | 0.044 | 0.199 | 0.000% | 0.007 | 0.046 | 0.202 | 0.000% | 0.009 | 0.056 | 0.219 | 0.000% | 0.017 | 0.107 | 0.325 | 0.000% | |
| EL62 | Ionia Nisia | 2 | 0.004 | 0.023 | 0.103 | 0.000% | 0.004 | 0.025 | 0.105 | 0.000% | 0.006 | 0.034 | 0.121 | 0.000% | 0.011 | 0.066 | 0.189 | 0.000% | |
| EL63 | Dytiki Ellada | 2 | 0.011 | 0.064 | 0.284 | 0.000% | 0.012 | 0.068 | 0.290 | 0.000% | 0.017 | 0.094 | 0.334 | 0.000% | 0.030 | 0.181 | 0.521 | 0.000% | |
| EL64 | Sterea Ellada | 2 | 0.007 | 0.039 | 0.172 | 0.000% | 0.007 | 0.041 | 0.176 | 0.000% | 0.010 | 0.057 | 0.203 | 0.000% | 0.018 | 0.110 | 0.316 | 0.000% | |
| EL65 | Peloponnisos | 3 | 0.006 | 0.029 | 0.123 | 0.000% | 0.007 | 0.031 | 0.128 | 0.000% | 0.010 | 0.052 | 0.164 | 0.000% | 0.018 | 0.104 | 0.285 | 0.000% | |
| CY00 | Kypros | 2 | 0.006 | 0.035 | 0.157 | 0.000% | 0.007 | 0.038 | 0.160 | 0.000% | 0.009 | 0.052 | 0.185 | 0.000% | 0.017 | 0.100 | 0.288 | 0.000% | |
| Hungary | HU | Hungary | |||||||||||||||||
| HU12 | Pest | 1 | 0.005 | 0.052 | 0.252 | 0.000% | 0.006 | 0.055 | 0.256 | 0.000% | 0.009 | 0.071 | 0.279 | 0.000% | 0.022 | 0.149 | 0.429 | 0.000% | |
| HU23 | Dél‐Dunántúl | 1 | 0.003 | 0.033 | 0.159 | 0.000% | 0.004 | 0.035 | 0.162 | 0.000% | 0.006 | 0.045 | 0.176 | 0.000% | 0.014 | 0.095 | 0.271 | 0.000% | |
| HU31 | Észak‐Magyarország | 1 | 0.005 | 0.051 | 0.245 | 0.000% | 0.006 | 0.054 | 0.249 | 0.000% | 0.009 | 0.069 | 0.271 | 0.000% | 0.021 | 0.145 | 0.417 | 0.000% | |
| HU32 | Észak‐Alföld | 1 | 0.005 | 0.053 | 0.255 | 0.000% | 0.006 | 0.056 | 0.260 | 0.000% | 0.009 | 0.072 | 0.283 | 0.000% | 0.022 | 0.152 | 0.435 | 0.000% | |
| HU33 | Dél‐Alföld | 1 | 0.003 | 0.030 | 0.143 | 0.000% | 0.003 | 0.031 | 0.146 | 0.000% | 0.005 | 0.040 | 0.159 | 0.000% | 0.012 | 0.085 | 0.244 | 0.000% | |
| Bulgaria | BG | Bulgaria | |||||||||||||||||
| BG31 | Severozapaden | 1 | 0.003 | 0.021 | 0.100 | 0.000% | 0.003 | 0.022 | 0.102 | 0.000% | 0.004 | 0.028 | 0.112 | 0.000% | 0.008 | 0.055 | 0.179 | 0.000% | |
| BG32 | Severen tsentralen | 1 | 0.004 | 0.031 | 0.150 | 0.000% | 0.004 | 0.033 | 0.152 | 0.000% | 0.006 | 0.041 | 0.168 | 0.000% | 0.012 | 0.083 | 0.268 | 0.000% | |
| BG33 | Severoiztochen | 1 | 0.002 | 0.015 | 0.074 | 0.000% | 0.002 | 0.016 | 0.075 | 0.000% | 0.003 | 0.020 | 0.083 | 0.000% | 0.006 | 0.041 | 0.132 | 0.000% | |
| BG34 | Yugoiztochen | 1 | 0.006 | 0.045 | 0.218 | 0.000% | 0.006 | 0.048 | 0.222 | 0.000% | 0.009 | 0.060 | 0.244 | 0.000% | 0.018 | 0.121 | 0.391 | 0.000% | |
| BG41 | Yugozapaden | 1 | 0.004 | 0.032 | 0.155 | 0.000% | 0.005 | 0.034 | 0.158 | 0.000% | 0.006 | 0.043 | 0.173 | 0.000% | 0.013 | 0.086 | 0.277 | 0.000% | |
| BG42 | Yuzhen tsentralen | 1 | 0.004 | 0.029 | 0.138 | 0.000% | 0.004 | 0.030 | 0.140 | 0.000% | 0.006 | 0.038 | 0.154 | 0.000% | 0.011 | 0.077 | 0.247 | 0.000% | |
| Romania | RO | Romania | |||||||||||||||||
| RO11 | Nord‐Vest | 1 | 0.008 | 0.082 | 0.431 | 0.000% | 0.009 | 0.087 | 0.441 | 0.000% | 0.014 | 0.113 | 0.489 | 0.000% | 0.028 | 0.231 | 0.819 | 0.003% | |
| RO22 | Sud‐Est | 1 | 0.009 | 0.093 | 0.493 | 0.000% | 0.010 | 0.099 | 0.504 | 0.000% | 0.015 | 0.130 | 0.559 | 0.000% | 0.032 | 0.265 | 0.936 | 0.051% | |
| RO31 | Sud ‐ Muntenia | 1 | 0.007 | 0.069 | 0.365 | 0.000% | 0.008 | 0.073 | 0.373 | 0.000% | 0.011 | 0.096 | 0.413 | 0.000% | 0.024 | 0.196 | 0.692 | 0.000% | |
| RO32 | Bucuresti – Ilfov | 1 | 0.018 | 0.179 | 0.944 | 0.204% | 0.020 | 0.190 | 0.966 | 0.221% | 0.030 | 0.248 | 1.071 | 0.352% | 0.062 | 0.507 | 1.793 | 3.266% | |
| RO41 | Sud‐Vest Oltenia | 1 | 0.005 | 0.048 | 0.253 | 0.000% | 0.005 | 0.051 | 0.259 | 0.000% | 0.008 | 0.067 | 0.287 | 0.000% | 0.017 | 0.136 | 0.480 | 0.000% | |
| RO42 | Vest | 1 | 0.005 | 0.049 | 0.259 | 0.000% | 0.005 | 0.052 | 0.265 | 0.000% | 0.008 | 0.068 | 0.294 | 0.000% | 0.017 | 0.139 | 0.492 | 0.000% | |
Table A.5.
Estimated number of T. leucotreta adults escaping from disposed infested cut roses in a 3.14‐km2 circle per decade in the residential areas of some example NUTS 2 regions
| NUTS2 region | Time between disposal and waste treatment | Winter | Spring | Summer | Autumn | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 5th perc | Median | 95th perc | 5th perc | Median | 95th perc | 5th perc | Median | 95th perc | 5th perc | Median | 95th perc | ||
| Andalusia | 3 days | 0.000 | 0.033 | 0.467 | 0.001 | 0.035 | 0.298 | 0.003 | 0.027 | 0.128 | 0.003 | 0.032 | 0.155 |
| 7 days | 0.000 | 0.033 | 0.468 | 0.001 | 0.037 | 0.303 | 0.005 | 0.036 | 0.144 | 0.005 | 0.038 | 0.164 | |
| 14 days | 0.000 | 0.034 | 0.474 | 0.001 | 0.050 | 0.339 | 0.016 | 0.109 | 0.307 | 0.010 | 0.067 | 0.217 | |
| 28 days | 0.000 | 0.044 | 0.516 | 0.002 | 0.116 | 0.602 | 0.031 | 0.206 | 0.571 | 0.034 | 0.226 | 0.624 | |
| Corse | 3 days | 0.028 | 0.280 | 1.351 | 0.032 | 0.317 | 1.583 | 0.030 | 0.245 | 1.152 | 0.025 | 0.280 | 1.351 |
| 7 days | 0.028 | 0.280 | 1.351 | 0.032 | 0.319 | 1.585 | 0.042 | 0.301 | 1.240 | 0.026 | 0.280 | 1.351 | |
| 14 days | 0.028 | 0.280 | 1.351 | 0.034 | 0.326 | 1.597 | 0.087 | 0.571 | 1.730 | 0.029 | 0.280 | 1.351 | |
| 28 days | 0.028 | 0.280 | 1.351 | 0.047 | 0.384 | 1.688 | 0.281 | 1.861 | 4.963 | 0.047 | 0.280 | 1.351 | |
| Liguria | 3 days | 0.028 | 0.279 | 1.335 | 0.026 | 0.265 | 1.296 | 0.021 | 0.197 | 0.922 | 0.025 | 0.253 | 1.215 |
| 7 days | 0.028 | 0.279 | 1.335 | 0.027 | 0.267 | 1.298 | 0.031 | 0.242 | 0.992 | 0.026 | 0.255 | 1.220 | |
| 14 days | 0.028 | 0.279 | 1.335 | 0.028 | 0.273 | 1.307 | 0.068 | 0.465 | 1.388 | 0.068 | 0.465 | 1.388 | |
| 28 days | 0.028 | 0.279 | 1.335 | 0.040 | 0.323 | 1.384 | 0.229 | 1.533 | 3.993 | 0.229 | 1.533 | 3.993 | |
| Sicily | 3 days | 0.023 | 0.226 | 1.082 | 0.022 | 0.216 | 1.052 | 0.019 | 0.164 | 0.754 | 0.023 | 0.211 | 0.993 |
| 7 days | 0.023 | 0.226 | 1.082 | 0.023 | 0.223 | 1.061 | 0.030 | 0.220 | 0.843 | 0.031 | 0.243 | 1.048 | |
| 14 days | 0.023 | 0.229 | 1.087 | 0.033 | 0.267 | 1.131 | 0.100 | 0.671 | 1.787 | 0.059 | 0.405 | 1.313 | |
| 28 days | 0.027 | 0.244 | 1.111 | 0.068 | 0.477 | 1.502 | 0.192 | 1.282 | 3.337 | 0.209 | 1.389 | 3.665 | |
| Malta | 3 days | 0.028 | 0.274 | 1.306 | 0.027 | 0.261 | 1.268 | 0.022 | 0.195 | 0.905 | 0.026 | 0.251 | 1.192 |
| 7 days | 0.028 | 0.274 | 1.306 | 0.027 | 0.265 | 1.274 | 0.033 | 0.250 | 0.993 | 0.029 | 0.263 | 1.213 | |
| 14 days | 0.028 | 0.274 | 1.306 | 0.033 | 0.286 | 1.307 | 0.086 | 0.585 | 1.627 | 0.045 | 0.336 | 1.328 | |
| 28 days | 0.029 | 0.275 | 1.309 | 0.054 | 0.398 | 1.487 | 0.231 | 1.543 | 4.016 | 0.113 | 0.771 | 2.151 | |
Table A.6.
African countries and Israel with their status of Thaumatotibia leucotreta (in EPPO Global Database, Internet: https://gd.eppo.int/taxon/ARGPLE/distribution, accessed: 1 June 2022; Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, online, accessed on 4 October 2022)
| Country | ISO Country code | Export of cut roses in 2011–2020 (Eurostat) | Status of T. leucotreta (EPPO) |
|---|---|---|---|
| Israel | IL | Yes | Present, restricted distribution |
| Angola | AO | Yes | Present, no details |
| Burkina Faso | BF | No | Present, no details |
| Burundi | BI | No | Present, no details |
| Benin | BJ | No | Present, no details |
| Rep.Democr. Congo | CD | No | Present, no details |
| Central African Republic | CF | No | Present, no details |
| Ivory Coast | CI | Yes | Present, no details |
| Cameroon | CM | No | Present, no details |
| Cape Verde | CV | No | Present, no details |
| Eritrea | ER | Yes | Present, no details |
| Ethiopia | ET | Yes | Present, no details |
| Ghana | GH | Yes | Present, no details |
| Gambia | GM | No | Present, no details |
| Kenya | KE | Yes | Present, no details |
| Madagascar | MG | No | Present, no details |
| Mali | ML | No | Present, no details |
| Mauritius | MU | Yes | Present, no details |
| Malawi | MW | Yes | Present, no details |
| Mozambique | MZ | No | Present, no details |
| Niger | NE | Yes | Present, no details |
| Nigeria | NG | Yes | Present, no details |
| Rwanda | RW | Yes | Present, no details |
| Sudan | SD | No | Present, no details |
| Sierra Leone | SL | No | Present, no details |
| Senegal | SN | No | Present, no details |
| Somalia | SO | No | Present, no details |
| Eswatini | SZ | Yes | Present, no details |
| Chad | TD | No | Present, no details |
| Togo | TG | No | Present, no details |
| Tanzania | TZ | Yes | Present, no details |
| Uganda | UG | Yes | Present, no details |
| South Africa | ZA | Yes | Present, no details |
| Zambia | ZM | Yes | Present, no details |
| Zimbabwe | ZW | Yes | Present, no details |
| Botswana | BW | No | No notification |
| Congo | CG | No | No notification |
| Algeria | DZ | No | No notification |
| Egypt | EG | Yes | No notification |
| Gabon | GA | No | No notification |
| Guinea | GN | No | No notification |
| Equatorial Guinea | GQ | No | No notification |
| Guinea‐Bissau | GW | No | No notification |
| Comoros | KM | No | No notification |
| Liberia | LR | No | No notification |
| Lesotho | LS | No | No notification |
| Libya | LY | No | No notification |
| Morocco | MA | Yes | No notification |
| Mauritania | MR | No | No notification |
| Namibia | NA | Yes | No notification |
| Seychelles | SC | No | No notification |
| South Sudan | SS | No | No notification |
| Sao Tome and Principe | ST | No | No notification |
| Tunisia | TN | Yes | No notification |
Table A.7.
Average export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to EU ordered by the average annual volume [in pcs] (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, online, accessed on 4 October 2022)
| Country | Average export 2011–2020 to Europe (EU27) | |
|---|---|---|
| Absolute (roses, pcs) | Relative (%) | |
| Total (selected countries of Africa and Israel) | 4,411,137,386 | 100% |
| Kenya | 2,406,079,285 | 54.55% |
| Ethiopia | 1,486,976,887 | 33.71% |
| Uganda | 238,429,156 | 5.41% |
| Zambia | 150,072,433 | 3.40% |
| Zimbabwe | 71,434,840 | 1.62% |
| Tanzania | 48,774,537 | 1.11% |
| Rwanda | 8,420,765 | 0.19% |
| South Africa | 492,771 | 0.01% |
| Morocco | 252,006 | 0.01% |
| Israel | 127,714 | 0.00% |
| Eritrea | 50,080 | 0.00% |
| Malawi | 16,898 | 0.00% |
| Mauritius | 4,796 | 0.00% |
| Côte d'Ivoire | 1,484 | 0.00% |
| Egypt | 1,085 | 0.00% |
| Nigeria | 1,038 | 0.00% |
| Niger | 558 | 0.00% |
| Namibia | 528 | 0.00% |
| Eswatini | 452 | 0.00% |
| Tunisia | 52 | 0.00% |
| Angola | 20 | 0.00% |
| Ghana | 1 | 0.00% |
| Burkina Faso | 0 | 0.00% |
| Burundi | 0 | 0.00% |
| Benin | 0 | 0.00% |
| Congo, Democratic Republic of | 0 | 0.00% |
| Central African Republic | 0 | 0.00% |
| Cameroon | 0 | 0.00% |
| Cabo Verde | 0 | 0.00% |
| Gambia | 0 | 0.00% |
| Madagascar | 0 | 0.00% |
| Mali | 0 | 0.00% |
| Mozambique | 0 | 0.00% |
| Sudan) | 0 | 0.00% |
| Sierra Leone | 0 | 0.00% |
| Senegal | 0 | 0.00% |
| Somalia | 0 | 0.00% |
| Chad | 0 | 0.00% |
| Togo | 0 | 0.00% |
Table A.8.
Definition of climate suitability classes in the pathway model using quartiles of the climate suitability index (number of pupae per year per NUTS2 region) in the physiologically based demographic model (PBDM, see Appendix B)
| No. pupae/year per NUTS2 region | Climate suitability class | Interpretation | Colour |
|---|---|---|---|
| 0 | 0 | Not suitable | |
| 1 ≤ × < 128 | 1 |
Low climate suitability
High climate suitability |
|
| 128 ≤ × < 255 | 2 | ||
| 255 ≤ × < 382 | 3 | ||
| 382 ≤ × < 509 | 4 |
Table A.9.
Average annual import of cut roses from African countries with reported presence of FCM and from Israel to EU27 during 2011–2020 by importing country (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, online, accessed on 4 October 2022)
| Country | Average annual import of cut roses from African countries with reported presence of FCM and from Israel to EU27 | |
|---|---|---|
| Absolute (roses, pcs) | Relative (%) | |
| Total (selected countries of Africa and Israel) | 4,411,137,386 | 100% |
| The Netherlands | 3,376,807,643 | 76.55% |
| Belgium | 710,493,824 | 16.11% |
| Germany | 276,623,163 | 6.27% |
| Sweden | 23,218,812 | 0.53% |
| France | 12,992,148 | 0.29% |
| Italy | 3,324,573 | 0.08% |
| Cyprus | 2,916,762 | 0.07% |
| Ireland | 1,661,139 | 0.04% |
| Greece | 1,082,122 | 0.02% |
| Croatia | 569,584 | 0.01% |
| Romania | 460,236 | 0.01% |
| Bulgaria | 406,617 | 0.01% |
| Spain | 232,484 | 0.01% |
| Luxembourg | 120,154 | 0.00% |
| Austria | 95,904 | 0.00% |
| Czechia | 74,929 | 0.00% |
| Slovenia | 26,769 | 0.00% |
| Poland | 10,880 | 0.00% |
| Malta | 8,229 | 0.00% |
| Portugal | 5,475 | 0.00% |
| Estonia | 3,168 | 0.00% |
| Denmark | 1,719 | 0.00% |
| Hungary | 672 | 0.00% |
| Lithuania | 206 | 0.00% |
| Finland | 100 | 0.00% |
| Latvia | 77 | 0.00% |
| Slovakia | 0 | 0.00% |
In bold: Countries in the climatically suitable area.
Table A.10.
Definition of seasons used in the temporal stratification of the pathway model.
| Season | Months |
|---|---|
| Winter | December–February |
| Spring | March–May |
| Summer | June–August |
| Autumn | September–November |
Table A.11.
Average direct import of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel into the countries of the EU with areas climatically suitable stratified by season (Eurostat, EU trade since 1988 by HS2‐4‐4‐4‐6 and CN8, CN 06031100, monthly statistics 12‐4‐2010‐4‐12.2020, online, accessed on 25 January 2023)
| Direct export of cut roses from African countries with FCM occurrence and from Israel to the EU areas climatically suitable | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Average seasonal direct trade of cut roses [pcs] | ||||||||||
| Season | Winter | Spring | Summer | Autumn | Annual | |||||
| Importer | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD |
| Spain & Portugal | 106,573 | 51,173 | 101,610 | 52,522 | 15,344 | 16,381 | 78,777 | 44,764 | 305,426 | 83,625 |
| France | 2,960,189 | 1,800,901 | 3,879,806 | 2,364,787 | 2,974,537 | 1,836,849 | 2,902,042 | 2,101,205 | 12,925,290 | 3,869,307 |
| Italy & Malta | 1,407,232 | 1,065,589 | 802,466 | 421,035 | 267,157 | 239,740 | 865,456 | 558,681 | 3,423,925 | 1,210,909 |
| Slovenia | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Croatia | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Greece & Cyprus | 1,109,223 | 312,163 | 846,886 | 368,115 | 914,804 | 549,670 | 1,032,039 | 314,822 | 3,915,333 | 783,485 |
| Hungary | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Bulgaria | 214,184 | 114,673 | 158,978 | 80,027 | 126,039 | 70,582 | 191,025 | 104,772 | 694,758 | 185,065 |
| Romania | 46,622 | 41,802 | 347,955 | 65,466 | 117,178 | 123,854 | 171,172 | 181,550 | 713,979 | 201,319 |
| Sum | 5,844,022 | 6,137,700 | 4,415,060 | 5,240,510 | 21,978,711 | |||||
Table A.12.
Average direct import of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Spain and Portugal stratified by season. Cells marked in grey are not used to calculate the mean and standard deviation (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Direct export of cut roses from African countries with FCM occurrence and from Israel to the EU area climatically suitable | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Season | Importer: Spain & Portugal | |||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 103,236 | 9,600 | 0 | 33,900 | 40,110 | 152,610 | 165,505 | 114,040 | 117,804 | 122,040 | 106,573 | 51,173 |
| Spring | 31,040 | 2,830 | 0 | 113,130 | 141,840 | 148,402 | 49,745 | 130,694 | 119,432 | 8,030 | 101,610 | 52,522 |
| Summer | 0 | 0 | 430 | 1,470 | 3,098 | 6,767 | 37,134 | 36,234 | 22,556 | 150 | 15,344 | 16,381 |
| Autumn | 0 | 340 | 750 | 37,704 | 90,035 | 91,020 | 62,432 | 157,974 | 91,453 | 20,820 | 78,777 | 44,764 |
| Total | 134,276 | 12,770 | 1,180 | 186,204 | 275,083 | 398,799 | 314,816 | 438,942 | 351,245 | 151,040 | ||
Table A.30.
Total and residential areas of NUTS2 region in the climatically suitable area of the EU (Eurostat, table LAN_USE_OVW, year: 2018, online, accessed on 20 February 2023)
| Country | NUTS code | Area | Residential area |
|---|---|---|---|
| (km2) | (km2) | ||
| Spain & Portugal | ESPT | ||
| ES11 | 29,571 | 680 | |
| ES12 | 10,601 | 147 | |
| ES13 | 5,326 | 100 | |
| ES21 | 7,229 | 154 | |
| ES22 | 10,391 | 47 | |
| ES23 | 5,045 | 27 | |
| ES24 | 47,722 | 106 | |
| ES30 | 8,031 | 438 | |
| ES41 | 94,225 | 446 | |
| ES42 | 79,458 | 475 | |
| ES43 | 41,634 | 280 | |
| ES51 | 32,110 | 606 | |
| ES52 | 23,255 | 702 | |
| ES53 | 4,990 | 428 | |
| ES61 | 87,600 | 952 | |
| ES62 | 11,314 | 315 | |
| ES63 | 20 | 20 | |
| ES64 | 14 | 14 | |
| PT11 | 21,287 | 655 | |
| PT15 | 4,997 | 115 | |
| PT16 | 28,200 | 680 | |
| PT17 | 3,015 | 226 | |
| PT18 | 31,604 | 191 |
| France | FR | ||
| FRG0 | 32,376 | 1,998 | |
| FRH0 | 27,471 | 1,971 | |
| FRI1 | 41,804 | 2,814 | |
| FRI3 | 25,967 | 1,158 | |
| FRJ1 | 27,643 | 1,112 | |
| FRJ2 | 45,603 | 1,803 | |
| FRK2 | 44,962 | 2,396 | |
| FRL0 | 31,839 | 2,097 | |
| FRM0 | 8,726 | 78 | |
| Italy & Malta | ITMT | ||
| ITC1 | 25,387 | 1,308 | |
| ITC3 | 5,416 | 162 | |
| ITC4 | 23,864 | 1,383 | |
| ITF1 | 10,833 | 255 | |
| ITF2 | 4,461 | 111 | |
| ITF3 | 13,670 | 679 | |
| ITF4 | 19,541 | 546 | |
| ITF5 | 10,073 | 138 | |
| ITF6 | 15,222 | 402 | |
| ITG1 | 25,833 | 639 | |
| ITG2 | 24,100 | 306 | |
| ITH2 | 6,207 | 90 | |
| ITH3 | 18,407 | 1,080 | |
| ITH4 | 7,862 | 462 | |
| ITH5 | 22,453 | 1,734 | |
| ITI1 | 22,988 | 607 | |
| ITI2 | 8,464 | 298 | |
| ITI3 | 9,401 | 320 | |
| ITI4 | 17,232 | 1,013 | |
| MT00 | 316 | 56 | |
| Slovenia | SI | ||
| SI03 | 12,433 | 275 | |
| SI04 | 7,840 | 146 | |
| Croatia | HR | ||
| HR02 | 23,220 | 297 | |
| HR03 | 24,705 | 316 | |
| HR05 | 2,911 | 167 | |
| HR06 | 8,028 | 461 |
| Greece & Cyprus | |||
| EL30 | 3,817 | 408 | |
| EL41 | 3,848 | 66 | |
| EL42 | 5,311 | 225 | |
| EL43 | 8,346 | 115 | |
| EL51 | 14,186 | 226 | |
| EL52 | 18,838 | 282 | |
| EL53 | 9,462 | 63 | |
| EL54 | 9,152 | 52 | |
| EL61 | 14,050 | 179 | |
| EL62 | 2,301 | 99 | |
| EL63 | 11,314 | 115 | |
| EL64 | 15,560 | 162 | |
| EL65 | 15,509 | 236 | |
| CY00 | 9,253 | 284 | |
| Hungary | HU | ||
| HU12 | 6,391 | 323 | |
| HU23 | 14,197 | 344 | |
| HU31 | 13,426 | 286 | |
| HU32 | 17,723 | 354 | |
| HU33 | 18,336 | 538 | |
| Bulgaria | BG | ||
| BG31 | 19,068 | 185 | |
| BG32 | 14,812 | 131 | |
| BG33 | 14,648 | 317 | |
| BG34 | 19,801 | 119 | |
| BG41 | 20,301 | 343 | |
| BG42 | 22,366 | 259 | |
| Romania | RO | ||
| RO11 | 34,160 | 435 | |
| RO22 | 35,774 | 355 | |
| RO31 | 34,467 | 586 | |
| RO32 | 1,804 | 181 | |
| RO41 | 29,206 | 556 | |
| RO42 | 32,042 | 503 |
Table A.31.
Average re‐export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel into the countries of the EU with areas climatically suitable via the Netherlands and Belgium stratified by season. (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Export of cut roses from the Netherlands and Belgium to the EU area climatically suitable | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Average seasonal trade of cut roses [pcs] | ||||||||||
| Season | Winter | Spring | Summer | Autumn | Annual | |||||
| Importer | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD |
| Spain & Portugal | 9,456,896 | 15,986,293 | 7,960,316 | 7,132,380 | 4,474,421 | 830,011 | 5,454,526 | 1,044,759 | 30,554,150 | 13,884,949 |
| France | 112,100,482 | 12,025,162 | 128,525,842 | 26,247,886 | 93,037,573 | 13,036,657 | 99,774,668 | 11,094,109 | 433,439,048 | 33,492,424 |
| Italy & Malta | 46,911,375 | 4,777,377 | 45,665,516 | 7,625,087 | 32,624,136 | 3,111,446 | 42,758,330 | 4,478,807 | 167,958,860 | 10,451,179 |
| Slovenia | 921,748 | 201,786 | 1,212,569 | 158,102 | 871,427 | 84,731 | 1,098,204 | 137,442 | 4,103,953 | 302,651 |
| Croatia | 1,788,736 | 1,122,441 | 2,455,304 | 1,379,491 | 1,905,345 | 903,247 | 2,139,576 | 1,068,565 | 8,350,921 | 21,78,469 |
| Greece & Cyprus | 3,057,764 | 796,735 | 2,151,447 | 761,785 | 1,461,848 | 632,963 | 1,741,536 | 769,181 | 8,418,517 | 1,465,958 |
| Hungary | 3,372,167 | 718,415 | 4,101,544 | 1,031,021 | 3,096,897 | 497,899 | 3,206,296 | 530,643 | 13,776,885 | 1,449,223 |
| Bulgaria | 714,168 | 223,717 | 823,484 | 310,909 | 718,250 | 397,692 | 835,305 | 474,415 | 3,104,691 | 713,977 |
| Romania | 5,678,353 | 4,218,837 | 8,493,255 | 6,079,992 | 7,144,377 | 4,528,549 | 7,864,685 | 5,354,353 | 29,858,426 | 9,568,094 |
| Sum | 184,001,688 | 201,389,277 | 145,334,275 | 164,873,126 | 699,565,450 | |||||
Table A.44.
Production surface of cut roses in the Netherlands for specific years (Dutch CBS: opendata.cbs.nl/statline/ keyword: ‘rozen’, online, accessed on 21 March 2023)
| Area of production of cut roses in the Netherlands | ||||
|---|---|---|---|---|
| Unit | (ha) | |||
| Year | 2010 | 2015 | 2020 | Mean |
| Total | 501 | 286 | 217 | 334 |
Table A.45.
Production surface of flowers and ornamentals of Belgium in comparison to the Netherlands for specific years (Eurostat: Crops by classes of utilised agricultural area [EF_LUS_ALLCROPS], online, accessed on 1 February 2023)
| Area of production of flowers and ornamentals | |||
|---|---|---|---|
| Unit | (ha) | ||
| Year | 2013 | 2016 | Mean |
| The Netherlands | 30,590 | 32,630 | 31,610 |
| Belgium | 1,510 | 5,270 | 3,390 |
| Factor: Sum/NL | 105% | 116% | 111% |
Table A.46.
Calculation of the dilution factor in the Dutch/Belgian cluster using two scenarios (with or without intra‐European trade)
| Dilution of cut roses from African countries with occurrence of FCM and from Israel by other import and own production | ||||||||
|---|---|---|---|---|---|---|---|---|
| Season | Scenario 1 | Scenario 2 | ||||||
| Import from African countries with FCM and Israel | Other import (World+EU27) | Own production | Low dilution factor | Import from African countries with FCM and Israel | Other import (World, not EU) | Own production | High dilution factor | |
| a | b1 | c | a/(a + b1 + c) | a | b2 | c | a/(a + b2 + c) | |
| Winter | 982,765,878 | 208,140,149 | 30,565,283 | 80% | 982,765,878 | 70,841,982 | 30,565,283 | 91% |
| Spring | 1,133,976,961 | 264,543,637 | 30,565,283 | 79% | 1,133,976,961 | 67,516,387 | 30,565,283 | 92% |
| Summer | 899,871,296 | 184,295,721 | 30,565,283 | 81% | 899,871,296 | 53,392,079 | 30,565,283 | 91% |
| Autumn | 1,017,821,932 | 217,535,357 | 30,565,283 | 80% | 1,017,821,932 | 71,917,058 | 30,565,283 | 91% |
| Total | 4,034,436,066 | 874,514,864 | 122,261,130 | 4,034,436,066 | 263,667,505 | 122,261,130 | ||
Table A.47.
Average re‐export (intra‐EU trade) of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel into the countries of the EU with areas climatically suitable via Germany and Luxembourg stratified by season (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses from Germany and Luxembourg to the EU area climatically suitable | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Average seasonal trade of cut roses (pcs) | ||||||||||
| Winter | Spring | Summer | Autumn | Annual | ||||||
| Importer | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD |
| Spain & Portugal | 0 | 0 | 167,808 | 97,037 | 0 | 0 | 0 | 0 | 169,461 | 93,508 |
| France | 175,465 | 187,776 | 235,545 | 242,386 | 131,374 | 142,615 | 567,938 | 1,284,961 | 1,440,994 | 979,315 |
| Italy & Malta | 155,077 | 250,723 | 177,537 | 298,559 | 123,696 | 302,040 | 157,018 | 370,555 | 856,180 | 454,646 |
| Slovenia | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Croatia | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Greece & Cyprus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hungary | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Bulgaria | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Romania | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sum | 330,542 | 580,891 | 255,071 | 724,956 | 2,466,636 | |||||
Table A.60.
Production surface of cut roses in Germany for specific years (DEStatis: Zierpflanzenanbau Report 2021, online, accessed on 1 February 2023)
| Area of production of cut roses in Germany | ||||
|---|---|---|---|---|
| Unit | (ha) | |||
| Year | 2012 | 2017 | 2021 | Mean |
| Total | 354 | 323 | 234 | 304 |
Table A.61.
Production surface of flowers and ornamentals of Luxembourg in comparison to Germany for specific years (Eurostat: Crops by classes of utilised agricultural area [EF_LUS_ALLCROPS], online, accessed on 1 February 2023)
| Area of production of flowers and ornamentals | |||
|---|---|---|---|
| Unit | [ha] | ||
| Year | 2013 | 2016 | Mean |
| Germany | 7,710 | 7,640 | 7,675 |
| Luxembourg | 0 | 0 | |
| Factor: Sum/NL | 100% | 100% | 100% |
Table A.62.
Calculation of the dilution factor in the German/Luxembourg cluster using two scenarios (with or without intra‐European trade)
| Season | Dilution of cut roses from African countries with occurrence of FCM and Israel by other import and own production | |||||||
|---|---|---|---|---|---|---|---|---|
| Scenario 1 | Scenario 2 | |||||||
| Import from African countries with FCM and Israel | Other import (World+EU27) | Own production | Low dilution factor | Import from African countries with FCM and Israel | Other import (World, not EU) | Own production | High dilution factor | |
| a | b1 | c | a/(a + b1 + c) | a | b2 | c | a/(a + b2 + c) | |
| Winter | 63,232,315 | 246,529,799 | 15,183,333 | 19% | 63,232,315 | 5,533,512 | 15,183,333 | 75% |
| Spring | 75,084,845 | 310,911,069 | 15,183,333 | 19% | 75,084,845 | 5,285,530 | 15,183,333 | 79% |
| Summer | 66,759,404 | 278,238,726 | 15,183,333 | 19% | 66,759,404 | 3,769,106 | 15,183,333 | 78% |
| Autumn | 67,821,871 | 274,707,989 | 15,183,333 | 19% | 67,821,871 | 4,823,631 | 15,183,333 | 77% |
| Total | 272,898,435 | 1,110,387,583 | 60,733,333 | 272,898,435 | 19,411,779 | 60,733,333 | ||
Table A.63.
Life‐stages of intercepted specimens of Thaumatotibia leucotreta on cut roses as reported by the Dutch NPPO in the years 2015–2022 (NVWA, 2022: Development stage of intercepted Thaumatotibia leucotreta specimens; personal communication, 12 October 2022)
| Intercepted life‐stage | Roses (2015–2018) | Roses (2019–2022) |
|---|---|---|
| (n = 364) | (n = 190) | |
| Eggs | 54 (15%) | 95 (50%) |
| L1 | 131 (36%) | 28 (15%) |
| L1 or L2 | – | 33 (17%) |
| L2 | 85 (23%) | 19 (10%) |
| L3 | 49 (13%) | 7 (4%) |
| L4 | 33 (9%) | 5 (3%) |
| L5 | 12 (3%) | 3 (2%) |
Table A.64.
Life‐stages of intercepted specimens of Thaumatotibia leucotreta on cut roses as reported by the Dutch NPPO in the years 2019–2022 from selected African countries (NVWA, 2022: Development stage of intercepted Thaumatotibia leucotreta specimens; personal communication, 12 October 2022)
| Intercepted life‐stage | Ethiopia (2019–2022) | Kenya (2019–2022) | Uganda (2019–2022) |
|---|---|---|---|
| On cut roses | (n = 38) | (n = 36) | (n = 88) |
| Eggs | 8 (21%) | 24 (67%) | 41 (47%) |
| L1 | 3 (8%) | 5 (13%) | 18 (20%) |
| L1 or L2 | 25 (66%) | 4 (11%) | 3 (3%) |
| L2 | – | 2 (6%) | 15 (17%) |
| L3 | 2 (5%) | – | 5 (6%) |
| L4 | – | 1 (3%) | 3 (3%) |
| L5 | – | – | 3 (3%) |
Table A.65.
Development time of FCM immatures developmental stages in artificial medium under different temperatures (Daiber 1979a,b,c, 1989). Proportion of eggs and portion of young larvae out of all larvae are calculated by EFSA
| Temperature | FCM immatures developmental stages | Sum of different periods | Proportion of eggs | Proportion of young larvae (L1 + L2) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Egg | Larval | Pupae | Egg to pupa | Egg to larva | Larva to pupa | Rel. to E to P | Rel. to E to L | Rel. to L to P | Rel. to L | |||||||
| [°C] | Development duration [d] | [%] | ||||||||||||||
| E | L (Total) | L1 | L1 + L2 | L2 | L3 | L4 | L5 | P | E + L + P | E + L | L + P | E/(E + L + P) | E/(E + L) | 0.4 L/(L + P) | ||
| 10.9 | 22 | assumed as 40% | ||||||||||||||
| 12.1 | 61 | 68 | 129 | 19% | ||||||||||||
| 15 | 45.6 | 45 | 90.6 | 20% | ||||||||||||
| 15.3 | 14 | 39 | 53 | 26% | ||||||||||||
| 17.9 | 11 | 21 | 22 | 54 | 32 | 43 | 20% | 34% | 20% | |||||||
| 20 | 7 | 18.8 | 25.8 | 27% | ||||||||||||
| 21.2 | 14 | 15 | 29 | 19% | ||||||||||||
| 23.8 | 6 | 12 | 15 | 33 | 18 | 27 | 18% | 33% | 18% | |||||||
| 25 | 11.6 | |||||||||||||||
| 30 | 4 | 7 | 10 | 21 | 11 | 17 | 19% | 36% | 16% | |||||||
| 35 | 3 | |||||||||||||||
Table A.66.
Intercepted life‐stages of FCM on cut roses reported from the Netherlands (NVWA, 2022: Development stage of intercepted Thaumatotibia leucotreta specimens; personal communication, 12 October 2022). Proportion of eggs and portion of young larvae out of all larvae are calculated by the EFSA
| Period | FCM immatures developmental stages | Sum of different periods | Proportion of eggs | Proportion of young larvae (L1 + L2) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Egg | Larval | Pupae | Egg to pupa | Egg to larva | Larva to pupa | Rel. to E to P | Rel. to E to L | Rel. to L to P | Rel. to L | |||||||
| Number of interceptions [−] | [%] | |||||||||||||||
| E | L (Total) | L1 | L1 + L2 | L2 | L3 | L4 | L5 | P | E + L + P | E + L | L + P | E/(E + L + P) | E/(E + L) | (L1 + L2)/(L + P) | (L1 + L2)/L | |
| Years (all countries) | ||||||||||||||||
| 2019–2022 | 95 | 95 | 28 | 33 | 19 | 7 | 5 | 3 | 190 | 50% | 84% | |||||
| 2015–2018 | 54 | 310 | 131 | 85 | 49 | 33 | 12 | 364 | 15% | |||||||
| Countries (2019–2022) | ||||||||||||||||
| Ethiopia | 8 | 30 | 3 | 25 | 2 | 38 | 21% | 93% | ||||||||
| Kenya | 24 | 12 | 5 | 4 | 2 | 1 | 36 | 67% | 92% | |||||||
| Uganda | 41 | 47 | 18 | 3 | 15 | 5 | 3 | 3 | 88 | 47% | 77% | |||||
Table A.67.
Result report on EKE question 1a
| Overview of the results of the Expert Knowledge Elicitation Question 1a | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | Proportion of infested roses, infested with eggs | ||||||||||||||
| Stratification | Eggs out of all life stages | ||||||||||||||
| Question | What is the proportion of infested roses infested with eggs at border control? | ||||||||||||||
| Unit | [%], theoretical restriction: 0–100% | ||||||||||||||
| Results | P1% | P2.5% | P5% | P10% | P16.7% | P25% | P33.3% | P50% | P66.7% | P75% | P83.3% | P90% | P95% | P97.5% | P99% |
| Elicited values | 25 | 40 | 50 | 60 | 70 | ||||||||||
| EKE results | 25.0 | 26.6 | 28.7 | 32.1 | 35.8 | 39.8 | 43.5 | 50.3 | 56.7 | 59.8 | 63.1 | 65.8 | 68.0 | 69.2 | 70.1 |
| Fitted distribution | BetaGeneral (1.5237, 1.2203, 23, 70.8) | ||||||||||||||
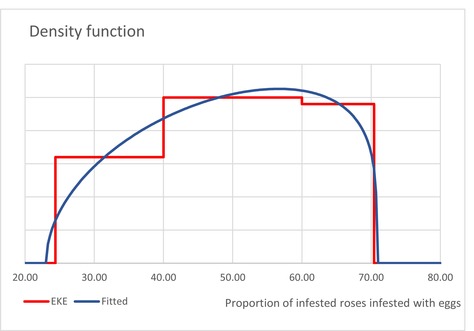
|
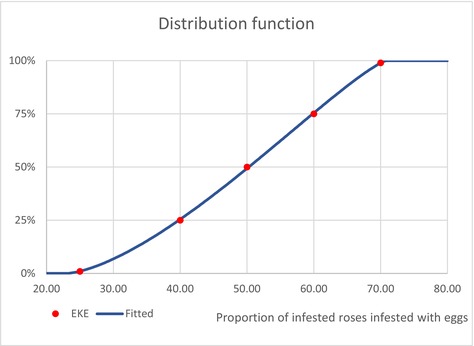
|
||||||||||||||
| Summary of the evidence used for the evaluation | |
| |
| Main uncertainties | |
| |
| Reasoning for a scenario which would lead to a reasonable high proportion | The judgement on the upper limit considers that
|
| Reasoning for a scenario which would lead to a reasonable low proportion | The judgement on the lower limit considers that
|
| Fair estimate as judgement on the weighted evidence | The judgement on the median considers that
|
| Precision of the judgement as description of remaining uncertainties | The judgement on the interquartile range considers that
|
| Experts | Antoon LOOMANS, Panos MILONAS, Klara NICOVA, Stella PAPANASTASIOU, Giuseppe STANCANELLI, Hans‐Hermann THULKE |
| Facilitator/Reporter | Olaf MOSBACH‐SCHULZ (facilitator) |
| Observers | na |
| Date and place of the EKE | Virtual meeting on 11 November 2022 |
Table A.68.
Result report on EKE question 1b
| Overview of the results of the Expert Knowledge Elicitation Question 1b | |||||||||||||||
| Parameter | Proportion of roses infested with larvae, infested with young larvae | ||||||||||||||
| Stratification | Young larvae (L1 + L2) out of all life stages without eggs | ||||||||||||||
| Question | Taking only the infested roses, which are not infested with eggs. What is the proportion of these infested roses, infested with young larvae (L1 or L2) at border control? | ||||||||||||||
| Unit | [%], theoretical restriction: 0–100% | ||||||||||||||
| Results | P1% | P2.5% | P5% | P10% | P16.7% | P25% | P33.3% | P50% | P66.7% | P75% | P83.3% | P90% | P95% | P97.5% | P99% |
| Elicited values | 40 | 55 | 70 | 75 | 85 | ||||||||||
| EKE results | 40.0 | 41.7 | 44.0 | 47.7 | 51.7 | 56.2 | 60.2 | 67.4 | 73.9 | 76.9 | 79.8 | 82.1 | 83.8 | 84.6 | 85.1 |
| Fitted distribution | BetaGeneral (1.4356, 0.99332, 38.1, 85.4) | ||||||||||||||
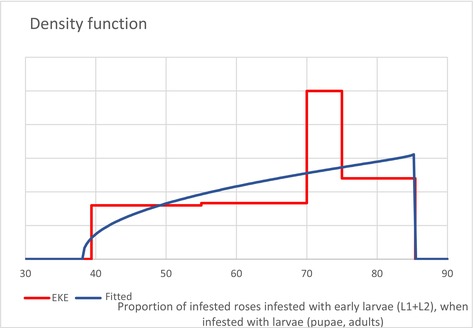
|
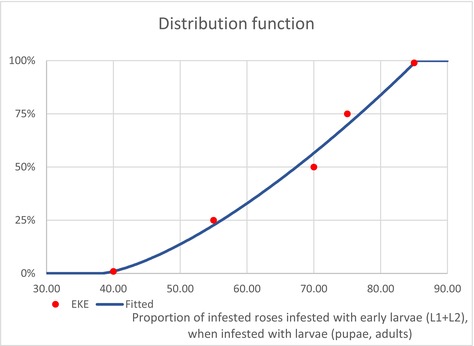
|
||||||||||||||
| Figure (a): Comparison of elicited and fitted values/density function to describe the remaining uncertainties of the parameter | Figure (b): Cumulative distribution function (CDF) of the likelihood of the parameter | ||||||||||||||
| Summary of the evidence used for the evaluation | |
| |
| Main uncertainties | |
| |
| Reasoning for a scenario which would lead to a reasonable high proportion | The judgement on the upper limit considers that
|
| Reasoning for a scenario which would lead to a reasonable low proportion | The judgement on the lower limit considers that
|
| Fair estimate as judgement on the weighted evidence | The judgement on the median considers that
|
| Precision of the judgement as description of remaining uncertainties | The judgement on the interquartile range considers that
|
| Experts | Antoon LOOMANS, Panos MILONAS, Klara NICOVA, Stella PAPANASTASIOU, Giuseppe STANCANELLI, Hans‐Hermann THULKE |
| Facilitator/Reporter | Olaf MOSBACH‐SCHULZ (facilitator) |
| Observers | na |
| Date and place of the EKE | Virtual meeting on 11 November 2022 |
Table A.69.
Distribution of infestation of cut roses at the EU boarder with different immature life stages of FCM
| Proportion at entry | Percentiles | ||||||||||||||
| Life stages | P1% | P2.5% | P5% | P10% | P16.7% | P25% | P33.3% | P50% | P66.7% | P75% | P83.3% | P90% | P95% | P97.5% | P99% |
| Eggs | 24.9% | 26.5% | 28.7% | 32.0% | 35.8% | 39.8% | 43.5% | 50.2% | 56.7% | 59.8% | 63.1% | 65.8% | 68.0% | 69.2% | 70.1% |
| L1 + L2 | 14.1% | 15.3% | 17.6% | 20.4% | 23.3% | 25.2% | 27.8% | 32.0% | 37.4% | 40.5% | 44.5% | 48.2% | 53.0% | 55.9% | 58.9% |
| L3 + L4 + L5 | 5.8% | 6.4% | 7.0% | 8.2% | 9.6% | 11.1% | 12.3% | 16.1% | 19.7% | 21.7% | 24.6% | 27.9% | 31.6% | 33.9% | 37.2% |
At import cut flowers from Africa are mainly infested with eggs and young larvae.
Table A.70.
Inspection rate of consignments of cut roses from Ethiopia, Kenya, Tanzania and Zambia at the Dutch border between 2018 and 2021 (NVWA, 2022: Data on inspection frequencies for cut roses 2018–2021; personal communication, 12 October 2022)
| Country of origin | Year | Consignments imported | No. of inspections | Inspection rate consignments | ||
|---|---|---|---|---|---|---|
| [no. cons.] | [no. cons.] | [% cons.] | ||||
| Estimate | 95%‐CI 1 | |||||
| Ethiopia | 2018 | 635 | 33 | 5.20% | ||
| Ethiopia | 2019 | 112 | 10 | 8.93% | ||
| Ethiopia | 2020 | 518 | 30 | 5.79% | ||
| Ethiopia | 2021 | 1,962 | 102 | 5.20% | ||
| Ethiopia | 2018–2021 | 3,227 | 175 | 5.42% | 4.67% | 6.26% |
| Kenya | 2018 | 58,335 | 4,019 | 6.89% | ||
| Kenya | 2019 | 52,892 | 5,732 | 10.84% | ||
| Kenya | 2020 | 42,273 | 5,439 | 12.87% | ||
| Kenya | 2021 | 38,030 | 4,589 | 12.07% | ||
| Kenya | 2018–2021 | 191,530 | 19,779 | 10.33% | 10.19% | 10.46% |
| Tanzania | 2018 | 1,663 | 284 | 17.08% | ||
| Tanzania | 2019 | 871 | 371 | 42.59% | ||
| Tanzania | 2020 | 195 | 112 | 57.44% | ||
| Tanzania | 2021 | 0 | 0 | |||
| Tanzania | 2018–2021 | 2,729 | 767 | 28.11% | 26.42% | 29.83% |
| Zambia | 2018 | 727 | 82 | 11.28% | ||
| Zambia | 2019 | 616 | 70 | 11.36% | ||
| Zambia | 2020 | 405 | 37 | 9.14% | ||
| Zambia | 2021 | 337 | 57 | 16.91% | ||
| Zambia | 2018–2021 | 2,085 | 246 | 11.80% | 10.44% | 13.26% |
| All | 2018–2021 | 199,571 | 20,967 | 10.51% | 10.37% | 10.64% |
Clopper–Pearson approximation (see below).
Table A.71.
Size and weight of consignments from Ethiopia, Kenya, Tanzania and Zambia at the Dutch border between 2018 and 2021 using trade data from Eurostat (EU trade since 1988 by HS2‐4‐6 and CN8, online, accessed on 4 October 2022)
| Country of origin | Import of cut roses of NL (Eurostat) | Average consignment size 1 | |||
|---|---|---|---|---|---|
| [No. roses] | [100 kg] | [No. roses /cons.] | [kg /cons.] | ||
| 50.7 g per rose | |||||
| Ethiopia | 2018 | 772,833,741 | 391,826.71 | 1,217,061 | 61,705 |
| Ethiopia | 2019 | 643,991,752 | 326,503.82 | 5,749,926 | 291,521 |
| Ethiopia | 2020 | 874,588,023 | 443,416.13 | 1,688,394 | 85,602 |
| Ethiopia | 2021 | 840,201,265 | 425,982.04 | 428,237 | 21,712 |
| Ethiopia | 2018–2021 | 3,131,614,781 | 1,587,729 | 970,442 | 49,201 |
| Kenya | 2018 | 2,091,366,487 | 1,060,322.81 | 35,851 | 1,818 |
| Kenya | 2019 | 2,153,153,091 | 1,091,648.62 | 40,708 | 2,064 |
| Kenya | 2020 | 1,840,848,751 | 933,310.32 | 43,547 | 2,208 |
| Kenya | 2021 | 2,017,377,550 | 1,022,810.42 | 53,047 | 2,689 |
| Kenya | 2018–2021 | 8,102,745,879 | 4,108,092 | 42,305 | 2,145 |
| Tanzania | 2018 | 45,025,113 | 22,827.73 | 27,075 | 1,373 |
| Tanzania | 2019 | 40,132,765 | 20,347.31 | 46,077 | 2,336 |
| Tanzania | 2020 | 15,263,161 | 7,738.42 | 78,273 | 3,968 |
| Tanzania | 2021 | ||||
| Tanzania | 2018–2021 | 100,421,039 | 50,913 | 36,798 | 1,866 |
| Zambia | 2018 | 85,761,147 | 43,480.90 | 117,966 | 5,981 |
| Zambia | 2019 | 83,997,625 | 42,586.80 | 136,360 | 6,913 |
| Zambia | 2020 | 102,281,036 | 51,856.49 | 252,546 | 12,804 |
| Zambia | 2021 | 97,089,926 | 49,224.59 | 288,101 | 14,607 |
| Zambia | 2018–2021 | 369,129,734 | 187,149 | 177,041 | 8,976 |
| All | 2018–2021 | 11,703,911,433 | 5,933,883 | 58,645 | 2,973 |
Number of consignments as indicated by the Dutch NPPO (see Table A.68).
Table A.72.
Sample size of cut roses at the Dutch border control, interpolated in steps of 200 (NVWA, 2022: Infestation rates of consignments of cut‐roses infested by T. leucotreta; personal communication, 12 October 2022. Note: the intermediate steps are calculated by EFSA)
| Consignment size | Sample size | |
|---|---|---|
| [roses] | ||
| From | to | |
| 400 | 10,000 | 200 |
| > 10,000 | 50,000 | 400 |
| > 50,000 | 100,000 | 600 |
| > 100,000 | 150,000 | 800 |
| > 150,000 | 200,000 | 1,000 |
| > 200,000 | 1,200 | |
Table A.73.
Number of specimens per infested sample (n = 217 infested consignments in the period 2019–2022) as reported by the Dutch NPPO for all countries and years (NVWA, 2022: Infestation rates of consignments of cut‐roses infested by T. leucotreta; personal communication, 12 October 2022)
| Number of T. leucotreta specimens per consignment | Fraction of consignments |
|---|---|
| (n = 217) | |
| 1 | 167 (77%) |
| 2 | 32 (15%) |
| 3 | 18 (8%) |
| 4 or more | 0 (0%) |
Table A.74.
Estimated infestation rate (including the 95% confidence interval (CI)) dependent from the sample size and number of specimens detected
| Consignment size | Sample size | Detected specimens | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| [roses] | 0 infested | 1 infested | 2 infested | 3 infested | ||||||||||
| From | To | Estimate | 95%‐CI 1 | Estimate | 95%‐CI 1 | Estimate | 95%‐CI 1 | Estimate | 95%‐CI 1 | |||||
| Infestation rate [% roses] | ||||||||||||||
| 400 | 10,000 | 200 | 0% | 0% | 1.487% | 0.50% | 0.013% | 2.75% | 1.00% | 0.12% | 3.57% | 1.50% | 0.310% | 4.32% |
| > 10,000 | 50,000 | 400 | 0% | 0% | 0.746% | 0.25% | 0.006% | 1.38% | 0.50% | 0.06% | 1.79% | 0.75% | 0.155% | 2.18% |
| > 50,000 | 100,000 | 600 | 0% | 0% | 0.498% | 0.17% | 0.004% | 0.93% | 0.33% | 0.04% | 1.20% | 0.50% | 0.103% | 1.45% |
| > 100,000 | 150,000 | 800 | 0% | 0% | 0.374% | 0.13% | 0.003% | 0.69% | 0.25% | 0.03% | 0.90% | 0.38% | 0.077% | 1.09% |
| > 150,000 | 200,000 | 1,000 | 0% | 0% | 0.299% | 0.10% | 0.003% | 0.56% | 0.20% | 0.02% | 0.72% | 0.30% | 0.062% | 0.87% |
| > 200,000 | 1,200 | 0% | 0% | 0.249% | 0.08% | 0.002% | 0.46% | 0.17% | 0.02% | 0.60% | 0.25% | 0.052% | 0.73% | |
Clopper–Pearson approximation (see below).
Table A.75.
Calculation of the 95% confidence intervals for Binomial(N,p) distributed rates with the Clopper–Pearson approximation. The exponential approximation for full presence/absence is less conservative than Clopper–Pearson (minimal change)
| Observation k out of N | Lower bound | Upper bound |
|---|---|---|
| 0 < k < N | BETA.INV (0.025, k, N – k + 1) | BETA.INV (0.975, k + 1, N – k) |
| k = 0 (full absence in the sample) | 0% | 1‐EXP(LN(0.05)/N) ‘Upper 95% level’ |
| k = N (full presence in the sample | EXP(LN(0.05)/N) ‘Lower 95% level’ | 100% |
Table A.76.
Estimated interception rate with Thaumatotibia leucotreta (including the 95% confidence interval [CI]) for selected African countries
| Country of Origin | Year | No. inspections | No. interceptions with FCM | Interception rate | ||
|---|---|---|---|---|---|---|
| Estimate | 95%‐CI 1 | |||||
| (No. cons.) | (No. cons.) | (% cons.) | (% cons.) | (% cons.) | ||
| Ethiopia | 2018 | 33 | 0 | 0.00% | 0.00% | 8.68% |
| Ethiopia | 2019 | 10 | 0 | 0.00% | 0.00% | 25.89% |
| Ethiopia | 2020 | 30 | 0 | 0.00% | 0.00% | 9.50% |
| Ethiopia | 2021 | 102 | 8 | 7.84% | 3.45% | 14.87% |
| Ethiopia | 2018–2021 | 175 | 8 | 4.57% | 1.99% | 8.81% |
| Kenya | 2018 | 4,019 | 23 | 0.57% | 0.36% | 0.86% |
| Kenya | 2019 | 5,732 | 40 | 0.70% | 0.50% | 0.95% |
| Kenya | 2020 | 5,439 | 33 | 0.61% | 0.42% | 0.85% |
| Kenya | 2021 | 4,589 | 40 | 0.87% | 0.62% | 1.19% |
| Kenya | 2018–2021 | 19,779 | 136 | 0.69% | 0.58% | 0.81% |
| Tanzania | 2018 | 284 | 14 | 4.93% | 2.72% | 8.13% |
| Tanzania | 2019 | 371 | 13 | 3.50% | 1.88% | 5.92% |
| Tanzania | 2020 | 112 | 3 | 2.68% | 0.56% | 7.63% |
| Tanzania | 2021 | |||||
| Tanzania | 2018–2021 | 767 | 30 | 3.91% | 2.65% | 5.54% |
| Zambia | 2018 | 82 | 4 | 4.88% | 1.34% | 12.02% |
| Zambia | 2019 | 70 | 4 | 5.71% | 1.58% | 13.99% |
| Zambia | 2020 | 37 | 1 | 2.70% | 0.07% | 14.16% |
| Zambia | 2021 | 57 | 7 | 12.28% | 5.08% | 23.68% |
| Zambia | 2018–2021 | 246 | 16 | 6.50% | 3.76% | 10.35% |
| All | 2018–2021 | 20,967 | 190 | 0.91% | 0.78% | 1.04% |
Clopper–Pearson approximation (see above).
Table A.77.
Schematic view on the infestation level
| Consignments checked at the EU border | ||||
|---|---|---|---|---|
| No of specimens | No FCM detected | 1 specimen detected | 2 specimens detected | 3 specimens detected |
| Model assumption | Infestation assumed below the limit of detection | Estimated infestation, 95% CI calculated | Estimated infestation, 95% CI calculated | Estimated infestation, 95% CI calculated |
| Probabilistic model | Randomly selected between 0% and the LOD | Randomly selected from the 95%CI | Randomly selected from the 95%CI | Randomly selected from the 95%CI |
| Merged according to the distribution of findings at the Dutch border | ||||
Table A.78.
Schematic view on the final infestation level
| Consignments arriving at the EU border | |||
|---|---|---|---|
| Level/result of the inspection | Not inspected | Checked without detection | Checked with detection |
| Model assumption | Infestation assumed to be similar to the inspected ones | Infestation assumed below the limit of detection | Rejected, not considered |
| Probabilistic model | See first scheme | Randomly selected between 0% and the LOD | |
| Merged according to the distribution of controls and findings at the Dutch border | |||
Table A.79.
Simulation of the overall infestation rate of cut roses imported to the Netherlands stratified by selected African countries
| Parameter | Unit | Country | |||||
|---|---|---|---|---|---|---|---|
| Ethiopia | Kenya | Tanzania | Zambia | All countries | |||
| Proportion of intercepted consignments | [% cons.] | ||||||
| Low (P2.5) | 1.99% | 0.58% | 2.65% | 3.76% | |||
| Est | 4.57% | 0.69% | 3.91% | 6.50% | |||
| High (P97.5) | 8.81% | 0.81% | 5.54% | 10.35% | |||
| Average consignment size | [roses] | 970,442 | 42,305 | 36,798 | 177,041 | ||
| Sample size | [roses] | 1,200 | 400 | 400 | 1,000 | ||
| Detection limit according to the sample size | [%] | ||||||
| High (P95) | 0.249% | 0.746% | 0.746% | 0.299% | |||
| Proportion of interceptions with 1 specimen | [%] | ||||||
| Constant | 77% | 77% | 77% | 77% | |||
| Infestation level of interceptions with 1 specimen | [%] | ||||||
| Low (P2.5) | 0.002% | 0.006% | 0.006% | 0.003% | |||
| Est | 0.083% | 0.250% | 0.250% | 0.100% | |||
| High (P97.5) | 0.463% | 1.385% | 1.385% | 0.556% | |||
| Proportion of interceptions with 2 specimens | [%] | ||||||
| Constant | 15% | 15% | 15% | 15% | |||
| Infestation level of interceptions with 2 specimens | [%] | ||||||
| Low (P2.5) | 0.020% | 0.061% | 0.061% | 0.024% | |||
| Est | 0.167% | 0.500% | 0.500% | 0.200% | |||
| High (P97.5) | 0.601% | 1.794% | 1.794% | 0.721% | |||
| Proportion of interceptions with 3 specimens | [%] | ||||||
| Constant | 8% | 8% | 8% | 8% | |||
| Infestation level of interceptions with 3 specimens | [%] | ||||||
| Low (P2.5) | 0.052% | 0.155% | 0.155% | 0.062% | |||
| Est | 0.250% | 0.750% | 0.750% | 0.300% | |||
| High (P97.5) | 0.729% | 2.176% | 2.176% | 0.874% | |||
| Infestation rate within the inspected consignments | [%] | ||||||
| P01 | 0.011% | 0.012% | 0.030% | 0.016% | |||
| P05 | 0.024% | 0.043% | 0.065% | 0.032% | |||
| Median | 0.136% | 0.395% | 0.403% | 0.165% | |||
| P95 | 0.248% | 0.746% | 0.743% | 0.297% | |||
| P99 | 0.260% | 0.777% | 0.776% | 0.313% | |||
| Total import of roses per year | [roses] | ||||||
| Min | 643,991,752 | 1,840,848,751 | 15,263,161 | 83,997,625 | |||
| Max | 874,588,023 | 2,153,153,091 | 45,025,113 | 102,281,036 | |||
| Inspection rate | [% cons] | ||||||
| Low (P2.5) | 4.67% | 10.19% | 26.42% | 10.44% | |||
| Est | 5.42% | 10.33% | 28.11% | 11.80% | |||
| High (P97.5) | 6.26% | 10.46% | 29.83% | 13.26% | |||
| Number of infested, imported roses per year | [roses] | ||||||
| P05 | 173,520 | 855,344 | 15,748 | 28,193 | 2,134,881 | ||
| Median | 1,021,268 | 7,868,961 | 108,735 | 151,369 | 9,162,766 | ||
| P95 | 1,917,593 | 14,902,594 | 262,553 | 276,725 | 16,284,936 | ||
| Number of imported roses per year | [roses] | ||||||
| P05 | 653,680,625 | 1,855,131,767 | 16,563,332 | 84,219,119 | 2,688,766,359 | ||
| Median | 757,116,193 | 1,995,552,436 | 29,810,984 | 92,374,548 | 2,874,815,411 | ||
| P95 | 860,638,364 | 2,136,040,972 | 43,034,339 | 100,521,627 | 3,061,979,365 | ||
| Infestation rate within total import | [%] | ||||||
| P01 | 0.011% | 0.012% | 0.025% | 0.015% | 0.041% | ||
| P05 | 0.023% | 0.043% | 0.058% | 0.031% | 0.074% | ||
| Median | 0.136% | 0.394% | 0.400% | 0.164% | 0.319% | ||
| P95 | 0.248% | 0.746% | 0.744% | 0.297% | 0.564% | ||
| P99 | 0.260% | 0.777% | 0.775% | 0.312% | 0.599% | ||
| Ethiopia | Kenya | Tanzania | Zambia | All countries | |||
Table A.80.
Result report on EKE question 2
| Overview of the results of the Expert Knowledge Elicitation Question 2 | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | Infestation rate of cut roses imported from Africa at border | ||||||||||||||
| Stratification | None (Total import from Africa) | ||||||||||||||
| Question | What is the proportion of cut roses (stems) imported from any African country, which are infested with any life stage of FCM directly after entering the EU for trade? | ||||||||||||||
| Unit | [%] | ||||||||||||||
| Results | P1% | P2.5% | P5% | P10% | P16.7% | P25% | P33.3% | P50% | P66.7% | P75% | P83.3% | P90% | P95% | P97.5% | P99% |
| Elicited values | 0.01% | 0.12% | 0.20% | 0.35% | 0.65% | ||||||||||
| EKE results | 0.01% | 0.02% | 0.03% | 0.05% | 0.08% | 0.11% | 0.14% | 0.21% | 0.29% | 0.34% | 0.40% | 0.46% | 0.54% | 0.59% | 0.65% |
| Fitted distribution | BetaG .00835) | ||||||||||||||
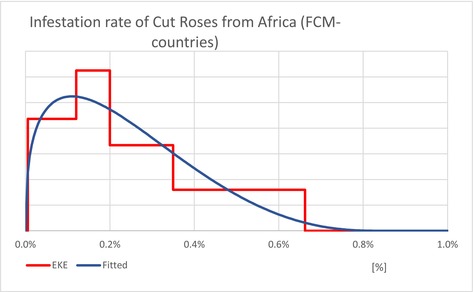
|
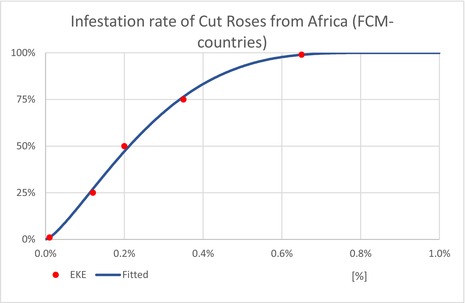
|
||||||||||||||
| Figure (a): Comparison of elicited and fitted values/density function to describe the remaining uncertainties of the parameter | Figure (b): Cumulative distribution function (CDF) of the likelihood of the parameter | ||||||||||||||
| Summary of the evidence used for the evaluation | |
| |
| Main uncertainties | |
| |
| Reasoning for a scenario which would lead to a reasonable high proportion | The judgement on the upper limit considers that
|
| Reasoning for a scenario which would lead to a reasonable low proportion | The judgement on the lower limit considers that
|
| Fair estimate as judgement on the weighted evidence | The judgement on the median considers that
|
| Precision of the judgement as description of remaining uncertainties | The judgement on the interquartile range considers that
|
| Experts | Antoon LOOMANS, Panos MILONAS, Klara NICOVA, Stella PAPANASTASIOU, Giuseppe STANCANELLI, Hans‐Hermann THULKE |
| Facilitator/Reporter | Olaf MOSBACH‐SCHULZ (facilitator) |
| Observers | na |
| Date and place of the EKE | Virtual meeting on 12 December 2022 |
Table A.81.
Number of specimens per infested sample (n = 217) as reported by the Dutch NPPO for all countries and years (nppo, 2022: Infestation rates of consignments of cut roses infested by T. leucotreta; personal communication, 12 October 2022) and as estimated by fitted Poisson distributions
| No. specs. per sample | Observation Dutch NPPO | Fitted Poisson distribution | Fitted Poisson distribution | Fitted Poisson distribution bunch of 10 roses | Fitted Poisson distribution individual roses |
|---|---|---|---|---|---|
| P(k | X ≥ 1) | P(k | X ≥ 1) | P(k) | P(k) | P(k) | |
| Condition: | k ≥ 1, n = 400 | k ≥ 1, n = 400 | n = 400 | n = 10 | n = 1 |
| k = | mu 400 = 0.57 | mu 400 = 0.57 | mu 10 = 0.01425 | mu 1 = 0.001425 | |
| 0 | Not reported | Conditional distribution | 57% | 98.585% | 99.858% |
| 1 | 77% | 74% | 32% | 1.405% | 0.142% |
| 2 | 15% | 21% | 9% | 0.010% | 0.000% |
| 3 | 8% | 4% | 2% | 0.000% | 0.000% |
| 4 | 0% | 1% | 0% | 0.000% | 0.000% |
| ≥ 5 | 0% | 0% | 0% | 0.000% | 0.000% |
| Mean no. specs | 1.31 | 1.31 | 0.57 | 0.0142 | 0.0014 |
Table A.82.
Development times of young larvae (L1 and L2), mature larvae (L3–L5) and pupae according to the literature (Daiber (1979a,b,c, 1989), Schwartz (1981), interpolation of larval stages according Daiber (1979a,b,c))
| Temperature | Development time [d] | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Eggs | Young larvae (L1–L2) | Mature larvae (L3–L5) | Pupae | |||||||||
| Min | Est | Max | Min | Est | Max | Min | Est | Max | Min | Est | Max | |
| 15 | 14.5 | 14.5 | 14.5 | 5.4 | 6.4 | 7.5 | 38.1 | 40.9 | 43.6 | 40.0 | 42.5 | 45.0 |
| 20 | 7.0 | 7.0 | 7.0 | 1.6 | 2.4 | 3.1 | 15.4 | 18.5 | 24.4 | 11.0 | 15.8 | 19.0 |
| 25 | 5.6 | 5.6 | 5.6 | 1.3 | 1.7 | 2.0 | 6.7 | 8.8 | 10.0 | 10.0 | 12.0 | 14.0 |
| 30 | 4.0 | 4.0 | 4.0 | 1.1 | 1.1 | 1.1 | 5.9 | 5.9 | 5.9 | 7.0 | 8.5 | 10.0 |
Table A.83.
Average seasonal temperatures at selected locations in the years 2011–2020 given as median and first and third quartile. Weather data as used for the physiologically based demographic model (PBDM, see Appendix B)
| Ambient temperatures | Location | Szeged (HU) | Archacon (FR) | Capo Vaticano (IT) | Cartagena (ES) | Fuengirola (ES) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Grey/Blue | Blue/Green | Green/Yellow | Yellow/Red | Red+ | ||||||||||||
| Season | P25 | Median | P75 | P25 | Median | P75 | P25 | Median | P75 | P25 | Median | P75 | P25 | Median | P75 | |
| Q1 | Winter | −2.0 | 1.3 | 4.3 | 5.0 | 7.5 | 10.0 | 8.5 | 10.4 | 11.8 | 10.6 | 12.0 | 13.4 | 12.3 | 13.5 | 14.7 |
| Q2 | Spring | 8.7 | 12.4 | 16.1 | 11.2 | 13.4 | 16.0 | 12.8 | 15.1 | 17.8 | 14.9 | 17.0 | 19.0 | 15.1 | 17.0 | 19.1 |
| Q3 | Summer | 19.2 | 21.7 | 24.2 | 19.1 | 20.8 | 22.6 | 23.3 | 25.2 | 26.9 | 24.2 | 25.5 | 26.8 | 23.1 | 24.7 | 26.2 |
| Q4 | Autumn | 8.2 | 12.7 | 16.6 | 11.5 | 15.7 | 18.5 | 15.8 | 18.8 | 21.6 | 16.9 | 20.8 | 23.3 | 17.2 | 20.4 | 22.9 |
Table A.84.
Average seasonal temperatures of the climate suitability classes in the physiologically based demographic model (PBDM) in the years 2011–2020 as used in the developmental model
| Climate suitability class | 1 | 2 | 3 | 4 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Average temperature | Blue | Green | Yellow | Red | ||||||||
| P01 | Median | P99 | P01 | Median | P99 | P01 | Median | P99 | P01 | Median | P99 | |
| Winter | −2.0 | 4.4 | 10.0 | 5.0 | 8.9 | 11.8 | 8.5 | 11.2 | 13.4 | 10.6 | 12.7 | 14.7 |
| Spring | 8.7 | 12.9 | 16.0 | 11.2 | 14.3 | 17.8 | 12.8 | 16.0 | 19.0 | 14.9 | 17.0 | 19.1 |
| Summer | 19.2 | 21.3 | 22.6 | 19.1 | 23.0 | 26.9 | 23.3 | 25.3 | 26.8 | 24.2 | 25.1 | 26.2 |
| Autumn | 8.2 | 14.2 | 18.5 | 11.5 | 17.2 | 21.6 | 15.8 | 19.8 | 23.3 | 16.9 | 20.6 | 22.9 |
Table A.85.
Definition of different scenarios for the time between initial disposal and waste treatment
| Treatment | ||
|---|---|---|
| Days after entry | Days after initial disposal | |
| Scenario 1 | 18 days | 3 days |
| Scenario 2 | 22 days | 7 days |
| Scenario 3 | 29 days | 14 days |
| Scenario 4 | 43 days | 28 days |
| Scenario 5 1 | 71 days | 56 days |
Scenario 5 was calculated but disregarded as worst‐case scenario.
Table A.86.
Median seasonal proportion of escaping adults before the waste treatment for four scenarios of the time between initial disposal and waste treatment
| Proportion adults | Season | Time between initial disposal and waste treatment | |||||
|---|---|---|---|---|---|---|---|
| Climate suitability class | 3 days | 7 days | 14 days | 28 days | 56 days | ||
| 1 | Blue | Winter | 0.05% | 0.05% | 0.05% | 0.05% | 0.05% |
| Spring | 0.05% | 0.11% | 0.39% | 2.67% | 13.1% | ||
| Summer | 0.43% | 3.22% | 17.8% | 96.3% | 100% | ||
| Autumn | 0.05% | 0.20% | 0.94% | 4.98% | 32.4% | ||
| 2 | Green | Winter | 0.05% | 0.05% | 0.05% | 0.12% | 0.27% |
| Spring | 0.08% | 0.26% | 1.29% | 6.44% | 47.1% | ||
| Summer | 0.53% | 4.06% | 27.2% | 99.9% | 100% | ||
| Autumn | 0.16% | 0.82% | 4.38% | 27.0% | 99.5% | ||
| 3 | Yellow | Winter | 0.05% | 0.07% | 0.21% | 0.97% | 5.12% |
| Spring | 0.09% | 0.46% | 2.95% | 15.1% | 91.2% | ||
| Summer | 0.73% | 5.05% | 43.9% | 100% | 100% | ||
| Autumn | 0.33% | 2.33% | 11.9% | 79.4% | 100% | ||
| 4 | Red | Winter | 0.05% | 0.12% | 0.38% | 2.82% | 14.1% |
| Spring | 0.16% | 0.82% | 4.62% | 28.5% | 99.9% | ||
| Summer | 0.73% | 5.09% | 43.9% | 100% | 100% | ||
| Autumn | 0.37% | 2.65% | 14.2% | 87.2% | 100% | ||
Natural mortality of the life stages is assumed (see Section A.9) during the time until the waste treatment. This includes a stable host suitability, and no ‘specific’ treatment of the disposed waste before the commercial waste treatment.
Table A.88.
Proportion of different types of waste treatments of commercial waste per country in the EU climatically suitable area (Eurostat, ENV_WASTRT (WST_OPER), 2020, online, downloaded on 1 February 2023)
| Country in the climatically suitable area | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Spain | Portugal | France | Italy | Malta | Slovenia | Croatia | Greece | Cyprus | Hungary | Bulgaria | Romania | |
| Treatment | Proportion of different treatments of commercial waste [%] | |||||||||||
| Landfill | 5% | 5% | 4% | 0% | 100% | 0% | 15% | 10% | 12% | 3% | 23% | 24% |
| Compost | 92% | 89% | 96% | 98% | 0% | 100% | 77% | 73% | 88% | 88% | 53% | 68% |
| Incineration | 3% | 6% | 0% | 2% | 0% | 0% | 8% | 17% | 0% | 8% | 24% | 8% |
| Total | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% |
Table A.89.
Proportion of different types of waste treatments of household waste per country in the EU climatically suitable area (Eurostat, ENV_WASTRT (WST_OPER), 2020, online, downloaded on 1 February 2023)
| Country in the climatically suitable area | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Spain | Portugal | France | Italy | Malta | Slovenia | Croatia | Greece | Cyprus | Hungary | Bulgaria | Romania | |
| Treatment | Proportion of different treatments of household waste [%] | |||||||||||
| Landfill | 72% | 63% | 36% | 3% | 100% | 96% | 93% | 95% | 100% | 77% | 100% | 100% |
| Compost | 10% | 15% | 0% | 74% | 0% | 4% | 7% | 5% | 0% | 6% | 0% | 0% |
| Incineration | 18% | 22% | 64% | 23% | 0% | 0% | 0% | 0% | 0% | 18% | 0% | 0% |
| Total | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% |
A.1.1. Results for different wasting pathways
The following table explores the pathways of escape and compares the escape from private compost with the collected waste with different waste collection scenarios: The timing from disposal to waste treatment varies from 3, 7, 14 to 28 days.
A.1.2. Results for the seasonal number of mated females
Escape of adults of T. leucotreta is more likely, when the time between initial disposal at the consumer and household waste treatment is prolonged, especially 14 or 28 days. But the effect is more pronounced, when the temperatures induce a faster development, which is especially in summer. In the scenario with 28d, the longer duration is also sufficient to increase the escape in spring and autumn. To support the interpretation, a reasonable clustering scenario is assumed (see Section 2.3.3), namely the escape of females T. leucotreta with possible mating partner in a bunch of 10 roses.
A.1.3. Results for the seasonal number of mated females for selected NUTS2 regions
Considering results at EU level, the highest estimated number of adults escapes and consequently also mating females occurs during summer (Figure A.1, Table A.3). When considering individual NUTS2 regions higher results may occur for specific regions (e.g. Andalusia [Figure A.3], Sicily [Figure A.4]) also in autumn esp. in the 28 days scenario.
A.1.3.1. Rhône‐Alpes (France)
Figure A.2.
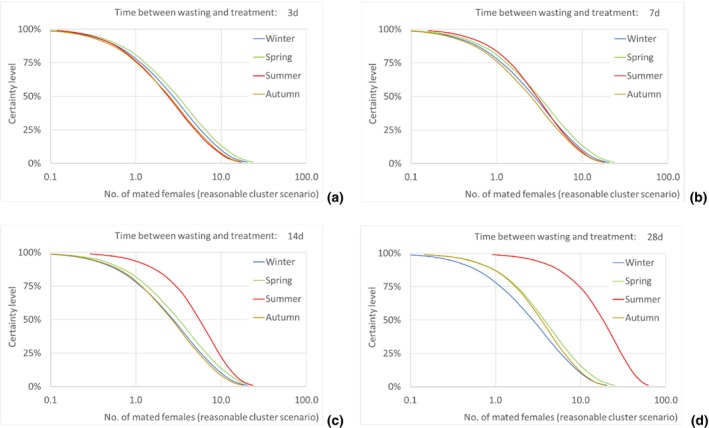
Average seasonal number of female T. leucotreta with possible mating partner in a bunch of 10 roses (Reasonable clustering scenario) per season (blue line = winter, green = spring, red = summer and yellow = autumn) in Rhône‐Alpes (France) with certainty curves for different scenarios between initial wasting and waste treatment: (a) 3 days, (b) 7 days, (c) 14 days and (d) 28 days
A.1.3.2. Andalusia (Spain)
Figure A.3.
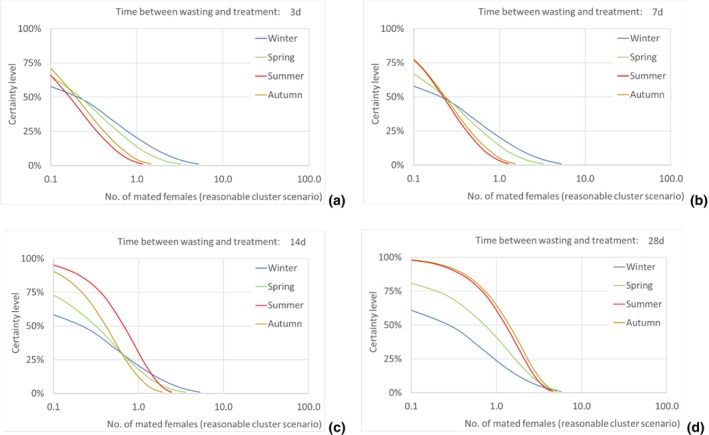
Average seasonal number of female T. leucotreta with possible mating partner in a bunch of 10 roses (Reasonable clustering scenario) per season (blue line = winter, green = spring, red = summer and yellow = autumn) in Andalusia (Spain) with certainty curves for different scenarios between initial wasting and waste treatment: (a) 3 days, (b) 7 days, (c) 14 days and (d) 28 days
A.1.3.3. Sicily (Italy)
A.1.4. Escape per 1 km circles in the residential areas of the NUTS2 regions within 10 days (scenario 3, opinion section 2.3.3)
The pathway model calculates the average annual number of T. leucotreta imported via cut roses from Africa (incl. Israel) and escaping to the environment in the different NUTS2 regions of the EU. NUTS2 regions with unfavourable climatic conditions for possible establishment are excluded. The spatial resolution assumes that the cut roses will be handled, used and wasted in residential areas. The temporal resolution considers trade and climatic conditions per season.
Due to lack of detailed information on the temporal and local appearance of the insects, the model does not estimate the likelihood of establishment, esp. the proportion of females getting mated, the proportion of mated females finding a suitable host and the proportion of populations successful establishing in the new environment.
Instead in Section 2.2.3, a reasonable scenario of clustering in bunches of 10 roses is defined to interpret the data in view of possible mating. In this section, the average number of T. leucotreta adults escaping within a circle with radius 1 km (approximate flying distance to find mating partners) and the usual timespan for mating of 10 days is calculated. The parameter is stratified by NUTS2 regions and described as median estimate with an 90% certainty interval. Finally, the likelihood is calculated, that the average number of escaped insects is equal or higher 2.
Thus, this calculation could be interpreted as (scenario 3) ‘best case scenario’, when no temporal or spatial clustering happens and all escaping adults are homogeneously distributed within the residential area of a NUTS2 area and throughout a year.
Excluded are timely and spatial variations during a year or within the residential areas. These could be triggered by
Clustered infestation of cut roses imported from Africa (incl. Israel): Infestation rates may vary between countries, production sites, production cycles and within a consignment. Therefore, single parts of a consignment may be free of T. leucotreta, while others shown a large proportion of infested roses, e.g. newly infested with eggs.
Specific import patterns: While the average monthly import during a year is quite constant, will be an increase of imported roses to specific festivities, e.g. Valentine's day, Mother's day, etc.
Collection points of private vegetal waste will lead to higher appearance of T. leucotreta adults during specific times and on specific locations.
While the assessment focusses on the residential areas the final distribution of wasted roses in the environment depends from the local circumstances. Areas with isolated houses may have a higher density of private compost sites, while agglomerated housing (e.g. flats) may have larger waste collection sites. Also, intermediate storage of organic waste may lead to higher appearance of FCM adults on specific sites.
For a successful transfer, it has however to be considered that a certain number of adult insects need to emerge at the same time and at the same place, within a climatically suitable region and in presence of suitable host plants, for a successful mating and reproduction to happen.
T. leucotreta adult males may respond to calling females over a distance of 1.0 km and mated females in urban areas may spread at maximum 1.5 km to locate host plants. Such distances generally match the size of the waste treatment plants, which are concentrated in few geographical locations and on limited areas in each NUT2 region. However, based on the simulation results, the number of the estimated escaping adults is expected to be higher in the residential areas in respect to the waste treatment facilities. This is because most of the escapes are estimated to occur during private composting and in the time from the initial disposal by the consumer until the waste treatment. It should be considered that adults escaping during private composting or in the time from disposal until waste treatment may be scattered over a large area limiting their potential for successful mating and reproduction.
To assess the transfer to a suitable host of the T. leucotreta adults escaped from the cut roses pathway, the Panel considered that the adults life span for T. leucotreta is ca. 2 weeks (Mkiga et al., 2019), and the majority of eggs are laid within the first 10 days of female adult stage (Daiber, 1980; Mkiga et al., 2019; EFSA, 2020); the area covered by males responding to calling females would be a circle with radius of ~ 1 km; given a sex ratio of approximately one male: two females, but at least one female and one male would be needed for a successful mating in a particular location; private gardens are mostly located in residential areas. Therefore, to provide an indication of the average number of adults escaping from cut roses that can mate within the flight distance reported for this species, the average number of adult escapes was also standardised to an area of 3.14 km2 over 10 days in the residential area of each NUTS2 region. Results are shown in Table A.4 and Figure A.5 below.
When the average numbers of adult escapes are standardised as discussed above in the residential area of each NUTS2 region, the highest values are observed in NUTS2 regions with high population densities in residential areas. For example, Corsica (France), Liguria (Italy) and Malta (Malta), which had a median of 23,948, 46,551 and 1,592 adult escapes per year, respectively, have the highest values when results are shown as average number of adult escapes standardised to an area of 3.14 km2 over 10 days in the residential area. For Corsica, the values vary from 0.2737 (90% uncertainty between 0.029 and 1.286322) to 0.75064 (90% uncertainty between 0.112 and 2.14375) when considering a time from initial disposal until waste treatment of 3 and 28 days, respectively. Similarly, for Liguria, the values vary from 0.25146 (90% uncertainty between 0.025 and 1.15592) to 0.6509 (90% uncertainty between 0.094 and 1.88955) from scenario 3 to 28 days. For Malta, the values vary from 0.243 (90% uncertainty between 0.026 and 1.16832) to 0.77865 (90% uncertainty between 0.113 and 2.098131) from scenario 3 to 28 days. Also in this case, when the period until waste treatment becomes longer, the number of escapes increases.
A.2. Pest status in Africa
The pathway model considers all African countries where Thaumatotibia leucotreta is reported as present according to EPPO. Israel is added to this list. Some countries do not export roses into the EU.
A.2.1. Export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel
Trade data for the years from 2011 to 2020 were used to estimate the average annual trade.
Main African countries with reported presence of Thaumatotibia leucotreta exporting cut roses to EU by their relative contributions are Kenya, Ethiopia, Uganda, Zambia and Zimbabwe.
A.3. Areas climatically suitable in Europe
The physiologically based demographic model (PBDM, see Appendix B) identified different climate suitability levels within the European Union by evaluating the potential of establishment of the pest from the physiological response of its developmental stages to climatic variables.
For the definition of the areas climatically suitable in Europe, the model version with the widest extension is used. This is the model with 6°C displacement of larvae mortality relative to adults.
The index for possible establishment ranges in Europe from 1 to 509 predicted pupae per year per NUTS2 region and was divided into four climate suitability classes using the quartiles.
The physiologically based demographic model is calculated on a 25 × 25 km grid, while the pathway model uses NUTS2 regions for its spatial stratification. A NUTS2 region was assigned the maximum climate suitability class of the grid cell covered by the region. This gives following areas of different climate suitability classes.
The countries with areas climatically suitable are Portugal, Spain, France, Italy, Malta, Slovenia, Croatia, Greece, Cyprus, Hungary, Bulgaria, Romania.
Due to lacking data on the internal distribution of cut roses within these countries, the model assumes that the roses will be distributed to the NUTS2 regions proportional to the population of the regions.
A.3.1. Import of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to EU
The Netherlands, Belgium and Germany are the main importer of cut roses in Europe, but most of the roses will be re‐exported to other European countries. Together, these three countries comprise more than 95% of the import from Africa.
The model therefore considers two main pathways into the areas climatically suitable:
Direct import from African countries with reported presence of Thaumatotibia leucotreta and from Israel to EU countries in the area climatically suitable
Re‐export of African roses from the Netherlands, Belgium and Germany to EU countries in the area climatically suitable
Re‐export of fresh flowers happens usually within few days. Therefore, the model assumes, that re‐export will only happens once. Nevertheless, the analysis of Intra‐European trade showed that:
Smaller countries (e.g. Portugal, Malta, Cyprus) in the area climatically suitable are substantially supplied by their neighbouring countries;
Between the Netherlands and Belgium, a larger trade was recognised, which indicates additional trade exceeding the national consumption.
In both cases, it is likely that the trade figures represent more economic relations (e.g. used airports for import), than real trade flows. To adapt the model to this uncertainty, the countries were additionally clustered in EU regions with assumed closer economic relationships:
The Netherland and Belgium (for re‐export);
Germany and Luxembourg (for re‐export);
Spain and Portugal (for import);
Italy and Malta (for import);
Greece and Cyprus (for import).
Possible re‐export from countries outside the EU is not considered due to low market share of other countries.
A.4. Direct import from Africa to the area climatically suitable
The trade data were taken from Eurostat (Table ‘EU trade since 1988 by HS2‐4‐6 and CN8’ [DS‐045409], downloaded on 24 January 2023 [Data update: 13 January 2023]) using the CN code 0603 11 00 for ‘Fresh roses as cut flowers and flower buds of a kind suitable for bouquets or for ornamental purposes’ (EC 2021/1832). The trade is retrieved in number of roses (‘Supplementary quantity’) on monthly values from December 2010 to November 2020.
In the analysis, seasons are aggregated and the mean and standard deviation of trade over the 10 years are calculated. Periods with missing values are not considered. Trade was assumed to be unsteady, if only the first years or only singular seasons are reported. In these cases, the direct trade, esp. to Slovenia, Croatia and Hungary, is assumed to be zero in the model.
In total, 22 million cut roses are annually exported directly from Africa to European countries with areas climatically suitable. The average seasonal volume ranges from 4.4 million in summer to 6.1 million in spring.
To cover the annual variation in the model, each seasonal trade volume is included by a normal distribution with the specific mean and standard deviation. Negative values are set to zero.
A.4.1. Direct import from Africa and Israel to EU Member States climatically suitable
A.4.1.1. Direct import from Africa and Israel to Spain and Portugal
The following tables show the detailed annual direct trade from Africa and Israel to the European country clusters.
A.4.1.2. Direct import from Africa and Israel to France
Table A.13.
Average direct import of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to France stratified by season (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Direct export of cut roses from African countries with FCM occurrence and from Israel to the EUI area climatically suitable | ||||||||||||
| Season | Importer: France | |||||||||||
| Year | ||||||||||||
| Winter | 5,596,280 | 6,722,266 | 2,372,482 | 832,821 | 2,676,646 | 2,269,597 | 2,085,767 | 2,492,516 | 2,885,476 | 1,668,034 | 2,960,189 | 1,800,901 |
| Spring | 7,769,782 | 6,497,010 | 1,473,821 | 701,959 | 6,327,428 | 2,577,148 | 3,242,526 | 2,597,895 | 2,640,660 | 4,969,828 | 3,879,806 | 2,364,787 |
| Sommer | 7,631,084 | 3,577,815 | 1,619,495 | 1,842,012 | 3,059,671 | 2,501,320 | 1,810,309 | 1,842,665 | 1,851,521 | 4,009,477 | 2,974,537 | 1,836,849 |
| Autumn | 8,507,193 | 2,454,707 | 1,236,657 | 2,754,378 | 2,346,986 | 2,116,546 | 2,008,757 | 1,777,857 | 1,828,043 | 3,989,292 | 2,902,042 | 2,101,205 |
| Total | 29,504,339 | 19,251,798 | 6,702,455 | 6,131,170 | 14,410,731 | 9,464,611 | 9,147,359 | 8,710,933 | 9,205,700 | 14,636,631 | ||
A.4.1.3. Direct import from Africa and Israel to Italy and Malta
Table A.14.
Average direct import of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Italy and Malta stratified by season (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Direct export of cut roses from African countries with FCM occurrence and from Israel to EU area climatically suitable | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Season | Importer: Italy & Malta | |||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 962,803 | 1,000,999 | 476,860 | 824,850 | 907,680 | 977,375 | 1,053,155 | 1,398,230 | 4,074,650 | 2,395,720 | 1,407,232 | 1,065,589 |
| Spring | 680,097 | 620,297 | 291,020 | 525,926 | 681,050 | 649,800 | 900,270 | 1,198,090 | 1,797,566 | 680,539 | 802,466 | 421,035 |
| Sommer | 134,720 | 116,120 | 3,400 | 24,020 | 162,990 | 354,570 | 273,480 | 257,690 | 698,340 | 646,240 | 267,157 | 239,740 |
| Autumn | 451,580 | 319,570 | 307,005 | 470,580 | 857,562 | 596,810 | 773,409 | 1,493,048 | 1,813,864 | 1,571,135 | 865,456 | 558,681 |
| Total | 2,229,200 | 2,056,986 | 1,078,285 | 1,845,376 | 2,609,282 | 2,578,555 | 3,000,314 | 4,347,058 | 8,384,420 | 5,293,634 | ||
A.4.1.4. Direct import from Africa and Israel to Slovenia
Table A.15.
Average direct import of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Slovenia stratified by season. Cells marked in grey are not used to calculate the mean and standard deviation (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 122010‐12.2020, online, accessed on 13 January 2023)
| Direct export of cut roses from African countries with FCM occurrence and from Israel to the EU area climatically suitable | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Season | Importer: Slovenia | |||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Spring | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sommer | 0 | 102,927 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Autumn | 0 | 164,759 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Total | 0 | 267,686 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
A.4.1.5. Direct import from Africa and Israel to Croatia
Table A.16.
Average direct import of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Croatia stratified by season. Cells marked in grey are not used to calculate the mean and standard deviation (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Direct export of cut roses from African countries with FCM occurrence and from Israel to the EU area climatically suitable | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Season | Importer: Croatia | |||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 863,030 | 540,730 | 416,740 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Spring | 828,155 | 750,534 | 568,232 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sommer | 468,570 | 355,471 | 106,253 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Autumn | 548,090 | 404,730 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Total | 2,707,845 | 2,051,465 | 1,091,225 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
A.4.1.6. Direct import from Africa and Israel to Greece and Cyprus
Table A.17.
Average direct import of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Greece and Cyprus stratified by season. (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Direct export of cut roses from African countries with FCM occurrence and from Israel to the EU area climatically suitable | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Season | Importer: Greece & Cyprus | |||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 1,032,800 | 710,450 | 675,060 | 859,005 | 1,217,925 | 977,642 | 1,308,958 | 1,242,602 | 1,451,601 | 1,616,185 | 1,109,223 | 312,163 |
| Spring | 536,791 | 620,195 | 467,460 | 736,928 | 1,007,870 | 975,032 | 1,239,143 | 1,242,926 | 1,351,166 | 291,344 | 846,886 | 368,115 |
| Summer | 528,500 | 494,050 | 537,142 | 727,347 | 759,907 | 1,087,659 | 1,202,638 | 1,270,758 | 2,191,819 | 348,223 | 914,804 | 549,670 |
| Autumn | 746,336 | 609,610 | 692,124 | 960,376 | 1,014,232 | 1,208,207 | 1,367,877 | 1,403,582 | 1,472,738 | 845,310 | 1,032,039 | 314,822 |
| Total | 2,844,427 | 2,434,305 | 2,371,786 | 3,283,656 | 3,999,934 | 4,248,540 | 5,118,616 | 5,159,868 | 6,467,324 | 3,101,062 | ||
A.4.1.7. Direct import from Africa and Israel to Hungary
Table A.18.
Average direct import of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Hungary stratified by season. Cells marked in grey are not used to calculate the mean and standard deviation. (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Direct export of cut roses from African countries with FCM occurrence and from Israel to the EU area climatically suitable | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Season | Importer: Croatia | |||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Spring | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Summer | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Autumn | 0 | 0 | 6,720 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Total | 0 | 0 | 6,720 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
A.4.1.8. Direct import from Africa and Israel to Bulgaria
Table A.19.
Average direct import of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Bulgaria stratified by season. Cells marked in grey are not used to calculate the mean and standard deviation. (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Direct export of cut roses from African countries with FCM occurrence and from Israel to the EU area climatically suitable | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Season | Importer: Bulgaria | |||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 15,000 | 34,700 | 0 | 0 | 149,200 | 331,218 | 157,080 | 242,930 | 190,490 | 53,720 | 214,184 | 114,673 |
| Spring | 11,500 | 22,210 | 2,090 | 24,070 | 100,410 | 209,210 | 196,016 | 135,124 | 154,130 | 34,220 | 158,978 | 80,027 |
| Summer | 0 | 0 | 0 | 73,520 | 59,205 | 161,090 | 183,290 | 102,660 | 123,950 | 0 | 126,039 | 70,582 |
| Autumn | 26,880 | 0 | 0 | 124,085 | 85,608 | 222,440 | 295,616 | 178,250 | 173,210 | 4,390 | 191,025 | 104,772 |
| Total | 221,675 | 394,423 | 923,958 | 832,002 | 658,964 | 641,780 | 92,330 | |||||
A.4.1.9. Direct import from Africa and Israel to Romania
Table A.20.
Average direct import of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Romania stratified by season. Cells marked in grey are not used to calculate the mean and standard deviation. (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12–2010‐12.2020, online, accessed on 13 January 2023)
| Direct export of cut roses from African countries with FCM occurrence and from Israel to the EU area climatically suitable | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Season | Importer: Romania | |||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 54,300 | 165,610 | 22,140 | 0 | 0 | 97,858 | 63,260 | 16,970 | 8,400 | 35,720 | 46,622 | 41,802 |
| Spring | 81,510 | 67,250 | 0 | 0 | 280 | 318,190 | 436,114 | 283,443 | 354,074 | 40,024 | 347,955 | 65,466 |
| Summer | 190,600 | 52,580 | 0 | 0 | 14,628 | 12,300 | 286,799 | 39,240 | 130,374 | 0 | 117,178 | 123,854 |
| Autumn | 245,975 | 109,400 | 0 | 0 | 124,450 | 45,430 | 408,375 | 13,970 | 216,911 | 0 | 171,172 | 181,550 |
| Total | 572,385 | 394,840 | 22,140 | 0 | 139,358 | 473,778 | 1,194,548 | 353,623 | 709,759 | 75,744 | ||
A.4.2. Inner‐national distribution of cut roses
No data on the further regional distribution of cut roses within the countries was available. Approximately the model takes the distribution of the population within the countries (clusters) as approximation for the distribution and consumption of cut roses. The resolution is set to NUTS2 level.
The population size on 1 January 2020 in European NUTS2 regions is taken from Eurostat (Table ‘Population on 1 January by age, sex and NUTS 2 region’ [DEMO_R_D2JAN], downloaded on 25 January 2023 [Version: 16 November 2022]). Missing data from Croatia were taken from the national CBS (podaci.dzs.hr/2022/en/29031, downloaded on 25 January 2023).
In the model only NUTS2 regions with climate suitability classes 1 to 4 are further considered.
A.4.2.1. NUTS2 regions climatically suitable in Spain and Portugal
Table A.21.
List of NUTS2 regions of Spain and Portugal with their absolute and relative population, and climate suitability class (Eurostat 2022, table DEMO_R_D2JAN, total population on 1st January 2020, online, accessed on 25 January 2023)
| NUTS code | Label | Population | Proportion | Climate suitability class |
|---|---|---|---|---|
| (−) | (%) | FCM | ||
| ESPTT | Spain and Portugal | 57,628,523 | 100.0% | |
| ES | Spain | 47,332,614 | 82.1% | |
| ES11 | Galicia | 2,702,592 | 4.7% | 1 |
| ES12 | Principado de Asturias | 1,018,899 | 1.8% | 1 |
| ES13 | Cantabria | 582,388 | 1.0% | 1 |
| ES21 | País Vasco | 2,189,138 | 3.8% | 1 |
| ES22 | Comunidad Foral de Navarra | 656,509 | 1.1% | 1 |
| ES23 | La Rioja | 315,931 | 0.5% | 1 |
| ES24 | Aragón | 1,330,333 | 2.3% | 1 |
| ES30 | Comunidad de Madrid | 6,747,068 | 11.7% | 1 |
| ES41 | Castilla y León | 2,401,307 | 4.2% | 1 |
| ES42 | Castilla‐la Mancha | 2,045,554 | 3.5% | 2 |
| ES43 | Extremadura | 1,061,979 | 1.8% | 2 |
| ES51 | Cataluña | 7,652,348 | 13.3% | 2 |
| ES52 | Comunitat Valenciana | 5,029,341 | 8.7% | 3 |
| ES53 | Illes Balears | 1,210,725 | 2.1% | 2 |
| ES61 | Andalucía | 8,478,083 | 14.7% | 4 |
| ES62 | Región de Murcia | 1,504,869 | 2.6% | 3 |
| ES63 | Ciudad de Ceuta | 84,085 | 0.1% | 3 |
| ES64 | Ciudad de Melilla | 84,473 | 0.1% | 2 |
| ES70 | Canarias | 2,236,992 | 3.9% | |
| PT | Portugal | 10,295,909 | 17.9% | |
| PT11 | Norte | 3,575,338 | 6.2% | 2 |
| PT15 | Algarve | 438,406 | 0.8% | 3 |
| PT16 | Centro (PT) | 2,217,285 | 3.8% | 3 |
| PT17 | Área Metropolitana de Lisboa | 2,863,272 | 5.0% | 3 |
| PT18 | Alentejo | 704,558 | 1.2% | 3 |
| PT20 | Região Autónoma dos Açores (PT) | 242,796 | 0.4% | |
| PT30 | Região Autónoma da Madeira (PT) | 254,254 | 0.4% |
A.4.2.2. NUTS2 regions climatically suitable in France
Table A.22.
List of NUTS2 regions of France with their absolute and relative population, and climate suitability class (Eurostat 2022, table DEMO_R_D2JAN, total population on 1st January 2020, online, accessed on 25 January 2023)
| NUTS code | Label | Population | Proportion | Climate suitability class |
|---|---|---|---|---|
| (−) | (%) | FCM | ||
| FR | France | 67,320,216 | 100.0% | |
| FR10 | Île de France | 12,291,557 | 18.3% | 0 |
| FRB0 | Centre ‐ Val de Loire | 2,565,726 | 3.8% | 0 |
| FRC1 | Bourgogne | 1,618,321 | 2.4% | 0 |
| FRC2 | Franche‐Comté | 1,176,196 | 1.7% | 0 |
| FRD1 | Basse‐Normandie | 1,463,606 | 2.2% | 0 |
| FRD2 | Haute‐Normandie | 1,849,826 | 2.7% | 0 |
| FRE1 | Nord‐Pas‐de‐Calais | 4,061,166 | 6.0% | 0 |
| FRE2 | Picardie | 1,926,629 | 2.9% | 0 |
| FRF1 | Alsace | 1,908,494 | 2.8% | 0 |
| FRF2 | Champagne‐Ardenne | 1,311,830 | 1.9% | 0 |
| FRF3 | Lorraine | 2,315,678 | 3.4% | 0 |
| FRG0 | Pays‐de‐la‐Loire | 3,818,421 | 5.7% | 1 |
| FRH0 | Bretagne | 3,358,524 | 5.0% | 1 |
| FRI1 | Aquitaine | 3,478,538 | 5.2% | 1 |
| FRI2 | Limousin | 726,253 | 1.1% | 0 |
| FRI3 | Poitou‐Charentes | 1,813,633 | 2.7% | 1 |
| FRJ1 | Languedoc‐Roussillon | 2,864,782 | 4.3% | 1 |
| FRJ2 | Midi‐Pyrénées | 3,087,068 | 4.6% | 1 |
| FRK1 | Auvergne | 1,371,820 | 2.0% | 0 |
| FRK2 | Rhône‐Alpes | 6,692,326 | 9.9% | 1 |
| FRL0 | Provence‐Alpes‐Côte d'Azur | 5,077,582 | 7.5% | 1 |
| FRM0 | Corse | 345,867 | 0.5% | 1 |
| FRY1 | Guadeloupe | 412,682 | 0.6% | |
| FRY2 | Martinique | 359,821 | 0.5% | |
| FRY3 | Guyane | 288,086 | 0.4% | |
| FRY4 | La Réunion | 856,858 | 1.3% | |
| FRY5 | Mayotte | 278,926 | 0.4% | |
| FRXX | Not regionalised/Unknown NUTS 2 | 0 | 0.0% |
A.4.2.3. NUTS2 regions climatically suitable in Italy and Malta
Table A.23.
List of NUTS2 regions of Italy and Malta with their absolute and relative population, and climate suitability class (Eurostat 2022, table DEMO_R_D2JAN, total population on 1st January 2020, online, accessed on 25 January 2023)
| NUTS code | Label | Population | Proportion | Climate suitability class |
|---|---|---|---|---|
| (−) | (%) | FCM | ||
| ITMT | Italy and Malta | 60,156,052 | 100.0% | |
| IT | Italy | 59,641,488 | 99.1% | |
| ITC1 | Piemonte | 4,311,217 | 7.2% | 1 |
| ITC2 | Valle d'Aosta/Vallée d'Aoste | 125,034 | 0.2% | 0 |
| ITC3 | Liguria | 1,524,826 | 2.5% | 1 |
| ITC4 | Lombardia | 10,027,602 | 16.7% | 1 |
| ITF1 | Abruzzo | 1,293,941 | 2.2% | 1 |
| ITF2 | Molise | 300,516 | 0.5% | 1 |
| ITF3 | Campania | 5,712,143 | 9.5% | 2 |
| ITF4 | Puglia | 3,953,305 | 6.6% | 2 |
| ITF5 | Basilicata | 553,254 | 0.9% | 1 |
| ITF6 | Calabria | 1,894,110 | 3.1% | 2 |
| ITG1 | Sicilia | 4,875,290 | 8.1% | 3 |
| ITG2 | Sardegna | 1,611,621 | 2.7% | 2 |
| ITH1 | Provincia Autonoma di Bolzano/Bozen | 532,644 | 0.9% | 0 |
| ITH2 | Provincia Autonoma di Trento | 545,425 | 0.9% | 1 |
| ITH3 | Veneto | 4,879,133 | 8.1% | 1 |
| ITH4 | Friuli‐Venezia Giulia | 1,206,216 | 2.0% | 1 |
| ITH5 | Emilia‐Romagna | 4,464,119 | 7.4% | 1 |
| ITI1 | Toscana | 3,692,555 | 6.1% | 2 |
| ITI2 | Umbria | 870,165 | 1.4% | 1 |
| ITI3 | Marche | 1,512,672 | 2.5% | 1 |
| ITI4 | Lazio | 5,755,700 | 9.6% | 2 |
| MT | Malta | 514,564 | 0.9% | |
| MT00 | Malta | 514,564 | 0.9% | 2 |
A.4.2.4. NUTS2 regions climatically suitable in Slovenia
Table A.24.
List of NUTS2 regions of Slovenia with their absolute and relative population, and climate suitability class (Eurostat 2022, table DEMO_R_D2JAN, total population on 1st January 2020, online, accessed on 25 January 2023)
| NUTS code | Label | Population | Proportion | Climate suitability class |
|---|---|---|---|---|
| (−) | (%) | FCM | ||
| SI | Slovenia | 2,095,861 | 100.0% | |
| SI03 | Vzhodna Slovenija | 1,100,012 | 52.5% | 1 |
| SI04 | Zahodna Slovenija | 995,849 | 47.5% | 1 |
A.4.2.5. NUTS2 regions climatically suitable in Croatia
Table A.25.
List of NUTS2 regions of France with their absolute and relative population, and climate suitability class (Croatian CBS, 2022: Population estimate of Croatia in mid‐2021, online: podaci.dzs.hr/2022/en/29031, accessed on 25 January 2023)
| NUTS code | Label | Population | Proportion | Climate suitability class |
|---|---|---|---|---|
| (−) | (%) | FCM | ||
| HR | Croatia | 3,878,981 | 100.0% | |
| HR02 | Pannonian Croatia | 1,022,966 | 26.4% | 1 |
| HR03 | Adriatic Croatia | 1,300,810 | 33.5% | 1 |
| HR04 | Continental Croatia | |||
| HR05 | City of Zagreb | 768,054 | 19.8% | 1 |
| HR06 | Northern Croatia | 787,151 | 20.3% | 1 |
A.4.2.6. NUTS2 regions climatically suitable in Greece and Cyprus
Table A.26.
List of NUTS2 regions of Greece and Cyprus with their absolute and relative population, and climate suitability class (Eurostat 2022, table DEMO_R_D2JAN, total population on 1st January 2020, online, accessed on 25 January 2023)
| NUTS code | Label | Population | Proportion | Climate suitability class |
|---|---|---|---|---|
| (−) | (%) | FCM | ||
| ELCY | Greece and Cyprus | 11,606,570 | 100.0% | |
| EL | Greece | 10,718,565 | 92.3% | |
| EL30 | Attiki | 3,738,901 | 32.2% | 3 |
| EL41 | Voreio Aigaio | 229,516 | 2.0% | 2 |
| EL42 | Notio Aigaio | 347,512 | 3.0% | 2 |
| EL43 | Kriti | 636,504 | 5.5% | 3 |
| EL51 | Anatoliki Makedonia, Thraki | 598,613 | 5.2% | 1 |
| EL52 | Kentriki Makedonia | 1,872,102 | 16.1% | 1 |
| EL53 | Dytiki Makedonia | 264,670 | 2.3% | 1 |
| EL54 | Ipeiros | 333,265 | 2.9% | 2 |
| EL61 | Thessalia | 715,115 | 6.2% | 1 |
| EL62 | Ionia Nisia | 203,149 | 1.8% | 2 |
| EL63 | Dytiki Ellada | 651,065 | 5.6% | 2 |
| EL64 | Sterea Ellada | 556,002 | 4.8% | 2 |
| EL65 | Peloponnisos | 572,151 | 4.9% | 3 |
| CY | Cyprus | 888,005 | 7.7% | |
| CY00 | Kypros | 888,005 | 7.7% | 2 |
A.4.2.7. NUTS2 regions climatically suitable in Hungary
Table A.27.
List of NUTS2 regions of Hungary with their absolute and relative population, and climate suitability class (Eurostat 2022, table DEMO_R_D2JAN, total population on 1 January 2020, online, accessed on 25 January 2023)
| NUTS code | Label | Population | Proportion | Climate suitability class |
|---|---|---|---|---|
| (−) | (%) | FCM | ||
| HU | Hungary | 9,769,526 | 100.0% | |
| HU11 | Budapest | 1,750,216 | 17.9% | 0 |
| HU12 | Pest | 1,297,102 | 13.3% | 1 |
| HU21 | Közép‐Dunántúl | 1,060,755 | 10.9% | 0 |
| HU22 | Nyugat‐Dunántúl | 994,549 | 10.2% | 0 |
| HU23 | Dél‐Dunántúl | 874,573 | 9.0% | 1 |
| HU31 | Észak‐Magyarország | 1,118,577 | 11.4% | 1 |
| HU32 | Észak‐Alföld | 1,442,660 | 14.8% | 1 |
| HU33 | Dél‐Alföld | 1,231,094 | 12.6% | 1 |
| HUXX | Not regionalised/Unknown NUTS 2 | 0 | 0.0% |
A.4.2.8. NUTS2 regions climatically suitable in Bulgaria
Table A.28.
List of NUTS2 regions of Bulgaria with their absolute and relative population, and climate suitability class (Eurostat 2022, table DEMO_R_D2JAN, total population on 1 January 2020, online, accessed on 25 January 2023)
| NUTS code | Label | Population | Proportion | Climate suitability class |
|---|---|---|---|---|
| (−) | [%] | FCM | ||
| BG | Bulgaria | 6,951,482 | 100.0% | |
| BG31 | Severozapaden | 728,157 | 10.5% | 1 |
| BG32 | Severen tsentralen | 773,450 | 11.1% | 1 |
| BG33 | Severoiztochen | 924,870 | 13.3% | 1 |
| BG34 | Yugoiztochen | 1,024,115 | 14.7% | 1 |
| BG41 | Yugozapaden | 2,094,260 | 30.1% | 1 |
| BG42 | Yuzhen tsentralen | 1,406,630 | 20.2% | 1 |
A.4.2.9. NUTS2 regions climatically suitable in Romania
Table A.29.
List of NUTS2 regions of Romania with their absolute and relative population, and climate suitability class (Eurostat 2022, table DEMO_R_D2JAN, total population on 1st January 2020, online, accessed on 25 January 2023)
| NUTS code | Label | Population | Proportion | Climate suitability class |
|---|---|---|---|---|
| (−) | (%) | FCM | ||
| RO | Romania | 19,328,838 | 100.0% | |
| RO11 | Nord‐Vest | 2,547,429 | 13.2% | 1 |
| RO12 | Centru | 2,314,826 | 12.0% | 0 |
| RO21 | Nord‐Est | 3,184,215 | 16.5% | 0 |
| RO22 | Sud‐Est | 2,377,101 | 12.3% | 1 |
| RO31 | Sud ‐ Muntenia | 2,901,376 | 15.0% | 1 |
| RO32 | Bucuresti ‐ Ilfov | 2,322,002 | 12.0% | 1 |
| RO41 | Sud‐Vest Oltenia | 1,910,409 | 9.9% | 1 |
| RO42 | Vest | 1,771,480 | 9.2% | 1 |
A.4.3. Area of NUTS2 regions climatically suitable
For the distribution of the escaping adults within a NUTS2 region the model focusses on the residential areas only.
A.4.4. Grading
Cut roses entering the European market are mainly packed in cardboard boxes and will be sorted and re‐packed into bunches of 10–20 stems directly after entering the EU. They need to comply with specific quality standards, which will be checked at the grading step. ‘Products which do not meet the requirements for pre‐treatment, minimum quality, bacteria content and ripeness are not traded and are destroyed if necessary.’ (Dutch Centre for the Promotion of Imports from developing countries [CBI], 2017. Exporting roses to Europe. Updated: 13 June 2017. Internet: www.cbi.eu/market-information/cut-flowers-foliage/roses/europe; accessed on 3 February 2023. Because the grading is a responsibility of the trader, parts of the process can be already done at the country of origin (e.g. export of sorted and packed to bunches).
Due to the absence of further detailed information the model assumes a loss of 2–5% of flowers at grading. This margin is frequently accepted for trade of perishable products. The uncertainty on the average proportion is modelled by a Uniform distribution.
It is assumed, that rejected flowers will be transferred into the commercial waste flow for vegetal waste.
A.5. Re‐export by the Netherland and Belgium to the climatically suitable area
Also the data on Inner‐European trade were taken from Eurostat (Table ‘EU trade since 1988 by HS2‐4‐6 and CN8’, accessed on 25 January 2023 [Data update: 13 January 2023]) using the CN code 0603 11 00 for ‘Fresh roses as cut flowers and flower buds of a kind suitable for bouquets or for ornamental purposes’ (EC 2021/1832). The trade is retrieved in number of roses (‘Supplementary quantity’) on monthly values from December 2010 to November 2020.
The export of cut roses from the Netherlands and Belgium to European countries with areas climatically suitable is on average 699 million per year. This is more than 30 times more than the direct import of these countries. The average seasonal volume ranges from 145 million in summer to 201 million in spring.
To cover the annual variation in the model, each seasonal trade volume is included by a Normal distribution with the specific mean and standard deviation.
A.5.1.1. Export from the Netherlands and Belgium to Spain and Portugal
Table A.32.
Average re‐export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Spain and Portugal via the Netherlands and Belgium stratified by season. (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Export of cut roses [pcs] from the Netherland and Belgium to the EU area climatically suitable | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Season | Importer: Spain and Portugal | |||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 54,922,392 | 5,652,691 | 4,688,797 | 3,626,780 | 4,709,680 | 4,094,909 | 4,601,809 | 3,546,874 | 4,525,384 | 4,199,646 | 9,456,896 | 15,986,293 |
| Spring | 9,570,556 | 7,305,652 | 5,183,524 | 5,084,602 | 5,337,739 | 27,603,086 | 4,911,252 | 4,860,443 | 6,892,104 | 2,854,199 | 7,960,316 | 7,132,380 |
| Summer | 6,418,915 | 4,438,117 | 3,529,361 | 3,876,374 | 4,371,803 | 4,466,202 | 4,193,624 | 3,792,502 | 5,253,900 | 4,403,415 | 4,474,421 | 830,011 |
| Autumn | 7,006,463 | 5,112,871 | 3,890,031 | 4,986,154 | 5,182,401 | 6,443,965 | 4,348,308 | 5,089,070 | 5,543,803 | 6,942,195 | 5,454,526 | 1,044,759 |
| Total | 77,918,326 | 22,509,331 | 17,291,713 | 17,573,910 | 19,601,623 | 42,608,162 | 18,054,993 | 17,288,889 | 22,215,191 | 18,399,455 | ||
A.5.1.2. Export from the Netherlands and Belgium to France
Table A.33.
Average re‐export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to France via the Netherlands and Belgium stratified by season. (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses (pcs) from the Netherland and Belgium to the EU climatically suitable area | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Importer: France | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 105,443,649 | 86,308,088 | 117,352,998 | 106,072,208 | 117,405,185 | 127,401,101 | 122,384,930 | 123,117,862 | 108,414,921 | 107,103,882 | 112,100,482 | 12,025,162 |
| Spring | 113,918,882 | 100,973,929 | 119,436,131 | 124,940,318 | 147,277,945 | 161,713,606 | 150,058,041 | 152,882,927 | 136,168,966 | 77,887,672 | 128,525,842 | 26,247,886 |
| Summer | 75,537,438 | 76,934,831 | 76,807,397 | 97,689,767 | 115,366,711 | 103,772,589 | 99,150,542 | 94,501,506 | 92,539,093 t | 98,075,858 | 93,037,573 | 13,036,657 |
| Autumn | 85,667,581 | 91,630,564 | 90,743,824 | 107,298,307 | 116,508,744 | 110,497,034 | 101,204,375 | 106,755,003 | 103,342,089 | 84,099,161 | 99,774,668 | 11,094,109 |
| Total | 380,567,550 | 355,847,412 | 404,340,350 | 436,000,600 | 4,96,558,585 | 503,384,330 | 472,797,888 | 477,257,298 | 440,465,069 | 367,166,573 | ||
A.5.1.3. Export from the Netherlands and Belgium to Italy and Malta
Table A.34.
Average re‐export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Italy and Malta via the Netherlands and Belgium stratified by season. (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses (pcs) from the Netherland and Belgium to the EU area climatically suitable | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Importer: Italy and Malta | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 51,212,506 | 45,487,867 | 47,078,115 | 44,951,207 | 52,564,530 | 51,367,942 | 52,371,942 | 43,205,992 | 39,486,198 | 41,387,448 | 46,911,375 | 4,777,377 |
| Spring | 44,302,035 | 48,945,386 | 47,739,292 | 45,469,677 | 49,642,989 | 56,152,297 | 49,865,702 | 44,871,286 | 42,820,139 | 26,846,361 | 45,665,516 | 7,625,087 |
| Summer | 30,466,710 | 30,816,086 | 36,928,206 | 30,549,912 | 36,447,464 | 35,939,689 | 34,650,285 | 31,792,558 | 27,875,089 | 30,775,364 | 32,624,136 | 3,111,446 |
| Autumn | 40,818,457 | 43,150,536 | 43,413,959 | 44,613,784 | 44,566,565 | 48,417,493 | 45,930,981 | 41,741,261 | 31,535,713 | 43,394,554 | 42,758,330 | 4,478,807 |
| Total | 166,799,708 | 168,399,875 | 175,159,572 | 165,584,580 | 183,221,548 | 191,877,421 | 182,818,910 | 161,611,097 | 141,717,139 | 142,403,727 | ||
A.5.1.4. Export from the Netherlands and Belgium to Slovenia
Table A.35.
Average re‐export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Slovenia via the Netherlands and Belgium stratified by season. (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses (pcs) from the Netherland and Belgium to the EU area climatically suitable | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Importer: Slovenia | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 1,005,363 | 942,239 | 892,867 | 860,816 | 759,867 | 726,592 | 855,575 | 791,601 | 942,687 | 1,439,872 | 921,748 | 201,786 |
| And Spring | 1,343,583 | 1,331,953 | 1,277,011 | 1,094,782 | 1,109,039 | 1,102,119 | 1,157,047 | 1,149,996 | 1,541,320 | 1,018,843 | 1,212,569 | 158,102 |
| Summer | 1,039,225 | 854,929 | 827,672 | 784,687 | 766,526 | 816,391 | 831,853 | 914,622 | 932,086 | 946,282 | 871,427 | 84,731 |
| Autumn | 1,213,301 | 1,125,320 | 1,058,102 | 966,095 | 961,370 | 1,077,297 | 983,052 | 1,056,319 | 1,123,349 | 1,417,831 | 1,098,204 | 137,442 |
| Total | 4,601,472 | 4,254,441 | 4,055,652 | 3,706,380 | 3,596,802 | 3,722,399 | 3,827,527 | 3,912,538 | 4,539,442 | 4,822,828 | ||
A.5.1.5. Export from the Netherlands and Belgium to Croatia
Table A.36.
Average re‐export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Croatia via the Netherlands and Belgium stratified by season. (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses (pcs) from the Netherlands and Belgium to the EU area climatically suitable | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Importer: Croatia | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 624,210 | 484,580 | 673,707 | 828,130 | 1,814,309 | 1,963,876 | 2,300,148 | 2,504,696 | 2,869,914 | 3,823,790 | 1,788,736 | 1,122,441 |
| Spring | 883,970 | 818,868 | 697,271 | 1,424,131 | 2,687,434 | 3,687,763 | 3,445,956 | 3,443,517 | 4,451,406 | 3,012,724 | 2,455,304 | 1,379,491 |
| Summer | 847,243 | 801,209 | 1,034,271 | 1,195,977 | 2,059,519 | 2,747,543 | 2,025,793 | 2,381,601 | 3,458,288 | 2,502,009 | 1,905,345 | 903,247 |
| Autumn | 750,690 | 791,794 | 1,012,756 | 1,689,809 | 2,200,260 | 2,857,706 | 2,166,501 | 3,055,162 | 3,848,748 | 3,022,336 | 2,139,576 | 1,068,565 |
| Total | 3,106,113 | 2,896,451 | 3,418,005 | 5,138,047 | 8,761,522 | 11,256,888 | 9,938,398 | 11,384,976 | 14,628,356 | 12,360,859 | ||
A.5.1.6. Export from the Netherlands and Belgium to Greece and Cyprus
Table A.37.
Average re‐export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Greece and Cyprus via the Netherlands and Belgium stratified by season (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses [pcs] from the Netherland and Belgium to the EU area climatically suitable | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Importer: Greece and Cyprus | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 3,058,945 | 3,155,845 | 1,948,872 | 1,918,740 | 2,374,786 | 2,932,047 | 3,720,640 | 3,381,805 | 3,720,117 | 4,365,838 | 3,057,764 | 796,735 |
| Spring | 2,332,618 | 2,483,458 | 1,222,767 | 973,875 | 1,936,915 | 2,265,068 | 2,369,365 | 2,949,135 | 3,454,441 | 1,526,823 | 2,151,447 | 761,785 |
| Summer | 849,296 | 835,480 | 713,876 | 972,249 | 1,457,777 | 1,443,578 | 1,853,957 | 2,118,312 | 2,635,520 | 1,738,431 | 1,461,848 | 632,963 |
| Autumn | 1,064,463 | 848,008 | 721,621 | 1,241,455 | 2,046,289 | 1,618,086 | 2,416,566 | 2,459,965 | 3,023,298 | 1,975,609 | 1,741,536 | 769,181 |
| Total | 7,305,322 | 7,322,791 | 4,607,136 | 5,106,319 | 7,815,767 | 8,258,779 | 10,360,528 | 10,909,217 | 12,833,376 | 9,606,701 | ||
A.5.1.7. Export from the Netherlands and Belgium to Hungary
Table A.38.
Average re‐export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Hungary via the Netherlands and Belgium stratified by season (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses [pcs] from the Netherlands and Belgium to the EU area climatically suitable | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Importer: Hungary | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 4,346,528 | 4,346,528 | 4,346,528 | 3,122,310 | 2,823,419 | 3,490,718 | 2,734,464 | 2,581,344 | 3,131,580 | 2,798,249 | 3,372,167 | 718,415 |
| Spring | 4,756,980 | 5,024,544 | 3,781,356 | 3,571,663 | 3,301,393 | 5,919,991 | 3,904,358 | 4,128,578 | 4,469,345 | 2,157,236 | 4,101,544 | 1,031,021 |
| Summer | 3,296,814 | 3,020,070 | 2,856,324 | 3,118,007 | 3,654,948 | 4,073,845 | 2,469,502 | 3,050,100 | 3,008,869 | 2,420,491 | 3,096,897 | 497,899 |
| Autumn | 3,787,750 | 2,740,039 | 3,101,069 | 3,191,286 | 3,843,916 | 3,395,119 | 2,470,416 | 3,802,608 | 3,310,171 | 2,420,583 | 3,206,296 | 530,643 |
| Total | 16,188,072 | 15,131,181 | 14,085,277 | 13,003,266 | 13,623,676 | 16,879,673 | 11,578,740 | 13,562,630 | 13,919,965 | 9,796,559 | ||
A.5.1.8. Export from the Netherlands and Belgium to Bulgaria
Table A.39.
Average re‐export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Bulgaria via the Netherlands and Belgium stratified by season. (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses [pcs] from the Netherlands and Belgium to the EU area climatically suitable | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Importer: Bulgaria | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 701,933 | 686,740 | 538,284 | 418,049 | 606,632 | 553,377 | 643,998 | 1,133,920 | 829,972 | 1,028,770 | 714,168 | 223,717 |
| Spring | 719,991 | 522,810 | 705,171 | 472,013 | 623,786 | 749,185 | 837,847 | 1,128,000 | 1,511,261 | 964,776 | 823,484 | 310,909 |
| Summer | 457,355 | 355,147 | 389,682 | 329,470 | 477,217 | 645,155 | 1,444,716 | 922,922 | 1,266,157 | 894,682 | 718,250 | 397,692 |
| Autumn | 562,702 | 477,338 | 522,199 | 415,767 | 527,285 | 747,746 | 1,423,528 | 862,931 | 1,879,353 | 934,199 | 835,305 | 474,415 |
| Total | 2,441,981 | 2,042,035 | 2,155,336 | 1,635,299 | 2,234,920 | 2,695,463 | 4,350,089 | 4,047,773 | 5,486,743 | 3,822,427 | ||
A.5.1.9. Export from the Netherlands and Belgium to Romania
Table A.40.
Average re‐export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Romania via the Netherlands and Belgium stratified by season (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses [pcs] from the Netherland and Belgium to the EU area climatically suitable | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Importer: Romania | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 2,326,982 | 1,683,713 | 2,872,126 | 2,008,723 | 1,988,658 | 6,640,686 | 5,915,151 | 8,859,692 | 11,640,742 | 12,847,059 | 5,678,353 | 4,218,837 |
| Spring | 2,476,565 | 4,347,120 | 2,552,544 | 3,282,221 | 2,981,559 | 10,397,611 | 11,460,874 | 14,759,952 | 18,758,298 | 13,915,809 | 8,493,255 | 6,079,992 |
| Summer | 2,255,004 | 3,154,482 | 4,017,982 | 3,430,715 | 2,870,267 | 7,640,417 | 9,884,183 | 12,849,892 | 13,545,425 | 11,795,401 | 7,144,377 | 4,528,549 |
| Autumn | 1,984,667 | 2,931,792 | 3,765,089 | 2,742,336 | 5,009,901 | 7,731,739 | 10,286,333 | 14,751,512 | 15,255,584 | 14,187,898 | 7,864,685 | 5,354,353 |
| Total | 9,043,218 | 12,117,107 | 13,207,741 | 11,463,995 | 12,850,385 | 32,410,453 | 37,546,541 | 51,221,048 | 59,200,049 | 52,746,167 | ||
A.5.2. Dilution of African cut roses during re‐export by the Netherlands and Belgium
The origin of the cut roses is not traced when these are being re‐exported for intra‐EU trade from the Netherlands and Belgium. A dilution may happen, when they are mixed with roses of other origins including the production in the Netherlands and Belgium itself.
To estimate the dilution, factor the model uses information on the import of cut roses from third countries outside the EU excluding the African import, and the total import of cut roses excluding the African import. The latter includes also import from other European countries.
While the Netherlands and Belgium imported on annual average 4,034 million cut roses from African FCM countries and 264 million from other third countries, the total import from Non‐African FCM countries is 875 million roses (incl. intra‐EU trade). The own production of cut roses in the Netherlands and Belgium is about 122 million.
A.5.2.1. Direct export of cut roses from African countries with FCM occurrence and from Israel to the Netherlands and Belgium
Table A.41.
Average direct export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to the Netherlands and Belgium stratified by season (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Direct export of cut roses (pcs) from African countries with FCM occurrence and from Israel to the Netherlands and Belgium | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Exporter: African countries with reported occurrence of FCM and Israel | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 968,311,592 | 899,907,368 | 962,253,601 | 912,705,433 | 1,002,526,366 | 1,045,167,474 | 998,528,528 | 991,120,020 | 1,042,930,826 | 1,004,207,567 | 982,765,878 | 48,403,322 |
| Spring | 1,088,359,634 | 1,078,252,708 | 1,082,907,378 | 1,160,242,521 | 1,194,317,742 | 1,252,616,965 | 1,216,071,008 | 1,126,489,283 | 1,225,796,204 | 914,716,166 | 1,133,976,961 | 99,723,029 |
| Summer | 856,247,226 | 792,848,979 | 803,373,275 | 1,009,336,075 | 976,441,600 | 938,395,539 | 974,945,987 | 841,497,757 | 883,725,177 | 921,901,345 | 899,871,296 | 75,856,641 |
| Autumn | 931,377,554 | 948,755,946 | 981,919,812 | 1,027,628,033 | 1,081,044,981 | 1,035,893,806 | 1,030,055,155 | 1,052,462,689 | 1,036,091,544 | 1,052,989,797 | 1,017,821,932 | 48,156,920 |
| Total | 3,844,296,006 | 3,719,765,001 | 3,830,454,066 | 4,109,912,062 | 4,254,330,689 | 4,272,073,784 | 4,219,600,678 | 4,011,569,749 | 4,188,543,751 | 3,893,814,875 | 4,034,436,066 | 201,558,934 |
A.5.2.2. Export from Non‐EU countries (different from African countries with FCM occurrence and Israel) to the Netherlands and Belgium
Table A.42.
Average export of cut roses from countries outside the EU (excluding the African countries with reported presence of Thaumatotibia leucotreta and Israel) to the Netherlands and Belgium stratified by season (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses [pcs] from Non‐EU countries (excluding African countries with FCM occurrence and Israel) to the Netherlands and Belgium | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Exporter: All countries outside EU27 (2020) (excluding African countries with FCM occurrence and Israel) | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 57,219,395 | 62,838,666 | 55,824,054 | 68,010,189 | 70,661,224 | 79,576,154 | 66,049,118 | 71,153,040 | 81,133,470 | 95,954,508 | 70,841,982 | 12,106,589 |
| Spring | 52,810,406 | 59,368,783 | 55,656,894 | 61,398,802 | 65,396,635 | 76,929,861 | 67,769,557 | 87,204,694 | 90,357,076 | 58,271,159 | 67,516,387 | 13,117,814 |
| Summer | 39,467,290 | 44,117,164 | 42,493,733 | 46,854,560 | 51,622,219 | 54,417,987 | 52,111,798 | 66,038,031 | 68,913,177 | 67,884,829 | 53,392,079 | 10,842,946 |
| Autumn | 56,227,276 | 58,780,418 | 59,426,662 | 63,269,178 | 67,875,321 | 72,597,561 | 72,829,050 | 84,295,425 | 85,670,656 | 98,199,028 | 71,917,058 | 13,744,875 |
| Total | 205,724,367 | 225,105,031 | 213,401,343 | 239,532,729 | 255,555,399 | 283,521,563 | 258,759,523 | 308,691,190 | 326,074,379 | 320,309,524 | 263,667,505 | 44,181,183 |
A.5.2.3. Export from all countries (including EU countries; excluding African countries with FCM occurrence and Israel) to the Netherlands and Belgium
Table A.43.
Average export of cut roses from all countries (including other EU countries; excluding African countries with reported presence of Thaumatotibia leucotreta and Israel) to the Netherlands and Belgium stratified by season (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses [pcs] from all countries (excluding African countries with FCM occurrence and Israel) to the Netherlands and Belgium | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Exporter: All countries including EU27 (2020) (excluding African countries with FCM occurrence and Israel) | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 74,503,091 | 119,991,131 | 121,163,875 | 130,573,999 | 141,665,463 | 424,630,986 | 360,752,981 | 214,006,436 | 151,287,708 | 342,825,815 | 208,140,149 | 122,562,989 |
| Spring | 80,363,073 | 131,168,647 | 148,345,013 | 157,541,324 | 423,672,557 | 485,353,485 | 464,134,848 | 250,543,206 | 179,912,356 | 324,401,863 | 264,543,637 | 149,636,638 |
| Summer | 89,406,636 | 116,909,664 | 112,385,598 | 113,030,775 | 409,059,708 | 422,248,395 | 112,722,488 | 139,778,234 | 134,146,359 | 193,269,355 | 184,295,721 | 125,004,409 |
| Autumn | 122,573,769 | 129,665,466 | 138,068,493 | 136,168,094 | 421,034,045 | 425,570,306 | 136,623,353 | 172,469,809 | 150,816,977 | 342,363,256 | 217,535,357 | 126,040,423 |
| Total | 366,846,569 | 497,734,908 | 519,962,979 | 537,314,192 | 1,395,431,773 | 1,757,803,172 | 1,074,233,670 | 776,797,685 | 616,163,400 | 1,202,860,289 | 874,514,864 | 461,235,861 |
A.5.2.4. Own production of cut roses in the Netherlands and Belgium
The national statistical office of the Netherlands (CBS: opendata.cbs.nl/statline/) reports the area of cut roses production for certain years in open fields and glasshouses.
Similar data were not available for Belgium. Instead, the relation of area for all flower and ornamental production (Eurostat: Crops by classes of utilised agricultural area) in the Netherland and Belgium was used to estimate a factor of 111% to extrapolate from the area of the Netherland to the area of both, the Netherlands and Belgium.
To convert the production area into the number of produced cut roses a productivity of 200,000 roses/ha was used. The estimation was informed by productivity figures of different types of roses, flowers per plant and plants per ha. The reported data ranges roughly from 150,000 to 320,000.
The mean values were used to calculate the average, annual production of cut roses in the Netherland and Belgium:
Finally, the production is equally distributed on all four seasons (30,565,283 pcs/season), reasoned by a higher consumption in spring, but an assumed additional outdoor production in summer.
A.5.2.5. Dilution
For the calculation of the dilution rate two scenarios were assessed:
In scenario 1 the Netherlands and Belgium get non‐infested roses from other countries than the African countries with FCM occurrence and Israel, including the intra‐EU trade, for re‐export. This gives a higher dilution and low dilution factor (proportion of cut roses from African countries with FCM occurrence and Israel in re‐export).
In scenario 2 the intra‐EU trade to the Netherlands and Belgium is not re‐exported to further countries. This gives a lower dilution and higher dilution factor.
In the model calculation a Uniform distribution between the low and high scenario is used to describe the uncertainty on the current proportion of cut roses from African countries with FCM occurrence and from Israel in re‐export.
A.6. Re‐export (intra‐EU trade) from Germany and Luxembourg to the EU area climatically suitable
Also, the data on Inner‐European trade were taken from Eurostat (Table ‘EU trade since 1988 by HS2‐4‐6 and CN8’ [DS‐045409], downloaded on 24th January 2023 [Data update: 13 January 2023]) using the CN code 0603 11 00 for ‘Fresh roses as cut flowers and flower buds of a kind suitable for bouquets or for ornamental purposes’ (EC 2021/1832). The trade is retrieved in number of roses (‘Supplementary quantity’) on monthly values from December 2010 to November 2020.
The export of cut roses from Germany and Luxembourg to EU countries with areas climatically suitable is limited to France, Italy and Malta. It comprises 1.8 million cut flowers in total. The average seasonal volume ranges from 0.34 million in summer to 0.42 million in spring. For all other countries the trade is unsteady and assumed to be zero in the model.
To cover the annual variation in the model, each seasonal trade volume is included by a Normal distribution with the specific mean and standard deviation.
A.6.1.1. Export from Germany and Luxembourg to Spain and Portugal
Table A.48.
Average re‐export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Spain and Portugal via Germany and Luxembourg stratified by season. Cells marked in grey are not used to calculate the mean and standard deviation (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses (pcs) from Germany and Luxembourg to the EU area climatically suitable | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Importer: Spain and Portugal | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 0 | 2,560 | 0 | 26,360 | 952,000 | 0 | 10 | 0 | 0 | 1 | 0 | 0 |
| Spring | 0 | 0 | 2 | 47,520 | 126,920 | 137,560 | 147,650 | 186,448 | 164,096 | 364,464 | 167,808 | 97,037 |
| Summer | 6,200 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| Autumn | 15 | 14 | 0 | 8,320 | 0 | 34,560 | 0 | 0 | 50 | 110,806 | 0 | 0 |
| Total | 6,215 | 2,574 | 2 | 82,200 | 1,078,921 | 172,120 | 147,661 | 186,448 | 164,146 | 475,271 | ||
A.6.1.2. Export from Germany and Luxembourg to France
Table A.49.
Average re‐export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to France via Germany and Luxembourg stratified by season. Cells marked in grey are not used to calculate the mean and standard deviation (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses [pcs] from Germany and Luxembourg to the EU area climatically suitable | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Importer: France | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 418,323 | 422,709 | 480,492 | 119,157 | 55,278 | 86,425 | 122,551 | 23,952 | 16,070 | 9,695 | 175,465 | 187,776 |
| Spring | 703,801 | 354,210 | 553,036 | 184,632 | 139,576 | 307,278 | 93,618 | 7,598 | 10,665 | 1,040 | 235,545 | 242,386 |
| Summer | 171,349 | 203,647 | 129,674 | 76,088 | 485,755 | 112,243 | 116,293 | 6,883 | 7,853 | 3,959 | 131,374 | 142,615 |
| Autumn | 448,670 | 547,480 | 146,687 | 129,470 | 4,186,056 | 133,551 | 73,318 | 4,272 | 3,707 | 6,167 | 567,938 | 1,284,961 |
| Total | 1,742,143 | 1,528,046 | 1,309,889 | 509,347 | 4,866,665 | 639,497 | 405,780 | 42,705 | 38,295 | 20,861 | ||
A.6.1.3. Export from Germany and Luxembourg to Italy and Malta
Table A.50.
Average re‐export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Italy and Malta via Germany and Luxembourg stratified by season. Cells marked in grey are not used to calculate the mean and standard deviation (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses (pcs) from Germany and Luxembourg to the EU area climatically suitable | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Importer: Italy and Malta | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 14,2173 | 18,132 | 116,884 | 2,616 | 90,308 | 180,388 | 2,024 | 260 | 157,418 | 840,569 | 155,077 | 250,723 |
| Spring | 47,981 | 5,907 | 52,842 | 2,149 | 2,050 | 21,883 | 45 | 488,427 | 255,100 | 898,987 | 177,537 | 298,559 |
| Summer | 19,213 | 3,149 | 17,411 | 3,186 | 2,299 | 133 | 4,591 | 3,493 | 222,278 | 961,210 | 123,696 | 302,040 |
| Autumn | 23,535 | 70,493 | 10,522 | 6,023 | 5,499 | 1,014 | 1,841 | 2 | 265,425 | 1,185,830 | 157,018 | 370,555 |
| Total | 232,902 | 97,681 | 197,659 | 13,974 | 100,156 | 203,418 | 8,501 | 492,182 | 900,221 | 3,886,596 | ||
A.6.1.4. Export from Germany and Luxembourg to Slovenia
Table A.51.
Average re‐export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Slovenia via Germany and Luxembourg stratified by season. Cells marked in grey are not used to calculate the mean and standard deviation (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses [pcs] from Germany and Luxembourg to the EU area climatically suitable | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Importer: Slovenia | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 8,462 | 10,390 | 9,320 | 9,240 | 2,280 | 0 | 0 | 3,150 | 0 | 0 | 0 | 0 |
| Spring | 16,380 | 22,754 | 1,600 | 18,230 | 600 | 0 | 2,110 | 0 | 4,225 | 0 | 0 | 0 |
| Summer | 1,050 | 10,940 | 7,500 | 480 | 0 | 0 | 0 | 0 | 817 | 1,340 | 0 | 0 |
| Autumn | 9,125 | 24,292 | 3,690 | 6,600 | 0 | 0 | 750 | 550 | 0 | 3,850 | 0 | 0 |
| Total | 35,017 | 68,376 | 22,110 | 34,550 | 2,880 | 0 | 2,860 | 3,700 | 5,042 | 5,190 | ||
A.6.1.5. Export from Germany and Luxembourg to Croatia
Table A.52.
Average re‐export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Croatia via Germany and Luxembourg stratified by season. Cells marked in grey are not used to calculate the mean and standard deviation (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses (pcs) from Germany and Luxembourg to the EU area climatically suitable | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Importer: Croatia | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 13,000 | 0 | 0 | 0 | 0 | 0 | 0 | 208,780 | 45,552 | 0 | 0 | 0 |
| Spring | 0 | 3,660 | 60 | 0 | 0 | 0 | 3,150 | 338,338 | 7,084 | 384 | 0 | 0 |
| Summer | 640 | 8,520 | 220 | 0 | 0 | 0 | 0 | 36,790 | 0 | 18,144 | 0 | 0 |
| Autumn | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 171,964 | 0 | 33,180 | 0 | 0 |
| Total | 13,640 | 12,180 | 280 | 0 | 0 | 0 | 3,150 | 755,872 | 52,636 | 51,708 | ||
A.6.1.6. Export from Germany and Luxembourg to Greece and Cyprus
Table A.53.
Average re‐export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Greece and Cyprus via Germany and Luxembourg stratified by season. Cells marked in grey are not used to calculate the mean and standard deviation (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses (pcs) from Germany and Luxembourg to the EU area climatically suitable | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Importer: Greece and Cyprus | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 22,225 | 100,193 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 12 | 0 | 0 |
| Spring | 8,080 | 25 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Summer | 200 | 25 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Autumn | 16,108 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Total | 46,613 | 100,243 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 12 | ||
A.6.1.7. Export from Germany and Luxembourg to Hungary
Table A.54.
Average re‐export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Hungary via Germany and Luxembourg stratified by season. Cells marked in grey are not used to calculate the mean and standard deviation (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses (pcs) from Germany and Luxembourg to the EU area climatically suitable | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Importer: Hungary | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 2,360 | 25,832 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Spring | 0 | 0 | 0 | 0 | 0 | 72,000 | 0 | 0 | 0 | 0 | 0 | 0 |
| Summer | 0 | 65,100 | 0 | 468 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Autumn | 0 | 200 | 14,378 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Total | 2,360 | 91,132 | 14,378 | 468 | 0 | 72,000 | 0 | 0 | 0 | 0 | ||
A.6.1.8. Export from Germany and Luxembourg to Bulgaria
Table A.55.
Average re‐export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Bulgaria via Germany and Luxembourg stratified by season. Cells marked in grey are not used to calculate the mean and standard deviation. (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses (pcs) from Germany and Luxembourg to the EU area climatically suitable | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Importer: Bulgaria | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 48,785 | 14,600 | 0 | 0 | 0 | 1,000 | 6,886 | 0 | 0 | 0 | 0 | 0 |
| Spring | 39,160 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Summer | 17,375 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 314 | 0 | 0 | 0 |
| Autumn | 23,655 | 0 | 0 | 0 | 0 | 12,060 | 0 | 0 | 433 | 0 | 0 | 0 |
| Total | 128,975 | 14,600 | 0 | 0 | 0 | 13,060 | 6,886 | 0 | 747 | 0 | ||
A.6.1.9. Export from Germany and Luxembourg to Romania
Table A.56.
Average re‐export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Romania via Germany and Luxembourg stratified by season. Cells marked in grey are not used to calculate the mean and standard deviation (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses [pcs] from Germany and Luxembourg to the EU area climatically suitable | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Importer: Romania | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 11,730 | 10,691 | 7,120 | 0 | 0 | 0 | 976,326 | 0 | 13,152 | 2,940 | 0 | 0 |
| Spring | 10,678 | 10,298 | 0 | 410 | 0 | 648,000 | 0 | 0 | 0 | 3,836 | 0 | 0 |
| Summer | 26,638 | 123,752 | 45,360 | 720 | 0 | 146,920 | 0 | 92,136 | 3,864 | 0 | 0 | 0 |
| Autumn | 50,540 | 140,532 | 0 | 340 | 0 | 2,298,240 | 233,280 | 110 | 0 | 2,926 | 0 | 0 |
| Total | 99,586 | 285,273 | 52,480 | 1,470 | 0 | 3,093,160 | 1,209,606 | 92,246 | 17,016 | 9,702 | ||
A.6.2. Dilution of African cut roses during re‐export by Germany and Luxembourg
Similar to the dilution in the Netherland and Belgium a corresponding factor was estimated for Germany and Luxembourg.
While Germany and Luxembourg imported on annual average 273 million cut roses from African countries with FCM occurrence and Israel and 19 million from other third countries, the total import from countries where FCM is absent is 1,110 million roses (including EU countries, esp. the Netherlands and Belgium). The own production of cut roses in the Germany and Luxembourg is about 61 million.
A.6.2.1. Direct export from African countries with FCM occurrence and Israel to Germany and Luxembourg
Table A.57.
Average direct export of cut roses from African countries with reported presence of Thaumatotibia leucotreta and from Israel to Germany and Luxembourg stratified by season (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Direct export of cut roses [pcs] from African countries with FCM occurrence and from Israel to Germany and Luxembourg | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Exporter: African countries with reported occurrence of FCM and Israel | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 66,183,111 | 68,038,967 | 63,033,999 | 58,479,160 | 65,858,076 | 61,729,455 | 57,970,454 | 61,629,019 | 52,061,581 | 77,339,328 | 63,232,315 | 6,812,138 |
| Spring | 77,470,073 | 77,402,267 | 70,941,533 | 69,233,447 | 86,184,881 | 74,817,527 | 76,730,243 | 71,251,024 | 84,301,324 | 62,516,130 | 75,084,845 | 7,040,119 |
| Summer | 70,033,416 | 60,103,519 | 70,507,279 | 64,234,869 | 73,989,473 | 68,801,088 | 72,301,862 | 59,117,096 | 74,002,658 | 54,502,781 | 66,759,404 | 6,861,288 |
| Autumn | 72,468,416 | 63,139,415 | 70,172,519 | 69,192,138 | 79,131,192 | 67,404,126 | 72,521,837 | 60,642,306 | 67,804,899 | 55,741,864 | 67,821,871 | 6,654,630 |
| Total | 286,155,016 | 268,684,168 | 274,655,330 | 261,139,614 | 305,163,622 | 272,752,196 | 279,524,396 | 252,639,445 | 278,170,462 | 250,100,103 | 272,898,435 | 16,254,249 |
A.6.2.2. Export from the non‐EU countries (different from African countries with FCM occurrence and Israel) to Germany and Luxembourg
Table A.58.
Average export of cut roses from countries outside the EU (excluding the African countries with reported presence of Thaumatotibia leucotreta and Israel) to Germany and Luxembourg stratified by season (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses [pcs] from Non‐EU countries (excluding African countries with FCM occurrence and Israel) to Germany and Luxembourg | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Exporter: All countries outside EU27 (2020) (excluding African countries with FCM occurrence and Israel) | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 10,381,450 | 8,329,155 | 6,366,583 | 7,355,775 | 5,769,171 | 4,104,950 | 4,112,952 | 3,661,479 | 2,858,588 | 2,395,018 | 5,533,512 | 2,581,063 |
| Spring | 8,805,197 | 7,412,268 | 7,106,249 | 7,435,264 | 6,223,055 | 3,571,263 | 4,144,927 | 3,761,527 | 3,110,066 | 1,285,485 | 5,285,530 | 2,427,199 |
| Summer | 6,051,951 | 4,883,054 | 4,743,628 | 6,279,209 | 4,236,482 | 3,236,675 | 2,875,200 | 2,620,271 | 1,870,404 | 894,182 | 3,769,106 | 1,769,923 |
| Autumn | 8,710,025 | 6,284,306 | 7,276,253 | 7,009,185 | 4,676,858 | 4,431,852 | 3,958,670 | 3,243,049 | 2,387,310 | 258,805 | 4,823,631 | 2,543,496 |
| Total | 33,948,623 | 26,908,783 | 25,492,713 | 28,079,433 | 20,905,566 | 15,344,740 | 15,091,749 | 13,286,326 | 10,226,368 | 4,833,490 | 19,411,779 | 9,137,744 |
A.6.2.3. Export from all countries (excluding African countries with FCM occurrence and Israel) to Germany and Luxembourg
Table A.59.
Average export of cut roses from all countries (including other EU countries; excluding African countries with reported presence of Thaumatotibia leucotreta and Israel) to Germany and Luxembourg stratified by season (Eurostat, EU trade since 1988 by HS2‐4‐6 and CN8, CN 06031100, monthly statistics 12‐2010‐12.2020, online, accessed on 13 January 2023)
| Season | Export of cut roses [pcs] from all countries (excluding African countries with FCM occurrence and Israel) to Germany and Luxembourg | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Exporter: All countries including EU27 (2020) (excluding African FCM countries and Israel) | ||||||||||||
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Mean | SD |
| Winter | 200,069,796 | 212,226,199 | 193,216,213 | 254,689,631 | 252,022,490 | 236,711,711 | 299,011,853 | 275,433,513 | 274,884,677 | 267,031,911 | 246,529,799 | 35,252,866 |
| Spring | 238,094,718 | 278,311,529 | 275,137,340 | 312,314,358 | 306,767,513 | 366,977,231 | 343,313,811 | 341,755,120 | 332,460,379 | 313,978,686 | 310,911,069 | 38,416,344 |
| Summer | 209,282,507 | 235,277,209 | 233,861,580 | 345,111,859 | 256,121,346 | 324,539,694 | 295,690,036 | 286,419,206 | 311,857,411 | 284,226,411 | 278,238,726 | 43,732,132 |
| Autumn | 205,049,967 | 224,776,785 | 255,457,710 | 273,628,807 | 255,962,295 | 299,543,671 | 306,875,394 | 310,841,076 | 294,433,230 | 320,510,954 | 274,707,989 | 38,747,980 |
| Total | 852,496,988 | 950,591,722 | 957,672,843 | 1,185,744,655 | 1,070,873,644 | 1,227,772,307 | 1,244,891,094 | 1,214,448,915 | 1,213,635,697 | 1,185,747,962 | 1,110,387,583 | 142,065,164 |
A.6.2.4. Own production of cut roses in Germany
The national statistical office of Germany (DEStatis:Zierpflanzenanbau Report 2021, downloaded on 1 February 2023) reports the area of cut roses production for certain years.
Similar data were not available for Luxembourg. Instead, the relation of area for all flower and ornamental production (Eurostat: Crops by classes of utilised agricultural area) in Germany and Luxembourg was used. It reports in 2016 no additional production area in Luxembourg.
To convert the production area into the number of produced cut roses, a productivity of 200,000 roses/ha was used. The estimation was informed by productivity figures of different types of roses, flowers per plant and plants per ha. The reported data ranges roughly from 150,000 to 320,000.
The mean values were used to calculate the average, annual production of cut roses in the Germany and Luxembourg:
Finally, the production is equally distributed on all four seasons (15,183,333 pcs/season), reasoned by a higher consumption in spring, but an assumed additional outdoor production in summer.
A.6.2.5. Dilution
For the calculation of the dilution rate, two scenarios were assessed:
• In scenario 1, Germany and Luxembourg get non‐infested roses from other countries than the African countries with FCM occurrence and Israel, including the intra‐EU trade, for re‐export. This gives a higher dilution and low dilution factor (proportion of African countries with FCM occurrence and Israel in intra‐EU trade re‐export).
• In scenario 2, the intra‐EU trade to Germany and Luxembourg is not re‐exported to further countries. This gives a lower dilution and higher dilution factor.
A.7. Infestation of cut roses at entry into the EU
A.7.1. Life stage of FCM at entry
The life stage at entry was assessed by Expert Knowledge Elicitation of two parameters:
• (Question 1a) the proportion of infested roses infested with eggs at border control;
• (Question 1b) the proportion of infested roses, which are not infested with eggs, but infested with young larvae (L1 or L2) at border control.
These life stages are difficult to detect. Especially early larval stages can only be detected by empty eggshells and entry holes.
It is assumed, that
• due to feeding damages and excrements the later larval stages (L3, L4 or L5) will be detected and sorted out at the place of origin;
• due to handling the pupae and adults are unlikely to be in imported cut flowers.
Following quantitative evidence was discussed during the EKE:
Cited from the Dutch report ‘Development stage of intercepted Thaumatotibia leucotreta specimens’ (NVWA, 2022: Development stage of intercepted Thaumatotibia leucotreta specimens; personal communication, 12 October 2022):
Results: Life stages of intercepted specimens of Thaumatotibia leucotreta on cut roses were recorded by the NPPO for 452 specimens in the period 2015–2018 and for 190 specimens in the period 2019–2022. The table below provides an overview of the life stages detected on cut roses. The results are summarised for all origins. The table indicates that, after T. leucotreta was regulated as a quarantine species, there has been a clear shift in the interceptions towards younger life‐stages. Since then, 50% of the interceptions concern eggs while the fraction of intercepted older larval stages (L2/L3 to L5) have decreased.
The table below provides an overview of the life stages detected at the entry points in the Netherlands on cut roses from the origins with most interceptions in in the period 2019–2022. For all origins, young life‐stages predominate, although the fraction of eggs varies.
Under the assumption of continuous infestation of the roses in the countries of origin in time and pressure, the development time of the (immature) life stages can be used as proxy for the proportion of infestations. For the larval life stages (L1–L5), equal durations were assumed in the calculation.
The table also shows that the profile of life stages is not influenced by the temperature.
No infestations with pupae or adults were recognised by the Dutch border control. All countries include interception from non‐African countries.
Combining the result of both elicitations gives the missing distribution of the later larval stages.
A.7.2. Infestation rate
The infestation rate of cut roses from African countries with reported occurrence of FCM (incl. Israel) at entry into the EU was assessed by Expert Knowledge Elicitation:
(Question 2) What is the proportion of cut roses (stems) imported from any African country, which are infested with any life stage of FCM directly after entering the EU for trade?
Following scenario was specified:
Definition ‘Infested rose’ A rose is considered infested, if one or more specimens of FCM (at any life stage) is attached to the rose
The infestation rate is assessed at time and place, when a consignment is entering Europe (place of control, e.g. airport, harbour) without any handling in Europe, e.g. grading, etc.
It is assumed that the profile of immature life stages of FCM infestation follows the result of EKE question 1
Some quantitative evidence was discussed and reviewed for the EKE.
Inspection rate
For the Dutch border inspection, the applied inspection rate is known for the years 2018–2021 (NVWA, 2022: Data on inspection frequencies for cut roses 2018–2021; personal communication, 12 October 2022):
For the countries with reduced inspection rate, the average inspection rate ranges between countries from 5.42% (Ethiopia) to 28.11% (Tanzania) with an overall rate of 10.51% (90%CI: 10.37–10.64%). Esp. for Uganda all consignments are inspected.
Sample size
The Dutch NPPO (NVWA, 2022: Infestation rates of consignments of cut roses infested by T. leucotreta; personal communication, 12 October 2022) indicates ‘The size of the infested consignments and sample size are not recorded in laboratory's database, so we cannot directly report the infestation rate of the inspected sample. However, we can provide an approximation of the infestation rate of T. leucotreta in intercepted consignments of cut roses. Based on import data from 2020, the median consignment weight of African roses is 635 kg. African roses a relatively light compared to European roses, being ca. 25 g per stem. This would amount to 25.400 roses in such a consignment. According to the inspection registry in the Netherlands, this would require a sample size of 400 inspected roses.’
This is not confirmed by the import data from Eurostat, which indicates a four to five times larger weight of the consignments, and two times larger number of roses [as Eurostat assumes a standard weight of 50.7 g/rose [Conversion factor for unit mass, CNVF2016, CN 06031100, online)].
Again, the average consignment sizes are varying between the countries from 36,798 roses (Tanzania) to 970,442 roses (Ethiopia) with an average of 58,645 roses per consignment.
The Dutch report gives only few indications on the actual sample size: ‘During import inspections in the Netherlands cut roses are inspected for the presence of T. leucotreta. The number of inspected roses is dependent on the size of the consignment and increases stepwise from 200 inspected roses in batches of 400–10,000 roses up to 1.200 inspected roses in batches of over 200,000 roses.’ (NVWA, 2022: Infestation rates of consignments of cut roses infested by T. leucotreta; personal communication, 12 October 2022).
By interpolation, we could assume following numbers of inspected roses.
Infestation sample
The number of specimens within an infested sample is described in the Dutch report (NVWA, 2022: Infestation rates of consignments of cut‐roses infested by T. leucotreta; personal communication, 12 October 2022) for all countries and years as in Table A.73 below.
Infestation per consignment
This would lead to following infestation rates within a consignment.
Interception rate
Finally, the Dutch report gives the interceptions with FCM per country and year (NVWA, 2022: Data on inspection frequencies for cut roses 2018–2021; personal communication, 12 October 2022). This gives following interception rates (see Table A.76 below).
The average interception rate (at consignment level) varies by country from 0.69% (Kenia) to 6.5% (Zambia) with an overall average of 0.91% (95%‐CI 0.78–1.04%).
Total infestation rate
To combine the different rates, a country‐specific approach was done for the full period from 2018 to 2021. Because individual data on the consignment size were not available, all calculations are based on the country‐specific average consignment size (according to Eurostat, see Table A.69).
The infestation rate within the inspected consignments is given by:
with
| Abbr. | Description | Remarks |
| rinspected | Infestation rate within the inspected consignments [% roses] | |
| pintercepted | Proportion of intercepted consignments | Triangular (P2.5, est, P97.5) |
| r0 specimens | Detection limit according to the sample size | Uniform (0, P95) |
| qk spec | Proportion of interceptions with k specimens | Constant |
| rk spec | Infestation level of interceptions with k specimens | Triangular (P2.5, est, P97.5) |
The total infestation rate is assuming that intercepted consignments are not imported, but the not inspected have the same infestation level as the inspected ones:
| Abbr. | Description | Remarks |
| rtotal | Infestation rate within total import [% roses] | |
| n | Total import of roses [no. roses] | Uniform (min, max) |
| u | Inspection rate | Triangular (P2.5, est, P97.5) |
| rinspected | Infestation rate within the inspected consignments [% roses] | Calculated |
| pintercepted | Proportion of intercepted consignments | Triangular (P2.5, est, P97.5) |
| r0 specimens | Detection limit according to the sample size | Uniform (0, P95) |
Simulation results
The infestation rate varies between the countries from 0.136% of the roses (Ethiopia) to 0.400% (Tanzania), with a trade‐weighted average of 0.319% (90%‐UI 0.074–0.564%).
Missing factors
Variation in trade, incl. size of consignments, different sampling numbers.
Detection level of visual inspection at border, esp. depending on the profile of life stages.
Correctness of assumed representative sampling at border control with interpolated sample size.
Calculations are done under the assumption that one specimen corresponds to one rose.
Calculations are limited to four countries and import only to NL.
Calculations are only for 4 years 2018–2021.
The simulated results and additional evidence were used to conduct an EKE on 12 December 2022 with following results.
A.7.2.1. Infestation rate estimated using a Poisson model
After the elicitation, the Panel reviewed the results and used a Poisson model to check the consistency of the elicited values.
The starting point were the observations from the Dutch NPPO on the number of specimens, when a sample indicates an interception (cp. Table A.73 [NVWA, 2022: Infestation rates of consignments of cut‐roses infested by T. leucotreta; personal communication, 12 October 2022]). On infested samples (n = 400 cut roses with one or more specimens), the NPPO reported on average 1.31 specimens (Table A.81, 2nd column).
Using the method of moments to fit a Poisson distribution to the observations under the condition of infested samples (k ≥ 1) leads to following equation:
and results in a mean of the unconditional Poisson distribution of mu400 = 0.57 (Table A.81, 4th column). Assuming that the specimens are uniformly distributed inside the sample would lead to a Poisson distribution for individual roses (n = 1, Table A.81, 6th column) with a mean value of:
The likelihood of infested roses (one or more specimens) can be calculated in this model as:
which is consistent with the elicitation results.
A.8. FCM development after entry
The time to develop different life stages of FCM is reported in literature (see Section 3.1.1 of the opinion). For the developmental model, following times were taken to form TRIANGULAR distributions for the uncertainties.
No development is assumed below 8°C.
The reciprocal value gives the developmental progress per day.
Values for temperatures between the given temperatures were linearly interpolated.
A.8.1. Seasonal ambient temperatures in the climatically suitable area
To estimate the climatic situation in the different climate suitability classes, five European locations were selected, which represent the situation at the limits of the classes (see table below).
These led to following temperature ranges, which were used in the developmental model to estimate the escape of FCM at different days after entry:
The detailed development over time is given by the profiles.
A.8.2. Proportion of insects developing to adults before the waste treatment
The waste model includes the proportion of adults, which will escape the cut rose, before it is treated by different forms of waste treatment. This proportion is influenced by the development time of Thaumatotibia leucotreta, esp. the temperatures in the compartments B (climatised) and C (ambient temperatures, depending on the season), and the time between initial disposal by the consumer and the waste treatment. Because of lacking information on the duration between disposal and treatment, four scenarios are defined: 3 days, 7 days, 14 days and 28 days. 3 days can be seen as best case, while 28 days is a reasonable worst case.
For the calculation, a mean time of 15 days until initial disposal wasting was used.
Which resulted in following escape rates of FCM adults before the waste treatment.
A.9. Waste treatment in the climatically suitable area
The main characteristics of the different treatments for the organic fraction of solid waste (landfill, composting, anaerobic digestion and incineration) are briefly described in this section. This information was used as part of the evidence for the expert judgement on: (i) Proportion of organic waste going to private composting (A.9.1), (ii) Survival rate of FCM at landfill (A.9.4), composting (A.9.5) and anaerobic digestion and incineration (A.9.6).
Landfill
According to the Landfill Directive (LD) (1999/31/EC, 2018/850/EC) landfill is defined as: ‘a waste disposal site for the deposit of the waste onto or into land (i.e. underground)’. The definition does not include (i) facilities where waste is unloaded in preparation for further transport for recovery, treatment or disposal elsewhere, (ii) storage of waste prior to recovery or treatment for a period less than 3 years as a general rule or (iii) storage of waste prior to disposal for a period less than 1 year.
Landfilling is considered as the least preferable option by the EU Waste Framework Directive (WFD) (2008/98/EC 2018/851/CE) and should be limited to the necessary minimum. Landfilling of bio‐waste is explicitly addressed in the LD which requires Member States (MS) to adopt national strategies for the progressive reduction of biodegradable municipal waste (BMW) going to landfill by means of in particular, recycling, composting, biogas production or material/energy recovery. According to the LD, by 2035, all MS should reduce the amount of municipal waste sent to landfill to 10% or less of the total municipal waste generated. As reported by the European Environment Agency (EEA), implementation of these directives so far resulted in a decrease in the landfill rate (waste sent to landfill as a proportion of waste generated) from 23% to 16% between 2010 and 2020 in EU‐27 and an overall 27.5% reduction of the total amount of waste sent to landfill in the same period. However, it should be noted that the municipal waste landfill rates are highly variable across MS with differences ranging from close to 0 to 90% (EEA) https://www.eea.europa.eu/ims/diversion-of-waste-from-landfill).
In article 6(a) of LD, it is specified that only waste that has been subjected to a pretreatment to reduce the volume or hazardous nature, facilitate its handling or enhance recovery may be placed into landfill. The Biodegradable municipal waste (BMW) going to landfill as part of the municipal solid waste (MSW) is therefore assumed to follow the typical landfilling operations for non‐hazardous waste involving: (1) waste dumping at the working face, (2) waste spreading, shredding and compaction and (3) waste covering (Chandrappa and Das, 2012).
Upon arrival at the landfill site, MSW is generally compacted using bulldozers or other compaction vehicle to spread the waste evenly and optimise the usable landfill space. The densely compacted waste should then be covered with at least 15 cm of soil or other approved material at the end of the working day to reduce odours, prevent dispersion of the waste and deter scavengers. Once the landfill has reached the permitted height, a final cover consisting of an engineered barrier of clay, soil or other materials to seal in the waste is constructed and laid over the whole landfill (Boase, 1999, Chen et al., 2011).
Industrial composting
Composting, defined as ‘the controlled biological decomposition of organic material in the presence of air to form a humus‐like material’. (EEA, online) involves % of the separately collected bio‐waste in Europe and consists of four main phases: (1) Reception and preparation (sorting, shredding, homogenising), (2) Aerobic degradation, (3) Maturation and (4) Finalisation (size classification, removal of impurities) with the las two steps not always involved (EC, 2018).
Most common industrial composting methods are (i) windrow, an outdoor composting system consisting in piles of compost that are aerated either manually or mechanically turning the piles; (ii) aerated static pile (ASP), a system that makes use of forced aeration to push or pull air through the pile core and (iii) in‐vessel composting, a system where the waste is enclosed in vessels or containers and environmental conditions are strictly controlled by the waste management facilities (Lim et al., 2017). A combination of the composting systems is also possible, e.g. the waste can first spend short time in in‐vessel for fast degradation followed by continuous degradation and stabilisation in a windrow (Lim et al., 2017).
Composting is a microbiological process facilitated by bacteria and fungi and under controlled conditions, it proceeds through three phases: (i) the mesophilic phase, normally lasting for a couple of days and normally reaching about 45°C, (ii) the thermophilic phase, characterised by an increase in the temperature up to about 70°C and lasting from a few days to several months (depending on the size of the system and the composition of the initial substrate) and, finally, (iii) a cooling and maturation phase (Nikoloudakis et al., 2018; Palaniveloo et al., 2020). The maturation time strongly depends on the conditions during composting and can take as long as 6–12 months (Chen et al., 2011) or as short as 14–21 days (Raabe, 2001).
Home composting
While the industrial composting is a well‐defined process taking place under controlled conditions in authorised plants, home composting is a gardening practise, carried out by citizens on a voluntary basis not entailing specific permits or obligations. No precise definition exists for home composting; the practice is normally understood as a cooler aerobic breakdown of domestic organic waste (dumping, feeding, etc.). The composting typically takes place in small‐scale composters and by ‘slow‐stack’ treatment methods (European bioplastics, 2015), for this reason, home composting is normally done by garden owners that can use the final product (i.e. the compost) as a fertiliser. Temperatures reached during home composting are generally lower than those reached in industrial settings, normally not controlled and following much more fluctuating pattern (Colón et al., 2010, Tatàno et al., 2015).
Due to the general lack of regulatory drivers to promote home composting at European level and the domestic nature of the practice, there is a substantial lack of data in terms of prevalence and volume of organic waste treated as home composting. A number of MS promote home composting as a strategy to reduce the amount of biodegradable waste going to landfill, but the uptake of this practice remains challenging to estimate and absence of harmonised collection of information makes country‐level data difficult to compare. For example, a proportion ranging from 5% to 10% of the households are reportedly active in home composting in the Netherlands while it seems that about 14% of the household waste treated biologically in 2012 was home composted in Sweden (Sulewski et al., 2021). Again, the statistical office of Slovenia reported 48% of people having a home composting system in 2015 (Žitnik and Vidic, 2016) while in Italy, the national aggregated value for the amount of waste destined for home composting in 2020 was 3.8% of the separately collected organic waste (ISPRA, 2021). Based on 2012 aggregate data from Austria, Denmark, Estonia, Finland, Germany, Ireland, Sweden and the UK, Stenmarck et al. estimated a home composting rate of 8% of the total household food waste (Stenmarck et al., 2016).
Anaerobic digestion
Anaerobic digestion (AD) is another decomposition process mediated by bacteria. It consists in a series of biochemical reactions where bacteria break down the organic matter of any substrate into a gaseous mixture (CH4, CO2, H2, H2S, etc.) in the absence of free oxygen. To ensure anaerobic environment, AD occurs in a closed tank or vessel, often called digester (Uddin and Wright, 2022).
The biochemical AD reactions can be divided into four distinct stages: (i) hydrolysis, (ii) acidogenesis, (iii) acetogenesis and (iv) methanogenesis (Uddin and Wright, 2022). The methanogenesis stage is anaerobic; therefore, the digesters are covered to ensure anaerobic conditions (Uddin and Wright, 2022). Common feedstock for AD (the biodegradable biomass materials used in AD) includes agricultural waste (including livestock manure), industrial agronomic residues and the organic fraction of municipal solid waste. Co‐digestion of different substrates is often used to avoid nutrient imbalances/process inhibition (Uddin and Wright, 2022). In order to enhance the performance of the AD processes, a range of mechanical, chemical, physical or thermal pretreatments can be used (alone or in combination) to enable the cell constituents of the organic matter to become easily available to microorganisms for biogas production (Uddin and Wright, 2022). Final products of AD are the biogas and the digestate, which is the unconverted substrate normally used as a soil fertiliser.
Optimal operating temperature ranges for AD are 55–60°C and 35–40°C for digestion operated by thermophilic and mesophilic methanogenic bacteria, respectively (Uddin and Wright, 2022). The average residence time of the substrate in the digester varies with the amount/type of substrate itself and the operating temperature (Uddin and Wright, 2022); typical retention times are in the range of 10–40 days and 12–14 days for mesophilic and thermophilic digesters, respectively (Verma, 2002, Monnet, 2004).
Incineration
In general, any incineration facility will incorporate the following processes: waste storage and handling, pretreatment and waste preparation, combustion, air pollution control and residue (ash) handling (NRC, 2000).
In practice, the incoming waste is usually tipped from collection vehicles into reception chambers where non‐combustible objects are removed before the transfer into a concrete waterproof storage chamber (Boase, 1999; Neuwahl et al., 2019). The whole delivery area may be enclosed to limit the odour, noise and emissions from the waste (Neuwahl et al., 2019). The storage chambers usually have a capacity of 3–5 days of plant operational throughput; however, this is dependent on local factors and the specific nature of the waste (Neuwahl et al., 2019). During combustion, the reaction temperature ranges from 800°C to 1,450°C.
A.9.1. Proportion of private compost
Table A87.
Result report on EKE question 3
| Overview of the results of the Expert Knowledge Elicitation Question 3 | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | Proportion of organic waste going to private composting | ||||||||||||||
| Stratification | Countries in the climatically suitable area/only one average of the climatically suitable area | ||||||||||||||
| Question | Compared to the waste counted in the statistics of Eurostat, what additional proportion of organic waste is going to private composting? | ||||||||||||||
| Unit | [%], theoretical restriction: 0–100% | ||||||||||||||
| Results | P1% | P2.5% | P5% | P10% | P16.7% | P25% | P33.3% | P50% | P66.7% | P75% | P83.3% | P90% | P95% | P97.5% | P99% |
| Elicited values | 2% | 8% | 13% | 25% | 40% | ||||||||||
| EKE results | 2.0% | 2.2% | 2.6% | 3.6% | 5.1% | 7.1% | 9.4% | 14% | 21% | 24% | 28% | 32% | 36% | 38% | 40% |
| Fitted distribution | BetaGeneral (0.85283, 1.555, 0.0188, 0.425) | ||||||||||||||
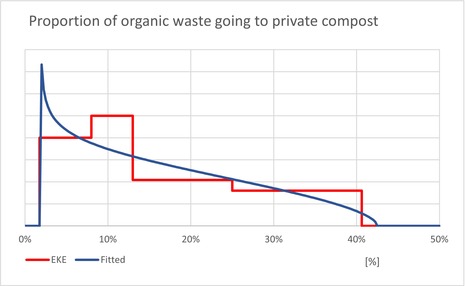
|
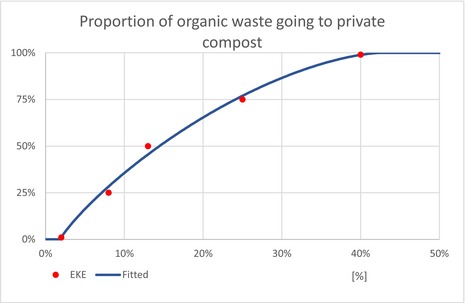
|
||||||||||||||
| Figure (a): Comparison of elicited and fitted values/density function to describe the remaining uncertainties of the parameter | Figure (b): Cumulative distribution function (CDF) of the likelihood of the parameter | ||||||||||||||
| Summary of the evidence used for the evaluation | |
| Main uncertainties | |
| |
| Reasoning for a scenario which would lead to a reasonable high proportion | The judgement on the upper limit considers that
|
| Reasoning for a scenario which would lead to a reasonable low proportion | The judgement on the lower limit considers that
|
| Fair estimate as judgement on the weighted evidence | The judgement on the median considers that
|
| Precision of the judgement as description of remaining uncertainties | The judgement on the interquartile range considers that
|
| Experts | Antoon LOOMANS, Panos MILONAS, Klara NICOVA, Giuseppe STANCANELLI, Hans‐Hermann THULKE |
| Facilitator/Reporter | Olaf MOSBACH‐SCHULZ (facilitator) |
| Observers | na |
| Date and place of the EKE | Virtual meeting on 9 January 2023 |
A.9.2. Proportion of treatments of commercial waste in the climatically suitable area
Eurostat reports the treatment of waste by waste category, hazardousness and waste management operations (Eurostat table: ENV_WASTRT (WST_OPER) in tonnes. The model uses most recent data of 2020 on ‘Vegetal waste’ to calculate the relative proportions:
• Landfill consists of ‘Disposal – landfill (D1, D5, D12)’; ‘Disposal – Other (D2‐D4, D6‐D7)’; and ‘Recovery – backfilling’
• Compost consists of ‘Recovery – recycling’
• Incineration, anaerobic digestion consist of ‘Disposal – incineration (D10)’; and ‘Recovery – Energy recovery (R1)’.
Hence, the model assumes, that in commercial situations the wasted cut flowers are only mixed with other vegetal waste.
A.9.3. Proportion of treatments of household waste in the climatically suitable area
As proxy for the proportion of different treatments of household waste, the model uses the same classification of Eurostat on household waste. Because the category ‘Recovery – recycling’ comprises several possibilities of recycling, this approach assumes similar recycling rates for all kind of household waste.
A.9.4. Result Survival rate of FCM at landfill
Table A.90.
report on EKE question 4a
| Overview of the results of the Expert Knowledge Elicitation Question 4a | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | Survival rate of FCM in household waste going to landfill | ||||||||||||||
| Stratification | |||||||||||||||
| Question | Assuming infestation of cut roses at the beginning of the waste treatment with later larval stages and pupae. What is the proportion of FCM which will develop to an adult and escape the waste treatment (landfill)? | ||||||||||||||
| Unit | [Escape out of 10,000] | ||||||||||||||
| Results | P1% | P2.5% | P5% | P10% | P16.7% | P25% | P33.3% | P50% | P66.7% | P75% | P83.3% | P90% | P95% | P97.5% | P99% |
| Elicited values | 0 | 10 | |||||||||||||
| EKE results | 0.1 | 0.3 | 0.5 | 1.0 | 1.7 | 2.5 | 3.4 | 5.1 | 6.7 | 7.6 | 8.4 | 9.1 | 9.6 | 9.8 | 10.0 |
| Fitted distribution | Uniform (0, 0.00101) | ||||||||||||||
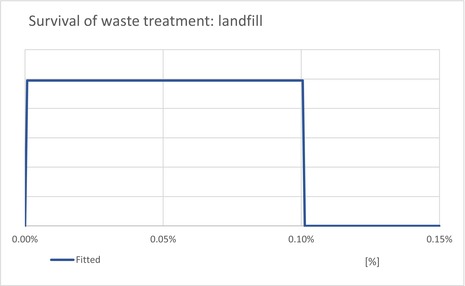
|
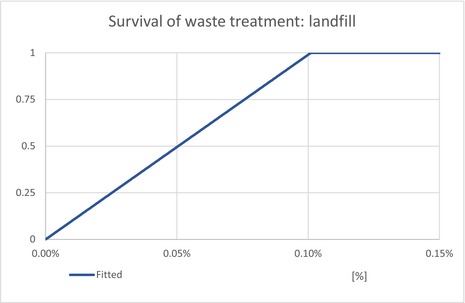
|
||||||||||||||
| Figure (a): Comparison of elicited and fitted values/density function to describe the remaining uncertainties of the parameter | Figure (b): Cumulative distribution function (CDF) of the likelihood of the parameter | ||||||||||||||
| Summary of the evidence used for the evaluation | |
| Main uncertainties | |
| |
| Reasoning for a scenario which would lead to a reasonable high proportion | The judgement on the upper limit considers that
|
| Reasoning for a scenario which would lead to a reasonable low proportion | The judgement on the lower limit considers that
|
| Fair estimate as judgement on the weighted evidence | The judgement on the median considers that
|
| Precision of the judgement as description of remaining uncertainties | The judgement on the interquartile range considers that
|
| Experts | Antoon LOOMANS, Panos MILONAS, Klara NICOVA, Stella PAPANASTASIOU, Giuseppe STANCANELLI, Hans‐Hermann THULKE |
| Facilitator/Reporter | Olaf MOSBACH‐SCHULZ (facilitator) |
| Observers | Na |
| Date and place of the EKE | Virtual meeting on 9 January 2023 |
A.9.5. Survival rate of FCM at composting
Table A.91.
Result report on EKE question 4b
| Overview of the results of the Expert Knowledge Elicitation Question 4b | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | Survival rate of FCM in household waste going to compost | ||||||||||||||
| Stratification | |||||||||||||||
| Question | Assuming infestation of cut roses at the beginning of the waste treatment with later larval stages and pupae. What is the proportion of FCM which will develop to an adult and escape the waste treatment (compost)? | ||||||||||||||
| Unit | [Escape out of 10,000] | ||||||||||||||
| Results | P1% | P2.5% | P5% | P10% | P16.7% | P25% | P33.3% | P50% | P66.7% | P75% | P83.3% | P90% | P95% | P97.5% | P99% |
| Elicited values | 0 | 1 | |||||||||||||
| EKE results | 0.01 | 0.03 | 0.05 | 0.10 | 0.17 | 0.25 | 0.34 | 0.51 | 0.67 | 0.76 | 0.84 | 0.91 | 0.96 | 0.98 | 1.00 |
| Fitted distribution | |||||||||||||||

|
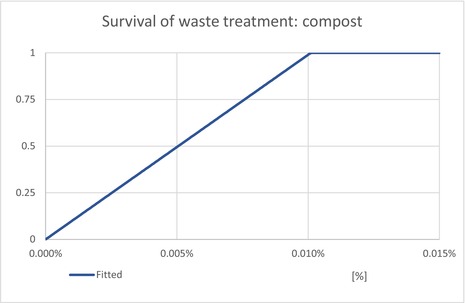
|
||||||||||||||
| Figure (a): Comparison of elicited and fitted values/density function to describe the remaining uncertainties of the parameter | Figure (b): Cumulative distribution function (CDF) of the likelihood of the parameter | ||||||||||||||
| Summary of the evidence used for the evaluation | |
| Main uncertainties | |
| |
| Reasoning for a scenario which would lead to a reasonable high proportion | The judgement on the upper limit considers that
|
| Reasoning for a scenario which would lead to a reasonable low proportion | The judgement on the lower limit considers that
|
| Fair estimate as judgement on the weighted evidence | The judgement on the median considers that
|
| Precision of the judgement as description of remaining uncertainties | The judgement on the interquartile range considers that
|
| Experts | Antoon LOOMANS, Panos MILONAS, Klara NICOVA, Stella PAPANASTASIOU, Giuseppe STANCANELLI, Hans‐Hermann THULKE |
| Facilitator/Reporter | Olaf MOSBACH‐SCHULZ (facilitator) |
| Observers | na |
| Date and place of the EKE | Virtual meeting on 9 January 2023 |
A.9.6. Survival rate of FCM at incineration and anaerobic digestion
Table A.92.
Result report on EKE question 4c
| Overview of the results of the Expert Knowledge Elicitation Question 4c | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | Survival rate of FCM in household waste going to incineration or anaerobic digestion | ||||||||||||||
| Stratification | |||||||||||||||
| Question | Assuming infestation of cut roses at the beginning of the waste treatment with later larval stages and pupae. What is the proportion of FCM which will develop to an adult and escape the waste treatment (incineration or anaerobic digestion)? | ||||||||||||||
| Unit | [Escape out of 10,000] | ||||||||||||||
| Results | P1% | P2.5% | P5% | P10% | P16.7% | P25% | P33.3% | P50% | P66.7% | P75% | P83.3% | P90% | P95% | P97.5% | P99% |
| Elicited values | 0 | 0 | |||||||||||||
| EKE results | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Fitted distribution | Constant(0) | ||||||||||||||
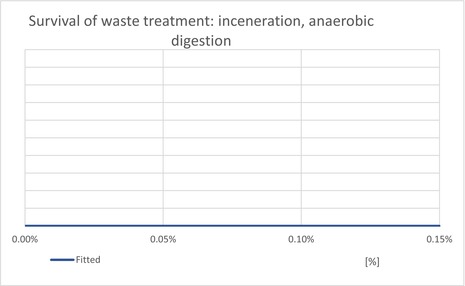
|
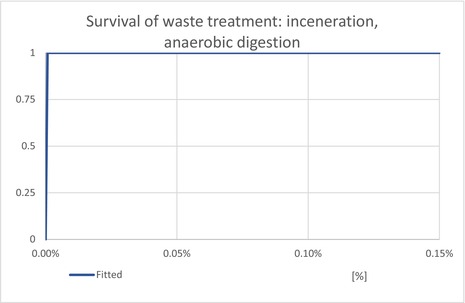
|
||||||||||||||
| Figure (a): Comparison of elicited and fitted values/density function to describe the remaining uncertainties of the parameter | Figure (b): Cumulative distribution function (CDF) of the likelihood of the parameter | ||||||||||||||
| Summary of the evidence used for the evaluation | |
| Main uncertainties | |
| |
| Reasoning for a scenario which would lead to a reasonable high proportion | The judgement on the upper limit considers that
|
| Reasoning for a scenario which would lead to a reasonable low proportion | The judgement on the lower limit considers that
|
| Fair estimate as judgement on the weighted evidence | The judgement on the median considers that
|
| Precision of the judgement as description of remaining uncertainties | The judgement on the interquartile range considers that
|
| Experts | Antoon LOOMANS, Panos MILONAS, Klara NICOVA, Stella PAPANASTASIOU, Giuseppe STANCANELLI, Hans‐Hermann THULKE |
| Facilitator/Reporter | Olaf MOSBACH‐SCHULZ (facilitator) |
| Observers | na |
| Date and place of the EKE | Virtual meeting on 9 January 2023 |
A.10. Natural developmental mortality
Finally, the number of escaped insects is corrected by a factor taking into account the mortality during the development of the larval and pupal stage. Numbers were taken from the reported mortality of larvae in citrus fruits between 25% and 37%. Assuming a similar natural mortality for the pupa stage gives an overall developmental mortality between 44% and 60%.
Appendix B – PBDM modelling of temperature‐driven dynamics of FCM Thaumatotibia leucotreta (Lepidoptera: Tortricidae), the false codling moth
Assessing the EU suitability for establishment of the exotic false codling moth (Thaumatotibia leucotreta) was a major part of the quantitative assessment of whether the importation of cut roses from East Africa provides a potential pathway for pest introduction in the EU. To determine this, a mechanistic temperature‐driven physiologically based demographic modelling (PBDM) was developed. The approach explicitly incorporates the mathematical description of the observed thermal biology of the pest to assess the phenology and dynamics of T. leucotreta populations in its native African range and its prospective range in the EU and the Mediterranean Basin.
Various modelling methods have been used to assess the suitability for potential establishment of pest species: Ecological niche models (ENMs) and physiologically based demographic models (PBDMs) are referenced in this text. ENMs characterise the ecological niche of T. leucotreta using aggregate weather and other factors in areas where the species has been recorded and use the model to predict the potential geographic range of the species in novel areas. The main advantage of ENM approaches is their relative simplicity to implement and low biological data requirements and, may provide an unbiased first screening of the potential range of invasive pests in new areas. The PBDM is used to capture the biology of T. leucotreta to predict prospectively its dynamics and distribution across vast geographic regions independent of occurrence records.
In its simplest form, the PBDM captures the time‐varying response of T. leucotreta to thermal variables as biodemographic functions (BDFs). The available data on the effects of temperature on T. leucotreta developmental rate, reproduction and mortality are illustrated in Box 1 and Section 3.1 of the opinion. These BDFs are imbedded in temperature‐driven population dynamics models and used to assess the EU suitability for potential establishment of T. leucotreta.
Age‐structured dynamics models that have been used to model the population dynamics of invasive species include the Manetsch (1976), Vansickle (1977) and forward Fokker‐Planck partial differential equation models (Buffoni and Pasquali, 2007). These models capture distributed maturation times of cohort members as observed in nature. The models also can imbed the time–temperature varying dynamics of holometabolous insect species having distinct stages: egg (e.g. e = eggs), larval (l), pupal (p) and adults (a) in an age specific manner. Individuals enter the first age class, say as an egg and develop through the life stages at temperature‐dependent rates exiting from the final age class, or via mortality from any age or stage.
The time invariant distributed‐maturation‐time demographic model (Manetsch, 1976) was used here to capture the dynamics of T. leucotreta. Absent extrinsic mortality, the theoretical distribution of cohort developmental times can be approximated using k age categories (i.e. Erlang parameter ), where var is the variance of average physiological developmental time D. The general form of the time invariant model is given for the ith of k age classes of a stage (left superscript s) with initial density in each age class . Using the notation of Di Cola et al. (1999, page 523) and ignoring the stage superscript, the age class dynamics is as follows:
| (1) |
where is the change in physiological age at time t and temperature T (see text Box 1), and is the net proportional loss rate that may be (+, −) and includes age‐specific components such as death rates, density‐dependent capacity‐related mortality and net immigration. In terms of flux, where , and
| (2) |
The density of the stage population at time t is
Potential total births b0(t) are the sum of adults' (stage A in Ak age classes) times the age specific reproduction () corrected for sex ratio (sr) and J limiting factors (0 ≤ fj < 1) such as temperature, relative humidity, nutrition with only temperature used in the T. leucotreta assessment:
| (3) |
The individuals enter the youngest age class (k = 1) of the first developmental stage, with forward flows (ageing) occurring between age classes and between stages, and adults exit as deaths latest at maximum adult age (i = Ak).
The numerical solution and the computations for the time invariant and time varying dynamics of the pest (and possibly any species in all trophic levels) are made with the same few lines of computer code (c.f. Abkin and Wolf, 1976; see Gutierrez, 1996, p. 157–159). Because of non‐linearities and the time varying nature, the model can only be evaluated numerically.
Regional analysis
The regional analysis of the geographic distribution and abundance of a species occurred as follows: (i) run the model using daily weather data (observed or climate model data) across all lattice cell in the landscape across all years, (ii) compute means, standard deviation and coefficient of variation for annual summary variables in each lattice cell and (iii) map the geo‐referenced data using GIS (e.g. GRASS, see Neteler et al., 2012).
Appendix C – Compartment model for FCM development in EU
The compartment model for Thaumatotibia leucotreta (Lepidoptera: Tortricidae) development in the EU is available in Excel format in the Supporting publication to this Scientific opinion.
Appendix D – Cut roses pathway model for import from Africa
1.
The cut roses pathway model from Africa is available in Excel format in the Supporting publication to this Scientific opinion.
Appendix E – EFSA Climate Suitability Analysis of Thaumatotibia leucotreta
1.
The EFSA Climate Suitability Analysis report is available on the Zenodo platform: https://doi.org/10.5281/zenodo.7648499 and it shall be cited as:
Rossi Eugenio, Campese Caterina, Maiorano Andrea, Picchi Malayka, Papanastassiou Stella, Muñoz Guajardo Irene and Stancanelli Giuseppe. (2023). EFSA Climate Suitability Analysis of Thaumatotibia leucotreta. Zenodo. https://doi.org/10.5281/zenodo.7648499
Supporting information
Compartment model for FCM development in EU
Cut roses pathway model for import from Africa
Suggested citation: EFSA PLH Panel (EFSA Panel on Plant Health) , Bragard, C ., Baptista, P ., Chatzivassiliou, E ., Di Serio, F ., Gonthier, P ., Jaques Miret, J. A ., Fejer Justesen, A ., MacLeod, A ., Magnusson, C. S ., Navas‐Cortes, J. A ., Parnell, S ., Potting, R ., Reignault, P. L ., Stefani, E ., Thulke, H. H ., Vicent Civera, A ., Van der Werf, W ., Yuen, J ., … Milonas, P . 2023. Assessment of the probability of introduction of Thaumatotibia leucotreta into the European Union with import of cut roses. EFSA Journal, 21(10), 1–166. 10.2903/j.efsa.2023.8107
Requestor: European Commission
Question number: EFSA‐Q‐2022‐00033
Panel members: Claude Bragard, Paula Baptista, Elisavet Chatzivassiliou, Francesco Di Serio, Paolo Gonthier, Josep Anton Jaques Miret, Annemarie Fejer Justesen, Alan MacLeod, Christer Sven Magnusson, Panagiotis Milonas, Juan A Navas‐Cortes, Stephen Parnell, Roel Potting, Philippe L Reignault, Emilio Stefani, Hans‐Hermann Thulke, Wopke Van der Werf, Antonio Vicent Civera, Jonathan Yuen and Lucia Zappalà.
Reproduction of the images listed below is prohibited and permission must be sought directly from the copyright holder: Copyright granted: Figure 27. By courtesy of Xingyu Li and Simon McKirdy, received by email on 13 January 2023.
Declarations of interest: If you wish to access the declaration of interests of any expert contributing to an EFSA scientific assessment, please contact interestmanagement@efsa.europa.eu.
Acknowledgements: EFSA and the EFSA Plant Health Panel wishes to thank the Netherlands Food and Consumer Product Safety Authority (NVWA) for provision of data and information on cut roses import, particularly regarding the sampling and inspection procedures, the infestation rates and the development stage of intercepted T. leucotreta specimens; the Belgian Federal Agency for the Safety of the Food Chain (FAVV‐AFSCA) for provision of data on interception and development stage of intercepted Thaumatotibia leucotreta specimens in cut roses import; the Kenya Plant Health Inspectorate Service (KEPHIS) for provision of report, data and information about T. leucotreta on cut roses and phytosanitary measures; the Ethiopian Agriculture Authority for provision of report on T. leucotreta on cut roses and phytosanitary measures; the Uganda Phytosanitary and Quarantine Inspection Service of the Ministry of Agriculture Animal Industry and Fisheries for provision of information on T. leucotreta on cut roses and phytosanitary measures. EFSA and the EFSA Plant Health Panel wish to thank the following for the support provided to this scientific output: Caterina Campese for her contribution to the systematic literature search on T. leucotreta distribution and the development of the Köppen–Geiger maps for T. leucotreta; Klara Nicova for her contribution on the collection and presentation of information and data on the waste management processes in the EU; Stella Papanastasiou for her contribution to the collection and analysis of data on T. leucotreta biology and occurrence from scientific literature and on T. leucotreta interceptions from the EUROPHYT and TRACES databases; Ewelina Czwienczek and Alzbeta Mikulova for their kind advice and precious support in the final editorial work.
Disclaimer: The designations employed and the presentation of material on the maps included in this Scientific Opinion do not imply the expression of any opinion whatsoever on the part of the European Food Safety Authority concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries.
EFSA may include images or other content for which it does not hold copyright. In such cases, EFSA indicates the copyright holder and users should seek permission to reproduce the content from the original source.
Adopted: 28 April 2023
Notes
Imported from specified countries of origin: Columbia, Ecuador, Ethiopia, Kenya, Tanzania, Zambia.
Definition of incursion (as per ISPM 5): ‘An isolated population of a pest recently detected in an area, not known to be established, but expected to survive for the immediate future [ICPM, 2003]’.
Establishment: Perpetuation, for the foreseeable future, of a pest within an area after entry (IPPC Secretariat, 2022).
‘Vegetal waste’ comprises ‘Green waste’ and ‘Vegetal waste of food production and products’ according EU waste statistics (EC 2150/2002, amended EC 849/2002 Annex III).
References
- Abkin MH and Wolf C, 1976. Computer library for agricultural systems simulation. Distributed delay routines: DEL, DELS, DELF, DELLF, DELVF, DELLVF. Department of Agricultural Economics, Michigan State University. Available online: https://pdf.usaid.gov/pdf_docs/pnaae013.pdf [Google Scholar]
- Adom M and Fening K, 2021. Pest status, bio‐ecology and management of the false codling moth, Thaumatotibia leucotreta (Meyrick) (Lepidoptera: Tortricidae) and its implication for international trade. Bulletin of Entomological Research, 111, 17–30. [DOI] [PubMed] [Google Scholar]
- Ali‐Ahmed D, Soltani N, Kellouche A and Mazouzi F, 2007. Effects of the soil texture and the burying depth of the larvae on some biological parameters of Ceratitis capitata (Diptera: Trypetidae). African Journal of Agricultural Research, 2, 105–111. [Google Scholar]
- Baker RHA, 2002. Predicting the limits to the potential distribution of alien crop pests. In: Hallman GJ and Schwalbe CP (eds.), Invasive Arthropods in Agriculture. Problems and solutions, pp. 207–241. Enfield, USA, Science Publishers Inc. 450 pp. [Google Scholar]
- Barker B and Coop C, 2019. False Codling Moth ‐ Thaumatotibia leucotreta (Lepidoptera: Tortricidae) ‐Phenology/Degree‐Day and Climate Suitability Model Analysis for USPEST.ORG. ‐ Prepared for USDA APHIS PPQ Version 1.0. 2 December 2019, 13 pp. Available online: https://uspest.org/CAPS/Thaumatotibia_leucotreta_white_paper.pdf
- Baughman WB, Nelson PN and Grieshop MJ, 2015. Impact of cultivation and subsequent Burial on Cydia pomonella (Lepidoptera: Tortricidae) and Conotrachelus nenuphar (Coleoptera: Curculionidae). Horticultural Entomology. Journal of Economic Entomology, 108, 1215–1220. 10.1093/jee/tov071 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boardman L, Grout TG and Terblanche JS, 2012. False codling moth Thaumatotibia leucotreta (Lepidoptera, Tortricidae) larvae are chill‐susceptible. Insect Science, 19, 315–328. 10.1111/j.1744-7917.2011.01464.x [DOI] [Google Scholar]
- Boardman L, Sørensen JG and Terblanche JS, 2013. Physiological responses to fluctuating thermal and hydration regimes in the chill susceptible insect, Thaumatotibia leucotreta . Journal of Insect Physiology, 59, 781–794. 10.1016/j.jinsphys.2013.05.005 [DOI] [PubMed] [Google Scholar]
- Boase CJ, 1999. Trends in urban refuse disposal: a pest's perspective. International Conference on Urban Pests, 83–98. [Google Scholar]
- Boersma N, 2018. Impacts of temperature variation on performance, life‐history and flight ability of the false codling moth, Thaumatotibia leucotreta (Lepidoptera: Tortricidae). Stellenbosch University, Stellenbosch. [Google Scholar]
- Buffoni G and Pasquali S, 2007. Structured population dynamics: continuous size and discontinuous stage structures. Journal of Mathematical Biology, 54, 555–595. 10.1007/s00285-006-0058-2 [DOI] [PubMed] [Google Scholar]
- Carrier Transicold . 2022. Available online: https://www.carrier.com/truck-trailer/ and https://www.youtube.com/embed/ylzV3jKlX20
- CBI (Centre for the Promotion of Imports from developing countries) , 2017. Exporting roses to Europe. Version of 13th June 2017, downloaded on 21st April 2022. Available online: https://www.cbi.eu/market-information/cut-flowers-foliage/roses/europe
- CBS , 2022. Landbouw; gewassen, dieren, grondgebruik en arbeid op nationaal niveau. Published September 30, 2022. Available online: https://opendata.cbs.nl/statline/#/CBS/nl/dataset/81302ned/table?dl=6BCD5 [Accessed: 12 October 2022]
- Chandrappa R and Das DB, 2012. In: Allan R, Forstner U and Salomons W (eds). Solid waste management: principles and practice. 1st Edition. Springer, Berlin, Heidelberg. 10.1007/978-3-642-28681-0 [DOI] [Google Scholar]
- Chen Z, Gong H, Zhang M, Wu W, Liu Y and Feng J, 2011. Impact of using high‐density polyethylene geomembrane layer as landfill intermediate cover on landfill gas extraction. Waste Management, 31, 1059–1064. 10.1016/j.wasman.2010.12.012 [DOI] [PubMed] [Google Scholar]
- COLEACP et al., 2020. Protocols for the management of the false codling moth (Thaumatotibia leucotreta) in roses in Kenya. Available online: https://fpeak.org/wp-content/uploads/2021/05/FCM-Manual.pdf [Accessed: 28 April 2023]
- Colón J, Martínez‐Blanco J, Gabarrell X, Artola A, Sánchez A, Rieradevall J and Font X, 2010. Environmental assessment of home composting. Resources, Conservation and Recycling, 54, 893–904. 10.1016/j.resconrec.2010.01.008 [DOI] [Google Scholar]
- Daiber CC, 1976. A survey of the false codling moth (Cryptophlebia leucotreta Meyr.) in peach orchards. Phytophylactica, 8, 96–102. [Google Scholar]
- Daiber CC, 1979a. A study of the biology of the false codling moth [Cryptophlebia leucotreta (Meyr.)]: The egg. Phytophylactica, 11, 129–132. 10.10520/AJA03701263_523 [DOI] [Google Scholar]
- Daiber CC, 1979b. A study of the biology of the false codling moth [Cryptophlebia leucotreta (Meyr.)]: The larva. Phytophylactica, 11, 141–144. 10.10520/AJA03701263_526 [DOI] [Google Scholar]
- Daiber CC, 1979c. A study of the biology of the false codling moth [Cryptophlebia leucotreta (Meyr.)]: The cocoon. Phytophylactica, 11, 151–157. 10.10520/AJA03701263_496 [DOI] [Google Scholar]
- Daiber CC, 1980. A study of the biology of the false codling moth Cryptophlebia leucotreta (Meyr.): the adult and generations during the year. Phytophylactica, 12, 187–194. 10.10520/AJA03701263_482 [DOI] [Google Scholar]
- Daiber K, 1989. Die falsche Obstmade, Cryptophlebia leucotreta (Meyr.) (Lepidoptera: Tortricidae), im südlichen Afrika/The false codling moth, Cryptophlebia leucotreta (Meyr.) (Lepidoptera: Tortricidae), in southern Africa. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz/Journal of Plant Diseases and Protection: 71–80.
- De Jager ZM, 2013. Biology and ecology of the false codling moth, Thaumatotibia leucotreta (Meyrick), Stellenbosch. Stellenbosch University. [Google Scholar]
- Di Cola G, Gilioli G and Baumgärtner J, 1999. Mathematical models for age‐structured population dynamics. In: Huffaker CB and Gutierrez AP (eds). Ecological entomology. 2nd Edition. Wiley. [Google Scholar]
- EEA , online. Available online: https://www.eea.europa.eu/help/glossary/eea-glossary/composting [Accessed: 30 September 2022]
- EFSA (European Food Safety Authority) , 2014. Guidance on Expert Knowledge Elicitation in Food and Feed Safety Risk Assessment. EFSA Journal 2014;12(6):3734, 278 pp. 10.2903/j.efsa.2014.3734 [DOI] [Google Scholar]
- EFSA (European Food Safety Authority) , Loomans AJM, van Noort T, Schenk M, Delbianco A and Vos S, 2020. Pest survey card on Thaumatotibia leucotreta . EFSA supporting publication 2020;17(8):EN‐1916, 26 pp. 10.2903/sp.efsa.2020.EN-1916 [DOI] [Google Scholar]
- EFSA (European Food Safety Authority) and Maiorano A, 2022. Scientific Report on the SCAN‐Clim: a tool to support pest climate suitability analysis based on climate classification. EFSA Journal 2022;20(2):7104, 18 pp. 10.2903/j.efsa.2022.7104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health) , Jeger M, Bragard C, Caffier D, Candresse T, Chatzivassiliou E, Dehnen‐Schmutz K, Gregoire J‐C, Jaques Miret JA, MacLeod A, Navajas Navarro M, Niere B, Parnell S, Potting R, Rafoss T, Rossi V, Urek G, Van Bruggen A, Van Der Werf W, West J, Winter S, Hart A, Schans J, Schrader G, Suffert M, Kertesz V, Kozelska S, Mannino MR, Mosbach‐Schulz O, Pautasso M, Stancanelli G, Tramontini S, Vos S and Gilioli G, 2018. Guidance on quantitative pest risk assessment. EFSA Journal 2018;16(8):5350, 86 pp. 10.2903/j.efsa.2018.5350 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EPPO , 2013. Pest risk analysis for Thaumatotibia leucotreta. EPPO, Paris (modified in 2021 to clarify the phytosanitary measures recommended). Available online: https://www.eppo.int/QUARANTINE/Pest_Risk_Analysis/PRA_intro.htm
- EPPO , 2014. EPPO Reporting Service no. 07–2014 Num. article: 2014/126. Available online: https://gd.eppo.int/reporting/article-3219 https://gd.eppo.int/reporting/article-3219 https://gd.eppo.int/reporting/article-3219 https://gd.eppo.int/reporting/article-3219
- EPPO , 2018. EPPO Reporting Service no. 08–2018 Num. article: 2018/160. Available online: https://gd.eppo.int/reporting/article-6354
- EPPO (European and Mediterranean Plant Protection Organization) , online. EPPO Global Database. Available online: https://gd.eppo.int/ [Accessed: 17 February 2023]
- EPPO (European and Mediterranean Plant Protection Organization) , 2010. Isolated finding of Thaumatotibia (Cryptophlebia) leucotreta on Capsicum chinensis in the Netherlands. In: EPPO Reporting Service, (2010/013). Available online: https://archives.eppo.int/EPPOReporting/2010/Rse-1001.pdf
- European bioplastics , 2015. Home composting, fact sheet. Available online: https://docs.european-bioplastics.org/publications/pp/EUBP_PP_Home_composting.pdf
- European Commission , 2018. Best Available Techniques (BAT). Reference document for waste treatment. pp 365–366.
- FloraNews , 2015. De Nederlandse roos in cijfers. Available online: https://www.floranews.com/UserFiles/Nederlandse%20roos%20in%20cijfers.pdf https://www.floranews.com/UserFiles/Nederlandse%20roos%20in%20cijfers.pdf
- Flower Watch , 2011. Quality through the chain: Roses from Africa via The Netherlands to European retail. Productschap Tuinbouw. Available online: https://www.flowerwatch.com/assets/files/Whitepaper-FlowerWatch-Quality-Standards.pdf [Google Scholar]
- Gilligan TM, Epstein ME and Hoffman KM, 2011. Discovery of False Codling Moth, Thaumatotibia leucotreta (Meyrick), in California (Lepidoptera: Tortricidae). Proceedings of the Entomological Society of Washington, 113, 426–435. [Google Scholar]
- Giliomee JH and Riedl H, 1998. A century of codling moth control in South Africa I. Historical perspective. Journal of the Southern African Society for Horticultural Sciences, 8, 27–31. [Google Scholar]
- GRASS Development Team , 2022. Geographic Resources Analysis Support System (GRASS) Software, Version 8.2.0. Open Source Geospatial Foundation. Available online: http://grass.osgeo.org [Google Scholar]
- Grimm V, Johnston ASA, Thulke HH, Forbes VE and Thorbek P, 2020. Three questions to ask before using model outputs for decision support. Nature Communications, 11, 4959. 10.1038/s41467-020-17785-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gutierrez AP, 1992. The physiological basis of ratio‐dependent predator‐prey theory: the metabolic pool model as a paradigm. Ecology, 73, 1552–1563. 10.2307/1940008 [DOI] [Google Scholar]
- Gutierrez AP, 1996. Applied population ecology: a supply‐demand approach. John Wiley and Sons. Available online: https://www.wiley.com/en-us/Applied+Population+Ecology%3A+A+Supply+Demand+Approach-p-9780471135869 [Google Scholar]
- Gutierrez AP and Baumgärtner JU, 1984. Multitrophic level models of predator‐prey energetics: I. Age‐specific energetics models—Pea aphid Acyrthosiphon pisum (Homoptera: Aphididae) as an example. Canadian Entomologist, 116, 924–932. 10.4039/Ent116923-7 [DOI] [Google Scholar]
- Gutierrez AP and Ponti L, 2013. Eradication of invasive species: Why the biology matters. Environmental Entomology, 42, 395–411. 10.1603/EN12018 [DOI] [PubMed] [Google Scholar]
- Gutierrez AP and Ponti L, 2023a. Analysis of invasive insects: links to climate change. In Ziska LH (ed.), Invasive Species and Global Climate Change, 2nd Edition. pp. 50–73. CAB International. 10.1079/9781800621459.0004 [DOI] [Google Scholar]
- Gutierrez AP and Ponti L, 2023b. Assessing and managing the impact of climate change on an invasive weed, yellow starthistle. In: Ziska LH (ed.), Invasive Species and Global Climate Change, 2nd Edition. pp. 260–281. CAB International. 10.1079/9781800621459.0015 [DOI] [Google Scholar]
- Gutierrez AP, Ponti L, Cooper ML, Gilioli G, Baumgärtner J and Duso C, 2012. Prospective analysis of the invasive potential of the European grapevine moth Lobesia botrana (Den. & Schiff.) in California. Agricultural and Forest Entomology, 14, 225–238. 10.1111/j.1461-9563.2011.00566.x [DOI] [Google Scholar]
- Gutierrez AP, Ponti L, d'Oultremont T and Ellis CK, 2008. Climate change effects on poikilotherm tritrophic interactions. Climatic Change, 87, S167–S192. 10.1007/s10584-007-9379-4 [DOI] [Google Scholar]
- Gutierrez AP, Ponti L and Gilioli G, 2010. Climate change effects on plant‐pest‐natural enemy interactions. In: Hillel D and Rosenzweig C (eds). Handbook of Climate Change and Agroecosystems: Impacts, Adaptation, and Mitigation. Imperial College Press. pp. 209–237. 10.1142/9781848166561_0012 [DOI] [Google Scholar]
- Harkema H, Paillart M, Lukasse L, Westra E and Hogeveen E, 2017. Transport and storage of cut roses: endless possibilities? Guide of practice for sea freight of cut roses developed within GreenCHAINge project (No. 1699). Wageningen Food & Biobased Research. 10.18174/401918 [DOI] [Google Scholar]
- Hofmeyr JH, Carpenter JE, Bloem S, Slabbert JP, Hofmeyr M and Groenewald SS, 2015. Development of the sterile insect technique to suppress false codling moth Thaumatotibia leucotreta (Lepidoptera: Tortricidae) in citrus fruit: research to implementation (Part 1). African Entomology, 23, 180–186. [Google Scholar]
- Huisman KJ and Koster JC, 2000. New and interesting Microlepidoptera from The Netherlands in particular from the years 1997 and 1998 (Lepidoptera). (Nieuwe en interessante Microlepidoptera uit Nederland in hoofdzaak van de jaren 1997 en 1998 (Lepidoptera)). Entomologische Berichten, 60, 193–216. [Google Scholar]
- Ichimura K, Taguchi M and Norikoshi R, 2006. Extension of the vase life in cut roses by treatment with glucose, isothiazolinonic germicide, citric acid and aluminum sulphate solution. Japan Agricultural Research Quarterly: JARQ, 40, 263–269. [Google Scholar]
- IPPC Secretariat . 2022. Glossary of phytosanitary terms. International Standard for Phytosanitary Measures No. 5. Rome. FAO on behalf of the Secretariat of the International Plant Protection Convention. Available online: https://www.fao.org/3/mc891e/mc891e.pdf
- ISPRA , 2021. Rapporto Rifiuti Urbani ‐ Edizione 2021. Available online: https://www.isprambiente.gov.it/it/pubblicazioni/rapporti/rapporto-rifiuti-urbani-edizione-2021 [Google Scholar]
- Karvonen J, 1983. Cryptophlebia leucotreta imported into Finland (Lepidoptera, Tortricidae). Notulae Entomologicae, 63, 94. [Google Scholar]
- Kelly A, 1914. The False codling moth (Enarmonia batrachopa, Meyrick) with particular reference to its attack upon acorns. South African Agricultural Journal, 8, 72–75. [Google Scholar]
- Knill‐Jones SA, 1994. Two species of micro‐lepidoptera new to the Isle of Wight. Entomologist's Record and Journal of Variation, 106, 114. [Google Scholar]
- Kottek M, Grieser J, Beck C, Rudolf B and Rubel F, 2006. World map of the Köppen‐Geiger climate classification updated. Meteorologische Zeitschrift, 15, 259–263. 10.1127/0941-2948/2006/0130 [DOI] [Google Scholar]
- Langmaid JR, 1996. Sitotroga cerealella (Olivier) (Lepidoptera: Gelechiidae) and Cryptophlebia leucotreta (Meyrick) (Lepidoptera: Tortricidae) at m.v. light in Hampshire. Entomologist's Gazette, 47, 50. [Google Scholar]
- Li X, Emery RN, Coupland GT, Ren Y and McKirdy SJ, 2022. Evaluation of the likelihood of establishing False Codling Moth (Thaumatotibia leucotreta) in Australia via the international cut flower market. Insects, 2022, 883. 10.3390/insects13100883 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lim LY, Bong CPC, Lee CT, Klemeš JJ, Sarmidi MR and Lim JS, 2017. Review on the current composting practices and the potential of improvement using two‐stage composting. Chemical Engineering Transactions, 61, 1051–1056. 10.3303/CET1761173 [DOI] [Google Scholar]
- Love CN, Moore SD and Hill MP, 2019. The role of abiotic factors in the pupation of Thaumatotibia leucotreta Meyrick (Lepidoptera: Tortricidae) in the soil. Agricultural and Forest Entomology, 21, 38–49. [Google Scholar]
- MacLeod A and Korycinska A, 2019. Detailing Köppen–Geiger climate zones at sub‐national to continental scale: a resource for pest risk analysis. EPPO Bulletin, 49, 73–82. 10.1111/eppo.12549 [DOI] [Google Scholar]
- Manetsch TJ, 1976. Time‐varying distributed delays and their use in aggregative models of large systems. IEEE Transactions on Systems, Man and Cybernetics, 6, 547–553. 10.1109/TSMC.1976.4309549 [DOI] [Google Scholar]
- Mikhailov A, 2020. Turbo, an improved rainbow colormap for visualization. Google AI Blog. Available online: https://ai.googleblog.com/2019/08/turbo-improved-rainbow-colormap-for.html [Accessed: 7 May 2021]. [Google Scholar]
- Milonas PG and Savopoulou‐Soultani M, 2000. Development, survivorship and reproduction of Adoxophyes orana (Lepidoptera: Tortricidae) at constant temperatures. Annals of the Entomological Society of America, 93, 96–102. 10.1603/0013-8746(2000)093[0096:DSAROA]2.0.CO;2 [DOI] [Google Scholar]
- Mkiga AM, Mohamed SA, du Plessis H, Khamis F and Ekesi S, 2019. Field and laboratory performance of False Codling Moth, Thaumatotibia leucotreta (Lepidoptera: Troticidae) on orange and selected vegetables. Insects, 10, 63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monnet F, 2004. An Introduction to Anaerobic Digestion of Organic Wastes. 2003. sl: Remade Scotland. Available online: https://www.cti2000.it/Bionett/BioG-2003-002%20IntroAnaerobicDigestion.pdf
- Moore SD, Kirkman W, Albertyn S and Hattingh V, 2016. Comparing the use of laboratory‐reared and field‐collected Thaumatotibia leucotreta (Lepidoptera: Tortricidae) larvae for demonstrating efficacy of postharvest cold treatments in citrus fruit. Journal of Economic Entomology, 109, 1571–1577. 10.1093/jee/tow137 [DOI] [PubMed] [Google Scholar]
- Moore SD, Peyper M, Kirkman W, Marsberg T, Albertyn S, Stephen PR, Thackeray SR, Grout TG, Sharp G, Sutton G and Hattingh V, 2022. Efficacy of various low temperature and exposure time combinations for Thaumatotibia leucotreta (Meyrick) (Lepidoptera: Tortricidae) larvae. Journal of Economic Entomology, 115, 1115–1128. 10.1093/jee/toac064 [DOI] [PubMed] [Google Scholar]
- Myburgh AC, 1965. Low temperature sterilization of false Codling Moth, Argyroploce leucotreta Meyr., in export citrus. Journal of the Entomological Society of Southern Africa, 28, 277–285. 10.10520/AJA00128789_3425 [DOI] [Google Scholar]
- National Research Council , 2000. Waste Incineration and Public Health. The National Academies Press. 10.17226/5803 [DOI] [PubMed] [Google Scholar]
- NBN Atlas , 1997. Human observation of Thaumatotibia leucotreta (Meyrick, 1913) | False Codling Moth recorded on 1997‐12‐31. Available online: https://records.nbnatlas.org/occurrences/633934f9-66db-4f52-8aef-eb7ced7e951f. [Accessed: 10 January 2023].
- Neteler M, Bowman MH, Landa M and Metz M, 2012. GRASS GIS: a multi‐purpose Open Source GIS. Environmental Modelling and Software, 31, 124–130. 10.1016/j.envsoft.2011.11.014 [DOI] [Google Scholar]
- Neuwahl F, Cusano G, Gómez Benavides J, Holbrook S and Roudier S, 2019. Best Available Techniques (BAT) Reference Document for Waste Incineration: Industrial Emissions Directive 2010/75/EU (Integrated Pollution Prevention and Control) (Issue KJ‐NA‐29971‐EN‐N (online)). Publications Office of the European Union. 10.2760/761437 [DOI] [Google Scholar]
- Nikoloudakis Y, Panagiotakis S, Manios T, Markakis E and Pallis E, 2018. Composting as a service: a real‐world IoT implementation. Future Internet, 10. 10.3390/fi10110107 [DOI] [Google Scholar]
- Odanga J, Mohamed S, Mwalusepo S, Olubayo F, Nyankanga R, Khamis F, Rwomushana I, Johansson T and Ekesi S, 2018. Spatial Distribution of Bactrocera dorsalis and Thaumatotibia leucotreta in Smallholder Avocado Orchards along Altitudinal Gradient of Taita Hills and Mount Kilimanjaro. Insects, 9, 71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palaniveloo K, Amran MA, Norhashim NA, Mohamad‐Fauzi N, Peng‐Hui F, Hui‐Wen L, Kai‐Lin Y and Razak SA, 2020. Food waste composting and microbial community structure profiling. Processes, 8, 723. [Google Scholar]
- Potting RPJ and van der Straten M, 2011. Pest Risk Analysis for Thaumatotibia leucotreta. Version 5 February 2011, 27 pp. Available online: https://www.researchgate.net/publication/277466540_Pest_Risk_Assessment_Thaumatotibia_leucotreta
- Raabe RD, 2001. The rapid composting method. Available online: https://vric.ucdavis.edu/pdf/compost_rapidcompost.pdf
- Rabobank , 2022. World Floriculture Map. Available online: https://research.rabobank.com/far/en/documents/175926_Rabobank_Flower-Map-2021_20211230.pdf
- Reda K and Szafir DA, 2021. Rainbows revisited: Modeling effective colormap design for graphical inference. IEEE Transactions on Visualization and Computer Graphics, 27, 1032–1042. 10.1109/TVCG.2020.3030439 [DOI] [PubMed] [Google Scholar]
- Rennwald E, 2022. Thaumatotibia leucotreta (Meyrick, 1913). Available online: https://lepiforum.org/wiki/page/Thaumatotibia_leucotreta
- Rogard J, 2015. Première signalisation en France de Thaumatotibia (Cryptophlebia) leucotetra [sic, recte: leucotreta] (Meyrick, 1913), le faux carpocapse (Lep. Tortricidae Olethreutinae). ‐ Oreina 30: 20. Available online: https://oreina.org/artemisiae/biblio/docpdf/Rogard2015-232.pdf [Accessed: 10 January 2023].
- Rossi E, Campese C, Maiorano A, Picchi M, Papanastassiou S, Muñoz Guajardo I and Stancanelli G, 2023. EFSA Climate Suitability Analysis of Thaumatotibia leucotreta . Zenodo. 10.5281/zenodo.7648499 [DOI] [Google Scholar]
- Royal Flora Auctions , 2021. Available online: https://www.royalfloraholland.com/en/news-2022/week-13/excellent-results-2021-concerns-about-the-future-of-the-floriculture-industry
- Ruane AC, Goldberg R and Chryssanthacopoulos J, 2015. Climate forcing datasets for agricultural modeling: Merged products for gap‐filling and historical climate series estimation. Agricultural and Forest Meteorology, 200, 233–248. 10.1016/j.agrformet.2014.09.016 [DOI] [Google Scholar]
- Rubel F, Brugger K, Haslinger K and Auer I, 2017. The climate of the European Alps: Shift of very high resolution Köppen‐Geiger climate zones 1800–2100. Meteorologische Zeitschrift, 26, 115–125. 10.1127/metz/2016/0816 [DOI] [Google Scholar]
- Schwartz A, 1981. 'n Bydrae tot die biologie en beheer van die valskodlingmot Cryptophlebia leucotreta (Meyr.) (Lepidoptera: Eucosmidae) op nawels, Stellenbosch: Stellenbosch University.
- Soderstrom EL, Brandl DG and Mackey B, 1990. Responses of codling moth (Lepidoptera: Tortricidae) life stages to high carbon dioxide or low oxygen atmospheres. Journal of Economic Entomology, 83, 472–475. [Google Scholar]
- Stenmarck A, Jensen C, Quested T, Moates G, Buksti M, Cseh B, Juul S, Parry A, Politano A, Redlingshofer B, Scherhaufer S, Silvennoinen K, Soethoudt H, Zübert C and Östergren K, 2016. Estimates of European food waste levels. Available online: https://www.eu-fusions.org/phocadownload/Publications/Estimates%20of%20European%20food%20waste%20levels.pdf
- Stotter RL and Terblanche JS, 2009. Low‐temperature tolerance of false codling moth Thaumatotibia leucotreta (Meyrick) (Lepidoptera: Tortricidae) in South Africa. Journal of Thermal Biology, 34, 320–325. 10.1016/j.jtherbio.2009.05.002 [DOI] [Google Scholar]
- Sulewski P, Kais K, Gołaś M, Rawa G, Urbańska K and Wąs A, 2021. Home bio‐waste composting for the circular economy. Energies, 14, 6164. 10.3390/en14196164 [DOI] [Google Scholar]
- Šumpich J, Liška J, Laštůvka A, Sitek J, Skyva J, Vávra J, Maršík L, Dvořák I, Žemlička M, Kabátek P, Laštůvka Z, Marek J, Marek S, Mikát M, Vacula D, Křivan V, Elsner G, Volf M, Jirgl T, Krejčík P, Hromádková V and Richter I, 2022. Faunistic records from The Czech Republic – 530: Lepidoptera. Klapalekiana, 58, 121–140. [Google Scholar]
- Šumpich J, Žemlička M, Liška J and Skyva J, 2021. On the lepidopteran fauna (Lepidoptera) of northern Bohemia – II. Sborník Severočeského Muzea, Přírodní Vědy, 39, 37–166. (in Czech). [Google Scholar]
- Svensson I, 2002. Anmärkningsvärda fynd av småfjärilar (Microlepidoptera) i Sverige 2001 (Remarkable records of Microlepidoptera in Sweden during 2001). Entomologisk Tidskrift, 123, 1–11. [Google Scholar]
- Tatàno F, Pagliaro G, Di Giovanni P, Floriani E and Mangani F, 2015. Biowaste home composting: experimental process monitoring and quality control. Waste Management, 38. 10.1016/j.wasman.2014.12.011 [DOI] [PubMed] [Google Scholar]
- Terblanche JS, de Jager Z, Boardman L and Addison P, 2014. Physiological traits suggest limited diapause response in false codling moth, Thaumatotibia leucotreta (Lepidoptera: Tortricidae). Journal of Applied Entomology, 138, 683–691. 10.1111/jen.12116 [DOI] [Google Scholar]
- Terblanche JS, Mitchell KA, Uys W, Short C and Boardman L, 2017. Thermal limits to survival and activity in two life stages of false codling moth Thaumatotibia leucotreta (Lepidoptera, Tortricidae). Physiological Entomology, 42, 379–388. 10.1111/phen.12210 [DOI] [Google Scholar]
- Uddin MM and Wright MM, 2022. Anaerobic digestion fundamentals, challenges, and technological advances. Physical Sciences Reviews. 10.1515/psr-2021-0068 [DOI] [Google Scholar]
- Uys W, 2014. Thermal limits of activity and survival in false codling moth, Thaumatotibia leucotreta (Lepidoptera: Tortricidae): Life‐stage and ramping rate effects [Thesis (MSc), Stellenbosch University]. Available online: https://scholar.sun.ac.za:443/handle/10019.1/95769
- Vansickle J, 1977. Attrition in distributed delay models. IEEE Transactions on Systems, Man, and Cybernetics, 7, 635–638. 10.1109/TSMC.1977.4309800 [DOI] [Google Scholar]
- VBN , 2017. Vereniging van Bloemenveilingen in Nederland t(he Dutch Flower Auctions Association). Available online: https://www.vbn.nl/site/wp-content/uploads/2017/04/Afzetsimulatie-snijbloemen-140122.pdf
- Venette RC, Davis EE, DaCosta M, Heisler H and Larson M. 2003. Mini Risk Assessment False codling moth, Thaumatotibia (=Cryptophlebia) leucotreta (Meyrick) [Lepidoptera: Tortricidae]. pp. 30. Available online: https://www.cdfa.ca.gov/plant/fcm/pdfs/publications/Venette_et_al_2003-FCM_PRA.pdf
- Verma S, 2002. Anaerobic digestion of biodegradable organics in municipal solid wastes. Columbia University, New York, NY, USA. Available online: https://gwcouncil.org/m-s-thesis-anaerobic-digestion-of-biodegradable-organics-in-municipal-solid-wastes/ [Google Scholar]
- Wysoki M, 1986. New records of Lepidopterous pests of macadamia in Israel. Phytoparasitica, 14, 147. [Google Scholar]
- Yakimova E, Kapchina‐Toteva V, Alexieva V, Sergiev I, Karanov E and Str B, 1996. Effect of chlorsulfuron (Glean‐75) and sucrose on some post‐harvest physiological events in cut flowers. Bulgarian Journal of Plant Physiology, 22, 74–87. [Google Scholar]
- Žitnik M and Vidic T, 2016. Food among waste. Available online: https://www.stat.si/statweb/File/DocSysFile/9206/FOOD_AMONG_WASTE_internet.pdf
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Compartment model for FCM development in EU
Cut roses pathway model for import from Africa