

Abstract
Animal models indicate that the endocannabinoid system (ECS) plays a modulatory role in stress and reward processing, both crucially impaired in addictive disorders. Preclinical findings showed endocannabinoid-modulated synaptic plasticity in reward brain networks linked to the metabotropic-glutamate-5 receptor (mGluR5), contributing to drug-reinforcing effects and drug-seeking behavior. Although animal models postulate a link between ECS and cocaine addiction, human translational studies are lacking. Here, we tested previous preclinical findings by investigating plasma endocannabinoids (eCBs) anandamide (AEA), 2-arachidonoylglycerol (2-AG), and the related N-acylethanolamines (NAEs) palmitoylethanolamide (PEA) and oleoylethanolamide (OEA), including their interaction with cerebral mGluR5, in chronic cocaine users (CU). We compared basal plasma concentrations between chronic CU (N = 103; 69 recreational CU and 34 dependent CU) and stimulant-naïve healthy controls (N = 92). Follow-up basal eCB/NAE plasma levels after 12 months were used for reliability and stability check (CU: N = 33; controls: N = 43). In an additional analysis using 11C-ABP688 positron emission tomography (PET) in a male subsample (CU: N = 18; controls: N = 16), we investigated the relationships between eCBs/NAEs and mGluR5 density in the brain. We found higher 2-AG plasma levels in dependent CU compared to controls and recreational CU. 2-AG levels were stable over time across all groups. In the PET-subsample, a positive association between 2-AG and mGluR5 brain density only in CU was found. Our results corroborate animal findings suggesting an alteration of the ECS in cocaine dependence and an association between peripheral 2-AG levels and cerebral mGluR5 in humans. Therefore, the ECS might be a promising pharmaco-therapeutic target for novel treatments of cocaine dependence.
Subject terms: Addiction, Molecular neuroscience
Introduction
Impaired reward behavior and inadequate stress response drive drug addiction development and maintenance. Accordingly, long-term maladaptive functional and structural synaptic plasticity caused by repeated drug exposure have been reported in brain networks related to reward behavior and stress response [1, 2]. Cocaine, as a non-selective monoamine transporter inhibitor, leads to elevated dopamine (DA) levels within the brain regions of the mesolimbic reward network in animals and humans (including projections from the ventral tegmental area [VTA] to the nucleus accumbens [NAc]), which has been linked to the highly addictive properties of cocaine [3, 4]. Animal models have shown that acute cocaine-induced activation of DA neurons in the NAc is specifically associated with the drug-reinforcing effects enhancing motivated and drug-seeking behavior [5–7]. Moreover, preclinical evidence suggests that repeated cocaine use entails drug-induced neuroplastic adaptations in mesolimbic DA neurons, causing drug-cue-related hyper-sensitivity and subsequently contributing to cocaine use maintenance and drug relapse even after a prolonged time of abstinence [8–10]. Human neuroimaging and pharmacological studies corroborate altered mesolimbic DA networks in chronic cocaine users (CU) contributing to the drug’s reinforcing and addictive effects [11–20].
In recent years, accumulating evidence mainly from preclinical studies indicates that the endocannabinoid system (ECS) plays a major role in reward processing. Specifically, long-term depression (LTD) causing synaptic plasticity in the mesolimbic DA network has been linked to the ECS underpinning drug-seeking and addictive behavior [21–24]. Animal models consistently showed endocannabinoid-mediated LTD in the VTA and NAc caused by the two main endocannabinoids (eCB), anandamide (AEA) and 2-arachidonoylglycerol (2-AG), activating the cannabinoid type-1 (CB1) receptor in the mesolimbic reward network. AEA and 2-AG are synthesized postsynaptically on-demand by the primary enzymes N-acylphosphatidylethanolamine specific phospholipase D and diacylglycerol lipase (DAGL), respectively. They act as retrograde neurotransmitters by activating presynaptic CB1 receptors entailing inhibition of presynaptic neurotransmitter release [25–29]. CB1 receptors are highly expressed throughout the mesolimbic DA network localized on terminals of glutamatergic and GABAergic neurons but not on DA neurons itself [30–32].
Stimulation of 2-AG-mediated LTD at GABAergic interneurons in the VTA resulting in disinhibition of DA neurons has been shown in animal models entailing increased DA release and projections to the NAc [33–35]. Preclinical findings showed that acute and repeated cocaine administration facilitates endocannabinoid-mediated LTD at GABAergic interneurons and abolishes endocannabinoid-mediated LTD at glutamatergic neurons, both resulting in increased DA release in the VTA and its projection to the NAc, contributing to the drug reinforcing effects [36–38]. Moreover, cocaine-induced activation of the endocannabinoid-LTD mechanism in the VTA has been recently linked to motivational behavior in rats, while the CB1 receptor inverse agonist rimonabant was able to block reward-seeking behavior [39]. In the NAc, a single in vivo cocaine administration in mice has been shown to abolish endocannabinoid-mediated LTD at terminals of glutamatergic neurons resulting in increased glutamate release and subsequently in increased excitation of DA cells [40].
Synaptic plasticity induced by endocannabinoid-mediated LTD has been further related to activation of postsynaptic metabotropic glutamate receptor 5 (mGluR5) in various brain regions in rodents [41–43]. Animal models showed that activating mGluR5 triggers synthesis and release of 2-AG through the DAGL pathway into the synaptic cleft [44]. Notably, a single administration of cocaine in mice has been shown to alter the functional interaction of endocannabinoid-mediated LTD and mGluR5 [40]. However, human findings using positron emission tomography (PET) imaging with the selective mGluR5 radio tracer 11C-ABP688 reported no differences in mGluR5 density between individuals with chronic CU and healthy controls [45], whereas other studies showed lower mGluR5 density in CU [46, 47].
Only one human study has investigated plasma endocannabinoids in individuals with cocaine use so far reporting elevated levels of AEA and the structurally related N-acylethanolamines (NAEs) palmitoylethanolamide (PEA), and oleoylethanolamide (OEA) but lower 2-AG levels in abstinent individuals with cocaine use disorder (CUD) compared to healthy controls. Of note, reduced 2-AG levels in CUD was likely primarily driven by co-morbid alcohol use disorder (AUD) [48]. Moreover, we recently found lower levels of OEA and PEA in hair samples of chronic CU compared to stimulant-naïve healthy controls, which might indicate tonic alterations in these lipid pathways [49]. However, basal eCB/NAE plasma levels of current, chronic CU and its interaction with mGluR5 have yet to be studied in humans.
Here, we analyzed basal eCB/NAE plasma levels in recreational CU, dependent CU, and healthy controls to identify specific lipid profiles related to cocaine use in humans. Additionally, we analyzed mGluR5 density of a subsample previously reported [45] to investigate potential cocaine-related associations between plasma eCB/NAE levels and mGluR5 density in an exploratory analysis. Based on reported animal model data and our recent findings [49], we expected alterations of eCB/NAE plasma levels primarily in dependent CU compared to controls, specifically for 2-AG, as well as an association between 2-AG and mGluR5 density in chronic CU.
Methods and materials
Participants
As part of the longitudinal Zurich Cocaine Cognition Study (ZuCo2St; [50, 51]), plasma eCB/NAE data of stimulant-naïve controls (n = 92) and chronic CU (n = 103; n = 69 recreational and n = 34 dependent users) were included. Additionally, we used eCB/NAE plasma data from the follow-up session after 12 months, including 43 stimulant-naïve healthy controls and 33 chronic CU from the first session to check for stability and reliability of eCB/NAE plasma levels over time. Chronic CU who participated in the follow-up assessment were further subdivided into individuals with increased (n = 16) and decreased (n = 17) cocaine use. Inclusion and exclusion criteria are described in the supplementary materials. Cocaine dependence was diagnosed following the DSM-IV criteria [52], with dependent CU fulfilling and recreational CU not meeting these criteria. Cocaine craving was assessed by the brief version of the Cocaine Craving Questionnaire (CCQ, [53]).
Eighteen male chronic CU and 17 cocaine-naïve controls from the ZuCo2St further underwent an additional PET procedure on a separate testing day to determine mGluR5 density (see below) with the same in- and exclusion criteria. Of this subsample, PET data of 18 male CU and 16 male controls were available with corresponding basal eCB/NAE plasma data.
The study has been carried out in accordance with the Declaration of Helsinki and was approved by the Cantonal Ethics Committee of Zurich (2022-00465, E-14/2009, 2009-0099/3). All participants provided written informed consent and were financially compensated for their participation.
Analysis of N-acylethanolamines and phyto-cannabinoids in plasma
Blood plasma samples were taken after the psychiatric interviews. Four NAEs, including the two eCBs, 2-AG and AEA, as well as OEA, and PEA and the plant-derived cannabinoids ∆9-tetrahydrocannabinol (THC) and cannabidiol (CBD) were extracted by liquid-liquid extraction and analyzed by isotope-dilution liquid chromatography/tandem mass spectrometry as previously described [54] using an Agilent 1200 high-performance liquid chromatography system coupled with an API 5000 triple quadruple mass spectrometer (AB Sciex). Because lipids such as endocannabinoids can readily cross the blood brain barrier, plasma endocannabinoid levels give reliable information about levels in the brain [55, 56].
PET imaging and analysis
PET imaging acquisition and preprocessing have been previously described in detail [45]. Briefly, data were acquired with the whole-body scanner Discovery STX PET/computed tomography scanner (GE Healthcare, Waukesha, USA) at the Department of Nuclear Medicine of the University Hospital Zurich. The mGluR5-selective radioligand 11C-ABP688 [57] was administered according to a bolus-infusion-protocol [58].
PET images were preprocessed using PMOD 3.307 software (PMOD Technologies, Zurich, Switzerland). Nine VOIs were generated based on the Montreal Neurological Institute brain templates: Prefrontal Cortex (PFC), hippocampal regions, anterior cingulate cortex (ACC), midcingulate cortex (MCC), amygdala, insula, caudate, putamen, and thalamus VOI, as well as the cerebellar reference VOI. Normalized values of distribution (Vnorm = CT[VOI]/CT[Cer]) of 11C-ABP688 uptake were calculated [59]. For more information see supplementary materials.
All procedures were kept consistent on the baseline, follow-up, and PET testing days, when blood samples were taken (see supplementary materials).
Statistical analysis
Statistical analyses were performed with IBM SPSS Statistics 28.0.1.1. and R version 4.2.1 [60] for Tukey HSD post-hoc analyses using multcomp R package. Pearson’s χ2-tests were carried out to analyze frequency data and quantitative between-group data were analyzed by analyses of variance (ANOVAs) with GROUP (controls, recreational, and dependent CU) as the fixed factor. For the main outcome variables of eCB/NAE plasma concentrations, we used analyses of covariance (ANCOVAs) with GROUP as the fixed factor to control for the confounding variables sex [61], age [62], and recent cannabis use, i.e., THC and CBD plasma levels as well as current alcohol dependence based on their previously reported influences on 2-AG [48, 63]. Repeated measure ANCOVA with sex and age as covariates and TIME (baseline and follow-up) as within-subject factor, and GROUP (controls, CU increase, and CU decrease) as between-subject factor as well as correlation analyses were used for a reliability check of eCB/NAE.
Potential associations between eCBs/NAEs and cocaine use variables (see Table 1) within chronic CU were investigated using stepwise linear regression analyses—forward and backward method (see supplementary materials). The influence of current alcohol dependence was specifically considered here given previously reported impact of co-morbid AUD on the association of 2-AG with CUD [48]. Moreover, we checked for potential associations between eCBs/NAEs and cannabis use variables (see Table 1) overall and within each group using stepwise linear regression analyses—forward and backward method (see supplementary materials).
Table 1.
Demographic data (means and standard deviations).
| Stimulant-naïve controls | Recreational cocaine users | Dependent cocaine users | Value | df1, df2 | p | |
|---|---|---|---|---|---|---|
| n = 92 | n = 69 | n = 34 | ||||
| Female/male | 26/66 | 18/51 | 9/25 | χ2 = 0.1 | 2 | 0.949 |
| Age | 30.55 (9.1) | 28.67 (7.6) | 33.97 (10.6)e | F = 4.1 | 2, 192 | 0.018 |
| Years of education | 10.80 (1.8) | 10.38 (1.7) | 9.49 (1.2)d,e | F = 7.9 | 2, 192 | <0.001 |
| Verbal IQ | 108.02 (12.1) | 103.13 (9.7)d | 100.91 (11.4)d | F = 6.6 | 2, 192 | 0.002 |
| BDI sum score | 4.16 (4.2) | 7.67 (6.6)d | 11.53 (8.2)d,e | F = 20.4 | 2, 192 | <0.001 |
| ADHD-SR | 7.51 (4.9) | 12.90 (9.1)d | 16.88 (8.4)d,e | F = 24.1 | 2, 192 | <0.001 |
| Smoking (y/n) | 66/26 | 57/12 | 27/7 | χ2 = 2.8 | 2 | 0.251 |
| Cigarettes/weeka | 84.14 (61.7) | 107.87 (55.9) | 135.04 (95.7)d | F = 5.8 | 2, 147 | 0.004 |
| Alcohol grams/week | 107.1 (115.6) | 170.70 (114.8) | 226.23 (322.0)d | F = 6.9 | 2, 192 | 0.001 |
| Alcohol dependence (y/n)c | - | 3/66 | 7/27 | χ2 = 6.9 | 1 | 0.009 |
| Cocaine | ||||||
| Cocaine craving | - | 19.14 (9.2) | 20.50 (11.4) | F = 0.4 | 1, 101 | 0.518 |
| Grams/week | - | 1.09 (1.4) | 5.47 (6.9) | F = 25.7 | 1, 101 | <0.001 |
| Cocaine abstinence in h | - | 643.40 (881.0) | 535.07 (814.6) | F = 0.4 | 1, 101 | 0.549 |
| Years of useb | - | 6 (0.75–17) | 9 (1–30) | F = 8.7 | 1, 101 | 0.004 |
| Hair concentration pg/mg | - | 3 070 (6 685.8) | 18 685.74 (21 478.2) | F = 30.7 | 1, 101 | <0.001 |
| Urine sample positive (y/n) | - | 9/59 | 13/21 | χ2 = 8.4 | 1 | 0.004 |
| Cannabis | ||||||
| Cannabis use (y/n) | 40/52 | 46/23d | 22/12d | χ2 = 10.0 | 2 | 0.007 |
| Cannabis dependence (y/n)c | 3/89 | 6/63 | 3/31 | χ2 = 2.52 | 2 | 0.283 |
| Grams per weeka | 1.21 (1.9) | 1.43 (2.4) | 2.49 (5.4) | F = 1.3 | 2, 105 | 0.286 |
| Years of usea,b | 7 (0–35) | 9 (0–26) | 12.5 (1.5–34)d,e | F = 6.2 | 2, 105 | 0.003 |
| Urine sample positive (y/n)a | 12/28 | 14/31 | 11/11 | χ2 = 2.9 | 2 | 0.232 |
| MDMA | ||||||
| MDMA use (y/n) | 1/91 | 22/47d | 10/24d | χ2 = 31.2 | 2 | <0.001 |
| Pills per weeka | - | 0.29 (0.4) | 1.22 (3.1) | F = 1.8 | 1, 28 | 0.186 |
| Years of usea,b | - | 4.5 (0–17) | 5 (0.5–18) | F = 1.1 | 1, 28 | 0.294 |
| Hair concentration pg/mga | - | 1 199.75 (2 336.5) | 673.00 (1 026) | F = 0.5 | 1, 28 | 0.504 |
Significant p-values are marked in bold.
ADHD attention-deficit/hyperactivity disorder, BDI Beck’s Depression Inventory, MDMA 3,4 methylenedioxymethamphetamine.
aWithin users.
bMedian (range).
cBased on DSM-IV criteria.
dTukey HSD post-hoc tests significant differences compared to controls.
eTukey HSD post-hoc tests significant differences compared to RCU.
For our secondary outcome, to determine associations between eCBs/NAEs and mGluR5 density, we used Spearman’s rank correlations within the chronic CU and control group separately and the Fisher’s r-to-z transformation to compare correlation coefficients, as well as binary logistic regression models (see supplementary materials).
False discovery rate (FDR) was applied to account for multiple comparisons when needed [64]. Cohen’s d effect size was calculated by the means and pooled standard deviations of the groups [65].
All plasma eCB/NAE levels were winsorized in order to account for outliers based on Wilcox [66] (see supplementary materials).
The statistical comparisons were carried out with a significance level of p < 0.05 (two-tailed).
Results
Participants’ demographic and substance use data are shown in Table 1.
Dependent CU shows elevated 2-AG plasma levels
ANCOVAs including sex, age, recent cannabis use (THC and CBD plasma levels), and current alcohol dependence as covariates and the dependent variables AEA, 2-AG, OEA, and PEA yielded a significant group effect only for 2-AG (F(2,187) = 4.3, p = 0.014, ηp2 = 0.04) showing highest basal 2-AG plasma levels in dependent CU (see Fig. 1a). Tukey HSD post-hoc comparisons identified significant elevated basal 2-AG plasma concentration in dependent CU compared to controls (p = 0.011, d = 0.61) and to recreational CU (p = 0.042, d = 0.52). In contrast, controls and recreational CU did not differ in 2-AG plasma levels (p = 0.825). No further group differences were found for AEA, PEA, and OEA (p’s > 0.165) as shown in Fig. 1b–d.
Fig. 1. Group differences in eCB/NAE plasma levels.

Dependent CU (DCU; dark blue) showed elevated basal plasma levels of a 2-arachidonylglycerol (2-AG) compared to controls (white) and recreational CU (RCU; light blue striped), whereas no group differences were found for b anandamide (AEA) c palmitoylethanolamide (PEA) d oleoylethanolamide (OEA). Bars represent means including individual data points, and error bars reflect standard error of the mean (SEM); corrected for age, sex, recent cannabis use, and alcohol dependence. Significant p-values marked with p < 0.05*, p < 0.01**.
2-AG plasma concentration was stable and reliable over time
Repeated measure ANCOVA with the dependent variable 2-AG controlling for age and sex showed no main effect of TIME (F(1,72) = 0.03, p = 0.870) as well as no TIME × GROUP interaction (F(2,72) = 0.051, p = 0.950). All three groups showed stable 2-AG levels over the twelve months period even if cocaine use was increased or decreased (see Supplementary materials Figure S2). Additionally, test–retest reliability was tested by correlation analyses with eCBs/NAEs plasma levels at baseline and follow-up (see supplementary materials Table S2). 2-AG plasma concentration at both time points was positively correlated over all participants and remained significant even after FDR-correction (pFDR = 0.016).
Elevated basal 2-AG levels are associated with higher cocaine craving in dependent CU, which is explained by current alcohol dependence
Stepwise linear regression analyses forward and backward within chronic CU for each eCB/NAE (AEA, 2-AG, OEA, and PEA) as the dependent variable and entering cocaine use variables (see Table 1) as regressors yielded exactly one significant association. Over all four regression models regarding each eCB/NAE, only CCQ sum score was identified as a significant regressor for basal 2-AG plasma level (F(1, 98) = 7.1, p = 0.009) even when controlling for age, sex, recent cannabis use (THC and CBD plasma levels), and alcohol dependence in the model (see supplementary materials Table S3). Separate analyses of recreational and dependent CU groups indicate that the negative association between 2-AG and CCQ sum score was driven by dependent CU (B = −0.41, t = −2.60, p = 0.015), explaining 38% of the variance of basal 2-AG plasma levels (F(5, 28) = 3.4, p = 0.015, R2 = 0.38), in contrast to recreational CU (B = −0.13, t = −1.05, p = 0.297). Association within recreational and dependent CU are shown in Supplementary Fig. S3a, and regression parameters of the models are shown in Supplementary Table S3.
To formally test whether current alcohol dependence and its interaction with 2-AG might influence the relationship between cocaine craving and 2-AG in dependent CU, we used a Linear Mixed Model (LMM) with CCQ sum score as the dependent variable and 2-AG, alcohol dependence, and the interaction 2-AG*alcohol dependence as factors. Results showed that the initial relationship between cocaine craving and 2-AG was primarily driven by alcohol dependence (F(1,34) = 11.36, p = 0.002) and its interaction with 2-AG (F(1,34) = 8.03, p = 0.008), whereas 2-AG alone was no longer a significant predictor for CCQ sum score in the model (F(1,34) = 2.62, p = 0.115). More precisely, results indicate that the association between cocaine craving and 2-AG was only present in dependent CU co-morbid with alcohol dependence (see supplementary materials Figure S3b). Additional LMM including cannabis dependence in the model was not significant (see supplementary materials Table S4).
Additional stepwise linear regression analyses forward and backward were performed to check for potential confounding effects of cannabis use on 2-AG over all subjects and within groups. Results yielded cannabis use (in the last 6 months) as a significant regressor for 2-AG only within dependent CU explaining approximately 15% of the variance (F(1, 32) = 6.7, p = 0.014). However, this regressor became non-significant after controlling for age, sex, recent cannabis use (THC and CBD plasma levels), and alcohol dependence in the model (see Supplementary materials Table S5). Moreover, adding cannabis use as an additional covariate in the group analysis for 2-AG did not change the results (F(2,186) = 4.7, p = 0.010, ηp2 = 0.05). Distributions of cannabis use in the last 6 months for each group are shown in the Supplementary materials Fig. S4.
Interaction between 2-AG and mGluR5 is specific to chronic cocaine use
Correlation analyses yielded significant positive correlations between 2-AG and region-specific mGluR5 VOIs within chronic CU showing significantly different correlation coefficients compared to controls for the brain structures insula, ACC, MCC, amygdala, thalamus, caudate, and hippocampal regions (see Table 2). Positive interactions within chronic CU are shown in Fig. 2a–g (scatterplots for the controls are shown in the Supplementary materials Fig. S5a-g). After controlling for multiple comparisons using FDR, only the correlation p-value of the thalamus region remained significant (see Table 2) while the other regions did not reach the significant threshold anymore even though showing clear statistical trends. To increase test power and to test for mGluR5 brain density overall, we performed an additional LMM including the fixed factors 2-AG and GROUP as well as its interaction 2-AG*GROUP and all nine cortical and subcortical mGluR5 VOIs for each subject as the dependent variable, with subjects and mGluR5 VOIs as random effect to control for subject and repeated level (VOIs within each subject) variability. LMM result showed a specific interaction effect of 2-AG*GROUP on mGluR5 density overall (F(1,34) = 6.33, p = 0.017), whereas 2-AG alone (F(1,34) = 1.29, p = 0.265) and GROUP (F(1,34) = 4.07, p = 0.052) did not reach the significant threshold. The 2-AG*GROUP interaction effect on mGluR5 remained significant even after including smoking and age into the model (F(1,34) = 4.41, p = 0.043) indicating a robust positive association between 2-AG and mGluR5 brain density overall only in the CU group (see Fig. 3). The model further yielded a significant main effect of smoking on mGluR5 density overall (F(1,34) = 5.97, p = 0.020) but not GROUP alone (F(1,34) = 2.63, p = 0.114) as we reported previously [45].
Table 2.
Spearman’s correlation coefficients for 2-AG plasma levels and mGluR5 density.
| mGluR5 density VOIs | r | p | Fisher’s z | |
|---|---|---|---|---|
| Cocaine users | Amygdala | 0.51 | 0.033 | 2.39 |
| Controls | −0.34 | 0.204 | p = 0.017 | |
| Cocaine users | Insula | 0.52 | 0.027 | 2.21 |
| Controls | −0.26 | 0.339 | p = 0.027 | |
| Cocaine users | ACC | 0.48 | 0.046 | 2.12 |
| Controls | −0.28 | 0.295 | p = 0.034 | |
| Cocaine users | MCC | 0.48 | 0.043 | 1.76 |
| Controls | −0.14 | 0.602 | p = 0.078 | |
| Cocaine users | Thalamus | 0.59 | 0.011a | 2.62 |
| Controls | −0.31 | 0.240 | p = 0.009a | |
| Cocaine users | Caudate | 0.56 | 0.016 | 2.56 |
| Controls | −0.33 | 0.217 | p = 0.011a | |
| Cocaine users | Putamen | 0.42 | 0.083 | 1.91 |
| Controls | −0.27 | 0.322 | p = 0.056 | |
| Cocaine users | Hippocampal region | 0.47 | 0.048 | 2.16 |
| Controls | −0.30 | 0.264 | p = 0.031 | |
| Cocaine users | PFC | 0.44 | 0.067 | 2.01 |
| Controls | −0.28 | 0.295 | p = 0.044 |
Significant p-values < 0.05 are shown in bold (uncorrected).
aSignificant after FDR-correction.
Fig. 2. Positive correlations between 2-AG and mGluR5 VOIs within chronic CU.

Within chronic cocaine users (n = 18), basal plasma levels of 2-AG were positively associated with mGluR5 density of the (a) amygdala, (b) thalamus, (c) hippocampal regions, (d) anterior cingulate cortex (ACC), (e) midcingulate cortex (MCC), (f) insula, and (g) caudate. After controlling for tobacco use and age, the interaction between 2-AG and mGluR5 was specifically evident in chronic cocaine users for the brain structures amygdala (a), thalamus (b), as well as hippocampal region (c), and as trend levels for the other brain structures (d–g).
Fig. 3. Interaction between 2-AG and mGluR5 density in chronic CU.
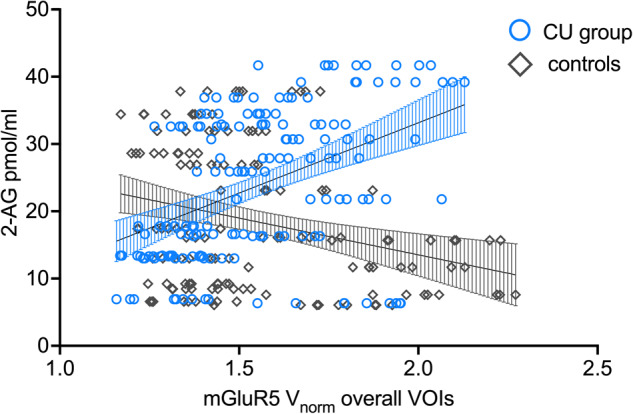
Linear mixed models yielded a specific interaction effect of 2-AG*GROUP on mGluR5 density overall indicating a positive association between 2-AG and mGluR5 density overall VOIs only in chronic cocaine users (CU; blue circles) but not in controls (gray diamonds).
To formally address whether the relationship between 2-AG and mGluR5 density was unique to chronic CU even after controlling for confounding variables age and smoking, we used binary logistic regression models with GROUP as the dependent variable. Robust interaction effects for chronic CU between 2-AG and mGluR5 density were found for the brain structures: thalamus (B = 1.09, Wald=4.61, p = 0.032, OR = 2.96, 95%CI [1.10; 7.98]), amygdala (B = 0.62, Wald=4.34, p = 0.037, OR = 1.85, 95%CI [1.04; 3.30]), and hippocampal regions (B = 0.77, Wald=4.39, p = 0.036, OR = 2.15, 95%CI [1.05; 4.40]). Insula, caudate, ACC, and MCC showed differences at trend levels but did not meet the significant threshold (see supplementary materials Table S6d–g). All logistic regression models are shown in the supplementary materials Table S6.
Discussion
Our findings indicate alterations of the ECS in individuals with cocaine dependence. Specifically, basal 2-AG was elevated in cocaine dependence compared to controls and at a trend level also to recreational users. In contrast, the latter two groups did not differ in basal 2-AG plasma concentration. Additional exploratory analyses of a male subgroup with mGluR5-specific PET yielded a positive association between basal 2-AG plasma levels and cerebral mGluR5 density only evident in chronic CU, corroborating animal findings and suggesting a specific 2-AG/mGluR5 interaction underpinning cocaine dependence also in humans.
Our findings suggest that 2-AG might play a crucial role in cocaine dependence in humans. This is consistent with recent animal models showing elevated 2-AG levels in the VTA after cocaine administration, contributing to the positive reinforcing and motivation-enhancing effects of the substance [36, 39, 67]. Although human plasma samples can only reflect peripheral eCB/NAE levels and do not allow direct assessment of brain-specific concentrations, lipids such as endocannabinoids are known to readily cross the blood-brain barrier and therefore plasma endocannabinoid levels are assumed to give reliable information about levels in the brain [55, 56]. Whereas a previous study did not find correlations between AEA levels in cerebrospinal fluid (CSF) and serum [68], recent human findings indicate that peripheral endocannabinoid plasma concentrations might reflect brain levels. Translational evidence from humans and mice indicate a similar relationship between fear behavior and AEA concentrations, genetically manipulated by the FAAH polymorphism, in both, the periphery as well as in the amygdala and prefrontal cortex [69]. Moreover, peripheral levels of AEA, OEA, and PEA have been recently reported to show a negative correlation with FAAH levels in the human brain using the FAAH tracer [C-11]CURB in patients with alcohol use disorder [70] indicating that peripheral plasma endocannabinoids might reflect central endocannabinoid dynamics. Therefore, we speculate that the elevated 2-AG levels found in individuals with cocaine dependence indicate higher response to the cocaine-rewarding effects as well as higher motivated behavior resulting in increased vulnerability to developing cocaine dependence. Given that i) cocaine use intensity (i.e., cocaine grams per week, cocaine years of use, cocaine abstinence, cocaine hair concentrations, and positive urine sample) did not significantly explain the 2-AG variance, ii) 2-AG levels were stable over time, and iii) 2-AG levels did not covary with changing cocaine use between baseline and follow-up, elevated basal 2-AG plasma levels might reflect a risk factor for cocaine dependence (e.g., reward behavior) rather than a dose-dependent consequence of chronic cocaine use. Accordingly, no correlations between 2-AG and cocaine addiction severity variables have been previously shown in abstinent cocaine users [48].
In contrast to our findings, the same study found reduced 2-AG plasma levels and elevated NAE levels in abstinent individuals with CUD compared to healthy controls. However, lower 2-AG plasma levels were only significantly different from controls in abstinent cocaine users with co-morbid AUD as the authors reported in the supplements, whereas AUD had no influence on NAE levels. Moreover, a high number of psychiatric co-morbidities such as mood (40%), anxiety (25%) and substance use disorder other than cocaine (86%) as well as personality disorders (33%) were reported for the abstinent CUD group, which had effects on eCB/NAE plasma levels and might further explain the different findings of 2-AG. Importantly, ROC curves showed that CUD was only a significant predictor for NAEs but not 2-AG indicating that the findings regarding lower 2-AG plasma levels in abstinent cocaine users might not be related to chronic cocaine use per se. Since we controlled for a variety of confounding variables, which have been reported to influence eCB/NAE, we believe that our analyses detected robust and specific effects of dependent cocaine use. However, the question of whether elevated 2-AG levels in individuals with cocaine dependence is due to long-term changes in synaptic plasticity caused by repeated cocaine use, as previously shown in animal models, or whether it is a pre-existing phenomenon, and 2-AG is thus a biomarker of vulnerability to cocaine dependence, requires further investigation. Our present analyses with follow-up data suggest stable 2-AG plasma levels over a period of at least 12 months, independent of increased or decreased cocaine use, which might be indicative of 2-AG being a biomarker (i.e., trait marker) or alternatively suggesting that cocaine-induced changes in 2-AG might be of limited reversibility. Future studies should, therefore, address our assumption that individuals with elevated basal 2-AG levels are more prone to the rewarding effects of cocaine (e.g., wanting and liking) and subsequently more vulnerable to developing cocaine dependence suggesting elevated 2-AG as a potential biomarker of cocaine use disorder.
Although we found higher basal 2-AG plasma levels in the cocaine-dependent group, cocaine craving was negatively associated with 2-AG plasma levels in dependent CU initially. This result would have been in contrast to previous animal findings reporting a positive association between stress-induced cocaine craving and phasic 2-AG response [71, 72]. However, the additional LMM showed that the relationship between 2-AG and cocaine craving was primarily driven by alcohol dependence and only found in individuals with co-morbid cocaine and alcohol dependence but not with cocaine dependence alone. Genetic and pharmacological downregulation of 2-AG signaling in rodent brain regions responsible for stress and affect regulation (e.g., amygdala, paraventricular hypothalamus, and prefrontal regions) has been linked to anxiety- and depressive-like effects typically observed during drug withdrawal [73, 74]. Therefore, one might speculate that our findings of lower 2-AG levels associated with higher cocaine craving in individuals with co-morbid cocaine and alcohol dependence may be a phasic response to the negative affective state of cocaine and/or alcohol withdrawal. However, no reliable measures of withdrawal and anxiety symptoms neither for cocaine nor for alcohol were available for the present dataset and future human studies should investigate the influence of alcohol and cocaine withdrawal on 2-AG and craving in AUD alone and co-morbid with CUD. In fact, since the relationship between 2-AG and cocaine craving was no longer evident in dependent CU without co-morbid alcohol dependence, the present results support our suggestion of elevated 2-AG levels as a stable biomarker unique for cocaine dependence.
Here, we found for the first time a relationship between 2-AG and mGluR5 density in humans, including brain regions involved in processing reward behavior and affect regulation (i.e., amygdala, thalamus, and hippocampal regions). This association was evident only in chronic CU but not in healthy controls and was strongest in the thalamus, where the correlation survived FDR-correction despite the small sample size. Moreover, the LMM including all nine mGluR5 density VOIs yielded a robust effect of 2-AG on mGluR5 density overall specific for chronic CU. Our present results are in line with previous animal models showing an involvement of mGluR5 and 2-AG in LTD throughout various brain regions [41–43] and are consistent with the notion of drug-induced long-term synaptic plasticity [1]. Notably, single in vivo cocaine administration has been previously reported to affect mGluR5 and endocannabinoid-LTD in mice [40], which might in the long run lead to alterations in this system mechanism and may further explain that our finding of the statistical interaction between 2-AG and mGluR5 was only evident in chronic CU. Recent animal models reported an association between the mGluR5-mediated 2-AG-LTD in the NAc and reward-seeking behavior, showing that drug-conditioned cues can amplify 2-AG-mediated LTD by excitatory afferents from the PFC impinging on D1-medium spiny neurons (D1-MSN) in the NAc and stimulating glutamate release [21, 75]. Subsequent activation of postsynaptic mGluR5-mediated 2-AG-LTD amplified LTD at glutamatergic neurons resulting in increased drug-seeking behavior, including craving. These findings are consistent with both previous preclinical results and recent human findings reporting impaired glutamate homeostasis in the NAc in chronic CU [76, 77]. Although we were not able to assess mGluR5 density in the ventral striatum (i.e., VTA and NAc) due to limited spatial resolution of PET imaging, and eCBs/NAEs can only be assessed in the periphery and not brain-structure-specific in humans, we nevertheless found a significant interaction between basal 2-AG and mGluR5 density in a variety of brain regions involved in reward and affect regulation, which was evident only in chronic CU. These findings indeed indicate that there might be a specific 2-AG/mGluR5-mechanism underpinning cocaine dependence in humans, which may contribute to increased craving behavior and vulnerability to cocaine relapse. Our results extend previous findings from our lab [45], indicating that cocaine use disorder might be rather linked to 2-AG and its interaction with mGluR5 than exclusively to mGluR5 density per se, which has been reported previously [46, 47]. Of note, our PET subsamples consist of male controls and CU and need to be tested in women as well. Future studies should address this issue and our assumption by using PET imaging techniques with selective tracers for enzymes and lipids related to 2-AG in both sexes. This will allow for detecting alterations of the ECS, specifically in the NAc, during craving.
We recently reported lower tonic OEA and PEA hair concentration in a subgroup of the present chronic CU compared to healthy controls, specifically pronounced in individuals showing cocaine dependence [49]. However, we did not find differences between groups in AEA and 2-AG hair concentrations. This might be caused by the fact that concentrations of OEA and PEA are generally higher than AEA and 2-AG in hair and other biological samples [55]. In addition, both OEA and PEA, although structurally related to AEA, serve different functions in the body. While OEA is a satiety compound and the main endogenous ligand to PPAR-α and thus deeply involved in the regulation of the energy metabolism [78], PEA is also considered an agonist at PPAR-α but also binds to the cannabinoid-like G-protein coupled receptors GPR55 and GPR119 and is suggested to be related to chronic inflammation and pain [79, 80]. Moreover, reported findings might also be related to phasic vs. tonic eCB/NAE release in the brain, which may be further supported by our null results of the correlation analyses between plasma and hair levels of eCBs/NAEs after FDR-correction (see supplementary materials Table S6). No association between hair and plasma eCB/NAE levels have been also recently reported [81].
The present findings are novel in the field of cocaine use disorder in humans and crucially contribute to a better mechanistic understanding of the role of the ECS in cocaine dependence. Specifically, 2-AG seems to be a critical player and might act as a biomarker for the vulnerability to develop cocaine use disorder and subsequently might be a promising pharmaco-therapeutic target for novel treatments of aiming to prevent development of cocaine use disorder, drug relapse, and improve cocaine abstinence.
Supplementary information
Acknowledgements
We thank Avery Nelson for his preliminary statistical analyses with parts of the present data set.
Author contributions
SLK and BBQ contributed to the final research question and hypotheses. SLK analyzed the data and wrote the paper. BBQ designed initial research and revised the paper. LMH, MV, and KHP were involved in data collection and AKK helped with the dataset. VT and SMA were involved in the PET study and supported initial PET data analysis. MRB performed the toxicological hair analysis. CB, FP, CR, and FML conducted the eCB/NAE plasma analyses.
Funding
This work was supported by the Brain & Behavior Research Foundation (Grant No. 30549; NARSAD Young Investigator Grant to SLK), the Uniscientia Foundation (Grant No. 195-2022 to SLK and BBQ), and the Swiss National Science Foundation (Grant No. PP00P1-123516/1 and PP00P1-146326 to BBQ). SLK was partially financed by the Swiss Foundation for Alcohol Use (Grant No. 322) to SLK and BBQ and later financed by the Swiss National Science Foundation (Grant No. TMPFP1_209804) to SLK.
Competing interests
FML is a shareholder of curantis UG (Ltd.). CR is a shareholder of lero bioscience UG (Ltd). BBQ received funding from Gilead Sciences. All other authors declare no conflict of interest.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
The online version contains supplementary material available at 10.1038/s41398-023-02628-7.
References
- 1.Luscher C, Malenka RC. Drug-evoked synaptic plasticity in addiction: from molecular changes to circuit remodeling. Neuron. 2011;69:650–63. doi: 10.1016/j.neuron.2011.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Wemm SE, Sinha R. Drug-induced stress responses and addiction risk and relapse. Neurobiol Stress. 2019;10:100148. doi: 10.1016/j.ynstr.2019.100148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ritz MC, Lamb RJ, Goldberg SR, Kuhar MJ. Cocaine receptors on dopamine transporters are related to self-administration of cocaine. Science. 1987;237:1219–23. doi: 10.1126/science.2820058. [DOI] [PubMed] [Google Scholar]
- 4.Hernandez L, Hoebel BG. Food reward and cocaine increase extracellular dopamine in the nucleus accumbens as measured by microdialysis. Life Sci. 1988;42:1705–12. doi: 10.1016/0024-3205(88)90036-7. [DOI] [PubMed] [Google Scholar]
- 5.Pettit HO, Justice JB., Jr Dopamine in the nucleus accumbens during cocaine self-administration as studied by in vivo microdialysis. Pharm Biochem Behav. 1989;34:899–904. doi: 10.1016/0091-3057(89)90291-8. [DOI] [PubMed] [Google Scholar]
- 6.Wise RA, Newton P, Leeb K, Burnette B, Pocock D, Justice JB., Jr Fluctuations in nucleus accumbens dopamine concentration during intravenous cocaine self-administration in rats. Psychopharmacology. 1995;120:10–20. doi: 10.1007/BF02246140. [DOI] [PubMed] [Google Scholar]
- 7.Gratton A, Wise RA. Drug- and behavior-associated changes in dopamine-related electrochemical signals during intravenous cocaine self-administration in rats. J Neurosci. 1994;14:4130–46. doi: 10.1523/JNEUROSCI.14-07-04130.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Thomas MJ, Kalivas PW, Shaham Y. Neuroplasticity in the mesolimbic dopamine system and cocaine addiction. Br J Pharm. 2008;154:327–42. doi: 10.1038/bjp.2008.77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Baumgartner U, Buchholz HG, Bellosevich A, Magerl W, Siessmeier T, Rolke R, et al. High opiate receptor binding potential in the human lateral pain system. Neuroimage. 2006;30:692–9. doi: 10.1016/j.neuroimage.2005.10.033. [DOI] [PubMed] [Google Scholar]
- 10.Kalivas PW, O’Brien C. Drug addiction as a pathology of staged neuroplasticity. Neuropsychopharmacology. 2008;33:166–80. doi: 10.1038/sj.npp.1301564. [DOI] [PubMed] [Google Scholar]
- 11.Gu H, Salmeron BJ, Ross TJ, Geng X, Zhan W, Stein EA, et al. Mesocorticolimbic circuits are impaired in chronic cocaine users as demonstrated by resting-state functional connectivity. Neuroimage. 2010;53:593–601. doi: 10.1016/j.neuroimage.2010.06.066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Breiter HC, Gollub RL, Weisskoff RM, Kennedy DN, Makris N, Berke JD, et al. Acute effects of cocaine on human brain activity and emotion. Neuron. 1997;19:591–611. doi: 10.1016/s0896-6273(00)80374-8. [DOI] [PubMed] [Google Scholar]
- 13.Konova AB, Moeller SJ, Tomasi D, Volkow ND, Goldstein RZ. Effects of methylphenidate on resting-state functional connectivity of the mesocorticolimbic dopamine pathways in cocaine addiction. JAMA Psychiatry. 2013;70:857–68. doi: 10.1001/jamapsychiatry.2013.1129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Volkow ND, Wang GJ, Fischman MW, Foltin R, Fowler JS, Franceschi D, et al. Effects of route of administration on cocaine induced dopamine transporter blockade in the human brain. Life Sci. 2000;67:1507–15. doi: 10.1016/s0024-3205(00)00731-1. [DOI] [PubMed] [Google Scholar]
- 15.Volkow ND, Wang GJ, Fowler JS, Logan J, Gatley SJ, Hitzemann R, et al. Decreased striatal dopaminergic responsiveness in detoxified cocaine-dependent subjects. Nature. 1997;386:830–3. doi: 10.1038/386830a0. [DOI] [PubMed] [Google Scholar]
- 16.Volkow ND, Fowler JS, Wolf AP, Schlyer D, Shiue CY, Alpert R, et al. Effects of chronic cocaine abuse on postsynaptic dopamine receptors. Am J Psychiatry. 1990;147:719–24. doi: 10.1176/ajp.147.6.719. [DOI] [PubMed] [Google Scholar]
- 17.Volkow ND, Fowler JS, Wang GJ, Hitzemann R, Logan J, Schlyer DJ, et al. Decreased dopamine D2 receptor availability is associated with reduced frontal metabolism in cocaine abusers. Synapse. 1993;14:169–77. doi: 10.1002/syn.890140210. [DOI] [PubMed] [Google Scholar]
- 18.Volkow N, Li TK. The neuroscience of addiction. Nat Neurosci. 2005;8:1429–30. doi: 10.1038/nn1105-1429. [DOI] [PubMed] [Google Scholar]
- 19.Martinez D, Narendran R, Foltin RW, Slifstein M, Hwang DR, Broft A, et al. Amphetamine-induced dopamine release: markedly blunted in cocaine dependence and predictive of the choice to self-administer cocaine. Am J Psychiatry. 2007;164:622–9. doi: 10.1176/ajp.2007.164.4.622. [DOI] [PubMed] [Google Scholar]
- 20.Martinez D, Carpenter KM, Liu F, Slifstein M, Broft A, Friedman AC, et al. Imaging dopamine transmission in cocaine dependence: link between neurochemistry and response to treatment. Am J Psychiatry. 2011;168:634–41. doi: 10.1176/appi.ajp.2010.10050748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Spanagel R. Cannabinoids and the endocannabinoid system in reward processing and addiction: from mechanisms to interventions. Dialogues Clin Neurosci. 2020;22:241–50. doi: 10.31887/DCNS.2020.22.3/rspanagel. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Parsons LH, Hurd YL. Endocannabinoid signalling in reward and addiction. Nat Rev Neurosci. 2015;16:579–94. doi: 10.1038/nrn4004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Moreira FA, Jupp B, Belin D, Dalley JW. Endocannabinoids and striatal function: implications for addiction-related behaviours. Behav Pharm. 2015;26:59–72. doi: 10.1097/FBP.0000000000000109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Everett TJ, Gomez DM, Hamilton LR, Oleson EB. Endocannabinoid modulation of dopamine release during reward seeking, interval timing, and avoidance. Prog Neuropsychopharmacol Biol Psychiatry. 2021;104:110031. doi: 10.1016/j.pnpbp.2020.110031. [DOI] [PubMed] [Google Scholar]
- 25.Blankman JL, Cravatt BF. Chemical probes of endocannabinoid metabolism. Pharm Rev. 2013;65:849–71. doi: 10.1124/pr.112.006387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Blankman JL, Simon GM, Cravatt BF. A comprehensive profile of brain enzymes that hydrolyze the endocannabinoid 2-arachidonoylglycerol. Chem Biol. 2007;14:1347–56. doi: 10.1016/j.chembiol.2007.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Deutsch DG, Chin SA. Enzymatic synthesis and degradation of anandamide, a cannabinoid receptor agonist. Biochem Pharm. 1993;46:791–6. doi: 10.1016/0006-2952(93)90486-g. [DOI] [PubMed] [Google Scholar]
- 28.Devane WA, Hanus L, Breuer A, Pertwee RG, Stevenson LA, Griffin G, et al. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science. 1992;258:1946–9. doi: 10.1126/science.1470919. [DOI] [PubMed] [Google Scholar]
- 29.Sugiura T, Kondo S, Sukagawa A, Nakane S, Shinoda A, Itoh K, et al. 2-Arachidonoylglycerol: a possible endogenous cannabinoid receptor ligand in brain. Biochem Biophys Res Commun. 1995;215:89–97. doi: 10.1006/bbrc.1995.2437. [DOI] [PubMed] [Google Scholar]
- 30.Herkenham M, Lynn AB, Little MD, Johnson MR, Melvin LS, de Costa BR, et al. Cannabinoid receptor localization in brain. Proc Natl Acad Sci USA. 1990;87:1932–6. doi: 10.1073/pnas.87.5.1932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mailleux P, Vanderhaeghen JJ. Distribution of neuronal cannabinoid receptor in the adult rat brain: a comparative receptor binding radioautography and in situ hybridization histochemistry. Neuroscience. 1992;48:655–68. doi: 10.1016/0306-4522(92)90409-u. [DOI] [PubMed] [Google Scholar]
- 32.Szabo B, Schlicker E. Effects of cannabinoids on neurotransmission. Handb Exp Pharm. 2005;168:327–65. doi: 10.1007/3-540-26573-2_11. [DOI] [PubMed] [Google Scholar]
- 33.Riegel AC, Lupica CR. Independent presynaptic and postsynaptic mechanisms regulate endocannabinoid signaling at multiple synapses in the ventral tegmental area. J Neurosci. 2004;24:11070–8. doi: 10.1523/JNEUROSCI.3695-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Szabo B, Muller T, Koch H. Effects of cannabinoids on dopamine release in the corpus striatum and the nucleus accumbens in vitro. J Neurochem. 1999;73:1084–9. doi: 10.1046/j.1471-4159.1999.0731084.x. [DOI] [PubMed] [Google Scholar]
- 35.Melis M, Pistis M, Perra S, Muntoni AL, Pillolla G, Gessa GL. Endocannabinoids mediate presynaptic inhibition of glutamatergic transmission in rat ventral tegmental area dopamine neurons through activation of CB1 receptors. J Neurosci. 2004;24:53–62. doi: 10.1523/JNEUROSCI.4503-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wang H, Treadway T, Covey DP, Cheer JF, Lupica CR. Cocaine-induced endocannabinoid mobilization in the ventral tegmental area. Cell Rep. 2015;12:1997–2008. doi: 10.1016/j.celrep.2015.08.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Pan B, Hillard CJ, Liu QS. Endocannabinoid signaling mediates cocaine-induced inhibitory synaptic plasticity in midbrain dopamine neurons. J Neurosci. 2008;28:1385–97. doi: 10.1523/JNEUROSCI.4033-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Wang R, Hausknecht KA, Gancarz-Kausch AM, Oubraim S, Shen RY, Haj-Dahmane S. Cocaine self-administration abolishes endocannabinoid-mediated long-term depression of glutamatergic synapses in the ventral tegmental area. Eur J Neurosci. 2020;52:4517–24. doi: 10.1111/ejn.14980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Engi SA, Beebe EJ, Ayvazian VM, Cruz FC, Cheer JF, Wenzel JM, et al. Cocaine-induced increases in motivation require 2-arachidonoylglycerol mobilization and CB1 receptor activation in the ventral tegmental area. Neuropharmacology. 2021;193:108625. doi: 10.1016/j.neuropharm.2021.108625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Fourgeaud L, Mato S, Bouchet D, Hemar A, Worley PF, Manzoni OJ. A single in vivo exposure to cocaine abolishes endocannabinoid-mediated long-term depression in the nucleus accumbens. J Neurosci. 2004;24:6939–45. doi: 10.1523/JNEUROSCI.0671-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Robbe D, Kopf M, Remaury A, Bockaert J, Manzoni OJ. Endogenous cannabinoids mediate long-term synaptic depression in the nucleus accumbens. Proc Natl Acad Sci USA. 2002;99:8384–8. doi: 10.1073/pnas.122149199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Puente N, Cui Y, Lassalle O, Lafourcade M, Georges F, Venance L, et al. Polymodal activation of the endocannabinoid system in the extended amygdala. Nat Neurosci. 2011;14:1542–7. doi: 10.1038/nn.2974. [DOI] [PubMed] [Google Scholar]
- 43.Nufer TM, Merrill C, Friend L, Hopkins Z, Boyce Z, Edwards JG. Expression of type I mGluRs predicts plasticity in the hippocampal stratum radiatum interneurons. Neurosci Lett. 2019;712:134472. doi: 10.1016/j.neulet.2019.134472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Piomelli D. More surprises lying ahead. The endocannabinoids keep us guessing. Neuropharmacology. 2014;76:228–34. doi: 10.1016/j.neuropharm.2013.07.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Hulka LM, Treyer V, Scheidegger M, Preller KH, Vonmoos M, Baumgartner MR, et al. Smoking but not cocaine use is associated with lower cerebral metabotropic glutamate receptor 5 density in humans. Mol Psychiatry. 2014;19:625–32. doi: 10.1038/mp.2013.51. [DOI] [PubMed] [Google Scholar]
- 46.Milella MS, Marengo L, Larcher K, Fotros A, Dagher A, Rosa-Neto P, et al. Limbic system mGluR5 availability in cocaine dependent subjects: a high-resolution PET [(11)C]ABP688 study. Neuroimage. 2014;98:195–202. doi: 10.1016/j.neuroimage.2014.04.061. [DOI] [PubMed] [Google Scholar]
- 47.Martinez D, Slifstein M, Nabulsi N, Grassetti A, Urban NB, Perez A, et al. Imaging glutamate homeostasis in cocaine addiction with the metabotropic glutamate receptor 5 positron emission tomography radiotracer [(11)C]ABP688 and magnetic resonance spectroscopy. Biol Psychiatry. 2014;75:165–71. doi: 10.1016/j.biopsych.2013.06.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Pavon FJ, Araos P, Pastor A, Calado M, Pedraz M, Campos-Cloute R, et al. Evaluation of plasma-free endocannabinoids and their congeners in abstinent cocaine addicts seeking outpatient treatment: impact of psychiatric co-morbidity. Addict Biol. 2013;18:955–69. doi: 10.1111/adb.12107. [DOI] [PubMed] [Google Scholar]
- 49.Voegel CD, Kroll SL, Schmid MW, Kexel AK, Baumgartner MR, Kraemer T, et al. Alterations of stress-related glucocorticoids and endocannabinoids in hair of chronic cocaine users. Int J Neuropsychopharmacol. 2022;25:226–37. doi: 10.1093/ijnp/pyab070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Schote AB, Jager K, Kroll SL, Vonmoos M, Hulka LM, Preller KH, et al. Glucocorticoid receptor gene variants and lower expression of NR3C1 are associated with cocaine use. Addict Biol. 2019;24:730–42. doi: 10.1111/adb.12632. [DOI] [PubMed] [Google Scholar]
- 51.Vonmoos M, Hulka LM, Preller KH, Jenni D, Baumgartner MR, Stohler R, et al. Cognitive dysfunctions in recreational and dependent cocaine users: role of attention-deficit hyperactivity disorder, craving and early age at onset. Br J Psychiatry. 2013;203:35–43. doi: 10.1192/bjp.bp.112.118091. [DOI] [PubMed] [Google Scholar]
- 52.American Psychiatric Association. Diagnostic and statistical manual of mental disorders: DSM-IV. 4th ed. American Psychiatric Association (APA): Washington, DC; 1994.
- 53.Sussner BD, Smelson DA, Rodrigues S, Kline A, Losonczy M, Ziedonis D. The validity and reliability of a brief measure of cocaine craving. Drug Alcohol Depend. 2006;83:233–7. doi: 10.1016/j.drugalcdep.2005.11.022. [DOI] [PubMed] [Google Scholar]
- 54.Couttas TA, Boost C, Pahlisch F, Sykorova EB, Leweke JE, Koethe D, et al. Simultaneous assessment of serum levels and pharmacologic effects of cannabinoids on endocannabinoids and n-acylethanolamines by liquid chromatography-tandem mass spectrometry. Cannabis Cannabinoid Res. 2023;8:657–69. doi: 10.1089/can.2021.0181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hillard CJ. Circulating endocannabinoids: from whence do they come and where are they going? Neuropsychopharmacology. 2018;43:155–72. doi: 10.1038/npp.2017.130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Banks WA, Kastin AJ, Rapoport SI. Permeability of the blood-brain barrier to circulating free fatty acids. In: S. Y, D.I. M, editors. Handbook of essential fatty acid biology. Totowa, NJ: Humana Press; 1997.
- 57.Ametamey SM, Treyer V, Streffer J, Wyss MT, Schmidt M, Blagoev M, et al. Human PET studies of metabotropic glutamate receptor subtype 5 with 11C-ABP688. J Nucl Med. 2007;48:247–52. [PubMed] [Google Scholar]
- 58.Burger C, Deschwanden A, Ametamey S, Johayem A, Mancosu B, Wyss M, et al. Evaluation of a bolus/infusion protocol for 11C-ABP688, a PET tracer for mGluR5. Nucl Med Biol. 2010;37:845–51. doi: 10.1016/j.nucmedbio.2010.04.107. [DOI] [PubMed] [Google Scholar]
- 59.Innis RB, Cunningham VJ, Delforge J, Fujita M, Gjedde A, Gunn RN, et al. Consensus nomenclature for in vivo imaging of reversibly binding radioligands. J Cereb Blood Flow Metab. 2007;27:1533–9. doi: 10.1038/sj.jcbfm.9600493. [DOI] [PubMed] [Google Scholar]
- 60.R Core Team. R: a language and environment for statistical computing. R Foundation for Statistical Computing: Vienna, Austria; 2018.
- 61.Vecchiarelli HA, Morena M, Lee TTY, Nastase AS, Aukema RJ, Leitl KD, et al. Sex and stressor modality influence acute stress-induced dynamic changes in corticolimbic endocannabinoid levels in adult Sprague Dawley rats. Neurobiol Stress. 2022;20:100470. doi: 10.1016/j.ynstr.2022.100470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Piyanova A, Lomazzo E, Bindila L, Lerner R, Albayram O, Ruhl T, et al. Age-related changes in the endocannabinoid system in the mouse hippocampus. Mech Ageing Dev. 2015;150:55–64. doi: 10.1016/j.mad.2015.08.005. [DOI] [PubMed] [Google Scholar]
- 63.Bequet F, Uzabiaga F, Desbazeille M, Ludwiczak P, Maftouh M, Picard C, et al. CB1 receptor-mediated control of the release of endocannabinoids (as assessed by microdialysis coupled with LC/MS) in the rat hypothalamus. Eur J Neurosci. 2007;26:3458–64. doi: 10.1111/j.1460-9568.2007.05900.x. [DOI] [PubMed] [Google Scholar]
- 64.Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc. 1995;57:289–300. [Google Scholar]
- 65.Cohen J. Statistical power analysis for the bevavioral sciences. 2 ed. Lawrence Erlbaum Associates: Hillsday, New Jersey; 1988.
- 66.Wilcox R. Modern statistics for the social and behavioral sciences: a practical introduction. CRC Press: Hoboken, NJ; 2011.
- 67.Nakamura Y, Dryanovski DI, Kimura Y, Jackson SN, Woods AS, Yasui Y, et al. Cocaine-induced endocannabinoid signaling mediated by sigma-1 receptors and extracellular vesicle secretion. Elife. 2019;8:e47209.. doi: 10.7554/eLife.47209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Koethe D, Giuffrida A, Schreiber D, Hellmich M, Schultze-Lutter F, Ruhrmann S, et al. Anandamide elevation in cerebrospinal fluid in initial prodromal states of psychosis. Br J Psychiatry. 2009;194:371–2. doi: 10.1192/bjp.bp.108.053843. [DOI] [PubMed] [Google Scholar]
- 69.Mayo LM, Asratian A, Linde J, Holm L, Natt D, Augier G, et al. Protective effects of elevated anandamide on stress and fear-related behaviors: translational evidence from humans and mice. Mol Psychiatry. 2020;25:993–1005. doi: 10.1038/s41380-018-0215-1. [DOI] [PubMed] [Google Scholar]
- 70.Best LM, Williams B, Le Foll B, Mansouri E, Bazinet RP, Lin L, et al. Lower brain fatty acid amide hydrolase in treatment-seeking patients with alcohol use disorder: a positron emission tomography study with [C-11]CURB. Neuropsychopharmacol. 2020;45:1286–96. doi: 10.1038/s41386-020-0606-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Caccamise A, Van Newenhizen E, Mantsch JR. Neurochemical mechanisms and neurocircuitry underlying the contribution of stress to cocaine seeking. J Neurochem. 2021;157:1697–713. doi: 10.1111/jnc.15340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.McReynolds JR, Doncheck EM, Li Y, Vranjkovic O, Graf EN, Ogasawara D, et al. Stress promotes drug seeking through glucocorticoid-dependent endocannabinoid mobilization in the prelimbic cortex. Biol Psychiatry. 2018;84:85–94. doi: 10.1016/j.biopsych.2017.09.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Bedse G, Hill MN, Patel S. 2-Arachidonoylglycerol modulation of anxiety and stress adaptation: from grass roots to novel therapeutics. Biol Psychiatry. 2020;88:520–30. doi: 10.1016/j.biopsych.2020.01.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Morena M, Patel S, Bains JS, Hill MN. Neurobiological interactions between stress and the endocannabinoid system. Neuropsychopharmacology. 2016;41:80–102. doi: 10.1038/npp.2015.166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Bilbao A, Neuhofer D, Sepers M, Wei SP, Eisenhardt M, Hertle S, et al. Endocannabinoid LTD in accumbal d1 neurons mediates reward-seeking behavior. iScience. 2020;23:100951. doi: 10.1016/j.isci.2020.100951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Kalivas PW. The glutamate homeostasis hypothesis of addiction. Nat Rev Neurosci. 2009;10:561–72. doi: 10.1038/nrn2515. [DOI] [PubMed] [Google Scholar]
- 77.Engeli EJE, Zoelch N, Hock A, Nordt C, Hulka LM, Kirschner M, et al. Impaired glutamate homeostasis in the nucleus accumbens in human cocaine addiction. Mol Psychiatry. 2021;26:5277–85. doi: 10.1038/s41380-020-0828-z. [DOI] [PubMed] [Google Scholar]
- 78.Gaetani S, Oveisi F, Piomelli D. Modulation of meal pattern in the rat by the anorexic lipid mediator oleoylethanolamide. Neuropsychopharmacology. 2003;28:1311–6. doi: 10.1038/sj.npp.1300166. [DOI] [PubMed] [Google Scholar]
- 79.Lo Verme J, Fu J, Astarita G, La Rana G, Russo R, Calignano A, et al. The nuclear receptor peroxisome proliferator-activated receptor-alpha mediates the anti-inflammatory actions of palmitoylethanolamide. Mol Pharm. 2005;67:15–9. doi: 10.1124/mol.104.006353. [DOI] [PubMed] [Google Scholar]
- 80.Petrosino S, Di, Marzo V. The pharmacology of palmitoylethanolamide and first data on the therapeutic efficacy of some of its new formulations. Br J Pharm. 2017;174:1349–65. doi: 10.1111/bph.13580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Barba SV, Kirschbaum C, Gao W. Endocannabinoid and perceived stress: association analysis of endocannabinoid levels in hair versus levels in plasma and urine. Biol Psychol. 2023;178:108541.. doi: 10.1016/j.biopsycho.2023.108541. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.