

Abstract
Culex (Barraudius) modestus (Ficalbi 1889) are found in temperate regions across Europe, Asia, and Northern Africa. These mosquitoes thrive during the summer and prefer to breed in permanent vegetative habitats such as rice paddies and marshes. Culex modestus feed on a wide range of bird species but are highly attracted to humans, which makes them a potential ‘bridge’ vector for enzootic pathogens. There is compelling evidence that Culex modestus is an efficient vector for West Nile virus, potentially capable of causing epidemics in humans and other mammals. This species is also a likely vector for Usutu virus, avian malaria (Plasmodium spp.), and parasitic heartworms (Dirofilaria spp.). Culex modestus can be morphologically identified at the larval and adult stages, and a distinctive phenotype of this species is their ability to overwinter. Despite the widespread establishment of this mosquito species and their role as vectors for human pathogens, we lack sufficient knowledge on this species to implement and evaluate targeted vector control measures. Since Culex modestus can be considered a potential public health threat, there is a need for a better understanding of this mosquito species.
Graphical Abstract

Supplementary Information
The online version contains supplementary material available at 10.1186/s13071-023-05997-6.
Keywords: Culex modestus, Mosquito, Vector, Arbovirus, Avian malaria, Canine filariasis
Background
Culex (Cx.) mosquitoes of the Culicidae family are important global vectors for human and animal pathogens, including arthropod-borne viruses (arboviruses) and eukaryotic parasites. In Europe, the northern house mosquito Cx. pipiens sensu lato has been traditionally considered the primary mosquito vector responsible for pathogen transmission. They are a known vector for West Nile virus, Japanese encephalitis virus, Rift Valley fever virus, Sindbis virus, Tahyna virus, dirofilarial worms, and avian malaria [1]. Yet, a lesser known member of the Culex genus—Cx. (Barraudius) modestus—may play an important role in pathogen transmission as well. Little is known about this vector, but evidence suggests that Cx. modestus is more anthropophilic (human-biting) and potentially more competent as a West Nile virus vector than Cx. pipiens s.l. mosquitoes. In this article we present a review of the literature, including what is known, and what is unknown, about the important but overlooked mosquito vector Cx. modestus.
Ecology
Geographic distribution
Culex modestus was first identified in 1889 in Ravenna, Italy [2], and has since been discovered across a wide landscape of countries in Europe, northern Africa, and Asia (Fig. 1). The current distribution of native Cx. modestus mosquitoes at the regional level in Europe can be found on the European Centre for Disease Prevention and Control (ECDC) website, where they provide regular updates on the distribution of native and invasive mosquito species [3].
Fig. 1.

Map of Culex modestus distribution (as of August 2023). Countries highlighted in blue represent those with at least one report of Cx. modestus, whereas those in grey indicate that Cx. modestus are absent or have not yet been reported. Sources used in the creation of this map can be found in the supplementary material (Additional file 2: Table S1). Created with Mapchart.net
The Cx. modestus genotype can be divided into lineages I and II based on haplotypes in the mitochondrial cytochrome oxidase 1 gene (COX1) [4, 5]. Lineage I has been found in Spain and Portugal, lineage II was found in Germany, Sweden, and the UK, and both lineages were found in Belgium, France, Serbia, and Denmark. A haplotype analysis suggested that Cx. modestus from the UK and Germany share a common origin from France, whilst another part of the UK population may have been derived from Serbia [5]. In Belgium, Cx. modestus were likely derived from the UK and Germany, possibly the result of several independent introduction events [4]. Phylogeny on the basis of rRNA intergenic spacers between the 28S rRNA gene (3’ end) and 18S rRNA gene (5’ end) on chromosome I revealed that Cx. modestus is highly evolutionarily distant from Cx. pipiens pipiens, Cx. pipiens molestus, and Cx. torrentium [6].
It would be interesting to investigate the origin of Cx. modestus and how they have spread inter- and cross-continentally. A small number of Cx. modestus were found breeding in used tires during a large national survey in Spain [7], alerting to a potential mode of dispersal to new areas. In a mosquito survey of international ships arriving to Hebei Province in China, more than half of the vessels were found with mosquitoes onboard [8]. Almost all the mosquitoes on these ships were Culex (97%), and Cx. modestus and Cx. tritaeniorhynchus were the dominant species. Sea travel could therefore be a likely route of migration for these species. Given the extensive establishment of Cx. modestus across Eurasia and northern Africa, potential introduction to new areas by global transport, and how the effects of climate change are expected to develop more suitable mosquito climates and habitats across the globe, this species will probably appear in new areas.
Habitat
In contrast to Cx. pipiens s.l., which can be found scattered across a wide range of ecological habitats, Cx. modestus are generally restricted to specific rural and agricultural breeding sites [9]. Their larvae are found in mostly permanent water bodies including rice fields [10–12], reedbeds [11, 13, 14], fish ponds [9, 15–17], wetlands [9, 11, 18], marshes [11, 19, 20], and woodlands [13] or deciduous forests [14]. They have been found co-habitating with other mosquito species, mostly those of the Anopheline and Culicine genera (Table 1).
Table 1.
Mosquito species found to co-habitate with Culex modestus larvae
| Genus | Species | Country | Habitat | Source |
|---|---|---|---|---|
| Aedes | Ae. (Ochlerotatus) caspius | Iran | Unspecified | [21] |
| Anopheles | An. claviger | Algeria | Peri-urban habitat | [22] |
| An. hyrcanus | Unspecified | Rice fields | [10] | |
| An. labranchiae | Algeria | Peri-urban habitat | [22] | |
| An. maculipennis s.l | Moldova | Unspecified | [23] | |
| An. melanoon | France | Rice fields and reed beds | [11] | |
| An. sacharovi | Unspecified | Rice fields | [10] | |
| Culex | Cx. longiareolata | Algeria | Peri-urban habitat | [22] |
| Cx. pipiens s.l | Algeria | Peri-urban habitat | [22] | |
| Belgium | Peri-urban habitat | [4] | ||
| Iran | Unspecified | [21] | ||
| Cx. territans | Moldova | Unspecified | [23] | |
| Cx. theileri |
Algeria Iran |
Peri-urban habitat Unspecified | [21, 22] | |
| Unspecified | Unspecified | [10] | ||
| Moldova | Unspecified | [23] | ||
| Cx. torrentium | Moldova | Unspecified | [23] | |
| Culiseta | Cs. annulata | Moldova | Unspecified | [23] |
| Cs. subochrea | Iran | Unspecified | [21] | |
| Uranotaenia | Ur. unguiculata | Unspecified | Unspecified | [10] |
| Moldova | Unspecified | [23] |
Specific details regarding the ecological and climatic factors that influence Cx. modestus breeding are based on a limited number of studies. A cross-sectional survey conducted across 46 locations in northwestern Iran described the larval habitats of Cx. modestus in detail [24]. Most Cx. modestus were found in natural habitats (85%) as opposed to artificial ones (15%), in water bodies that were permanent (90%) rather than temporary (10%). Most were found in water that was slow running (73%) as opposed to stagnant (27%) and transparent (98%) versus opaque (2%). Most habitats contained vegetation (85%) as opposed to no vegetation (15%), with a bed type not based on clay or sand (55%). Most larvae were found in semi-shade (60%) compared to sunny (4%) or shade (36%). Similarly, Cx. modestus in Algeria could only be found at peri-urban as opposed to urban habitats [22]. The peri-urban sites were described as having open, permanent, stagnant, and clean water with an average pH of 7.7. In contrast, a different larval sampling survey in Iran found more Cx. modestus in urban than rural areas; however, only a small number of Cx. modestus was recovered and represented only a small proportion of the overall mosquito population (2.4%, n = 33) [25]. The ability to grow in urban settings suggests that Cx. modestus are adaptable to diverse environments.
In a random sampling of adult mosquitoes at a nature reserve in Spain, the abundance of Cx. modestus was positively associated with inundation area (i.e. water surface) and hydroperiod (i.e. water permanence) [20]. The Cx. modestus displayed a preference for hydroperiods of > 150–200 days/year. In a 7-year surveillance study of adult mosquitoes in the Piedmont Region of Italy, proximity to rice fields was positively correlated to population clusters and abundance of Cx. modestus [26]. Elevation and distance from breeding sites, on the other hand, were negatively associated with abundance. When comparing two mosquito seasons—a wetter season with a colder average temperature (2002: 20.5 °C, 512 mm) vs. a drier season with a warmer average temperature (2003: 23 °C, 122 mm)—more Cx. modestus were captured during the wetter and colder season [26], demonstrating the importance of water surface area for Cx. modestus breeding.
The abundance of Cx. modestus was shown to be affected by agricultural changes. In fact, the rapid proliferation and subsequent near elimination of Cx. modestus are well described in the Camargue region of France [27]. Culex modestus was rarely reported in the Camargue until a massive upscale in rice cultivation following World War II. The population of Cx. modestus as well as An. hyrcanus became widespread and abundant throughout the region where they were considered major pests and nuisance biters. In that period, increases in mosquito density could thus be attributed to changes in paddy surface area, which suggests that reed marshes alone cannot sustain populations of Cx. modestus to the same extent as rice paddies. In later years, rice cultivation in France started to decrease and insecticide use in remaining paddies was implemented to combat pests. Consequently, the mosquito populations in these fields declined. Since the start of the twenty-first century, insecticide use has been slowly replaced by more sustainable pest control methods, resulting in a gradual increase in mosquito populations once again.
Biology
Lifecycle
Populations of Cx. modestus bloom during the warm summer months. Most studies that measured levels of Cx. modestus across different periods captured larger numbers in the months of July and August [19, 26, 28–35]. One study in Romania captured more Cx. modestus in late August or early September [36], because, interestingly, while more Cx. pipiens s.l. were captured over the summer months, towards the end of the season the proportion of Cx. modestus exceeded that of Cx. pipiens s.l. mosquitoes. In France, more Cx. modestus were captured in June or July [37]. One sampling study from Spain found a significantly higher proportion captured in marshlands in June (98%) than in any other habitat or time between March and November [20]. Likewise, a study from Moldova also captured a higher number in June (49.4%) than any other time between May and September [38]. A sampling study in Algeria found Cx. modestus from May to October with temperatures ranging from 17.9 at night to 31.4 °C during the day and rainfall between 0 and 66 mm [22].
Little is known about the life cycle of Cx. modestus and what sets it apart from other members of the Culex genus. Culex modestus has been reported to overwinter (diapause) [13, 39], a phenomenon characterized by the ability of female mosquitoes to ‘hibernate’ during the cold months of the year. Like Cx. pipiens pipiens and Culex tarsalis, female Cx. modestus enter diapause when temperatures begin to fall after the summer [40], which encourages female mosquitoes to fly to secluded shelters such as caves or bunkers. When outdoor temperatures begin to rise again, females exit diapause and seek a blood meal to produce a batch of eggs. This adaptation to the cold raises concern over potential arbovirus carry-over between transmission seasons [41].
The potential for Cx. modestus to produce a first batch of eggs without a prior blood meal (autogeny) was briefly mentioned in one research article in which larvae were captured in the Padurea Domneasca reserve in Moldova and reared to adulthood in the laboratory [23]. The researchers observed that female adults deposited a first batch of viable eggs after sugar feeding without a prior blood meal, and the same was observed in the next generation. This is currently the only report of autogeny in Cx. modestus.
Three research groups have reported the rearing of Cx. modestus in the laboratory. A colony from Lattes, France, was maintained at 26 ± 1 °C with > 50% RH and a 16-h photoperiod with 1.5 h of simulated dawn/dusk crepuscular periods [42, 43]. Larvae were reared in pans containing tap water and yeast tablets [43], while adults were provided with 10% sugar solution and heparinized rabbit blood for egg production [42, 43]. A laboratory colony of Cx. modestus (Beijing strain) in China was maintained at 26 ± 1 °C and 75 ± 5% RH under a 14-h photoperiod [44]. In a study from Greece that used F0 adults reared from larvae, the larvae were captured and consequently reared in the laboratory in pans (53.3 × 40.6 cm) filled with deionized water and aerated with aquarium pumps [45]. Larvae were fed a mixture of liver and yeast powder medium (2:3 ratio) under natural light-dark conditions at 25 °C and > 60% RH, and adult mosquitoes were provided with 10% sugar solution.
Host preference
Culex modestus are known to feed on birds and mammals interchangeably, which could make this species a good ‘bridge’ vector for enzootic pathogens such as West Nile virus. While it has been suggested that Cx. modestus displays a preference for feeding on birds (ornithophily) [46], certain data from literature show a strong disposition for human biting. A study evaluating human biting willingness in mosquitoes found that Cx. modestus and other human-biting (anthropophilic) species such as Anopheles plumbeus, Aedes vexans, Ae. (Ochlerotatus) sticticus, and Cx. pipiens molestus were positively correlated to human biting (> 0.7 human biting willingness), whereas species such as Cx. pipiens pipiens and Culex territans were negatively correlated (< 0.1 human biting willingness) [47]. The relative biting risk index (RBRI) for Cx. modestus (0.07 RBRI) was higher than that of 38 out of 42 species included in the study but lower than for Ae. vexans (> 0.4 RBRI), Aedes (Ochlerotatus) annulipes (0.25 RBRI), and Ae. (Ochlerotatus) sticticus (> 0.1 RBRI) [47]. Another study observed that Cx. modestus were more attracted to humans than to birds, whereas Cx. pipiens s.l. were more attracted to birds [48]. Recent evidence therefore suggests that Cx. modestus displays more anthropophilic or mammalophilic rather than ornithophilic feeding behaviour.
Known hosts of Cx. modestus are based on a limited number of studies (Table 2). Culex modestus has been shown to feed on freshwater dwellers such as ducks, geese, and herons; mammals and farmland animals such as dogs, chickens, cattle, and horses; and even birds of prey (i.e. Western Marsh harrier) and reptiles. Given the natural aquatic habitats of Cx. modestus in areas such as rice paddies and marshes, it is fitting that they feed on animals from the same habitats, such as wild aquatic birds or rural farmland animals. It is interesting that some studies have found this species to be more attracted to humans than to birds, but blood-meal host identification on field-captured mosquitoes found more mosquitoes that fed on birds than mammals. This suggests that the feeding preference of Cx. modestus depends on host abundance and availability in their current habitats.
Table 2.
Host sources of Culex modestus based on blood-meal identification or host-feeding studies
| Class | Scientific name | Common name | Country | Source |
|---|---|---|---|---|
| Bird (Aves) | Anas acuta | Northern pintail | Spain | [100] |
| Anas platyrhynchos | Mallard | Czech Republic | [9] | |
| Spain | [46, 100] | |||
| Anas strepera | Gadwall | Czech Republic | [9] | |
| Spain | [100] | |||
| Anas sp. | Duck (unspecified) | China | [53] | |
| France | [30] | |||
| Spain | [101] | |||
| Anser anser | Greylag goose | Czech Republic | [9] | |
| Spain | [46, 100] | |||
| Ardea cinerea | Grey heron | Spain | [100] | |
| Branta canadensis | Canada goose | Spain | [100] | |
| Chen caerulescens | Snow goose | Spain | [100] | |
| Circus aeruginosus | Western Marsh harrier | Czech Republic | [9] | |
| Cygnus olor | Mute swan | United Kingdom | [28] | |
| Egretta garzetta | Little egret | Spain | [100] | |
| Emberiza citrinella | Yellowhammer | Czech Republic | [9] | |
| Erithacus rubecula | European robin | Czech Republic | [9] | |
| Falco vespertinus | Red-footed falcon | Hungary | [96] | |
| Fringilla coelebs | Eurasian chaffinch | Czech Republic | [9] | |
| Gallus gallus | Red junglefowl | Spain | [46, 100] | |
| Gallus sp. | Chicken (unspecified) | Russia | [87] | |
| Hirundo rustica | Barn swallow | Czech Republic | [9] | |
| United Kingdom | [28] | |||
| Muscicapa striata | Spotted flycatcher | Czech Republic | [9] | |
| Nycticorax nycticorax | Black-crowned night heron | Czech Republic | [9] | |
| Parus caeruleus | Blue tit | Czech Republic | [9] | |
| Parus major | Great tit | Czech Republic | [9] | |
| Passer domesticus | House sparrow | Czech Republic | [9] | |
| Sturnus vulgaris | Common starling | Czech Republic | [9] | |
| Tadorna ferruginea | Ruddy shelduck | Spain | [100] | |
| Tadorna tadorna | Common shelduck | Spain | [100] | |
| Turdus merula | Eurasian blackbird | Czech Republic | [9] | |
| Unspecified | Unspecified | Spain | [34, 46] | |
| Mammal (Mammalia) | Bos taurus | Cow | China | [33] |
| Russia | [87] | |||
| Canis familiaris | Dog | Italy | [91] | |
| Russia | [87] | |||
| Equus caballus | Horse | France | [30] | |
| Russia | [87] | |||
| Homo sapiens | Human | Russia | [87] | |
| UK | [51] | |||
| Sus domesticus | Pig | China | [33] | |
| Unspecified | Unspecified | Spain | [34, 46] | |
| Reptile (Reptilia) | Unspecified | Unspecified | Spain | [34] |
Host-seeking behaviour
Understanding the host-seeking behaviour of mosquitoes can help to elucidate their vectorial capacity and determine potential target areas for vector and pest control. Culex modestus are nuisance biters described as having painful bites [10, 49, 50]. The hourly human biting rate of Cx. modestus captured in England was found to be 2.5 with a range of 0–55 [51], but most likely the daily bites and frequency of feeding will depend on the local mosquito abundance and climate. Culex modestus are nocturnal feeders, but different studies have reported differences in peak biting activity. One research group observed different host-seeking activities depending on the mode of trapping [32]. With CO2-baited traps, the host-seeking pattern was unimodal with peak trapping in the evening (87.0%) and to a lesser extent during the night (12.2%) or early morning (0.7%). With human landing catches (HLCs), host-seeking activity was bimodal with most adult mosquitoes still captured in the evening (84.9%) but a non-negligible proportion captured in the morning (15.1%). The same study found that the timing of the sunset as well as relative humidity (RH) influenced the initiation of flight and peak flight activity in this species. In another study, the biting activity of Cx. modestus ranged from – 0.5 to + 2 h from sunset, with peak biting at + 1 h [51]. A different study observed that Cx. modestus fed exclusively at night with peak activity from 22:00 to 00:00, but the timing in relation to the sunset was not reported [52]. Finally, one article reported a higher number of Cx. modestus captured on average at dusk (21:00 to 23:00) than at dawn (05:00 to 07:00), but the difference in mean number captured per night was small (n = ~ 180 at dusk, n = ~ 120 at dawn) [53].
Limited evidence on the flight range of Cx. modestus suggests that they have a restricted travel radius. Compared to Cx. pipiens s.l., Cx. modestus was less likely to stray from the shorelines of breeding sites [9]. The radial distance of active dispersion around an emergence site for Cx. modestus was estimated to be 700 m, shorter than that of An. melanoon (1000 m) which co-habitated in the same breeding sites [11]. A trapping study that captured mosquitoes using traps placed at different distances from the ground measured a mean flying height of 2.26 m (± 0.57) for Cx. modestus, which was lower than that of Cx. pipiens s.l. (2.66 m ± 0.90) but higher than for Coquillettidia richiardii (2.00 m ± 1.00), Aedes detritus (1.98 m ± 0.71), and Ae. caspius (1.74 m ± 0.24) [54]. A different study observed that more Cx. modestus were captured in traps placed 5 m above the ground than at 1 or 3 m, suggesting that this altitude reflected the ornithophilic behaviour of the species [53].
Observations of Cx. modestus in Volgograd, Russia, found that they could feed interchangeably indoors (endophagy) and outdoors (exophagy) [48]. Culex modestus fed indoors more frequently than Cx. pipiens s.l., and they fed more often on humans and other mammals both indoors and outdoors than Cx. pipiens s.l. mosquitoes. As the Cx. modestus in this study were found in both rural and urban areas, more research is needed to understand their feeding behaviour across different habitats in other locations.
Microbiome
Unravelling the microbiome of mosquito vectors can help to identify potential targets for control. A prime example of a microbe used in the biological control of mosquitoes is Wolbachia pipientis, an intracellular bacterium which has been successfully used as an antiviral strategy against dengue virus in endemic areas [55]. While Wolbachia is known to persistently infect Culex spp. [56], particularly Cx. pipiens s.l., which has a high prevalence of Wolbachia, current evidence suggests that most Cx. modestus do not carry this endosymbiont [57, 58]. Mosquitoes from Eastern Europe (7% prevalence rate) [58] and Italy (unknown prevalence rate) [57] have been found infected with Wolbachia, while one study with a low sample size (n = 11) did not find any Wolbachia-infected Cx. modestus from Belgium [59]. It would be interesting to investigate whether Wolbachia can modify arbovirus infection in Cx. modestus, as it has been shown to do for other species [60].
Viral metagenomic analyses of field-captured mosquitoes have identified insect-specific viruses (ISVs) belonging to Cx. modestus captured from Belgium [4] and eastern Macedonia and northern Greece [61] (Table 3). The latter identified several core ISVs that overlapped between Cx. modestus and Cx. theileri, An. melanoon, and Uranotaenia unguiculata, which have been found to co-habitate together. The potential role of these ISVs on the life history, survival, and vector competence of Cx. modestus mosquitoes is not known.
Table 3.
Insect-specific viruses found in Culex modestus
| Type | Family | Name | Source |
|---|---|---|---|
| (-)ssRNA | Orthomyxoviridae | Culex orthomyxo-like virus | [4, 61] |
| dsRNA | Chrysoviridae | Hubei chryso-like virus 1 | [61] |
| Partitiviridae | Atrato Partiti-like virus 3 | [4] | |
| Beihai partiti-like virus 2 | [4, 61] | ||
| Sonnbo virus | [4] | ||
| ( +)ssRNA | Picornaviridae | Ista virus | [4] |
| Virgaviridae | Alexandroupolis virga-like virus | [61] | |
| dsRNA | Totiviridae | Culex inatomii totivirus | [4, 61] |
| Fitzroy Crossing toti-like virus | [4] | ||
| N/A | Unclassified | Yongsan negev-like virus 1 | [4] |
Microsporidians of the genus Cristulospora have been found in Cx. modestus from Uzbekistan [62], and the arthropod parasite Crithidia brevicula was found in mosquitoes from the Czech Republic [63]. It has been shown that Cx. modestus larvae can be infected with the parasitic mermithid nematode Romanomermis iyengari [64] and the fungal parasite Coelomomyces iliensis [65]. Further research is needed to characterize the complete microbiome of Cx. modestus, including the bacteriome and mycobiome, across a wider geographical landscape.
Morphology and Identification
Morphology
Upon its discovery by Ficalbi in 1889, Cx. modestus was described using the phrase “zanzara di colorito modesto”, which translates to “mosquito of modest colouring”. This is likely the origin of the modestus name, as this species is small with homogeneously brown features. The morphology of Cx. modestus and other members of the Barraudius Edwards subgenus is described in detail by Becker et al. 2010 [10].
The unique characteristics of Cx. modestus that are used for their morphological identification can be found in the terga, hind legs, male gonocoxite, and larval siphons [10]. The terga display dark triangular patches over the longitudinal abdominal bands (Fig. 2A). In the hind legs, tarsomere I is distinctly shorter than the hind tibia (Fig. 2B) [10]. A microscopic image of adult Cx. modestus compared to Cx. pipiens pipiens can be found in the supplemental material (Additional file 1: Fig. S1). The male gonostylus is long, comprising half the length of the gonocoxite, and the ventral arm of the aedeagus is short, not extending beyond the apex of the paraproct (Fig. 2C–D) [10]. In the larval siphon, the tufts 1-S are arranged in a “ventral zig zag row” towards the apex of the siphon, with one tuft usually inserted in the pecten, and the saddle seta 1-X have two or three branches (Fig. 2E) [10]. Due to general similarities in morphology, Cx. modestus can be confused with Ae. cinereus or Cx. martini [10].
Fig. 2.
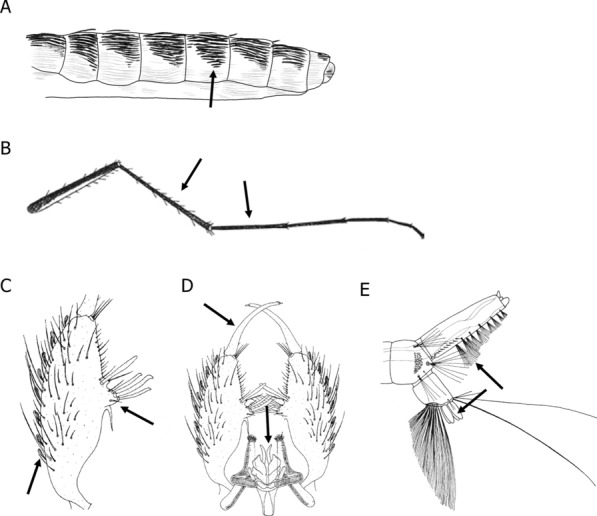
Identifiable characteristics of Culex modestus. A Schematic of adult abdomen; B adult hind leg; C male gonocoxite; D male hypopygium; E larval siphon. Schematic A made using Procreate.com. Schematics B–E were reproduced and adapted from Becker et al. 2010 [10] with permission from Springer Nature. The black arrows indicate the unique identifiable features
Molecular identification
Currently, the primary method of molecular detection of Cx. modestus is DNA barcoding. This entails PCR amplification of a target sequence such as the mitochondrial COX1 or ribosomal ITS or rDNA genes to produce a “universal” fragment, which, when sequenced, can be used to identify a range of insect species [66]. A common barcoding target is the 710-bp fragment in COX1 described by Folmer et al. [67]. COX1 is the largest of the mitochondrial subunit-encoding genes and is highly conserved with a slow rate of nucleotide substitution, making it an excellent target for insect species identification. Diagnostic enzyme markers are another target for molecular identification, as malate dehydrogenase 2 and adenylate kinase have been identified as suitable allozyme markers for the taxonomy of Cx. modestus [68].
Other Culex species have several molecular markers that can be used for identification, including microsatellite loci (e.g. CQ11), nuclear genes such as ace-2 [69], and the ribosomal ITS region. While morphological identification is considered the gold standard method for mosquito identification, morpho-taxonomy can be highly time-consuming and often misleading if samples are handled improperly or if performed by an inexperienced taxonomist. For large-scale studies capturing high volumes of mosquitoes, a PCR- or RT-PCR-based method of identification would be ideal.
Disease transmission
Vector incrimination is necessary to determine if an arthropod species is a natural vector of disease. Following the same principle as Koch’s postulates, vector incrimination determines if there is an association in space and time between the arthropod species and human cases of the disease, evidence of direct contact between the arthropod species and the humans, and evidence that the arthropod can harbour and transmit the pathogen [70]. While there is significant evidence of Cx. modestus attraction and willingness to bite humans, eco-epidemiological modelling data for this species are lacking. Understanding vector population dynamics and transmission risk over space and time is needed to anticipate potential outbreaks and evaluate the impact of vector control or outbreak mitigation strategies.
The ability of a species to acquire, maintain, and transmit a pathogen is defined as vector competence. Vector competence studies of mosquitoes typically involve measures of infection rate, dissemination rate, and transmission rate as the proportion of mosquitoes with pathogens in the body, head, and saliva, respectively. Mosquitoes with detectable pathogens in the saliva are considered competent vectors, with the proportion of competent mosquitoes in a species expressed as the transmission efficiency. Ideally, mosquito vector competence is determined using living mosquitoes to determine the presence of infectious pathogens in the saliva. Vector competence studies are preferable to the detection of DNA or RNA in field-captured mosquitoes, as the detection of nucleic acid alone is not sufficient evidence to implicate an insect as a vector. However, to understand the true risk of mosquito-borne disease transmission in a given area, measures of vector competence of a species can be used to determine their vectorial capacity. Vectorial capacity, or the Ross-Macdonald model, is defined as the number of infectious mosquito bites per day, a rate comparable to the basic reproduction number (R0) [71]. Vectorial capacity includes the human biting rate, the ratio of mosquitoes to humans (or animals), the vector’s daily probability of survival, the extrinsic incubation period of the pathogen, and the vector competence. Currently, there are no estimated measures of Cx. modestus vectorial capacity anywhere in the world, but measures of vector competence can serve as a proxy for estimated transmission risk.
Culex modestus is a potential vector for several pathogenic viruses and parasites (Table 4). Evidence from epidemiological modelling, vector competence studies, and field detection data support the case that Cx. modestus is a competent vector for West Nile virus. Additionally, the evidence that this species is a vector for Usutu virus, avian malaria, and dirofilarial worms is compelling. However, several pathogens have only been detected via RNA or DNA screenings of field-captured Cx. modestus, including Japanese encephalitis virus [33], Sindbis virus [72], Kyzylagach virus [73], Tahyna virus [17, 39, 74, 75], Batai virus [76], Lednice virus [77], Ebinur Lake virus (previously named Abbey Lake virus) [78, 79], Zaliv Terpeniya virus [80], Banna virus [74, 81], and Haemoproteus sp. [34]. However, whether Cx. modestus is a competent vector for these pathogens requires further investigation.
Table 4.
Reports of vector competence or molecular detection of infectious pathogens in Culex modestus
| Family | Pathogen | Detection method | Mosquito type | Country | Source | |
|---|---|---|---|---|---|---|
| Virus | Flaviviridae | West Nile virus | RNA detection | Field | Czech Republic | [9, 15, 16, 102] |
| RNA detection | Field | Italy | [103] | |||
| RNA detection | Field | Kazakhstan | [104] | |||
| RNA detection | Field | Romania | [72] | |||
| RNA detection | Field | Russia | [48, 87] | |||
| Vector competence | Lab | China | [44] | |||
| Vector competence | Lab | France | [42, 43] | |||
| Usutu virus | RNA detection | Field | Czech Republic | [16, 89] | ||
| Vector competence | Field | Belgium | [59] | |||
| Japanese encephalitis virus | RNA detection | Field | China | [33] | ||
| Alphaviridae | Sindbis virus | RNA detection | Field | Romania | [72] | |
| Kyzylagach virus | RNA detection | Field | Czech Republic | [73] | ||
| Peribunyaviridae | Tahyna virus | Immunological detection | Field | France | [39] | |
| Immunological & RNA detection | Field | China | [74] | |||
| Immunological detection | Field | Czech Republic | [17, 75] | |||
| Batai virus | RNA detection | Field | Germany | [76] | ||
| Lednice virus | RNA detection | Field | Czech Republic | [77] | ||
| Ebinur Lake virus (Abbey Lake virus) | RNA detection | Field | China | [78, 79] | ||
| Phenuiviridae | Zaliv Terpeniya virus | RNA detection | Field | Azerbaijan | [80] | |
| Reoviridae | Banna virus | RNA detection | Field | China | [74, 81] | |
| Eukaryota | Onchocercidae | Dirofilaria immitis | Vector competence | Field | Italy | [90] |
| Plasmodiidae | Plasmodium relictum | Vector competence | Lab | Bulgaria | [14] | |
| Plasmodium sp. | DNA detection | Field | Romania | [92] | ||
| DNA detection | Field | Spain | [34] | |||
| Haemoproteidae | Haemoproteus sp. | DNA detection | Field | Spain | [34] | |
| Trypanosomatidae | Trypanosoma corvi/culicavium | Microscopic examination | Field | Czech Republic | [63] |
West Nile virus (WNV) is a member of the Flaviviridae family and the Japanese encephalitis serocomplex. The virus is maintained in an enzootic transmission cycle between avians and mosquito vectors and can cause severe neuroinvasive disease in humans and other mammals. WNV was first discovered in Uganda in 1937 and is widespread across the globe, causing seasonal outbreaks in temperate regions. The largest recorded outbreak of WNV occurred in the USA in 2002 with more than 4156 human cases, 2942 reports of meningoencephalitis, and 284 deaths [82]. Climatic and environmental shifts driven by the effects of climate change are expected to increase the incidence and spread of WNV in the future [83]. Epidemiological data from Southern France suggested that Cx. modestus was the main amplifier of WNV over Cx. pipiens s.l. [84], though equine WNV outbreaks in Italy were significantly associated with suitable habitats for both Cx. pipiens s.l. and Cx. modestus [85]. In contrast to these studies, Cx. modestus was estimated to be a less significant amplifier of equine WNV in Croatia than Cx. pipiens s.l. and Ae. vexans due to their low abundance [86]. The estimated relative risk for WNV transmission was on average 35.2 for Cx. pipiens s.l., 65.4 for Ae. vexans, and only 2.0 for Cx. modestus. In a study that used laboratory-colonized mosquitoes from France, Cx. modestus was shown to have a higher vector competence for WNV than Cx. pipiens pipiens [43]. The dissemination and transmission rates were 89.2% and 54.5% for Cx. modestus and 38.5% and 15.8% for Cx. pipiens pipiens, respectively. In contrast, a different study of laboratory colonies reported a lower WNV vector competence in Cx. modestus (35% transmission rate) compared to Cx. pipiens pallens from Beijing, China (48% transmission rate) [44]. A surveillance study in Russia found a higher WNV infection rate in field-caught Cx. modestus than in Cx. pipiens pipiens (2.72% vs. 0.79%), despite having a much lower sample size (370 Cx. modestus vs. 1261 Culex pipiens pipiens) [87]. In Romania, Cx. modestus had an infection rate of 1.81 per 1000 mosquitoes for WNV, whereas Cx. pipiens s.l. had an infection rate of 0.77 per 1000 mosquitoes [72]. Culex modestus from Russia had an infection rate estimate of 0.24 per 1000 mosquitoes, lower than that of Cx. pipiens s.l. (0.51 per 1000 mosquitoes) [48]. These data combined suggest Cx. modestus may play an important role in WNV circulation.
Usutu virus (USUV) is another flavivirus antigenically similar to WNV. USUV rarely causes symptomatic infections in humans but can result in rapid and destructive outbreaks among birds [88]. RNA of Usutu virus has been detected in pools of Cx. modestus captured from the Czech Republic [16, 89]. Another study using field-captured mosquitoes from Belgium investigated the vector competence of Cx. modestus to USUV [59]. The sample size of Cx. modestus included in the latter study was low (n = 5) but the transmission potential for USUV was high (20% transmission efficiency), suggesting that Cx. modestus could be potent vectors for USUV. The co-circulation of WNV and USUV in the same habitat has been reported in the Czech Republic [16], but given that these two arboviruses share considerable geographical overlap, co-circulation is likely not an uncommon occurrence. Because of their preferred habitat for rural wetland areas, farmers, hunters, and people living around ponds and farmland animals, such as horses, may have a higher risk of WNV or USUV infection by Cx. modestus [9].
Culex modestus has been implicated as a vector of the parasitic heartworm Dirofilaria immitis with a vector efficiency index of 2.0, higher than that of An. maculipennis (1.4) and Cx. pipiens s.l. (1.2), but lower than in Ae. caspius (6.3) [90]. Data suggest that these four species including Cx. modestus were primarily responsible for the transmission of canine filariasis in Piedmont, Italy [91]. Field Cx. modestus captured from Spain were found positive for avian malaria (Plasmodium sp., 0.5–6.3% prevalence) and Haemoproteus sp. (unknown prevalence) [34]. In mosquitoes captured from Romania, one positive pool of Cx. modestus containing Plasmodium sp. was found [92]. In addition, there is evidence from a laboratory investigation that a small number of Cx. modestus could transmit Plasmodium relictum sporozoites in the salivary glands [14]. In a survey of blood-sucking insects in the Czech Republic, Cx. modestus were found carrying the avian trypanosome Trypanosoma corvi/culicavium [63].
Capture and control
Trapping methods
Wild Cx. modestus have been captured using a variety of different trapping methods, including HLCs [32, 51, 52], BG-Sentinel traps baited with CO2 [4, 13, 59, 93], CDC light traps baited with CO2 [12, 15, 26, 52, 53, 61], Mosquito Magnet Pro traps [19, 28, 29], and larval dipping [18, 19, 21, 29, 94]. Other trapping methods include CO2-baited traps (undescribed) [32, 95], CO2-baited CDC traps without light [35], CDC traps without light baited with a Japanese quail or rabbit [9], CO2-baited CDC traps with sentinel birds (Japanese quail or chicken) or sentinel mammals (rabbit or guinea pig) [35], the EVS/CO2 Mosquito Trap [14], and a resting box baited with Red-footed falcon nestlings [96].
In studies that have compared trapping methods, either a majority or an equal number of Cx. modestus adults could be captured with HLCs or CO2-baited traps. The only exception is a study performed in the UK which captured significantly higher proportions of Cx. modestus with the Mosquito Magnet Pro than with resting boxes [28]. Regarding Cx. pipiens s.l., more Cx. modestus were captured with HLCs than with CO2-baited traps in Italy [32]. In France, more Cx. modestus were captured with CO2-baited traps (38 mosquitoes per trap night) and HLCs (33 mosquitoes per trap night) than with pigeon-baited traps (0.3 mosquitoes per trap night), and no Cx. modestus were caught using resting boxes (336 trap nights) or gravid traps (176 trap nights) [95]. Similar to Cx. modestus, more Cx. pipiens s.l. were captured using CO2-baited traps than the pigeon-baited traps (78 vs. 7 mosquitoes per trap night), despite this species being highly ornithophilic, suggesting that CO2-baited traps are more attractive to these species than the use of live birds. Overall, the CO2-baited traps collected two-thirds of all mosquitoes as well as sampled all the 14 species identified in the study [95]. A much lower number of Cx. pipiens s.l. were caught with HLCs than Cx. modestus (3 vs. 33 mosquitoes per trap night), suggesting that Cx. modestus is more attracted to humans than Cx. pipiens s.l. [95]. Similarly, a study from China captured an equal number of Cx. modestus in CO2-baited CDC light traps (n = 260) and HLCs (n = 258) [52]. Interestingly, although Cx. modestus could be captured using human bait, no Cx. pipiens molestus were caught using this technique, despite the anthropophilic behaviour of the molestus biotype, whereas they could be caught with CO2-baited CDC light traps (n = 267) [52].
Traps can be coupled with lures or attractants to enhance mosquito attraction. CO2, released by dry ice or fermentation, simulates animal respiration, which is a long-distance attractant for mosquitoes. BG lure and BG Sweetscent (Biogents AG, Regensburg, Germany) mimic the scent of human and/or animal skin and can be coupled with CO2-baited and/or light traps. The Mosquito Magnet Pro trap uses Octenol as a lure, which selectively attracts mammalophilic species. Culex modestus was the dominant mosquito species captured using the Mosquito Magnet Pro in a surveillance study in the UK compared to Cx. pipiens s.l./Cx. torrentium, suggesting that Cx. modestus is more attracted to mammalian hosts than the latter species [29]. In a different study, CDC light traps (without light) baited with CO2 from dry ice captured 97% of all Cx. modestus mosquitoes compared to horse urine, acetone, O-1-octen-3-ol, and ammonium hydroxide [93].
Depending on the purpose of the research, when aiming to capture high densities of Cx. modestus, CO2-baited traps could be the best choice as they are less labour-intensive than HLCs. When the aim is to capture host-seeking adults that are specifically human-biting, HLC is the most suitable method; otherwise, the use of a trap coupled with a human- or animal-scented lure would be appropriate.
Insecticides and insecticide resistance
In the event of an arbovirus outbreak, interventions should be carefully chosen to target mosquito populations responsible for transmission. Insecticides delivered through, for example, space spraying can be used to kill adult vectors, while other interventions such as larval source management can be used to reduce the density of vector populations. Currently, insecticide research specifically targeting Cx. modestus is scarce. Exposure to 5 mg/l avermectin-impregnated fine plant powder was significantly toxic to Cx. modestus larvae [97]. Two water-based pyrethroid formulations (Aqua-K-Othrine, 2% deltamethrin, and Pesguard S102, 10% d-phenothrin) were tested for their potential use in aerial spraying to control adult mosquitoes, of which both pyrethroids showed efficacy against Cx. modestus and other riceland mosquitoes in Greece [45]. Space spraying campaigns to target adult host-seeking mosquitoes would be best carried out during the peak hours of the night when the mosquitoes are most active [52].
The widespread use of insecticides for the control of agricultural pests and disease vectors has led to a global rise in genetic and phenotypic insecticide resistance in mosquitoes. Only one study from Belgium has reported the presence of insecticide resistance markers in Cx. modestus [98]. In 51 captured mosquitoes, knockdown resistance to pyrethroids and DDT (L1014F) was 43%, while the acetylcholinesterase-1 substitution conferring resistance to organophosphates and carbamates (G119S) was 3.9% [98]. Insecticide-resistant mosquito populations may be less susceptible to vector control responses that are insecticide-based, and there is also evidence that resistance mutations can increase arbovirus dissemination in the mosquito [99]. It is important to monitor insecticide resistance in areas with known or suspected mosquito-borne pathogen transmission.
Conclusion
Despite their extensive presence and capacity to spread human pathogens, little is known about Cx. modestus. These mosquitoes are widespread across the European continent and parts of Asia and northern Africa, but their origin and migration patterns are insufficiently described. Culex modestus are adapted to temperate regions, thriving during the warm summer months whilst able to survive cold winters. This species feeds on a wide range of bird species, but nonetheless appears to be highly attracted to humans, suggesting it could be a model ‘bridge’ vector for enzootic pathogens. While this species is likely an intermediate vector for West Nile virus, Usutu virus, canine filariasis, and avian malaria, more research is needed to implicate Cx. modestus in the transmission of other viruses and parasites. For example, Sindbis virus, Tahyna virus, and Trypanosoma sp. have been detected in field-captured Cx. modestus, but no studies on their vector competence for these pathogens have been carried out. Culex modestus are known to harbour insect-specific viruses, but limited knowledge is available on the bacteria or fungi that colonize them. It would be especially interesting to investigate whether Wolbachia has any effect on the life history traits or vector competence for this species. While Cx. modestus appears to be a rural pest, it remains unclear whether these mosquitoes prefer feeding indoors or outdoors and how well they survive in more urban areas. Mosquitoes captured from Belgium were found to be insecticide-resistant to all four major classes of insecticides used in public health, while it is unknown if mosquitoes from other areas carry these adaptations, potentially complicating future vector control strategies. Although Cx. modestus is a potential vector for human pathogens, there is still a significant lack of baseline knowledge on this species. More research on the biology and behaviour of Cx. modestus is thus urgently needed. As a priority, the vectorial capacity of Cx. modestus to WNV in hotspot areas should be determined (including measures of mosquito abundance, human-biting rate, etc.) to implicate this species as a capable WNV vector. Mosquito abundance studies can be done in parallel to research into the behaviour of Cx. modestus, for example to determine feeding patterns (endophagy vs. exophagy), resting preferences (endophily vs. exophily), and breeding sites (urban vs. rural), which can be used to determine optimal target sites for mosquito control. Finally, if this species is confirmed as an efficient vector of WNV, cost-effective methods of reducing populations of Cx. modestus, such as larval source reduction or insecticide-based interventions, should be investigated for WNV outbreak prevention and mitigation.
Supplementary Information
Additional file 1: Fig. S1. Comparison of A Culex pipiens pipiens and B Culex modestus. The black arrows indicate the unique characteristics for morphological identification of Culex modestus.
Additional file 2: Table S1. Studies with reports of Culex modestus by country.
Acknowledgements
We thank Xin Zhang, Louna Haliche, and Alex Calzoni for their translations of non-English publications, Alekxander Binderup for his help with finding references, and Lander De Coninck for his advice regarding the mosquito virome.
Abbreviations
- Ae.
Aedes
- An.
Anopheles
- CDC
Centers for Disease Prevention and Control
- COX1
Cytochrome oxidase 1
- Cs.
Culiseta
- Cx.
Culex
- ECDC
European Centre for Disease Prevention and Control
- HLC
Human landing catch
- ISV
Insect-specific virus
- RBRI
Relative biting risk index
- RH
Relative humidity
- S.l.
Sensu lato
- Ur.
Uranotaenia
- USUV
Usutu virus
- WNV
West Nile virus
Author contributions
AS conducted the review and wrote the manuscript; LD edited the manuscript. Both authors read and approved the final manuscript.
Funding
Not applicable.
Availability of data and materials
All data generated or analyzed during this study are included in this published article and its supplementary information files.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Culex pipiens—Factsheet for experts. 2020. https://www.ecdc.europa.eu/en/infectious-disease-topics/related-public-health-topics/disease-vectors/facts/mosquito-factsheets
- 2.Ficalbi E. Notizie preventive sulle zanzare italiane. IV. Nota preventiva. Descrizione di una specie nuova Zanzara di colorito modesto Culex modestus, n sp. Bull Della Soc Entomol Ital. 1890;21:93–94. [Google Scholar]
- 3.European Centre for Disease Prevention and Control (ECDC). Mosquito maps. 2023
- 4.Wang L, Rosales Rosas AL, De Coninck L, et al. Establishment of Culex modestus in belgium and a glance into the virome of belgian mosquito species. mSphere. 2021 doi: 10.1128/mSphere.01229-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hernández-Triana L, Brugman V, Pramual P, et al. Genetic diversity and population structure of Culex modestus across Europe: does recent appearance in the United Kingdom reveal a tendency for geographical spread? Med Vet Entomol. 2019;1:34. doi: 10.1111/mve.12412. [DOI] [PubMed] [Google Scholar]
- 6.Shaikevich EV, Zagoskin MV, Mukha DV. Comparative characteristics of the intergenic spacer of the ribosomal RNA gene cluster in mosquitoes of the genus Culex (Diptera: Culicidae) Mol Biol. 2013;47:364–372. doi: 10.7868/s0026898413030129. [DOI] [PubMed] [Google Scholar]
- 7.Roiz D, Eritja R, Escosa R, et al. A survey of mosquitoes breeding in used tires in Spain for the detection of imported potential vector species. J Vector Ecol. 2007;32:10. doi: 10.3376/1081-1710(2007)32[10:asombi]2.0.co;2. [DOI] [PubMed] [Google Scholar]
- 8.Nie WZ, Li JC, Li DX, et al. Mosquitoes found aboard ships arriving at Qinhuangdao Port PR. China. Med Entomol Zool. 2004;55:333–5. [Google Scholar]
- 9.Radrova J, Seblova V, Votypka J. Feeding behavior and spatial distribution of Culex Mosquitoes (Diptera: Culicidae) in Wetland Areas of the Czech Republic. J Med Entomol. 2013;50:1097–1104. doi: 10.1603/me13029. [DOI] [PubMed] [Google Scholar]
- 10.Becker N, Petrić D, Zgomba M, et al. Mosquitoes and their control. 2. Heidelberg: Springer Nature; 2010. pp. 1–577. [Google Scholar]
- 11.Cailly P, Balenghien T, Ezanno P, et al. Role of the repartition of wetland breeding sites on the spatial distribution of Anopheles and Culex, human disease vectors in Southern France. Parasit Vectors. 2011;4:65. doi: 10.1186/1756-3305-4-65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chaskopoulou A, Dovas CI, Chaintoutis SC, et al. Detection and early warning of West Nile virus circulation in Central Macedonia, Greece, using sentinel chickens and mosquitoes. Vector-Borne Zoonotic Dis. 2013;13:723–732. doi: 10.1089/vbz.2012.1176. [DOI] [PubMed] [Google Scholar]
- 13.Rudolf I, Šikutová S, Šebesta O, et al. Overwintering of Culex modestus and other mosquito species in a reedbed ecosystem, including arbovirus findings. J Am Mosq Control Assoc. 2020;36:257–260. doi: 10.2987/20-6949.1. [DOI] [PubMed] [Google Scholar]
- 14.Dimitrov D, Bobeva A, Marinov MP, et al. First evidence for development of Plasmodium relictum (Grassi and Feletti, 1891) sporozoites in the salivary glands of Culex modestus Ficalbi, 1889. Parasitol Res. 2023;0123456789:1–5. doi: 10.1007/s00436-023-07853-z. [DOI] [PubMed] [Google Scholar]
- 15.Rudolf I, Bakonyi T, Šebesta O, et al. West Nile virus lineage 2 isolated from Culex modestus mosquitoes in the Czech Republic, 2013: expansion of the European WNV endemic area to the North? Eurosurveillance. 2014;19:20867. doi: 10.2807/1560-7917.es2014.19.31.20867. [DOI] [PubMed] [Google Scholar]
- 16.Rudolf I, Bakonyi T, Šebesta O, et al. Co-circulation of Usutu virus and West Nile virus in a reed bed ecosystem. Parasit Vectors. 2015;8:1–5. doi: 10.1186/s13071-015-1139-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Danielová V, Holubová J. Two more mosquito species proved as vectors of Tahyna virus in Czechoslovakia. Folia Parasitol . 1977;24:187–189. [PubMed] [Google Scholar]
- 18.Pradel JA, Martin T, Rey D, et al. Is Culex modestus (Diptera: Culicidae), vector of West Nile Virus, Spreading in the Dombes Area, France? J Med Entomol. 2009;46:1269–1281. doi: 10.1603/033.046.0604. [DOI] [PubMed] [Google Scholar]
- 19.Golding N, Nunn MA, Medlock JM, et al. West Nile virus vector Culex modestus established in southern England. Parasit Vectors. 2012;5:32. doi: 10.1186/1756-3305-5-32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Roiz D, Ruiz S, Soriguer R, et al. Landscape effects on the presence, abundance and diversity of mosquitoes in mediterranean wetlands. PLoS ONE. 2015 doi: 10.1371/journal.pone.0128112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Saghafipour A, Abai MR, Farzinnia B, et al. Mosquito (Diptera: Culicidae) fauna of Qom Province. Iran J Arthropod-Borne Dis. 2012;6:54–61. [PMC free article] [PubMed] [Google Scholar]
- 22.Arroussi DER, Bouaziz A, Boudjelida H. Mosquito survey reveals the first record of Aedes (Diptera: Culicidae) species in urban area, Annaba district. Northeastern Algeria Pol J Entomol. 2021;90:14–26. [Google Scholar]
- 23.Sulesco TM, Toderas LG, Uspenskaia IG, et al. Larval habitats diversity and distribution of the mosquito (Diptera: Culicidae) Species in the Republic of Moldova. J Med Entomol. 2015;52:1299–1308. doi: 10.1093/jme/tjv142. [DOI] [PubMed] [Google Scholar]
- 24.Adham D, Moradi-Asl E, Vatandoost H, et al. Ecological niche modeling of West Nile virus vector in northwest of Iran. Oman Med J. 2019;34:514–520. doi: 10.5001/omj.2019.94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Khoobdel M, Keshavarzi D, Hassan Mossa-Kazemi S, et al. Species diversity of mosquitoes of the Genus Culex (Diptera, Culicidae) in the coastal areas of the Persian Gulf. AIMS Public Health. 2019;6:99–106. doi: 10.3934/publichealth.2019.2.99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Bisanzio D, Giacobini M, Bertolotti L, et al. Spatio-temporal patterns of distribution of West Nile virus vectors in eastern Piedmont Region, Italy. Parasit Vectors. 2011;4:230. doi: 10.1186/1756-3305-4-230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ponçon N, Balenghien T, Toty C, et al. Effects of local anthropogenic changes on potential malaria vector Anopheles hyrcanus and West Nile virus vector Culex modestus, Camargue. France Emerg Infect Dis. 2007;13:1810–1815. doi: 10.3201/eid1312.070730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Brugman VA, Hernández-Triana LM, England ME, et al. Blood-feeding patterns of native mosquitoes and insights into their potential role as pathogen vectors in the Thames estuary region of the United Kingdom. Parasit Vectors. 2017;10:1–12. doi: 10.1186/s13071-017-2098-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Vaux AGC, Gibson G, Hernandez-Triana LM, et al. Enhanced West Nile virus surveillance in the North Kent marshes UK. Parasit Vectors. 2015 doi: 10.1186/s13071-015-0705-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Balenghien T, Fouque F, Sabatier P, et al. Horse-, Bird-, and human-seeking behavior and seasonal abundance of mosquitoes in a West Nile Virus Focus of Southern France. J Med Entomol. 2006;43:936–946. doi: 10.1603/0022-2585(2006)43[936:hbahba]2.0.co;2. [DOI] [PubMed] [Google Scholar]
- 31.Šebesta O, Gelbič I, Peško J. Seasonal dynamics of mosquito occurrence in the lower Dyje River Basin at the Czech-Slovak-Austrian border. Ital J Zool. 2013;80:125–138. [Google Scholar]
- 32.Veronesi R, Gentile G, Carrieri M, et al. Seasonal pattern of daily activity of Aedes caspius, Aedes detritus, Culex modestus, and Culex pipiens in the Po delta of northern Italy and significance for vector-borne disease risk assessment. J Vector Ecol. 2012;37:49–61. doi: 10.1111/j.1948-7134.2012.00199.x. [DOI] [PubMed] [Google Scholar]
- 33.Zeng X. Isolation of Japanese B encephalitis virus from Culex (Barraudius) modestus Ficalbi in Shenyang area (author’s transl) Zhonghua Yu Fang Yi Xue Za Zhi. 1980;14:209–210. [PubMed] [Google Scholar]
- 34.Ferraguti M, de la Puente JM, Muñoz J, et al. Avian Plasmodium in Culex and Ochlerotatus mosquitoes from Southern Spain: effects of season and host-feeding source on parasite dynamics. PLoS ONE. 2013;8:e66237. doi: 10.1371/journal.pone.0066237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Votýpka J, Seblová V, Rádrová J. Spread of the West Nile virus vector Culex modestus and the potential malaria vector Anopheles hyrcanus in central Europe. J Vector Ecol. 2008;33:269–277. doi: 10.3376/1081-1710-33.2.269. [DOI] [PubMed] [Google Scholar]
- 36.Cotar AI, Falcuta E, Prioteasa LF, et al. Transmission dynamics of the west nile virus in mosquito vector populations under the influence of weather factors in the Danube Delta. Romania EcoHealth. 2016;13:796–807. doi: 10.1007/s10393-016-1176-y. [DOI] [PubMed] [Google Scholar]
- 37.Ponçon N, Toty C, L'Ambert G, et al. Population dynamics of pest mosquitoes and potential malaria and West Nile virus vectors in relation to climatic factors and human activities in the Camargue. France Med Vet Entomol. 2007;21:350–357. doi: 10.1111/j.1365-2915.2007.00701.x. [DOI] [PubMed] [Google Scholar]
- 38.Sulesco T, Toderas L, Toderas I. A recent survey of the mosquito (Diptera: Culicidae) fauna and seasonal human biting activity in the city of Chisinau Moldova. J Eur Mosq Control Assoc. 2013;31:1–7. [Google Scholar]
- 39.Chippaux A, Rageau J, Mouchet J. [Hibernation of arbovirus Tahyna in Culex modestus Fic in France] C R Acad Hebd Seances. Acad Sci D. 1970;270:1648–1650. [PubMed] [Google Scholar]
- 40.Schaefer CH, Washino RK. Synthesis of energy for overwintering in natural populations of the mosquito Culex tarsalis. Comp Biochem Physiol. 1970;35:503–506. [Google Scholar]
- 41.Nasci RS, Savage HM, White DJ, et al. West Nile virus in overwintering Culex mosquitoes, New York City, 2000. Emerg Infect Dis. 2001;7:742. doi: 10.3201/eid0704.010426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Balenghien T, Vazeille M, Reiter P, et al. Evidence of laboratory vector competence of Culex modestus for West Nile virus. J Am Mosq Control Assoc. 2007;23:233–236. doi: 10.2987/8756-971X(2007)23[233:EOLVCO]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 43.Balenghien T, Vazeille M, Grandadam M, et al. Vector competence of some French Culex and Aedes mosquitoes for West Nile Virus. Vector-Borne Zoonotic Dis. 2008;8:589–595. doi: 10.1089/vbz.2007.0266. [DOI] [PubMed] [Google Scholar]
- 44.Jiang SF, Zhang YM, Guo XX, et al. Experimental studies on comparison of the potential vector competence of four species of Culex mosquitoes in China to transmit West Nile Virus. J Med Entomol. 2010;47:788–790. doi: 10.1603/me08292. [DOI] [PubMed] [Google Scholar]
- 45.Chaskopoulou A, Latham MD, Pereira RM, et al. Efficacy of aerial ultra-low volume applications of two novel water-based formulations of unsynergized pyrethroids against riceland mosquitoes in Greece. J Am Mosq Control Assoc. 2011;27:414–422. doi: 10.2987/11-6177.1. [DOI] [PubMed] [Google Scholar]
- 46.Muñoz J, Ruiz S, Soriguer R, et al. Feeding patterns of potential West Nile virus vectors in South-West Spain. PLoS ONE. 2012 doi: 10.1371/journal.pone.0039549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Trájer AJ. Which mosquitoes (Diptera: Culicidae) are candidates for DNA extraction in forensic practice? J Forensic Leg Med. 2018;1:183–191. doi: 10.1016/j.jflm.2018.07.002. [DOI] [PubMed] [Google Scholar]
- 48.Platonov AE, Fedorova MV, Karan LS, et al. Epidemiology of West Nile infection in Volgograd, Russia, in relation to climate change and mosquito (Diptera: Culicidae) bionomics. Parasitol Res. 2008;103:45–53. doi: 10.1007/s00436-008-1050-0. [DOI] [PubMed] [Google Scholar]
- 49.Marshall JF. Records of Culex (Barraudius) modestus Ficalbi (Diptera, Culicidae) obtained in the south of England. Nature. 1945;156:172–173. [Google Scholar]
- 50.Callot J, Van-Ty D. Localités françaises nouvelles pour Culex (Culex) theileri, Culex (Barraudius) modestus et Culex (Neoculex) impudicus. Ann Parasitol Hum Comparée. 2017;1942:142–150. [Google Scholar]
- 51.Brugman VA, England ME, Stoner J, et al. How often do mosquitoes bite humans in southern England? A standardised summer trial at four sites reveals spatial, temporal and site-related variation in biting rates. Parasit Vectors. 2017;10:420. doi: 10.1186/s13071-017-2360-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Wang ZM, Xing D, Wu ZM, et al. Biting activity and host attractancy of mosquitoes (Diptera: Culicidae) in Manzhouli. China J Med Entomol. 2012;49:1283–1288. doi: 10.1603/me11131. [DOI] [PubMed] [Google Scholar]
- 53.Guo XX, Zhang YM, Li CX, et al. Host-seeking behavior and arbovirus detection in mosquitoes of habahe county, Xinjiang Uigur Autonomous Region. China J Am Mosq Control Assoc. 2015;31:329–335. doi: 10.2987/moco-31-04-329-335.1. [DOI] [PubMed] [Google Scholar]
- 54.Bellini R, Veronesi R, Draghetti S, et al. Study on the flying height of Aedes caspius and Culex pipiens females in the Po delta area. Italy J Am Mosq Control Assoc. 1997;13:356–360. [PubMed] [Google Scholar]
- 55.Mohd Nordin NR, Arsad S, Hilmi Mahmud M, et al. Wolbachia in dengue control: a systematic review. Maced J Med Sci. 2022 doi: 10.3889/oamjms.2022.9014. [DOI] [Google Scholar]
- 56.Bergman A, Hesson JC. Wolbachia prevalence in the vector species Culex pipiens and Culex torrentium in a Sindbis virus-endemic region of Sweden. Parasit Vectors. 2021;14:428. doi: 10.1186/s13071-021-04937-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Ricci I, Cancrini G, Gabrielli S, et al. Searching for Wolbachia (Rickettsiales: Rickettsiaceae) in Mosquitoes (Diptera: Culicidae): large polymerase chain reaction survey and new identifications. J Med Entomol. 2002;39:562–567. doi: 10.1603/0022-2585-39.4.562. [DOI] [PubMed] [Google Scholar]
- 58.Shaikevich E, Bogacheva A, Ganushkina L. Dirofilaria and Wolbachia in mosquitoes (Diptera: Culicidae) in central European Russia and on the Black Sea coast. Parasite. 2019 doi: 10.1051/parasite/2019002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Soto A, De Coninck L, Devlies AS, et al. Belgian Culex pipiens pipiens are competent vectors for West Nile virus while Culex modestus are competent vectors for Usutu virus. PLoS Negl Trop Dis. 2023;17:e0011649. doi: 10.1371/journal.pntd.0011649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Ant TH, Mancini MV, McNamara CJ, et al. Wolbachia-Virus interactions and arbovirus control through population replacement in mosquitoes. Pathog Glob Health. 2023;117:245–258. doi: 10.1080/20477724.2022.2117939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Konstantinidis K, Dovrolis N, Kouvela A, et al. Defining virus-carrier networks that shape the composition of the mosquito core virome of a local ecosystem. Virus Evol. 2022;8:036. doi: 10.1093/ve/veac036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Khodzhaeva LF, Issi IV. A new genus of microsporidans Cristulospora gen n (Amblyospiridae) with 3 new species from blood-sucking mosquitoes in Uzbekistan. Parazitologiia. 1989;23:140–5. [PubMed] [Google Scholar]
- 63.Svobodova M, VOLF P, Votypka J. Trypanosomatids in ornithophilic bloodsucking Diptera. Med Vet Entomol. 2015;29:444–447. doi: 10.1111/mve.12130. [DOI] [PubMed] [Google Scholar]
- 64.Pridantseva EA, Lebedeva NI, Shcherban’ ZP, Kadyrova MK. An evaluation of the possibility of using Romanomermis iyengari Welch mermithids for mosquito control in Uzbekistan. Med Parazitol (Mosk). 1990;1:15–7. [PubMed] [Google Scholar]
- 65.Dzerzhinskiĭ VA, Dubitskiĭ AM, Deshevykh ND. Appearance of variants in the artificial infection of larvae of the blood-sucking mosquito, Culex modestus, infested with the entomopathogenic fungus, Coelomomyces iliensis. Parazitologiia. 1975;9:540–542. [PubMed] [Google Scholar]
- 66.Kumar Barik T. Molecular identification of mosquito vectors and their management. Berlin: Springer Nature; 2020. [Google Scholar]
- 67.Folmer O, Black M, Hoeh W, et al. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotechnol. 1994;3:294–299. [PubMed] [Google Scholar]
- 68.Weitzel T, Collado A, Jöst A, et al. Genetic differentiation of populations within the Culex pipiens complex and phylogeny of related species. J Am Mosq Control Assoc. 2009;25:6–17. doi: 10.2987/08-5699.1. [DOI] [PubMed] [Google Scholar]
- 69.Smith JL, Fonseca DM. Rapid assays for identification of members of the Culex (Culex) pipiens complex, their hybrids, and other sibling species (Diptera: Culicidae) Am J Trop Med Hyg. 2004;70:339–345. [PubMed] [Google Scholar]
- 70.Beier JC. Vector incrimination and entomological inoculation rates. Methods Mol Med. 2002;72:3–11. doi: 10.1385/1-59259-271-6:01. [DOI] [PubMed] [Google Scholar]
- 71.Macdonald G. The epidemiology and control of Malaria. London: Oxford University Press; 1957. p. 201. [Google Scholar]
- 72.Crivei LA, Moutailler S, Gonzalez G, et al. Detection of West Nile Virus Lineage 2 in Eastern Romania and first identification of Sindbis Virus RNA in mosquitoes analyzed using high-throughput microfluidic real-time PCR. Viruses. 2023 doi: 10.3390/v15010186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Šikutová S, Dočkal P, Straková P, et al. First record of mosquito-borne kyzylagach virus in central Europe. Viruses. 2020 doi: 10.3390/v12121445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Cao Y, Fu S, Tian Z, et al. Distribution of mosquitoes and mosquito-borne arboviruses in Inner Mongolia. China Vector-Borne Zoonotic Dis. 2011;11:1577–1581. doi: 10.1089/vbz.2010.0262. [DOI] [PubMed] [Google Scholar]
- 75.Hubálek Z, Rudolf I, Bakonyi T, et al. Mosquito (Diptera: Culicidae) surveillance for arboviruses in an area endemic for West Nile (Lineage Rabensburg) and Tahyna viruses in Central Europe. J Med Entomol. 2010;47:466–472. doi: 10.1603/me09219. [DOI] [PubMed] [Google Scholar]
- 76.Scheuch D, Schäfer M, Eiden M, et al. Detection of Usutu, Sindbis, and Batai Viruses in Mosquitoes (Diptera: Culicidae) collected in Germany, 2011–2016. Viruses. 2018;10:389. doi: 10.3390/v10070389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Danielová V, Hájková Z, Kolman JM, et al. Results of the virological examination of mosquitoes in southern Moravia in 1962–1964. Cesk Epidemiol Mikrobiol Imunol. 1966;15:178–184. [PubMed] [Google Scholar]
- 78.Liu R, Zhang G, Sun X, et al. Isolation and molecular characterization on Abbey Lake Orthobunyavirus (Bunyaviridae) in Xinjiang, China. Zhonghua Liu Xing Bing Xue Za Zhi Zhonghua Liuxingbingxue Zazhi. 2014;1:939–942. [PubMed] [Google Scholar]
- 79.Xia H, Liu R, Zhao L, et al. Characterization of Ebinur Lake Virus and Its human seroprevalence at the China-Kazakhstan Border. Front Microbiol. 2020 doi: 10.3389/fmicb.2019.03111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.L’vov DK, Al’khovskiĭ SV, Shchelkanov MI, et al. Genetic characterization of the Zaliv Terpeniya virus (ZTV, Bunyaviridae, Phlebovirus, Uukuniemi serogroup) strains isolated from the ticks Ixodes (Ceratixodes) uriae White, 1852, obligate parasites of the Alcidae birds, in high latitudes of Northern Eurasia and the mosquitoes Culex modestus Ficalbi, 1889, in subtropics Transcaucasus] Vopr Virusol. 2014;59:12–8. [PubMed] [Google Scholar]
- 81.Cao YX, Fu SH, Tian ZF, et al. Isolation and identification of Banna virus from mosquito for the first time in Inner Mongolia. Zhonghua Shi Yan He Lin Chuang Bing Du Xue Za Zhi. 2009;23:106–108. [PubMed] [Google Scholar]
- 82.O’Leary DR, Marfin AA, Montgomery SP, et al. The epidemic of West Nile Virus in the United States, 2002. Vector-Borne Zoonotic Dis. 2004;4:61–70. doi: 10.1089/153036604773083004. [DOI] [PubMed] [Google Scholar]
- 83.Heidecke J, Schettini AL, Rocklöv J. West Nile virus eco-epidemiology and climate change. PLOS Clim. 2023;2:e0000129. [Google Scholar]
- 84.Tran A, L’Ambert G, Balança G, et al. An integrative eco-epidemiological analysis of West Nile Virus transmission. EcoHealth. 2017;14:474–489. doi: 10.1007/s10393-017-1249-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Mughini-Gras L, Mulatti P, Severini F, et al. Ecological niche modelling of potential west nile virus vector mosquito species and their geographical association with equine epizootics in Italy. EcoHealth. 2014;11:120–132. doi: 10.1007/s10393-013-0878-7. [DOI] [PubMed] [Google Scholar]
- 86.Vignjević G, Vrućina I, Sestak I, et al. Equine seroprevalence rates as an additional indicator for a more accurate risk assessment of the West Nile virus transmission. Coll Antropol. 2013;37:949–956. [PubMed] [Google Scholar]
- 87.Fyodorova MV, Savage HM, Lopatina JV, et al. Evaluation of potential West Nile Virus vectors in volgograd Region, Russia, 2003 (Diptera: Culicidae): species composition, bloodmeal host utilization, and virus infection rates of mosquitoes. J Med Entomol. 2006;43:552–563. doi: 10.1603/0022-2585(2006)43[552:eopwnv]2.0.co;2. [DOI] [PubMed] [Google Scholar]
- 88.Rijks JM, Kik ML, Slaterus R, et al. Widespread Usutu virus outbreak in birds in the Netherlands, 2016. Eurosurveillance. 2016;21:30391. doi: 10.2807/1560-7917.ES.2016.21.45.30391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Hönig V, Palus M, Kaspar T, et al. Multiple lineages of Usutu virus (Flaviviridae, Flavivirus) in blackbirds (Turdus merula) and mosquitoes (Culex pipiens, Cx modestus) in the Czech Republic (2016–2019) Microorganisms. 2019;7:568. doi: 10.3390/microorganisms7110568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Rossi L, Pollono F, Meneguz P, et al. Four species of mosquito as possible vectors for Dirofilaria immitis piedmont rice-fields. Parassitologia. 1999;41:537–542. [PubMed] [Google Scholar]
- 91.Pollono F, Rossi L, Cancrini G. Research on Culicidae attracted to dog bait in piedmont. Parassitologia. 1998;40:439–445. [PubMed] [Google Scholar]
- 92.Ionicǎ AM, Zittra C, Wimmer V, et al. Mosquitoes in the danube delta: searching for vectors of filarioid helminths and avian malaria. Parasit Vectors. 2017 doi: 10.1186/s13071-017-2264-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Merdić E, Jeličić Ž, Krčmar S, et al. Efficacy of mosquito attractants in various habitats of a floodplain. Biologia. 2010;65:545–551. [Google Scholar]
- 94.Freitas FB, Novo MT, Esteves A, et al. Species composition and WNV screening of mosquitoes from lagoons in a wetland area of the Algarve Portugal. Front Physiol. 2012 doi: 10.3389/fphys.2011.00122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.L’Ambert G, Ferré JB, Schaffner F, et al. Comparison of different trapping methods for surveillance of mosquito vectors of West Nile virus in Rhône Delta. France J Vector Ecol. 2012;37:269–275. doi: 10.1111/j.1948-7134.2012.00227.x. [DOI] [PubMed] [Google Scholar]
- 96.Soltész Z, Erdélyi K, Bakonyi T, et al. West Nile virus host-vector-pathogen interactions in a colonial raptor. Parasit Vectors. 2017;10:10. doi: 10.1186/s13071-017-2394-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Belevich O, Yurchenko Y, Alekseev A, et al. Toxic effects of fine plant powder impregnated with avermectins on Mosquito Larvae and nontarget aquatic invertebrates. J Med Entomol. 2021;58:773–780. doi: 10.1093/jme/tjaa227. [DOI] [PubMed] [Google Scholar]
- 98.Wang L, Soto A, Remue L, et al. First report of mutations associated with pyrethroid (L1014F) and organophosphate (G119S) resistance in Belgian Culex (Diptera: Culicidae) mosquitoes. J Med Entomol. 2022;59:2072–2079. doi: 10.1093/jme/tjac138. [DOI] [PubMed] [Google Scholar]
- 99.Atyame CM, Alout H, Mousson L, et al. Insecticide resistance genes affect Culex quinquefasciatus vector competence for West Nile virus. Proc R Soc B Biol Sci. 2019;286:20182273. doi: 10.1098/rspb.2018.2273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Alcaide M, Rico C, Ruiz S, et al. Disentangling vector-borne transmission networks a universal DNA barcoding method to identify vertebrate hosts from arthropod bloodmeals. PLoS ONE. 2009 doi: 10.1371/journal.pone.0007092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Martínez-De La Puente J, Ruiz S, Soriguer R, et al. Effect of blood meal digestion and DNA extraction protocol on the success of blood meal source determination in the malaria vector Anopheles atroparvus. Malar J. 2013;12:109. doi: 10.1186/1475-2875-12-109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Rudolf I, Blažejová H, Šebesta O, et al. West Nile virus (lineage 2) in mosquitoes in southern Moravia—awaiting the first autochthonous human cases. Epidemiol Mikrobiol Imunol. 2018;67:44–46. [PubMed] [Google Scholar]
- 103.Mancini G, Montarsi F, Calzolari M, et al. Mosquito species involved in the circulation of West Nile and Usutu viruses in Italy. Vet Ital. 2017;53:97–110. doi: 10.12834/VetIt.114.933.4764.2. [DOI] [PubMed] [Google Scholar]
- 104.Nurmakhanov T, Sansyzbaev Y, Atshabar B, et al. Phylogenetic characteristics of West Nile virus isolated from Culex modestus mosquitoes in West Kazakhstan. Front Public Health. 2021;12:8. doi: 10.3389/fpubh.2020.575187. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1: Fig. S1. Comparison of A Culex pipiens pipiens and B Culex modestus. The black arrows indicate the unique characteristics for morphological identification of Culex modestus.
Additional file 2: Table S1. Studies with reports of Culex modestus by country.
Data Availability Statement
All data generated or analyzed during this study are included in this published article and its supplementary information files.