

Abstract
Antimicrobial resistance is a major threat to human health globally. Antrodia camphorata was grown in a malt/yeast extract broth liquid medium for 15 days. Then, 4-L fermentation broth was harvested, yielding 7.13 g of the ethyl acetate extract. By tracing the antimicrobial activity, 12.22 mg of the antimicrobial compound was isolated. The structure of 5-methyl-benzo [1,3]-dioxole-4,7-diol (MBBD) was elucidated using NMR and MS data analyses. The antibacterial activity of MBBD was detected through the microbroth dilution method. MBBD exhibited broad-spectrum antibacterial activity. The minimum inhibitory concentration (MIC) range of MBBD for drug-resistant pathogenic bacteria was 64–256 μg/mL, with the lowest MIC observed for Acinetobacter baumannii (64 μg/mL), followed by Pseudomonas aeruginosa (MIC = 128 μg/mL). Klebsiella pneumoniae, Staphylococcus aureus, Enterococcus faecalis, and Escherichia coli were also sensitive, with an MIC of 256 μg/mL. The MIC range of MBBD against 10 foodborne pathogens was 12.5–100 μg/mL. Based on the results of this study, MBBD exhibits broad-spectrum antibacterial activity, particularly demonstrating excellent inhibitory effects against A. baumannii. MBBD will be good candidates for new antimicrobial drugs.
Introduction
Globally, the spread of antibiotic-resistant bacteria is a substantial threat causing morbidity and mortality [1, 2]. Drug-resistant bacteria directly caused 1.27 million deaths worldwide in 2019. As many as 4.95 million deaths were caused by drug-resistant bacterial infections, making drug resistance the third leading cause of human death [3]. ESKAPE (Enterococcus faecalis, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Escherichia coli) are the most common opportunistic pathogens causing nosocomial infections [4]. ESKAPE-induced infections are often associated with high morbidity, mortality, and treatment costs. These pathogens are highly mutagenic, rapidly transmissible, and exhibit severe drug resistance [5, 6]. Types and mechanisms of bacterial resistance are complex and belong to four main categories: impaired drug penetration, altered drug targets, drug inactivation, and active efflux. Notably, the majority of conventional antibiotics target specific bacterial surfaces or structures that can contribute to resistance development [7, 8]. The rate of antibiotic development has not matched with the increasing rate of bacterial resistance. Thus, new antibacterial drugs need to be urgently discovered and developed for tackling the problem of bacterial resistance. Fungi produce a diverse array of secondary metabolites, such as antimicrobial peptides, polyketides, phenolics, and terpenoids. These metabolites exhibit various antibacterial activities. For instance, chaetochromin A, a compound isolated from the secondary metabolite of Chaetomium gracile, exhibits significant antibacterial activity against E. coli and S. aureus [9]. Other researchers obtained Bionectin D from the ethyl acetate extract of the endophytic fungus Bionectria sp. Y1085, which was isolated from the plant Huperzia serrata. Similarly, Bionectin D exhibited significant antibacterial activity against both E. coli and S. aureus [10]. The new endophytic fungus, Curvularia sp. T12, was isolated from the medicinal plant Rauwolfia macrophylla. Large-scale fermentation and subsequent extraction of the crude extract yielded two compounds, 2’-deoxyribolactone and hexylitaconic acid. These compounds exhibited significant inhibitory effects against E. coli, P. agarici, and S. warneri [11]. Additionally, previous reports have indicated that dothideomycetide A, a compound isolated from the endophytic fungus Dothideomycete sp. of a Thai medicinal plant Tiliacora triandra, exhibits antibacterial activity against methicillin-resistant S. aureus strains [12]. However, it is worth noting that the cultivation process for fungi is concise and uncomplicated, making it economically feasible. This process also demonstrates a notable level of versatility, as it can be finely tuned through the addition of precursors, elicitors, specialized enzymes, and modifiers, thereby significantly enhancing the production of bioactive compounds [13]. The utilization of fungal metabolites for the management of bacterial infections and addressing antibacterial resistance holds promising potential.
Antrodia camphorata is a unique, valuable, and endemic mushroom species containing various active constituents, including terpenoids, maleic and succinic acid derivatives, benzenoids, benzoquinone derivatives, lignans, and polysaccharides [14, 15]. These secondary metabolites exhibit varied biological activities, such as anticancer, antidiabetic, immunomodulatory, anti-inflammatory, antiviral, antiallergic, and antioxidant activities [16–18]. Furthermore, A. camphorata has been reported to possess antibacterial bioactive. Methyl antcinate and antcins, which are extracted from A. camphorata fruiting bodies, have displayed potent inhibitory action against Helicobacter pylori, the bacterium causing gastritis [19]. Both ethyl acetate and chloroform extracts of A. camphorata significantly inhibited oral bacteria such as Streptococcus mutants and Porphyromonas gingivalis [20]. However, reports on the antibacterial activity of A. camphorata extracts against drug-resistant bacteria and foodborne pathogens are lacking. Therefore, this study investigated the antimicrobial activity of A. camphorata fermentation broth and isolated, purified, and identified the antimicrobial compounds present in the broth by using an antimicrobial activity tracking method.
Materials and methods
Fungal culture and growth conditions
The A. camphorata strain used in this experiment was provided by the Yunnan Academy of Forestry and Grassland Science (Kunming, Yunnan, China). The fungal species were identified using ITS, β-tubulin gene fragments, and elongation factor 1α gene fragments, with the strain designated as YAFAC008 [21]. The A. camphorata strain was inoculated in PDA and cultivated in a constant temperature incubator at 26°C with 60% humidity for 12 days, followed by storage in a refrigerator at 4°C. Subsequently, 100 mL of malt/yeast extract medium was added to each of 40 triangular flasks with a volume of 250 mL, and they were autoclaved for sterilization. After cooling to room temperature, precise 0.5 cm2 of A. camphorata mycelia were excised from the mycelium’s edge and placed into each triangular flask under aseptic conditions. The flasks were then placed in a shaker and incubated at 28°C with agitation at 150 r/min for 15 days (S1 Fig).
Pathogenic bacteria and antibiotics used in antimicrobial experiments
The pathogenic bacteria used in the experiment included: drug-resistant pathogenic bacteria (A. baumannii, P. aeruginosa, K. pneumoniae, S. aureus, E. faecalis, and E. coli) that were purchased from the Guangdong Bacterial Resistance Monitoring and Quality Control Centre. The foodborne pathogens included: Bacillus cereus (BC), B. lentus (BL), B. pumilus (BP), B. subtilis (BS), Streptococcus agalactiae (SA), Shigella flexneri (SF), Micrococcus luteus (ML), Vibrio parahaemolyticus (VB), Staphylococcus haemolyticus (SH), and Salmonella paratyphi B (SP) were purchased from Henan Industrial Microbial Strain Engineering Technology Research Centre (S2 Table). The disease-causing bacteria were inoculated in Luria–Bertani liquid medium and incubated at 37°C for 24 h. The cultured bacteria were coated with sterile saline in a 10-fold gradient dilution for colony counting. Then, each bacterial solution was diluted with saline to a bacterial concentration of 1 × 106 Colony-Forming Units per Milliliter (CFU/mL) according to the colony counting results.
The antibiotics, including chloramphenicol, gentamycin, ampicillin, streptomycin, tetracycline, and kanamycin, were purchased from Kunming Shuoyang Technology Company. Each antibiotic was weighed around 50.00 mg, placed in a centrifuge tube, and dissolved by adding 1.0 mL of Dimethyl Sulfoxide (DMSO) to ensure that the concentration of each antibiotic was 50 mg/mL.
Preparation of A. camphorata extracts
After the incubation period, A. camphorata fermentation broth was filtered to separate the mycelium, and the pH of the broth was adjusted to 1–2 (S2 Fig). Ethyl acetate was added at a 1:1 volume ratio, shaken and mixed, sonicated for 60 min, and let stand for 8 hours. The lower layer of water was removed, and the top layer of ethyl acetate was dried using a rotary evaporator and weighed.
Antibacterial activity of A. camphorata extracts
The paper diffusion method was used to test antibacterial activity [22]. Using a sterile cotton swab, 250 μL of the bacterial suspension was applied to a 4-mm solid agar-containing plate (diameter: 90 mm). Sterile filter paper discs (diameter: 6 mm) were affixed to the bacteria-containing agar plates, and 10 μL of the A. camphorata extract (50 mg/mL dissolved with DMSO) was added dropwise. The diameter of the clear inhibition zone (mm) around the round paper slice of the drop-spiked extract was measured after 24 h of incubation at 37°C. In all experiments, a negative control (10 μL DMSO) was included; the positive control used the aforementioned six standard antibiotics (50 mg/mL).
Isolation and identification of compounds
In the liquid fermentation culture of 4 L of A. camphorata, the ethyl acetate extract was prepared at the end of the culture period. Equal masses of the 100–200-mesh silica gel were stirred with the extract to dry. The extract weighed 10 times the weight of the 200–300-mesh silica gel, filled in a normal phase silica gel column (length: 60 cm, diameter: 6 cm), and eluted with a gradient of petroleum ether and ethyl acetate, with 1 L of each gradient. The elute was concentrated to obtain a normal-phase silica gel column separation fraction, and this fraction was assayed separately for antibacterial activity. The fraction with antibacterial activity was used as the target fraction of the separated compounds. The target components were subjected to thin-layer chromatography, and similar components were combined. The monomeric compounds were carried out on the Agilent 1260 High-Performance Liquid Chromatography (HPLC) system (Agilent Technologies, Wilmington, Germany) equipped with an Agilent four-unit pump (CA, Germany), a 7125 injector (CA, Germany) and a G1314A UV detector. The analytical column used was a Zorbax Extend-C18 column (4.6 mm × 250 mm, 5 μm, Agilent, Germany). The absorbance was measured at 254 nm. Elution was carried out at a flow rate of 200 μL/min at 30°C. The mobile phase consisted of A (water) and B (100% acetonitrile): 0–90 min, 100%–0% A, and 0%–100% B. The pure monomeric compounds were subjected to nuclear magnetic resonance (NMR) and mass spectrometry (MS) to identify their structures. 1HNMR and 13C-NMR spectra were recorded on Bruker AVANCE III 400 MHz (Bruker BioSpin GmbH, Rheinstetten, Germany) instruments, using tetramethylsilane (TMS) as an internal standard: chemical shifts (δ) are given in ppm, coupling constants (J) in Hz, and the solvent signals were used as references (CDCl3: δC = 77.2 ppm, residual CHCl3 in CDCl3: δH = 7.26 ppm; CD3OD: δC = 49.0 ppm, residual CH3OH in CD3OD: δH = 4.78 ppm). MS data were obtained in the ESI mode on API Qstar Pulsar instrument.
Determination of Minimum Inhibitory Concentration (MIC) of compound MBBD
The microbroth dilution method was used to determine the MIC of the compound MBBD against the tested bacteria [23]. MBBD dissolved in DMSO was diluted with sterile saline in a twofold gradient. Then, 100 μL of MBBD solution at different dilutions (100, 50, 25, and 12.5 μg/mL) and each suspension of foodborne pathogens were pipetted into small wells of 96-well plates to determine the MIC of MBBD against the foodborne pathogens. Later, 100 μL of MBBD solution at different dilutions (1024, 512, 256, 128, 64, 32, 16, 8, 4, 2, and 1 μg/mL) and the suspension of each drug-resistant pathogenic bacteria were pipetted into the wells of a 96-well plate to determine the MIC of MBBD against the drug-resistant pathogenic bacteria. The wells spiked with 100 μL DMSO were used as the solvent control group, the wells spiked with 100 μL sterile saline were used as the blank control group, and six replicates were established for each group. The 96-well plates were placed in a shaker and incubated at 37°C and 120 r/min for 24 h. At 600 nm, the bacterial growth levels were measured using an enzyme marker. MIC was defined as the lowest dilution of MBBD with an absorbance of ≤0.1.
Cytotoxicity assay of MBBD
The cytotoxic activity of MBBD was tested using the MTS method [23]. The cell lines included were leukemia HL-60, lung cancer A-S49, liver cancer SMMC-7721, breast cancer MCF-7, colon cancer SW480, and human normal lung epithelial cells BEAS-2B. The cells were grown at 37°C in 5% CO2 in Dulbecco’s Modified Eagle Medium supplemented with 4.5 g/L glucose, 2 mM L-glutamine, 1% penicillin-streptomycin, and 10% fetal bovine serum. The cells were seeded at a density of 1.5 × 104 cells/well in 96-well culture plates overnight and then treated with MBBD at 40 μM. After 24 h of incubation, a culture medium containing MTS (1 mg/mL) was added to each well, and the plate was incubated for another 2 h. Consecutively, the medium was removed, and DMSO was added to extract the MTS formazan. The absorbance of each well was measured using an enzyme-linked immunosorbent assay reader at 492 nm. Three blank replicate wells (a mixture of 20 μL of MTS solution and 100 μL of culture solution) and two positive controls of cisplatin DDP and taxol were used for each experiment.
Statistical analysis
The means were compared using the SPSS for Windows statistical software package. The experiment was repeated three times, with the results presented as the mean ± standard deviation of three independent experiments.
Results
Comparing the antimicrobial activity of A. camphorata extracts with the positive control
Drug-resistant pathogenic bacteria exhibited resistance to at least one of the six tested antibiotics. All the tested bacteria demonstrated resistance to ampicillin without exhibiting any discernible inhibition zone. Multidrug resistance was defined as a lack of susceptibility to at least one agent in three or more categories of antibiotics. A. baumannii and P. aeruginosa exhibited resistance to multiple antibiotics, with resistance rates of 6/6 and 5/6, respectively. In contrast, E. coli demonstrated resistance to 2/6 antibiotics, while S. aureus, E. faecalis, and K. pneumoniae each displayed a resistance rate of 1/6. None of the six antibiotics at a concentration of 50.00 mg/mL showed inhibitory effects on A. baumannii (Table 1 and S3 Fig). A. camphorata extracts exhibited remarkably potent inhibition against A. baumannii, maintaining its effectiveness even when the mass concentration was reduced to 12.50 mg/mL (Fig 1).
Table 1. Comparing the antimicrobial activity of A. camphorata extracts with the positive control.
| Species | Chloramphenicol | Gentamycin | Ampicillin | Streptomycin | Tetracycline | Kanamycin | A. camphorata extract | Antibiotic Rate |
|---|---|---|---|---|---|---|---|---|
| E. faecalis | 27.0±0.4 | 10.5±0.6 | ND | 18±0.7 | 12.5±0.7 | 15.0±0.8 | 10.3±0.5 | 1/6 |
| S. aureus | 24.5±0.3 | 12.0±0.5 | ND | 17.2±0.6 | 13.5±0.3 | 12.0±0.4 | 10.3±0.5 | 1/6 |
| P. aeruginosa | ND | ND | ND | ND | 8.5±0.6 | ND | 10.4±0.3 | 5/6 |
| A. baumannii | ND | ND | ND | ND | ND | ND | 13.5±0.4* | 6/6 |
| K. pneumoniae | 28±0.2 | 13.0±0.3 | ND | 17.2±0.5 | 13.1±0.3 | 18.2±0.6 | 10.1±0.1 | 1/6 |
| E. coli | ND | 14.2±0.3 | ND | 16.5±0.2 | 14.5±0.4 | 11.8±0.8 | 13.9±0.4 | 2/6 |
ND: not detected; The mass concentration of each antibiotic and the extract of A. camphorata: 50.00 mg/mL; Values are the mean of n = 3 experiments, Inhibition zone diameter, measured in millimeters.
Fig 1. Antibacterial activity of A. camphorata extracts compared with 5 antibiotics.

Gen. represents Gentamicin, Amp. represents Ampicillin, Str. represents Streptomycin, Tet. represents Tetracycline, Kan. represents Kanamycin, BD represents A. camphorata extracts (50.00 mg/mL), BD-1 represents A. camphorata extracts (25.00 mg/mL), BD-2 represents A. camphorata extracts (12.50 mg/mL).
Antibacterial activity of A. camphorata extracts against foodborne pathogens
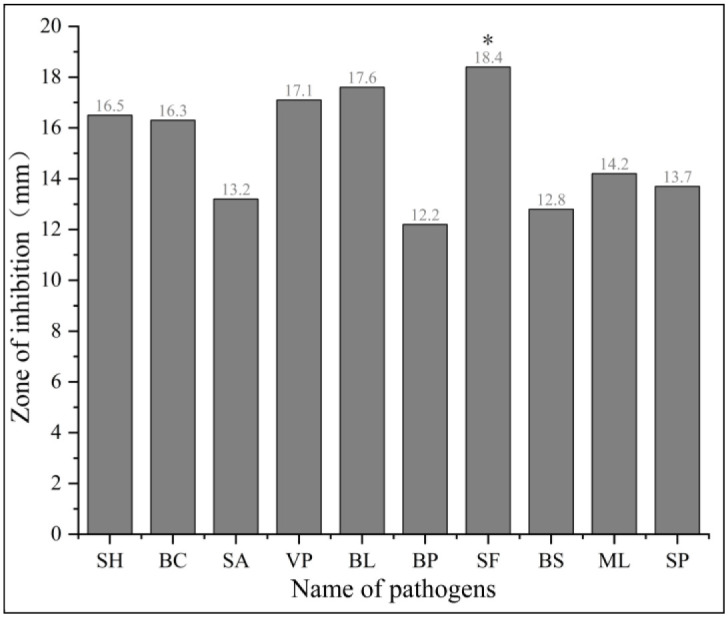
Using the paper diffusion method, the antibacterial activity of the ethyl acetate extract of A. camphorata against 10 foodborne pathogens was evaluated. The extract showed significant inhibitory effect against all 10 tested bacteria at 50 mg/mL concentration, with the highest inhibitory effect observed against Shigella flexneri, which had an inhibition zone diameter of 18.3 mm. However, the extract’s effect on Bacillus pumilus was less potent, and an inhibition zone diameter of 12.3 mm was observed. The inhibitory activity of the remaining eight bacteria was better, with the inhibition zone ranging from 12.8 to 17.6 mm (Fig 2). These findings indicate that liquid fermentation of A. camphorata can generate a diverse array of active antibacterial substances with broad-spectrum activity.
Fig 2. Antibacterial activity of A. camphorata extracts against foodborne pathogens.
Isolation and identification of the antibacterial compound MBBD
The fermentation broth of A. camphorata was subjected to extraction by using ethyl acetate and concentrated using a rotary evaporator to yield 7.13 g of the extract from 4-L broth. The extract was fractionated through normal-phase silica gel column chromatography, each fraction was tested for antibacterial activity. When eluted with a mixture of petroleum ether and ethyl acetate in a 100:7 ratio, a white crystalline concentrate with antibacterial activity was obtained. The fraction containing the crystals was further purified through recrystallization. Preparative HPLC separation (95:5, methanol-water) led to the isolation of compounds AC-1 (12.22 mg, retention time = 1.526 min) and AC-2 (4.2 mg, retention time = 1.632 min). Antibacterial assays revealed that compound AC-1 had significant inhibitory activity against the tested bacteria, whereas compound AC-2 did not exhibit any activity (Table 2). Compound AC-1 was identified as MBBD by using NMR and MS methods, and its structural formula was determined to be 5-methyl-1,3-benzodioxole-4,7-diol (S4–S8 Figs).
Table 2. Separation and chemical analysis of A. camphorata extracts.
| Compound | Retention time(min) | Peak area | Peak height | Peak width | Symmetry factor | Peak area ratio (%) |
|---|---|---|---|---|---|---|
| AC-1 | 1.526 | 2056.383 | 646.470 | 0.0510 | 1.041 | 89.724 |
| AC-2 | 1.632 | 235.509 | 71.345 | 0.0505 | 0.896 | 10.276 |
MIC of MBBD against pathogenic bacteria
The compound MBBD has impressive antibacterial activity against various bacteria and well against both resistant pathogenic bacteria and foodborne pathogens. The microbroth dilution method is a standard technique for determining the MIC of an antibacterial compound against a pathogenic microbe. In this study, the MIC values of MBBD against foodborne pathogens ranged from 12.5 to 100 μg/mL. The lowest MIC value (12.5 μg/mL) was observed against S. haemolyticus and V. parahaemolyticus, followed by B. cereus (MIC = 25 μg/mL), B. pumilus, B. subtilis, and Streptococcus agalactiae (MIC = 50 μg/mL). However, the inhibition of B. lentus, Salmonella paratyphi B, Shigella flexneri, and M. luteus was relatively weak, with an MIC of 100 μg/mL (Fig 3). Moreover, the inhibitory effect of MBBD against drug-resistant pathogenic bacteria was also significant, with the lowest MIC value (64 μg/mL) observed against A. baumannii, followed by P. aeruginosa (MIC = 128 μg/mL). The MIC values of MBBD against K. pneumoniae, S. aureus, E. faecalis, and E. coli were 256 μg/mL (Fig 4). The consistent results obtained in triplicate are reassuring and suggest the robustness of the experiment.
Fig 3. MIC of compound MBBD against foodborne pathogens.

Values are the mean of n = 3 experiments.
Fig 4. MIC of compound MBBD against resistant pathogenic bacteria.

Data are reported as means ± SD of triplicate wells. Values are the mean of n = 3 experiments.
Cytotoxicity of MBBD
MBBD was insensitive to five types of human cancer cells and exhibited no cytotoxicity against human normal lung epithelial cells BEAS-2B (S9 Fig). The MTS assay showed that the cell inhibition rates of MBBD (40 μM) against HL-60, A-S49, SMMC-7721, MCF-7, SW480, and BEAS-2B were all <20% (Fig 5). This suggests that MBBD can potentially serve as a therapeutic agent that selectively targets pathogenic bacteria without affecting healthy human cells. Furthermore, the positive controls cisplatin DDP and taxol significantly inhibited the tested human cancer cells (Fig 6).
Fig 5. Cell inhibition rate of compound MBBD (40μM).

Fig 6. Cell inhibition rate of positive control group DDP and taxol.

Discussion
A. baumannii is a pathogenic bacterium that commonly causes infections in clinical settings, and its resistance to antibiotics has been increasing due to acquired resistance and antibiotic misuse. Owing to the emergence of multidrug-resistant A. baumannii (MDRAB), extensively drug-resistant A. baumannii, and even pan-drug-resistant A. baumannii, the number of antibiotics available for clinical treatment is limited, which has thus created an urgent need for novel antibiotics [24]. Polymyxin B is currently the drug of choice for treating MDRAB, but its use is limited because of nephrotoxicity, and drug resistance has been reported recently [25]. Cefiderocol, a new iron-carrying cephalosporin, is in phase III clinical trials and has shown robust activity against MDRAB. However, cefiderocol, when used alone, may result in the development of drug-resistant bacteria and may have some side effects, such as diarrhea, rash, blood in urine, a positive urine test for red blood cells, and an increased plasma white blood cell count [26–28].
Research into the development of new antibacterial drugs from fungi is gaining momentum because of the abundant resources available, lower adverse effects, and lower likelihood of drug resistance. Studies have revealed that altenusin, a compound isolated from the endophytic fungus Alternaria tenuissima PC-005 of Polygonum cephalicum, exhibited good antibacterial activity against A. baumannii with a MIC value of 125 μg/mL [29]. Similarly, researchers isolated an active compound, Flindersine (2,6-dihydro-2,2-dimethyl-5H-pyrano [3,2-c] quinoline-5-one-9cl), from the ethyl acetate extract of Toddalia asiatica. This compound exhibited MIC values of 125 μg/ml against A. baumannii [30]. Additionally, a newly discovered naturally-occurring hydroquinone, α,2,5-trihydroxyacetophenone, was extracted from the marine-derived fungus Pseudopithomyces maydicus PSU-AMF350. This compound demonstrated antibacterial efficacy against A. baumannii, with an MIC value of 200 μg/mL [31]. In this study, MBBD, isolated and purified from the A. camphorata fermentation broth, exhibited a significant inhibitory effect on A. baumannii with a MIC value of 64 μg/mL. Additionally, it exhibited a stronger antibacterial effect than altenusin, Flindersine and α2,5-trihydroxyacetophenone. Notably, the potential use of MBBD as an antibiotic for A. baumannii treatment is promising and could offer a new approach to combat this challenging pathogen.
MBBD belongs to the benzodioxole class and was first isolated from A. camphorata fermentation broth by Taiwanese researchers in 2011. It exhibited good antioxidant activity [32]. In the same year, researchers successfully isolated and purified another benzodioxole-like compound known as SY1 (4,7-dimethoxy-5-methyl-1,3-benzodioxole) from A. camphorata substrates. SY1 exhibited anticancer activity [33]. Benzodioxole structures are less common in fungi, but they are more common in plants. For example, apiol, sanguinarine (SAG), chelirubine, coptisine, berberine, and podophyllotoxin (Fig 7) are known to contain benzodioxole analogs with antibacterial properties. Berberine, when combined with other antibiotics, effectively killed MRSA by inhibiting phenol-soluble regulatory protein aggregation into amyloid fibrils, thereby disrupting MRSA biofilms [34]. SAG, which is found in the Papaveraceae, Berberidaceae, and Ranunculaceae families, possesses antioxidant, anti-inflammatory, and antiproliferative properties against various malignancies. In 2020, a study reported a significant inhibitory effect of SAG against Providencia rettgeri, with a MIC value of 7.8 μg/mL [35]. Chelerythrine (CHE) is a natural benzophenanthridine alkaloid derived from traditional medicinal plants of the Papaveraceae family and T. asiatica [36]. In prior research, CHE isolated from T. asiatica roots exhibited antibacterial activity against β-lactamase-resistant Staphylococcus by disrupting the bacterial cell wall, cell membrane, and inhibiting protein biosynthesis [37]. Subsequent investigations conducted by other scientists demonstrated that CHE also hindered the growth of carbapenem-resistant Serratia marcescens (MIC = 125 μg/mL /mL), leading to disruptions in cell membrane integrity and significant alterations in cell morphology [38]. In our study, MBBD displayed significant inhibitory action against both ESKAPE pathogens (MIC = 64–256 μg/mL) and foodborne pathogens (MIC = 12.5–100 μg/mL). These studies indicate that benzodioxole analogs have a wide range of biological activity. In summary, MBBD containing the benzodioxole structure can be considered an effective approach for the prevention or inhibition of A. baumannii. Furthermore, it exhibits no toxicity toward human normal lung epithelial cells, potentially making it a comparatively safer option. However, achieving large-scale production of MBBD poses several significant challenges that require careful consideration. First and foremost, identifying a culture medium capable of promptly and efficiently inducing MBBD production in A. camphorata is of paramount importance. Additionally, gaining a comprehensive understanding of the entire biosynthetic pathway, including all relevant enzymes and genes, necessitates the application of ’omics’ methodologies, which encompass genomics, transcriptomics, proteomics, and metabolomics. This comprehensive elucidation is poised to facilitate precise regulatory and manipulative strategies for optimizing the biosynthetic process, ultimately resulting in enhanced production efficiency [39]. Significantly, prior investigations have unveiled distinct benzodioxole derivatives demonstrating noteworthy anticancer and antibacterial properties, hinting at a conceivable correlation between these effects and the compounds’ DNA-binding capabilities [40]. As our research progresses, we are dedicated to embarking on a comprehensive exploration of the potential mechanisms underlying MBBD’s antibacterial activity.
Fig 7. Chemical structure of MBBD and its analogs.

Nowadays, the fruiting bodies of A. camphorata are in great demand and very expensive due to host specificity, rarity in nature, and the difficulty of artificial cultivation. Malic acid and succinic acid derivatives are the primary bioactive compounds in the liquid fermentation of A. camphorata. Among these compounds, antrodin A exhibits excellent inhibitory activity against the protease of the hepatitis C virus, while antrodin B and C demonstrate significant cytotoxicity against lung cancer cell lines [41]. Liquid fermentation allows for better control of the cultivation environment. Some researchers have found that supplementing the culture medium with p-hydroxybenzoic acid, eugenol, and coenzyme Q0 during the liquid fermentation of A. camphorata can promote the production of antroquinonol, a compound with remarkable anticancer properties [42]. Additionally, the addition of petroleum ether extracts from camphor trees to the culture medium has been shown to significantly increase the total triterpene yield of A. camphorata. Therefore, deep fermentation technology stands out due to its advantages, including short cycles, high efficiency, low cost, and ease of scalable production [43–45]. In this study, A. camphorata was cultured using liquid fermentation because it is less time-consuming and cost-effective. MBBD (122.8 mg/4 L) with broad-spectrum antibacterial activity was isolated and purified from the A. camphorata fermentation broth. Therefore, liquid fermentation of A. camphorata has the potential to yield numerous antimicrobial compounds. In conclusion, MBBD can serve as a reference for developing drugs to combat A. baumannii and can play a crucial role in the application of liquid fermentation of A. camphorata and the development of new antibacterial drugs.
Conclusion
In this study, compound MBBD was isolated and identified from the fermentation broth of A. camphorata. MBBD exhibited inhibitory activity against drug-resistant bacterial strains, including A. baumannii, P. aeruginosa, K. pneumoniae, S. aureus, E. faecalis, E. coli, as well as 10 foodborne pathogens. In particular, it demonstrated a significant inhibitory action against highly resistant A. baumannii. Additionally, MBBD was non-toxic to the normal human lung epithelial cells BEAS-2B. MBBD exhibits broad-spectrum antibacterial activity, particularly demonstrating excellent inhibitory effects against A. baumannii. MBBD may be good candidates for new antimicrobial drugs.
Supporting information
(DOCX)
(DOCX)
Gen. represents Gentamicin, Amp. represents Ampicillin, Str. represents Streptomycin, Tet. represents Tetracycline, Kan. represents Kanamycin, BD represents A. camphorata extracts (50.00 mg/mL), BD-1 represents A. camphorata extracts (25.00 mg/mL), BD-2 represents A. camphorata extracts (12.50 mg/mL). The figures, from left to right and top to bottom, represent A. baumannii, S. aureus, P. aeruginosa, E. coli, K. pneumoniae, and E. faecalis.
(DOCX)
(DOCX)
(DOCX)
(DOCX)
(DOCX)
(DOCX)
MBBD was not cytotoxic to human normal lung epithelial cells BEAS-2B.
(DOCX)
Data Availability
All relevant data are within the manuscript and its Supporting information files.
Funding Statement
Wang Yi received the award of General Project of the Basic Research Program in Yunnan Province (202101AT070218), the National Natural Science Foundation of China (31860177), the Reserve Talents for Young and Middle-aged Academic and Technical Leaders of the Yunnan Province (202205AC160044), and Yunnan Key Laboratory for Fungal Diversity and Green Development (E03A311261-3). Zhen Yuan received the award of General Project of the Basic Research Program in Yunnan Province(202101AT070044), the National Natural Science Foundation of China (2160736), Major Project of the Agricultural Basic Research Program in Yunnan Province (202101BD070001-020), and the Yunnan Talent Support Program Young Talent (XDYC-QNRC-2022-0245) The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Tacconelli E., Carrara E., Savoldi A., Harbarth S., Mendelson M., Monnet D.L., et al. (2018). Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis. 18, 318–327. doi: 10.1016/S1473-3099(17)30753-3 . [DOI] [PubMed] [Google Scholar]
- 2.Wang Y., Luo Q., Xiao T., Zhu Y., Xiao Y. (2022). Impact of polymyxin resistance on virulence and fitness among clinically important Gram-negative bacteria. Engineering. 13, 178–185. doi: 10.1016/j.eng.2020.11.005 [DOI] [Google Scholar]
- 3.Murray C.J., Ikuta K.S., Sharara F., Swetschinski L., Aguilar G.R., Gray A., et al. (2022). Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. The Lancet. 399, 629–655. doi: 10.1016/S0140-6736(21)02724-0 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.De Oliveira D. M., Forde B. M., Kidd T. J., Harris P. N., Schembri M. A., Beatson S. A., et al. (2020). Antimicrobial resistance in ESKAPE pathogens. Clinical microbiology reviews, 33(3), e00181–19. doi: 10.1128/CMR.00181-19 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Mulani M. S., Kamble E. E., Kumkar S. N., Tawre M. S., Pardesi K. R. (2019). Emerging strategies to combat ESKAPE pathogens in the era of antimicrobial resistance: a review. Frontiers in microbiology, 10, 539. doi: 10.3389/fmicb.2019.00539 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Vrancianu C. O., Gheorghe I., Dobre E. G., Barbu I. C., Cristian R. E., Popa M., et al. (2020). Emerging strategies to combat β-lactamase producing ESKAPE pathogens. International journal of molecular sciences, 21(22), 8527. doi: 10.3390/ijms21228527 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lai C. K., Ng R. W., Leung S. S., Hui M., Ip M. (2022). Overcoming the rising incidence and evolving mechanisms of antibiotic resistance by novel drug delivery approaches–an overview. Advanced Drug Delivery Reviews, 181, 114078. doi: 10.1016/j.addr.2021.114078 . [DOI] [PubMed] [Google Scholar]
- 8.Huemer M., Mairpady Shambat S., Brugger S. D., Zinkernagel A. S. (2020). Antibiotic resistance and persistence—Implications for human health and treatment perspectives. EMBO reports, 21(12), e51034. doi: 10.15252/embr.202051034 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Chovanová K., Zámocký M. (2016). Detection of the antibacterial effect of Chaetomium cochliodes Palliser CCM F-232 based on agar plugs and unprocessed fungal substances from cultivation media. Biologia, 71(11), 1204–1211. doi: 10.1515/biolog-2016-0153 [DOI] [Google Scholar]
- 10.Yang Y. H., Yang D. S., Li G. H, Pu X. J, Mo M. H, Zhao P. J. (2019). Antibacterial diketopiperazines from an endophytic fungus Bionectria sp. Y1085. J Antibiot (Tokyo). doi: 10.1038/s41429-019-0209-5 . [DOI] [PubMed] [Google Scholar]
- 11.Kaaniche F., Hamed A., Abdel-Razek A. S., Wibberg. D, Abdissa N., El Euch I. Z., et al. (2019). Bioactive secondary metabolites from new endophytic fungus Curvularia. sp isolated from Rauwolfia macrophylla. PLoS One. doi: 10.1371/journal.pone.0217627 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Senadeera S.P., Wiyakrutta S., Mahidol C., Ruchirawat S., Kittakoop P. (2012). A novel tricyclic polyketide and its biosynthetic precursor azaphilone derivatives from the endophytic fungus Dothideomycete sp. Organic and Biomolecular Chemistry. 10, 7220–7226. doi: 10.1039/c2ob25959a . [DOI] [PubMed] [Google Scholar]
- 13.Singh A., Singh D. K., Kharwar R. N., White J. F., Gond S. K. (2021). Fungal endophytes as efficient sources of plant-derived bioactive compounds and their prospective applications in natural product drug discovery: Insights, avenues, and challenges. Microorganisms. doi: 10.3390/microorganisms9010197 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Geethangili M., Tzeng Y. M. (2011). Review of pharmacological effects of Antrodia camphorata and its bioactive compounds. Evidence-based complementary and alternative medicine, 2011. doi: 10.1093/ecam/nep108 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kuang Y., Li B., Wang Z., Qiao X., Ye M. (2021). Terpenoids from the medicinal mushroom Antrodia camphorata: Chemistry and medicinal potential. Natural Product Reports. 38, 83–102. doi: 10.1039/d0np00023j . [DOI] [PubMed] [Google Scholar]
- 16.Chuang S. Y., Chen C. Y., Yang S. C., Alalaiwe A., Lin C. H., Fang J. Y., et al. (2021). 2, 4-dimethoxy-6-methylbenzene-1, 3-diol, a benzenoid from Antrodia cinnamomea, Mitigates Psoriasiform inflammation by suppressing MAPK/NF-κB phosphorylation and GDAP1L1/Drp1 translocation. Frontiers in Immunology, 12, 664425. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Chang C. J., Lu C. C., Lin C. S., Martel J., Ko Y. F., Ojcius D. M., et al. (2018). Antrodia cinnamomea reduces obesity and modulates the gut microbiota in high-fat diet-fed mice. International journal of obesity, 42(2), 231–243. doi: 10.1038/ijo.2017.149 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Chao T. Y., Hsieh C. C., Hsu S. M., Wan C. H., Lian G. T., Tseng Y. H., et al. (2021). Ergostatrien-3β-ol (EK100) from Antrodia camphorata attenuates oxidative stress, inflammation, and liver injury in vitro and in vivo. Preventive Nutrition and Food Science, 26(1), 58. doi: 10.3746/pnf.20-21.26.1.58 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Geethangili M., Fang S.H., Lai C.H., Rao Y.K., Lien H.M., Tzeng Y.M. (2010) Inhibitory effect of Antrodia camphorata constituents on the Helicobacter pylori-associated gastric inflammation. Food Chemistry. 119(1), 149–153. doi: 10.1016/j.foodchem.2009.06.006 [DOI] [Google Scholar]
- 20.Lien H. M., Tseng C. J., Huang C. L., Lin Y. T., Chen C. C., Lai Y. Y. (2014). Antimicrobial activity of Antrodia camphorata extracts against oral bacteria. PLoS One, 9(8), e105286. doi: 10.13-71/journal.pone.0105286 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zhang Z., Wang Y., Yuan X. L., Luo Y. N., Luo M. N., Zheng Y. (2022). Effects of culture mechanism of Cinnamomum kanehirae and C. camphora on the expression of genes related to terpene biosynthesis in Antrodia cinnamomea. Mycobiology, 50(2), 121–131. doi: 10.1080/12298093.2022.2059156 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Salam M.A., Al-Amin M.Y., Pawar J.S, Akhter N, Lucy I.B. (2023). Conventional methods and future trends in antimicrobial susceptibility testing. Saudi J Biol Sci. doi: 10.1016/j.sjbs.2023.103582 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Guo L.F., Liu G.R., Liu L. (2020). Caryophyllene-type sesquiterpenoids and α-furanones from the plant endophytic fungus Pestalotiopsis theae. Chin J Nat Med. doi: 10.1016/S1875-5364(20)30032-7 . [DOI] [PubMed] [Google Scholar]
- 24.Lin M.F., Lan C.Y. (2014) Antimicrobial resistance in Acinetobacter baumannii: From bench to bedside. World Journal of Clinical Cases: WJCC. 2(12), 787. doi: 10.12998/wjcc.v2.i12.787 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Nation R.L., Rigatto M.H.P., Falci D.R., Zavascki A.P. (2019). Polymyxin acute kidney injury: dosing and other strategies to reduce toxicity. Antibiotics. 8(1), 24. doi: 10.3390/antibiotics8010024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Zhanel G. G., Golden A. R., Zelenitsky S., Wiebe K., Lawrence C. K., Adam H. J., et al. (2019). Cefiderocol: a siderophore cephalosporin with activity against carbapenem-resistant and multidrug-resistant gram-negative bacilli. Drugs, 79, 271–289. doi: 10.1007/s40265-019-1055-2 . [DOI] [PubMed] [Google Scholar]
- 27.Portsmouth S., van Veenhuyzen D., Echols R., Machida M., Ferreira J. C. A., Ariyasu M., et al. (2018). Cefiderocol versus imipenem-cilastatin for the treatment of complicated urinary tract infections caused by Gram-negative uropathogens: a phase 2, randomised, double-blind, non-inferiority trial. The Lancet Infectious Diseases,18(12), 1319–1328. doi: 10.1016/S1473-3099(18)30554-1 . [DOI] [PubMed] [Google Scholar]
- 28.Katsube T., Saisho Y., Shimada J., Furuie H. (2019). Intrapulmonary pharmacokinetics of cefiderocol, a novel siderophore cephalosporin, in healthy adult subjects. Journal of Antimicrobial Chemotherapy, 74(7), 1971–1974. doi: 10.1093/jac/dkz123 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Liu J., Zhang C. L., Zhang Q. Y. (2018). Study on the active ingredients of endophytic fungus Alternaria tenuissima PC-005 against drugresistant bacteria. Zhongnan Pharmacy, 16(6), 741–744. [Google Scholar]
- 30.Duraipandiyan V., Ignacimuthu S. (2009). Antibacterial and antifungal activity of Flindersine isolated from the traditional medicinal plant, Toddalia asiatica (L.) Lam. Journal of ethnopharmacology. doi: 10.1016/j.jep.2009.02.020 . [DOI] [PubMed] [Google Scholar]
- 31.Ningsih B.N.S., Rukachaisirikul V., Pansrinun S., Phongpaichit S., Preedanon S., Sakayaroj J. (2022). New aromatic polyketides from the marine-derived fungus Pseudopithomyces maydicus PSU-AMF350 and their antimicrobial activity. Natural Product Research, 36(19), 4982–4989. [DOI] [PubMed] [Google Scholar]
- 32.Wu M.D., Cheng M.J., Wang W.Y., Huang H.C., Yuan G.F., Chen J.J., et al. (2011). Antioxidant activities of extracts and metabolites isolated from the fungus Antrodia cinnamomea. Natural Product Research. 25(16), 1488–1496. doi: 10.1080/14786410903132563 . [DOI] [PubMed] [Google Scholar]
- 33.Lien H.M., Lin H.W., Wang Y.J., Chen L.C., Yang D.Y., Lai Y.Y., et al. (2011). Inhibition of anchorage-independent proliferation and G0/G1 cell-cycle regulation in human colorectal carcinoma cells by 4, 7-dimethoxy-5-methyl-l, 3-benzodioxole isolated from the fruiting body of Antrodia camphorate. Evidence-Based Complementary and Alternative Medicine. 2011. doi: 10.1093/ecam/nep020 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Chu M., Zhang M. B., Liu Y. C., Kang J. R., Chu Z. Y., Yin K. L., et al. (2016). Role of berberine in the treatment of methicillin-resistant Staphylococcus aureus infections. Scientific reports, 6(1), 24748. doi: 10.1038/srep24748 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Zhang Q., Lyu Y., Huang J., Zhang X., Yu N., Wen Z., Chen S. (2020). Antibacterial activity and mechanism of sanguinarine against Providencia rettgeri in vitro. PeerJ, 8, e9543. doi: 10.7717/peerj.9543 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.He N., Wang P., Wang P., Ma C., Kang W. (2018). Antibacterial mechanism of chelerythrine isolated from root of Toddalia asiatica (Linn) Lam. BMC Complementary and Alternative Medicine, 18(1), 1–9. doi: 10.1186/s12906-018-2317-3 .. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.He H., Zhuo R., Dai J., Wang X., Huang X., Wang H., et al. (2020). Chelerythrine induces apoptosis via ROS‐mediated endoplasmic reticulum stress and STAT3 pathways in human renal cell carcinoma. Journal of cellular and molecular medicine, 24(1), 50–60. doi: 10.1111/jcmm.14295 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Qian W. D., Huang J., Zhang J. N., Li X. C., Kong Y., Wang T., et al. (2021). Antimicrobial and Antibiofilm Activities and Mechanism of Action of Chelerythrine Against Carbapenem-Resistant Serratia marcescens In Vitro. Microbial Drug Resistance, 27(8), 1105–1116. doi: 10.1089/mdr.2020.0207 . [DOI] [PubMed] [Google Scholar]
- 39.Hillman E. T., Readnour L. R., Solomon K. V. (2017). Exploiting the natural product potential of fungi with integrated-omics and synthetic biology approaches. Current Opinion in Systems Biology. doi: 10.1016/j.coisb.2017.07.010 [DOI] [Google Scholar]
- 40.Gupta D. S., Rao B. G., Bommaka K. M., Raghavendra M. N., Aleti S. (2016). Eco-sustainable synthesis and biological evaluation of 2-phenyl 1,3-benzodioxole derivatives as anticancer, DNA binding and antibacterial agents. Arabian Journal of Chemistry. doi: 10.1016/j.arabjc.2014.08.004 [DOI] [Google Scholar]
- 41.Phuong D. T., Ma C. M., Hattori M., Jin J. S. (2009). Inhibitory effects of antrodins A-E from Antrodia cinnamomea and their metabolites on hepatitis C virus protease. Phytother Res. doi: 10.1002/ptr.2657 . [DOI] [PubMed] [Google Scholar]
- 42.Hu Y. D., Lu R. Q., Liao X. R, Zhang B. B, Xu G. R. (2016). Stimulating the biosynthesis of antroquinonol by addition of effectors and soybean oil in submerged fermentation of Antrodia camphorata. Biotechnol Appl Biochem. doi: 10.1002/bab.1387 [DOI] [PubMed] [Google Scholar]
- 43.Lu Z. M., Geng Y., Li H. X., Sun Q., Shi J. S., Xu Z. H. (2014). Alpha-terpineol promotes triterpenoid production of Antrodia cinnamomea in submerged culture. FEMS Microbiol Lett. doi: 10.1111/1574-6968.12545 . [DOI] [PubMed] [Google Scholar]
- 44.Xia Y., Zhou X., Liang L., Liu X., Li H., Xiong Z., et al. (2022). Genetic evidence for the requirements of antroquinonol biosynthesis by Antrodia camphorata during liquid-state fermentation. Journal of Industrial Microbiology and Biotechnology, 49(1), kuab086. doi: 10.1093/jimb/kuab086 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Meng L., Luo B., Yang Y., Faruque M. O., Zhang J., Li X., et al. (2021). Addition of vegetable oil to improve triterpenoids production in liquid fermentation of medicinal fungus Antrodia cinnamomea. Journal of Fungi, 7(11), 926. doi: 10.3390/jof7110926 . [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOCX)
(DOCX)
Gen. represents Gentamicin, Amp. represents Ampicillin, Str. represents Streptomycin, Tet. represents Tetracycline, Kan. represents Kanamycin, BD represents A. camphorata extracts (50.00 mg/mL), BD-1 represents A. camphorata extracts (25.00 mg/mL), BD-2 represents A. camphorata extracts (12.50 mg/mL). The figures, from left to right and top to bottom, represent A. baumannii, S. aureus, P. aeruginosa, E. coli, K. pneumoniae, and E. faecalis.
(DOCX)
(DOCX)
(DOCX)
(DOCX)
(DOCX)
(DOCX)
MBBD was not cytotoxic to human normal lung epithelial cells BEAS-2B.
(DOCX)
Data Availability Statement
All relevant data are within the manuscript and its Supporting information files.