

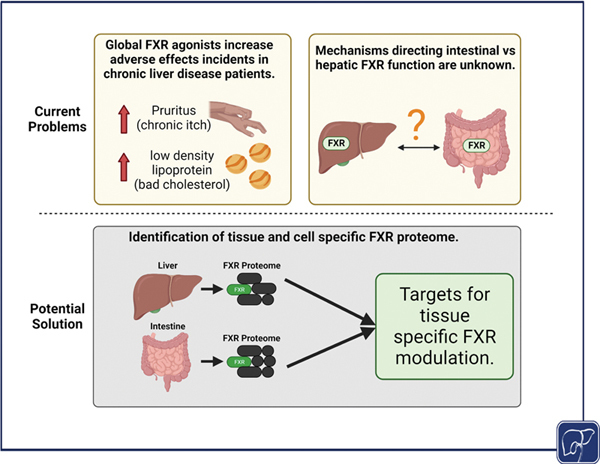
Abstract
Chronic liver diseases encompass a wide spectrum of hepatic maladies that often result in cholestasis or altered bile acid secretion and regulation. Incidence and cost of care for many chronic liver diseases are rising in the United States with few Food and Drug Administration-approved drugs available for patient treatment. Farnesoid X receptor (FXR) is the master regulator of bile acid homeostasis with an important role in lipid and glucose metabolism and inflammation. FXR has served as an attractive target for management of cholestasis and fibrosis; however, global FXR agonism results in adverse effects in liver disease patients, severely affecting quality of life. In this review, we highlight seminal studies and recent updates on the FXR proteome and identify gaps in knowledge that are essential for tissue-specific FXR modulation. In conclusion, one of the greatest unmet needs in the field is understanding the underlying mechanism of intestinal versus hepatic FXR function.
Keywords: bile acids, farnesoid X receptor, proteome, tissue-specific FXR functions
FXR
FXR is a member of the nuclear receptor superfamily and is widely recognized as the master regulator of bile acid synthesis and transport. 7 8 9 10 11 First discovered as a binding partner for retinoid X receptor (RXR), 12 FXR is highly expressed in the liver and intestine, where it carries out a major role in suppressing bile acid synthesis via downstream effectors, fibroblast growth factor (FGF) 15 (murine ortholog of human FGF19), and to a less extent small heterodimer partner (SHP). 13 14 Four isoforms of FXR (FXRα1–4) arise in humans and mice with alternative splicing of a 4-amino acid extension of the DNA binding domain, which separates FXRα1 and FXRα3 isoforms from FXRα2 and FXRα4. 15 16 17 Human liver preferentially expresses FXRα1 and FXRα2, while mouse liver tissue preferentially expresses FXRα2 or FXRα4, and human and mouse intestines preferentially express FXRα3 and FXRα4. 15 17 In both human and mouse livers, hepatic FXRα2 is the dominant driver of FXR agonism functions. 15 18 It is still unclear if FXR is a type I (cytoplasmic) or type II (nuclear) nuclear receptor, but its transcriptional activation has been extensively studied in the liver and intestine.
Canonical Function
In the gastrointestinal tract, FXR is highly expressed in the distal ileum and is critical in regulating enterohepatic bile acid homeostasis, including suppressing bile acid synthesis, and promoting bile acid transport. In the intestine, FXR is activated by bile acids to initiate the expression and secretion of FGF15/19 into portal circulation. Intestinal FXR activation regulates enterohepatic bile acid circulation through complex regulation of intestinal bile acid transporters, specifically promoting efflux and inhibiting influx of bile acids. In the ileum, apical sodium bile acid transporter expression is decreased while the expression of fatty acid binding protein 6 and organic solute transporter α and β expression are increased. 19 20 21 Further, FXR promotes epithelial layer integrity following activation through increased intestinal tight junction protein expression 22 and mucus production. 23 Moreover, FXR may modulate the ceramide production in the ileum to regulate metabolic diseases. 24 25 It is important to note that bile acids are metabolized and modified by the intestinal microbiome and there is a mutual relationship between bile acids and microbiome composition. 26 Bacteria create secondary bile acids via deconjugation, dihydroxylation at carbon 7, oxidation, and epimerization of primary bile acids to dampen antimicrobial function, alter intestinal immune microenvironment, and improve bacterial fitness. 26 27
In the liver, circulating FGF15/FGF19 binds to hepatic β-klotho and FGF receptor 4 dimer to inhibit gene expression of cytochrome P450 7a1 ( Cyp7a1 /CYP7A1) and 8b1 ( Cyp8b1 /CYP8B1), ultimately suppressing bile acid synthesis. 13 14 28 29 Circulating bile acids activate hepatic FXR leading to induction of SHP that mainly functions to inhibit Cyp8b1 expression. 14 29 30 31 32 Hepatocyte canalicular bile acid efflux transporter, bile salt export pump (BSEP), and sinusoidal uptake transporter, sodium taurocholate co-transporting polypeptide, are both regulated by hepatic FXR activation, serving as the main mechanism for hepatic bile acids to be transported from portal circulation into the bile canaliculi. 14 33 Hepatic FXR activation also results in reduced fatty acid synthesis 30 34 35 and hepatic inflammation. 36 37 38 Since CYP7A1 mediates the rate-limiting step of bile acid synthesis, and CYP8B1 determines bile acid hydrophobicity, it is generally considered that intestinal FXR is critical for regulating the bile acid pool and hydrophobicity, while hepatic FXR is critical in determining hydrophobicity of bile acids. 14 39 40 There are major differences between the murine and human bile acid speciation, which lends complexity to current studies of bile acid effects in disease states. 41 Overall, humans display a hydrophobic bile acid pool and mice exhibit hydrophilic bile acid pool with unique bile acid species, muricholic acids. 42 CYP2C70 has been identified as the enzyme responsible for α- and β-muricholic acid formation from chenodeoxycholic acid. 41 Murine models of CYP2C70 deficiency demonstrate a humanized bile acid pool with increased hepatic damage that is ameliorated following FXR activation. 41 43
Noncanonical Function
In recent years, our understanding of the impact of FXR activation has expanded from the enterohepatic system. FXR activation has been found to reduce lung macrophage activation following nitrogen mustard exposure 44 and increase β-oxidative gene expression in cardiomyocytes. 45 In the brain, FXR expression is correlated with Alzheimer's disease and loss of FXR reduces β-amyloid-induced brain injury. 46 FXR increases water reabsorption and promotes renal medullary collecting duct cell survival, ultimately affecting urine concentration during dehydration. 47 Besides, adipose-specific overexpression of FXR promotes brown adipose tissue whitening and fibrosis. 48 There is little information on FXR function in important sensory cells like cholangiocytes, tuft cells in the intestine, or chromaffin cells of the adrenals (PMID: 24068255, PMID: 35245089, PMID: 17963822). The ubiquitous expression of FXR in various organs, while less than in hepatocytes and ileal enterocytes, makes it crucial to understand FXR activation in a whole-body setting (PMID: 36988391). 44 45 46 47 49
FXR Function in Disease
The role of FXR in intestinal inflammation and fibrosis has been of increasing interest. Whole-body activation of FXR with obeticholic acid (OCA) in mice reduces dextran sodium sulfate (DSS) and trinitrobenzenesulfonic acid-induced colitis including immune cell infiltration and inflammatory cytokine expression. 22 Further, OCA, also known as INT-747, reduces proinflammatory cytokine secretion in activated mononuclear cells and monocytes derived from inflammatory bowel disease patients. 22 Notably, murine models of whole-body FXR loss demonstrate an enhanced inflammatory phenotype following DSS treatment with increased innate lymphoid cell presence within the damaged intestine and increased inflammatory cytokine expression. 50 Similarly, inhibition of ileal FXR by Parabacteroides distasonis improves hepatic fibrosis in mice fed methionine and choline-deficient diet. 51 Prophylactic FXR activation in the intestine, with tissue-specific FXR agonist fexaramine, prevents DSS-induced intestinal villus damage, serum interleukin 17 (IL-17) secretion, and immune cell infiltration of the intestine. 50 Function of fexaramine, and other fex-derivatives, is thought to be gastrointestinal-specific with heterogeneity of FXR activation depending on route of administration. 52 53 Oral administration of fexaramine is able to activate ileal FXR, with little to no activation in other colon, liver, and kidney, which is likely due to its increased interactions with helix 3 of the FXR protein and deeper penetration and filling of the ligand binding pocket due to fexaramine's hydrophobic rings and larger volume. 52 53 Fexaramine's intestine-specific activation is likely due to poor absorption into circulation. 53
While ileal FXR activation is widely regarded to contribute to hepatic function, liver FXR activation may also influence gut permeability. Hepatic FXR loss results in increased colonic mucus secretion and enhanced bacterial response gene expression profile. 23 Further, loss of hepatic FXR shifts the microbiome toward mucosal protection by reducing abundance of mucin-degrading genera ( Turicibacter ) and increasing abundance of mucus barrier-enforcing bacteria ( Roseburia, Bifidobacterium, and Clostridium sensu stricto 1 ). 23 FXR activation antagonizes nuclear factor kappa B (NF-κB) signaling which results in reduced hepatic inflammation. 37 Mice lacking FXR display increased hepatic inflammation following treatment with lipopolysaccharide, a bacterial cell wall component, which is ameliorated following transfection with FXRα2-adenovirus. 37 FXR activation prevents NF-κB activity through interference of NF-κB and DNA binding. 37
The effect of FXR activation on hepatic fibrosis is considered disease-dependent. 54 55 56 57 Loss of FXR has been shown to have no effect on hepatic fibrosis in mice following carbon tetrachloride treatment, a classical model of liver injury; however, in common bile duct ligated and 3,5-diethoxycarbonyl-1,4-dihydrocollidine-fed mice, loss of FXR directs protection against portal fibrosis in the liver. 54 FXR expression, as shown by immunohistochemistry, is found mainly in hepatocytes and cholangiocytes and minimally in murine myofibroblasts. 54 Conversely, it has been shown that FXR activation by OCA attenuates collagen deposition, α smooth muscle actin positive staining, and hepatic hydroxyproline content in mice treated with carbon tetrachloride and rats treated with thioacetamide. 55 56 57 The protective effects of FXR activation are thought to result from SMAD3 and FXR interaction following FXR activation. 55 Together these data suggest that FXR activation may have indirect effects on fibrosis, and loss of FXR improves portal fibrosis while global FXR activation improves noncholestatic hepatic fibrosis.
Known Mechanisms and Interactions of the FXR Proteome
Due to the synergistic roles of bile acids in lipid and glucose homeostasis, FXR regulation of bile acid synthesis and transport, and FXR antagonism effects on inflammation, FXR has been extensively researched as a therapeutic target for chronic liver diseases. This pursuit of global FXR agonists can be controversial in the context of disease treatment, largely due to our knowledge gaps in understanding mechanisms underlying tissue-specific FXR functions. 58 59 60
Originally speculated to be an independent bile acid sensor, the complex role of FXR cofactors in directing tissue-specific FXR response has been of growing interest. 61 62 63 FXR can inhibit gene expression of apolipoprotein A-I (ApoA-I) as a monomer or homodimer 64 ; however, FXR transcriptional activation is regarded to be a direct result of heterodimerization with other transcriptional regulators like RXR α (RXRα). 65 66 Interestingly, several factors are now shown to interact with and regulate FXR function. Hereon, we summarize a few of the suspected, and confirmed, members of the FXR proteome in the liver and intestine.
FXR Binding Partners
RXRα
RXRα is a nuclear receptor and promiscuous binding partner discovered to be the “missing factor” in various nuclear receptor transcriptional activity. 67 RXR isoforms, α, β, and γ, 68 are activated by 9- cis -retinoic acid and can act as a homodimer to activate the transcription of target genes. Heterodimerization of RXRs with other nuclear receptors can result in nonpermissive or silent partner function, which cannot be activated by RXR agonists, or in a permissive function, responding to ligand activation of either RXR or its partner nuclear receptor. 67 69 The FXR/RXR complex activated by RXR's endogenous ligands (e.g., 9- cis -retinoic acid) increased FXR-mediated transcriptional activation following activation by synthetic agonists (e.g., WAY-362450), suggesting that RXR activation promotes transcriptional activity of their permissive partners. 65 Like other RXR/nuclear receptor complexes, FXR/RXR heterodimer facilitates transactivation by binding to target sequences with RXR binding to 5′ half-site and its partner binding to the 3′ half-site of target sequences/response elements. 67 Interestingly, it has been found that FXR binding of the SHP promoter requires FXR interaction with the liver receptor homologue 1 (LRH-1) response element without LRH-1 binding; however, 9- cis -retinoic acid-dependent SHP expression requires RXRα occupation of the inverted repeat separated by 1 nucleotide (IR-1) site for subsequent SHP expression. 70 It was originally found that the FXR/RXR heterodimer can bind to the IR-1 sequence with high affinity; however, changes to the half-site sequences, spacing nucleotide, and flanking nucleotides are also bound by this heterodimer, shown pictorially in Fig. 1 . 71 While the FXR/RXR complex upholds many known behaviors of nuclear receptor interactions, their differential expressions may rely on unique site binding and cofactor recruitment at the time of ligand activation. 72
Fig. 1.
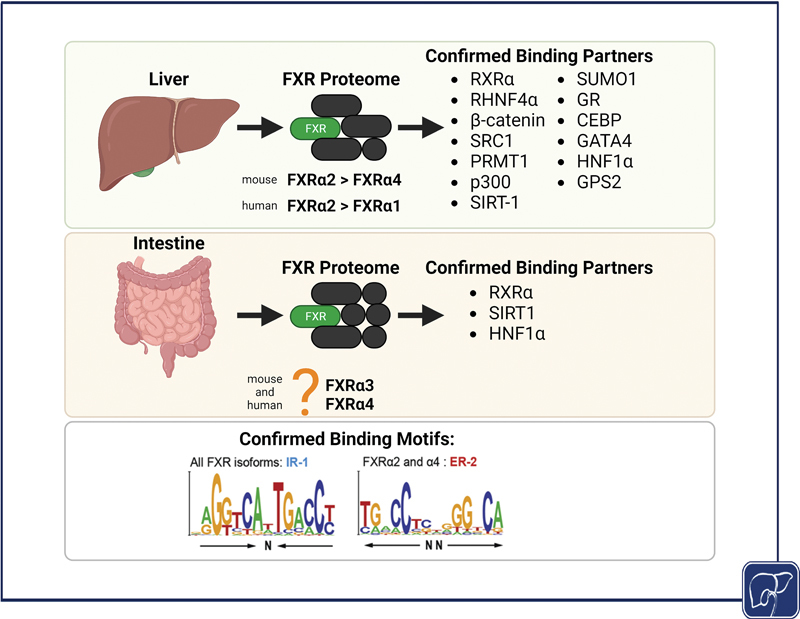
Current understanding of the FXR proteome. The liver in human and mice preferentially expresses FXRα2 to perform ligand-activated transcriptional activity. It is unknown which FXR isoform, FXRα3 or FXRα4, is preferentially expressed in the intestine. All FXR isoforms bind to the IR-1 motif while only FXR2 and FXR4 have been shown to bind the ER-2 DNA binding motifs. Identification of confirmed binding partners of the hepatic and intestinal FXR proteome, and studies on confirmed DNA binding motifs the FXR isoforms, may provide ideal targets for tissue-specific FXR therapeutics. Figure created with biorender.com and confirmed DNA binding motif sequences are repurposed with permission. 15
Hepatocyte Nuclear Factor 4 α
Hepatocyte nuclear factor 4 α (HNF4α) is an orphan nuclear receptor that is highly expressed in epithelial tissues of digestive organs such as the liver and intestine. HNF4α plays essential roles in enterohepatic development, hepatic metabolism, and regulation of hepatocyte cell fate of hepatic progenitor cells. 73 74 75 HNF4α is known to interact with other transcription factors to induce transcriptional regulation. 76 HNF4α and FXR share many target genes related to bile acid synthesis, albeit their actions are in an opposing manner, as HNF4α normally promotes, whereas FXR suppresses, the expression of genes in bile acid synthesis. 77 HNF4 regulates bile acid conjugation through expression of bile acid-CoA:amino acid N -acyltransferase and bile acid-CoA ligase. 78 The existence of the FXR/HNF4α complex has been established in mouse 77 and human hepatocytes. 79 Despite these findings, the mechanism or biological significance of the interaction between FXR and HNF4α remains unclear.
HNF4α not only interacts with FXR but also induces FXR gene expression. In the fasting state, peroxisome proliferator-activated receptor-gamma coactivator-1 α (PGC-1α) coactivates HNF4α to induce FXR transcription, favoring isoforms FXRα3 and FXRα4. 80 FXR competitively binds PGC-1α to inhibit transcriptional activation of sulfotransferase family 1E member 1 ( Sult1e1 ) gene by HNF4α. 81 HNF4α and LRH-1 interaction keeps Cyp7a1 gene in a transcriptionally active state, which can be reversed by SHP-FGF15/19-mediated suppression. Specifically, SHP inhibits LRH1 activity to prevent FGF15/19 activation of ERK and JNK pathways that activate Cyp7a1 gene transcription. 14 82 83 84 SHP has been shown to directly interact with HNF4α at the cysteine sulfinic acid decarboxylase promoter to inhibit its transcription and reduce downstream taurine production. 85 Liver zonation transcriptomics data found that Cyp7a1 expression is limited to pericentral hepatocytes, specifically referred to as layer 1, while HNF4α and FXR are equally expressed through all hepatocyte zones, layers 1 to 9. 86 Ubiquitous expression of HNF4α and FXR throughout the hepatocyte zones indicates that their downstream effects may rely on cofactor or ligand binding.
FXR Cofactors
It has been long suspected that FXR cofactors, unable to bind DNA but able to bind nuclear receptors, influence tissue-specific FXR activation. These regulatory cofactors often function in histone modification or chromatin remodeling capacities, inherently affecting the transcription of FXR target genes through their coactivator or corepressor function. Below we detail key studies that identify and investigate FXR/cofactor complexes in vitro and in vivo.
Cofactors via Posttranslational Modifications of FXR (PMRT, p300, SIRT1, SUMO1, SRC1, O-GlcNac Transferase)
Posttranslational modifications of proteins are important for cell homeostasis, proliferation, and stress response. Posttranslational modifications of FXR direct its function by altering DNA binding, ligand binding, heterodimer formation, and subcellular localization. 87 Protein arginine methyl-transferase type I (PMRT1), p300, and sirtuin 1 (SIRT1) can regulate transcription through methylation, acetylation, and deacetylation, respectively, of histone and nonhistone proteins. 88 89 90 Further, SIRT1 can interact with p300 to repress its transcriptional regulatory activity. 91 Small ubiquitin-like modifier (SUMO) proteins direct protein–protein interaction and cellular localization of nuclear receptors. 87 The steroid receptor coactivator 1 (SRC1) initiates p160 SRC family protein recruitment to regulate nuclear receptor function. 92 FXR is subject to methylation, phosphorylation, acetylation, SUMOylation, and O-GlcNAcylation at various sites including lysine 67, 122, and 127 and glutamate 277 and AF1 domain. 88 93
Methylation of FXR and FXR target gene histones by PRMT1 is essential for FXR activation. 88 Following treatment with a synthetic bile acid, OCA, FXR recruits PRMT1, which methylates histone H4 protein near promoter regions of BSEP and SHP . 88 This FXR activation is ablated in the presence of methylation inhibitors, indicating that methylation is important in regulating FXR transcriptional activity. Reduced methylation of FXR target promoter regions results in decreased FXR transcriptional activity and subsequently the level of conjugated bile acids in the liver. 94 Conversely, FXR transcriptional regulation of bile acid homeostasis requires phosphorylation by nonreceptor tyrosine kinase, Src, at tyrosine 67. 95 Phospho-defective FXR, or Src downregulation, disrupts the expression of FXR target genes and impairs bile acid homeostasis following cholic acid feeding in wild-type (WT) mice. 95
Acetylation of FXR and histones at the Shp /SHP promoter initiates SHP gene expression following FXR activation in mouse livers and HepG2 cells. 89 The recruitment of p300 is FXR-dependent as shown in FXR null mice who lack p300 recruitment and its subsequent acetylation at the Shp promoter. Interestingly, acetylation of FXR at lysine 157 and lysine 217 by p300 prevents the FXR/RXRα dimer formation. 66 Mutations at these acetylation sites result in retained RXR binding and ablated p300 acetylation. Further, it has been found that inhibition of p300, in vitro, resulted in increased ApoA-I and reduced G-6-Pase and phosphoenolpyruvate carboxykinase expression, which was unaffected by FXR activation with GW4064. 89 SIRT1 deacetylation of FXR promotes FXR/RXRα dimer formation with increased FXR transactivation. 66 Deletion of intestinal SIRT1 decreases FXR/HNF1α complex formation resulting in reduced bile acid transport and increased hepatic bile acid synthesis. 96
Hepatic fibrosis resolution remains a key goal in liver disease research. OCA has been shown to be an effective prophylactic treatment against fibrosis. 97 Activated hepatic stellate cells (HSCs) display increased FXR SUMOylation, which renders FXR unable to bind OCA. Prevention of FXR SUMOylation, in combination with OCA treatment, effectively reduces HSC activation and hepatic fibrosis formation in mice. 97 In addition, FXR/SUMO1 complex formation decreases FXR binding and recruitment to the BSEP and SHP promoters in HepG2 cells. 98
SRC1, along with other co-activators such as PGC-1α, is responsible for hepatocyte differentiation, metabolism, and homeostasis via HNF4α regulation. 99 Due to HNF4α-directed expression of Cyp7a1 , it is unsurprising that SRC1 impacts FXR activity. SRC1 interacts with the FXR ligand binding domain following the formation of the FXR/RXR complex. 100
FXR transcriptional activity is regulated by glucose and O-linked-N-acetylglucosaminylation (O-GlcNAc) of the N-terminus of the AF1 domain. 93 O-GlcNAc transferase regulates FXR activity during fasting and feeding through O-GlcNAcylation at serine 72 in murine FXRα1 and human FXRα3 and serine 62 in human FXRα2. 93 Further, O-GlcNAc transferase can also modify carbohydrate-responsive element-binding protein (ChREBP) to interact with O-GlcNAc–FXR under high glucose concentrations to express glycolysis and lipogenesis genes. 101 However, in the presence of bile acids, regardless of high glucose levels, ChREBP-target gene expression is inhibited. 101 In human hepatocytes, ligand activation of FXR inhibits glucose transcription of ChREBP genes. 79
Taken together, studies demonstrate that FXR transactivation is not only cofactor-dependent but driven by posttranslational modifications of FXR and target gene environments, including epigenetic modifications. Due to the complex nature of FXR regulation, targeting of individual cofactors will likely need to be disease- or cell-dependent. Understanding of FXR posttranslational modifications, as well as cofactors that induce them, will provide key insights into the regulation of FXR transcriptional activity in a tissue-specific manner.
Beta Catenin
Beta catenin (β-catenin) is a well-known and evolutionary conserved protein shown to be important in tight-junction formation, cell proliferation, and is integral to the Wnt signaling cascade. 102 In the liver, β-catenin regulates liver homeostasis, injury repair, and tumorigenesis, and protein expression is mainly found in pericentral hepatocytes. 103 104 105 106 While a relationship has been identified, the molecular mechanism of β-catenin and FXR interactions is undefined. In mouse models of hepatocellular carcinoma (HCC), FXR and β-catenin expression patterns display an inverse relationship. 107 108 HCC patients display decreased FXR expression, 108 while β-catenin expression increases in HCC patients and human-derived HCC cell lines 109 110 compared with controls.
In mouse hepatocytes, it is thought that β-catenin sequesters FXR resulting in reduced FXR availability to promote bile acid efflux via regulating bile acid transporter expression and coactivating pregnane X receptor to regulate Cyp3a11 gene expression. 111 GW4064 treatment in β-catenin knockout (KO) mice, subjected to bile duct ligation, demonstrates increased RXRα and FXR binding in hepatocytes. 111 Similarly, GW4064 treatment, in an α-naphthyl isothiocyanate model of biliary injury, shows increased FXR binding to RXRα and reduced β-catenin binding to FXR. 112 Bile duct ligation of transgenic mice overexpressing hepatocyte S45D- β-catenin and low-density lipoprotein receptor 5/6 double KO mice with deficient hepatocyte Wnt signaling demonstrates similar FXR/β-catenin complex levels to WT mice following immunoprecipitation pulldown. 113 Contradictory to these findings, patients with primary sclerosing cholangitis display reduced β-catenin protein expression and mRNA expression of SHP, FXR target gene, and Cyp7a1 , SHP target gene. 104 However, there could be other mechanisms to downregulate the expression of these genes, such as inflammation. Taken together, these studies indicate that the FXR/β-catenin complex inhibits hepatocyte FXR function, and due to peri-central protein expression pattern of β-catenin, exploration of interzonal and portal hepatocyte FXR should be further studied. 103 106 However, the formation of this complex may be transient, depending on injury caused by experimental cholestasis model, hepatocyte zonation, or ligand activation.
G Protein Pathway Suppressor 2
G protein pathway suppressor 2 (GPS2) is an epigenetic modifier and is considered one of the core subunits, along with silencing mediator of retinoid and thyroid receptors and nuclear receptor corepressor (NCOR), of the chromatin corepressor complex. 114 115 The role of GPS2 is critical in regulating transcription, e.g., the regulation of macrophage plasticity is conducted by tightly regulated chromatin remodeling and transcription regulation via the chromatin corepressor complex containing GPS2. 116 In murine models of nonalcoholic steatohepatitis (NASH), GPS2 has been shown to promote steatosis by antagonizing peroxisome proliferator-activated receptor α (PPARα) transcriptional activity with the corepressor, NCOR. 114 It has also been shown that hepatocyte GPS2 is required for hepatitis C virus replication in Huh-7 cell lines. 117
For bile acid regulation, GPS2 manifested a gene-specific regulation of CYP7A1 and CYP8B1 expression. Where functions to enhance SHP-mediated suppression of CYP7A1 gene transcription, GPS2 can recruit P300/CREB binding protein complex to the HNF4α response element and interact with FXR to form an enhancer/promoter loop for increased expression of CYP8B1 in HepG2 cells. 118 While little is known about the role of hepatic GPS2 in cholestasis, further investigation of the FXR/GPS2 complex may provide insight into its regulation of FXR activity.
Glucocorticoid Receptor
Glucocorticoid receptor (GR), a member of the nuclear receptor superfamily, is activated by glucocorticoids in the cytoplasm and translocates to the nucleus to activate various transcriptional pathways. 119 GR activation promotes anti-inflammatory signaling but can lead to cholestasis and insulin resistance. 120 The formation of FXR/GR complex prevents FXR-directed SHP expression through recruitment of C-terminal binding protein to the SHP promoter in HepG2 cells. 120 Hepatic GR activation increases autocrine regulation of Cyp7a1 through FGF21 secretion 121 and activation of FXR increases glucocorticoid secretion in WT mice. 122 More investigation is required to understand the FXR and GR interaction during FXR activation.
Recent Advances of the FXR Proteome
Utilization of global FXR agonists in primary biliary cholangitis and NASH patients remains controversial due to severe adverse effects such as pruritus, fatigue, and increased serum low-density lipoprotein. 123 124 125 In preclinical settings, inhibition of mast cell FXR reduces serum histamine levels and prevents bile duct damage in a murine model of mast cell-induced cholestasis. 58 In a murine model of nonalcoholic fatty liver disease, caffeic acid phenethyl ester treatment reduces steatosis through decreased bacterial bile salt hydrolase activity and increased tauro-β-muricholic acid, an endogenous FXR antagonist. 126 To prevent off-target effects of FXR agonism, the field must turn to understanding the tissue- and cell-specific roles of FXR.
Various research groups have explored the FXR interactome through chromatin immunoprecipitation (ChIP) with a greater focus on hepatic 15 127 128 than intestinal FXR. 129 Below we briefly describe seminal studies on the FXR proteome.
ChIP Insights
In humans, the dominance of FXR isoforms in the liver affects FXR activation responses. 15 Diseased livers from patients with NASH, cirrhosis, and HCC have increased FXRα1 isoform expression with preferential binding to the IR-1 DNA motif. 15 IR-1 binding by FXRα1 regulates bile acid metabolism/transport and inflammatory signaling. Patients with healthy or steatotic livers express increased FXRα2 with increased binding to everted repeat spaced by 2 nucleotides (ER-2) binding motif, shown pictorially in Fig. 1 . 15 In vitro exploration of HepG2 cells overexpressing FXRα1 or FXRα2 confirms preferential binding to IR-1 or ER-2 regulatory regions, respectively.
In mice, FXR binds IR-1 motifs at intergenic and intron regions, with additional clusters of FXR binding within 1–2 kb of transcription start sites. 128 129 130 FXR Re-ChIP analysis demonstrates that FXR/RXR co-occupancy of the SHP promoter is unchanged following FXR activation, despite a marked increase in SHP mRNA expression. 128 In normal and obese mice treated with GW4064, activated FXR represses a large amount of binding motifs identified by ChIP sequencing (ChIP-seq), which challenges previous understanding that SHP represses genes following FXR activation. 128 In vitro, FXR/RXR transcriptional activity increases with LRH-1 transfection and FXR/LRH-1 complex has been detected following co-immunoprecipitation. 130 Based on these findings, FXR transcriptional activation may depend on isoform expression, cofactor interaction, disease setting, and ligand binding.
A recent study of the hepatic FXR proteome demonstrates that cistrome, epigenetic, and protein forces regulate the specific biological pathways studied in various disease models. 127 Based on analysis of publicly available databases, LRH-1, retinoic acid receptor α (RARα), and GA-binding protein (GABPA) interact with FXR to direct its intracellular protein trafficking, protein metabolism, and cell cycle functions. 127 Conversely, Foxa1/2, nuclear factor interleukin 3 (NFIL3), RAR-related orphan receptor α (RORα), GR, NCOR1, and HNF1α interact with FXR to regulate lipid and steroid, amino acid, and carbohydrate metabolism. 127 It is important to recognize that many transcriptional regulators are shared between these two sets of FXR functions. In the WT mouse liver, only complexes with CCAAT/enhancer-binding protein β (CEBP), GATA binding protein 4 (GATA4), HNF1α, GR, and RXRα are confirmed to interact with FXR following rapid immunoprecipitation mass spectrometry of endogenous proteins, also called RIME. 127
One of the greatest unmet needs in the field is understanding the regulation of intestinal versus hepatic FXR function. Enterohepatic ChIP-seq reveals that only 11% of total FXR DNA binding sites are shared between the liver and intestine accounting for 1,713 genes. 129 Moreover, FXR binds 4,248 unique genes in the liver and 3,406 unique genes in the intestine. 129 The most enriched liver transcription pathways include metabolic and biosynthetic processes while the intestine is enriched for catalytic activity and oxidoreductase activity following FXR activation in WT mice. It has been found that mouse livers contain IR-1 DNA motifs while intestine presents with both IR-1 and ER-2. 129 These results suggest an organ-specific transcriptome is dependent on DNA regulatory element motifs. Further investigation of FXR proteome formation, duration, and ligand dependency, in liver and intestine, will allow researchers to develop targeted therapeutics to enhance specific FXR functions.
Discussion
We have outlined the concerted efforts of transcriptional regulators in the diverse functions of FXR activation in the liver and intestine, summarized in Tables 1 and 2 , and highlighted known binding partners of tissue-specific FXR isoforms, summarized in Fig. 1 . However, to the best of our knowledge, few intestinal FXR proteome studies have been published to date. Increased focus on defining the intestinal FXR proteome may assist in identifying FXR protein complexes for therapeutic functions due to DNA binding heterogeneity and potential unique protein interactions. Similarly, there is little knowledge of the FXR proteome in key bile acid facing cells like cholangiocytes, endothelial cells, and renal cells. Understanding FXR function through its binding partners in these few but impactful cells will help researchers attenuate adverse effects of global FXR agonism. Continued practice of open-access ChIP-seq datasets, as done with the FXR super-signaling atlas that combines multiple single datasets into an interactive platform, 131 can inspire researchers to solve the FXR proteome puzzle.
Table 1. Posttranslational modifications of FXR.
| Enzyme, modification | Modification target | Function | Reference |
|---|---|---|---|
| PMRT1, methylation | Promoter region | Increases BSEP and SHP mRNA expression Increases FXR transcriptional activity Increases conjugated bile acids (liver) |
88 94 |
| p300, acetylation | Promoter region, FXR lysine 157 and lysine 217 |
Increases SHP expression Prevents FXR/RXRα dimerization |
66 89 |
| SIRT-1, deacetylation | FXR | Increases FXR transcriptional activity Promotes FXR/RXRα dimerization |
66 |
| SUMO1, SUMOylation | FXR | Decreases FXR binding to BSEP and SHP promoters | 98 |
| Src kinase, phosphorylation | FXR tyrosine 67 | Increases FXR transcriptional activity | 95 |
| O-linked-N-acetylglucosamine transferase, O-GlcNAc | FXR serine 62 or 72, isoform-dependent | Increases glycolytic and lipogenic gene expression (in absence of FXR ligands) | 79 93 101 |
Abbreviation: FXR, farnesoid X receptor.
Table 2. FXR binding partners.
| Binding partners | Detection method | Function | Reference |
|---|---|---|---|
| RXRα | EMSA, ChIP, co-IP, ALPHA | Increases FXR transcriptional activity | 65 67 70 |
| HNF4α | ChIP-Seq | Unknown biological significance | 77 |
| β-catenin | ChIP | Inhibits FXR transcription through inhibitory complex formation | 104 113 |
| GPS-2 | Yeast two-hybrid interaction screening | Increases Cyp7a1 and Cyp8b1 expression | 118 |
| GR | ChIP, Co-IP | Represses FXR transcriptional activity and reduced hepatic gluconeogenesis | 120 121 |
| SRC1 | Protein crystallization | SRC1 binds FXR ligand binding domain in FXR/RXRα complex | 100 |
Abbreviations: ALPHA, amplified luminescence proximity homogenous assay; ChIP, chromatin immunoprecipitation; ChIP-seq, ChIP sequencing; Co-IP, co-immunoprecipitation; EMSA, electrophoretic mobility shift assay; FXR, farnesoid X receptor.
Conclusion
While we believe that deciphering tissue-specific FXR proteomes is the key to understanding the tissue-specific FXR function, the role of chromatin structure, FXR isoform expression, hepatocyte liver zonation, and DNA binding affinity cannot be ignored. The recruitment of FXR activators results in histone modification and chromatin remodeling, beyond the initial euchromatin opening by tissue-specific pioneer factors, to allow the expression of target genes. Moreover, FXR isoform expression and their protein and DNA binding affinity also impact FXR transcriptional activity and ligand activation. Liver zonation may influence FXR function through cofactor expression, ligand secretion, and downstream FXR gene expression. Effort must be made to combine research in chromatin environment, DNA binding motifs, and proteome analysis to push the field of nuclear receptor biology forward.
Funding Statement
Funding This work was supported by the National Institutes of Health (grants: ES007148, ES029258, DK122725, GM135258, AND GM093854), the Department of Veteran Affairs (BX002741), and the Momental Foundation Mistletoe Research Fellowship (FP00032129). The authors would like to thank Rulaiha Elizabeth Taylor, Zakiyah Henry, and Dr. Bo Kong for their support to this work and Dr. Saskia van Mil for her permission to reuse two figure panels of FXR binding motif sequences found in Fig. 1 .
Footnotes
Conflict of Interest None declared.
Lay Summary.
Occurrence and treatment cost of chronic liver disease are increasing in the United States with few Food and Drug Administration-approved drugs available for patients. A common symptom of liver disease is reduced or blocked bile flow from the liver, which is regulated by farnesoid X receptor (FXR), a nuclear receptor protein that is important for regulating liver function. FXR must bind other proteins to control bile acid synthesis and bile flow and has unique organ-dependent roles. Understanding how FXR activity is controlled in different organs is an urgent unmet need in liver and intestinal disease research. In this review, we summarize the first findings of FXR-associated proteins and highlight recent studies addressing the knowledge gap for organ-specific FXR research.
References
- 1.Hirode G, Saab S, Wong R J. Trends in the burden of chronic liver disease among hospitalized US adults. JAMA Netw Open. 2020;3(04):e201997. doi: 10.1001/jamanetworkopen.2020.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Abeysekera K WM, Macpherson I, Glyn-Owen K et al. Community pathways for the early detection and risk stratification of chronic liver disease: a narrative systematic review. Lancet Gastroenterol Hepatol. 2022;7(08):770–780. doi: 10.1016/S2468-1253(22)00020-6. [DOI] [PubMed] [Google Scholar]
- 3.Boyer J L. Bile formation and secretion. Compr Physiol. 2013;3(03):1035–1078. doi: 10.1002/cphy.c120027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Anson M L. The denaturation of proteins by synthetic detergents and bile salts. J Gen Physiol. 1939;23(02):239–246. doi: 10.1085/jgp.23.2.239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hofmann A F. Micellar solubilization of fatty acids and monoglycerides by bile salt solutions. Nature. 1961;190:1106–1107. doi: 10.1038/1901106a0. [DOI] [PubMed] [Google Scholar]
- 6.Jones H, Alpini G, Francis H. Bile acid signaling and biliary functions. Acta Pharm Sin B. 2015;5(02):123–128. doi: 10.1016/j.apsb.2015.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Chiang J Y, Kimmel R, Weinberger C, Stroup D. Farnesoid X receptor responds to bile acids and represses cholesterol 7alpha-hydroxylase gene (CYP7A1) transcription. J Biol Chem. 2000;275(15):10918–10924. doi: 10.1074/jbc.275.15.10918. [DOI] [PubMed] [Google Scholar]
- 8.Makishima M, Okamoto A Y, Repa J Jet al. Identification of a nuclear receptor for bile acids Science 1999284(5418):1362–1365. [DOI] [PubMed] [Google Scholar]
- 9.Parks D J, Blanchard S G, Bledsoe R Ket al. Bile acids: natural ligands for an orphan nuclear receptor Science 1999284(5418):1365–1368. [DOI] [PubMed] [Google Scholar]
- 10.Wang H, Chen J, Hollister K, Sowers L C, Forman B M. Endogenous bile acids are ligands for the nuclear receptor FXR/BAR. Mol Cell. 1999;3(05):543–553. doi: 10.1016/s1097-2765(00)80348-2. [DOI] [PubMed] [Google Scholar]
- 11.Forman B M, Goode E, Chen J et al. Identification of a nuclear receptor that is activated by farnesol metabolites. Cell. 1995;81(05):687–693. doi: 10.1016/0092-8674(95)90530-8. [DOI] [PubMed] [Google Scholar]
- 12.Seol W, Choi H S, Moore D D. Isolation of proteins that interact specifically with the retinoid X receptor: two novel orphan receptors. Mol Endocrinol. 1995;9(01):72–85. doi: 10.1210/mend.9.1.7760852. [DOI] [PubMed] [Google Scholar]
- 13.Inagaki T, Choi M, Moschetta A et al. Fibroblast growth factor 15 functions as an enterohepatic signal to regulate bile acid homeostasis. Cell Metab. 2005;2(04):217–225. doi: 10.1016/j.cmet.2005.09.001. [DOI] [PubMed] [Google Scholar]
- 14.Kong B, Wang L, Chiang J Y, Zhang Y, Klaassen C D, Guo G L. Mechanism of tissue-specific farnesoid X receptor in suppressing the expression of genes in bile-acid synthesis in mice. Hepatology. 2012;56(03):1034–1043. doi: 10.1002/hep.25740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ramos Pittol J M, Milona A, Morris I et al. FXR isoforms control different metabolic functions in liver cells via binding to specific DNA motifs. Gastroenterology. 2020;159(05):1.853E13–1.865E13. doi: 10.1053/j.gastro.2020.07.036. [DOI] [PubMed] [Google Scholar]
- 16.Huber R M, Murphy K, Miao Bet al. Generation of multiple farnesoid-X-receptor isoforms through the use of alternative promoters Gene 2002290(1–2):35–43. [DOI] [PubMed] [Google Scholar]
- 17.Zhang Y, Kast-Woelbern H R, Edwards P A. Natural structural variants of the nuclear receptor farnesoid X receptor affect transcriptional activation. J Biol Chem. 2003;278(01):104–110. doi: 10.1074/jbc.M209505200. [DOI] [PubMed] [Google Scholar]
- 18.Boesjes M, Bloks V W, Hageman J et al. Hepatic farnesoid X-receptor isoforms α2 and α4 differentially modulate bile salt and lipoprotein metabolism in mice. PLoS One. 2014;9(12):e115028. doi: 10.1371/journal.pone.0115028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Chen F, Ma L, Dawson P A et al. Liver receptor homologue-1 mediates species- and cell line-specific bile acid-dependent negative feedback regulation of the apical sodium-dependent bile acid transporter. J Biol Chem. 2003;278(22):19909–19916. doi: 10.1074/jbc.M207903200. [DOI] [PubMed] [Google Scholar]
- 20.Dawson P A, Haywood J, Craddock A L et al. Targeted deletion of the ileal bile acid transporter eliminates enterohepatic cycling of bile acids in mice. J Biol Chem. 2003;278(36):33920–33927. doi: 10.1074/jbc.M306370200. [DOI] [PubMed] [Google Scholar]
- 21.Grober J, Zaghini I, Fujii H et al. Identification of a bile acid-responsive element in the human ileal bile acid-binding protein gene. Involvement of the farnesoid X receptor/9-cis-retinoic acid receptor heterodimer. J Biol Chem. 1999;274(42):29749–29754. doi: 10.1074/jbc.274.42.29749. [DOI] [PubMed] [Google Scholar]
- 22.Gadaleta R M, van Erpecum K J, Oldenburg B et al. Farnesoid X receptor activation inhibits inflammation and preserves the intestinal barrier in inflammatory bowel disease. Gut. 2011;60(04):463–472. doi: 10.1136/gut.2010.212159. [DOI] [PubMed] [Google Scholar]
- 23.Ijssennagger N, van Rooijen K S, Magnúsdóttir S et al. Ablation of liver Fxr results in an increased colonic mucus barrier in mice . JHEP Rep. 2021;3(05):100344. doi: 10.1016/j.jhepr.2021.100344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Jiang C, Xie C, Li F et al. Intestinal farnesoid X receptor signaling promotes nonalcoholic fatty liver disease. J Clin Invest. 2015;125(01):386–402. doi: 10.1172/JCI76738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Jiang C, Xie C, Lv Y et al. Intestine-selective farnesoid X receptor inhibition improves obesity-related metabolic dysfunction. Nat Commun. 2015;6:10166. doi: 10.1038/ncomms10166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Guzior D V, Quinn R A. Review: microbial transformations of human bile acids. Microbiome. 2021;9(01):140. doi: 10.1186/s40168-021-01101-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zhang Y, Gao X, Gao S et al. Effect of gut flora mediated-bile acid metabolism on intestinal immune microenvironment. Immunology. 2023 doi: 10.1111/imm.13672. [DOI] [PubMed] [Google Scholar]
- 28.Song K H, Li T, Owsley E, Strom S, Chiang J Y. Bile acids activate fibroblast growth factor 19 signaling in human hepatocytes to inhibit cholesterol 7alpha-hydroxylase gene expression. Hepatology. 2009;49(01):297–305. doi: 10.1002/hep.22627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kim I, Ahn S H, Inagaki T et al. Differential regulation of bile acid homeostasis by the farnesoid X receptor in liver and intestine. J Lipid Res. 2007;48(12):2664–2672. doi: 10.1194/jlr.M700330-JLR200. [DOI] [PubMed] [Google Scholar]
- 30.Goodwin B, Jones S A, Price R R et al. A regulatory cascade of the nuclear receptors FXR, SHP-1, and LRH-1 represses bile acid biosynthesis. Mol Cell. 2000;6(03):517–526. doi: 10.1016/s1097-2765(00)00051-4. [DOI] [PubMed] [Google Scholar]
- 31.Lu T T, Makishima M, Repa J J et al. Molecular basis for feedback regulation of bile acid synthesis by nuclear receptors. Mol Cell. 2000;6(03):507–515. doi: 10.1016/s1097-2765(00)00050-2. [DOI] [PubMed] [Google Scholar]
- 32.Miao J, Choi S E, Seok S M et al. Ligand-dependent regulation of the activity of the orphan nuclear receptor, small heterodimer partner (SHP), in the repression of bile acid biosynthetic CYP7A1 and CYP8B1 genes. Mol Endocrinol. 2011;25(07):1159–1169. doi: 10.1210/me.2011-0033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Rizzo G, Renga B, Mencarelli A, Pellicciari R, Fiorucci S. Role of FXR in regulating bile acid homeostasis and relevance for human diseases. Curr Drug Targets Immune Endocr Metabol Disord. 2005;5(03):289–303. doi: 10.2174/1568008054863781. [DOI] [PubMed] [Google Scholar]
- 34.Clifford B L, Sedgeman L R, Williams K J et al. FXR activation protects against NAFLD via bile-acid-dependent reductions in lipid absorption. Cell Metab. 2021;33(08):1.671E7–1.684E7. doi: 10.1016/j.cmet.2021.06.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Watanabe M, Houten S M, Wang L et al. Bile acids lower triglyceride levels via a pathway involving FXR, SHP, and SREBP-1c. J Clin Invest. 2004;113(10):1408–1418. doi: 10.1172/JCI21025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Gai Z, Visentin M, Gui T et al. Effects of farnesoid X receptor activation on arachidonic acid metabolism, NF-kB signaling, and hepatic inflammation. Mol Pharmacol. 2018;94(02):802–811. doi: 10.1124/mol.117.111047. [DOI] [PubMed] [Google Scholar]
- 37.Wang Y D, Chen W D, Wang M, Yu D, Forman B M, Huang W. Farnesoid X receptor antagonizes nuclear factor kappaB in hepatic inflammatory response. Hepatology. 2008;48(05):1632–1643. doi: 10.1002/hep.22519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Xu Z, Huang G, Gong W et al. FXR ligands protect against hepatocellular inflammation via SOCS3 induction. Cell Signal. 2012;24(08):1658–1664. doi: 10.1016/j.cellsig.2012.04.015. [DOI] [PubMed] [Google Scholar]
- 39.Li T, Matozel M, Boehme S et al. Overexpression of cholesterol 7α-hydroxylase promotes hepatic bile acid synthesis and secretion and maintains cholesterol homeostasis. Hepatology. 2011;53(03):996–1006. doi: 10.1002/hep.24107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Pandak W M, Bohdan P, Franklund C et al. Expression of sterol 12alpha-hydroxylase alters bile acid pool composition in primary rat hepatocytes and in vivo. Gastroenterology. 2001;120(07):1801–1809. doi: 10.1053/gast.2001.24833. [DOI] [PubMed] [Google Scholar]
- 41.de Boer J F, Verkade E, Mulder N L et al. A human-like bile acid pool induced by deletion of hepatic Cyp2c70 modulates effects of FXR activation in mice . J Lipid Res. 2020;61(03):291–305. doi: 10.1194/jlr.RA119000243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Li J, Dawson P A. Animal models to study bile acid metabolism. Biochim Biophys Acta Mol Basis Dis. 2019;1865(05):895–911. doi: 10.1016/j.bbadis.2018.05.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.de Boer J F, de Vries H D, Palmiotti A et al. Cholangiopathy and biliary fibrosis in Cyp2c70-deficient mice are fully reversed by ursodeoxycholic acid. Cell Mol Gastroenterol Hepatol. 2021;11(04):1045–1069. doi: 10.1016/j.jcmgh.2020.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Murray A, Banota T, Guo G L et al. Farnesoid X receptor regulates lung macrophage activation and injury following nitrogen mustard exposure. Toxicol Appl Pharmacol. 2022;454:116208. doi: 10.1016/j.taap.2022.116208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Guo G L, Santamarina-Fojo S, Akiyama T E et al. Effects of FXR in foam-cell formation and atherosclerosis development. Biochim Biophys Acta. 2006;1761(12):1401–1409. doi: 10.1016/j.bbalip.2006.09.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Yan N, Yan T, Xia Y, Hao H, Wang G, Gonzalez F J. The pathophysiological function of non-gastrointestinal farnesoid X receptor. Pharmacol Ther. 2021;226:107867. doi: 10.1016/j.pharmthera.2021.107867. [DOI] [PubMed] [Google Scholar]
- 47.Guo Y, Xie G, Zhang X. Role of FXR in renal physiology and kidney diseases. Int J Mol Sci. 2023;24(03):24. doi: 10.3390/ijms24032408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Yang J, de Vries H D, Mayeuf-Louchart A et al. Role of bile acid receptor FXR in development and function of brown adipose tissue. Biochim Biophys Acta Mol Cell Biol Lipids. 2023;1868(02):159257. doi: 10.1016/j.bbalip.2022.159257. [DOI] [PubMed] [Google Scholar]
- 49.Ding L, Yang L, Wang Z, Huang W. Bile acid nuclear receptor FXR and digestive system diseases. Acta Pharm Sin B. 2015;5(02):135–144. doi: 10.1016/j.apsb.2015.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Fu T, Li Y, Oh T G et al. FXR mediates ILC-intrinsic responses to intestinal inflammation. Proc Natl Acad Sci U S A. 2022;119(51):e2213041119. doi: 10.1073/pnas.2213041119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Zhao Q, Dai M Y, Huang R Y et al. Parabacteroides distasonis ameliorates hepatic fibrosis potentially via modulating intestinal bile acid metabolism and hepatocyte pyroptosis in male mice. Nat Commun. 2023;14(01):1829. doi: 10.1038/s41467-023-37459-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Downes M, Verdecia M A, Roecker A J et al. A chemical, genetic, and structural analysis of the nuclear bile acid receptor FXR. Mol Cell. 2003;11(04):1079–1092. doi: 10.1016/s1097-2765(03)00104-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Fang S, Suh J M, Reilly S M et al. Intestinal FXR agonism promotes adipose tissue browning and reduces obesity and insulin resistance. Nat Med. 2015;21(02):159–165. doi: 10.1038/nm.3760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Fickert P, Fuchsbichler A, Moustafa T et al. Farnesoid X receptor critically determines the fibrotic response in mice but is expressed to a low extent in human hepatic stellate cells and periductal myofibroblasts. Am J Pathol. 2009;175(06):2392–2405. doi: 10.2353/ajpath.2009.090114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Fan Y Y, Ding W, Zhang C, Fu L, Xu D X, Chen X. Obeticholic acid prevents carbon tetrachloride-induced liver fibrosis through interaction between farnesoid X receptor and Smad3. Int Immunopharmacol. 2019;77:105911. doi: 10.1016/j.intimp.2019.105911. [DOI] [PubMed] [Google Scholar]
- 56.Zhou J, Huang N, Guo Y et al. Combined obeticholic acid and apoptosis inhibitor treatment alleviates liver fibrosis. Acta Pharm Sin B. 2019;9(03):526–536. doi: 10.1016/j.apsb.2018.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Verbeke L, Mannaerts I, Schierwagen R et al. FXR agonist obeticholic acid reduces hepatic inflammation and fibrosis in a rat model of toxic cirrhosis. Sci Rep. 2016;6:33453. doi: 10.1038/srep33453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Meadows V, Kennedy L, Ekser B et al. Mast cells regulate ductular reaction and intestinal inflammation in cholestasis through farnesoid X receptor signaling. Hepatology. 2021;74(05):2684–2698. doi: 10.1002/hep.32028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kjærgaard K, Frisch K, Sørensen M et al. Obeticholic acid improves hepatic bile acid excretion in patients with primary biliary cholangitis. J Hepatol. 2021;74(01):58–65. doi: 10.1016/j.jhep.2020.07.028. [DOI] [PubMed] [Google Scholar]
- 60.Mudaliar S, Henry R R, Sanyal A J et al. Efficacy and safety of the farnesoid X receptor agonist obeticholic acid in patients with type 2 diabetes and nonalcoholic fatty liver disease. Gastroenterology. 2013;145(03):5740–820. doi: 10.1053/j.gastro.2013.05.042. [DOI] [PubMed] [Google Scholar]
- 61.Eloranta J J, Kullak-Ublick G A. Coordinate transcriptional regulation of bile acid homeostasis and drug metabolism. Arch Biochem Biophys. 2005;433(02):397–412. doi: 10.1016/j.abb.2004.09.019. [DOI] [PubMed] [Google Scholar]
- 62.Nettles K W, Greene G L. Nuclear receptor ligands and cofactor recruitment: is there a coactivator “on deck”? Mol Cell. 2003;11(04):850–851. doi: 10.1016/s1097-2765(03)00133-3. [DOI] [PubMed] [Google Scholar]
- 63.Henry Z, Meadows V, Guo G L. FXR and NASH: an avenue for tissue-specific regulation. Hepatol Commun. 2023;7(05):7. doi: 10.1097/HC9.0000000000000127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Claudel T, Sturm E, Duez H et al. Bile acid-activated nuclear receptor FXR suppresses apolipoprotein A-I transcription via a negative FXR response element. J Clin Invest. 2002;109(07):961–971. doi: 10.1172/JCI14505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Zheng W, Lu Y, Tian S et al. Structural insights into the heterodimeric complex of the nuclear receptors FXR and RXR. J Biol Chem. 2018;293(32):12535–12541. doi: 10.1074/jbc.RA118.004188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Kemper J K, Xiao Z, Ponugoti B et al. FXR acetylation is normally dynamically regulated by p300 and SIRT1 but constitutively elevated in metabolic disease states. Cell Metab. 2009;10(05):392–404. doi: 10.1016/j.cmet.2009.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Mangelsdorf D J, Evans R M. The RXR heterodimers and orphan receptors. Cell. 1995;83(06):841–850. doi: 10.1016/0092-8674(95)90200-7. [DOI] [PubMed] [Google Scholar]
- 68.Wagner C E, Jurutka P W, Marshall P A, Heck M C. Retinoid X receptor selective agonists and their synthetic methods. Curr Top Med Chem. 2017;17(06):742–767. doi: 10.2174/1568026616666160617091559. [DOI] [PubMed] [Google Scholar]
- 69.Shulman A I, Larson C, Mangelsdorf D J, Ranganathan R. Structural determinants of allosteric ligand activation in RXR heterodimers. Cell. 2004;116(03):417–429. doi: 10.1016/s0092-8674(04)00119-9. [DOI] [PubMed] [Google Scholar]
- 70.Hoeke M O, Heegsma J, Hoekstra M, Moshage H, Faber K N. Human FXR regulates SHP expression through direct binding to an LRH-1 binding site, independent of an IR-1 and LRH-1. PLoS One. 2014;9(02):e88011. doi: 10.1371/journal.pone.0088011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Laffitte B A, Kast H R, Nguyen C M, Zavacki A M, Moore D D, Edwards P A. Identification of the DNA binding specificity and potential target genes for the farnesoid X-activated receptor. J Biol Chem. 2000;275(14):10638–10647. doi: 10.1074/jbc.275.14.10638. [DOI] [PubMed] [Google Scholar]
- 72.Jiang L, Liu X, Liang X et al. Structural basis of the farnesoid X receptor/retinoid X receptor heterodimer on inverted repeat DNA. Comput Struct Biotechnol J. 2023;21:3149–3157. doi: 10.1016/j.csbj.2023.05.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Thakur A, Wong J CH, Wang E Y et al. Hepatocyte nuclear factor 4-alpha is essential for the active epigenetic state at enhancers in mouse liver. Hepatology. 2019;70(04):1360–1376. doi: 10.1002/hep.30631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Bookout A L, Jeong Y, Downes M, Yu R T, Evans R M, Mangelsdorf D J. Anatomical profiling of nuclear receptor expression reveals a hierarchical transcriptional network. Cell. 2006;126(04):789–799. doi: 10.1016/j.cell.2006.06.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Yamagata K, Furuta H, Oda Net al. Mutations in the hepatocyte nuclear factor-4alpha gene in maturity-onset diabetes of the young (MODY1) Nature 1996384(6608):458–460. [DOI] [PubMed] [Google Scholar]
- 76.Yeh M M, Bosch D E, Daoud S S. Role of hepatocyte nuclear factor 4-alpha in gastrointestinal and liver diseases. World J Gastroenterol. 2019;25(30):4074–4091. doi: 10.3748/wjg.v25.i30.4074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Thomas A M, Hart S N, Li G et al. Hepatocyte nuclear factor 4 alpha and farnesoid X receptor co-regulates gene transcription in mouse livers on a genome-wide scale. Pharm Res. 2013;30(09):2188–2198. doi: 10.1007/s11095-013-1006-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Inoue Y, Yu A M, Inoue J, Gonzalez F J. Hepatocyte nuclear factor 4alpha is a central regulator of bile acid conjugation. J Biol Chem. 2004;279(04):2480–2489. doi: 10.1074/jbc.M311015200. [DOI] [PubMed] [Google Scholar]
- 79.Caron S, Huaman Samanez C, Dehondt H et al. Farnesoid X receptor inhibits the transcriptional activity of carbohydrate response element binding protein in human hepatocytes. Mol Cell Biol. 2013;33(11):2202–2211. doi: 10.1128/MCB.01004-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Zhang Y, Castellani L W, Sinal C J, Gonzalez F J, Edwards P A. Peroxisome proliferator-activated receptor-gamma coactivator 1alpha (PGC-1alpha) regulates triglyceride metabolism by activation of the nuclear receptor FXR. Genes Dev. 2004;18(02):157–169. doi: 10.1101/gad.1138104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Wang S, Yuan X, Lu D, Guo L, Wu B. Farnesoid X receptor regulates SULT1E1 expression through inhibition of PGC1α binding to HNF4α. Biochem Pharmacol. 2017;145:202–209. doi: 10.1016/j.bcp.2017.08.023. [DOI] [PubMed] [Google Scholar]
- 82.Kir S, Zhang Y, Gerard R D, Kliewer S A, Mangelsdorf D J. Nuclear receptors HNF4α and LRH-1 cooperate in regulating Cyp7a1 in vivo. J Biol Chem. 2012;287(49):41334–41341. doi: 10.1074/jbc.M112.421834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Li T, Jahan A, Chiang J Y. Bile acids and cytokines inhibit the human cholesterol 7 alpha-hydroxylase gene via the JNK/c-jun pathway in human liver cells. Hepatology. 2006;43(06):1202–1210. doi: 10.1002/hep.21183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Gupta S, Stravitz R T, Dent P, Hylemon P B. Down-regulation of cholesterol 7alpha-hydroxylase (CYP7A1) gene expression by bile acids in primary rat hepatocytes is mediated by the c-Jun N-terminal kinase pathway. J Biol Chem. 2001;276(19):15816–15822. doi: 10.1074/jbc.M010878200. [DOI] [PubMed] [Google Scholar]
- 85.Wang Y, Matye D, Nguyen N, Zhang Y, Li T. HNF4α regulates CSAD to couple hepatic taurine production to bile acid synthesis in mice. Gene Expr. 2018;18(03):187–196. doi: 10.3727/105221618X15277685544442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Halpern K B, Shenhav R, Matcovitch-Natan Oet al. Single-cell spatial reconstruction reveals global division of labour in the mammalian liver Nature 2017542(7641):352–356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Appelman M D, van der Veen S W, van Mil S WC. Post-translational modifications of FXR; implications for cholestasis and obesity-related disorders. Front Endocrinol (Lausanne) 2021;12:729828. doi: 10.3389/fendo.2021.729828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Rizzo G, Renga B, Antonelli E, Passeri D, Pellicciari R, Fiorucci S. The methyl transferase PRMT1 functions as co-activator of farnesoid X receptor (FXR)/9-cis retinoid X receptor and regulates transcription of FXR responsive genes. Mol Pharmacol. 2005;68(02):551–558. doi: 10.1124/mol.105.012104. [DOI] [PubMed] [Google Scholar]
- 89.Fang S, Tsang S, Jones R et al. The p300 acetylase is critical for ligand-activated farnesoid X receptor (FXR) induction of SHP. J Biol Chem. 2008;283(50):35086–35095. doi: 10.1074/jbc.M803531200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Rahman S, Islam R. Mammalian Sirt1: insights on its biological functions. Cell Commun Signal. 2011;9:11. doi: 10.1186/1478-811X-9-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Bouras T, Fu M, Sauve A A et al. SIRT1 deacetylation and repression of p300 involves lysine residues 1020/1024 within the cell cycle regulatory domain 1. J Biol Chem. 2005;280(11):10264–10276. doi: 10.1074/jbc.M408748200. [DOI] [PubMed] [Google Scholar]
- 92.Walsh C A, Qin L, Tien J C, Young L S, Xu J. The function of steroid receptor coactivator-1 in normal tissues and cancer. Int J Biol Sci. 2012;8(04):470–485. doi: 10.7150/ijbs.4125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Berrabah W, Aumercier P, Gheeraert C et al. Glucose sensing O-GlcNAcylation pathway regulates the nuclear bile acid receptor farnesoid X receptor (FXR) Hepatology. 2014;59(05):2022–2033. doi: 10.1002/hep.26710. [DOI] [PubMed] [Google Scholar]
- 94.Cabrerizo R, Castaño G O, Burgueño A L et al. Promoter DNA methylation of farnesoid X receptor and pregnane X receptor modulates the intrahepatic cholestasis of pregnancy phenotype. PLoS One. 2014;9(01):e87697. doi: 10.1371/journal.pone.0087697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Byun S, Kim D H, Ryerson D et al. Postprandial FGF19-induced phosphorylation by Src is critical for FXR function in bile acid homeostasis. Nat Commun. 2018;9(01):2590. doi: 10.1038/s41467-018-04697-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Kazgan N, Metukuri M R, Purushotham A et al. Intestine-specific deletion of SIRT1 in mice impairs DCoH2-HNF-1α-FXR signaling and alters systemic bile acid homeostasis. Gastroenterology. 2014;146(04):1006–1016. doi: 10.1053/j.gastro.2013.12.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Zhou J, Cui S, He Q et al. SUMOylation inhibitors synergize with FXR agonists in combating liver fibrosis. Nat Commun. 2020;11(01):240. doi: 10.1038/s41467-019-14138-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Balasubramaniyan N, Luo Y, Sun A Q, Suchy F J. SUMOylation of the farnesoid X receptor (FXR) regulates the expression of FXR target genes. J Biol Chem. 2013;288(19):13850–13862. doi: 10.1074/jbc.M112.443937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Martínez-Jiménez C P, Gómez-Lechón M J, Castell J V, Jover R. Underexpressed coactivators PGC1alpha and SRC1 impair hepatocyte nuclear factor 4 alpha function and promote dedifferentiation in human hepatoma cells. J Biol Chem. 2006;281(40):29840–29849. doi: 10.1074/jbc.M604046200. [DOI] [PubMed] [Google Scholar]
- 100.Wang N, Zou Q, Xu J, Zhang J, Liu J. Ligand binding and heterodimerization with retinoid X receptor α (RXRα) induce farnesoid X receptor (FXR) conformational changes affecting coactivator binding. J Biol Chem. 2018;293(47):18180–18191. doi: 10.1074/jbc.RA118.004652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Benhamed F, Filhoulaud G, Caron S, Lefebvre P, Staels B, Postic C. O-GlcNAcylation links ChREBP and FXR to glucose-sensing. Front Endocrinol (Lausanne) 2015;5:230. doi: 10.3389/fendo.2014.00230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Valenta T, Hausmann G, Basler K. The many faces and functions of β-catenin. EMBO J. 2012;31(12):2714–2736. doi: 10.1038/emboj.2012.150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Goel C, Monga S P, Nejak-Bowen K. Role and regulation of Wnt/β-catenin in hepatic perivenous zonation and physiological homeostasis. Am J Pathol. 2022;192(01):4–17. doi: 10.1016/j.ajpath.2021.09.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Ayers M, Liu S, Singhi A D, Kosar K, Cornuet P, Nejak-Bowen K. Changes in beta-catenin expression and activation during progression of primary sclerosing cholangitis predict disease recurrence. Sci Rep. 2022;12(01):206. doi: 10.1038/s41598-021-04358-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Zhang S, Zhang J, Evert K et al. The hippo effector transcriptional coactivator with PDZ-binding motif cooperates with oncogenic β-catenin to induce hepatoblastoma development in mice and humans. Am J Pathol. 2020;190(07):1397–1413. doi: 10.1016/j.ajpath.2020.03.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Zummo F P, Berthier A, Gheeraert C et al. A time- and space-resolved nuclear receptor atlas in mouse liver. J Mol Endocrinol. 2023;71(01):71. doi: 10.1530/JME-23-0017. [DOI] [PubMed] [Google Scholar]
- 107.Kim I, Morimura K, Shah Y, Yang Q, Ward J M, Gonzalez F J. Spontaneous hepatocarcinogenesis in farnesoid X receptor-null mice. Carcinogenesis. 2007;28(05):940–946. doi: 10.1093/carcin/bgl249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Wolfe A, Thomas A, Edwards G, Jaseja R, Guo G L, Apte U. Increased activation of the Wnt/β-catenin pathway in spontaneous hepatocellular carcinoma observed in farnesoid X receptor knockout mice. J Pharmacol Exp Ther. 2011;338(01):12–21. doi: 10.1124/jpet.111.179390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Xu C, Xu Z, Zhang Y, Evert M, Calvisi D F, Chen X. β-Catenin signaling in hepatocellular carcinoma. J Clin Invest. 2022;132(04):132. doi: 10.1172/JCI154515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Liu X, Zhang X, Ji L, Gu J, Zhou M, Chen S. Farnesoid X receptor associates with β-catenin and inhibits its activity in hepatocellular carcinoma. Oncotarget. 2015;6(06):4226–4238. doi: 10.18632/oncotarget.2899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Thompson M D, Moghe A, Cornuet P et al. β-Catenin regulation of farnesoid X receptor signaling and bile acid metabolism during murine cholestasis. Hepatology. 2018;67(03):955–971. doi: 10.1002/hep.29371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Liu J, Liu J, Meng C et al. NRF2 and FXR dual signaling pathways cooperatively regulate the effects of oleanolic acid on cholestatic liver injury. Phytomedicine. 2023;108:154529. doi: 10.1016/j.phymed.2022.154529. [DOI] [PubMed] [Google Scholar]
- 113.Zhang R, Nakao T, Luo J et al. Activation of WNT/beta-catenin signaling and regulation of the farnesoid X receptor/beta-catenin complex after murine bile duct ligation. Hepatol Commun. 2019;3(12):1642–1655. doi: 10.1002/hep4.1430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Liang N, Damdimopoulos A, Goñi S et al. Hepatocyte-specific loss of GPS2 in mice reduces non-alcoholic steatohepatitis via activation of PPARα. Nat Commun. 2019;10(01):1684. doi: 10.1038/s41467-019-09524-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Huang Z, Liang N, Goñi S et al. The corepressors GPS2 and SMRT control enhancer and silencer remodeling via eRNA transcription during inflammatory activation of macrophages. Mol Cell. 2021;81(05):9.53E11–9.68E11. doi: 10.1016/j.molcel.2020.12.040. [DOI] [PubMed] [Google Scholar]
- 116.Fan R, Toubal A, Goñi S et al. Loss of the co-repressor GPS2 sensitizes macrophage activation upon metabolic stress induced by obesity and type 2 diabetes. Nat Med. 2016;22(07):780–791. doi: 10.1038/nm.4114. [DOI] [PubMed] [Google Scholar]
- 117.Xu G, Xin X, Zheng C. GPS2 is required for the association of NS5A with VAP-A and hepatitis C virus replication. PLoS One. 2013;8(11):e78195. doi: 10.1371/journal.pone.0078195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Sanyal S, Båvner A, Haroniti A et al. Involvement of corepressor complex subunit GPS2 in transcriptional pathways governing human bile acid biosynthesis. Proc Natl Acad Sci U S A. 2007;104(40):15665–15670. doi: 10.1073/pnas.0706736104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Petta I, Dejager L, Ballegeer M et al. The interactome of the glucocorticoid receptor and its influence on the actions of glucocorticoids in combatting inflammatory and infectious diseases. Microbiol Mol Biol Rev. 2016;80(02):495–522. doi: 10.1128/MMBR.00064-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Lu Y, Zhang Z, Xiong X et al. Glucocorticoids promote hepatic cholestasis in mice by inhibiting the transcriptional activity of the farnesoid X receptor. Gastroenterology. 2012;143(06):1630–1.64E11. doi: 10.1053/j.gastro.2012.08.029. [DOI] [PubMed] [Google Scholar]
- 121.Al-Aqil F A, Monte M J, Peleteiro-Vigil Aet al. Interaction of glucocorticoids with FXR/FGF19/FGF21-mediated ileum-liver crosstalk Biochim Biophys Acta Mol Basis Dis 20181864(9, Pt B):2927–2937. [DOI] [PubMed] [Google Scholar]
- 122.Hoekstra M, van der Sluis R J, Li Z, Oosterveer M H, Groen A K, Van Berkel T J.FXR agonist GW4064 increases plasma glucocorticoid levels in C57BL/6 mice Mol Cell Endocrinol 2012362(1–2):69–75. [DOI] [PubMed] [Google Scholar]
- 123.REGENERATE Study Investigators Younossi Z M, Ratziu V, Loomba Ret al. Obeticholic acid for the treatment of non-alcoholic steatohepatitis: interim analysis from a multicentre, randomised, placebo-controlled phase 3 trial Lancet 2019394(10215):2184–2196. [DOI] [PubMed] [Google Scholar]
- 124.POISE Study Group . Nevens F, Andreone P, Mazzella G et al. A placebo-controlled trial of obeticholic acid in primary biliary cholangitis. N Engl J Med. 2016;375(07):631–643. doi: 10.1056/NEJMoa1509840. [DOI] [PubMed] [Google Scholar]
- 125.Xu J, Wang Y, Khoshdeli M et al. IL-31 levels correlate with pruritus in patients with cholestatic and metabolic liver diseases and is farnesoid X receptor responsive in NASH. Hepatology. 2023;77(01):20–32. doi: 10.1002/hep.32599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Zhong X C, Liu Y M, Gao X X et al. Caffeic acid phenethyl ester suppresses intestinal FXR signaling and ameliorates nonalcoholic fatty liver disease by inhibiting bacterial bile salt hydrolase activity. Acta Pharmacol Sin. 2023;44(01):145–156. doi: 10.1038/s41401-022-00921-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Dubois-Chevalier J, Dubois V, Dehondt H et al. The logic of transcriptional regulator recruitment architecture at cis -regulatory modules controlling liver functions . Genome Res. 2017;27(06):985–996. doi: 10.1101/gr.217075.116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Lee J, Seok S, Yu P et al. Genomic analysis of hepatic farnesoid X receptor binding sites reveals altered binding in obesity and direct gene repression by farnesoid X receptor in mice. Hepatology. 2012;56(01):108–117. doi: 10.1002/hep.25609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Thomas A M, Hart S N, Kong B, Fang J, Zhong X B, Guo G L. Genome-wide tissue-specific farnesoid X receptor binding in mouse liver and intestine. Hepatology. 2010;51(04):1410–1419. doi: 10.1002/hep.23450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Chong H K, Infante A M, Seo Y K et al. Genome-wide interrogation of hepatic FXR reveals an asymmetric IR-1 motif and synergy with LRH-1. Nucleic Acids Res. 2010;38(18):6007–6017. doi: 10.1093/nar/gkq397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Jungwirth E, Panzitt K, Marschall H U, Wagner M, Thallinger G G. A comprehensive FXR signaling atlas derived from pooled ChIP-seq data. Stud Health Technol Inform. 2019;260:105–112. [PubMed] [Google Scholar]