

Significance
Most animals have specific brain neurons that regulate sleep-wake cycles and other aspects of circadian behavior. Drosophila has only about 150 of these clock neurons. Despite their small numbers, they have remarkably diverse anatomy and gene expression profiles. To address the different functions of these neurons, we used highly specific and efficient CRISPR-based methods to create cell type–specific disruptions of three traditional circadian genes. We were able to assign the function of the photoreceptor cryptochrome to two tiny subsets of clock neurons. In addition, two independent methods assigned the neuropeptide PDF (pigment dispersing factor) to the adult stage. In summary, we find that the CRISPR-based methods are very efficient at studying adult-specific functions of genes in small, discrete sets of neurons.
Keywords: adult-specific CRISPR, cell type–specific CRISPR, circadian, clock neurons, cryptochrome
Abstract
Circadian behavioral rhythms in Drosophila melanogaster are regulated by about 75 pairs of brain neurons. They all express the core clock genes but have distinct functions and gene expression profiles. To understand the importance of these distinct molecular programs, neuron-specific gene manipulations are essential. Although RNAi based methods are standard to manipulate gene expression in a cell-specific manner, they are often ineffective, especially in assays involving smaller numbers of neurons or weaker Gal4 drivers. We and others recently exploited a neuron-specific CRISPR-based method to mutagenize genes within circadian neurons. Here, we further explore this approach to mutagenize three well-studied clock genes: the transcription factor gene vrille, the photoreceptor gene Cryptochrome (cry), and the neuropeptide gene Pdf (pigment dispersing factor). The CRISPR-based strategy not only reproduced their known phenotypes but also assigned cry function for different light-mediated phenotypes to discrete, different subsets of clock neurons. We further tested two recently published methods for temporal regulation in adult neurons, inducible Cas9 and the auxin-inducible gene expression system. The results were not identical, but both approaches successfully showed that the adult-specific knockout of the neuropeptide Pdf reproduces the canonical loss-of-function mutant phenotypes. In summary, a CRISPR-based strategy is a highly effective, reliable, and general method to temporally manipulate gene function in specific adult neurons.
Organismal behavior involves sensing the environment, internal processing, and motor actions. In multicellular organisms, this process is regulated largely by cells of the nervous system which often have defined roles as parts of bigger networks and circuits. One behavior that is almost ubiquitous to organisms on earth is aligning activity bouts to the daily rotations of the earth. The underlying mechanism is circadian rhythmicity.
In Drosophila, circadian clocks tick away in about 75 pairs of central brain clock neurons. These neurons all express the core circadian machinery, which includes the transcription factors CLOCK/CYCLE and the circadian inhibitory proteins PER/TIM. Rhythmic expression of these genes is important for maintaining circadian rhythmicity, and together they form a core transcription–translation feedback loop whose activity can be entrained by environmental cues such as light (1–3). Clock neurons also express several other proteins, such as the transcription factors VRILLE (VRI) and PAR-domain Protein 1 (PDP1); their rhythmic expression is important for circadian behavior (4, 5).
Many additional genes are specifically expressed within clock neurons and also contribute to rhythmic behavior. Pigment dispersing factor (Pdf) encodes a circadian neuropeptide expressed exclusively within the small and large LNvs (sLNvs and lLNvs); Pdf mutants are arrhythmic and show increased sleep (6, 7). Another gene expressed by roughly half the clock neurons coding for the photoreceptor cryptochrome is cry (8–11). Wild-type flies are arrhythmic in constant light, whereas cry mutants are rhythmic in this condition (12). Despite the specificity in expression and probably function, few studies have explored the cell type–specific functions of these genes (13), and the cellular focus of many of circadian genes remains unknown.
Functional roles of specific circadian neurons have been broadly associated with the 7 to 8 subgroups based on anatomical location. For example, the PDF-positive sLNvs are key regulators of rhythmicity under constant darkness and regulate morning activity. Another group of neurons, the six LNds along with the 5th sLNv, regulate evening activity and thus were labeled evening cells (14–17). Yet, other clock neuron subgroups play accessory roles in regulating rhythmicity and even sleep. They include the PDF-positive lLNvs, the DN1s, the DN2s, the LPNs and the mysterious DN3s (18–22).
Recent single-cell sequencing indicates at least 17 distinct subgroups, about twice as many as originally identified based on anatomical location (23). It is likely that many of these new subgroups are also functionally distinct. Fortunately, many of them can now be accessed through judicious use of the Gal4/Upstream Activating Sequence (UAS) system (24, 25), which allows for cell type–specific perturbations. This is because there are multiple Gal4 and split Gal4 drivers available to access specific circadian neurons (26, 27).
These drivers can be combined with the standard method of UAS-RNA interference (RNAi) to knockdown gene expression. Although this approach works well in many cases and has been invaluable for cell type–specific perturbations, it is often variable and usually induces partial knockdown (28, 29). Fortunately, it is now possible to disrupt genes completely and in the same cell-specific manner using Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)-Cas9-based gene editing (30, 31). Cas9 is an enzyme that induces a double-stranded break at a specific genomic location determined by the guideRNA (gRNA) sequence (32). The error prone repair machinery endogenous to all cells usually leads to mutations known as indels (insertions-deletions). These range from a missense mutation to a small addition or deletion causing frameshift mutations, often disrupting protein function (31).
Earlier adaptations of this method achieved cell specificity by expressing the Cas9 protein in a Gal4-dependent manner while ubiquitously expressing a single-gRNA against a gene of interest. The method was found to be more consistent and effective than RNAi in inducing loss of function phenotypes (29, 33). More recent developments such as multiplexing guides against a gene in an array under UAS control further improved both the efficiency and cell type specificity (33, 34). This improved method worked well for disrupting the functions of the core clock genes period (per) and timeless (tim) in circadian neurons (35, 36), and it was more recently extended to the disruption of genes encoding G-protein-coupled receptors in these neurons (37).
Here, we implement this method to further understand the roles of three circadian behavior-associated genes: vri, cry, and Pdf. We chose to generate a CRISPR-based line against vri because there are no successful RNAi data in the literature, and we chose cry and Pdf to address precise cell type–specific functions. Lastly, the CRISPR/Cas9 strategy was combined with two recent methods for temporal regulation, which successfully restricted gene editing and the resulting phenotype to specific neurons in just the adult stage. The results taken together substantially add to previous circadian analyses and identify specific subsets of clock neurons within which these genes function to regulate behavior.
Results
CRISPR-Mediated Loss of VRI from Clock Neurons Leads to a Short Circadian Period.
Circadian behavioral rhythms are primarily regulated by a molecular program operating in clock neurons. One component of this program is the transcription factor VRI encoded by the essential gene vrille (vri). Homozygous mutants are developmentally lethal (38), and hemizygous mutants are strongly rhythmic but with a shorter free-running circadian period (4). Cycling of VRI in clock neurons is likely important as constant overexpression in clock-expressing cells causes arrhythmicity and a longer circadian period (4). However, an RNAi against vri in clock neurons did not affect circadian behavior (39). We therefore developed a line with CRISPR/Cas9-based 3×-gRNAs targeting the vri gene under UAS-control (UAS-vri-g). When expressed along with the reduced expression UAS-Cas9.P2 (29) in clock neurons and the CLK856-Gal4 driver, there was potent cell type–specific loss of VRI-staining (Fig. 1 A and B and SI Appendix, Fig. S1A). Importantly, the effect was shown to be specific as only VRI levels in Gal4-labeled cells were impacted; VRI levels in the DN3 neurons were unaffected (SI Appendix, Fig. S1A) as expected since most of them are not labeled by the CLK856-Gal4 driver (23). These flies had normal behavior under 12-h light:12-h dark (LD) conditions, but with a small evening peak advance compared to the control strains (SI Appendix, Fig. S1 B and C). Consistent with this phenotype, the flies were completely rhythmic in constant darkness (DD) but with a significantly shorter circadian period (Fig. 1 C–G). These phenotypes are identical to those of a recent vri mutant (40) indicating that they derive from the clock neurons.
Fig. 1.
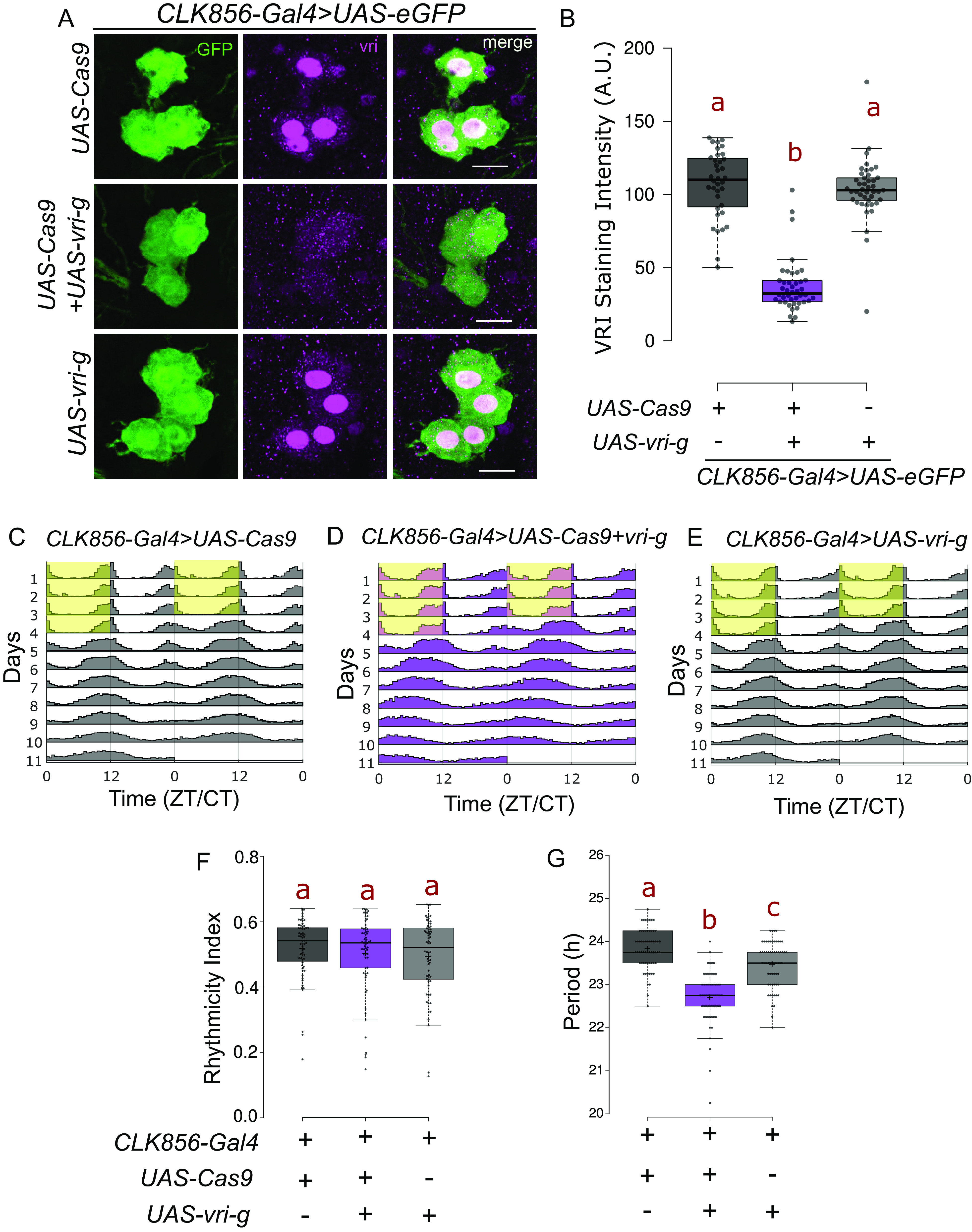
Loss of VRI from clock neurons leads to a shorter circadian period. (A) Representative images of lLNV neurons stained for GFP and VRI at ZT19. There is loss of (nuclear) VRI staining in cells expressing both UAS-Cas9 and UAS-3×-guides against vri (UAS-vri-g). Some nonnuclear staining is visible in both control and mutant cells, which could be due to background reactivity of the antibody. (The scale bar represents 10 µm.) (B) Quantification of VRI staining intensity from lLNv neurons represented as a boxplot, n ≥ 36 cells from at least 12 hemibrains per genotype, letters represent statistically distinct groups; P < 0.001, Kruskal–Wallis test followed by a post hoc Dunn’s test. (C–E) Actograms represent double-plotted average activity of flies from an experiment across multiple days. Yellow panel indicates lights ON. (F) Rhythmicity index for individual flies plotted as a boxplot, n ≥ 62 per genotype from at least two independent experiments. No statistical difference was observed among the groups tested (P = 0.6, Kruskal–Wallis test). (G) Free running period under constant darkness for individual rhythmic flies (RI > 0.25) plotted as a boxplot, letters represent statistically distinct groups; P < 0.01, Kruskal–Wallis test followed by a post hoc Dunn’s test.
CRY Functions Independently in Two Subsets of Clock Neurons.
We next addressed the circadian gene cry, which encodes the photoreceptor Cryptochrome (CRY). Wild-type flies are arrhythmic under constant bright light conditions, cry mutants in contrast remain rhythmic in constant light (12). This classic phenotype is reproduced when cry was mutated with UAS-Cas9.P2 and 3×-gRNA against cry (UAS-cry-g) specifically in clock neurons using the CLK856-Gal4 driver, which also led to loss of nearly all CRY staining from all clock neurons (SI Appendix, Fig. S3). Whereas only about 10 percent of the control flies were rhythmic in constant light (rhythmicity index > 0.25), 90 percent of the cry-mutated flies were rhythmic (Fig. 2 A–C and G and SI Appendix, Table S1). These flies were also rhythmic in constant darkness, like the controls (SI Appendix, Fig. S2) and like traditional cry mutants (12).
Fig. 2.

CRY function in two discrete subsets of clock neurons can regulate rhythmicity under constant light conditions. (A–F) Actograms represent double-plotted average activity of flies from an experiment across multiple days in constant light. Flies expressing both UAS-Cas9 and UAS-3×-guides against cry (UAS-cry-g) in clock neurons are rhythmic in constant light conditions. (G) Rhythmicity index of individual flies quantified for LL2-7 represented by a boxplot, n ≥ 51 per genotype from at least two independent experiments. Letters represent statistically distinct groups; P < 0.001, Kruskal–Wallis test followed by a post hoc Dunn’s test. (H) Period under constant light for individual rhythmic flies (rhythmicity index > 0.25) quantified for LL2-7 represented by a boxplot, letters represent statistically distinct groups; P < 0.01, Kruskal–Wallis test followed by a post hoc Dunn’s test. (I and J) Phase shifts in hours in response to a 60-min light pulse at indicated times (n ≥ 26). Phase difference plotted as means ± SEM of differences from DD3-5. Same control flies are plotted in (I and J).
As CLK856-Gal4 is expressed in most clock neurons, we then asked if the constant light phenotype could be assigned to a more specific subset of circadian neurons. CRISPR-mediated cell type–specific mutagenesis of cry led to loss of CRY staining from specific neurons (SI Appendix, Fig. S3). All PDF neurons are CRY positive (8, 11), but disrupting cry in PDF cells with the Pdf-Gal4 (7) had only a very small effect on constant light arrhythmicity (Fig. 2 D and G and SI Appendix, Table S1). In contrast, flies in which cry was mutated in the cells marked by the dvPdf-Gal4 were fully rhythmic, indistinguishable from CRY loss in all circadian neurons (Fig. 2G and SI Appendix, Table S1). This is interesting because the dvPdf-Gal4 labels only 5 more cells per hemibrain than Pdf-Gal4, and only 2 of these 5 cells are CRY positive: the 5th sLNv and the one ITP-positive LNd (41, 42).
Ongoing work in our lab has characterized two Janelia split-Gal4 drivers, ss00639-GAL4 and ss00773-Gal4. They both specifically label one CRY+ LNd and the 5th sLNv in addition to very few additional cells (SI Appendix, Fig. S4). Expression of CRISPR reagents against cry with the ss00639-Gal4 driver also caused complete rhythmicity in constant light; more than 90% of the flies were rhythmic (Fig. 2 E and G and SI Appendix, Table S1). Similar but less strong results were seen with ss00773-Gal4 (Fig. 2G and SI Appendix, Table S1). A third Gal4 driver, MB122B-Gal4, also expresses in these same two cells along with two additional LNd cells (43, 44), and mutating cry in cells labeled by this driver resulted in about 80% rhythmic flies in constant light (Fig. 2G and SI Appendix, Table S1).
In addition to the lateral cells, CRY is expressed in the DN1p neurons labeled by the CLK4.1-Gal4 driver (19, 45). Mutating cry in cells labeled by this driver also resulted in potent rhythmicity; nearly 75% of these flies were rhythmic in constant light (Fig. 2 F and G and SI Appendix, Table S1). These data taken together identify two sets of circadian neurons that can regulate rhythmicity independently under constant light in the absence of CRY: two evening cells consisting of the 5th sLNv and the single ITP-positive LNd as well as the DN1p neurons.
To address possible mechanisms underlying the different groups of clock neurons and their regulation of constant light rhythmicity, we examined the circadian period of all rhythmic genotypes. Interestingly, cry mutants in all clock neurons have a circadian period of 24.7 ± 0.7 h in constant light, about an hour longer than their constant darkness period of 23.6 ± 0.08 h (Fig. 2H and SI Appendix, Fig. S2E). Mutating CRY in the evening cells with any of the three Gal4s gave rise to a similar period length of greater than 24 h, ~25 h (SI Appendix, Table S1, green boxes in Fig. 2H). Surprisingly, mutating CRY only in the DN1ps caused a dramatically shorter constant light period of ~22 h (SI Appendix, Table S1 and Fig. 2H). These data indicate that specific clock neurons mediate the constant light effect and that there are interesting differences between the two discrete sets of relevant neurons (Discussion).
In addition to being rhythmic under constant light, cry mutants are also deficient in their response to light pulses (46, 47). Control flies experience a delay phase shift in response to a light pulse in the early night at ZT15, an advance phase in response to a light pulse in the late night at ZT21 but little change with a light pulse in the middle of the night at ZT18 (46). The unresponsive cry mutant phenotype was reproduced by loss of CRY function specifically in clock neurons (Fig. 2I). Interestingly, flies with loss of CRY from PDF neurons were also completely deficient in the ZT15 light pulse-mediated phase delay but still maintained some phase advance induced by the ZT21 light pulse. In addition, when CRY function was perturbed in both PDF neurons and evening cells using the dvPDF-Gal4, the phase advance as well as the phase delay response was abolished (Fig. 2I). However, loss of CRY either in the evening cells alone or in the DN1p neurons did not affect the phase response (Fig. 2J). These data further indicate that CRY functions in discrete sets of clock neurons to mediate diverse light responses (Discussion).
PDF Regulates Activity and Sleep from Both the lLNvs and sLNvs.
A key circadian molecule beyond the core clock is the neuropeptide pigment dispersing factor (PDF); it is encoded by the gene Pdf. Null mutants of the gene (Pdf 01) are largely arrhythmic in constant darkness (7). PDF is expressed in only 8 pairs of lateral neurons in the adult brain, four sLNvs and four lLNvs ventral lateral neurons per hemibrain. The lLNvs express very high levels of PDF, which can be easily visualized within their long arborizations. They extend to the optic lobes and to the other side of the brain. The sLNvs extend a long dorsal process, which is also marked by PDF staining [SI Appendix, Fig. S5A, (48)].
A UAS-3×-gRNA line against Pdf (UAS-Pdf-g) in combination with the Pdf-Gal4 driver that labels all PDF-producing neurons caused loss of PDF from the cell bodies and projections of most PDF neurons (Fig. 3 A and B and SI Appendix, Fig. S5B). There was however residual staining in some lLNvs (Fig. 3 A, Bottom and SI Appendix, Fig. S5B), which likely reflects some residual PDF due to its very high levels in these cells prior to the mutagenesis. Nonetheless, this strain was completely arrhythmic in constant darkness whereas the parental control strains had normal rhythms (SI Appendix, Fig. S6 A–C and F). The very few rhythmic flies also had a shorter period phenotype (SI Appendix, Fig. S6I).
Fig. 3.
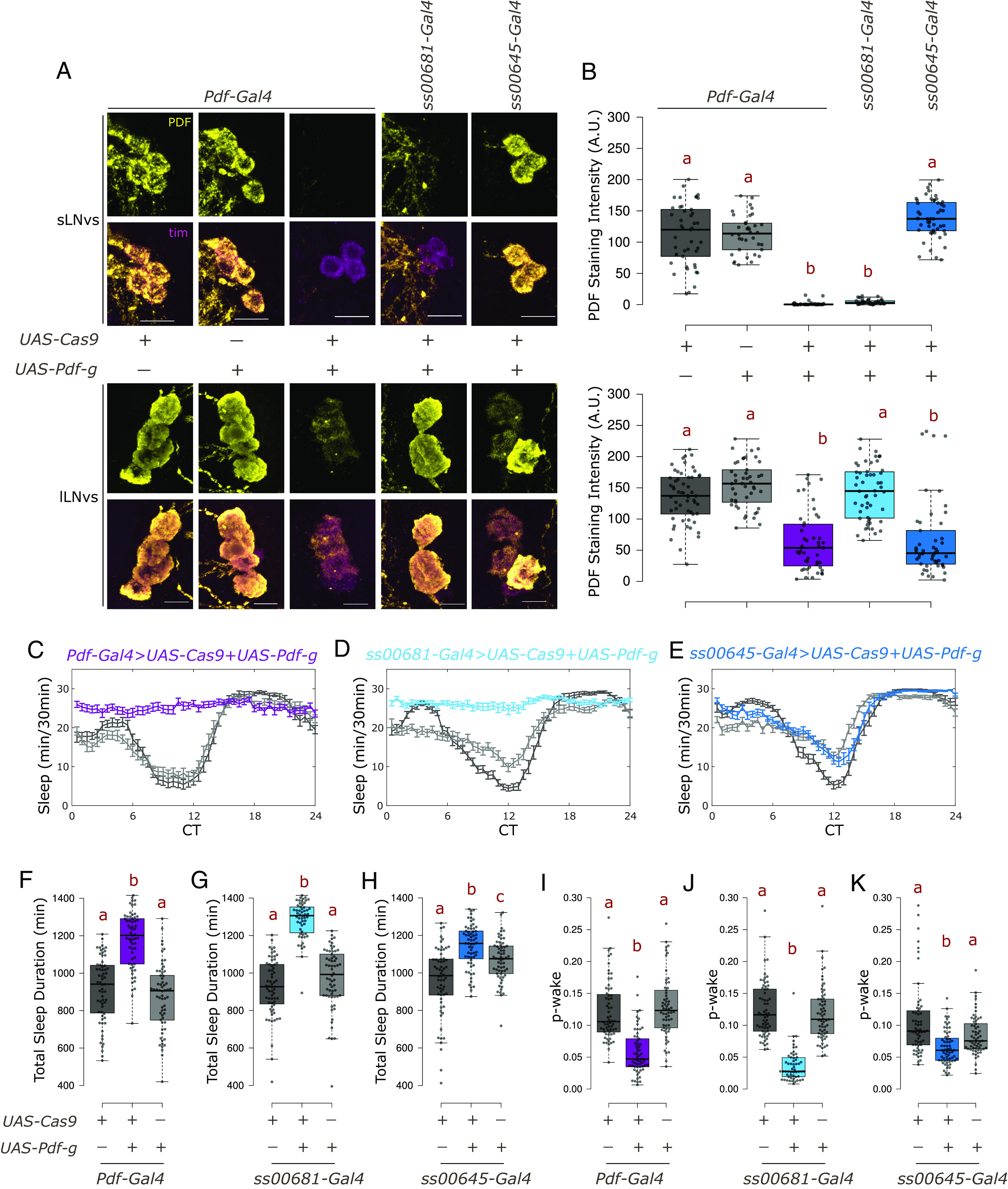
PDF from sLNvs maintains wakefulness under constant darkness. (A) Representative images of sLNvs (Top) and lLNvs (Bottom) stained for TIM and PDF at ZT18. TIM labels all clock neurons. There is loss of PDF staining in cells expressing both UAS-Cas9 and UAS-3×-guides against Pdf (UAS-Pdf-g). (The scale bar represents 10 µm.) (B) Quantification of PDF staining intensity from sLNv (Top) and lLNv (Bottom) neurons represented with boxplots, n ≥ 36 cells from at least 11 hemibrains per genotype. Letters represent statistically distinct groups; P < 0.001, Kruskal–Wallis test followed by a post hoc Dunn’s test. (C–E) Sleep plots representing average sleep of flies from DD1-4 in 30-min bins. UAS-Cas9 alone control is indicated in dark gray, UAS-Pdf-g alone control in light gray whereas the experimental genotype expressing both is represented by color indicated above the plots. (F–H) Total sleep duration in a day for individual flies quantified (from DD1-4) represented by a boxplot, n ≥ 55 per genotype from at least two independent experiments. Letters represent statistically distinct groups, P < 0.01; Kruskal–Wallis test followed by a post hoc Dunn’s test. (I–K) p-wake over a 24-h period for individual flies quantified (from DD1-4) represented by a boxplot, letters represent statistically distinct groups; P < 0.001, Kruskal–Wallis test followed by a post hoc Dunn’s test.
We then used previously described split-Gal4 lines generated by the Rubin lab at Janelia (36) to specifically target only sLNvs or only lLNvs. The CRISPR-Cas9-based method was again highly effective in targeting PDF levels only in the cells targeted by the respective Gal4s. Expression of the gRNA and Cas9 in sLNv-specific driver (ss00681-Gal4) led to near complete loss of PDF staining from 100% of the sLNvs examined and their dorsal processes, with no effect on the lLNv cell bodies or on their processes. Similarly, expressing the CRISPR reagents using the lLNv-specific driver (ss00645-Gal4) significantly reduced PDF expression from lLNv cell bodies and processes, without affecting PDF levels in the sLNvs or their dorsal process (Fig. 3 A and B and SI Appendix, Fig. S5B). However, the effect on the lLNvs was not as strong: about 20% of the neurons (1 to 2 neurons from about half the hemibrains) still had unchanged PDF staining. This is very likely because the ss00645-Gal4 variably labels 3 to 4 lLNvs per hemisphere. Notably however, there remains some residual staining in the lLNvs even with Pdf-Gal4 (Fig. 3A and Fig. 3 B, Bottom), probably due to the higher levels of PDF in the lLNvs (Discussion). As expected based on earlier studies (13), loss of PDF from the sLNvs but not from the lLNvs caused arrhythmicity in constant darkness (SI Appendix, Fig. S6 D–H).
Loss of PDF from both small and large PDF neurons also caused substantially increased sleep under constant darkness (Fig. 3 C and F), and targeting the guides specifically to the small cells caused a similar sleep increase (Fig. 3 D and G). This increase in sleep is not because of a lack of the ability to move as these flies are even more active than the controls when awake (SI Appendix, Table S2). Although a role for PDF neurons in sleep and arousal has been described (6, 49), CRISPR-mediated loss of PDF specifically from the sLNvs here is the most direct evidence that the sLNvs regulate sleep. Loss of PDF from only the large cells also increased sleep but to a much smaller extent (Fig. 3 E and H). Although the difference between the small and large cells may be due to the residual large cell PDF that escapes knockdown, or perhaps to the few lLNvs not labeled by the split-Gal4 driver, the very significant reduction in PDF signal from lLNv cell bodies and processes in most brains (Fig. 3 A and B, Bottom and SI Appendix, Fig. S5B) makes it more likely that the lLNvs have a smaller role in regulating sleep than the sLNvs in constant darkness.
An analysis of sleep structure indicated a substantial decrease in p-wake (the instantaneous probability that the flies will awaken from sleep), but no effect on p-doze [the instantaneous probability that awake flies will fall asleep (50)] (Fig. 3 I–K and SI Appendix, Table S2). These data are consistent with a wake-promoting role of PDF (6) and identify PDF from the sLNvs as the primary regulator.
Temporally Regulated CRISPR-Cas9 Mutagenesis Assigns PDF Function to Adult Neurons.
The function of several circadian genes has not yet been definitively assigned to the adult fly, so some of them might have a substantial developmental role. Traditional methods of temporal regulation of Gal4 activity such as GeneSwitch (51) or the temperature-sensitive Gal80 system (52) are not ideal for behavior experiments as they either involve toxic drugs or dramatic shifts in temperature, both of which severely impact the standard locomotor behavior assay (53, 54). Pairing with the temperature sensitive Gal80 system also resulted in leaky mutagenesis (33). We therefore assessed two more recently developed methods, one specific to CRISPR and the other more general, to mutagenize genes of interest adult-specifically.
The first was the inducible Cas9 method developed by Port et al. (33). It is a UAS-based method: The Cas9 sequence is preceded by a Green Fluorescent Protein (GFP) cassette, which is followed by a STOP codon flanked by FRT sites (hereafter called STOP-Cas9). Pairing with a heat-shock induced flp transgene allows for temporal control of mutagenesis as Cas9 expression occurs only post heat shock (SI Appendix, Fig. S7A). We adapted this method for adult-specific induction by using three consecutive heat shocks (referred to below as “heat shock”; see Materials and Methods). Since these heat shocks are temporally separated by several days from the behavior analyses, the heat shocks are not expected to alter circadian behavior. The STOP-Cas9 transgene was combined with Pdf-Gal4, UAS-Pdf-g, a UAS-mRFP transgene to label the PDF cells as well as a heat shock-induced flp encoding hsFLPG5 (SI Appendix, Fig. S7A).
In control flies without UAS-Pdf-g, heat shock did not affect the levels of PDF (Fig. 4 A and B and SI Appendix, Fig. S7B) and did not negatively impact rhythmic behavior (Fig. 4 C and D). Importantly, flies expressing all the transgenes including the UAS-gRNA but without heat shock showed no loss of PDF signal (Fig. 4 A and B and SI Appendix, Fig. S7B) and no defects in behavior (Fig. 4 C–F); this indicates no leaky expression without heat shock. In the same flies two weeks post heat shock, PDF staining was lost from all sLNvs in ~60% of the hemibrains analyzed, whereas some mosaicism was observed in the remaining 40% of the hemibrains; 1 to 2 neurons still expressed both GFP and PDF (Fig. 4 A and B and SI Appendix, Fig. S7B). A similar fraction of the heat shocked mutant flies, about 38 percent, were rhythmic in constant darkness (Fig. 4D and SI Appendix, Fig. S7C), indicating an adult-specific requirement of PDF for rhythmicity. The ca. 60 percent arrhythmic flies also showed an adult-specific effect of PDF on sleep (Fig. 4E).
Fig. 4.
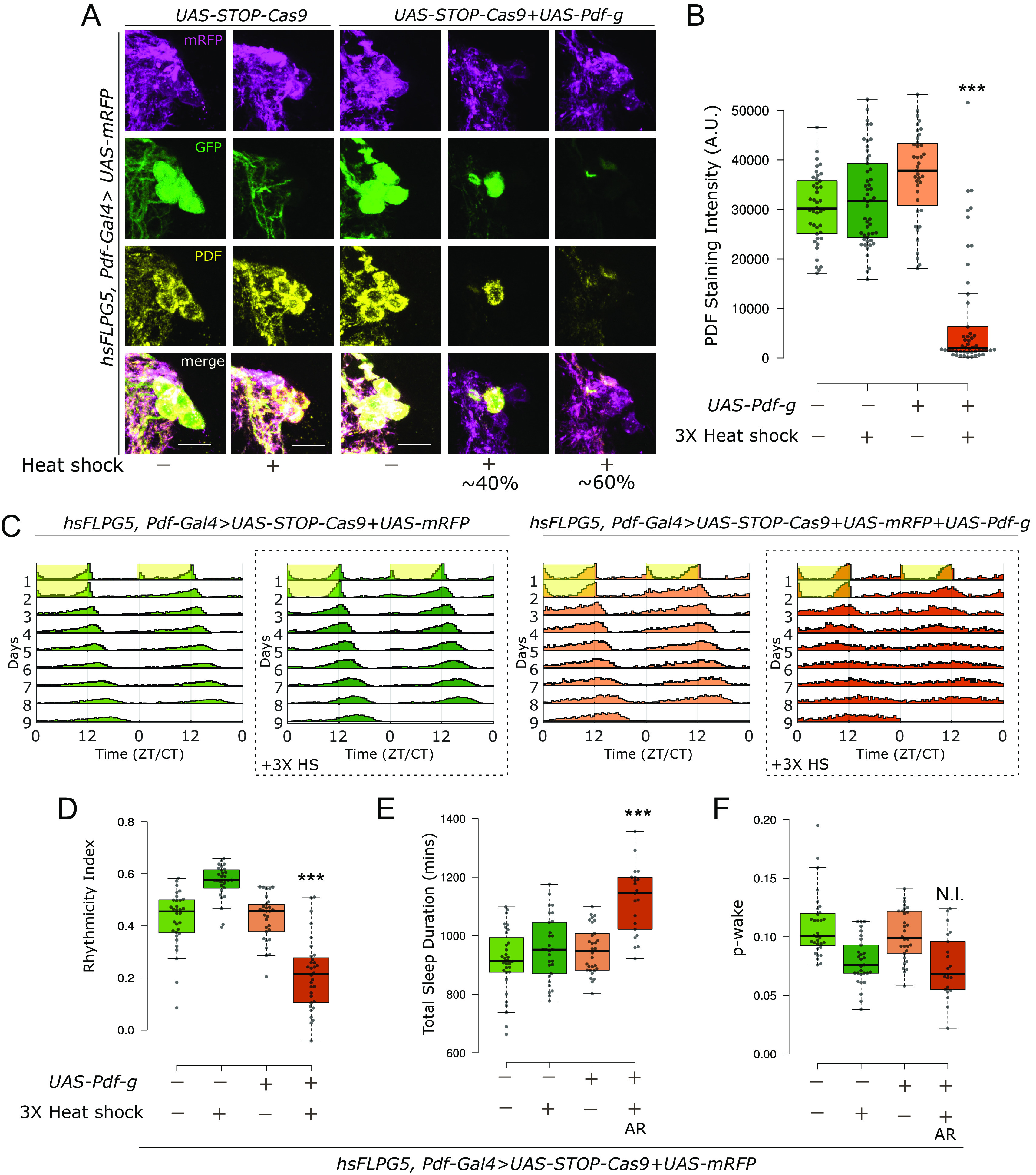
Adult-specific loss of PDF using an inducible Cas9 system results in loss of rhythmicity and increased sleep. (A) Representative images of sLNvs stained for GFP, mRFP and PDF. 2 wk post heat shock, there is loss of PDF staining in cells expressing both UAS-Cas9 and UAS-Pdf-g. In about 60% of the hemibrains analyzed, PDF staining was lost from all sLNvs, whereas in the remaining 40% hemibrains, some mosaicism was observed as PDF staining was unchanged in 1 to 2 sLNvs. (The scale bar represents 10 µm.) (B) Quantification of PDF staining intensity from sLNvs represented with boxplots, n ≥ 39 cells from at least 12 hemibrains per genotype/condition. There is a significant effect of both variables on the staining intensity and a significant interaction when CRISPR reagents for inducible CRISPR for Pdf were combined with 3× heat shock (Two-way ANOVA; *** Interaction P < 0.001). (C) Actograms represent double-plotted average activity of flies from an experiment across multiple days. Yellow panel indicates lights ON. Panels in dotted boxes indicate heat-shocked flies. (D) Rhythmicity index for individual flies quantified (from DD2-7) represented by a boxplot, n ≥ 29 per genotype. There is a significant effect of both variables on rhythmicity and a significant interaction when CRISPR reagents for inducible CRISPR for Pdf were combined with 3× heat shock (Two-way ANOVA; *** Interaction P < 0.001). (E) Total sleep duration in a day for individual flies quantified for DD1-4 represented by a boxplot. For the group expressing both UAS-Cas9 and UAS-Pdf-g with heat-shock, only data from the arrhythmic flies are shown and used for statistics (indicated by AR). Also see SI Appendix, Fig. S5D. There is a significant effect of both variables on sleep duration and a significant interaction when CRISPR reagents for inducible CRISPR for Pdf were combined with 3× heat shock (Two-way ANOVA; *** Interaction P < 0.001). (F) p-wake over a 24-h period for individual flies quantified (from DD1-4) represented by a boxplot. For the group expressing both UAS-Cas9 and UAS-Pdf-g with heat-shock, only data from the arrhythmic flies are shown and used for statistics (indicated by AR). Also see SI Appendix, Fig. S5D. Only heat-shock had a significant effect on p-wake and there was no interaction between the variables (Two-way ANOVA; N.I. = No Interaction, p = 0.31).
The remaining rhythmic flies likely reflect mosaicism, as PDF from 1 to 2 neurons might be sufficient to maintain some level of rhythmicity. This is supported by the fact that the rhythmicity and sleep phenotypes were correlated in the mutant flies (SI Appendix, Fig. S7D). Surprisingly, heat shock alone had a strong effect on the sleep structure and lowered p-wake significantly in the controls. Therefore, no conclusions could be drawn about the adult-specific effect of PDF in maintaining p-wake (Fig. 4F). These data indicate that the heat-shock-based methods are compatible with studying circadian behavior but may be less suitable for studying sleep behavior.
We also combined CRISPR-Cas9 mutagenesis with a more general method of temporal regulation that affects the Gal4 and therefore controls all UAS-based strategies, the recently developed auxin-inducible gene expression system [AGES, (55)]. It uses the traditional Gal4 repressor Gal80, which has been engineered to be regulated by auxin. Auxin is nontoxic and does not affect fly lifespan at the required concentrations (55). We combined the AGES system with the regular Cas9, UAS-Cas9.P2, along with UAS-Pdf-g and UAS-mRFP to label the LNvs with the Pdf-Gal4 driver (SI Appendix, Fig. S8A).
AGES sequesters GAL4 and thus inhibits expression from all UAS-transgenes without auxin feeding [SI Appendix, Fig. S8A, (55)]. Indeed, there is no to very low mRFP expression in cell bodies or processes without auxin feeding (Fig. 5A and SI Appendix, Fig. S8B). However, expression must be nonzero, as some PDF mutagenesis is observed without auxin feeding (~twofold reduction in PDF levels, Fig. 5 A and B). Not surprisingly, auxin feeding leads to a much bigger effect—near loss of all PDF from cell bodies (>10-fold reduction) as well as processes (Fig. 5 A and B and SI Appendix, Fig. S8B).
Fig. 5.
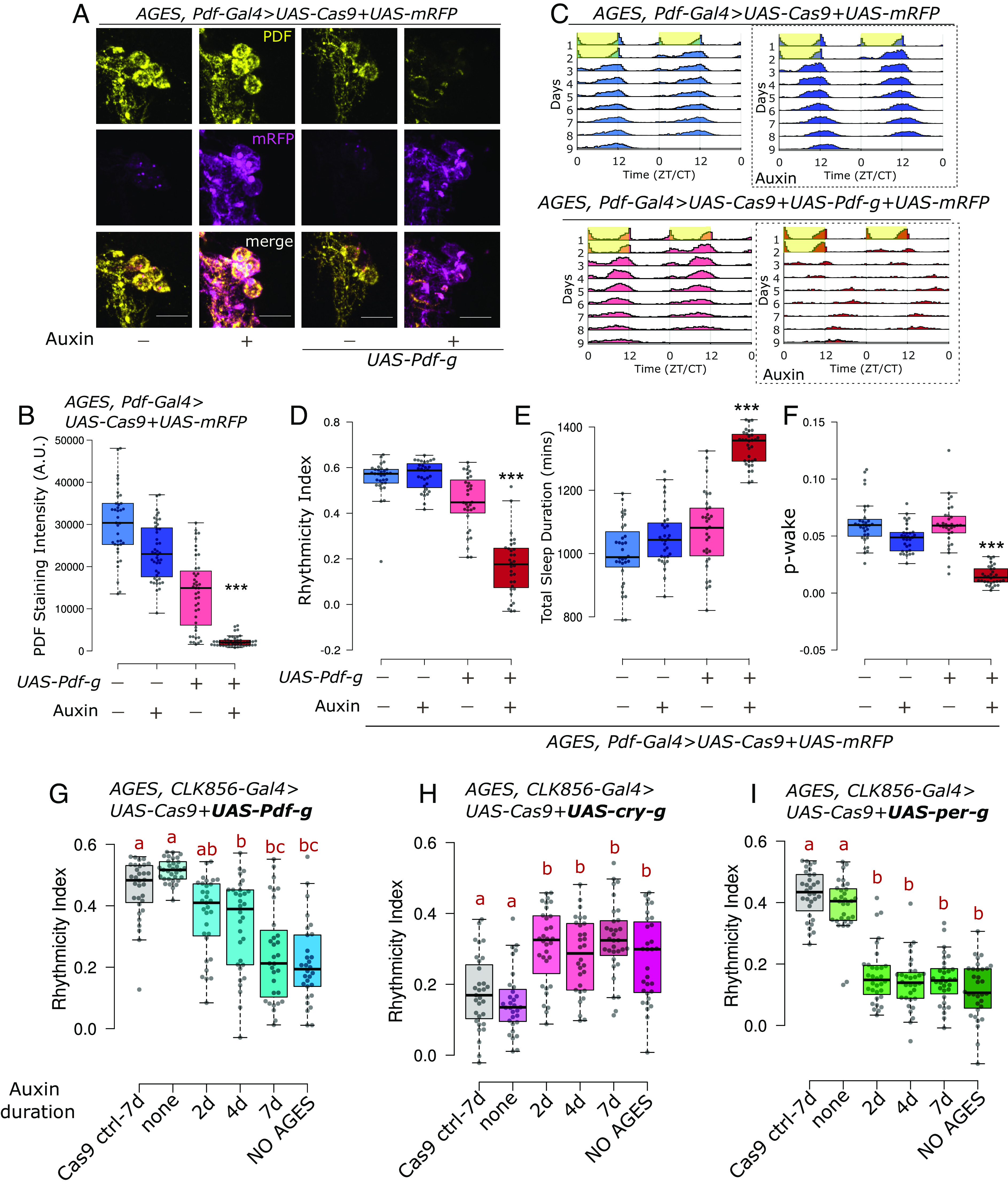
Adult-specific loss of PDF with AGES leads to loss of rhythmicity and wakefulness. (A) Representative images of sLNvs stained for mRFP and PDF. Upon feeding 10 mM auxin for 2 wk, there is loss of PDF staining in cells expressing both UAS-Cas9 and UAS-Pdf-g. (The scale bar represents 10 µm.) (B) Quantification of PDF staining intensity from sLNvs is represented with boxplots, n ≥ 39 cells from at least 11 hemibrains per genotype/condition. There is a significant effect of both variables on PDF staining intensity and a significant interaction when AGES and CRISPR reagents for Pdf were combined with auxin feeding (Two-way ANOVA; *** Interaction P < 0.001). (C) Actograms represent double-plotted average activity of flies from an experiment across multiple days. Yellow panels indicate lights ON. Panels in dotted boxes indicate auxin feeding. (D) Rhythmicity index for individual flies quantified (from DD2-7) represented by a boxplot, n ≥ 31 per genotype. There is a significant effect of both variables on rhythmicity and a significant interaction when AGES and CRISPR reagents for Pdf were combined with auxin feeding (Two-way ANOVA; *** Interaction P < 0.001). (E) Total sleep duration in a day for individual flies quantified (from DD1-4) represented by a boxplot. There is a significant effect of both variables on sleep duration and a significant interaction when AGES and CRISPR reagents for Pdf were combined with auxin feeding (Two-way ANOVA; *** Interaction P < 0.001). (F) p-wake over a 24-h period for individual flies quantified (from DD1-4) represented by a boxplot. There is a significant effect of both variables on p-wake and a significant interaction when AGES and CRISPR reagents for Pdf were combined with auxin feeding (Two-way ANOVA; *** Interaction P < 0.001). (G–I) Rhythmicity index for individual flies quantified (from DD2-6) represented by a boxplot, n ≥ 30 per genotype. Flies of the indicated genotypes were fed auxin for either 2, 4 or 7 d and compared with flies that were fed no auxin, or with flies with mutagenesis throughout development (NO AGES). Letters represent statistically distinct groups; Kruskal–Wallis test followed by a post hoc Dunn’s test, P < 0.003.
The small-scale leaky mutagenesis without auxin feeding had only a minor effect on rhythmicity and sleep (Fig. 5 C–E). However, auxin feeding caused a very strong effect on both rhythmicity and sleep just like the effect on PDF staining (Fig. 5 D and E and SI Appendix, Fig. S8C), and adult-specific loss of PDF lowered p-wake (Fig. 5F). Auxin feeding alone in control flies had a very minor effect on PDF staining (20% reduction in cell bodies) and no effect on circadian behavior and sleep duration (Fig. 5 A–E and SI Appendix, Fig. S8 B and C). There was also a significant interaction between auxin feeding and expression of CRISPR reagents against PDF on p-wake (P < 0.001, Two-way ANOVA; Fig. 5F). Combining AGES with the CRISPR system therefore indicate that most if not all aspects of PDF function on circadian behavior and sleep are adult specific.
We then assayed the time required for adult-specific-CRISPR-mediated mutagenesis. Flies expressing UAS-Cas9.P2 and UAS-Pdf-g along with AGES were fed 10 mM auxin for 2, 4, and 7 d before testing for behavior on food without auxin. Two days of auxin feeding did not significantly alter rhythmicity of the controls, but there was a gradual reduction in rhythmicity from 4 to 7 d of auxin feeding (Fig. 5G). To further tease apart the required time for CRISPR mutagenesis from the effects of protein turnover, we used the same paradigm to test two proteins that undergo daily degradation—CRY and PER. For both proteins, 2 d of auxin feeding was sufficient to cause a phenotype indistinguishable from the perturbations through development (Fig. 5 H and I), indicating that no more than 2 d are required for mutagenesis.
Discussion
In this study we generated UAS-3×-gRNA lines for neuron-specific perturbation of three key circadian genes and combined them with defined Gal4 drivers to assay their function in specific subsets of clock neurons. We could reproduce the mutant phenotypes of vri, cry, and Pdf in known neurons and additionally identified some cell type–specific functions of these genes. The approach worked very well with specific and weak Gal4 drivers, which allowed the identification of two subsets of circadian neurons, within which CRY acts independently to regulate rhythmicity under constant light conditions. Finally, these reagents were compatible with existing tools and shown to be highly effective with two independent methods for temporal regulation. The results and lines described here will be of use to the circadian community, but we hope that the detailed validation of the method will appeal to a much wider population of fly neuroscientists and encourage them to switch from the standard but less dependable RNAi methodology to this more effective and consistent method for the spatial and temporal knockout of genes in adult fly neurons.
Our data show that this cell type–specific CRISPR method works well with highly expressed genes like Pdf and even with weak split-Gal4 drivers. It is also ideal for addressing the adult function of essential genes such as vri, by perturbing their expression levels and function in inessential cells as we show here for circadian neurons. In this context, the functions of many Drosophila genes are defined only in developmental contexts, even though these same genes are expressed again in adults. For example, we recently found that genes encoding GPCRs, transcription factors, and cell surface molecules are broadly yet specifically expressed in clock and dopaminergic neurons. Studying their function in these specific adult neurons is beginning to help define the molecular underpinnings of neural circuits and behavior (23, 56). Toward these goals, we have generated and begun to use a library of gRNA lines against all GPCRs expressed in Drosophila (37). Moreover, several lines with 2×-gRNA generated by Port and Boutros (33) are now publicly available. Such resources should also make this method a good choice for tissue and cell type–specific reverse genetic screens.
Combining multiple RNAi lines is not common, perhaps because of the suspected dilution effects of multiple UAS lines. In contrast, this CRISPR-Cas9 method does not require high expression levels, so combining gRNAs against multiple genes, for example to study enhancer and suppressor phenotypes, should be possible and would extend the approach in an interesting direction.
Another Application of This Method Should Be Cell-Specific Gene-Interaction Studies. VRI operates as a part of the molecular transcription–translation feedback loop that operates in clock neurons (57). Hemizygous mutants of vri are rhythmic with a short circadian period (4). Concurrently, a recent study generated a new homozygous mutant that has normal rhythms with a short circadian period despite dramatically reduced VRI levels in the clock neurons (40). These results are in complete agreement with ours (Fig. 1 C–G). It is also possible that very low levels of VRI in clock neurons is sufficient to maintain rhythms like in the whole-body mutant (40). Although unlikely in our view, this interpretation is consistent with an earlier study (58) that used cell-specific flip outs of a rescue transgene in a mutant vri background and found that VRI in PDF neurons regulates rhythmicity. Although overexpression of VRI also results in considerable loss of rhythms (4), simple loss of function via these CRISPR-Cas9-based methods is less prone to misinterpretation than gain-of-function experiments. In the future, guides can be targeted to specific subsets of circadian and non-circadian cells to further define the function of VRI in the regulation of development and behavior.
The CRY photoreceptor along with the compound eyes and H-B eyelets are the three pathways through which flies entrain to environmental light cues (59). It is therefore not surprising that cry mutants are still able to light entrain normally. However, these mutants are rhythmic under constant bright light conditions, indicating a special role for CRY under prolonged light exposure conditions (12). CRY is expressed in several different clock neurons; they include the PDF neurons, a subset of evening cells (5th sLNv+3 LNds) and a subset of the DN1ps (8). Interestingly, we find that CRY only functions in the evening cells and the DN1ps to regulate the response to constant light. This conclusion agrees with previous results indicating that evening cells (60, 61) and dorsal neurons (60, 62) are the principal drivers of rhythmicity under constant light conditions. QUASIMODO (QSM) is another protein that mediates light input to the circadian clock, and flies mutant for QSM in dorsal neurons maintain rhythmicity in constant light (63), further emphasizing the role of dorsal neurons in mediating rhythmicity under constant light conditions.
Our results also agree with earlier results that PDF-positive LNvs have no more than a minor role in responding to constant light (61, 62). Because the sLNvs are directly downstream of the light responsive H-B eyelets (64), these neurons may be constantly electrically stimulated and their molecular clock therefore poisoned by constant light even without CRY. In contrast, loss of CRY in either of the two other major subsets of CRY-positive neurons, the two evening cells (5th sLNv, CRY+ and ITP+ LNd) as well as the DN1ps, is sufficient to maintain rhythmicity in constant light. These data indicate that CRY may be the primary source of light information in these neurons; perhaps they are critical for adapting to altered photoperiods such as longer summer days.
Interestingly, CRY functions in a different subset of clock neurons, the PDF neurons, to effect light pulse-mediated phase delays in the early night. Phase delays induced by a light pulse in the late night also requires CRY in PDF neurons but additionally requires CRY in evening neurons (Fig. 2I). A CRY requirement for phase responses in PDF neurons has been previously described (65), and a role for the evening neurons in phase response behavior has also been suggested (60). As neuronal activation of PDF neurons at different times of day can mimic a light-mediated phase response curve in a CRY-independent manner (66), CRY photon capture may impact neuronal firing, a result consistent with direct observation (67), so altered electrical properties of the CRY mutants might also contribute to the observed light-mediated phenotypes.
Expression of UAS-Cas9 and UAS-Pdf-g with Pdf-Gal4 resulted in complete loss of PDF from sLNvs but only partial loss from the lLNvs (Fig. 3 A and B). The higher levels of PDF in lLNvs than in sLNvs is one possible explanation, but persistent PDF in lLNvs might also reflect the duration of expression of the CRISPR tools in the different cells. The sLNvs have an early larval origin, well before the lLNvs appear in the mid-late pupal stage (48). This temporal pattern is recapitulated by Pdf-Gal4 (7), allowing sLNvs a much longer time for mutagenesis and subsequent protein turnover.
PDF is a key circadian neuropeptide in Drosophila, and the small PDF neurons (sLNvs) are critical for circadian behavior (14, 15). This is because PDF from the sLNvs is a key regulator of rhythmicity in constant darkness (13). The results here show that loss of PDF from these neurons also has a major effect on sleep structure, namely, a decrease in p-wake. Loss of PDF from the lLNvs has a qualitatively similar but much smaller quantitative decrease in p-wake (Fig. 3 I–K). This indicates that PDF from both sources is wake-promoting, consistent with the fact that activation of lLNvs mediates arousal (18, 49). A greater role for lLNvs has been described in promoting wake through GABA signaling under light–dark conditions (68, 69). It is therefore possible that the lLNvs play a larger role in maintaining wakefulness under LD conditions, whereas rhythmic PDF release from the sLNvs in constant darkness promotes wake as well as maintains rhythmicity.
Two independent methods indicate that PDF functions adult specifically to regulate rhythmicity and sleep. Although both worked effectively, the two strategies have distinguishing features. The inducible Cas9 system was very robust, i.e., there was no leaky expression, which provides high confidence in the adult-specific conclusion. However, the induction was incomplete, leading to some mosaicism and considerable variability in the resulting phenotypes (SI Appendix, Fig. S7A). Perhaps this could be solved by further altering the heat shock regimen, especially for rhythmicity as this assay is insensitive to the brief heat shock. Effects on sleep were more problematic, as even sleep in the control flies was impacted by heat shock (Fig. 4F). It is presently uncertain if the effect of heat shock alone on sleep structure is strain-specific or a more general effect.
In contrast to heat shock, the AGES system was highly efficient but exhibited some background mutagenesis in the absence of auxin at least with the PDF-Gal4 driver. As the AGES system was highly effective at inhibiting expression of a fluorescent protein marker (Fig. 5A and SI Appendix, Fig. S8B), the background mutagenesis likely reflects the ability of even very low levels of Cas9 and gRNA expression to induce mutagenesis as well as the sensitivity of this assay. The lack of a similar leaky phenotype with the CLK856-Gal4 driver (Fig. 5G) suggests that it is Gal4-specific. Perhaps the CLK856-Gal4 driver only labels the PDF neurons at a later developmental stage, allowing less time to affect the leaky phenotype. Although mutagenesis sensitivity is a potential liability in this specific context, it is a generally positive feature of the CRISPR-based system: it does not require that drivers be highly expressed to mediate efficient mutagenesis. In the context of AGES, the use of lower expression Cas9 variants (33) might circumvent this issue. Importantly, auxin exposure alone had no detectable effect on circadian or sleep behavior (Fig. 5 E and F). Thus, we find auxin exposure to be a less disruptive perturbation than heat shock and hence a mode of CRISPR induction more compatible for behavior analyses. Moreover, temporal regulation via AGES allows combining with other UAS transgenes for experiments like complementation analysis. Although the current version of AGES is incompatible with the many Gal80-insensitive split-Gal4 lines (25), it can be used with new Gal80-sensitive split-Gal4 lines (70). In summary, both temporal methods are effective, and we hope this exploration of their differences will help researchers choose the one best suited to their needs.
Materials and Methods
Fly Stocks and Rearing.
All flies were raised on a standard cornmeal media supplemented with yeast in a temperature-controlled incubator at 25 °C in 12:12 Light:Dark cycles. The genotypes of all fly strains used in this study are listed in SI Appendix, Table S3. Details on fly lines generated in this study are described in SI Appendix, Materials and Methods, Generation of Fly Lines.
Locomotor Activity and Sleep Behavior.
For behavior analysis of flies with throughout (not adult-specific) perturbations, 0 to 3-d-old male flies of the appropriate genotype were collected and aged till they were about 2 wk old at 25 °C in 12:12 LD, with food changes every 3 to 4 d. The flies were then loaded into behavior tubes containing food (4% sucrose and 2% agar) and loaded onto Drosophila Activity Monitors (DAM; TriKinetics Inc., Waltham, MA). These DAMs were placed in light boxes with programmable Light-emitting Diode (LED) intensities inside a temperature-controlled incubator set to 25 °C. Flies were entrained for 2 to 3 d with 12-h:12-h lights ON (500 lx): lights OFF before being switched to either constant darkness or constant light (500 lx). Circadian rhythmicity and sleep analysis was performed using the 2020 version of Sleep and Circadian Analysis MATLAB Program (SCAMP) developed by Christopher G. Vecsey (71).Details of locomotor behavior analysis are described in SI Appendix, Materials and Methods, Locomotor Activity and Sleep Behavior.
Immunohistochemistry.
Whole flies of the same genotype and age as the behavior experiment cohort were fixed in 4% paraformaldehyde. Dissected brains were washed and blocked with 10% Normal Goat Serum (Jackson labs). The following primary antibodies were used diluted in blocking buffer– chicken anti-GFP (Abcam ab13970; 1:1,000), mouse anti-PDF (DSHB-PDF C7; 1:1,000), guinea pig anti-VRI [gift of Paul Hardin; 1:3,000 (72)], rat anti-TIM (Rosbash lab; 1:250), rabbit anti-dsRed (Takara Bio- 632393; 1:300), rabbit anti-PER (Rosbash lab, 1:1,000), rat anti-CRY (Rosbash lab, 1:200) and rat anti-RFP (Proteintech-5f8; 1:1,000). Further details on image acquisition and analysis can be found in SI Appendix, Materials and Methods, Immunohistochemistry and Image Analysis.
See also, SI Appendix, Materials and Methods, Data Representation and Statistics.
Supplementary Material
Appendix 01 (PDF)
Dataset S01 (XLSX)
Acknowledgments
We thank Michelle Lin for assistance with maintaining fly stocks. We thank Matthias Schlichting and all members of the Rosbash lab for useful discussions. We thank Paul Garrity, Justin Blau and Katharine Abruzzi for helpful comments on the manuscript. We thank Paul Hardin for the VRI antibody. We thank Heather Dionne, Aljoscha Nern and Gerry Rubin (Janelia Research campus) for the split-Gal4 lines, and Filip Port and Michael Boutros for the STOP-Cas9 flies. Stocks obtained from the Bloomington Drosophila Stock Center (NIH P40OD018537) and Vienna Drosophila Resource Center were used in this study. This work was supported by the Howard Hughes Medical Institute.
Author contributions
S.R. and M.R. designed research; S.R., D.S., and J.Q.L. performed research; S.R. and J.Q.L. contributed new reagents/analytic tools; S.R. and D.S. analyzed data; J.Q.L. edited the paper; and S.R. and M.R. wrote the paper.
Competing interests
The authors declare no competing interest.
Footnotes
Preprint: Biorxiv (https://www.biorxiv.org/content/10.1101/2023.02.23.529279v2).
This article is a PNAS Direct Submission.
Data, Materials, and Software Availability
All data are included in the article and/or supporting information.
Supporting Information
References
- 1.Allada R., Chung B. Y., Circadian organization of behavior and physiology in Drosophila. Annu. Rev. Physiol. 72, 605–624 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hardin P. E., Molecular genetic analysis of circadian timekeeping in Drosophila. Adv. Genet. 74, 141–173 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Helfrich-Förster C., The neuroarchitecture of the circadian clock in the brain of Drosophila melanogaster. Microsc. Res. Tech. 62, 94–102 (2003). [DOI] [PubMed] [Google Scholar]
- 4.Blau J., Young M. W., Cycling vrille expression is required for a functional Drosophila clock. Cell 99, 661–671 (1999). [DOI] [PubMed] [Google Scholar]
- 5.Benito J., Zheng H., Hardin P. E., PDP1ε functions downstream of the circadian oscillator to mediate behavioral rhythms. J. Neurosci. 27, 2539–2547 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Parisky K. M., et al. , PDF cells are a GABA-responsive wake-promoting component of the Drosophila sleep circuit. Neuron 60, 672–682 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Renn S. C., Park J. H., Rosbash M., Hall J. C., Taghert P. H., A pdf neuropeptide gene mutation and ablation of PDF neurons each cause severe abnormalities of behavioral circadian rhythms in Drosophila. Cell 99, 791–802 (1999). [DOI] [PubMed] [Google Scholar]
- 8.Benito J., Houl J. H., Roman G. W., Hardin P. E., The blue light photoreceptor CRYPTOCHROME is expressed in a subset of circadian oscillator neurons in the Drosophila CNS. J. Biol. Rhythms 23, 296–307 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Emery P., So W. V., Kaneko M., Hall J. C., Rosbash M., CRY, a Drosophila clock and light-regulated cryptochrome, is a major contributor to circadian rhythm resetting and photosensitivity. Cell 95, 669–679 (1998). [DOI] [PubMed] [Google Scholar]
- 10.Stanewsky R., et al. , The cryb mutation identifies cryptochrome as a circadian photoreceptor in Drosophila. Cell 95, 681–692 (1998). [DOI] [PubMed] [Google Scholar]
- 11.Yoshii T., Todo T., Wülbeck C., Stanewsky R., Helfrich-Förster C., Cryptochrome is present in the compound eyes and a subset of Drosophila’s clock neurons. J. Comp. Neurol. 508, 952–966 (2008). [DOI] [PubMed] [Google Scholar]
- 12.Emery P., Stanewsky R., Hall J. C., Rosbash M., A unique circadian-rhythm photoreceptor. Nature 404, 456–457 (2000). [DOI] [PubMed] [Google Scholar]
- 13.Shafer O. T., Taghert P. H., RNA-interference knockdown of Drosophila pigment dispersing factor in neuronal subsets: The anatomical basis of a neuropeptide’s circadian functions. PLOS One 4, e8298 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Grima B., Chélot E., Xia R., Rouyer F., Morning and evening peaks of activity rely on different clock neurons of the Drosophila brain. Nature 431, 869–873 (2004). [DOI] [PubMed] [Google Scholar]
- 15.Stoleru D., Peng Y., Agosto J., Rosbash M., Coupled oscillators control morning and evening locomotor behaviour of Drosophila. Nature 431, 862–868 (2004). [DOI] [PubMed] [Google Scholar]
- 16.Yao Z., Shafer O. T., The Drosophila circadian clock is a variably coupled network of multiple peptidergic units. Science 343, 1516–1520 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Dubowy C., Sehgal A., Circadian rhythms and sleep in Drosophila melanogaster. Genetics 205, 1373–1397 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Shang Y., Griffith L. C., Rosbash M., Light-arousal and circadian photoreception circuits intersect at the large PDF cells of the Drosophila brain. Proc. Natl. Acad. Sci. U.S.A. 105, 19587–19594 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zhang L., et al. , DN1p circadian neurons coordinate acute light and PDF inputs to produce robust daily behavior in Drosophila. Curr. Biol. 20, 591–599 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Guo F., et al. , Circadian neuron feedback controls the Drosophila sleep–activity profile. Nature 536, 292–297 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Guo F., Holla M., Díaz M. M., Rosbash M., A circadian output circuit controls sleep-wake arousal in Drosophila. Neuron 100, 624–635.e4 (2018). [DOI] [PubMed] [Google Scholar]
- 22.King A. N., Sehgal A., Molecular and circuit mechanisms mediating circadian clock output in the Drosophila brain. Eur. J. Neurosci. 51, 268–281 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ma D., et al. , A transcriptomic taxonomy of Drosophila circadian neurons around the clock. ELife 10, e63056 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Brand A. H., Perrimon N., Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Dev. Camb. Engl. 118, 401–415 (1993). [DOI] [PubMed] [Google Scholar]
- 25.Luan H., Peabody N. C., Vinson C. R., White B. H., Refined spatial manipulation of neuronal function by combinatorial restriction of transgene expression. Neuron 52, 425–436 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Meissner G. W., et al. , A searchable image resource of Drosophila GAL4-driver expression patterns with single neuron resolution. Elife 12, e80660 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sekiguchi M., Inoue K., Yang T., Luo D.-G., Yoshii T., A catalog of GAL4 drivers for labeling and manipulating circadian clock neurons in Drosophila melanogaster. J. Biol. Rhythms 35, 207–213 (2020). [DOI] [PubMed] [Google Scholar]
- 28.Dietzl G., et al. , A genome-wide transgenic RNAi library for conditional gene inactivation in Drosophila. Nature 448, 151–156 (2007). [DOI] [PubMed] [Google Scholar]
- 29.Meltzer H., et al. , Tissue-specific (ts)CRISPR as an efficient strategy for in vivo screening in Drosophila. Nat. Commun. 10, 2113 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Port F., Chen H.-M., Lee T., Bullock S. L., Optimized CRISPR/Cas tools for efficient germline and somatic genome engineering in Drosophila. Proc. Natl. Acad. Sci. U.S.A. 111, E2967–E2976 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Port F., Boutros M., Tissue-specific CRISPR-Cas9 screening in Drosophila. Methods Mol. Biol. 2540, 157–176 (2022). [DOI] [PubMed] [Google Scholar]
- 32.Jinek M., et al. , A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science 337, 816–821 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Port F., et al. , A large-scale resource for tissue-specific CRISPR mutagenesis in Drosophila. eLife 9, e53865 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Port F., Bullock S. L., Augmenting CRISPR applications in Drosophila with tRNA-flanked sgRNAs. Nat. Methods 13, 852–854 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Delventhal R., et al. , Dissection of central clock function in Drosophila through cell-specific CRISPR-mediated clock gene disruption. ELife 8, e48308 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Schlichting M., Díaz M. M., Xin J., Rosbash M., Neuron-specific knockouts indicate the importance of network communication to Drosophila rhythmicity. ELife 8, e48301 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Schlichting M., et al. , Dopamine and GPCR-mediated modulation of DN1 clock neurons gates the circadian timing of sleep. Proc. Natl. Acad. Sci. U.S.A. 119, e2206066119 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.George H., Terracol R., The vrille gene of Drosophila is a maternal enhancer of decapentaplegic and encodes a new member of the bZIP family of transcription factors. Genetics 146, 1345–1363 (1997). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Matsumoto A., et al. , A functional genomics strategy reveals clockwork orange as a transcriptional regulator in the Drosophila circadian clock. Genes Dev. 21, 1687–1700 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Gunawardhana K. L., Rivas G. B. S., Caster C., Hardin P. E., Crosstalk between vrille transcripts, proteins, and regulatory elements controlling circadian rhythms and development in Drosophila. iScience 24, 101893 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Bahn J. H., Lee G., Park J. H., Comparative analysis of Pdf-mediated circadian behaviors between Drosophila melanogaster and D. virilis. Genetics 181, 965–975 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Schubert F. K., Hagedorn N., Yoshii T., Helfrich-Förster C., Rieger D., Neuroanatomical details of the lateral neurons of Drosophila melanogaster support their functional role in the circadian system. J. Comp. Neurol. 526, 1209–1231 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Bulthuis N., Spontak K. R., Kleeman B., Cavanaugh D. J., Neuronal activity in non-LNv clock cells is required to produce free-running rest: Activity rhythms in Drosophila. J. Biol. Rhythms 34, 249–271 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Guo F., Chen X., Rosbash M., Temporal calcium profiling of specific circadian neurons in freely moving flies. Proc. Natl. Acad. Sci. U.S.A. 114, E8780–E8787 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Zhang Y., Liu Y., Bilodeau-Wentworth D., Hardin P. E., Emery P., Light and temperature control the contribution of specific DN1 neurons to Drosophila circadian behavior. Curr. Biol. 20, 600–605 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kistenpfennig C., Hirsh J., Yoshii T., Helfrich-Förster C., Phase-shifting the fruit fly clock without cryptochrome. J. Biol. Rhythms 27, 117–125 (2012). [DOI] [PubMed] [Google Scholar]
- 47.Vinayak P., et al. , Exquisite light sensitivity of Drosophila melanogaster cryptochrome. PLOS Genet. 9, e1003615 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Helfrich-Förster C., Development of pigment-dispersing hormone-immunoreactive neurons in the nervous system of Drosophila melanogaster. J. Comp. Neurol. 380, 335–354 (1997). [DOI] [PubMed] [Google Scholar]
- 49.Sheeba V., et al. , Large ventral lateral neurons modulate arousal and sleep in Drosophila. Curr. Biol. 18, 1537–1545 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Wiggin T. D., et al. , Covert sleep-related biological processes are revealed by probabilistic analysis in Drosophila. Proc. Natl. Acad. Sci. U.S.A. 117, 10024–10034 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Osterwalder T., Yoon K. S., White B. H., Keshishian H., A conditional tissue-specific transgene expression system using inducible GAL4. Proc. Natl. Acad. Sci. U.S.A. 98, 12596–12601 (2001). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.McGuire S. E., Mao Z., Davis R. L., Spatiotemporal gene expression targeting with the TARGET and gene-switch systems in Drosophila. Sci. STKE 2004, pl6 (2004). [DOI] [PubMed] [Google Scholar]
- 53.Li Q., Stavropoulos N., Evaluation of ligand-inducible expression systems for conditional neuronal manipulations of sleep in Drosophila. G3 (Bethesda) 6, 3351–3359 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Parisky K. M., Agosto Rivera J. L., Donelson N. C., Kotecha S., Griffith L. C., Reorganization of sleep by temperature in Drosophila requires light, the homeostat, and the circadian clock. Curr. Biol. 26, 882–892 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.McClure C. D., et al. , An auxin-inducible, GAL4-compatible, gene expression system for Drosophila. eLife 11, e67598 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ma D., et al. , Neural connectivity molecules best identify the heterogeneous clock and dopaminergic cell types in the Drosophila adult brain. Sci. Adv. 9, eade8500 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Cyran S. A., et al. , vrille, Pdp1, and dClock Form a second feedback loop in the Drosophila circadian clock. Cell 112, 329–341 (2003). [DOI] [PubMed] [Google Scholar]
- 58.Gunawardhana K. L., Hardin P. E., VRILLE controls PDF neuropeptide accumulation and arborization rhythms in small ventrolateral neurons to drive rhythmic behavior in Drosophila. Curr. Biol. 27, 3442–3453.e4 (2017). [DOI] [PubMed] [Google Scholar]
- 59.Helfrich-Förster C., Winter C., Hofbauer A., Hall J. C., Stanewsky R., The circadian clock of fruit flies is blind after elimination of all known photoreceptors. Neuron 30, 249–261 (2001). [DOI] [PubMed] [Google Scholar]
- 60.Stoleru D., et al. , The Drosophila circadian network is a seasonal timer. Cell 129, 207–219 (2007). [DOI] [PubMed] [Google Scholar]
- 61.Picot M., Cusumano P., Klarsfeld A., Ueda R., Rouyer F., Light activates output from evening neurons and inhibits output from morning neurons in the Drosophila circadian clock. PLoS Biol. 5, e315 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Murad A., Emery-Le M., Emery P., A subset of dorsal neurons modulates circadian behavior and light responses in Drosophila. Neuron 53, 689–701 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Chen K. F., Peschel N., Zavodska R., Sehadova H., Stanewsky R., QUASIMODO, a Novel GPI-anchored zona pellucida protein involved in light input to the Drosophila circadian clock. Curr. Biol. 21, 719–729 (2011). [DOI] [PubMed] [Google Scholar]
- 64.Schlichting M., et al. , A neural network underlying circadian entrainment and photoperiodic adjustment of sleep and activity in Drosophila. J. Neurosci. 36, 9084–9096 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Tang C.-H.A., Hinteregger E., Shang Y., Rosbash M., Light-mediated TIM degradation within Drosophila Pacemaker neurons (s-LNvs) is neither necessary nor sufficient for delay zone phase shifts. Neuron 66, 378–385 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Guo F., Cerullo I., Chen X., Rosbash M., PDF neuron firing phase-shifts key circadian activity neurons in Drosophila. eLife 3, e02780 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Fogle K. J., Parson K. G., Dahm N. A., Holmes T. C., CRYPTOCHROME is a blue-light sensor that regulates neuronal firing rate. Science 331, 1409–1413 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Liu S., et al. , WIDE AWAKE mediates the circadian timing of sleep onset. Neuron 82, 151–166 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Li Y., et al. , Drosophila neuroligin 4 regulates sleep through modulating GABA transmission. J. Neurosci. 33, 15545–15554 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Ewen-Campen B., et al. , split-inteinGal4 provides intersectional genetic labeling that is fully repressible by Gal80. bioRxiv [Preprint] (2023). 10.1101/2023.03.24.534001 (Accessed 15 May 2023). [DOI] [PMC free article] [PubMed]
- 71.Donelson N., et al. , High-resolution positional tracking for long-term analysis of Drosophila Sleep and locomotion using the “tracker” program. PLOS One 7, e37250 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Glossop N. R. J., et al. , VRILLE feeds back to control circadian transcription of Clock in the Drosophila circadian oscillator. Neuron 37, 249–261 (2003). [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Appendix 01 (PDF)
Dataset S01 (XLSX)
Data Availability Statement
All data are included in the article and/or supporting information.