

Abstract
Pure cultures of Geobacter sulfurreducens and other Fe(III)-reducing bacteria accumulated hydrogen to partial pressures of 5 to 70 Pa with acetate, butyrate, benzoate, ethanol, lactate, or glucose as the electron donor if electron release to an acceptor was limiting. G. sulfurreducens coupled acetate oxidation with electron transfer to an anaerobic partner bacterium in the absence of ferric iron or other electron acceptors. Cocultures of G. sulfurreducens and Wolinella succinogenes with nitrate as the electron acceptor degraded acetate efficiently and grew with doubling times of 6 to 8 h. The hydrogen partial pressures in these acetate-degrading cocultures were considerably lower, in the range of 0.02 to 0.04 Pa. From these values and the concentrations of the other reactants, it was calculated that in this cooperation the free energy change available to G. sulfurreducens should be about −53 kJ per mol of acetate oxidized, assuming complete conversion of acetate to CO2 and H2. However, growth yields (18.5 g of dry mass per mol of acetate for the coculture, about 14 g for G. sulfurreducens) indicated considerably higher energy gains. These yield data, measurement of hydrogen production rates, and calculation of the diffusive hydrogen flux indicated that electron transfer in these cocultures may not proceed exclusively via interspecies hydrogen transfer but may also proceed through an alternative carrier system with higher redox potential, e.g., a c-type cytochrome that was found to be excreted by G. sulfurreducens into the culture fluid. Syntrophic acetate degradation was also possible with G. sulfurreducens and Desulfovibrio desulfuricans CSN but only with nitrate as electron acceptor. These cultures produced cell yields of 4.5 g of dry mass per mol of acetate, to which both partners contributed at about equal rates. These results demonstrate that some Fe(III)-reducing bacteria can oxidize organic compounds under Fe(III) limitation with the production of hydrogen, and they provide the first example of rapid acetate oxidation via interspecies electron transfer at moderate temperature.
The bacterial reduction of Fe(III) (ferric iron) to Fe(II) (ferrous iron) is a process of major geochemical significance in anoxic aquatic environments such as groundwater aquifers and marine or freshwater sediments (9–11, 16). Iron(III)-reducing bacteria use ferric iron as the terminal electron acceptor for the oxidation of organic compounds to CO2 or other oxidized metabolites, and they conserve energy for growth at least partially by electron transport phosphorylation (15). In the absence of ferric iron, various ferric iron-reducing bacteria can use alternative electron acceptors, e.g., nitrate, fumarate, and elemental sulfur, for the oxidation of organic substrates (12). In the absence of any of these electron acceptors, ferric iron-reducing bacteria of the genera Geobacter, Desulfuromonas, Geovibrio, and Shewanella cannot switch to a fermentative type of metabolism. Transfer of electrons to protons to form hydrogen as the fermentation product is most often limited by the overall reaction energetics and would require hydrogen removal at low partial pressure (3, 14, 19, 21).
Especially with acetate as the substrate, the energy span of fermentation to CO2 and hydrogen is so small that the reverse reaction (homoacetogenesis) rather than the forward reaction most often proceeds. Nonetheless, a syntrophic coculture of a fermenting bacterium and a methanogenic bacterium which oxidized acetate at reasonable rates at 58°C was isolated (8, 28). The elevated temperature favors acetate cleavage (21). Recently, syntrophic acetate degradation has also been reported to occur at 37°C in cocultures of the newly described Clostridium ultunense with hydrogenotrophic methanogens, although at an extremely low rate (about 30 μM per day) (22).
In this paper, we document that Geobacter sulfurreducens can oxidize acetate in syntrophic cooperation with partners such as Wolinella succinogenes or Desulfovibrio desulfuricans at 30°C with nitrate as the electron acceptor.
MATERIALS AND METHODS
Sources of strains.
W. succinogenes (DSM 1740) was kindly supplied by Steve Goodwin, University of Massachusetts, Amherst. Methanospirillum hungatei (DSM 864) and D. desulfuricans CSN (DSM 9104) were from the culture collection of the Microbial Ecology Laboratory, Konstanz, Germany. G. sulfurreducens (ATCC 51573), G. metallireducens (ATCC 53774), G. hydrogenophilus (ATCC 51590), Shewanella alga BRY (ATCC 51181), Shewanella sp. strain GC-29, and Desulfuromonas palmitatis (ATCC 51701) were from the Department of Microbiology, University of Massachusetts, Amherst.
Media and cultivation.
For cocultures of G. sulfurreducens with W. succinogenes, a cysteine-reduced bicarbonate and phosphate-buffered medium was used with the following composition (in grams per liter): KH2PO4, 0.45; K2HPO4, 0.2; NH4Cl, 0.3; CaCl2 · 2H2O, 0.04; MgSO4 · 7H2O, 0.1; NaCl, 1.0; NaHCO3, 1.8; Na2CO3, 0.5; resazurin, 0.004; cysteine-HCl · H2O, 0.18. A syntrophic association between G. sulfurreducens and W. succinogenes was established on acetate-nitrate medium by inoculating 10% of the G. sulfurreducens bacteria from a fumarate-limited acetate-fumarate medium and 10% of the W. succinogenes bacteria from a formate-limited formate-nitrate medium. After 1 week of growth, 10% of this preculture was used as the inoculum for the experiment described here. Acetate, nitrate, H2, and the optical density were monitored regularly. The experiment was carried out in duplicate. To obtain a nondisturbed time course, care was taken to avoid temperature changes while sampling.
Cocultures of G. sulfurreducens with M. hungatei and D. desulfuricans CSN were grown in a similar manner in sulfide-reduced bicarbonate-buffered mineral medium (27) with acetate as the only source of organic carbon and electrons.
Analytical techniques.
Nitrate was analyzed with Dionex DX100 ion chromatograph equipped with a conductivity detector and an AS4A-SC column. Acetate was analyzed by high-pressure liquid chromatography with a Bio-Rad Aminex HPX-87H column and with 5 mM sulfuric acid as the eluent; the absorbance was monitored at 210 nm. Hydrogen and carbon monoxide were quantified on a TraceAnalytical Reduction Gas Detector.
Ammonia was determined by the phenol-hypochlorite method (25). Bacterial growth in anaerobic test tubes was monitored by turbidity measurement at 570 nm by placing the test tubes directly into a Bausch & Lomb spectrophotometer.
RESULTS
H2 production by pure cultures of iron-reducing bacteria.
Pure cultures of various species of iron-reducing bacteria were tested for their potential to produce molecular hydrogen as a side product after electron acceptor-limited growth. Except for G. metallireducens GS15, all the strains tested produced molecular hydrogen up to significant pressures (Table 1). G. metallireducens was also the only strain tested that did not consume molecular hydrogen when given ferric iron or other suitable electron acceptors. Obviously, only the hydrogen-consuming (hydrogenase-positive) ferric iron reducers could also produce molecular hydrogen.
TABLE 1.
Hydrogen concentrations in the headspace of pure cultures of Fe(III)-reducing bacteria
| Species | Strain | Electron donor and concn (mM)a | Electron acceptor and concn (mM)a | H2 (Pa) |
|---|---|---|---|---|
| G. sulfurreducens | PCA | Acetate (20) | Fumarate (20) | 40 |
| G. hydrogenophilus | H-2 | Acetate (20) | Fumarate (20) | 73 |
| G. hydrogenophilus | H-2 | Butyrate (10) | Fe(III) citrate (100) | 18 |
| G. hydrogenophilus | H-2 | Benzoate (5) | Fe(III) citrate (100) | 2.6 |
| G. hydrogenophilus | H-2 | Ethanol (20) | Fumarate (20) | 11 |
| G. metallireducens | GS15 | Acetate (20) | Nitrate (10) | 0.17 |
| Shewanella alga | BRY | Lactate (20) | Fumarate (20) | 43 |
| Shewanella sp. | GC29 | Glucose (20) | Fumarate (20) | 5.5 |
| Shewanella sp. | GC29 | Lactate (20) | Fumarate (20) | 7.0 |
| Desulfuromonas palmitatis | SDBY | Lactate (20) | Fumarate (20) | 14 |
The concentrations shown are the maximum observed. They varied by about 200%, depending on the culture conditions and age.
The production of hydrogen by G. sulfurreducens (4) during acetate degradation was investigated in more detail. After electron acceptor-limited growth with 20 mM acetate and 20 mM fumarate, the hydrogen partial pressure was approximately 40 Pa. After the headspace of the culture vessel was flushed with N2, the production of hydrogen was monitored (Fig. 1). The culture produced hydrogen until approximately the previous partial pressure (35 to 40 Pa) was reached. The maximum rate of hydrogen production in this experiment was very low (12.5 nmol per liter per min), compared to the rate of acetate degradation by the growing culture at similar density before fumarate depletion (>70 μmol per liter per min).
FIG. 1.

Time course of hydrogen production (▪) after replacing the headspace of the flask with N2 after 5 and 58 h. After 20 h, the vessel was placed on an orbital shaker at 100 rpm.
Acetate degradation in cocultures of G. sulfurreducens and W. succinogenes.
To test whether removal of hydrogen could allow quantitative oxidation of acetate via interspecies hydrogen transfer, G. sulfurreducens was cocultivated with W. succinogenes in the presence of nitrate as the terminal electron acceptor. W. succinogenes was chosen because it cannot oxidize acetate and is an efficient hydrogen-consuming anaerobe reducing nitrate to ammonia as the electron-accepting reaction. The coculture degraded acetate concomitantly with stoichiometric reduction of nitrate to ammonia (Fig. 2), while pure cultures of G. sulfurreducens or W. succinogenes did not degrade acetate with nitrate as the electron acceptor.
FIG. 2.

Time course of acetate oxidation by the coculture of G. sulfurreducens and W. succinogenes with nitrate as the electron acceptor. ▴, acetate; •, nitrate; ▪, optical density at 570 nm (OD 570).
The doubling time of the coculture with W. succinogenes was about 18 h at the beginning of the batch culture and was significantly shorter (6 to 8 h) toward the end of growth (Fig. 3). The coculture produced 18.5 ± 3.2 g of cell dry mass per mol of acetate oxidized. Microscopic examination revealed that both organisms were present, with clearly greater cell numbers of G. sulfurreducens (the cell number ratio of G. sulfurreducens to W. succinogenes was 2.1:1 to 4.5:1). Because of the larger cell size of G. sulfurreducens, the total biomass produced by this organism was about four to nine times greater than that of W. succinogenes. Pure cultures of W. succinogenes reached growth yields of 3.3 to 4.5 g per mol of hydrogen when cultivated with hydrogen and limiting amounts of nitrate (2 or 5 mM). Similar yield data were obtained with formate as an alternative electron donor or with fumarate as an alternative electron acceptor; these values agreed basically with earlier published data (1, 2, 13).
FIG. 3.

Half-logarithmic plot of a growth curve similar to that shown in Fig. 2.
During growth of the coculture, the hydrogen partial pressure was always below 0.04 Pa and was at times below 0.02 Pa. At the end of growth, the hydrogen concentration stayed at levels higher than 30 Pa in nitrate-limited cultures and lower than 0.02 Pa in acetate-limited cultures. In the supernatants of such cocultures, a c-type cytochrome was detected at up to 200 nM; this compound is characterized in a separate report (23).
Acetate degradation in cocultures of G. sulfurreducens with D. desulfuricans or M. hungatei.
To test whether G. sulfurreducens can also transfer reducing equivalents from acetate oxidation to other hydrogen-consuming anaerobes, D. desulfuricans CSN was tested as a potential electron acceptor. This bacterium can link the oxidation of hydrogen to the reduction of either sulfate or nitrate (7). With sulfate as the electron acceptor, cocultures of G. sulfurreducens and D. desulfuricans never produced sulfide or degraded acetate, even when tested in dense cell suspensions or incubated for more than 6 weeks. No growth occurred in such cocultures, although the hydrogen partial pressure was constantly lower than 0.2 Pa. However, if nitrate replaced sulfate as the electron acceptor the same coculture degraded acetate (Fig. 4) with concomitant reduction of nitrate to ammonia but at a considerably lower rate than that for the G. sulfurreducens-W. succinogenes coculture (doubling time, about 30 h). An extended lag period was often observed when these two species were coupled, probably due to delayed induction of the nitrate-reducing enzyme system. During this lag phase, the medium was oxidized and resazurin in the medium turned pink. The cell yield of these cocultures was 4.5 ± 1.1 g of dry cell mass per mol of acetate oxidized. Cocultures of G. sulfurreducens and the hydrogenotrophic methanogen M. hungatei did not degrade acetate to a measurable extent over 6 weeks of incubation. The hydrogen partial pressure was relatively high (between 1.2 and 3 Pa) in these cocultures.
FIG. 4.

Time course of acetate oxidation with nitrate as the electron acceptor by the coculture of G. sulfurreducens and D. desulfuricans CSN. ▴, acetate; ▪, optical density at 578 nm (OD 578).
DISCUSSION
In the present report, hydrogen formation from organic substrates by Fe(III)-reducing bacteria is documented for the first time. Production of hydrogen from acetate appeared to be most interesting because acetate is a major intermediate in anaerobic biomass degradation and because hydrogen formation from acetate could be linked to hydrogen utilization in a syntrophic oxidation process. Mesophilic conversion of acetate to CO2 and H2 is also of special interest because of the small amount of Gibbs free energy released.
Energetic considerations.
If interspecies electron transfer in our cocultures was based exclusively on molecular hydrogen as carrier (“interspecies hydrogen transfer”) the following equations would apply:
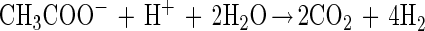 |
1 |
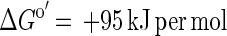 |
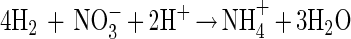 |
2 |
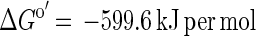 |
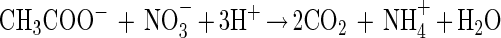 |
3 |
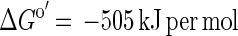 |
Conversion of acetate to H2 plus CO2 is endergonic under standard conditions (equation 1) and requires continuous removal of hydrogen by the hydrogen-consuming reaction (equation 2) to make the combined reaction exergonic for both partners (equation 3). Under the conditions applied in our cultures (acetate, 10 mM; CO2, 10 kPa), the first reaction becomes exergonic at a hydrogen partial pressure lower than 10 Pa. G. sulfurreducens in pure culture produced hydrogen in acceptor-limited cultures up to a pressure of 40 Pa. This level indicates a remnant energy potential in the cell of +4 kJ per mol of hydrogen. Obviously, the cells could not proliferate or grow under these conditions.
In growing cocultures of G. sulfurreducens and W. succinogenes, hydrogen partial pressures of 0.02 to 0.04 Pa were measured, shifting the Gibbs free energy of acetate conversion to H2 plus CO2 to approximately ΔG′ = −53 kJ per mol acetate. This small amount of free energy would allow synthesis of about two-thirds of an ATP unit, assuming that 1 mol of ATP requires approximately −70 kJ per mol under physiological conditions in actively growing cells (19, 21, 26). The cell yields obtained were far higher than expected from these considerations: the coculture yielded 18.5 g of dry cell mass per mol of acetate, and about 80% of this cell mass (14 g) was G. sulfurreducens cells. Even under optimal conditions, the yield of cell matter synthesized per mol of ATP formed is up to 10 g per mol of ATP on mineral media with a fatty acid as the sole substrate mostly in the range of 5 g per mol (24). The partner organism W. succinogenes in pure culture formed 3.6 to 4.5 g of cell mass per mol of hydrogen. Thus, one would expect 14.4 to 18 g of W. succinogenes cell mass per mol of acetate if the electrons were transferred exclusively through hydrogen, but the measured contribution to the total cell yield of the mixed culture was only about 20%, i.e., 4.5 g of cell mass. Obviously, the energy yield of G. sulfurreducens is considerably higher and that of W. succinogenes is lower than expected for electron transfer exclusively through hydrogen. We conclude that electrons in this coculture have to be transferred to a significant extent through a separate carrier system with a more positive redox potential than that of the proton/hydrogen couple.
Also, the coculture of G. sulfurreducens with D. desulfuricans degraded acetate in syntrophic cooperation only if nitrate served as the electron acceptor. Growth and acetate oxidation in this coculture were far slower than in the coculture with W. succinogenes, but the hydrogen partial pressure during this conversion was in the same range (0.02 Pa) as in the other coculture. The energy gain for acetate conversion to CO2 and H2 by G. sulfurreducens can be calculated at −53 kJ per mol of acetate, and the total cell yield for this coculture was 4.5 g per mol of acetate. In the microscopic picture, both bacteria were present at about equal mass fractions, confirming that each partner bacterium obtained less than 1 ATP unit per reaction run.
The G. sulfurreducens-D. desulfuricans coculture did not couple with sulfate as the electron acceptor, although such a reaction is theoretically possible:
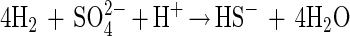 |
4 |
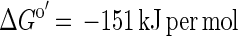 |
and, combining equations 1 and 4,
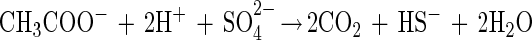 |
5 |
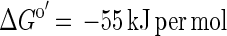 |
Such a syntrophically cooperating coculture has been described previously (17); however, this culture grows very slowly, with doubling times of more than 7 days. Obviously, G. sulfurreducens did not couple efficiently enough to the partner bacterium to perform this energetically difficult reaction (in our culture with 10 mM acetate, it yielded ΔG′ = −45 kJ per mol of acetate, just enough energy for one-third of an ATP unit per reaction run for each partner [18]).
The reason why acetate oxidation with sulfate failed could also be the presence of up to 4 mM sulfide in the culture medium, which was found to completely inhibit acetate degradation by G. sulfurreducens in the presence of its preferred electron acceptor, fumarate. During the growth of successful cocultures and of pure strains of G. sulfurreducens in media containing the redox indicator resazurin, the medium turned from colorless to pink, indicating that a redox potential more positive than that of the sulfate/sulfide couple was established. The production of sulfide keeps the redox potential lower and might be an additional factor preventing an effective coupling of acetate oxidation with sulfate reduction.
The energetic situation of syntrophic conversion of acetate to methane and CO2 is even worse:
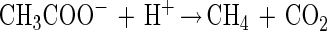 |
6 |
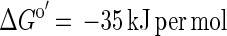 |
In our culture with 10 mM acetate, the ΔG′ is −25 kJ per mol, which can hardly feed two partner bacteria. This reaction can be operated efficiently by a syntrophic culture only at enhanced temperatures, e.g., 58°C (21, 28).
Kinetic considerations.
The observed syntrophic oxidation of acetate by the cocultures described here first suggested that electrons were transferred between the two partners through interspecies hydrogen transfer. The energetic considerations mentioned above postulate a further carrier system of higher redox potential. This argument is supported by calculations of diffusive hydrogen flux in our cocultures. In Table 2, we compare metabolic activities and electron transfer rates between the partner bacteria in both growing cocultures with a calculated diffusive hydrogen flux between the partners involved. Even assuming a maximal hydrogen concentration on the side of the producer equivalent to the hydrogen concentration accumulated in pure culture (Table 1), the calculated hydrogen flux can account only for 5.6 to 8.2% of the electron flow between the two partner organisms. The real diffusive hydrogen flux is probably far lower because the actual hydrogen concentration at the producer cell surface should be at least 1 order of magnitude lower than assumed above. This view is further supported by the low hydrogen release rate observed with G. sulfurreducens in pure culture (Fig. 1), although this might be due to insufficient induction of the hydrogenase system after growth with acetate plus fumarate. One may argue that the hydrogen flux through the liquid medium proceeds not only by diffusive transport but also by convective transport. The diffusion distance used in our calculations as the average distance between freely suspended cells at the densities indicated is equivalent to twice the diffusive boundary (Stern) layer (10 μm) that surrounds both the producer and the consumer cells and precludes convective transport. However, highly motile cells such as Wolinella cells may cause microscopically detectable turbulences and thus accelerate convective hydrogen flux between the partner bacteria.
TABLE 2.
Comparison of metabolic activity of G. sulfurreducens in coculture with either W. succinogenes or D. desulfuricans with the calculated electron flux through hydrogena
| Parameter | Value for coculture of:
|
Unit | |
|---|---|---|---|
| Geobacter and Wolinella | Geobacter and Desulfovibrio | ||
| Doubling time | 6–8 | 30 | h |
| μ | 0.099 | 0.023 | h−1 |
| Yieldb | 14 | 2.3 | g · mol−1 |
| Acetate oxidation rate | 120 | 167 | μmol · min−1 · g (dry mass)−1 |
| Geobacter single-cell surface areac | 4.3 | 4.3 | μm2 |
| At late log phase | |||
| OD570d | 0.6 | 0.2 | |
| Cell density | 180 | 60 | mg (dry mass) · liter−1 |
| Acetate oxidizing activity | 21.6 | 10.0 | μmol · min−1 · liter−1 |
| Cell no. | 1.2 × 108 | 0.4 × 108 | ml−1 |
| Total cell surface area in culture | 5.16 | 1.72 | cm2 · ml−1 |
| Average distance (d) | 20 | 22 | μm |
| Calculated H2 flux for C1 = 302 nMe and C2 = 0.15 nM | 1.78 | 0.56 | μmol · min−1 · liter−1 |
| Contribution to calculated electron flow in the active coculture | 8.2 | 5.6 | % |
Fluxes were calculated from Fick’s first law: J = A · D · (C1 − C2) · d−1, where A is the area over which the metabolite diffuses, i.e., the total producer cell surface, D is the diffusion constant (3.81 × 10−5 cm2 s−1 for H2 in H2O at 25°C [6]), (C1 − C2) is the concentration difference between source and sink, and d is the diffusion distance.
Only the yield of G. sulfurreducens was considered here.
The surface area of a single cell is calculated as that of a cylinder with a diameter of 0.5 μm and a height of 2.5 μm (4).
OD570, optical density at 570 nm.
Water under 1 atm of H2 at 30°C contains 0.75 mmol H2 in solution (5); 40 Pa of H2 in the gas phase is in equilibrium with 302 nM H2 in solution.
Formate has been suggested repeatedly as an alternative electron-shuttling system in interspecies electron transfer (19). For the cocultures considered here, it would not be a real alternative because the energetic problems arising are the same as with hydrogen, since the standard redox potentials are nearly the same (Eo′ = −430 and −414 mV, respectively). Also with respect to diffusion kinetics, formate would pose the same problems as hydrogen: the formate concentration energetically equivalent to the observed hydrogen concentrations in the cocultures (0.04 to 40 Pa) is in the range of 10 nM to 10 μM (20), and the electron flow through this carrier could be only in the same range as that of hydrogen. We did not measure formate concentrations in our cocultures, because no reliable techniques are available for formate detection in this concentration range.
We observed in pure cultures and in cocultures of G. sulfurreducens an accumulation of a c-type cytochrome up to 200 nM, which has so far been observed only associated with the cells (4) and will be described in detail in a separate paper (23). Since this carrier has a standard redox potential of −167 mV, it may act as an efficient electron shuttle system just in that potential range that would fit into our energetic considerations mentioned above. The increasing growth rate observed in the coculture with W. succinogenes at increased cell densities (Fig. 3) could be explained by the excretion of such a carrier, proportional to the cell density. Our results presented here provide indirect evidence for the operation of such a carrier system in the described cocultures.
ACKNOWLEDGMENTS
We thank Sabine Seeliger for providing information on the purified extracellular c-type cytochrome of G. sulfurreducens.
This study was supported by grants from the University of Konstanz, the Deutsche Forschungsgemeinschaft, Bonn-Bad Godesberg, and the Fonds der Chemischen Industrie, Frankfurt, Germany.
REFERENCES
- 1.Bokranz M, Katz J, Schröder I, Roberton A M, Kröger A. Energy metabolism and biosynthesis of Vibrio succinogenes. Arch Microbiol. 1983;135:36–41. [Google Scholar]
- 2.Bronder M, Mell H, Stupperich E, Kröger A. Biosynthetic pathways of Vibrio succinogenes growing with fumarate as terminal electron acceptor and sole carbon source. Arch Microbiol. 1982;131:216–223. doi: 10.1007/BF00405882. [DOI] [PubMed] [Google Scholar]
- 3.Bryant M P. Microbial methane production—theoretical aspects. J Anim Sci. 1979;48:193–201. [Google Scholar]
- 4.Caccavo F, Jr, Lonergan D J, Lovley D R, Davis M, Stolz J F, McInerney M J. Geobacter sulfurreducens sp. nov., a hydrogen- and acetate-oxidizing dissimilatory metal-reducing microorganism. Appl Environ Microbiol. 1994;60:3752–3759. doi: 10.1128/aem.60.10.3752-3759.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Dawson R M C, Elliott D C, Elliott W H, Jones K M. Data for biochemical research. 2nd ed. Oxford, United Kingdom: Clarendon Press; 1969. [Google Scholar]
- 6.Gertz K H, Loeschcke H H. Bestimmung des Diffusionskoeffizienten von H2, O2, N2, und He in Wasser und Blutserum bei konstant gehaltener Konvektion. Z Naturforsch Teil B. 1954;9:1–9. [Google Scholar]
- 7.Krekeler D, Cypionka H. The preferred electron acceptor of Desulfovibrio desulfuricans CSN. FEMS Microbiol Ecol. 1995;17:271. [Google Scholar]
- 8.Lee M J, Zinder S H. Isolation and characterization of a thermophilic bacterium which oxidizes acetate in syntrophic association with a methanogen and which grows acetogenically on H2-CO2. Appl Environ Microbiol. 1988;54:124–129. doi: 10.1128/aem.54.1.124-129.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lovley D R. Dissimilatory metal reduction. Annu Rev Microbiol. 1993;47:263–290. doi: 10.1146/annurev.mi.47.100193.001403. [DOI] [PubMed] [Google Scholar]
- 10.Lovley D R. Microbial reduction of iron, manganese and other metals. Adv Agron. 1995;54:175–231. [Google Scholar]
- 11.Lovley D R, Chapelle F H. Deep subsurface microbial processes. Rev Geophys. 1995;33:365–381. [Google Scholar]
- 12.Lovley D R, Coates J D, Saffarini D A, Lonergan D J. Dissimilatory iron reduction. In: Winkelmann G, Carrano C J, editors. Iron and related transition metals in microbial metabolism. Chur, Switzerland: Harwood Academic Publishers; 1997. pp. 187–215. [Google Scholar]
- 13.Macy J M, Schröder I, Thauer R K, Kröger A. Growth of Wolinella succinogenes on H2S plus fumarate and on formate plus sulfur as energy sources. Arch Microbiol. 1986;144:147–150. [Google Scholar]
- 14.McInerney M J. Anaerobic degradation of proteins and lipids. In: Zehnder A J B, editor. Biology of anaerobic microorganisms. New York, N.Y: John Wiley & Sons, Inc.; 1988. pp. 373–415. [Google Scholar]
- 15.Myers C R, Nealson K H. Respiration-linked proton translocation coupled to anaerobic reduction of manganese(IV) and iron(III) in Shewanella putrefaciens MR-1. J Bacteriol. 1990;172:6232–6238. doi: 10.1128/jb.172.11.6232-6238.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Nealson K H, Saffarini D A. Iron and manganese in anaerobic respiration: environmental significance, physiology, and regulation. Annu Rev Microbiol. 1994;48:311–343. doi: 10.1146/annurev.mi.48.100194.001523. [DOI] [PubMed] [Google Scholar]
- 17.Rozanova E, Galushko A, Nazina T. An acetate-decomposing sulphidogenic syntrophic association. In: Belaich J P, Bruschi M, Garcia J-L, editors. Microbiology and biochemistry of strict anaerobes involved in interspecies hydrogen transfer. New York, N.Y: Plenum Press; 1990. pp. 469–470. [Google Scholar]
- 18.Schink B. Conservation of small amounts of energy in fermenting bacteria. In: Finn R K, Praeve P, editors. Biotechnology, focus 2. Munich, Germany: Hanser Publishers; 1990. pp. 63–89. [Google Scholar]
- 19.Schink B. Syntrophism among prokaryotes. In: Balows A, Truper H G, Dworkin M, Schleifer K H, editors. The prokaryotes. 2nd ed. New York, N.Y: Springer-Verlag; 1991. pp. 276–299. [Google Scholar]
- 20.Schink B. Diversity, ecology, and isolation of acetogenic bacteria. In: Drake H L, editor. Acetogenesis. New York, N.Y: Chapman & Hall; 1994. pp. 197–235. [Google Scholar]
- 21.Schink B. Energetics of syntrophic cooperation in methanogenic degradation. Microbiol Mol Biol Rev. 1997;61:262–280. doi: 10.1128/mmbr.61.2.262-280.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Schnuerer A, Schink B, Svensson B H. Clostridium ultunense sp. nov., a mesophilic bacterium oxidizing acetate in syntrophic association with a hydrogenotrophic methanogenic bacterium. Int J Syst Bacteriol. 1997;46:1145–1152. doi: 10.1099/00207713-46-4-1145. [DOI] [PubMed] [Google Scholar]
- 23.Seeliger, S., R. Cord-Ruwisch, and B. Schink. A periplasmic and extracellular c-type cytochrome of Geobacter sulfurreducens acts as ferric iron reductase and as electron carrier to other acceptors or to partner bacteria. Submitted for publication. [DOI] [PMC free article] [PubMed]
- 24.Stouthamer A H. The search for correlation between theoretical and experimental growth yields. Int Rev Biochem. 1979;21:1–47. [Google Scholar]
- 25.Strickland J D H, Parsons T R. A practical handbook of seawater analysis. Ottawa, Ontario, Canada: Fisheries Research Board of Canada; 1972. [Google Scholar]
- 26.Thauer R K, Jungermann K, Decker K. Energy conservation in chemotrophic anaerobic bacteria. Bacteriol Rev. 1977;41:100–180. doi: 10.1128/br.41.1.100-180.1977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Widdel F, Pfennig N. Studies on dissimilatory sulfate-reducing bacteria that decompose fatty acids. I. Isolation of a new sulfate reducer enriched with acetate from saline environments. Description of Desulfobacter postgatei gen. nov. sp. nov. Arch Microbiol. 1981;129:395–400. doi: 10.1007/BF00406470. [DOI] [PubMed] [Google Scholar]
- 28.Zinder S H, Koch M. Non-aceticlastic methanogenesis from acetate: acetate oxidation by a thermophilic syntrophic coculture. Arch Microbiol. 1984;138:263–272. [Google Scholar]