

Abstract
Assays of photosynthesis were conducted with a biofilm population of a cyanobacterium, a Synechococcus sp., growing at ∼70°C in a Yellowstone National Park hot spring to test whether cells growing near the upper temperature limit of photosynthetic life are optimally adapted to their mean environmental temperature. Cell suspensions were assayed at 70, 65, and 55°C while being simultaneously exposed to modified solar environments, including reduction of total irradiance and exclusion of UV radiation. Carbon fixation was greatest at 65°C, while 70 and 55°C were always supraoptimal and suboptimal for photosynthesis, respectively. The degree of temperature stress was dependent upon light intensity, and this light-dependent temperature effect may involve both reduced quantum efficiency at subsaturating irradiances and a lower saturating irradiance at both supraoptimal and suboptimal temperatures. The Synechococcus sp. was also more susceptible to UV inhibition of photosynthesis at nonoptimal temperatures. These results suggest that this population is persisting at a nearly lethal temperature and is consequently subject to greater damage by both visible and UV radiation, but it is speculated that these cells may be avoiding competition with other photoautotrophs under these nonoptimal conditions. In separate experiments monitoring diurnal patterns of photosynthesis, cells exhibited peak productivity during the morning, followed by an afternoon decline. No recovery of photosynthesis was observed during the remaining daytime, and carbon fixation was always UV inhibited under conditions of photosynthetically saturating light.
Unicellular cyanobacteria formerly assigned to the morphospecies Synechococcus cf. lividus Copeland are major components of microbial mat communities in alkaline hot springs of the Western hemisphere and East Asia at temperatures ranging from approximately 45 to 73°C, the upper temperature limit of photosynthetic life (2, 6, 7). It is the only cyanobacterial morphospecies found above 64°C. Early studies in laboratory culture of clones isolated from an Oregon hot spring have provided evidence for the existence of multiple strains of this morphospecies that vary in optimal growth temperature and growth temperature range (31), and recent investigations suggest that these temperature strains may have diverged at the 16S rRNA sequence level (11, 28).
The effects of temperature on photosynthesis and growth of oxygenic phototrophs have been well described (8, 10). Reduction in photosynthesis at supraoptimal but nonlethal temperature is the product of both increased photorespiration as a result of higher affinity of ribulose-1,5-bisphosphate carboxylase/oxygenase (RUBISCO) for O2 with increasing temperature and lowered activity of RUBISCO and/or its activase (8, 10, 21). The lethal temperature has been found in plants and algae to be determined by the thermal stability of the thylakoid membrane, of which photosystem II is the most labile component (10). Suboptimal temperature effects on photosynthesis include rate limitation by enzymatic reactions due to thermodynamic constraints, limitation for inorganic phosphate, and feedback inhibition resulting from accumulation of photosynthetic end products (10).
The question of whether populations of Synechococcus sp. growing at the upper limit of their range are optimally adapted to local means or extremes in their thermal environment has not been resolved by previous studies. After investigating inorganic carbon fixation by a Synechococcus sp. collected from different temperatures along the outflow of Mushroom Spring, Lower Geyser Basin, Yellowstone National Park, T. D. Brock and M. L. Brock (2–4) reported a positive relationship between the mean temperature of collection and the optimal temperature for photosynthesis. It was concluded that populations of these cyanobacteria have evolved such that their optimal temperature for growth and photosynthesis matches mean environmental temperature, even at 70°C, near the upper limit of their range. Data obtained in laboratory culture, however, indicate that optimal temperature for both growth and photosynthesis of a clone isolated from a sample collected near the upper temperature limit from Hunter’s Hot Springs, Oreg., ranges from 63 to 67°C, lower than the mean environmental temperature of >70°C (25). These laboratory results suggest that these bacteria are subjected to chronic temperature stress in nature.
Resolution of this issue has implications for our understanding of the limits of physiological tolerance and adaptation in these bacteria. In order to determine whether a population of Synechococcus sp. from Octopus Spring, Lower Geyser Basin, Yellowstone National Park, which was growing near the upper temperature limit for photosynthetic life, was optimally adapted photosynthetically to its local thermal environment, photosynthesis was measured during the summers of 1995 and 1996 at three temperatures: 70, 65, and 55°C. The first was the approximate mean temperature of the environment, and these temperature treatments were hypothesized to be supraoptimal, optimal, and suboptimal for photosynthesis, respectively, based on the laboratory culture data of Meeks and Castenholz (25). Cells were simultaneously exposed to modified solar environments, including reduction of total irradiance and exclusion of UV radiation, in order to investigate whether and how temperature-dependent effects on photosynthetic uptake interact with these factors (e.g., whether effects of nonoptimal temperature are exacerbated with increasing irradiance). In addition, we evaluated whether the Octopus Spring high-temperature Synechococcus sp. was subject to diurnal inhibition of photosynthesis and whether inhibition, if observed, had a UV radiation component.
MATERIALS AND METHODS
Research site.
Octopus Spring is located in the Lower Geyser Basin of Yellowstone National Park, Wyo., in the White Creek drainage area near Great Fountain Geyser. It is an alkaline spring, with a measured pH of 8.3 at its source (3), and a hard siliceous sinter has been deposited along its outflow channels. Additional water chemistry data are presented by Brock (3). A perennial biofilm of the unicellular cyanobacterium Synechococcus cf. lividus Copeland develops at ∼72°C, with the mean upper temperature limit of its range measured at 71.7°C during the summer of 1996. At the collection site, where the mean temperature was ∼70°C, this morphospecies was the sole phototroph identifiable by light microscopy. Filaments of the photoheterotrophic green nonsulfur bacterium Chloroflexus cf. aurantiacus were not present.
Temperature and solar measurements.
Collection site temperature was monitored with a StowAway XTI (Onset Computer Corp.) temperature logger programmed to read every 16 s. Visible and near-infrared irradiance (in watts per square meter) was measured during experiments with an LI-1000 data logger equipped with a pyranometer (LICOR). UVA (320 to 400 nm) and UVB (280 to 320 nm) radiation were measured with an IL-1700 research radiometer (International Instruments, Inc.). Time points are reported in standard time.
Temperature, irradiance, and UV effects on photosynthesis.
The design described below was employed three times during the summers of 1995 and 1996. Synechococcus sp. biofilm was harvested from Octopus Spring with sterile 10-ml syringes, transferred to 16-oz (473-ml) Nalgene bottles, and stored in a thermos case filled with ∼70°C water until returned to the University of Georgia research trailer, West Yellowstone, Mont. Cell suspensions were easily homogenized with a syringe and diluted with fresh Octopus Spring water to an A750 of between 0.036 and 0.046. These absorbance values lie within a range of values (0.025 to 0.420) over which the concentration of the cell suspension has no effect on experimental results (data not shown). A total of 7.5 ml of suspension was delivered to 2-oz (59-ml) Whirl-Pak bags (Nasco), previously found to be nontoxic to Synechococcus sp. at these temperatures (28). Excess suspension was stored at −20°C for chlorophyll a (Chl a) quantification (described below). Aqueous NaH14CO3 was added to each bag in a semidarkened room to a final activity of 0.062 μCi ml−1. Duplicate bags were incubated outdoors in water baths at each of three temperatures (70 ± 1, 65 ± 1, and 55 ± 1°C), four irradiances (0, 1, 2, and 3 neutral density screens), and two UV radiation treatments (UV+ and UV−), provided by a cellulose diacetate control filter and a styrene filter (K-lite UVF CS; Multicraft Plastics, Eugene, Oreg.) which blocks ∼99% of radiation below 400 nm, respectively, for a total of 24 experimental treatments. Both UV filters transmitted visible irradiance equally (∼85% transmittance). Duplicate dark controls were also provided for each temperature. Experimental incubation was initiated within 90 min of cell collection for a duration of 60 min, a period over which 14C uptake is linear at these cell densities (data not shown). Experiments were terminated by transferring the bag contents to 20-ml scintillation vials containing 0.5 ml of formalin. These were stored in the dark at 4°C until returned to the University of Oregon, where subsamples were filtered onto GN-6 filters (Gelman), acidified with 3 ml of 2% (vol/vol) concentrated HCl, and rinsed with double-distilled water. Filters were transferred to new scintillation vials, which were subsequently filled with 8 ml of EcoLume scintillation cocktail (ICN) and stored overnight in the dark at 4°C. Counts per minute were determined with a Beckman LS6000SE scintillation counter and were normalized to cell Chl a content, determined spectrophotometrically according to the method of Lenz and Zeitschel (22).
Effects of fluctuating temperature on photosynthesis.
To determine the effects of fluctuating temperature on photosynthesis, procedures were performed as described above, with the following exceptions. Quadruplicate bags were incubated under cellulose diacetate control filters directly in the spring at one of three temperature treatments, each of which fluctuated because of an ebb-surge cycle at Octopus Spring: 67 to 74 (the collection site), 62 to 70, and 52 to 55°C.
Diurnal and UV effects on photosynthesis.
The following experiment was conducted at Octopus Spring twice in 1996. Procedures were performed as described above, with the following exceptions. Quadruplicate bags were incubated directly in the spring for 40 min (24 June 1996) or 60 min (2 July 1996) under one of three UV treatments (UV+, UVB−, and UV−) provided by the cellulose diacetate control filter, a polyester filter which primarily blocks UVB (Cadillac Plastics, Baltimore, Md.; 46% transmittance at 330 nm, <1% transmittance at 310 nm), and a total-UV-screening styrene filter, respectively. Filters were placed above the collection site, with the position from upstream to downstream at each time point determined by a random number generator. Counts per minute were normalized to both cell Chl a content and cell number as determined with a hemocytometer counting chamber.
Statistical analyses.
All analysis of variance (ANOVA) and analysis of covariance (ANCOVA) models were created and pairwise comparisons were made with SuperANOVA (1). Temperature means in the three-way ANOVA model were contrasted with F tests by using an F[1,24] distribution, while other pairwise comparisons were made with Bonferroni-Dunn tests.
Mean and 95% interval estimates of both total UV and UVB inhibition were obtained with the Bliss theorem (17). Mean percent inhibition is given by 100(1 − R), where R = (Xa/Xb)/(1 − g), Xa is the estimated mean counts per minute per microgram of Chl a per hour for the control treatment, and Xb is the estimated mean for either the UVB-excluded or total-UV-excluded treatments. For observations na = nb, g = (t2S2)/nb Xb2, where t is the value of Student’s t0.975 based on na + nb − 2 df and S2 is the error mean square of the model (see below). The 95% intervals are given by ±{tS/Xb [1/na (1 − g) + R2/nb]1/2}/(1 − g). The resultant intervals are technically fiducial limits rather than confidence limits, although the two are expected to be identical under these conditions (33). For the UV treatments at a given temperature treatment in the three-way ANOVA experiment, S was determined for a one-way ANCOVA model with irradiance, quantified as mean flux in watts per square meter penetrating the Whirl-Pak bag, as a covariate. The relationship between carbon assimilation rate and irradiance was curvilinear, fitting a second-degree polynomial better than a straight line, as determined by an F test. A similar technique was used to obtain S in the pooled estimates of UV inhibition in the diurnal experimental design, with incubation start time as the covariate in this case. Again, the relationship between the dependent variable and the covariate was better described by a second-degree polynomial. For individual time points, S was estimated from the original ANOVA model. Estimates of UV inhibition with intervals which did not overlap zero were considered to be significantly different from zero, and pairs of estimates with nonoverlapping intervals were considered to be significantly different from each other.
α, the slope describing change in the rate of photosynthesis in response to increasing light at light-limiting irradiances, and Isat, the saturating irradiance for photosynthesis, were estimated by fitting the data with the parabola P = α I − [(α I)2/4Pmax] (modified from Jassby and Platt [20]), where P is carbon uptake rate (counts per minute per microgram of Chl a per hour), I is irradiance (watts per square meter), and Pmax is maximal carbon uptake rate. Curve fitting was performed with SigmaPlot, version 5.0 (19), according to the Marquadt-Levenberg algorithm. Means and standard errors of Isat were estimated by 2Pmax/α with Tukey’s jackknife (33). The 95% confidence intervals (CIs) were determined assuming the appropriate t distribution.
RESULTS
Temperature profile for the Octopus Spring research site.
Temperature data collected at the Octopus Spring research site between 29 June 1996 and 2 July 1996 are shown in Fig. 1. The mean temperature (± standard error) was 69.6 ± 0.03°C. The observed fluctuation around the mean is primarily due to an ebb-and-surge cycle in the spring with a period of approximately 4.5 min.
FIG. 1.

Temperature profile of the Octopus Spring research site. Approximately 64 h of data was collected between 29 June 1996 and 2 July 1996.
Effect of temperature on photosynthesis.
Carbon uptake rate data from three experiments in which cells collected from the Octopus Spring site were exposed to combinations of different temperatures, total solar irradiances, and UV treatments are presented in Fig. 2. Dark control uptake values were low (Fig. 2), and thus, the observed values primarily reflect photosynthetic uptake at all temperatures. F test values for the effect of temperature in a three-way ANOVA were highly significant in all experiments (P < 0.001 in all cases), with within-temperature means summarized in Table 1. Carbon assimilation rates increased in the order 65°C > 70°C > 55°C for all experiments. Thus, the greatest carbon uptake was always observed at 65°C, a lower temperature than the measured mean temperature for the site, which indicates that the mean environmental temperature is supraoptimal for photosynthesis by Synechococcus sp. cells growing in Octopus Spring near the upper temperature limit for photosynthetic life.
FIG. 2.

Chl-normalized carbon uptake rate of Synechococcus sp. from the Octopus Spring research site on the morning of 8 July 1995 (A), the afternoon of 8 July 1995 (B), and on 25 June 1996 (C). Cells were assayed at a series of irradiances under the following conditions: 65°C, UV− (▾); 65°C, UV+ (▿); 70°C, UV− (•); 70°C, UV+ (○); 55°C, UV− (■); and 55°C, UV+ (□). Error bars represent standard errors.
TABLE 1.
Carbon uptake rate of the Octopus Spring Synechococcus sp. at different temperatures
| Expt | Carbon uptake rate (cpm μg of Chl a−1 h−1) ata:
|
||
|---|---|---|---|
| 70°C | 65°C | 55°C | |
| 8 July 1995 (1st [morning]) | 2305.1 ± 214.04 (a) | 3921.6 ± 361.39 (b) | 1643.6 ± 121.75 (c) |
| 8 July 1995 (2nd [afternoon]) | 1449.4 ± 100.47 (a) | 1734.3 ± 141.04 (b) | 669.3 ± 51.73 (c) |
| 25 June 1996 | 3084.9 ± 222.34 (a) | 4319.6 ± 285.00 (b) | 1703.8 ± 64.79 (c) |
Values are means ± standard errors (n = 8 observations for each estimate). Different letters within an experiment indicate significantly different treatments in F[1,24] tests.
A similar pattern was observed for the experiment in which photosynthesis was measured for cells collected from the research site and incubated at three different sites along the Octopus Spring outflow channel. The temperature ranges of the sites were 67 to 74, (the collection site), 62 to 70, and 52 to 55°C. The mean uptake rates at these sites were 6,512.4 ± 229.46, 8,360.6 ± 144.37, and 954.6 ± 112.92 cpm μg of Chl a−1 h−1, respectively.
Photosynthesis versus irradiance curves and the interaction of temperature and irradiance.
The effect of irradiance was highly significant for all experiments (P < 0.001; Fig. 2). However, there was also a highly significant temperature-by-irradiance interaction term in these experiments (P < 0.01 in all cases), which indicates that cells incubated at different temperatures are responding differently to changes in solar irradiance, i.e., have different photosynthesis versus irradiance curves. To investigate this issue further, the temperature dependence of photosynthesis was estimated across irradiance treatments by
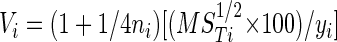 |
where ni is the number of observations at irradiance treatment i, MSTi is the among-temperature mean square for irradiance treatment i (i.e., the among-temperature variance component in a two-way ANOVA model for i), yi is the mean Chl-normalized carbon uptake at irradiance treatment i, and i = 0, 1, 2, and 3 neutral density screens, the four irradiance treatments. (1 + 1/4n) is a correction factor which removes bias from the estimate of V (33). V accounts for differences in mean carbon uptake among the irradiance treatments of an experiment by expressing the amount of variation as a percentage of the mean and is thus analogous to a coefficient of variation. Since increasing values of V indicate increasing disparity in photosynthetic performance among temperatures, V may therefore also be thought of as an estimate of the magnitude of temperature stress on photosynthesis outside of the optimal temperature range. V was calculated for all i within each of the three experiments and then treated as the dependent variable in an ANCOVA model with the experiment as the factor and mean visible irradiance as the covariate. Since the effect of the experiment was not significant (F[2,9] = 0.022), the data from the experiments were pooled, and these pooled data were regressed on mean visible irradiance. A second-degree polynomial fit the data significantly better than a straight line (F[1,12] = 31.862; P < 0.001), which indicates a curvilinear relationship between the temperature dependence index and visible irradiance (Fig. 3). This relationship suggests both that the carbon assimilation rate becomes increasingly temperature dependent with increasing irradiance at subsaturating irradiances and that this temperature dependence plateaus at higher irradiances.
FIG. 3.

Temperature dependence index (V) as a function of irradiance. Values of V correspond to the amount of variation in photosynthesis among temperatures expressed as a percentage of mean carbon assimilation rate and were plotted against solar irradiance for 8 July 1995 (first experiment [morning]) (○), 8 July 1995 (second experiment [afternoon]) (•), and 25 June 1996 (▿). The data were fit with a second-degree polynomial as described in the text.
The observed pattern of changing temperature effects with change in irradiance may reflect underlying differences among incubation temperatures in α (the initial slope of the photosynthesis versus irradiance curve at light-limiting irradiances), in Isat (the saturating irradiance for photosynthesis), or in both of these parameters. Estimates of α and Isat are provided in Table 2. No significant differences in these estimates, as determined by the overlap of 95% CIs, were found between UV+ and UV− treatments at any of the temperatures, and so these data were subsequently pooled to obtain single estimates of α and Isat for each temperature treatment within an experiment (Table 2).
TABLE 2.
Estimates of photosynthesis versus irradiance curve parameters α and Isat for Synechococcus sp. incubated at different temperatures
| Expt | Temp (°C) | UV treatment | Result for:
|
|
|---|---|---|---|---|
| α ± 95% CIa | Isat ± 95% CIb | |||
| 8 July 1995 (1st [morning]) | 70 | + | 11.0 ± 1.8 | 487.3 ± 108.7 |
| − | 16.9 ± 5.5 | 404.1 ± 96.2 | ||
| Pooled | 14.0 ± 3.1 | 438.9 ± 32.3 | ||
| 65 | + | 20.1 ± 4.3 | 524.6 ± 146.8 | |
| − | 18.4 ± 4.5 | 553.5 ± 215.5 | ||
| Pooled | 19.3 ± 2.7 | 553.0 ± 47.4 | ||
| 55 | + | 7.2 ± 1.7 | 483.3 ± 58.1 | |
| − | 10.6 ± 2.0 | 486.4 ± 67.9 | ||
| Pooled | 8.6 ± 1.9 | 481.3 ± 44.1 | ||
| 8 July 1995 (2nd [afternoon]) | 70 | + | 6.0 ± 1.5 | 541.0 ± 228.1 |
| − | 7.4 ± 1.4 | 576.0 ± 84.5 | ||
| Pooled | 6.7 ± 1.2 | 566.2 ± 59.8 | ||
| 65 | + | 7.4 ± 1.0 | 558.9 ± 141.6 | |
| − | 8.8 ± 1.1 | 571.8 ± 48.3 | ||
| Pooled | 8.1 ± 0.9 | 574.4 ± 41.0 | ||
| 55 | + | 2.9 ± 1.1 | 457.5 ± 150.5 | |
| − | 4.1 ± 0.5 | 523.6 ± 29.9 | ||
| Pooled | 3.5 ± 0.9 | 493.3 ± 53.1 | ||
| 25 June 1996 | 70 | + | 12.7 ± 2.9 | 563.6 ± 94.2 |
| − | 16.6 ± 3.1 | 549.1 ± 118.5 | ||
| Pooled | 14.6 ± 2.4 | 556.9 ± 34.6 | ||
| 65 | + | 19.6 ± 2.7 | 581.2 ± 90.9 | |
| − | 17.8 ± 1.6 | 655.9 ± 61.1 | ||
| Pooled | 18.6 ± 1.6 | 614.2 ± 29.1 | ||
| 55 | + | 7.7 ± 1.5 | 536.2 ± 59.0 | |
| − | 8.6 ± 1.3 | 557.4 ± 41.4 | ||
| Pooled | 8.2 ± 1.0 | 542.9 ± 22.2 | ||
Counts per minute per microgram of Chl a per hour per watt per square meter.
Watts per square meter.
α was greater in the 65°C treatment than at 70°C in the experiment of 25 June 1996, as indicated by nonoverlapping CIs (Table 2). A lower α at 70°C for the first experiment of 8 July 1995 (morning) is also suggested by nonoverlapping 90% CIs of the two temperature treatments. No difference was observed for the second experiment of 8 July 1995 (afternoon). α was always lower at 55°C than at 65 or 70°C.
The 95% CIs for estimates of Isat at 65 and 70°C for the first experiment of 8 July 1995 do not overlap, which indicates a higher saturating irradiance for photosynthesis at 65°C (Table 2). There is weaker evidence for a greater Isat at 65°C for 25 June 1996 provided by nonoverlapping 90% intervals. Isat at 65°C is greater than at 55°C for 25 June 1996, but estimates of Isat never differed between 70 and 55°C. No difference in Isat was found among any of the temperatures for the second experiment of 8 July 1995.
UV inhibition of photosynthesis.
Estimates of mean UVA and UVB fluxes and their associated standard errors were 30.3 ± 4.81 and 2.3 ± 0.38 W m−2, respectively, for the first experiment of 8 July 1995. These values were 34.7 ± 1.16 and 2.7 ± 0.12 W m−2 for the second experiment of 8 July 1995, and they were 38.8 ± 0.38 and 3.1 ± 0.13 W m−2 for the experiment of 25 June 1996.
UV inhibition data for the three temperatures are presented in Fig. 2 and Table 3. Since an interaction between irradiance and UV treatments was never observed (P > 0.20 in all cases), irradiance and UV treatments were pooled for a single estimate of UV inhibition at each temperature (see Materials and Methods). The three temperature treatments exhibited different susceptibilities to UV inhibition (Table 3). Octopus Spring cells were consistently inhibited by UV radiation at a suboptimal temperature (55°C). UV inhibition was observed at 70°C for the second experiment of 8 July 1995 and 25 June 1996 and at 65°C only for the second experiment of 8 July 1995. The UV effect at 65°C occurred only when UV inhibition was detected at the other two temperatures as well. No significant differences in magnitude of UV inhibition were observed at temperatures exhibiting UV-induced depression of carbon assimilation, with estimated means ranging from 13.5 to 35.4% (Table 3).
TABLE 3.
UV inhibition of carbon uptake by the Octopus Spring Synechococcus sp.
| Expt | % UV inhibition (mean ± 95% FI) at:
|
||
|---|---|---|---|
| 70°C | 65°C | 55°C | |
| 8 July 1995 (1st [morning]) | NSa | NS | 30.4 ± 12.6 |
| 8 July 1995 (2nd [afternoon]) | 18.9 ± 14.8 | 16.3 ± 10.7 | 35.4 ± 13.8 |
| 25 June 1996 | 19.9 ± 17.8 | NS | 13.5 ± 6.6 |
NS, FI overlaps zero.
Diurnal patterns of photosynthesis at Octopus Spring.
Figure 4 shows carbon uptake data obtained for experiments conducted on separate days in which cells were incubated at different times of day at the Octopus Spring collection site. Again, dark uptake values were very low compared with light-incubated values (Fig. 4). Diurnal variation in photosynthesis was indicated by a very highly significant effect of time of day (P < 0.0001) in both experiments. In the experiment of 24 June 1996 (Fig. 4A), this effect resulted from two sources. The first was a period of decreasing photosynthetic rate from an observed morning peak (Fig. 4; 9:40 > 11:00 > 12:20 by Bonferroni-Dunn tests) until the onset of a period of unchanging rate coincident with peak solar irradiance (Fig. 4A; 12:20 = 13:40 = 15:00 = 16:20 by Bonferroni-Dunn tests). The second was a decline in productivity during late afternoon (Fig. 4A; 16:20 > 17:35; P < 0.0001), at which time solar irradiance had fallen to levels found to be subsaturating for photosynthesis in the water bath experiments (Table 2). Data from the experiment on 2 July 1996 (Fig. 4B) are qualitatively similar to those from 24 June 1996 over the portion of the diel cycle where the two experiments overlap, although the difference between the observed maximum in carbon uptake and the afternoon depression was not as great. Both suggest a morning peak in photosynthetic rate and a period of a depressed but stable rate initiated near the onset of maximal solar irradiance, while neither shows a recovery of photosynthesis following this maximum. This pattern is also observed if data are normalized to cell number (data not shown).
FIG. 4.

Solar radiation data and Chl-normalized carbon uptake rate by Synechococcus sp. incubated at different times of day at the Octopus Spring research site on 24 June 1996 (A) and 2 July 1996 (B). Cell treatments were UV− (▿), UVB− (•), UV+ (○), and a dark control (▾). Error bars represent standard errors.
Diurnal patterns of UV inhibition at Octopus Spring.
A very highly significant effect of UV radiation was found in both experiments (P < 0.0001). In the experiment from 24 June 1996, total UV inhibition was detected and remained at a uniform, relative level until late afternoon (Fig. 4 and Table 4). This is confirmed both by the overlapping fiducial limits and by a lack of interaction between UV effect and incubation start time once the data between time points 1 through 5 were corrected for proportionality of UV treatments (Tukey’s test for nonadditivity [33]: F[1,7] = 11.73; P < 0.025) by logarithmic transformation (P > 0.10). The pooled estimate for total UV inhibition over time points 1 through 5 is 33.1% ± 5.3%, while that for UVB inhibition is 21.4% ± 6.6%. The subsequent disappearance of a UV effect in the late afternoon indicates an interaction between time of day and UV treatment (P < 0.0001). Similar magnitudes of total UV inhibition were seen in the experiment of 2 July 1996 (Fig. 4 and Table 4). The 95% fiducial limits for estimates of percent inhibition overlap for the four time points, and the pooled estimate is 37.2% ± 5.2%. A very highly significant interaction occurred between effect of UV-B and incubation start time (P < 0.001). This was due to lack of a significant effect at time point 2 as well as greater inhibition at time point 3. No recovery from UV inhibition was observed (Fig. 4).
TABLE 4.
Diurnal patterns of carbon uptake and UV inhibition of photosynthesis of the Octopus Spring Synechococcus sp. biofilm at 70°C
| Expt | Start time | % UV inhibition (mean ± 95% FI)a
|
|
|---|---|---|---|
| Total UV | UVB | ||
| 24 June 1996 | 9:40 | 31.5 ± 8.8 | 18.3 ± 12.5 |
| 11:00 | 37.7 ± 4.6 | 27.0 ± 6.2 | |
| 12:20 | 20.6 ± 19.1 | NS | |
| 13:40 | 35.9 ± 10.0 | NS | |
| 15:00 | 34.3 ± 15.2 | 27.5 ± 9.3 | |
| 16:20 | NSb | NS | |
| 17:35 | NS | NS | |
| 2 July 1996 | 7:50 | 32.9 ± 7.9 | 18.5 ± 7.1 |
| 9:50 | 36.0 ± 5.2 | NS | |
| 11:55 | 48.0 ± 14.8 | 35.1 ± 9.1 | |
| 13:55 | 33.8 ± 6.0 | 17.5 ± 7.7 | |
Percent reduction in uptake due to total UV or UVB inclusion.
NS, FI overlaps zero.
DISCUSSION
It has been hypothesized by Brock (2–4) that the hot spring Synechococcus sp. growing near the upper temperature limit for photosynthetic life has evolved to optimize photosynthesis and growth at the average temperature of the environment. However, our data indicate that cells of the Octopus Spring Synechococcus sp. collected at 70°C did not optimize photosynthesis at their mean environmental temperature, since photosynthesis was greater at 65°C (Table 1). This result was consistent across the 1995 and 1996 field seasons (Table 1). In addition, an increase in photosynthetic rate at temperatures lower than that of the collection site was obtained not only at constant water bath temperatures but also under periodically fluctuating temperature conditions in Octopus Spring. These data agree with those obtained for a laboratory clone of the Synechococcus high-temperature ecotype, for which photosynthesis and growth rate had an identical optimal range of 63 to 67°C, lower than the temperature of the environment from which the clone had been isolated (>70°C) (25).
While it is evident that this biofilm is under high-temperature stress, it is possible that these Synechococcus cells may benefit from life under nearly lethal conditions by avoiding competition with other photoautotrophs. Support for this interpretation is provided by a study of growth temperature ranges of temperature strains of Synechococcus sp. from Hunter’s Hot Springs, Oreg. (31). The clone capable of growing at 70°C or above grew best at ∼65°C, as found by Meeks and Castenholz (25), but its growth rate at this temperature was lower than that of other clones that could grow at 65°C but not at 70°C. This observation suggests that the strain capable of growing at the upper temperature limit would be outcompeted in nature at its optimal growth temperature. If this strain evolved from an ancestral Synechococcus sp. which grew at lower temperatures, it is possible that it has sacrificed growth rate at its optimum in the process of extending its temperature range to higher temperatures.
Data from Ferris and Ward (11) may support the above suggestion. In their profiles of microbial community structure at Octopus Spring using denaturing gradient gel electrophoresis of 16S rRNA gene fragments amplified from mat DNA, only a single cyanobacterial sequence type (A′) was recovered between 68 and 72°C during the summer of 1995, and this sequence is closely related to those recovered at lower temperatures (11). Still, more information is needed to evaluate the competition avoidance hypothesis presented above.
The impact of nonoptimal temperature on photosynthesis depends upon irradiance level (Fig. 3), but since this interaction represents only changing relative magnitudes of temperature effects with irradiance, it does not complicate the conclusion that 70 and 55°C are supraoptimal and suboptimal for photosynthesis, respectively. Similarly, Ibelings (18) found that elevated temperatures were more damaging to photosynthesis of a planktonic cyanobacterial surface bloom when cells were simultaneously exposed to high irradiance.
Solar data collected during this study (e.g., Fig. 4) indicate that these cells can be exposed for much of the day to light conditions under which supraoptimal environmental temperature has a great effect on photosynthesis (Fig. 3). This is due in part to the elevation of the site (∼2,450 m) as well as to the direct exposure resulting from the exclusion of shading vegetation by the high surrounding soil temperature. Consequently, cells of Synechococcus sp. maintain lower cell Chl a contents during the summer than cells acclimated to lower irradiances and thus appear yellow throughout the vertical profile of the biofilm (5).
This study has uncovered two possible causes of this interaction between temperature and irradiance. The first is a lower α, i.e., decreased quantum efficiency at subsaturating irradiances, at both supraoptimal and suboptimal temperatures (Table 2). Meeks and Castenholz (26, 27) also found a lower α at supraoptimal temperature in Synechococcus sp. clone H-Xf and indicated that this is likely the result of lowered RUBISCO activity at a nearly lethal temperature. A lower α at supraoptimal temperature has also been observed in sea-ice diatoms in McMurdo Sound, Antarctica (30), and in the kelp Laminaria saccharina (8). There was also an indication that cells at 70°C may saturate photosynthesis at lower irradiances than they would under more optimal conditions (Table 2), which would result in greater susceptibility to photoinhibition as a result of becoming exposed to excess light energy at lower irradiances.
Cells were also more susceptible to UV inhibition of photosynthesis at nonoptimal temperatures (Table 3). Inhibition has been widely reported for oxygenic phototrophs from a variety of habitats (16). As with visible wavelengths, the Octopus Spring high-temperature Synechococcus sp. is subjected to high summertime fluence rates of UV radiation, with UVA and UVB irradiance levels exceeding 40 and 3 W m−2, respectively, at peak irradiance on clear days (e.g., Fig. 4). These cells also lack UV-protective compounds, such as scytonemin, which has been shown to protect cells from UVA (320 to 400 nm) (12, 13), and mycosporine-like amino acids, the presence of which is positively correlated with UVB (280 to 320 nm) resistance in cyanobacteria (14, 15). Thus, an additional consequence of life at nearly lethal temperature (70°C) appears to be greater susceptibility to UV-induced reduction in photosynthesis.
Photosynthesis in the Octopus Spring biofilm at 70°C varied diurnally (Fig. 4). Such variation has also been observed in both freshwater and marine phytoplankton communities (9, 23, 24, 29, 32, 34–36). Most studies found mid- to late-morning peaks in photosynthetic activity, although Newhouse et al. (29) reported an afternoon maximum for neritic plankton off Waikiki Beach, and recovery of photosynthetic activity generally occurred overnight or early the next morning. In this study, no recovery was observed during the course of either experiment. Maximal photosynthesis was observed as early as 8:00, but solar irradiance had already attained levels nearing the estimated saturating irradiance for Synechococcus sp. at 70°C by this time (Table 2). Whether recovery in this biofilm takes place overnight and/or early the next morning under subsaturating irradiances is yet to be determined.
This is the first study to partition patterns of diurnal productivity into visible irradiance and UV radiation components. UV inhibition of photosynthesis by both UVA and UVB wavebands was found in both experiments. In the experiment of 24 June 1996, UV inhibition disappeared in the late afternoon, the same time that visible irradiance became light limiting for photosynthesis, which suggests that UV inhibition may not occur unless light is saturating. Thus, this may be evidence of a UV × irradiance interaction which we were not able to detect in our water bath experiments (see Results). If this is the case, it is possible that these cells may have a lower saturating irradiance for photosynthesis under high-temperature stress and thus be more susceptible to UV inhibition.
In summary, these results have implications for our understanding of the evolution of thermal tolerance in the cyanobacteria. Since many nonphotosynthetic prokaryotes can grow above 73°C, Brock (2) suggested that the critical upper temperature for photosynthetic life is determined by an inherent limitation in the stability of the photosynthetic apparatus which constrains evolution and thus has prevented the invasion of habitat exceeding 73°C by photosynthetic microorganisms. It was also proposed by Brock (2) that the high-temperature Synechococcus sp. optimizes growth and photosynthesis at temperatures up to this critical boundary, with the implication that temperatures above the optimum are lethal. This type of temperature response profile has been described for the motile thermophilic cyanobacterium Oscillatoria terebriformis (6). In contrast, evidence presented in this study and by Meeks and Castenholz (25) indicates that photosynthesis by the high-temperature ecotype of Synechococcus sp. declines gradually over a range of supraoptimal temperatures before reaching the critical limit. The evolution of the temperature response profile of photosynthesis in the high-temperature Synechococcus sp. therefore appears to have taken a different path from that proposed by Brock (2).
ACKNOWLEDGMENTS
We are grateful to Miriam E. Martin for assistance during the 1996 field season and to John Kelly and John Willis for helpful discussion. We also thank Richard Wiegert for the use of the University of Georgia research trailer in West Yellowstone, Montana, and the Yellowstone Center for Resources, Yellowstone National Park, for permission to conduct research within the park. We thank three anonymous reviewers for their comments.
This work was supported by U.S. National Science Foundation grants IBN-9219273 and IBN-9630674 to R.W.C.
REFERENCES
- 1.Abacus Concepts. SuperANOVA. Berkeley, Calif: Abacus Concepts, Inc.; 1989. [Google Scholar]
- 2.Brock T D. Micro-organisms adapted to high temperatures. Nature. 1967;214:882–885. doi: 10.1038/214882a0. [DOI] [PubMed] [Google Scholar]
- 3.Brock T D. Thermophilic microorganisms and life at high temperatures. New York, N.Y: Springer-Verlag; 1978. [Google Scholar]
- 4.Brock T D, Brock M L. Relationship between environmental temperature and optimum temperature of bacteria along a hot spring thermal gradient. J Appl Bacteriol. 1968;31:54–58. [Google Scholar]
- 5.Brock T D, Brock M L. Effect of light intensity on photosynthesis by thermal algae adapted to natural and reduced sunlight. Limnol Oceanogr. 1969;14:334–341. [Google Scholar]
- 6.Castenholz R W. Thermophilic blue-green algae and the thermal environment. Bacteriol Rev. 1969;33:476–504. doi: 10.1128/br.33.4.476-504.1969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Castenholz R W. Endemism and biodiversity of thermophilic cyanobacteria. Nova Hedwigia. 1996;112:33–47. [Google Scholar]
- 8.Davison I R. Environmental effects on algal photosynthesis: temperature. J Phycol. 1991;27:2–8. [Google Scholar]
- 9.Doty M S, Oguri M. Evidence for a photosynthetic daily periodicity. Limnol Oceanogr. 1957;2:37–40. [Google Scholar]
- 10.Falk S, Maxwell D P, Laudenbach D E, Hunter N P A. Photosynthetic adjustment to temperature. In: Baker N R, editor. Photosynthesis and the environment. Vol. 5. Dordrecht, The Netherlands: Kluwer Academic Publishers; 1996. pp. 367–385. [Google Scholar]
- 11.Ferris M J, Ward D M. Seasonal distributions of dominant 16S rRNA-defined populations in a hot spring microbial mat examined by denaturing gradient gel electrophoresis. Appl Environ Microbiol. 1997;63:1375–1381. doi: 10.1128/aem.63.4.1375-1381.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Garcia-Pichel F, Castenholz R W. Characterization and biological implications of scytonemin, a cyanobacterial sheath pigment. J Phycol. 1991;27:395–409. [Google Scholar]
- 13.Garcia-Pichel F, Sherry N D, Castenholz R W. Evidence for an ultraviolet sunscreen role of the extracellular pigment scytonemin in the terrestrial cyanobacterium Chlorogloeopsis sp. Photochem Photobiol. 1992;56:17–23. doi: 10.1111/j.1751-1097.1992.tb09596.x. [DOI] [PubMed] [Google Scholar]
- 14.Garcia-Pichel F, Castenholz R W. Occurrence of UV-absorbing, mycosporine-like compounds among cyanobacterial isolates and an estimate of their screening capacity. Appl Environ Microbiol. 1993;59:163–169. doi: 10.1128/aem.59.1.163-169.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Garcia-Pichel F, Wingard C E, Castenholz R W. Evidence regarding the UV sunscreen role of a mycosporine-like compound in the cyanobacterium Gloeocapsa sp. Appl Environ Microbiol. 1993;59:170–176. doi: 10.1128/aem.59.1.170-176.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Holm-Hansen O, Lubin D, Helbling E W. Ultraviolet radiation and its effects on organisms in aquatic environments. In: Young A R, Björn L, Mohan J, Nultsch W, editors. Environmental UV photobiology. New York, N.Y: Plenum Press; 1993. [Google Scholar]
- 17.Hubert J J. Bioassay. 3rd ed. Dubuque, Iowa: Kendall/Hunt Publishing Co.; 1992. [Google Scholar]
- 18.Ibelings B W. Changes in photosynthesis in response to combined irradiance and temperature stress in cyanobacterial surface waterblooms. J Phycol. 1996;32:549–557. [Google Scholar]
- 19.Jandel Scientific. SigmaPlot. San Rafael, Calif: Jandel Corporation; 1995. [Google Scholar]
- 20.Jassby A D, Platt T. Mathematical formulation of the relationship between photosynthesis and light for phytoplankton. Limnol Oceanogr. 1976;21:540–547. [Google Scholar]
- 21.Leegood R C, Edwards G E. Carbon metabolism and photorespiration: temperature dependence in relation to other environmental factors. In: Baker N R, editor. Photosynthesis and the environment. Vol. 5. Dordrecht, The Netherlands: Kluwer Academic Publishers; 1996. pp. 191–221. [Google Scholar]
- 22.Lenz J, Zeitschel B. Zur Bestimmung des Extinktionkoeffizienten für Chlorophyll a in Methanol. Kiel Meeresforsch. 1968;24:41–50. [Google Scholar]
- 23.Lorenzen C J. Diurnal variation in photosynthetic activity of natural phytoplankton populations. Limnol Oceanogr. 1963;8:56–62. [Google Scholar]
- 24.McAllister C D. Measurements of diurnal variation in productivity at Ocean Station “P.”. Limnol Oceanogr. 1963;8:289–292. [Google Scholar]
- 25.Meeks J C, Castenholz R W. Growth and photosynthesis in an extreme thermophile, Synechococcus lividus (Cyanophyta) Arch Mikrobiol. 1971;78:25–41. doi: 10.1007/BF00409086. [DOI] [PubMed] [Google Scholar]
- 26.Meeks J C, Castenholz R W. Photosynthetic properties of the extreme thermophile Synechococcus lividus. I. Effect of temperature on fluorescence and enhancement of CO2 assimilation. J Therm Biol. 1978;3:11–18. [Google Scholar]
- 27.Meeks J C, Castenholz R W. Photosynthetic properties of the extreme thermophile Synechococcus lividus. II. Stoichiometry between oxygen evolution and CO2 assimilation. J Therm Biol. 1978;3:19–24. [Google Scholar]
- 28.Miller, S. R., C. E. Wingard, and R. W. Castenholz. Unpublished data.
- 29.Newhouse J, Doty M S, Tsuda R T. Some diurnal features of a neritic surface plankton population. Limnol Oceanogr. 1967;12:207–212. [Google Scholar]
- 30.Palmisano A C, SooHoo J B, Sullivan C W. Effects of four environmental variables on photosynthesis-irradiance relationships in Antarctic sea-ice microalgae. Mar Biol. 1987;94:299–306. [Google Scholar]
- 31.Peary J A, Castenholz R W. Temperature strains of a thermophilic blue-green alga. Nature. 1964;202:720–721. [Google Scholar]
- 32.Shimada B M. Diurnal fluctuations in photosynthetic rate and chlorophyll “a” content of phytoplankton from Eastern Pacific waters. Limnol Oceanogr. 1958;3:336–339. [Google Scholar]
- 33.Sokal R R, Rohlf F J. Biometry: the principles and practice of statistics in biological research. 3rd ed. New York, N.Y: W. H. Freeman; 1995. [Google Scholar]
- 34.Taguchi S. Short-term variability of photosynthesis in natural marine phytoplankton populations. Mar Biol. 1976;37:197–207. [Google Scholar]
- 35.Verduin J. Daytime variations in phytoplankton photosynthesis. Limnol Oceanogr. 1957;2:333–336. [Google Scholar]
- 36.Vincent W F, Neale P J, Richerson P J. Photoinhibition: algal responses to bright light during diel stratification and mixing in a tropical alpine lake. J Phycol. 1984;20:201–211. [Google Scholar]