

Abstract
Understanding functional connectivity of the amygdala with other brain regions, especially task modulated connectivity, is a critical step toward understanding the role of the amygdala in emotional processes and the interactions between emotion and cognition. The present study performed coordinate-based meta-analysis on studies of task modulated connectivity of the amygdala which used psychophysiological interaction (PPI) analysis. We first analyzed 49 PPI studies on different types of tasks using activation likelihood estimation (ALE) meta-analysis. Widespread cortical and subcortical regions showed consistent task modulated connectivity with the amygdala, including the medial frontal cortex, bilateral insula, anterior cingulate, fusiform gyrus, parahippocampal gyrus, thalamus, and basal ganglia. These regions were in general overlapped with those showed coactivations with the amygdala, suggesting that these regions and amygdala are not only activated together, but also show different levels of interactions during tasks. Further analyses with subsets of PPI studies revealed task specific functional connectivities with the amygdala that were modulated by fear processing, face processing, and emotion regulation. These results suggest a dynamic modulation of connectivity upon task demands, and provide new insights on the functions of the amygdala in different affective and cognitive processes. The meta-analytic approach on PPI studies may offer a framework toward systematical examinations of task modulated connectivity.
Keywords: Amygdala, Connectivity, Dynamic connectivity, fMRI, Psychophysiological interaction, Task modulation
Introduction
The amygdala is a critical structure in the brain which has extensive and widespread connections with cortical regions (Young et al. 1994). Anatomically, the amygdala is anteriorly connected to the orbital frontal lobe and anterior temporal lobe via the uncinate fasciculus, posteriorly connected to the occipital and temporal lobes via the inferior longitudinal fasciculus, and bilaterally interconnected via the anterior commissure (Catani and Thiebaut de Schotten 2008; Catani et al. 2002). Functionally, the amygdala is mainly involved in emotional processing (Adolphs et al. 1994; Phan et al. 2002), especially for fear and disgust (Costafreda et al. 2008). It also interacts with other brain systems to affect different levels of cognitive processes from unconscious visual processing (Morris et al. 1998; Whalen et al. 1998), attention (Anderson and Phelps 2001), face processing (Mende-Siedlecki et al. 2013), to memory (Adolphs et al. 2005; Sarter and Markowitsch 1985), social judgment (Adolphs et al. 1998), and decision-making (Bechara et al. 2003) [see Phelps (2006) for a review]. To understand the functions of amygdala in these diverse cognitive and affective processes, it is critical to study the task modulated connectivity between the amygdala and other brain regions that involved in different processes (Pessoa 2014).
Many studies have demonstrated spatially restricted regions that showed functional connectivity with the amygdala in resting-state, including the parahippocampus, anterior cingulate, and orbital frontal cortex (Hahn et al. 2011; Lowe et al. 1998; Ma et al. 2010; Sripada et al. 2012). However, Roy et al. demonstrated widely distributed regions that showed positive or negative functional connectivity with the amygdala (Roy et al. 2009). The discrepancy may be due to that Roy et al. (2009) applied global signal regression, which shifts the overall distribution of correlations (Murphy et al. 2009; Saad et al. 2012). Although these studies on brain anatomical connectivity and endogenous connectivity have portrayed the infrastructure of brain communications, it is still largely unknown how different cognitive and affective processes modulate amygdala connectivity. Because maintaining functional communications is costly in terms of energy consumption, certain functional communication may be dynamic and only take place upon task demands (Bullmore and Sporns 2012). We therefore hypothesize that upon task request, the amygdala will be dynamically connected to regions that are involved in the task.
The amygdala is usually coactivated by different tasks with various brain regions. For example, emotion processing generally involves the amygdala as well as the medial prefrontal cortex and basal ganglia (Phan et al. 2002; Vytal and Hamann 2010). The amygdala, together with the fusiform gyrus and superior temporal sulcus generally participate in face processing (Laird et al. 2015; Mende-Siedlecki et al. 2013). Along this line, a study of coactivations of the amygdala from hundreds of studies has shown widespread regions in the brain that correspond well with animal studies of amygdala structural connectivity (Robinson et al. 2010). However, co-occurrence of two regions in one study doesn’t necessarily mean these regions have increased connectivity during the task. It is also possible that the two regions are activated by the same task but work independently to each other. Direct modeling of task related connectivity is necessary to examine whether the coactivated regions are more functionally connected during a task.
Psychophysiological interaction (PPI) is a widely used method to study task related functional connectivity on fMRI data (Friston et al. 1997). This method uses linear regression model to examine the interaction between a time series of a seed region (physiological variable) and a task (psychological variable). Using voxel-wise analysis, regions that showed task modulated connectivity with the seed region could be identified. Later modification includes adding deconvolution to take into account of hemodynamic response delays (Gitelman et al. 2003). The PPI method was validated by several simulation studies (Kim and Horwitz 2008; McLaren et al. 2012), especially for block designed experiments (Cisler et al. 2013). Using the amygdala as a seed region, a large number of studies have already examined amygdala connectivity that is modulated by variety of tasks. This enables us to systematically study task modulated connectivity of the amygdala, and especially how different tasks modulate amygdala connectivity with different brain regions.
In the current study, we used a meta-analytic approach to synthesize published PPI studies on task modulated amygdala connectivity. Specifically, we performed coordinate-based meta-analysis using activation likelihood estimation (ALE) (Turkeltaub et al. 2002). ALE is a voxel-wise meta-analytic approach to identify brain regions that were consistently reported in published papers. It treats reported lists of coordinates as a distribution of “activation” probabilities, and estimates likelihood of “activations” across studies. We adopted the ALE method in the current study to estimate brain regions that consistently conveyed task modulated connectivity with the amygdala in published literatures. We first ask whether there are regions that demonstrated consistent task modulated functional connectivity with the amygdala across studies. Next, we retrospectively grouped these PPI studies into specific task domains based on the numbers of available studies in a domain. We identified three task domains, fear processing, face processing, and emotion regulation. In addition to the involvement of amygdala in the three task domains, each task domain also recruits different sets of brain regions. Fear processing showed consistent activations in the insula, anterior cingulate cortex, and fusiform gyrus (Vytal and Hamann 2010). Face processing has been shown to recruit distributed networks upon task paradigms (Laird et al. 2015). And emotion regulation recruits the supplementary motor area, inferior frontal gyrus, and middle temporal cortex (Frank et al. 2014; Kohn et al. 2014). We performed meta-analysis separately on the three task domains, and predict that different tasks would modulate amygdala connectivity to different brain regions that are involved in the corresponding tasks.
Materials and methods
Article selection
Initial literature searches were conducted using PubMed and Google Scholar in September 2014. Two searches were performed using each search engine, (1) exact phrase “psychophysiological interaction” combined with “amygdala”, and (2) exact phrase “psycho-physiological interaction” combined with “amygdala”. Both fMRI and positron emission tomography (PET) studies were included. The PubMed search returned 48 and 2 papers, respectively, while, the Google Scholar search returned 739 and 120 papers, respectively. Papers published after the initial searches were identified by PubMed generated RSS (rich site summary) feeds and Google Scholar Alerts using the same keywords. At a later stage, we also searched the abbreviation term “PPI” combined with “amygdala” in PubMed for potential missing papers. All papers were carefully examined, and then were included or excluded for subsequent meta-analysis based on the following criteria. (1) They were research articles, but not reviews, conference abstracts or theses. (2) The amygdala was used as the seed in PPI analysis. (3) Results represented mentally healthy human subjects. If a study analyzed two groups of subjects, e.g., a group of patients and a group of healthy controls, the study would be included if the results of healthy controls were independently reported. (4) The studies that examined within subject effects (between experiment manipulations) but not group differences (genotypes, individual differences) were included. (5) The studies that explicitly reported stereoscopic coordinates were included. (6) The studies were excluded if results were reported with small volume correction (SVC). As a result, 49 papers were identified in total (Table 1).
Table 1.
List of papers that are included in the meta-analysis of psychophysiological interaction analysis of the amygdala
| Study# | First author | Year | n | Modality | Task domain | Seeds |
|---|---|---|---|---|---|---|
|
| ||||||
| 1 | Akitsuki | 2009 | 23 | fMRI | Empathy | Left |
| 2 | Amting | 2009 | 15 | fMRI | Cognitive control | Averaged |
| 3 | Amting | 2010 | 16 | fMRI | Emotion processing | Right |
| 4 | Banks | 2007 | 14 | fMRI | Emotion regulation | Left |
| 5a | Bruneau | 2015 | 18 | fMRI | Empathy | Averaged |
| 5b | Bruneau | 2015 | 21 | fMRI | Empathy | Averaged |
| 6 | Cohen | 2008 | 15 | fMRI | Learning | Averaged |
| 7 | Comte | 2014 | 33 | fMRI | Emotion processing | Right |
| 8 | Decety | 2011 | 22 | fMRI | Perspective taking | Left |
| 9 | Erk | 2010 | 17 | fMRI | Emotion regulation | Left |
| 10 | Fakra | 2008 | 14 | fMRI | Emotion processing | Both |
| 11 | Foland | 2008 | 9 | fMRI | Emotion processing | Left |
| 12 | Gianaros | 2012 | 97 | fMRI | Cognitive control | Right |
| 13 | Gold | 2014 | 17 | fMRI | Conditioning | Right |
| 14 | Grabenhorst | 2013 | 13 | fMRI | Food choice | Right |
| 15 | Hermans | 2012 | 18 | fMRI | Emotion processing | Averaged |
| 16 | Herrmann | 2015 | 38 | fMRI | Emotion processing | Right |
| 17 | Iidaka | 2001 | 12 | fMRI | Emotion processing | Both |
| 18 | Kanske | 2011 | 30 | fMRI | Emotion regulation | Left |
| 19 | Kienast | 2008 | 13 | fMRI | Emotion processing | Left |
| 20 | Koelsch | 2013 | 18 | fMRI | Emotion processing | Both |
| 21 | Larson | 2009 | 17 | fMRI | Shape processing | Left |
| 22 | Maquet | 1998 | 7 | PET | Sleep | Both |
| 23 | Meier | 2015 | 13 | fMRI | Emotion processing | Averaged |
| 24 | Mende-Siedlecki | 2013 | 215 | fMRI | Face processing | Both |
| 25 | Molapour | 2015 | 20 | fmri | Conditioning | Right |
| 26 | Monk | 2008 | 12 | fMRI | Emotion processing | Right |
| 27 | Morris | 1998 | 5 | PET | Emotion processing | Left |
| 28 | Morris | 2001 | 10 | PET | Food picture processing | Left |
| 29 | Mukherjee | 2014 | 24 | fMRI | Social judgment | Both |
| 30 | Murray | 2014 | 17 | fMRI | Memory encoding | Left |
| 31 | Pasley | 2004 | 7 | fMRI | Face processing | Left |
| 32 | Pichon | 2012 | 20 | fMRI | Emotion processing | Right |
| 33 | Ponz | 2010 | 9 | fMRI | Conditioning | Right |
| 34 | Sato | 2016 | 35 | fMRI | Emotion processing | Right |
| 35 | Schienle | 2013 | 34 | fMRI | Emotion processing | Both |
| 36 | Schmitgen | 2016 | 21 | fMRI | Theory of mind | Right |
| 37 | Skelly | 2012 | 18 | fMRI | Face processing | Both |
| 38 | Spielberg | 2014 | 41 | fMRI | Emotion processing | Both |
| 39 | Sripada | 2014 | 49 | fMRI | Emotion regulation | Right |
| 40 | Stegmayer | 2014 | 18 | fMRI | Working memory | Right |
| 41 | Sterpenich | 2006 | 30 | fMRI | Memory retrieval | Right |
| 42 | Stevens | 2013 | 20 | fMRI | Emotion processing | Both |
| 43 | Tottenham | 2012 | 25 | fMRI | Face processing | Left |
| 44 | Troiani | 2014 | 12 | fMRI | Face processing | Left |
| 45 | van Wingen | 2011 | 57 | fMRI | Emotion processing | Right |
| 46 | Williams | 2006 | 15 | fMRI | Emotion processing | Both |
| 47 | Winecoff | 2008 | 42 | fMRI | Emotion regulation | Both |
| 48 | Xu | 2013 | 20 | fMRI | Decision-making | Averaged |
| 49 | Yoder | 2015 | 43 | fMRI | Violence processing | Rightb |
‘Left’ and ‘right’ mean that the papers used the left or right amygdala as a seed region in their analysis, respectively. ‘Both’ means the papers used both left and right amygdala seeds as seeds in their analysis. While ‘averaged’ means only one seed that averaged between the left and right amygdala was used in their analyses
Basolateral complex of the amygdala (BLA) and central subnucleus (CE) were analyzed separately in this paper. The coordinates of the two seeds were pooled in the current analysis
It should be noted that studies using SVC are usually included in meta-analysis if such studies constitute only a small portion of the included studies. However, in the case of PPI analysis, since the effect size of an interaction is typically smaller than the main effect of task activation, a large proportion of PPI papers utilized SVC. Including these papers would bias the results toward subjectively selected regions. Therefore, we did not include any papers that used SVC in the current meta-analysis.
Activation likelihood estimation analysis
Some studies make multiple comparisons of different experimental effects from the same sample of subjects. To avoid bias from multiple contrasts in one study, we chose the one that is supposed to have larger effect to include in the current analysis. For example, Williams et al. (2006) compared fearful and neutral facial expressions in both conscious and unconscious conditions (Williams et al. 2006), and we used the contrast in the conscious condition in the current analysis. The specific contrasts used in the current analysis are listed in the supplementary materials. Bruneau et al. (2015) recruited two independent samples in their studies 1 and 2, therefore, were treated as two independent studies. The included papers used anatomically or functionally defined left or right amygdala as seeds, or averaged the left and right regions as one seed. Because the main goal of the current study is to examine task modulated connectivity between any parts of the amygdala and other brain regions, we pooled left and right amygdala results together if separate results were reported. We later performed separate analysis on left and right amygdala PPI effects to explore potential lateralized effects. Lastly, one study used the basolateral amygdala (BLA) and central nucleus (CE) as separate seeds (Yoder et al. 2015). The results of these two subnuclei were pooled in the current analysis.
Ginger ALE 2.3 (Laird et al. 2005a, b) was used for ALE analysis, with random effects model (Eickhoff et al. 2009), and non-additive algorithm for within contrast calculation (Turkeltaub et al. 2012). The analyses were performed in MNI space (Montreal Neurological Institute). For papers that reported results in Talairach space (Talairach and Tournoux 1988), the coordinates were converted into MNI space using Lancaster’s transformation (Laird et al. 2010; Lancaster et al. 2007). First, we asked which regions in the brain showed consistent task modulated connectivity with the amygdala. Since the PPI effects are a relative measure of connectivity differences between two task conditions, positive and negative effect are relative and somehow arbitrary. Therefore, we pooled the coordinates of positive and negative effects together regardless of the signs to examine regions that showed consistent task modulated connectivity with the amygdala. A total of 550 foci of 50 independent experiments from the 49 papers were used for this ALE analysis (see supplementary Table S1 for a full list of all contrasts). The resulting map was first approached using a uncorrected threshold p < 0.01, and a cluster-level threshold p < 0.05 was used to identify significant clusters (Eickhoff et al. 2012).
Since the functional lateralization of the amygdala has been well documented (Baas et al. 2004; Lanteaume et al. 2007), it is possible that the left and right amygdala show different patterns of task modulated connectivity. To explore this, we performed separate ALE analyses on the left and right amygdala and contrast analysis between them. 26 experiments that used the left amygdala and 29 experiments that used the right amygdala as seeds were used to perform subtraction ALE analysis (see supplementary Tables S2 and S3 for the list of all the experiments). The same statistical threshold of uncorrected p < 0.01 combined with cluster-level p < 0.05 was used to determine significant results.
Next, we examined the task modulations of amygdala connectivity for specific task domains. Two authors (DX, HJ) independently identified the specific cognitive processes by carefully checking the exact meaning of each contrast in each study and classified them into three groups, i.e., fear processing, face processing, and emotion regulation. The classification criteria were as follows: (a) For “fear processing”, 14 studies were included (paper #: 3, 13, 15, 20, 23, 26, 27, 31, 32, 38, 42, 43, 45, and 46). Classification criteria were that these studies compared PPI effects between fearful or threatening stimuli conditions with neutral or positive stimuli conditions. For such a specific effect, we were able to differentiate positive and negative effects. Twelve experiments of 95 foci reported increased connectivity with the amygdala during fear processing, while 8 experiments of 40 foci reported decreased connectivity with the amygdala (see supplementary Table S4 for a full list of all contrasts). ALE analyses of positive and negative effects were conducted separately. (b) For “face processing”, six papers were included (paper #: 24, 31, 37, 38, 44, and 45). Classification criteria were that these studies compared PPI effects between face stimuli conditions with other stimuli conditions. Five experiments of 70 foci showed positive modulations of face processing on connectivity with the amygdala (see supplementary Table S5 for a full list of all contrasts). Only two experiments conveyed negative modulations of face processing on connectivity with the amygdala. Therefore, only ALE analysis on positive effects was conducted. (c) For “emotion regulation”, five papers were included (paper #: 4, 9, 18, 39, and 47). Classification criteria were that these studies compared PPI effects between reappraise conditions with maintain or distraction conditions. Five experiments of 49 foci from the five papers showed greater connectivity with the amygdala in the reappraisal condition compared with the maintain condition (see supplementary Table S6 for a full list of all contrasts). Only one contrast demonstrated less connectivity with the amygdala in the reappraisal condition than in the maintain condition. Therefore, the negative effect was not analyzed. Statistical significant results were identified using a combined threshold of uncorrected p < 0.01 and cluster-level p < 0.05. We noted that because the numbers of included studies were small, occasionally some resultant clusters were only contributed by one single study. Therefore, we only reported clusters that were contributed by two or more studies.
Coactivation analysis of the amygdala
Our hypothesis is that the regions that are commonly activated with the amygdala will show task modulated connectivity with the amygdala. To directly demonstrate the overlap, we also performed coactivation analysis, which examined co-occurrence of brain regions with the amygdala in different tasks. The analysis is an updated version Robinson et al. (2010). Studies with activations within the amygdala were searched in the BrainMap database using Sleuth 2.3.2 (Laird et al. 2005a, b). The amygdala was defined by Talairach atlas in Sleuth. 8064 coordinates from 662 experiments of 418 studies were identified that reported clusters within the amygdala mask. These coordinates were used to perform an ALE analysis. The same statistical threshold of uncorrected p <0.01 combined with cluster-level p <0.05 was used to identify significant clusters.
Results
Overall task modulations of amygdala connectivity
The first ALE analysis of all 49 studies with pooled positive and negative results indicated widespread cortical and subcortical regions that showed consistent task modulated connectivity with the amygdala (Fig. 1; Table 2). These regions included the medial frontal gyrus (Brodmann’s area, BA 6/32), anterior cingulate (BA 32), bilateral inferior frontal gyrus/insula (BA 47/13), thalamus, basal ganglia nuclei (putamen, medial globus pallidus, and caudate), left middle temporal gyrus (BA 21), bilateral fusiform gyrus (BA 37) extending to the cerebellum, right parahippocampal gyrus/hippocampus (BA 36), and left middle/superior frontal gyrus (BA 10). The numbers of studies that contributed to each cluster ranged from 3 to 13 out of the total 50 independent experiments (see Table 2 for details).
Fig. 1.
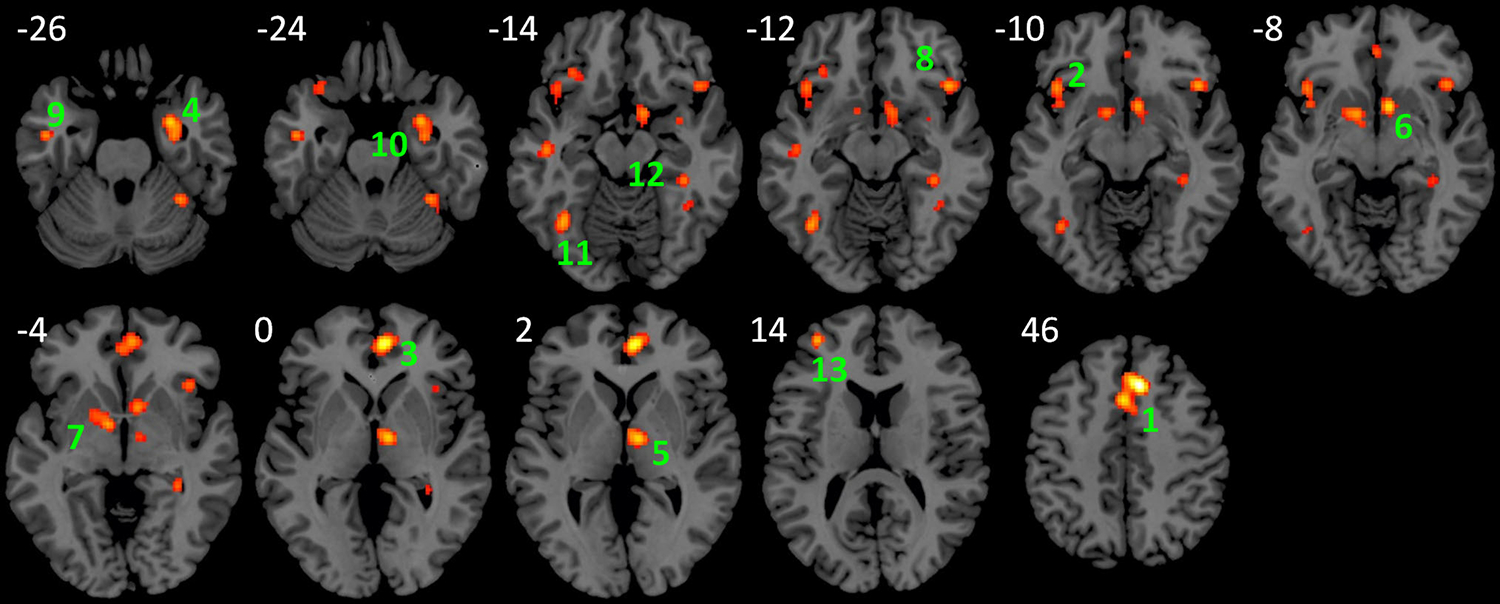
Regions that showed consistent task modulation of amygdala connectivity across 50 experiments of PPI analyses using the amygdala as seeds. The activation likelihood map was approached at uncorrected p < 0.01 and a cluster-level p < 0.05. Numbers near each slice represent MNI (Montreal Neurological Institute) z coordinates
Table 2.
Regions that showed task modulated connectivity with the amygdala
| Cluster# | Volume (mm3) | Extrema | x | y | z | Label | Contributed studies |
|---|---|---|---|---|---|---|---|
|
| |||||||
| 1 | 3424 | 0.028085 | 8 | 24 | 46 | Medial Frontal Gyrus, BA 6 | 1, 5b, 6, 7, 8, 18, 23, 24, 37, 39, 40, 45, 49 |
| 0.020473 | −2 | 14 | 46 | Medial Frontal Gyrus, BA 32 | |||
| 0.017329 | 4 | 8 | 54 | Medial Frontal Gyrus, BA 6 | |||
| 0.011548 | 8 | 26 | 36 | Cingulate Gyrus, BA 32 | |||
| 2 | 1960 | 0.016587 | −42 | 20 | −10 | L. Inferior Frontal Gyrus, BA 47 | 6, 8, 13, 18, 26, 35, 37, 43, 44 |
| 0.014766 | −40 | 20 | −20 | L. Inferior Frontal Gyrus, BA 47 | |||
| 0.014041 | −32 | 30 | −14 | L. Inferior Frontal Gyrus, BA 47 | |||
| 0.013599 | −40 | 12 | −8 | L. Insula, BA 13 | |||
| 3 | 1832 | 0.024385 | 6 | 48 | 0 | Anterior Cingulate, BA 32 | 7, 9, 13, 33, 37, 43, 45 |
| 0.014786 | −2 | 44 | −6 | Anterior Cingulate, BA 32 | |||
| 4 | 1752 | 0.019154 | 28 | 0 | −26 | R. Amygdala | 5b, 8, 12, 18, 24, 29, 35, 37 |
| 0.016753 | 30 | −8 | −26 | R. Amygdala | |||
| 0.01353 | 34 | 2 | −16 | R. Putamen | |||
| 5 | 1208 | 0.019103 | 8 | −10 | 2 | R. Thalamus | 12, 15, 20, 37, 46 |
| 6 | 1192 | 0.019494 | 8 | 10 | −8 | R. Caudate Head | 5b, 12, 23, 37, 39, 43 |
| 0.016312 | 10 | 6 | −14 | R. Sub-lobar | |||
| 7 | 1080 | 0.017705 | −10 | 0 | −4 | L. Medial Globus Pallidus | 6, 12, 37, 39 |
| 0.016955 | −12 | 4 | −6 | L. Medial Globus Pallidus | |||
| 8 | 872 | 0.017468 | 44 | 22 | −12 | R. Inferior Frontal Gyrus, BA 47 | 1, 13, 18, 23, 43 |
| 9 | 800 | 0.016139 | −48 | −8 | −26 | L. Middle Temporal Gyrus, BA 21 | 5b, 6, 12 |
| 0.015733 | −48 | −16 | −14 | L. Sub-Gyral, BA 21 | |||
| 10 | 760 | 0.016609 | 34 | −46 | −24 | R. Cerebellum, Anterior Lobe | 29, 34, 38, 40 |
| 0.012444 | 38 | −50 | −14 | R. Fusiform Gyrus, BA 37 | |||
| 11 | 712 | 0.017816 | −38 | −60 | −14 | L. Fusiform Gyrus, BA 37 | 16, 24, 29, 37 |
| 12 | 664 | 0.015137 | 36 | −34 | −14 | R. Parahippocampal Gyrus, BA 36 | 12, 21, 37 |
| 0.014701 | 32 | −38 | −4 | R. Hippocampus | |||
| 13 | 664 | 0.017654 | −36 | 50 | 14 | L. Middle Frontal Gyrus, BA 10 | 47, 48, 49 |
| 0.016305 | −34 | 52 | 8 | L. Middle Frontal Gyrus, BA 10 | |||
| 0.011056 | −28 | 52 | 18 | L. Superior Frontal Gyrus, BA 10 | |||
The clusters were approached at uncorrected p < 0.01 and a cluster-level p < 0.05. x, y, and z coordinates are given in MNI space L left, R right, BA Brodmann’s area
Separate ALE analyses of PPI studies using the left and right amygdala as seeds showed seemingly different patterns of task modulated amygdala connectivity (Supplementary Figure S1 and Tables S7 and S8). However, direct subtraction analysis did not show significant clusters between the two sets of studies under the preselected threshold of uncorrected p < 0.01 and cluster-level p < 0.05.
Task specific modulations of amygdala connectivity
The first analysis identified brain regions that showed increased and decreased connectivity with the amygdala in fear processing (Fig. 2a; Table 3). Six clusters were identified to show increased connectivity with the amygdala, including the bilateral insula/inferior frontal gyrus (BA 13/47/10) and anterior cingulate/medial frontal gyrus (BA 32/24/10). Nine clusters were identified to show decreased connectivity with the amygdala, while only four of them were contributed by two or more studies. They were located in the thalamus, left middle/superior temporal gyrus (BA 21/22), posterior cingulate (BA 23), and right inferior frontal gyrus (BA 47). The studies that contributed to each of these clusters are listed in Table 3.
Fig. 2.
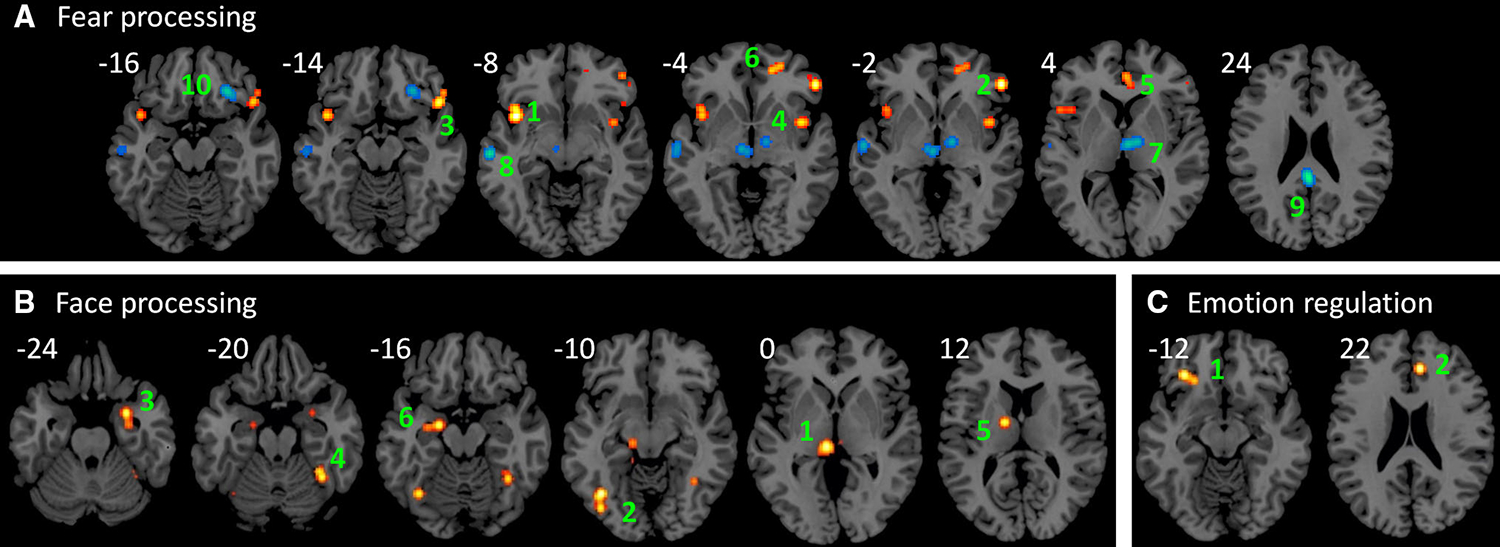
Regions that showed consistent increased (hot color) and decreased (winter color) connectivity with the amygdala in fear processing (a), face processing (b), and emotion regulation (c). The activation likelihood maps were aaproached at uncorrected p < 0.01 and a cluster-level p < 0.05. Numbers near each slice represent MNI (Montreal Neurological Institute) z coordinates
Table 3.
List of clusters that showed increased and decreased functional connectivity with the amygdala during fear processing
| Cluster# | Volume (mm3) | Extrema | x | y | z | Label | Contributed studies |
|---|---|---|---|---|---|---|---|
|
| |||||||
| Increased connectivity | |||||||
| 1 | 2520 | 0.013302 | −40 | 10 | −8 | L. Insula, BA 13 | 13, 23, 44 |
| 0.01063 | −40 | 16 | −8 | L. Insula, BA 13 | |||
| 0.008269 | −52 | 16 | 6 | L. Precentral Gyrus, BA 44 | |||
| 0.007905 | −42 | 16 | 0 | L. Insula, BA 13 | |||
| 2 | 1104 | 0.012847 | 52 | 36 | −2 | R. Inferior Frontal Gyrus | 13, 23 |
| 0.008165 | 46 | 44 | −8 | R. Sub-Gyral, BA 10 | |||
| 3 | 944 | 0.011367 | 48 | 22 | −14 | R. Inferior Frontal Gyrus, BA 47 | 13, 23 |
| 0.00815 | 52 | 30 | −14 | R. Inferior Frontal Gyrus, BA 47 | |||
| 4 | 736 | 0.00991 | 40 | 6 | −4 | R. Claustrum | 3, 44 |
| 5 | 648 | 0.008639 | 2 | 44 | 4 | L. Anterior Cingulate, BA 32 | 13, 23 |
| 0.007927 | 6 | 36 | 2 | R. Anterior Cingulate, BA 24 | |||
| 6 | 632 | 0.009047 | 16 | 48 | −4 | R. Anterior Cingulate, BA 32 | 13, 42 |
| 0.008658 | 24 | 52 | −4 | R. Medial Frontal Gyrus, BA 10 | |||
| Decreased connectivity | −60 | −12 | 0 | L. Superior Temporal Gyrus, BA 22 | |||
| 7 | 2952 | 0.008864 | 4 | −12 | 4 | R. Thalamus | 20, 46 |
| 0.008337 | 10 | −10 | 4 | R. Thalamus | |||
| 0.007903 | 4 | −14 | −2 | L. Thalamus | |||
| 0.007745 | −8 | −16 | −4 | L. Thalamus | |||
| 8 | 1784 | 0.010431 | −62 | −20 | −8 | L. Middle Temporal Gyrus, BA 21 | 3, 28, 38 |
| 0.007852 | −60 | −12 | 0 | L. Superior Temporal Gyrus, BA 22 | |||
| 9 | 1376 | 0.011565 | −2 | −38 | 24 | L. Posterior Cingulate, BA 23 | 38, 45 |
| 10 | 1320 | 0.009312 | 26 | 32 | −16 | R. Inferior Frontal Gyrus, BA 47 | 26, 32 |
The clusters were approached at uncorrected p < 0.01 and a cluster-level p < 0.05. x, y, and z coordinates are given in MNI space L left, R right, BA Brodmann’s area
The second analysis revealed brain regions that showed increased connectivity with the amygdala in face processing (Fig. 2b; Table 4). Six clusters in the thalamus/mid-brain, left fusiform gyrus (BA 37/19), parahippocampal gyrus (BA 34), right cerebellum extending to the fusiform gyrus, and left amygdala showed increased connectivity with the amygdala in face processing. All the clusters were contributed by two or more studies (Table 4).
Table 4.
List of clusters that showed increased functional connectivity with the amygdala during face processing
| Cluster# | Volume (mm3) | Extrema | x | y | z | Label | Contributed studies |
|---|---|---|---|---|---|---|---|
|
| |||||||
| Increased connectivity | |||||||
| 1 | 2608 | 0.011684 | −8 | −26 | 0 | L. Thalamus | 31, 37, 44 |
| 0.009499 | −14 | −36 | −6 | L. Parahippocampal Gyrus, BA 30 | |||
| 0.008935 | −14 | −22 | −8 | L. Substania Nigra | |||
| 0.008413 | 4 | −22 | −4 | R. Red Nucleus | |||
| 2 | 1616 | 0.01243 | −40 | −64 | −10 | L. Fusiform Gyrus, BA 37 | 24, 37, 38 |
| 0.009832 | −40 | −76 | −10 | L. Fusiform Gyrus, BA 19 | |||
| 3 | 1136 | 0.01109 | 28 | 2 | −24 | R. Parahippocampal Gyrus, BA 34 | 24, 37 |
| 0.008339 | 30 | −8 | −26 | R. Amygdala | |||
| 0.008276 | 30 | −2 | −34 | R. Uncus, BA 28 | |||
| 4 | 1048 | 0.01028 | 36 | −50 | −20 | R. Cerebellum, Anterior Lobe | 37, 38 |
| 0.009768 | 34 | −46 | −20 | R, Cerebellum, Anterior Lobe | |||
| 0.008725 | 36 | −54 | −12 | R. Cerebellum, Posterior Lobe | |||
| 5 | 832 | 0.011125 | −14 | −6 | 12 | L. Thalamus, Ventral Anterior Nucleus | 24, 37 |
| 0.008367 | −12 | 0 | 20 | L. Caudate Body | |||
| 6 | 816 | 0.01109 | −20 | −8 | −16 | L. Amygdala | 24, 37 |
| 0.008339 | −30 | −10 | −14 | L. Amygdala | |||
The clusters were approached at uncorrected p < 0.01 and a cluster-level p < 0.05. x, y, and z coordinates are given in MNI space L left, R right, BA Brodmann’s area
The third analysis examined brain regions that showed increased connectivity with the amygdala in emotion regulation. Five clusters were observed, but only two clusters were contributed by two studies (Fig. 2c; Table 5). The first was located in the left inferior frontal gyrus (BA 47), and the second was located in the cingulate gyrus (BA 32).
Table 5.
List of clusters that showed increased functional connectivity with the amygdala during emotion regulation
| Cluster# | Volume (mm3) | Extrema | x | y | z | Label | Contributed studies |
|---|---|---|---|---|---|---|---|
|
| |||||||
| Increased connectivity | |||||||
| 1 | 960 | 0.00916 | −32 | 32 | −12 | L. Inferior Frontal Gyrus, BA 47 | 4, 18 |
| 0.007762 | −24 | 28 | −14 | L. Inferior Frontal Gyrus, BA 47 | |||
| 2 | 920 | 0.009991 | 8 | 38 | 22 | Cingulate Gyrus, BA 32 | 4, 39 |
| 0.007628 | 8 | 44 | 32 | Medial Frontal Gyrus, BA 6 | |||
The clusters were approached at uncorrected p < 0.01 and a cluster-level p < 0.05. x, y, and z coordinates are given in MNI space L left, R right, BA Brodmann’s area
Coactivations of the amygdala
Lastly, we examined coactivations of the amygdala by performing an ALE analysis on studies that reported activations in the amygdala. The analysis identified 6 clusters (Fig. 3). The first mega cluster covered widespread regions including the bilateral fusiform gyrus, bilateral inferior occipital gyrus, bilateral middle/superior temporal gyrus, bilateral inferior/middle frontal gyrus, anterior cingulate, bilateral insula, basal ganglia and thalamus. The remaining clusters were located in the anterior cingulate, medial frontal gyrus, and posterior cingulate. A list of all the clusters is reported in supplementary Table S9.
Fig. 3.
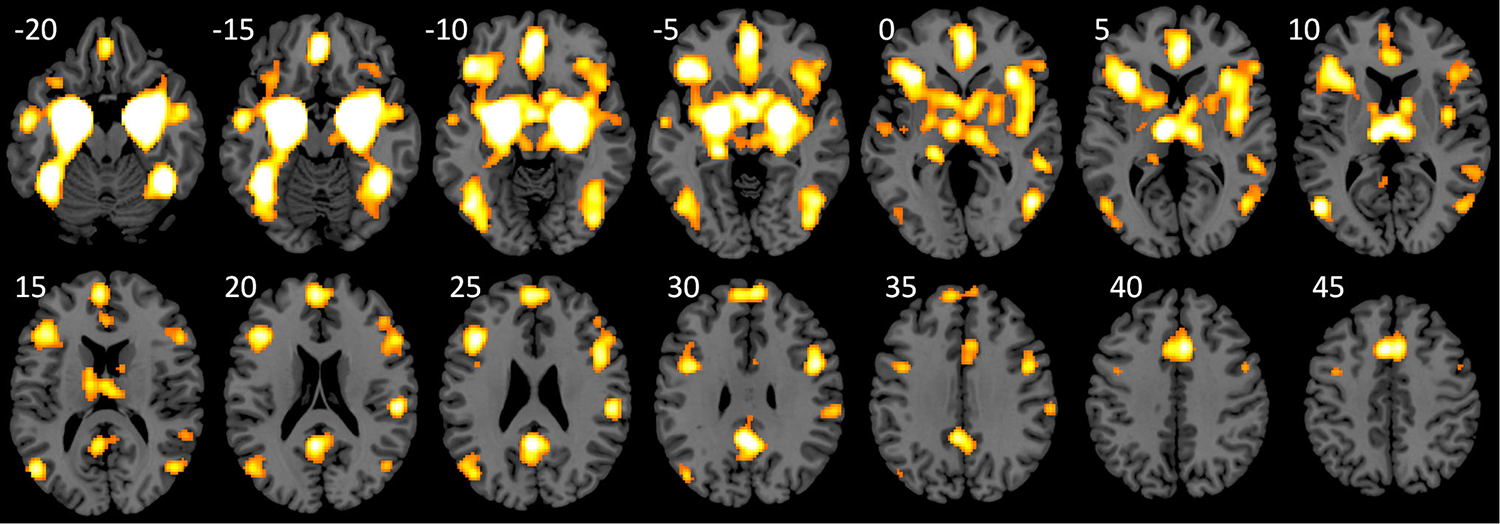
Regions that showed coactivations with the amygdala. The activation likelihood map was approached at uncorrected p < 0.01 and a cluster-level p < 0.05. Numbers near each slice represent MNI (Montreal Neurological Institute) z coordinates
Discussion
By performing coordinate-based meta-analysis on PPI studies using the amygdala as seeds, the current study first identified regions whose connectivity with the amygdala were consistently modulated by different tasks. Secondly, we showed that different cognitive and affective processes, i.e., fear processing and face processing, modulated amygdala connectivity with different brain regions.
The main meta-analysis of all 49 studies identified widespread brain regions that showed task modulated connectivity with the amygdala. These regions can be grouped into four sets based on their anatomical locations. The first is the dorsal medial frontal region (BA 6/32). Animal studies suggested anatomical projection from the region BA 6 to the amygdala (Robinson et al. 2010). And this region also shows significant resting-state connectivity (Hahn et al. 2011) and coactivation with the amygdala. The second set consists of ventral frontal regions, such as the bilateral inferior frontal gyrus, insula, and medial pre-frontal cortex/anterior cingulate cortex, which are anatomically connected to the amygdala via the uncinate fasciculus (Catani and Thiebaut de Schotten 2008). This set of regions also shows baseline functional connectivity in resting-state (Hahn et al. 2011; Roy et al. 2009) and coactivations with the amygdala. The third set includes subcortical regions, such as the thalamus, basal ganglia, and adjacent parahippocampal gyrus. The anatomical connections between these regions to the amygdala are difficult to be investigated using human DTI, but are evident in animal studies (Aggleton et al. 1980; Packard and Teather 1998). There regions also show baseline functional connectivity in resting-state (Hahn et al. 2011; Roy et al. 2009) and extensive coactivations with the amygdala. And the last set consists of posterior visual related regions, including the bilateral fusiform gyrus, which are anatomically connected to the amygdala via the inferior longitudinal fasciculus. These regions also show coactivations with the amygdala, however, they did not show reliable baseline functional connectivity with the amygdala in resting-state (Hahn et al. 2011; Roy et al. 2009). We note that several clusters in the current results were labeled as the cerebellum, but the major parts of the clusters are actually located in the adjacent fusiform gyrus. This spatial displacement may result from smoothness of fMRI data. But based on the current knowledge of amygdala connectivity, we regard these clusters as located in the fusiform gyrus but not cerebellum. It seems that the general patterns of anatomical connectivity, resting-state connectivity, coactivations, and task modulated connectivity of the amygdala agree with each other. But the spatial extend of resting-state functional connectivity of the amygdala is limited (Hahn et al. 2011; Ma et al. 2010; Sripada et al. 2012). It is consistent with the economic theory of brain connectivity stating that maintaining of long range connectivity in resting-state is costly (Bullmore and Sporns 2012). Therefore, certain long range functional communications, e.g., between the amygdala and fusiform gyrus, may only take place upon task demands.
In addition to the overall pattern, the current analysis also identified specific amygdala connectivity that is modulated by fear processing, face processing, and emotion regulation. Specifically, fear processing increases amygdala connectivity to the bilateral insula/inferior frontal gyrus (BA 13/47) and orbitofrontal cortex (anterior cingulate/medial frontal gyrus, BA 10/24/32), and reduces amygdala connectivity to the thalamus, middle temporal gyrus (BA 21/22), posterior cingulate (BA 23), and inferior frontal gyrus (BA 47). The orbitofrontal cortex is a key brain structure that is responsible for decision-making and emotion representations (Bechara 2000). Many studies have reported activations in the orbitofrontal cortex during fear processing (Fusar-Poli et al. 2009; Sladky et al. 2015). The current results suggest increased functional interactions between the orbital frontal regions and the amygdala during fear processing. Although the directionality of the increased connectivity is difficult to infer from PPI analysis, a study using dynamic causal modeling (DCM) (Friston et al. 2003) implied a downregulation of the orbitofrontal cortex to the amygdala (Sladky et al. 2015). The insula was originally thought to be specifically involved in the processing of disgust (Phillips et al. 1997). However, later studies showed that the insula was also involved in fear processing (Schienle et al. 2002; Vytal and Hamann 2010). The bilateral insula/inferior frontal gyrus together with dorsal anterior cingulate was defined as the salience network, which is responsible for processing saliency information (Seeley et al. 2007). The results highlight the interactions between the general purpose insula and the amygdala in fear processing. The current results also conveyed decreased connectivity between the thalamus and amygdala. The thalamic cluster was located in the anterior portion of the thalamus, and may correspond to the medial dorsal nucleus. The decreased connectivity may indicate an inhibitory modulation between these two nuclei during fear processing. We note that both of the contributing studies (Koelsch et al. 2013; Williams et al. 2006) involved conscious processing of fear. In the same study, it has been shown that unconscious processing of fear showed increased connectivity between the amygdala and posterior portion of the thalamus (possibly the pulvinar and lateral geniculate nucleus) (Williams et al. 2006). This may suggest a functional dissociation between different thalamic nuclei and the amygdala in fear processing.
The amygdala contains neurons specifically for face processing, even for a face that does not produce emotional response (Leonard et al. 1985). The current analysis demonstrated that the amygdala showed increased functional interactions with other face processing regions, such as the parahippocampal gyrus and fusiform gyrus (Mende-Siedlecki et al. 2013). An electrophysiological study on monkey showed that the response latencies of the amygdala face neurons are longer than face selective neurons in the superior temporal sulcus (Leonard et al. 1985). However, a DCM study on human subjects suggested bidirectional interactions between the amygdala and face regions during face processing (Herrington et al. 2011). We note that three out of five studies included in the current face analysis compared fearful face with other non-face objects. So the increased connectivity might result from either face or fear processing. However, one study (Mende-Siedlecki et al. 2013) indeed compared neutral face with other stimuli, and contributed to the clusters in the fusiform gyrus, parahippocampal gyrus, and thalamus. Given the strong involvement of these regions in face processing, it is highly likely that the connectivity between these regions to the amygdala was modulated by face processing, but not fear processing.
Emotion regulation involves cognitive regulations of emotional experiences. In addition to the amygdala, fMRI studies of emotion regulation usually report increased activation in cognitive control regions, such as lateral frontal cortex and anterior cingulate (Frank et al. 2014; Kohn et al. 2014). Along this line, the current study demonstrated increased connectivity between the inferior frontal gyrus and amygdala and between the anterior cingulate/medial frontal gyrus and amygdala, which may reflect a downregulation of these cognitive control regions to the amygdala. However, the causal influence needs further confirmation using methods such as DCM.
The current results demonstrated to some extent that different task domains modulated connectivity between the amygdala with different brain regions. However, the specificity of the task modulated connectivity seems quite limited, i.e., multiple tasks might modulate the same connectivity. For instance, thirteen studies contributed to the medial frontal cortex cluster (Table 2), which is the largest cluster in the main meta-analysis. The tasks used in these studies range from emotion processing, to cognitive processes such as face processing and working memory, and to higher level social processes such as empathy and perspective taking. Therefore, it is not a one-to-one mapping between function and connectivity (Pessoa 2014). Actually, the dorsal medial frontal cortex is one of the regions that are highly likely to be reported in neuroimaging studies (Yarkoni et al. 2011). It is possible that brain regions such as the medial frontal cortex could support multiple functions, so that the communications between the medial frontal cortex and amygdala could also support multiple functions. Alternatively, the interaction between two brain regions may support a specific cognitive or affective process, which may be part of many tasks. If we could decompose different tasks into underlying cognitive and affective processes, it might be possible to map a certain process to a connection. Fine coding of behavioral domains are needed [e.g. (Fox et al. 2005; Turner and Laird, 2012)] in future works to provide better mappings between connections and functions.
Several lines of studies have suggested functional lateralization of the amygdala. More brain imaging studies on emotions reported activations in the left amygdala than in the right amygdala (Baas et al. 2004), Direct simulation of the left amygdala could induce either pleasant or unpleasant emotions, but simulating the right amygdala could only induce negative emotions (Lanteaume et al. 2007). However, resting-state connectivity of the left and right amygdala shows similar spatial distributions (Roy et al. 2009). The current analysis demonstrated that task modulated amygdala turned out to be quite different for the left and right amygdala (supplementary Figure S1, Table S7 and S8). This might suggest different functional roles of the left and right amygdala in different tasks. We noted, however, that direct comparisons between the PPI effects of the left and right amygdala did not show significant results. The lateralization of task modulated amygdala connectivity is interesting, but further studies are certainly needed to confirm the observation.
When performing literature search for the meta-analysis, we identified several papers that examined individual differences of task modulated amygdala connectivity. These papers were not included in the current analysis, but they suggest several factors that may moderate task modulated amygdala connectivity. First, several studies have demonstrated that even though mean effects of task modulated connectivity with the amygdala were small, task modulated connectivity showed substantial individual differences (Cremers et al. 2010; Lee et al. 2012; Passamonti et al. 2008, 2009). These individual differences were shown to be correlated with many factors such as age, personality traits, and motivations (Cremers et al. 2010; Passamonti et al. 2008, 2009; Wu et al. 2016; Yoder et al. 2015). Second, there are a bunch of studies that showed genetic modulations of task modulated amygdala connectivity, including a serotonin transporter gene (5-HTTLPR) (Friedel et al. 2009; Lemogne et al. 2011; Madsen et al. 2016; Roiser et al. 2009), a dopamine D2 receptor gene (DRD2) (Blasi et al. 2009), a Catechol-O-methyltransferase (COMT) gene (Val158Met) (Rasch et al. 2010), a oxytocin receptor gene (Puglia et al. 2015), a noradrenergic system gene (ADRA2B) (Rasch et al. 2009), and a microRNA gene (MIR137) (Mothersill et al. 2014). For example, the 5-HTTLPR genotypes modulated task related connectivity between the amygdala and ACC in emotion regulation (Lemogne et al. 2011) and decision-making (Roiser et al. 2009). And last, higher level (three-way) interaction models have been proposed as a way to further study whether task modulation of connectivity between two regions was modulated by a third region, i.e., psychophysio-physiological interaction (PPPI) (Stamatakis et al. 2005). A recent study demonstrated that the task modulated connectivity between the amygdala and widespread networks were further modulated by the basal forebrain (Gorka et al. 2015). In short, task modulated connectivity of the amygdala show substantial individual differences, which may be modulated by many genetic factors and other subcortical nuclei.
The ALE method was originally developed to examine spatial consistency of brain activations studies (Turkeltaub et al. 2002). In principle, it can be applied to study spatial consistencies of any kinds of reported coordinates. For example, it has also been used to study consistent results of brain anatomical differences derived from voxel-based morphometry (VBM) studies (Chan et al. 2011; Di et al. 2009, 2014; Ellison-Wright et al. 2008) and diffusion tensor imaging (DTI) studies (Ellison-Wright and Bullmore 2009). A similar coordinate-based meta-analytic approach, i.e., kernel density analysis, has also been used to study consistent resting-state functional connectivity in major depressive disorder (Kaiser et al. 2015). To our knowledge, the current study is the first one to apply ALE method to PPI studies. The meta-analytic approach used in the current analysis could be applied to other brain structures, such as the thalamus, and could also be used to map a task to the whole brain connectome. The “task connectome” provides a systematic framework towards understanding of dynamic connectivity in different task contexts.
Conclusion
We demonstrated brain regions that consistently show task modulated connectivity with the amygdala. Fear processing, face processing, and emotion regulation selectively modulate amygdala connectivity to the regions that are responsible for these processes. However, the functional specificity of task modulated connectivity is quite limited. The meta-analytic approach on PPI studies may provide a systematic approach to map task modulated connectivity onto the functional brain connectome.
Supplementary Material
Acknowledgments
We thank Dr. Suril Gohel for his insightful comments on an earlier version of this manuscript. This research was supported by Grants from NIH R01AG032088, R01DA038895, and NSFC (National Natural Science Foundation of China) 31100747.
References
- Adolphs R, Tranel D, Damasio H, Damasio A (1994) Impaired recognition of emotion in facial expressions following bilateral damage to the human amygdala. Nature 372:669–672. doi: 10.1038/372669a0 [DOI] [PubMed] [Google Scholar]
- Adolphs R, Tranel D, Damasio AR (1998) The human amygdala in social judgment. Nature 393:470–474. doi: 10.1038/30982 [DOI] [PubMed] [Google Scholar]
- Adolphs R, Tranel D, Buchanan TW (2005) Amygdala damage impairs emotional memory for gist but not details of complex stimuli. Nat Neurosci 8:512–518. doi: 10.1038/nn1413 [DOI] [PubMed] [Google Scholar]
- Aggleton JP, Burton MJ, Passingham RE (1980) Cortical and subcortical afferents to the amygdala of the rhesus monkey (Macaca mulatta). Brain Res 190:347–368. doi: 10.1016/0006-8993(80)90279-6 [DOI] [PubMed] [Google Scholar]
- Anderson AK, Phelps EA (2001) Lesions of the human amygdala impair enhanced perception of emotionally salient events. Nature 411:305–309. doi: 10.1038/35077083 [DOI] [PubMed] [Google Scholar]
- Baas D, Aleman A, Kahn RS (2004) Lateralization of amygdala activation: a systematic review of functional neuroimaging studies. Brain Res Rev 45:96–103. doi: 10.1016/j.brainresrev.2004.02.004 [DOI] [PubMed] [Google Scholar]
- Bechara A (2000) Emotion, decision making and the orbitofrontal cortex. Cereb Cortex 10:295–307. doi: 10.1093/cercor/10.3.295 [DOI] [PubMed] [Google Scholar]
- Bechara A, Damasio H, Damasio AR (2003) Role of the amygdala in decision-making. Ann N Y Acad Sci 985:356–369. doi: 10.1111/j.1749-6632.2003.tb07094.x [DOI] [PubMed] [Google Scholar]
- Blasi G, Lo Bianco L, Taurisano P, Gelao B, Romano R, Fazio L, Papazacharias A, Di Giorgio A, Caforio G, Rampino A, Masellis R, Papp A, Ursini G, Sinibaldi L, Popolizio T, Sadee W, Bertolino A (2009) Functional variation of the dopamine D2 receptor gene is associated with emotional control as well as brain activity and connectivity during emotion processing in humans. J Neurosci 29:14812–14819. doi: 10.1523/JNEUROSCI.3609-09.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bruneau EG, Jacoby N, Saxe R (2015) Empathic control through coordinated interaction of amygdala, theory of mind and extended pain matrix brain regions. Neuroimage 114:105–119. doi: 10.1016/j.neuroimage.2015.04.034 [DOI] [PubMed] [Google Scholar]
- Bullmore E, Sporns O (2012) The economy of brain network organization. Nat Rev Neurosci 13:336–349. doi: 10.1038/nrn3214 [DOI] [PubMed] [Google Scholar]
- Catani M, Thiebaut de Schotten M (2008) A diffusion tensor imaging tractography atlas for virtual in vivo dissections. Cortex 44:1105–1132. doi: 10.1016/j.cortex.2008.05.004 [DOI] [PubMed] [Google Scholar]
- Catani M, Howard RJ, Pajevic S, Jones DK (2002) Virtual in vivo interactive dissection of white matter fasciculi in the human brain. Neuroimage 17:77–94. doi: 10.1006/nimg.2002.1136 [DOI] [PubMed] [Google Scholar]
- Chan RCK, Di X, McAlonan GM, Gong Q (2011) Brain anatomical abnormalities in high-risk individuals, first-episode, and chronic schizophrenia: an activation likelihood estimation meta-analysis of illness progression. Schizophr Bull 37:177–188. doi: 10.1093/schbul/sbp073 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cisler JM, Bush K, Steele JS (2013) A comparison of statistical methods for detecting context-modulated functional connectivity in fMRI. Neuroimage [DOI] [PMC free article] [PubMed] [Google Scholar]
- Costafreda SG, Brammer MJ, David AS, Fu CHY (2008) Predictors of amygdala activation during the processing of emotional stimuli: a meta-analysis of 385 PET and fMRI studies. Brain Res Rev 58:57–70. doi: 10.1016/j.brainresrev.2007.10.012 [DOI] [PubMed] [Google Scholar]
- Cremers HR, Demenescu LR, Aleman A, Renken R, van Tol M-J, van der Wee NJA, Veltman DJ, Roelofs K (2010) Neuroticism modulates amygdala-prefrontal connectivity in response to negative emotional facial expressions. Neuroimage 49:963–970. doi: 10.1016/j.neuroimage.2009.08.023 [DOI] [PubMed] [Google Scholar]
- Di X, Chan RCK, Gong Q (2009) White matter reduction in patients with schizophrenia as revealed by voxel-based morphometry: an activation likelihood estimation meta-analysis. Prog Neuropsychopharmacol Biol Psychiatry 33:1390–1394. doi: 10.1016/j.pnpbp.2009.08.020 [DOI] [PubMed] [Google Scholar]
- Di X, Rypma B, Biswal BB (2014) Correspondence of executive function related functional and anatomical alterations in aging brain. Prog Neuropsychopharmacol Biol Psychiatry 48:41–50. doi: 10.1016/j.pnpbp.2013.09.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eickhoff SB, Laird AR, Grefkes C, Wang LE, Zilles K, Fox PT (2009) Coordinate-based activation likelihood estimation meta-analysis of neuroimaging data: a random-effects approach based on empirical estimates of spatial uncertainty. Hum Brain Mapp 30:2907–2926. doi: 10.1002/hbm.20718 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eickhoff SB, Bzdok D, Laird AR, Kurth F, Fox PT (2012) Activation likelihood estimation meta-analysis revisited. Neuroimage 59:2349–2361. doi: 10.1016/j.neuroimage.2011.09.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellison-Wright I, Bullmore E (2009) Meta-analysis of diffusion tensor imaging studies in schizophrenia. Schizophr Res 108:3–10. doi: 10.1016/j.schres.2008.11.021 [DOI] [PubMed] [Google Scholar]
- Ellison-Wright I, Glahn DC, Laird AR, Thelen SM, Bullmore E (2008) The anatomy of first-episode and chronic schizophrenia: an anatomical likelihood estimation meta-analysis. Am J Psychiatry 165:1015–1023. doi: 10.1176/appi.ajp.2008.07101562 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox PT, Laird AR, Fox SP, Fox PM, Uecker AM, Crank M, Koenig SF, Lancaster JL (2005) BrainMap taxonomy of experimental design: description and evaluation. Hum Brain Mapp 25:185–198. doi: 10.1002/hbm.20141 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frank DW, Dewitt M, Hudgens-Haney M, Schaeffer DJ, Ball BH, Schwartz N, Hussein AA, Smart LM, Sabatinelli D (2014) Emotion regulation: quantitative meta-analysis of functional activation and deactivation. Neurosci Biobehav Rev 45:202–211. doi: 10.1016/j.neubiorev.2014.06.010 [DOI] [PubMed] [Google Scholar]
- Friedel E, Schlagenhauf F, Sterzer P, Park SQ, Bermpohl F, Ströhle A, Stoy M, Puls I, Hägele C, Wrase J, Büchel C, Heinz A (2009) 5-HTT genotype effect on prefrontal-amygdala coupling differs between major depression and controls. Psychopharmacology 205:261–271. doi: 10.1007/s00213-009-1536-1 [DOI] [PubMed] [Google Scholar]
- Friston KJ, Buechel C, Fink GR, Morris J, Rolls E, Dolan RJ (1997) Psychophysiological and modulatory interactions in neuroimaging. Neuroimage 6:218–229 [DOI] [PubMed] [Google Scholar]
- Friston KJ, Harrison L, Penny W (2003) Dynamic causal modelling. Neuroimage 19:1273–1302. doi: 10.1016/S1053-8119(03)00202-7 [DOI] [PubMed] [Google Scholar]
- Fusar-Poli P, Placentino A, Carletti F, Landi P, Allen P, Surguladze S, Benedetti F, Abbamonte M, Gasparotti R, Barale F, Perez J, McGuire P, Politi P (2009) Functional atlas of emotional faces processing: a voxel-based meta-analysis of 105 functional magnetic resonance imaging studies. J Psychiatry Neurosci 34:418–432 [PMC free article] [PubMed] [Google Scholar]
- Gitelman DR, Penny WD, Ashburner J, Friston KJ (2003) Modeling regional and psychophysiologic interactions in fMRI: the importance of hemodynamic deconvolution. Neuroimage 19:200–207 [DOI] [PubMed] [Google Scholar]
- Gorka AX, Knodt AR, Hariri AR (2015) Basal forebrain moderates the magnitude of task-dependent amygdala functional connectivity. Soc Cogn Affect Neurosci 10:501–507. doi: 10.1093/scan/nsu080 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hahn A, Stein P, Windischberger C, Weissenbacher A, Spindelegger C, Moser E, Kasper S, Lanzenberger R (2011) Reduced resting-state functional connectivity between amygdala and orbitofrontal cortex in social anxiety disorder. Neuroimage 56:881–889. doi: 10.1016/j.neuroimage.2011.02.064 [DOI] [PubMed] [Google Scholar]
- Herrington JD, Taylor JM, Grupe DW, Curby KM, Schultz RT (2011) Bidirectional communication between amygdala and fusiform gyrus during facial recognition. Neuroimage 56:2348–2355. doi: 10.1016/j.neuroimage.2011.03.072 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaiser RH, Andrews-Hanna JR, Wager TD, Pizzagalli DA (2015) Large-scale network dysfunction in major depressive disorder. JAMA Psychiatry. doi: 10.1001/jamapsychiatry.2015.0071 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim J, Horwitz B (2008) Investigating the neural basis for fMRI-based functional connectivity in a blocked design: application to interregional correlations and psycho-physiological interactions. Magn Reson Imaging 26:583–593. doi: 10.1016/j.mri.2007.10.011 [DOI] [PubMed] [Google Scholar]
- Koelsch S, Skouras S, Fritz T, Herrera P, Bonhage C, Küssner MB, Jacobs AM (2013) The roles of superficial amygdala and auditory cortex in music-evoked fear and joy. Neuroimage 81:49–60. doi: 10.1016/j.neuroimage.2013.05.008 [DOI] [PubMed] [Google Scholar]
- Kohn N, Eickhoff SB, Scheller M, Laird AR, Fox PT, Habel U (2014) Neural network of cognitive emotion regulation—an ALE meta-analysis and MACM analysis. Neuroimage 87:345–355. doi: 10.1016/j.neuroimage.2013.11.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laird AR, Fox PM, Price CJ, Glahn DC, Uecker AM, Lancaster JL, Turkeltaub PE, Kochunov P, Fox PT (2005a) ALE meta-analysis: controlling the false discovery rate and performing statistical contrasts. Hum Brain Mapp 25:155–164. doi: 10.1002/hbm.20136 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laird AR, Lancaster JL, Fox PT (2005b) BrainMap: the social evolution of a human brain mapping database. Neuroinformatics 3:65–78 [DOI] [PubMed] [Google Scholar]
- Laird AR, Robinson JL, McMillan KM, Tordesillas-Gutiérrez D, Moran ST, Gonzales SM, Ray KL, Franklin C, Glahn DC, Fox PT, Lancaster JL (2010) Comparison of the disparity between Talairach and MNI coordinates in functional neuroimaging data: validation of the Lancaster transform. Neuroimage 51:677–683. doi: 10.1016/j.neuroimage.2010.02.048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laird AR, Riedel MC, Sutherland MT, Eickhoff SB, Ray KL, Uecker AM, Fox PM, Turner JA, Fox PT (2015) Neural architecture underlying classification of face perception paradigms. Neuroimage 119:70–80. doi: 10.1016/j.neuroimage.2015.06.044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lancaster JL, Tordesillas-Gutiérrez D, Martinez M, Salinas F, Evans A, Zilles K, Mazziotta JC, Fox PT (2007) Bias between MNI and Talairach coordinates analyzed using the ICBM-152 brain template. Hum Brain Mapp 28:1194–1205. doi: 10.1002/hbm.20345 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lanteaume L, Khalfa S, Regis J, Marquis P, Chauvel P, Bartolomei F (2007) Emotion induction after direct intracerebral stimulations of human amygdala. Cereb Cortex 17:1307–1313. doi: 10.1093/cercor/bhl041 [DOI] [PubMed] [Google Scholar]
- Lee H, Heller AS, van Reekum CM, Nelson B, Davidson RJ (2012) Amygdala-prefrontal coupling underlies individual differences in emotion regulation. Neuroimage 62:1575–1581. doi: 10.1016/j.neuroimage.2012.05.044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lemogne C, Gorwood P, Boni C, Pessiglione M, Lehéricy S, Fossati P (2011) Cognitive appraisal and life stress moderate the effects of the 5-HTTLPR polymorphism on amygdala reactivity. Hum Brain Mapp 32:1856–1867. doi: 10.1002/hbm.21150 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leonard CM, Rolls ET, Wilson FAW, Baylis GC (1985) Neurons in the amygdala of the monkey with responses selective for faces. Behav Brain Res 15:159–176. doi: 10.1016/0166-4328(85)90062-2 [DOI] [PubMed] [Google Scholar]
- Lowe MJ, Mock BJ, Sorenson JA (1998) Functional connectivity in single and multislice echo planar imaging using resting-state fluctuations. Neuroimage 7:119–132. doi: 10.1006/nimg.1997.0315 [DOI] [PubMed] [Google Scholar]
- Ma N, Liu Y, Li N, Wang C-X, Zhang H, Jiang X-F, Xu H-S, Fu X-M, Hu X, Zhang D-R (2010) Addiction related alteration in resting-state brain connectivity. Neuroimage 49:738–744. doi: 10.1016/j.neuroimage.2009.08.037 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Madsen MK, Mc Mahon B, Andersen SB, Siebner HR, Knudsen GM, Fisher PM (2016) Threat-related amygdala functional connectivity is associated with 5-HTTLPR genotype and neuroticism. Soc Cogn Affect Neurosci 11:140–149. doi: 10.1093/scan/nsv098 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McLaren DG, Ries ML, Xu G, Johnson SC (2012) A generalized form of context-dependent psychophysiological interactions (gPPI): a comparison to standard approaches. Neuroimage 61:1277–1286. doi: 10.1016/j.neuroimage.2012.03.068 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mende-Siedlecki P, Verosky SC, Turk-Browne NB, Todorov A (2013) Robust selectivity for faces in the human amygdala in the absence of expressions. J Cogn Neurosci 25:2086–2106. doi: 10.1162/jocn_a_00469 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morris JS, Ohman A, Dolan RJ (1998) Conscious and unconscious emotional learning in the human amygdala. Nature 393:467–470. doi: 10.1038/30976 [DOI] [PubMed] [Google Scholar]
- Mothersill O, Morris DW, Kelly S, Rose EJ, Fahey C, O’Brien C, Lyne R, Reilly R, Gill M, Corvin AP, Donohoe G (2014) Effects of MIR137 on fronto-amygdala functional connectivity. Neuroimage 90:189–195. doi: 10.1016/j.neuroimage.2013.12.019 [DOI] [PubMed] [Google Scholar]
- Murphy K, Birn RM, Handwerker DA, Jones TB, Bandettini PA (2009) The impact of global signal regression on resting state correlations: are anti-correlated networks introduced? Neuroimage 44:893–905. doi: 10.1016/j.neuroimage.2008.09.036 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Packard MG, Teather LA (1998) Amygdala modulation of multiple memory systems: hippocampus and caudate-putamen. Neurobiol Learn Mem 69:163–203. doi: 10.1006/nlme.1997.3815 [DOI] [PubMed] [Google Scholar]
- Passamonti L, Rowe JB, Ewbank M, Hampshire A, Keane J, Calder AJ (2008) Connectivity from the ventral anterior cingulate to the amygdala is modulated by appetitive motivation in response to facial signals of aggression. Neuroimage 43:562–570. doi: 10.1016/j.neuroimage.2008.07.045 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Passamonti L, Rowe JB, Schwarzbauer C, Ewbank MP, von dem Hagen E, Calder AJ (2009) Personality predicts the brain’s response to viewing appetizing foods: the neural basis of a risk factor for overeating. J Neurosci 29:43–51. doi: 10.1523/JNEUROSCI.4966-08.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pessoa L (2014) Understanding brain networks and brain organization. Phys Life Rev 11:400–435. doi: 10.1016/j.plrev.2014.03.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phan KL, Wager T, Taylor SF, Liberzon I (2002) Functional neuroanatomy of emotion: a meta-analysis of emotion activation studies in PET and fMRI. Neuroimage 16:331–348. doi: 10.1006/nimg.2002.1087 [DOI] [PubMed] [Google Scholar]
- Phelps EA (2006) Emotion and cognition: insights from studies of the human amygdala. Annu Rev Psychol 57:27–53. doi: 10.1146/annurev.psych.56.091103.070234 [DOI] [PubMed] [Google Scholar]
- Phillips ML, Young AW, Senior C, Brammer M, Andrew C, Calder AJ, Bullmore ET, Perrett DI, Rowland D, Williams SC, Gray JA, David AS (1997) A specific neural substrate for perceiving facial expressions of disgust. Nature 389:495–498. doi: 10.1038/39051 [DOI] [PubMed] [Google Scholar]
- Puglia MH, Lillard TS, Morris JP, Connelly JJ (2015) Epigenetic modification of the oxytocin receptor gene influences the perception of anger and fear in the human brain. Proc Natl Acad Sci USA. 1422096112. doi: 10.1073/pnas.1422096112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rasch B, Spalek K, Buholzer S, Luechinger R, Boesiger P, Papassotiropoulos A, de Quervain DJ-F (2009) A genetic variation of the noradrenergic system is related to differential amygdala activation during encoding of emotional memories. Proc Natl Acad Sci USA 106:19191–19196. doi: 10.1073/pnas.0907425106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rasch B, Spalek K, Buholzer S, Luechinger R, Boesiger P, de Quervain DJ-F, Papassotiropoulos A (2010) Aversive stimuli lead to differential amygdala activation and connectivity patterns depending on catechol-O-methyltransferase Val158Met genotype. Neuroimage 52:1712–1719. doi: 10.1016/j.neuroimage.2010.05.054 [DOI] [PubMed] [Google Scholar]
- Robinson JL, Laird AR, Glahn DC, Lovallo WR, Fox PT (2010) Metaanalytic connectivity modeling: delineating the functional connectivity of the human amygdala. Hum Brain Mapp 31:173–184. doi: 10.1002/hbm.20854 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roiser JP, de Martino B, Tan GCY, Kumaran D, Seymour B, Wood NW, Dolan RJ (2009) A genetically mediated bias in decision making driven by failure of amygdala control. J Neurosci 29:5985–5991. doi: 10.1523/JNEUROSCI.0407-09.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roy AK, Shehzad Z, Margulies DS, Kelly AMC, Uddin LQ, Gotimer K, Biswal BB, Castellanos FX, Milham MP (2009) Functional connectivity of the human amygdala using resting state fMRI. Neuroimage 45:614–626. doi: 10.1016/j.neuroimage.2008.11.030 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saad ZS, Gotts SJ, Murphy K, Chen G, Jo HJ, Martin A, Cox RW (2012) Trouble at rest: how correlation patterns and group differences become distorted after global signal regression. Brain Connect 2:25–32. doi: 10.1089/brain.2012.0080 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sarter M, Markowitsch HJ (1985) Involvement of the amygdala in learning and memory: a critical review, with emphasis on anatomical relations. Behav Neurosci 99:342–380. doi: 10.1037/0735-7044.99.2.342 [DOI] [PubMed] [Google Scholar]
- Schienle A, Stark R, Walter B, Blecker C, Ott U, Kirsch P, Sammer G, Vaitl D (2002) The insula is not specifically involved in disgust processing: an fMRI study. NeuroReport 13:2023–2026. doi: 10.1097/00001756-200211150-00006 [DOI] [PubMed] [Google Scholar]
- Seeley WW, Menon V, Schatzberg AF, Keller J, Glover GH, Kenna H, Reiss AL, Greicius MD (2007) Dissociable intrinsic connectivity networks for salience processing and executive control. J Neurosci 27:2349–2356. doi: 10.1523/JNEUROSCI.5587-06.2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sladky R, Hoflich A, Kublbock M, Kraus C, Baldinger P, Moser E, Lanzenberger R, Windischberger C (2015) Disrupted effective connectivity between the amygdala and orbitofrontal cortex in social anxiety disorder during emotion discrimination revealed by dynamic causal modeling for fMRI. Cereb Cortex 25:895–903. doi: 10.1093/cercor/bht279 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sripada RK, King AP, Garfinkel SN, Wang X, Sripada CS, Welsh RC, Liberzon I (2012) Altered resting-state amygdala functional connectivity in men with posttraumatic stress disorder. J Psychiatry Neurosci 37:241–249. doi: 10.1503/jpn.110069 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stamatakis EA, Marslen-Wilson WD, Tyler LK, Fletcher PC (2005) Cingulate control of fronto-temporal integration reflects linguistic demands: a three-way interaction in functional connectivity. Neuroimage 28:115–121. doi: 10.1016/j.neuroimage.2005.06.012 [DOI] [PubMed] [Google Scholar]
- Talairach J, Tournoux P (1988) Co-planar stereotaxic atlas of the human brain. Thieme, New York [Google Scholar]
- Turkeltaub PE, Eden GF, Jones KM, Zeffiro TA (2002) Meta-analysis of the functional neuroanatomy of single-word reading: method and validation. Neuroimage 16:765–780. doi: 10.1006/nimg.2002.1131 [DOI] [PubMed] [Google Scholar]
- Turkeltaub PE, Eickhoff SB, Laird AR, Fox M, Wiener M, Fox P (2012) Minimizing within-experiment and within-group effects in activation likelihood estimation meta-analyses. Hum Brain Mapp 33:1–13. doi: 10.1002/hbm.21186 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turner JA, Laird AR (2012) The cognitive paradigm ontology: design and application. Neuroinformatics 10:57–66. doi: 10.1007/s12021-011-9126-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vytal K, Hamann S (2010) Neuroimaging support for discrete neural correlates of basic emotions: a voxel-based meta-analysis. J Cogn Neurosci 22:2864–2885. doi: 10.1162/jocn.2009.21366 [DOI] [PubMed] [Google Scholar]
- Whalen PJ, Rauch SL, Etcoff NL, McInerney SC, Lee MB, Jenike MA (1998) Masked presentations of emotional facial expressions modulate amygdala activity without explicit knowledge. J Neurosci 18:411–418. 9412517 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams LM, Das P, Liddell BJ, Kemp AH, Rennie CJ, Gordon E (2006) Mode of functional connectivity in amygdala pathways dissociates level of awareness for signals of fear. J Neurosci 26:9264–9271. doi: 10.1523/JNEUROSCI.1016-06.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu M, Kujawa A, Lu LH, Fitzgerald DA, Klumpp H, Fitzgerald KD, Monk CS, Phan KL (2016) Age-related changes in amygdala-frontal connectivity during emotional face processing from childhood into young adulthood. Hum Brain Mapp. doi: 10.1002/hbm.23129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yarkoni T, Poldrack RA, Nichols TE, Van Essen DC, Wager TD (2011) Large-scale automated synthesis of human functional neuroimaging data. Nat Methods 8:665–670. doi: 10.1038/nmeth.1635 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoder KJ, Porges EC, Decety J (2015) Amygdala subnuclei connectivity in response to violence reveals unique influences of individual differences in psychopathic traits in a nonforensic sample. Hum Brain Mapp 36:1417–1428. doi: 10.1002/hbm.22712 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Young MP, Scannell JW, Burns GAPC, Blakemore C (1994) Analysis of connectivity: neural systems in the cerebral cortex. Rev Neurosci 5:227–250. doi: 10.1515/REVNEURO.1994.5.3.227 [DOI] [PubMed] [Google Scholar]
Appendix: Studies that were included in the current meta-analysis
- 1.Akitsuki Y, Decety J (2009) Social context and perceived agency affects empathy for pain: an event-related fMRI investigation. Neuroimage 47:722–34. [DOI] [PubMed] [Google Scholar]
- 2.Amting JM, Miller JE, Chow M, Mitchell DG V (2009) Getting mixed messages: the impact of conflicting social signals on the brain’s target emotional response. Neuroimage 47:1950–9. [DOI] [PubMed] [Google Scholar]
- 3.Amting JM, Greening SG, Mitchell DG V (2010) Multiple mechanisms of consciousness: the neural correlates of emotional awareness. J Neurosci 30:10039–47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Banks SJ, Eddy KT, Angstadt M, et al. (2007) Amygdala-frontal connectivity during emotion regulation. Soc Cogn Affect Neurosci 2:303–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bruneau EG, Jacoby N, Saxe R (2015) Empathic control through coordinated interaction of amygdala, theory of mind and extended pain matrix brain regions. Neuroimage 114:105–119. [DOI] [PubMed] [Google Scholar]
- 6.Cohen MX, Elger CE, Weber B (2008) Amygdala tractography predicts functional connectivity and learning during feedback-guided decision-making. Neuroimage 39:1396–407. [DOI] [PubMed] [Google Scholar]
- 7.Comte M, Schön D, Coull JT, et al. (2016) Dissociating bottom-up and top-down mechanisms in the cortico-limbic system during emotion processing. Cereb Cortex 26:144–155. [DOI] [PubMed] [Google Scholar]
- 8.Decety J, Porges EC (2011) Imagining being the agent of actions that carry different moral consequences: an fMRI study. Neuropsychologia 49:2994–3001. [DOI] [PubMed] [Google Scholar]
- 9.Erk S, Mikschl A, Stier S, et al. (2010) Acute and sustained effects of cognitive emotion regulation in major depression. J Neurosci 30:15726–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fakra E, Salgado-Pineda P, Delaveau P, et al. (2008) Neural bases of different cognitive strategies for facial affect processing in schizophrenia. Schizophr Res 100:191–205. [DOI] [PubMed] [Google Scholar]
- 11.Foland LC, Altshuler LL, Bookheimer SY, et al. (2008) Evidence for deficient modulation of amygdala response by prefrontal cortex in bipolar mania. Psychiatry Res 162:27–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Gianaros PJ, Onyewuenyi IC, Sheu LK, et al. (2012) Brain systems for baroreflex suppression during stress in humans. Hum Brain Mapp 33:1700–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gold AL, Morey RA, McCarthy G (2015) Amygdala-prefrontal cortex functional connectivity during threat-induced anxiety and goal distraction. Biol Psychiatry 77:394–403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Grabenhorst F, Schulte FP, Maderwald S, Brand M (2013) Food labels promote healthy choices by a decision bias in the amygdala. Neuroimage 74:152–63. [DOI] [PubMed] [Google Scholar]
- 15.Hermans EJ, Henckens MJAG, Roelofs K, Fernández s (2012) Fear bradycardia and activation of the human periaqueductal grey. Neuroimage 66C:278–287. [DOI] [PubMed] [Google Scholar]
- 16.Herrmann MJ, Boehme S, Becker MPI, Tupak SV., Guhn A, Schmidt B, et al. (2016): Phasic and sustained brain responses in the amygdala and the bed nucleus of the stria terminalis during threat anticipation. Hum Brain Mapp 37:1091–1102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Iidaka T, Omori M, Murata T, et al. (2001) Neural interaction of the amygdala with the prefrontal and temporal cortices in the processing of facial expressions as revealed by fMRI. J Cogn Neurosci 13:1035–47. [DOI] [PubMed] [Google Scholar]
- 18.Kanske P, Heissler J, Schönfelder S, et al. (2011) How to regulate emotion? Neural networks for reappraisal and distraction. Cereb Cortex 21:1379–88. [DOI] [PubMed] [Google Scholar]
- 19.Kienast T, Hariri AR, Schlagenhauf F, et al. (2008) Dopamine in amygdala gates limbic processing of aversive stimuli in humans. Nat Neurosci 11:1381–2. [DOI] [PubMed] [Google Scholar]
- 20.Koelsch S, Skouras S, Fritz T, et al. (2013) The roles of superficial amygdala and auditory cortex in music-evoked fear and joy. Neuroimage 81:49–60. [DOI] [PubMed] [Google Scholar]
- 21.Larson CL, Aronoff J, Sarinopoulos IC, Zhu DC (2009) Recognizing threat: a simple geometric shape activates neural circuitry for threat detection. J Cogn Neurosci 21:1523–35. [DOI] [PubMed] [Google Scholar]
- 22.Maquet P, Phillips C (1998) Functional brain imaging of human sleep. J Sleep Res 7 Suppl 1:42–7. [DOI] [PubMed] [Google Scholar]
- 23.Meier ML, Stämpfli P, Vrana A, Humphreys BK, Seifritz E, Hotz-Boendermaker S (2015): Fear avoidance beliefs in back pain-free subjects are reflected by amygdala-cingulate responses. Front Hum Neurosci. 9:424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mende-Siedlecki P, Verosky SC, Turk-Browne NB, Todorov A (2013) Robust selectivity for faces in the human amygdala in the absence of expressions. J Cogn Neurosci 25:2086–106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Molapour T, Golkar A, Navarrete CD, Haaker J, Olsson A (2015) Neural correlates of biased social fear learning and interaction in an intergroup context. Neuroimage. 121:171–183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Monk CS, Telzer EH, Mogg K, et al. (2008) Amygdala and ventrolateral prefrontal cortex activation to masked angry faces in children and adolescents with generalized anxiety disorder. Arch Gen Psychiatry 65:568–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Morris JS, Friston KJ, Büchel C, et al. (1998) A neuromodulatory role for the human amygdala in processing emotional facial expressions. Brain 121(Pt 1):47–57. [DOI] [PubMed] [Google Scholar]
- 28.Morris JS, Dolan RJ (2001) Involvement of human amygdala and orbitofrontal cortex in hunger-enhanced memory for food stimuli. J Neurosci 21:5304–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Mukherjee P, Whalley HC, McKirdy JW, et al. (2014) Altered amygdala connectivity within the social brain in schizophrenia. Schizophr Bull 40:152–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Murray BD, Kensinger EA (2014) The route to an integrative associative memory is influenced by emotion. PLoS One 9:e82372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Pasley BN, Mayes LC, Schultz RT (2004) Subcortical discrimination of unperceived objects during binocular rivalry. Neuron 42:163–72. [DOI] [PubMed] [Google Scholar]
- 32.Pichon S, Rieger SW, Vuilleumier P (2012) Persistent affective biases in human amygdala response following implicit priming with negative emotion concepts. Neuroimage 62:1610–21. [DOI] [PubMed] [Google Scholar]
- 33.Ponz A, Khatami R, Poryazova R, et al. (2010) Reduced amygdala activity during aversive conditioning in human narcolepsy. Ann Neurol 67:394–8. [DOI] [PubMed] [Google Scholar]
- 34.Sato W, Kochiyama T, Uono S, Yoshikawa S, Toichi M (2016) Direction of amygdala–neocortex interaction during dynamic facial expression processing. Cereb Cortex. bhw036. [DOI] [PubMed] [Google Scholar]
- 35.Schienle A, Ubel S, Schöngaßner F, et al. (2014) Disgust regulation via placebo: an fMRI study. Soc Cogn Affect Neurosci 9:985–990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Schmitgen MM, Walter H, Drost S, Rückl S, Schnell K (2016): Stimulus-dependent amygdala involvement in affective theory of mind generation. Neuroimage. 129:450–459. [DOI] [PubMed] [Google Scholar]
- 37.Skelly LR, Decety J (2012) Passive and motivated perception of emotional faces: qualitative and quantitative changes in the face processing network. PLoS One 7:e40371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Spielberg JM, Forbes EE, Ladouceur CD, et al. (2015) Pubertal testosterone influences threat-related amygdala-orbitofrontal cortex coupling. Soc Cogn Affect Neurosci 10:408–415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sripada C, Angstadt M, Kessler D, et al. (2014) Volitional regulation of emotions produces distributed alterations in connectivity between visual, attention control, and default networks. Neuroimage 89:110–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Stegmayer K, Usher J, Trost S, et al. (2015) Disturbed cortico-amygdalar functional connectivity as pathophysiological correlate of working memory deficits in bipolar affective disorder. Eur Arch Psychiatry Clin Neurosci 265:303–311. [DOI] [PubMed] [Google Scholar]
- 41.Sterpenich V, D’Argembeau A, Desseilles M, et al. (2006) The locus ceruleus is involved in the successful retrieval of emotional memories in humans. J Neurosci 26:7416–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Stevens JS, Jovanovic T, Fani N, et al. (2013) Disrupted amygdala-prefrontal functional connectivity in civilian women with posttraumatic stress disorder. J Psychiatr Res 47:1469–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Tottenham N, Shapiro M, Telzer EH, Humphreys KL (2012) Amygdala response to mother. Dev Sci 15:307–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Troiani V, Price ET, Schultz RT (2014) Unseen fearful faces promote amygdala guidance of attention. Soc Cogn Affect Neurosci 9:133–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Van Wingen GA, Geuze E, Vermetten E, Fernández G (2011) Perceived threat predicts the neural sequelae of combat stress. Mol Psychiatry 16:664–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Williams LM, Das P, Liddell BJ, et al. (2006) Mode of functional connectivity in amygdala pathways dissociates level of awareness for signals of fear. J Neurosci 26:9264–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Winecoff A, Labar KS, Madden DJ, et al. (2011) Cognitive and neural contributors to emotion regulation in aging. Soc Cogn Affect Neurosci 6:165–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Xu P, Gu R, Broster LS, et al. (2013) Neural basis of emotional decision making in trait anxiety. J Neurosci 33:18641–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Yoder KJ, Porges EC, Decety J (2015) Amygdala subnuclei connectivity in response to violence reveals unique influences of individual differences in psychopathic traits in a nonforensic sample. Hum Brain Mapp. 36:1417–1428. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.