

Abstract
Recent trends in methamphetamine (METH) misuse and overdose suggest society is inadvertently overlooking a brewing METH crisis. In the past decade, psychostimulant-related lethal overdoses and hospitalizations have skyrocketed 127 and 245%, respectively. Unlike the opioid crisis, no pharmaceutical interventions are available for treating METH use disorder or reversing overdose. Herein, we report the first active vaccine that offers protection from lethal (+)-METH challenge in male Swiss Webster mice. This vaccine formulation of (S)MLMH-TT adjuvanted with CpG ODN 1826 + alum successfully raised anti-METH antibodies in high titers, reduced (+)-METH distribution to the brain, and lowered (+)-METH-associated stereotypies in a hyperlocomotion assay. A comparison of enantiomeric haptens and the racemate elucidated the importance of employing (S)-stereochemistry in METH hapten design for optimal protection.
Methamphetamine (METH), an illicit, psychostimulatory phenethylamine, produces a highly addictive, euphoric high by releasing high concentrations of dopamine, serotonin and norepinephrine in the brain.1 Dopamine, in particular, motivates and reinforces drug-taking behaviors that may ultimately produce METH use disorder. The 2017 National Survey on Drug Use and Health (NSDUH) reported 1.6 million past year users in the United States; included in this number are nearly one million Americans with clinically significant METH use disorder.2 Strikingly, this number represents a 29% increase in the incidence of METH use disorder compared to 2016.2
While the threat of METH-induced overdose is not as widely publicized as the opioid crisis, rates of psychostimulant-associated overdose death have risen since 2010. METH, now being described as the “forgotten killer”, is responsible for more overdose deaths in Texas and Colorado than opioids, and in California, the number of psychostimulant-related lethal overdoses rose by 127% from 2008 to 2013. For comparison, the number of opioid-related overdose deaths rose by 8.4%.3
Unlike the opioid crisis, there are no pharmaceutical options for the treatment of METH use disorder or overdose reversal. Because the pharmacology of METH is complicated by the involvement of multiple monoamine neurotransmitters, a traditional approach to therapeutic design, namely receptor modulation, proves challenging. Alternatively, our laboratory has leveraged the field of immunopharmacotherapy with the goal of developing an active vaccine for METH; an active vaccine uses a METH analogue, the hapten, to generate high concentration, high affinity antibodies that sequester drug in circulation. To date, nine experimental METH vaccines have been reported by our laboratory and others, but none have advanced past in vivo, murine models. Herein, we report a key advancement in METH vaccine development by designing the first vaccine that imparts protection from lethal doses of METH.
Three components are required in the design of a small molecule conjugate vaccine: the hapten, the immunogenic carrier protein, and an adjuvant system. Extensive efforts in our laboratory previously identified optimal carrier protein and adjuvant systems, tetanus toxoid (TT) and CpG ODN 1826 + alum, respectively, both in the context of METH and other drugs of abuse. With these components optimized, we focused on hapten design in this work to deliver a clinically viable, anti-METH vaccine candidate. The fundamental requirements in hapten design aim to chemically append a linker through which the immunogenic carrier can be covalently conjugated, all while maintaining as much of the native drug structure as possible.
Previous METH hapten design evaluated phenyl (red) and N-methyl (blue) substituted haptens (Figure 1).4−12 Later generation vaccines in both approaches generated antibodies with nanomolar Kd affinity for (+)-METH, reduced (+)-METH distribution to the CNS, and attenuated (+)-METH-induced self-administration, hyperthermia, conditioned place preference, and/or hyperlocomotion in murine models. Despite these promising early results, no active vaccine has reached clinical trials. The major limitation to currently available METH vaccines is surmountable antibody protection, a caveat common to all drugs of abuse vaccines. For example, MH6, a Janda Laboratory vaccine that we advanced furthest in preclinical development, lost efficacy in a self-administration model.13 In an effort to address surmountability, the current investigation sought to identify a vaccine efficacious against lethal doses of (+)-METH. We propose that identifying a vaccine candidate more equip to protect against the highest concentrations of (+)-METH, a scenario representative of drug seekers increasing dosages to achieve a high, will better translate to clinical utility.
Figure 1.
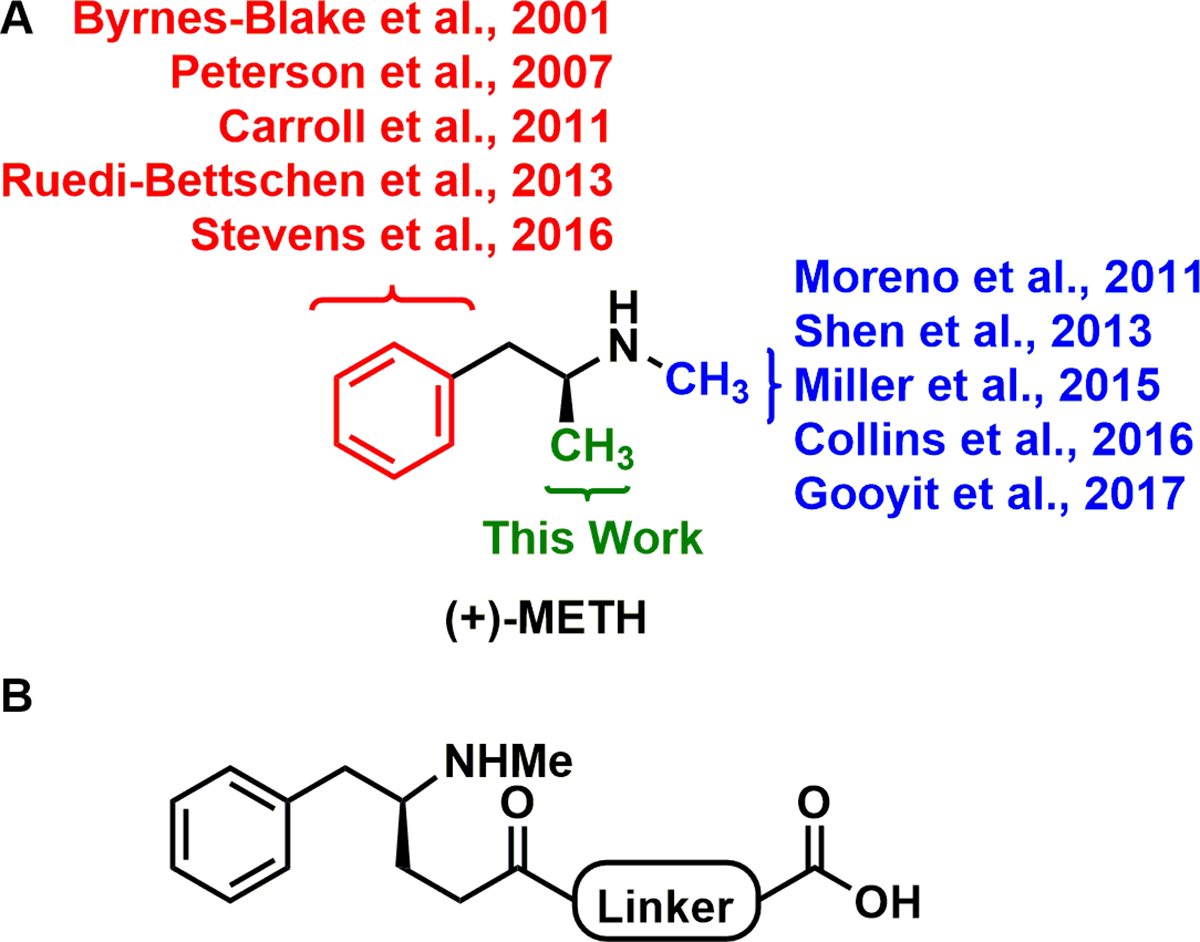
Structure of (+)-methamphetamine and the (S)-MLMH hapten. (A) Previous efforts to develop an active METH vaccine have evaluated conjugation to immunogenic proteins via the phenyl (red) and N-methyl (blue) substructures of the drug. References for these efforts are identified.4−10 (B) This work investigated modifications of the methyl substructure for the first time.
METH is a particularly challenging target for immunopharmacotherapy because of its low molecular weight structure (150 kDa), limited chemical-epitopes for antibody recognition, and similarity to endogenous monoamines. In this study, we reasoned that modifying the methyl substructure (Figure 1, green), being the smallest chemical-epitope of the native METH scaffold, would least influence the specificity of the generated antibodies to the drug. In our initial design, three linker identities were considered, 7-propionamidoheptanoic acid, (3-propionamidopropanoyl)glycine], and 5-oxo-5-((2-oxo-2-((5-propionamidopentyl)amino)ethyl)amino)pentanoic acid (Figure S1), before ultimately arriving at 7-propionamidoheptanoic acid to garnish the optimal series of methyl-linked METH haptens (MLMHs), (S)MLMH, (R)MLMH, and (R/S)MLMH. This work marks the first comparison of enantiomeric and racemic haptens. METH exists in two stereoisomers; the dextrorotatory isomer [(+)-METH/(S)-METH] is significantly more potent than (−)-METH.14 Yet, despite this knowledge, the field of immunopharmacotherapy had yet to compare enantiomeric and racemic haptens head-to-head.
The (S), (R), and (R/S) MLMH haptens were synthesized as depicted in Scheme 1 from their respective N-Boc-phenylalaninals (d, l, or racemic) with overall yields of 9.2% (S), 10.5% (R), and 8.9% (R/S). To conjugate the haptens to TT, carboxylic acids were first activated with N-hydroxysulfo-succinimide (sulfo-NHS) using 1-ethyl-3-(3-(dimethylamino)-propyl) carbodiimide (EDCI), then treated with TT to afford the desired hapten-TT immunoconjugates.
Scheme 1. Synthesis of METH Haptens.
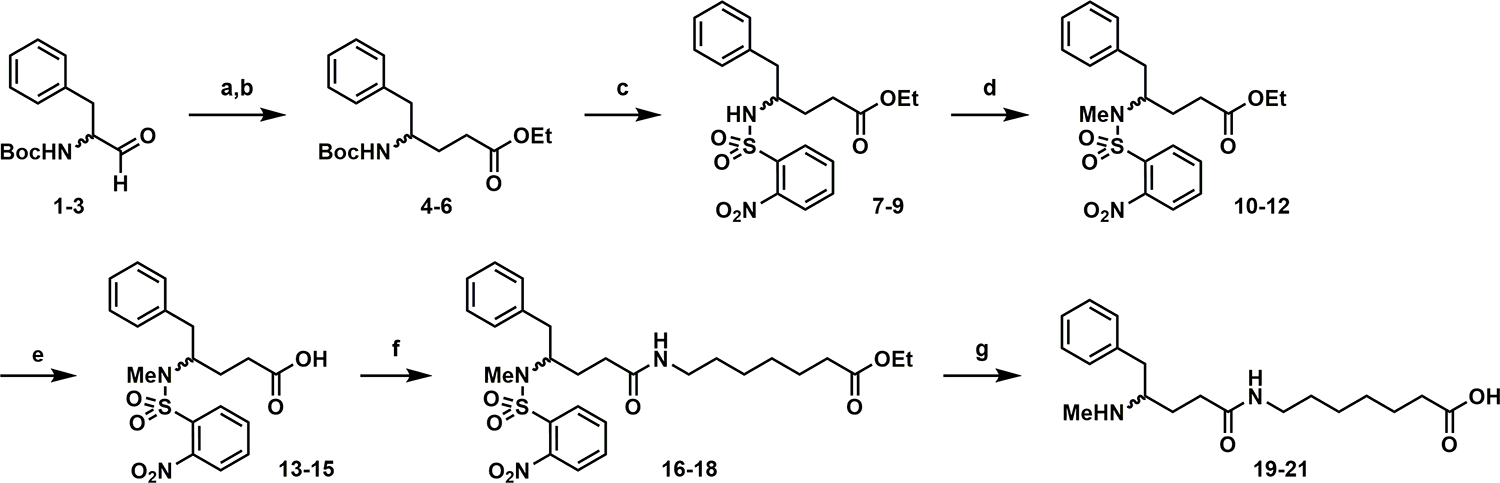
aReagents and Conditions: (a) Ph2P = CHCO2Et, DCM, 0 °C to rt, (S) 90%, (R) 90%, (R/S) 76%; (b) H2, Pd/C, EtOH, rt, (S) 85%, (R) 98%, (R/S) 95%; (c) TFA, DCM, rt, then NsCl, DIPEA, DCM, 0 °C to rt, (S) 54%, (R) 52%, (R/S) 86%; (d) MeI, Cs2CO3, DMF, rt, (S) 91%, (R) 79%, (R/S) 81%; (e) NaOH, MeOH, rt, (S) quant., (R) 94%, (R/S) quant.; (f) ethyl 7-aminoheptanoate, HATU, DIPEA, DMF, rt, (S) 94%, (R) 89%, (R/S) 93%; (g) 4-tBuPhSH, Cs2CO3, DMF, then NaOH, MeOH, (S) 26%, (R) 34%, (R/S) 19%.
As discussed, vide supra, the MLMH-TT immunoconjugates were formulated with alum and CpG ODN 1826 adjuvants, then administered intraperitoneally (i.p.) to Swiss Webster mice at 0, 2, and 4 weeks. Antibody titers were assessed by enzyme-linked immunosorbent assay (ELISA) against the corresponding hapten-BSA conjugate, namely (S)MLMH, (R)MLMH, or (R/S)MLMH-BSA (Figure 2A). Overall, robust antibody titers were observed for the experimental MLMH vaccines in comparison to vehicle (TT and alum + CpG ODN 1826). Importantly, higher titers were observed as the immunization study continued, evidence of a mounting immune response. Notably, the increase in titers for the (S)MLMH vaccine between the first and second bleeds was more pronounced than the change associated with (R)MLMH and (R/S)MLMH vaccines, possibly alluding to a stronger immune response. To quantify antibody IC50’s for (+)-METH raised by the (S)MLMH-TT, (R)MLMH-TT, and (R/S)MLMH-TT vaccines, surface plasmon resonance (SPR) was performed (Table S1). (S)MLMH-BSA, (R)MLMH-BSA, and (R/S)MLMH-BSA were immobilized on a SPR chip, and binding to (S)MLMH, (R)MLMH, and (R/S)MLMH serum antibodies was measured in the competitive environment of (+)-METH. As anticipated, hapten chirality was critical as antibodies raised by (S)MLMH-TT exhibited 10-fold higher affinity for (+)-METH compared to (R)MLMH-TT and (R/S)MLMH-TT raised antibodies (~0.2 vs ~2 μM).
Figure 2.
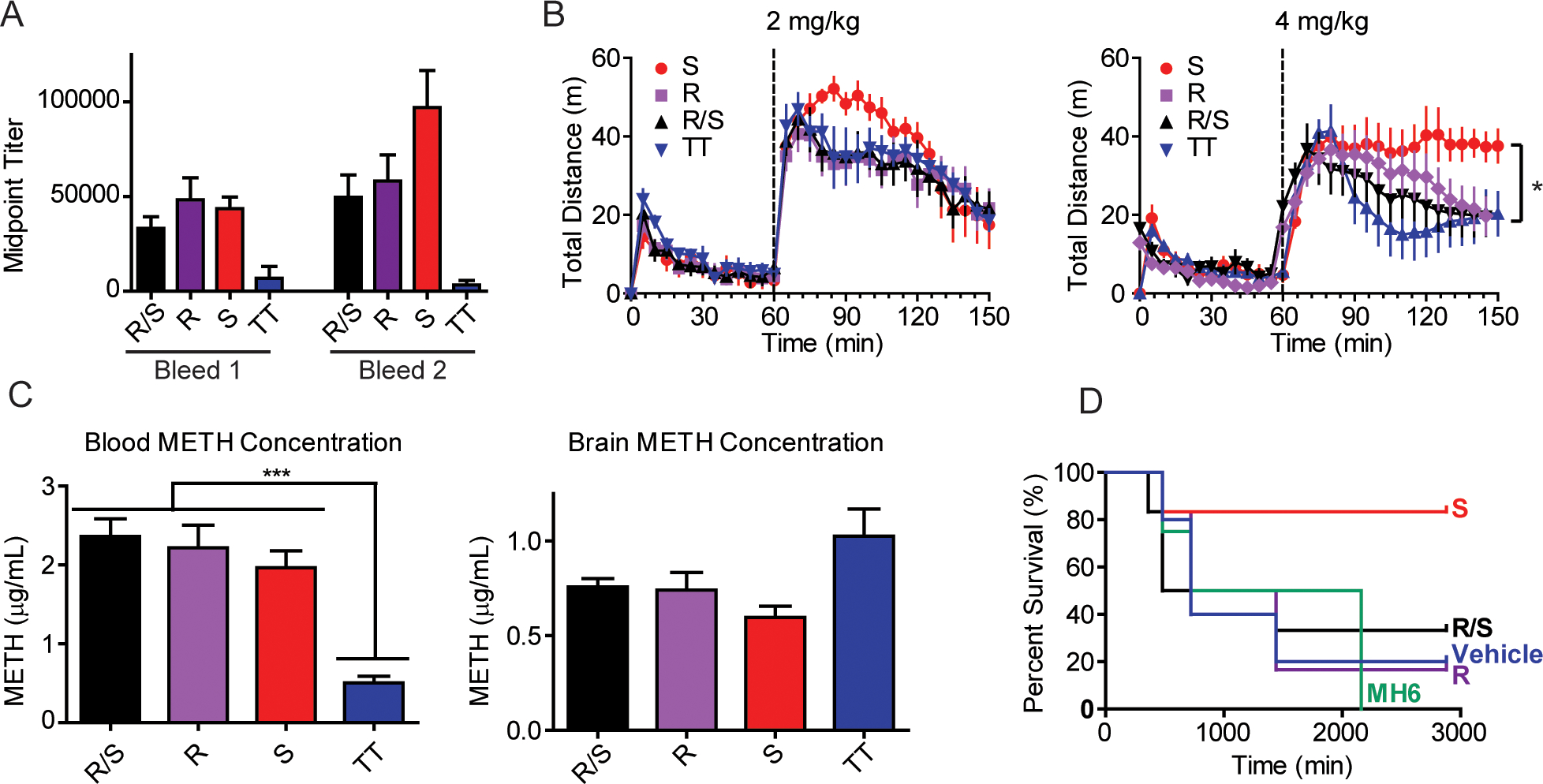
Evaluation of MLMH vaccines. (A) Anti-METH antibody midpoint titers measured by ELISA using serum from vaccinated mice (n = 6/group) on days 21 (bleed 1) and 35 (bleed 2). Bars denote means ± SEM (B) (+)-METH-induced hyperlocomotor activity in vaccinated mice (n = 6/group) at 2 and 4 mg/kg. Bars denote means ± SEM *P < 0.05 determined by one-way ANOVA. (C) Concentration of (+)-METH in the blood and brains of vaccinated and naive mice (n = 6/group). Bars denote means ± SEM ***P < 0.001 determined by one-way ANOVA. (D) Kaplan−Meier curve for survival in vaccinated groups (n = 5 or 6/group). The efficacy of MH6-TT, an anti-METH vaccine previously reported by the Janda Laboratory, was evaluated and shown for comparison.
As a stimulant, METH induces a quantifiable increase in rodent locomotor activity upon exposure with a ceiling of stereotypy. Stereotypies are repetitive movements that may present as adverse events in response to high concentrations of drug challenge. Colloquially, these behaviors would be referred to as “tweaking”. Using locomotor behavior as a model, vaccinated mice were challenged with (+)-METH at 1, 2, and 4 mg/kg concentrations and hyperlocomotor activity was evaluated using ANY-Maze video tracking software (Stoelting Co.). At low concentrations, vaccine efficacy can be measured by an ability to block METH-induced hyperlocomotion; whereas at high concentrations, vaccine efficiency is gauged by an ability to prevent stereotypy. The control group of mice vaccinated with TT and alum + CpG ODN 1826 alone exhibited elevated hyperlocomotor activity upon (+)-METH challenge at 1 mg/kg and the appearance of minor stereotypy and dramatic stereotypy at 2 mg/kg and 4 mg/kg, respectively (Figures 2B and S2). METH induced stereotypy presents as a freezing behavior in mice, where the animals rear on their hind legs and aggressively clean their face. Counterintuitively, this behavior is opposite to stimulant-induced hyperlocomotor activity and appears as a sharp decline in hyperlocomotion after drug exposure. Mice experiencing normal elimination of (+)-METH show a gradual return to baseline. Analysis of experimental vaccine cohorts demonstrated that all vaccines convey minimal protection to (+)-METH at low 1 mg/kg doses as animals exhibited equivalent locomotor activity to vehicle immunized mice. However, at 2 mg/kg and especially 4 mg/kg, (R)MLMH-TT, (R/S)MLMH-TT and vehicle immunized mice exhibited stereotypy, whereas (S)MLMH-TT immunized mice exhibited minimal to no adverse effects. Taken together, this finding is significant in that (S)MLMH-TT vaccinated mice are protected from high, potentially lethal (+)-METH concentrations.
To confirm these findings, biodistribution of (+)-METH was investigated. In this study, vaccinated and naive mice werë treated with 4 mg/kg (+)-METH, then sacrificed after 15 min by decapitation. Drug concentrations in the blood and brain were quantified by liquid chromatography mass spectrometry. Ultimately, significantly elevated levels of (+)-METH were observed in the plasma of (S)MLMH-TT, (R)MLMH-TT, and (R/S)MLMH-TT vaccinated mice versus vehicle vaccinated mice. Analogously, though not significant, reduced levels of (+)-METH were detected in the brains of (S)MLMH-TT, (R)MLMH-TT, and (R/S)MLMH-TT vaccinated mice versus vehicle. This biodistribution data demonstrated that vaccine-generated antibodies effectively scavenged free drug in the blood (Figure 2C). Specifically, mice vaccinated with (S)MLMH-TT, (R)MLMH-TT, and (R/S)MLMH-TT appropriated nearly 3.9, 4.4, and 4.7 times more (+)-METH in the blood than unvaccinated mice.
Based on these results and the findings of the hyperlocomotor activity assay, we probed the ability of these MLMH vaccines to protect against (+)-METH lethal challenge. To date, no report has described the successful use of an active vaccine to protect against METH overdose. Vehicle mice dosed i.p. with 20 mg/kg of (+)-METH exhibited a 20% survival rate at 48 h, while (R)MLMH-TT vaccinated mice exhibited 16.7% surivial, (R/S)MLMH-TT vaccinated mice exhibited 33.3% survival, and (S)MLMH-TT vaccinated mice exhibited 83.3% survival (Figure 2D). The MLMH vaccines significantly outperformed MH6-TT, an anti-METH vaccine that had previously exhibited promising results in modifying (+)-METH induced behaviors.7,13,15,16 Importantly, this report marks the first direct comparison of two anti-METH vaccines of unique classes, namely N-methyl linked and methyl-linked METH haptens, and the first evaluation of MH6-TT in a lethality study (see SI for a complete set of MH6 data vs the MLMH series).
Taken together, the results of ELISA, SPR, behavioral testing, biodistribution and lethal challenge revealed that conjugating METH immunogens via the methyl substructure (MLMH) is a successful strategy in METH hapten design. Importantly, a direct comparison of (S) and (R) enantiomers and the racemate unequivocally defined (S) stereochemistry as a fundamental requirement of hapten design. While each vaccine, (S)MLMH-TT, (R)MLMH-TT, and (R/S)MLMH-TT, generated antibodies and affected blood/brain partitioning, only the (S)MLMH-TT vaccine effectively prevented (+)-METH-associated stereotypy and lethality. These results suggest that (S)MLMH-TT presents a viable candidate for further consideration in more advanced rodent and nonhuman primate preclinical models.
Supplementary Material
ACKNOWLEDGMENTS
Research was supported by the National Institutes of Health grants R01 DA024705 (K.D.J.) and F32 DA044692 (M.E.O.), and the Skaggs Institute for Chemical Biology (K.D.J.).
Footnotes
The authors declare no competing financial interest.
REFERENCES
- (1).Sulzer D; Sonders MS; Poulsen NW; Galli A Mechanisms of neurotransmitter release by amphetamines: a review. Prog. Neurobiol. 2005, 75 (6), 406–33. [DOI] [PubMed] [Google Scholar]
- (2).Substance Abuse and Mental Health Services Administration Results from the 2016 National Survey on Drug Use and Health: Detailed Tables (HHS Publication No. SMA 17-5044, NSDUH Series H-52). https://www.samhsa.gov/data/report/results-2016-national-survey-drug-use-and-health-detailed-tables (accessed June 9, 2019). [Google Scholar]
- (3).Gorman A Opioids Are In the Spotlight. But Meth Hospitalizations Are Surging. https://khn.org/news/overshadowed-by-opioids-meth-is-back-and-hospitalizations-surge/ (accessed June 9, 2019). [Google Scholar]
- (4).Byrnes-Blake KA; Carroll FI; Abraham P; Owens SM Generation of anti-(+)methamphetamine antibodies is not impeded by (+)methamphetamine administration during active immunization of rats. Int. Immunopharmacol. 2001, 1 (2), 329–38. [DOI] [PubMed] [Google Scholar]
- (5).Carroll FI; Blough BE; Pidaparthi RR; Abraham P; Gong PK; Deng L; Huang X; Gunnell M; Lay JO Jr.; Peterson EC; Owens SM Synthesis of mercapto-(+)-methamphetamine haptens and their use for obtaining improved epitope density on (+)-methamphetamine conjugate vaccines. J. Med. Chem. 2011, 54 (14), 5221–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (6).Peterson EC; Gunnell M; Che Y; Goforth RL; Carroll FI; Henry R; Liu H; Owens SM Using hapten design to discover therapeutic monoclonal antibodies for treating methamphetamine abuse. J. Pharmacol. Exp. Ther. 2007, 322 (1), 30−9. [DOI] [PubMed] [Google Scholar]
- (7).Moreno AY; Mayorov AV; Janda KD Impact of distinct chemical structures for the development of a methamphetamine vaccine. J. Am. Chem. Soc. 2011, 133 (17), 6587–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (8).Shen XY; Kosten TA; Lopez AY; Kinsey BM; Kosten TR; Orson FM A vaccine against methamphetamine attenuates its behavioral effects in mice. Drug Alcohol Depend. 2013, 129 (1−2), 41–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (9).Collins KC; Schlosburg JE; Bremer PT; Janda KD Methamphetamine Vaccines: Improvement through Hapten Design. J. Med. Chem. 2016, 59 (8), 3878–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (10).Gooyit M; Miranda PO; Wenthur CJ; Ducime A; Janda KD Influencing Antibody-Mediated Attenuation of Methamphetamine CNS Distribution through Vaccine Linker Design. ACS Chem. Neurosci. 2017, 8 (3), 468–472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (11).Ruedi-Bettschen D; Wood SL; Gunnell MG; West CM; Pidaparthi RR; Carroll FI; Blough BE; Owens SM Vaccination protects rats from methamphetamine-induced impairment of behavioral responding for food. Vaccine 2013, 31 (41), 4596–4602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (12).Stevens MW; Gunnell MG; Tawney R; Owens SM Optimization of a methamphetamine conjugate vaccine for antibody production in mice. Int. Immunopharmacol. 2016, 35, 137–141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (13).Miller ML; Aarde SM; Moreno AY; Creehan KM; Janda KD; Taffe MA Effects of active anti-methamphetamine vaccination on intravenous self-administration in rats. Drug Alcohol Depend. 2015, 153, 29–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (14).Harris JE; Baldessarinig RJ Uptake of (3H)-catecholamines by homogenates of rat corpus striatum and cerebral cortex: effects of amphetamine analogues. Neuropharmacology 1973, 12 (7), 669–679. [DOI] [PubMed] [Google Scholar]
- (15).Miller ML; Moreno AY; Aarde SM; Creehan KM; Vandewater SA; Vaillancourt BD; Wright MJ Jr.; Janda KD; Taffe MA A methamphetamine vaccine attenuates methamphetamine-induced disruptions in thermoregulation and activity in rats. Biol. Psychiatry 2013, 73 (8), 721–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (16).Nguyen JD; Bremer PT; Hwang CS; Vandewater SA; Collins KC; Creehan KM; Janda KD; Taffe MA Effective active vaccination against methamphetamine in female rats. Drug Alcohol Depend. 2017, 175, 179–186. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.