

Abstract
A tandem pair of nearly identical genes from Serpulina hyodysenteriae (B204) were cloned and sequenced. The full open reading frame of one gene and the partial open reading frame of the neighboring gene appear to encode secreted proteins which are homologous to, yet distinct from, the 39-kDa extracytoplasmic protein purified from the membrane fraction of S. hyodysenteriae. We have designated these newly identified genes vspA and vspB (for variable surface protein).
Serpulina (Treponema) hyodysenteriae, a gram-negative anaerobic spirochete, is the etiologic agent of swine dysentery (5, 19, 21). After ingestion the pathogen colonizes the colon of the infected animal, commonly a postweaning pig, leading to disease characterized by severe mucohemorrhagic diarrhea, dehydration, rapid weight loss, and, in some cases, death. In recent years several genes have been cloned from S. hyodysenteriae which may be involved in the colonization of the swine colon and the pathogenicity of the spirochete or in the generation of a protective immune response by the host. These include the flaA1, flaB1, and flaB2 genes, encoding components of the spirochete’s periplasmic flagella (4, 8, 9, 15, 16); the smpA gene, encoding an outer membrane lipoprotein (22); and the hlyA, hlyB, and hlyC genes, encoding hemolysins (7, 14, 20).
We have previously described a rapid method for releasing extracytoplasmic proteins (ECP) from S. hyodysenteriae (B204) with the nonionic detergent Tween 20 and the subsequent subfractionation and purification of several of these proteins by centrifugation, differential urea solubility, and reverse-phase chromatography (4). One ECP, characterized by its partitioning during subfractionation, sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis, and amino-terminal sequencing, was a sedimentable, urea-insoluble 39-kDa protein. In this paper we report the identification of two closely related and contiguous genes which have striking homology to each other and are predicted to have marked homology to the still-unidentified structural gene for the 39-kDa ECP we have purified.
S. hyodysenteriae B204, obtained from Joann Kinyon, Iowa State University, was grown anaerobically (80% nitrogen, 10% carbon dioxide, 10% hydrogen) from a 5 to 10% inoculum in a Braun Biostat M fermentor at 37°C, pH 6.8, in Difco brain heart infusion broth (37 g/liter) supplemented with 5% heat-inactivated fetal bovine serum, 0.5% dextrose, and 20 mg of spectinomycin (Sigma)/liter. Cells were harvested by centrifugation in late log phase (A600 = 1.5 to 2) 18 to 24 h after inoculation, with a yield of approximately 10 g (wet weight) per liter.
Protein biochemistry.
Enzymatic iodination of surface proteins of intact S. hyodysenteriae cells was performed essentially as described by Marchalonis et al. (12). Twenty milliliters of an exponentially growing culture of S. hyodysenteriae (approximately 5 × 108 cells/ml) was centrifuged, washed with 0.5 volume of phosphate-buffered saline (PBS) (10 mM sodium phosphate [pH 7.2], 150 mM NaCl), and resuspended in a final volume of 2 ml of PBS. First, 25 μl of 0.5 M sodium phosphate (pH 7.3) was added, then 10 μl of lactoperoxidase (CalBiochem) (made up to 200 IU/ml in PBS), then 40 μl of 10 μM hydrogen peroxide, and finally 450 μCi of 125I (20 mCi/ml in 0.1 N NaOH). The mixture was incubated at 30°C for 5 min, another 40 μl of hydrogen peroxide was added, and after an additional 10 min of incubation the reaction was stopped by the addition of 400 μg of tyrosine (40 μl of a 10 mg/ml solution in PBS). A whole-cell lysate sample of iodinated S. hyodysenteriae was electrophoresed on a 12% acrylamide gel, dried, and then exposed on X-ray film for 2 h before development. We observed one predominant iodinated component (Fig. 1), whose electrophoretic mobility matched that of the Coomassie-staining 39-kDa protein purified from S. hyodysenteriae (4). Subsequent extraction of these labeled cells with Tween 20 and urea showed that this same iodinated protein partitioned like the 39-kDa protein (specifically, it is released by Tween 20 from the cells and is insoluble following extraction with 6 M urea [data not shown]). Trace amounts of iodinated protein components of both higher and lower molecular masses were also noted.
FIG. 1.
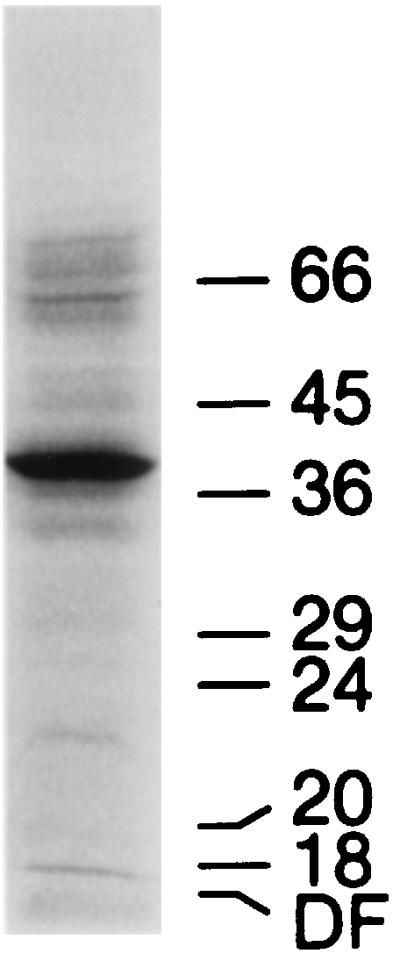
Autoradiography of a whole-cell lysate sample of iodinated S. hyodysenteriae cells after gel electrophoresis with the buffer system of Laemmli (10). The numbers correspond to the MW (in thousands) of prestained protein markers (Diversified Biotech). DF, dye front.
The cell pellets were washed with 10 mM potassium acetate and 150 mM potassium chloride (pH 4.75) (at 4°C) and used immediately or frozen at −20°C. As previously described (4), the cell pellet was resuspended in acetate buffer, pH 4.75, and extracted with detergent (1% [wt/vol] Tween 20) and the released proteins were collected by ultracentrifugation, extracted with urea, and centrifuged again to collect the urea-insoluble 39-kDa ECP. The urea-insoluble 39-kDa ECP was further purified by size exclusion chromatography on Sepharose 6B resin (BioRad) in 25 mM Tris-Cl (pH 6.8)–1% SDS and concentrated by precipitation with acetone prior to digestion with 1 μg of endoproteinase Lys-C (Boehringer Mannheim) per 75 μg of purified protein in 50 mM Tris-Cl (pH 8.5)–0.1% SDS for 4 to 12 h at 37°C. Peptide fragments were purified on an analytical C4 column developed with a gradient from 0 to 100% acetonitrile-isopropanol (2:1) and 0.1% trifluoroacetic acid and monitored at 214 nm. Peptide sequencing was performed by sequential Edman degradation on an automated gas phase sequencer (Applied Biosystems). Seven peptide fragments derived from the 39-kDa ECP were sequenced, including its N terminus and six internal LysC digest-generated peptides, which yielded a sequence of 213 residues out of approximately 350 predicted for a 39-kDa protein. These data are presented in Fig. 2.
FIG. 2.

Comparison of amino acid sequences for actual and predicted 39-kDa products. The LysC peptide fragments of the 39-kDa protein are presented on the top line. Question marks indicate sequence ambiguity at the specific cycle position. By the numbering system used in the figure, the peptides sequenced correspond to (i) Met1 to Arg41, (ii) Pro132 to Lys166, (iii) Thr174 to Ala220, (iv) Ala266 to Lys296, (v) His297 to Lys311, (vi) Val312 to Asn341, and (vii) Arg346 to Gln371. The sequences are aligned for best fit, with possible gaps or deletions between proteins indicated by dashed lines. Conserved sequences are shaded, the apparent variable regions (six are identified) are underlined (solid bar) and numbered, and apparent hydrophobic regions are underlined (broken bar).
Molecular cloning and analysis.
S. hyodysenteriae B204 DNA was prepared from a 1-liter cell pellet of early-log-phase cells (optical density at 600 nm = 0.5) and purified in CsCl gradients according to the method of Maniatis et al. (11). Linkers containing an EcoRI restriction site were blunt end ligated to an AluI partial digest of the S. hyodysenteriae DNA, and a library was created by cloning these fragments into the EcoRI site of lambda phage gt11 (Promega). The Escherichia coli strain Y1090r− (Promega) was used to screen the lambda gt11 phage library, and strains JM83 and DH5α were used for subcloning and DNA sequencing. The cloning of the vspA gene was done with the (degenerate) oligonucleotides COD555 [ATG-TA(T/C)-GG(T/C/A/G)-GA(T/C)-AG(T/C/ A/G)-GA], derived from amino acids Met1 to Asp6, and COD553 [TGG-AT(T/C/A)-GA(T/C)-TT(T/C)-TT(T/C/A/ G)-AC], derived from amino acids Trp8 to Thr13 of the 39-kDa ECP. Oligonucleotide probes were synthesized with an automated DNA synthesizer (Biosearch 8700) and purified by acrylamide gel prior to labeling with [γ-32P]dATP (New England Nuclear) and T4 polynucleotide kinase (Boehringer Mannheim). Degenerate oligonucleotides or plasmid subclone fragments were labeled by nick translation and used to probe Southern blots or to screen the phage library (11). A phage containing a 1.5-kb EcoRI fragment which hybridized to both oligonucleotide probes was identified, and its insert was subcloned into pUC19 (pTrep106) and sequenced by the dideoxy termination method of Sanger et al. (17). It was found to contain an extended open reading frame (ORF-1) which encoded 386 amino acids as shown in Fig. 3. The first 21 amino acids encoded by the ORF were typical of those corresponding to cleavable signal peptides directing transmembrane transport (23). The following 365 codons are predicted to encode a protein whose molecular weight (MW) is 41,000. Codon 22 of ORF-1 matches the N-terminal Met determined for the purified 39-kDa protein, consistent with a signal peptide-processing event between Gly21 and Met22 (23). Glycine or other small amino acids are typically found adjacent to the new N termini generated by signal peptidase processing. From Met22 onward the predicted polypeptide sequence matched 36 of the 40 amino acids obtained from the amino terminus of the 39-kDa ECP (Fig. 2). However, the four mismatches noted could not be reconciled with the nucleotide sequence of ORF-1 in pTrep106.
FIG. 3.


Nucleotide sequence of vspA and vspB genes from S. hyodysenteriae (length, 2,091 nucleotides). Insert sequences from two overlapping plasmids, pTrep106 and pTrep330, were combined, as indicated by arrows adjacent to Alu cloning sites (∗). The underlined sequences (AGGT) upstream of the ORFs are apparent RBSs. The underlined sequences following ORF-1 form an apparent stem-loop structure. The first amino acid obtained from the isolated 39-kDa protein (Met, indicated with a #) is an apparent site of posttranslational signal peptide processing for both vsp gene products.
The beginning of a possible second ORF, ORF-2, in pTrep106 was also identified. This ORF began 156 bp after the stop codon found at the end of ORF-1 and was disrupted 121 bp (40 amino acids) later at the 3′ AluI cloning site. The first 21 codons of ORF-2 differed from those of ORF-1 at only one codon (no. 4) within the predicted signal peptide region. The next 19 codons of ORF-2 were identical to those of ORF-1. A second phage hybridizing to a (nick-translated) unique 500-bp SphI-BclI fragment from pTrep106 (encoding the first 150 amino acids of the product of vspA) was isolated, and a 1.3-kb EcoRI fragment was subcloned and sequenced. This subclone (pTrep 330) contained a 700-bp overlap with the upstream pTrep106 cloned fragment and thus was able to extend the sequence information for ORF-2 by an additional 600 bp.
A comparison of the actual peptide sequence determined for the isolated 39-kDa protein to the predicted sequences for ORF-1 and ORF-2 is shown in Fig. 2. It would appear that neither of the cloned genes encodes the protein actually isolated from S. hyodysenteriae, as there were 22 mismatches out of 124 residues identified. Interestingly, most of the mismatches were clustered together. Further examination of the predicted protein sequences suggests at least six regions of variability (defined as ≥6 consecutive residue mismatches or 8 mismatches out of 10 residues) between the products of ORF-1 and ORF-2, two of which are also regions of nonhomology with the limited peptide sequence determined for the 39-kDa protein. These variable regions vary from 6 to 15 amino acids in size, and they also appear to be evenly spaced (separated by 27 to 32 amino acids) within the coding sequence. Five hydrophobic regions of 20 residues or more occur within the product of the ORF of vspA: one is at the N terminus (signal peptide), and the other four are evenly spaced throughout the coding sequence (Fig. 2) and may be membrane-spanning domains.
Both cloned ORFs are preceded by a likely ribosome binding site (RBS) within the sequence AGGTA, with the 5′ guanine at position −10 for both genes. The upstream sequences of these two genes are identical at 41 of 44 nucleotides. The genes also have in common a Pribnow consensus sequence (E. coli “−10 region”) of TATAAT about 25 bp preceding the RBS. The gene corresponding to ORF-2 appears to have a −35 promoter sequence (TTTACA) located 16 bp upstream of the −10 sequence. The upstream promoter sequences for the gene sequence corresponding to ORF-1 are not contained in pTrep106. We also note an inverted repeat sequence downstream from the termination codon for the ORF-1 gene which could form a stem-loop structure similar to that observed for the flaB1 flagellar gene of S. hyodysenteriae (4), which is possibly involved with rho-independent transcription termination.
Concluding remarks.
Surface iodination of S. hyodysenteriae identified a predominant component of the cell which had the same apparent MW and extraction properties as the 39-kDa protein released from whole cells by treatment with Tween 20 (4). It is likely that this 39-kDa protein is also one of the several surface proteins iodinated on cells incubated in neutral-pH buffer as reported by Wannemuehler et al. (24). We sought to obtain both its amino acid and nucleotide sequences in hopes of revealing a possible role this protein might play in either the pathogenesis of infection with S. hyodysenteriae or the stimulation of protective antibodies in animals which have recovered (and become immune to further infection). Such a role has already been suggested for a 16-kDa envelope antigen of S. hyodysenteriae (18). To our surprise, we found a tandem pair of closely related genes, neither of whose products precisely matches the amino acid sequence obtained from the 39-kDa ECP. We believe that this protein is the product of yet another closely related gene, an idea supported by the observation that some of the oligomer probes used in the cloning were found to hybridize to more than one region of a Southern blot of HindIII-digested genomic DNA. Since neither of the cloned fragments within pTrep106 or pTrep330 contains a HindIII site, the hybridizing regions of both of the cloned genes should reside on a single genomic HindIII fragment.
Our results indicated that the cloned genes encode apparently secreted proteins which have several potential membrane-spanning domains and share significant identity with the predominant surface-accessible membrane protein of S. hyodysenteriae. This led us to identify the cloned genes as vspA and vspB (for variable surface protein) with the expectation that an additional homologous gene(s) will be identified which should accurately encode the purified 39-kDa protein. Nucleotide sequence data also suggest that both of these genes have regulatory elements which may suffice for expression within S. hyodysenteriae. We have found no homology between these gene sequences and other bacterial genes reported in the literature.
Multiple copies of highly conserved genes are unusual in bacteria. Relevant examples from the literature include a group of nine virulence genes (linkage unknown) within Haemophilus influenzae with tandem tetranucleotide repeats (6); three neighboring, related surface antigen ORFs in Methanosarcina mazei with large 42- and 85-amino-acid repeat units (13); and a pair of genes encoding similar sized, secreted proteases in Erwinia chrysanthemi (2). Repeated epitopes are likely to be needed for maintaining functional consistency between duplicated genes, while regions of variability may play a role in antigenic variation as a protective response (reviewed in reference 3). Examples of this include the multiple flagellin genes in Proteus mirabilis, which have regions of sequence divergence thought to be exposed to the environment and thus to contribute to antigenic variation and perhaps host immune system evasion (1). Alternatively, antigenic diversity could arise from cellular recombination events between tandemly linked genes as described for Campylobacter jejuni (25). Within S. hyodysenteriae there is evidence of a family of at least three related flagellar genes (flaB) (8, 26); however, these genes do not appear to be closely linked. Thus, this is the first report of tandemly linked homologous genes within this bacterial pathogen. We do not yet have evidence indicating whether low-level expression of the other vsp genes may occur in culture or during swine intestinal infection. The potential for functional and antigenic diversity represented in a multiple vsp gene family makes this family of proteins an interesting subject for further investigation.
Nucleotide sequence accession number.
The nucleotide sequence from the cloned S. hyodysenteriae DNA has been deposited in the GenBank library of DNA sequences under accession no. AF012102.
REFERENCES
- 1.Belas R, Flaherty D. Sequence and genetic analysis of flagellin-encoding genes from Proteus mirabilis. Gene. 1994;128:33–41. doi: 10.1016/0378-1119(94)90230-5. [DOI] [PubMed] [Google Scholar]
- 2.Boyd C, Keen N T. Characterization of the prtA and prtB genes of Erwinia chrysanthemi EC16. Gene. 1993;133:115–118. doi: 10.1016/0378-1119(93)90234-t. [DOI] [PubMed] [Google Scholar]
- 3.Brunham R C, Plummer F A, Stephens R S. Bacterial antigenic variation, host immune response, and pathogen-host coevolution. Infect Immun. 1993;61:2273–2276. doi: 10.1128/iai.61.6.2273-2276.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Gabe J D, Chang R-J, Slomiany R, Andrews W H, McCaman M T. Isolation of extracytoplasmic proteins from Serpulina hyodysenteriae B204 and molecular cloning of the flaB1 gene encoding a 38-kilodalton flagellar protein. Infect Immun. 1995;63:142–148. doi: 10.1128/iai.63.1.142-148.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Harris D L, Glock R D, Christensten C R, Kinyon J M. Swine dysentery. I. Inoculation of pigs with Treponema hyodysenteriae (new species) and reproduction of the disease. Vet Med Small Anim Clin. 1972;67:61–64. [PubMed] [Google Scholar]
- 6.Hood D W, Deadman M E, Jennings M P, Bisercic M, Fleischmann R D, Venter J C, Moxon E R. DNA repeats identify novel virulence genes in Haemophilus influenzae. Proc Natl Acad Sci USA. 1996;93:11121–11125. doi: 10.1073/pnas.93.20.11121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hyatt D R, ter Huurne A A H M, van der Zeijst B A M, Joens L A. Reduced virulence of Serpulina hyodysenteriae hemolysin-negative mutants in pigs and their potential to protect pigs against challenge with a virulent strain. Infect Immun. 1994;62:2244–2248. doi: 10.1128/iai.62.6.2244-2248.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Koopman M B H, Baats E, de Leeuw O S, Van der Zeijst B A M, Kusters J G. Molecular analysis of a flagellar core protein gene of Serpulina (Treponema) hyodysenteriae. J Gen Microbiol. 1993;139:1701–1706. doi: 10.1099/00221287-139-8-1701. [DOI] [PubMed] [Google Scholar]
- 9.Koopman M B H, de Leeuw O S, Van der Zeijst B A M, Kusters J G. Cloning and DNA sequence analysis of a Serpulina (Treponema) hyodysenteriae gene encoding a periplasmic flagellar sheath protein. Infect Immun. 1992;60:2920–2925. doi: 10.1128/iai.60.7.2920-2925.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Laemmli U K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature (London) 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- 11.Maniatis T, Fritsch E F, Sambrook J. Molecular cloning: a laboratory manual. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory; 1982. [Google Scholar]
- 12.Marchalonis J, Cone R E, Santer V. A probe for accessible surface proteins of normal and neoplastic lymphocytes. Biochem J. 1971;124:921–927. doi: 10.1042/bj1240921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Mayerhofer L E, Conway de Macario E, Macario A J L. Conservation and variability in Archaea: protein antigens with tandem repeats encoded by a cluster of genes with common motifs in Methanosarcina mazei S-6. Gene. 1995;165:87–91. doi: 10.1016/0378-1119(95)00524-a. [DOI] [PubMed] [Google Scholar]
- 14.Muir S, Koopman M B H, Libby S J, Joens L A, Heffron F, Kusters J G. Cloning and expression of a Serpula (Treponema) hyodysenteriae hemolysin gene. Infect Immun. 1992;60:529–535. doi: 10.1128/iai.60.2.529-535.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Rosey E L, Kennedy M J, Petrella D K, Ulrich R G, Yancey R J., Jr Inactivation of Serpulina hyodysenteriae flaA1 and flaB1 periplasmic flagellar genes by electroporation-mediated allelic exchange. J Bacteriol. 1995;177:5959–5970. doi: 10.1128/jb.177.20.5959-5970.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Rosey E L, Kennedy M J, Yancey R J., Jr Dual flaA1 flaB1 mutant of Serpulina hyodysenteriae expressing periplasmic flagella is severely attenuated in a murine model of swine dysentery. Infect Immun. 1996;64:4154–4162. doi: 10.1128/iai.64.10.4154-4162.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Sanger F, Nicklen S, Coulson A R. DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci USA. 1977;74:5463–5467. doi: 10.1073/pnas.74.12.5463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sellwood R, Kent K A, Burrows M R, Lysons R J, Bland A P. Antibodies to a common outer envelope antigen of Treponema hyodysenteriae with antibacterial activity. J Gen Microbiol. 1989;135:2249–2257. doi: 10.1099/00221287-135-8-2249. [DOI] [PubMed] [Google Scholar]
- 19.Taylor D J, Alexander T J L. The production of dysentery in swine by feeding cultures containing a spirochete. Br Vet J. 1971;127:58–61. doi: 10.1016/s0007-1935(17)37282-2. [DOI] [PubMed] [Google Scholar]
- 20.ter Huurne A A H M, Muir S, van Houten M, van der Zeijst B A M, Gaastra W, Kusters J G. Characterization of three putative Serpulina hyodysenteriae hemolysins. Microb Pathog. 1994;16:269–282. doi: 10.1006/mpat.1994.1028. [DOI] [PubMed] [Google Scholar]
- 21.ter Huurne A A H M, Gaastra W. Swine dysentery: more unknown than known. Vet Microbiol. 1995;46:347–360. doi: 10.1016/0378-1135(95)00049-g. [DOI] [PubMed] [Google Scholar]
- 22.Thomas W, Sellwood R. Molecular cloning, expression, and DNA sequence analysis of the gene that encodes the 16-kilodalton outer membrane lipoprotein of Serpulina hyodysenteriae. Infect Immun. 1993;61:1136–1140. doi: 10.1128/iai.61.3.1136-1140.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Von Heijne G. Patterns of amino acids near signal-sequence cleavage sites. Eur J Biochem. 1986;133:17–21. doi: 10.1111/j.1432-1033.1983.tb07424.x. [DOI] [PubMed] [Google Scholar]
- 24.Wannemuehler M J, Hubbard R D, Greer J M. Characterization of the major outer membrane antigens of Treponema hyodysenteriae. Infect Immun. 1988;56:3032–3039. doi: 10.1128/iai.56.12.3032-3039.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wassenaar T M, Fry B N, van der Zeijst B A M. Variation of the flagellin gene locus of Campylobacter jejuni by recombination and horizontal gene transfer. Microbiology. 1995;141:95–101. doi: 10.1099/00221287-141-1-95. [DOI] [PubMed] [Google Scholar]
- 26.Zuerner R L, Stanton T B. Physical and genetic map of the Serpulina hyodysenteriae B78T chromosome. J Bacteriol. 1994;176:1087–1092. doi: 10.1128/jb.176.4.1087-1092.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]