

Abstract
This study investigated the dietary effects of coated l-ascorbic acid (LA) on growth, feed utilization, survival, serum biochemical indices, immunity, antioxidant capacity, and intestinal and hepatopancreatic histology of the pre-adult red swamp crayfish. Four isoproteinous and isolipidic diets were formulated to contain several LA levels as 0, 1300, 1600, and 1900 mg/kg and designated as control (LA0), LA13, LA16, and LA19, respectively. However, the analyzed LA concentrations in diets were 0.00, 199.57, 360.45, and 487.50 mg/kg in LA0, LA13, LA16, and LA19, respectively. Triplicate treatments of crayfish (21.60 ± 0.14 g) were fed the test diets and reared in fiberglass tanks with a density of 20 individuals per each for eight weeks. Results revealed that all LA treatments had significantly enhanced growth performance compared to the control. Of interest, the LA16 treatment recorded the highest final tank biomass, biomass gain, total feed intake, condition factor, and muscle yield among the other treatments. The tank feed conversion ratio was significantly decreased in LA treatments compared to the control. Moreover, dietary LA16 and LA19 had significantly higher survival rates (93.3%) compared to (85.0%) in the LA0 group. All dietary doses of LA significantly increased serum parameters (total protein, albumin, globulin, lysozyme activity) and respiratory burst activity compared to the LA0 treatment. Dietary LA16 significantly boosted the hepatopancreatic antioxidant capacity, manifested by decreased malondialdehyde concentrations, increased catalase, superoxide dismutase, and glutathione peroxidase enzyme activities, and reduced glutathione content compared to the LA-free diet. A normal histoarchitecture of the hepatopancreatic tubules was found in all LA treatments except with some minor degenerative changes in the tubular lumen, and hepatopancreatic cells associated with enlarged nuclei were found in the LA19. However, normal intestinal histoarchitecture was found in all treatments with no recorded intestinal lesions. Of interest, the polynomial regression performed on the analyzed LA concentrations suggested that 380 mg/kg would be suitable to provide maximal biomass gain for pre-adult crayfish. In conclusion, results revealed that coated LA could enhance the growth, immunity, and antioxidant capacity of pre-adult red swamp crayfish, suggesting its potential as a functional and necessary micronutrient for crayfish diets.
Subject terms: Cell biology, Immunology, Physiology, Structural biology, Zoology
Introduction
Aquaculture has been greatly developed in many countries worldwide to provide human beings with nutritious proteinous food from diverse fish and crustacean species1–3. Nowadays and in the coming years, with the continuously elevated fish prices that are associated with world inflation, researchers have directed toward finding new and cost-effective fish or crustacean species to be farmed to find new proteinous sources for human beings4,5. Red swamp crayfish (Procambarus clarkii) is a freshwater crustacean that has a natural distribution in the south-central part of America and northern Mexico, but it has been introduced to several countries, where it may become an invasive species6. It was farmed in China and then distributed to other geographic areas. In the 1980s, it was introduced into Egypt as an exotic species, and it spread to the river Nile and its tributaries and lakes7. The red swamp crayfish is characterized by its delicious taste, high muscle protein, and other nutritional attributes that give this species a high market value in various countries across the globe8.
In aquaculture, the nutritional supplementation of vitamin C, particularly its traditional form, l-ascorbic acid (LA), has gained particular attention in the last decade because of its vital importance for farmed aquatic organisms9. Awareness was given to LA because of its well-known functionality and health benefits as a necessary micronutrient for improving the health status and physiological responses of many finfish and crustacean species10. Notably, it was reported that aquatic organisms differ in their LA requirements from livestock animals as animals can create LA from glucuronic acid, while finfish and crustaceans have a deficit of the l-gulonolactone oxidase enzyme required for LA biosynthesis11. This makes them depend mainly on a constant supply through their formulated diets12. On the other hand, LA is a water-soluble vitamin and sensitive to being degraded and losing its functionality (and bioactivity) during the processing and storage of the formulated aquafeed. Moreover, leaching losses may also occur in LA and most water-soluble vitamins13. Thus, researchers conducted several studies to find alternative LA forms to keep its activities in aquafeed, such as using coated LA14,15 or its derivatives10,16.
Generally, LA is necessary for all aquatic animals for growth, reproduction, wound healing, immunity, and disease resistance9. It is also required for the biosynthesis of collagen and several hormones and has an essential role in mitigating the toxic impacts of pollutants17. For red swamp crayfish, dietary LA is vital for growth, immunity, and reproductive performance18. In shrimps, it was found that dietary LA deficiency has led to poor growth and feed conversion, dropped molting frequencies, incomplete molting cycle, exoskeleton lesions, reduced stress tolerance, impaired disease resistance, and elevated mortalities14.
The nutritional requirement of LA differs among crustaceans. For example, it was shown that supplemental LA (100 mg/kg) promoted the growth, antioxidant capacity, and digestive enzyme activities of freshwater prawn (Macrobrachium malcolmsonii)19. However, 130 mg/kg of ascorbic acid 2-monophosphate was optimal for Whiteleg prawn (Penaeus vannamei) in the early post-larval stages of life20. Hence, its efficacy is also associated with the supplementation dose, forms, and life stages of aquatic animals9. Several factors should be considered to optimize the LA requirements for fish and crustaceans, such as differences in requirements among species21, size and developmental stage12,22, LA forms23, among several others. Hence, all these factors should be considered during the supplementation of LA in diets prepared for fish and crustaceans.
A few research studies were conducted to evaluate LA effects on red swamp crayfish. Kong et al.15 clarified that coated LA (265.67 mg/kg) was optimal for the maximum growth of P. clarkii juveniles (7.0 g body weight). Another study found that dietary l-ascorbyl-2-monophosphate (300.95 mg /kg) was optimal for P. clarkii juveniles (0.7 g body weight)16. This study aims to determine the impacts of supplemental LA levels on the performances of pre-adult P. clarkii. Thus, its effects on growth, survival, muscle composition, hemolymph biochemical indices, non-specific immune responses, oxidative stress biomarkers, and intestinal and hepatopancreatic histology were investigated.
Material and methods
Ethical consent
Experiments accomplished in the existing research have been licensed by the Local Experimental Animal Care Committee, Faculty of Agriculture, Alexandria University. Ethical Consent was obtained from the Institutional Animal Care and Use Committee at Alexandria University with Approval Code (AU-08/22/12/28/3/121). All methods in this study were performed in accordance with the relevant guidelines and regulations and ARRIVE guidelines (https://arriveguidelines.org).
Preparation of the experimental diets
Coated vitamin C (l-Ascorbic Acid; LA), with a purity of ≥ 99.0% crystalline powder, was commercially obtained from Sigma-Aldrich® agent (Egypt). Four isoproteinous and isolipidic diets were formulated, whereas the basal diet was supplemented with different LA levels as 0, 1300, 1600, and 1900 mg/kg and labeled as LA0 (control), LA13, LA16, and LA19, respectively. The raw dietary ingredients were thoroughly crushed into fine particles to prepare the test diets and then strained via a mesh sieve. The chosen LA levels were added to the diets at the expense of microcrystalline cellulose. The LA levels were mixed with the raw diet components. A suitable amount of water and oil was added to each kg feed during the mixing procedures to moisten the diets and get a dough. The generated dough was then passed across a meat mincer to form pelletized diets (2 mm diameter). The formulated test diets were dried in the air, preserved in Ziplock bags, and kept in a refrigerator (− 20 °C) until utilized. The LA concentrations in the formulated LA-supplemented diets were analyzed directly by the high-performance liquid chromatography (HPLC) method, as explained by Shiau and Hsu24 and Wang et al.25. The analyzed LA concentrations of the test diets were 0.00, 199.57, 360.45, and 487.50 mg/kg in the LA0, LA13, LA16, and LA19 diets, respectively.
Chemical (nutrient) compositions of the test feeds
Information about the feed formulations, ingredients, and chemical compositions of the test feeds is given in Table 1. The analysis was conducted in line with the Association of Analytical Chemists (AOAC) procedures26. Crude protein (CP; %) was examined using Kjeldahl’s method for nitrogen determination. Crude fiber (CF; %) was evaluated according to the Weende method, which is based on the digestion of non-cellulosic compounds by sulfuric acid and potassium hydroxide. Ash (%) levels were assessed after the sample burning at 550 °C in a muffle furnace for one day. Moisture (%) content was assessed after the sample drying to a constant weight at 105 °C. Crude lipid (CL; %) content was evaluated by ether extraction using a Soxtec apparatus System.
Table 1.
Ingredients and proximate chemical composition (%) of the experimental diets supplied with various l-ascorbic acid (LA) levels.
| Feed ingredients | Diets with different LA levels | |||
|---|---|---|---|---|
| LA0 (Control) | LA13 | LA16 | LA19 | |
| Fish meal (FM)⁕ | 10.0 | 10.0 | 10.0 | 10.0 |
| Soybean meal (SBM)# | 35.0 | 35.0 | 35.0 | 35.0 |
| Corn gluten meal (60% CP) | 4.8 | 4.8 | 4.8 | 4.8 |
| Rice bran (14% CP) | 11.50 | 11.5 | 11.5 | 11.5 |
| Yellow corn meal | 14.5 | 14.5 | 14.5 | 14.5 |
| Wheat bran | 9.0 | 9.0 | 9.0 | 9.0 |
| Wheat flour | 12.0 | 12.0 | 12.0 | 12.0 |
| Sunflower oil | 0.70 | 0.70 | 0.70 | 0.70 |
| Microcrystalline cellulose | 1.70 | 1.57 | 1.54 | 1.51 |
| l-ascorbic acida | 0.0 | 0.13 | 0.16 | 0.19 |
| Vitamin and mineral premix b | 0.30 | 0.30 | 0.30 | 0.30 |
| Dicalcium phosphateb | 0.50 | 0.50 | 0.50 | 0.50 |
| Total | 100 | 100 | 100 | 100 |
| Proximate chemical analysis (% on DM basis) | ||||
| Dry matter (DM) | 89.64 | 89.65 | 89.65 | 89.65 |
| Crude protein (CP) | 30.00 | 30.01 | 30.02 | 30.02 |
| Crude lipids (CL) | 5.8 | 5.8 | 5.8 | 5.8 |
| Ash | 5.91 | 5.92 | 5.92 | 5.92 |
| Crude fiber (CF) | 5.31 | 5.31 | 5.31 | 5.31 |
| Nitrogen free extract (NFE)c | 52.98 | 52.95 | 52.95 | 52.94 |
| Analyzed LA (mg/kg) | 0.00 | 199.57 | 360.45 | 487.50 |
*Danish FM (72% CP), obtained from TripleNine Fish Protein, DK-6700 (Denmark).
#Egyptian soybean flour (44.8% CP), obtained from Cargill Trading Egypt Co. (Egypt).
al-ascorbic acid (LA), with a purity of ≥ 99.0%, crystalline powder, purchased from Sigma-Aldrich® agent (Egypt).
bPurchased from AGRI-VET for Manufacturing Vitamins and Feed additives (10th of Ramadan City A2, Egypt). Vitamin and Mineral composition (per kg premix mixture): Vitamin premix including 700 mg (vitamin B1); 3500 mg (vitamin B2); 1000 mg (vitamin B6); 7 mg (vitamin B12); 8,000,000 IU (vitamin A); 2,000,000 IU (vitamin D3); 7000 mg (vitamin E); 1500 mg (vitamin K3); 50 mg (biotin); 700 mg (folic acid); 20,000 mg (nicotinic acid), and 7000 mg (pantothenic acid). Mineral premix including 40 g (Zinc); 20 g (Iron); 2.7 g (Copper); 0.34 g (Iodine); 53 g (Manganese); 70 mg (Selenium); 70 mg (Cobalt) and Calcium carbonate as carrier up to 1 kg.
cNFE = 100 − (CP + CL + CF + Ash).
Experimental animals: Collection, acclimation, and rearing conditions
Pre-adult red swamp crayfish were collected from fertilized ponds at WorldFish Center, Abbassa, Egypt. Animals were transferred into the rearing tanks and then left for acclimatization to the new environments. Crayfish were fed a pelleted basal diet (LA0; control diet without supplements) during a 2-week adaptation period. Following the acclimatization, 240 crayfish (ca 21 g in weight and 6.5 cm initial length) were bulk weighed in groups of 20 and transferred to fiberglass tanks (120 cm × 80 cm; water depth 30 cm) to create triplicates for each feed treatment (LA0, LA13, LA16, and LA19). Crayfish were assembled, bulk-weighed, and allotted into an indoor system in the Wet Laboratory belonging to the WorldFish Center, Abbassa, Egypt. Each tank was supported with 4 PLA water pipes to act as a shelter for raising crayfish and to prevent cannibalism and predation. Animals were deprived of feeding on the basal diet for one day (24 h) before starting to feed on the test diets. During the trial, crayfish were hand-fed on the corresponding formerly prepared test diets to apparent satiety three times daily (08.00 a.m., 12.00 a.m., and 3.00 p.m.) for eight weeks (56 days). The quantity of feed ingested per tank was recorded daily. Moreover, the leftover meal was garnered by siphoning 2 h post-feeding and then oven-dried at 60 °C to accurately evaluate the tank feed intake. During the feeding trial, the water in each tank was kept at a depth of 30 cm with constant aeration to preserve the water quality adequate for rearing crayfish. Crayfish in all rearing tanks were examined daily throughout the feeding period, and dead animals were counted, recorded, and then discarded using complete hygienic disposal methods. Siphoning was conducted every two days, and water renewal was performed by exchanging 1/3 of the water volume and replacing it with new, well-aerated, de-chlorinated tap water. The siphoning was performed to remove feces and newly molted shells before offering a new feed. The water quality indices were monitored weekly, and their average values were sustained at 26.50 ± 0.5 °C for temperature, 6.5 ± 0.03 for pH, 7.5 ± 1.0 mg/L for dissolved oxygen, and 0.067 ± 0.033 mg/L for ammonia-N. The lighting regime was an 8 h light to 16 h dark cycle.
Sampling procedures
Animals in all treatments were starved for 24 h after the feeding trial ended. Before the sampling procedures, crayfish were subjected to hypothermic anesthesia via compression in an ice tank for 15 min27.
Hemolymph sampling
The hemolymph samples were obtained directly after the piercing of the heart of the crayfish. Three hemolymph samples (n = 3) were taken from each treatment. In this step, an equal volume of hemolymph from three crayfish per tank was pooled, homogenously mixed, and considered one sample. Two sets of hemolymph samples were collected using a 1-mL sterile syringe. The first set was collected and pooled from 3 crayfish per tank and was drawn into a sterile Eppendorf tube containing the same volume of the cooled anticoagulant. This sample was used to evaluate the respiratory burst activity (RBA) of the hemocytes. The second set was collected and pooled from another 3 crayfish per tank and was drawn without anticoagulant into a sterile Eppendorf tube and used for serum separation. To collect the serum, hemolymph samples were left on ice for 6 h in a standing position to separate the supernatant. The supernatant samples were then separated after centrifugation at 3000×g for 10 min at 4 °C. These samples were collected, pooled, refrigerated, and then stored at − 20 °C until being used to evaluate serum biochemical assays.
Tissue sampling
Following the hemolymph sampling procedure, the bloodless crayfish from each tank were placed on ice and then aseptically dissected to get different tissues (hepatopancreas, tail muscle, and intestines). Samples obtained from the tail muscle (n = 3) were weighed to evaluate the muscle yield (%). Further, the tail muscle samples were cryopreserved and held at − 20 °C to assess the proximate chemical composition analysis. The hepatopancreas samples (n = 3) were held at − 20 °C to be frozen before being used to prepare homogenate samples. The homogenate samples were prepared from 100 mg of the thawed hepatopancreas sample, homogenized well in sterile, cold physiological buffer saline (PBS) (at a volume ratio of 1:9) by a tissue homogenizer, and then centrifuged (5000×g for 10 min at 4 °C). After centrifugation, sediment was discarded, and the supernatant was collected and used to evaluate the oxidative stress biomarkers. Other hepatopancreas (n = 3) and intestinal samples (n = 3) were washed in sterile PBS and then rapidly fixed in a neutral and freshly prepared 10% formalin solution for histological analysis.
Evaluation of growth, feed utilization, morphology indices, and survival %
After the end of the feeding duration (56 days), crayfish were deprived of feeding on LA-supplemented diets for 24 h as a period of starvation. This step is essential to remove any ingested food from the gastrointestinal tract (GIT) of crayfish, according to Davis and Robinson28. Crayfish from each tank were collected into plastic buckets, counted (to know their final number), and then bulk weighed. The total final biomass, biomass gain, and total feed intake have been estimated. The tank feed conversion ratio was also evaluated as the total tank feed intake split by the tank biomass gain.
To calculate the morphology indices such as muscle yield (%) and condition factor (CF, g/cm3), three crayfish were taken from those present in each tank. Then, individual final body length (cm), individual final weight (g), and individual final muscle weight (g) were evaluated. Muscle yield and CF were assessed as follows:
To calculate the crayfish survival rate (SR; %), crayfish per tank were totaled and counted. Then, SR (%) was evaluated according to the following equation:
Proximate chemical composition of crayfish muscles
The proximate chemical composition of the abdominal muscles of the crayfish from different treatments was conducted in line with the AOAC procedures26, whereas CP (%), ash (%), moisture (%), and CL (%) were evaluated as previously described in "Chemical (nutrient) compositions of the test feeds" section.
Serum biochemical indices
Serum biochemical indices such as alanine aminotransferase (ALT; U/L), aspartate aminotransferase (AST; U/L), total protein (TP; g/dL), albumin (ALB; g/dL), and globulin (GLO; g/dL) were assayed using commercially purchased diagnostic kits (Spinreact, S.A., Gerona, Spain). Serum ALT and AST enzyme activities were measured in line with the method illustrated by Reitman and Frankel29. Hemolymph TP concentrations were assessed per the methodology explained by Bradford30, with bovine serum ALB as the standard. Hemolymph ALB was estimated in accordance with the method depicted by Doumas et al.31. Hemolymph GLO levels were calculated by subtracting ALB from TP content.
Non-specific immune assays
Serum lysozyme (LZM) activities were assayed by the turbidimetric method, as illustrated by Ellis32. Serum LYZ activity was assayed using Micrococcus lysodeikticus (Sigma-Aldrich, USA) suspension. One LYZ unit was outlined as the volume of the serum sample causing a reduction in the absorbance of 0.001 per min at 530 nm. The hemocyte respiratory burst activity (RBA) in the hemolymph samples was evaluated by using the nitro blue tetrazolium (NBT) test as a method used to determine superoxide anion production. RBA was then quantified via a spectrophotometer at OD = 630 nm as per the protocol illustrated by Song and Hsieh33.
Oxidative stress biomarkers
The supernatant resultant from centrifugation of the hepatopancreatic homogenate was used for evaluation of the enzymatic antioxidant mechanisms by measuring the superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx) enzyme activities. Moreover, the reduced glutathione (GSH) and malondialdehyde (MDA) concentrations were also assayed. The diagnostic kits used for the evaluation of CAT and SOD enzyme activities were commercially purchased from MyBioSource Inc. (San Diego, California, USA) as per the directions provided by the producer and following the methods labeled by Aebi34 and McCord and Fridovich35. The diagnostic kits used for evaluating GPx activity and MDA and reduced GSH concentrations were commercially purchased from Biodiagnostic Co. (Giza, Egypt) as per the directions provided by the manufacturer. Malondialdehyde (MDA) concentrations were evaluated (as lipid peroxidation end product) in the hepatopancreatic tissues at an OD of 532 nm by thiobarbituric acid substances (TBARS) method as per the assay previously explained by Ohkawa et al.36 and Ke et al.37. All parameters were measured in triplicates. The enzyme assays were defined as U/ mg protein. Reduced GSH content was described as µmol/mg protein. MDA concentrations were expressed as nmol/mg protein.
Histological procedures
The intestines and hepatopancreatic tissues of crayfish samples were immersed in 10% buffered formalin solution for 1–2 days, dehydrated in a series of graded ethyl alcohol (75%, 85%, 90%, 95%, 100%), cleared in xylene, and then embedded in paraffin to make paraffin blocks38. Ten cross-sections (5 μm thickness) were taken from each sample by using a rotary microtome and then stained with H&E. The histological assessment was performed on several photomicrographs captured of the histological sections using a digital camera (Leica EC3, Germany) joined with a light microscope (Leica DM500). Ten microscopic fields are randomly selected for each sample for evaluation. The shape and size of the hepatopancreatic epithelial cells, tubular lumen, and nuclei were examined. The shape and arrangement of the intestinal epithelial cells, intestinal lumen, and intestinal layers (epithelium, lamina propria, submucosa, and muscularis layer) were examined. The abnormal histological structure and lesions were judged by a professional histopathologist for qualitative evaluation of the results. Abnormal hepatopancreatic structure (epithelial vacuolation, degenerated cells, degenerated tubules, and enlarged nucleus) and intestinal inflammatory signs were recorded.
Statistical analyses
A one-way ANOVA was used to determine the dietary effects of supplemental LA levels on the measured parameters of crayfish. Data were expressed as means ± SE. Differences between means were examined at a 5% probability level using Duncan’s test as a post hoc test. Moreover, P < 0.05 was considered a statistically significant difference. Linear and quadratic regression analyses were conducted on the crayfish's growth, morphology indices, survival, and muscle chemical composition to assess the relationship between dietary LA levels and the measured parameters. Best fitted Cubic regression using a polynomial model was performed against the analyzed LA levels to determine the optimum level for maximum biomass gain, as clarified by Zeitoun et al.39.
Results
Effects of LA on growth, feed utilization, and morphology indices
The growth, feed utilization, and morphology indices of crayfish-fed diets supplemented with various LA levels for eight weeks are presented in Table 2. The final tank biomass, biomass gain, and total feed intake were linearly and quadratically increased (P < 0.05) in LA treatments compared to those fed the LA-free diet. Moreover, variations within different treatments showed that their highest levels were noticed in the LA16. The tank FCR significantly decreased in all LA treatments compared to the controls, and LA treatments showed non-significant differences. The morphology indices (CF and muscle yield) and SR (%) were linearly and quadratically increased in LA treatments compared to LA0. Of interest, the muscle yield and CF peaked in the LA16 compared to the other treatments. Furthermore, the highest SR (%) was found in the LA16 and LA19 treatments, with non-significant differences. The polynomial regression analysis of biomass gain and the analyzed LA concentrations suggests that dietary LA of 380 mg/kg would be optimum for maximal biomass gain of pre-adult crayfish (Fig. 1).
Table 2.
Growth performance, feed utilization, and morphology indices of red swamp crayfish fed diets supplemented with various levels of l-ascorbic acid for 56 days.
| Parameters | Experimental groups | PSEa | P value | ||||
|---|---|---|---|---|---|---|---|
| LA0 | LA13 | LA16 | LA19 | Linear | Quadratic | ||
| Initial biomass (g/tank) | 433.3 | 432.0 | 434.6 | 434.7 | 2.84 | 0.881 | 0.960 |
| Final biomass (g/tank) | 674.9 c | 831.6 b | 931.5 a | 839.2 b | 29.90 | 0.001 | 0.003 |
| Biomass gain (g/tank) | 241.6 c | 399.6 b | 496.8 a | 404.5 b | 30.25 | 0.002 | 0.004 |
| Total feed intake (g feed/tank) | 533.8 c | 789.8 b | 972.2 a | 799.0 b | 50.14 | 0.002 | 0.003 |
| Tank feed conversion ratio (FCR) | 2.21 a | 1.98 b | 1.96 b | 1.98 b | 0.041 | 0.008 | 0.021 |
| Condition factor (CF; g/cm3) | 2.24 c | 2.81 b | 4.04 a | 3.12 ab | 0.260 | 0.041 | 0.058 |
| Muscle yield (%) | 9.8 b | 11.4 ab | 14.1 a | 11.1 ab | 0.724 | 0.022 | 0.056 |
| Survival rate (SR; %) | 85.0 c | 91.7 b | 93.3 a | 93.3 a | 1.35 | 0.004 | 0.020 |
The values of Means that not sharing a common letter in the same row are significantly statistically different at P < 0.05.
aPooled standard error of the means.
Figure 1.
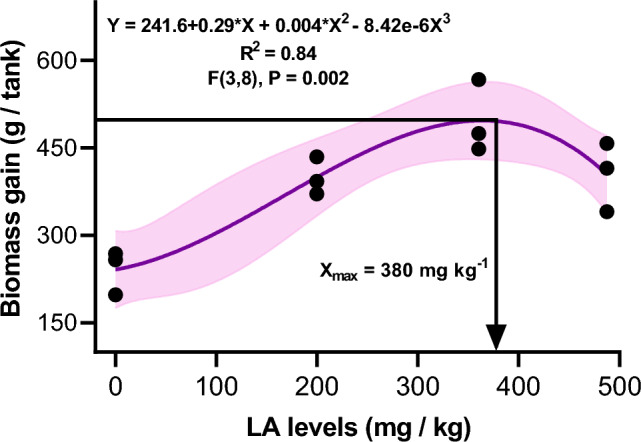
The relationship biomass gain (g/tank) of red swamp crayfish and the analyzed l-ascorbic acid (LA) levels in diets, as described by polynomial regression. Data were expressed as means ± S.E. (n = 3 replicates).
Effects of LA on proximate composition of crayfish muscles
The effects of dietary LA on the chemical analysis of the crayfish muscles are presented in Table 3. The CP content in the crayfish muscles was linearly and quadratically increased and was significantly elevated in all LA treatments compared with those fed on the LA-free diet (Table 3); however, the CP content in all LA treatments showed non-significant differences between their values (P > 0.05). On the other hand, moisture (%) and CL (%) were decreased linearly and quadratically alongside the increasing dietary LA supplementation. Of interest, the CL (%) was not significantly affected among LA13, LA16, and LA19 treatments (P > 0.05). However, ash (%) was linearly and quadratically elevated in LA16 and LA19 treatments compared to other treatments.
Table 3.
Proximate chemical composition (% on fresh weight basis) of the muscles of red swamp crayfish fed diets supplemented with various levels of l-ascorbic acid for 56 days.
| Parameters | Dietary l-ascorbic acid levels (% feed) | PSEa | P value | ||||
|---|---|---|---|---|---|---|---|
| LA0 | LA13 | LA16 | LA19 | Linear | Quadratic | ||
| Moisture (%) | 73.2 a | 72.4 ab | 71.7 b | 71.4 b | 0.286 | 0.019 | 0.072 |
| Crude protein (CP; %) | 13.9 b | 15.7 a | 15.9 a | 15.9 a | 0.290 | < 0.001 | 0.002 |
| Crude lipids (CL; %) | 7.5 a | 6.1 b | 6.2 b | 6.10 b | 0.187 | < 0.001 | < 0.001 |
| Ash (%) | 1.2 b | 1.2 b | 1.4 a | 1.50 a | 0.038 | 0.005 | 0.001 |
The values of Means that not sharing a common letter in the same row are significantly statistically different at P < 0.05.
aPooled standard error of the means.
Effects of LA on serum biochemical indices
Serum biochemical indices of crayfish fed on LA-supplied diets for 56 days are elucidated in Fig. 2. Compared with the LA0, the TP (Fig. 2A), ALB (Fig. 2B), and GLO (Fig. 2C) contents were significantly increased in all LA treatments. Moreover, their uppermost levels were found in the LA16 compared with the other treatments. Conversely, ALT (Fig. 2D) and AST (Fig. 2E) enzyme activities were significantly decreased in all LA treatments with increasing dietary LA levels in a dose-dependent trend compared to those fed on an LA-free diet.
Figure 2.

The serum biochemical indices include total protein (g/dL; A), albumin (g/dL; B), globulin (g/dL; C), ALT (U/L; D), and AST (U/L; E) in red swamp crayfish fed diets supplemented with various levels of l-ascorbic acid (LA) for 56 days (n = 3). Data were expressed as means ± SE. Bars labeled with different letters are statistically significantly different at P < 0.05.
Effects of LA on non-specific immunity
Dietary LA significantly increased serum LYZ (Fig. 3A) and RBA activities in the hemolymph (Fig. 3B) of crayfish compared with those reared in LA0, and their highest levels were found in LA16. However, their levels showed non-significant differences in the LA13 and LA19 treatments.
Figure 3.

The immune indices include serum lysozyme activity (A) and respiratory burst activity (RBA; B) of red swamp crayfish fed diets supplemented with various levels of l-ascorbic acid (LA) for 56 days (n = 3). Data were expressed as means ± SE. Bars labeled with different letters are statistically significantly different at P < 0.05.
Effects of LA on hepatopancreatic oxidative stress biomarkers
The impacts of LA supplementation on the hepatopancreatic MDA concentrations and antioxidant capacity of crayfish are presented in Fig. 4. The enzymatic antioxidant mechanisms, including CAT (Fig. 4A), SOD (Fig. 4B), and GPx (Fig. 4C) enzyme activities as well as the reduced GSH contents (Fig. 4D) of crayfish fed on LA-supplemented diets were significantly increased in response to the dietary LA levels. Furthermore, the highest CAT, SOD, GPx, and GSH levels were found in the LA16. In contrast, their lowest levels were found in LA0 (Fig. 4). The hepatopancreatic MDA concentrations were decreased significantly with regard to dietary LA supplementation levels. Their highest values were found in crayfish fed the LA-free diet (Fig. 4E).
Figure 4.

The oxidative stress biomarkers, including CAT (A), SOD (B), GPx (C), reduced GSH content (D), and MDA concentration (E) in the hepatopancreatic tissues of red swamp crayfish fed diets supplemented with various levels of l-ascorbic acid (LA) for 56 days (n = 3). Data were expressed as means ± SE. Bars labeled with different letters are statistically significantly different at P < 0.05.
Effects of dietary LA on hepatopancreatic histology
The impacts of supplemental LA on the hepatopancreatic histology of crayfish in comparison to those reared in control are shown in Fig. 5. The hepatopancreas exhibited well-organized glandular structures and tubular lumens in the LA0, LA13, and LA16 treatments with tightly arranged and intact hepatopancreatic tubules. Moreover, the hepatopancreatic cells (HC) were easily recognized and uniform in shape and size. However, the hepatopancreatic structures in LA19 were characterized by prominent epithelial vacuolation, mildly degenerated hepatopancreatic cells, degenerated hepatopancreatic tubules, lumen dilatation, and an enlarged hepatopancreatic nucleus.
Figure 5.

Representative photomicrographs of the hepatopancreatic tubules of red swamp crayfish fed diets supplemented with various levels of l-ascorbic acid (LA) for 56 days. Tissue sections were stained with H & E stain (X 10), scale bar = 20 µm. Hepatic cells (HC), Hemal space (HS), and Tubule lumen (TL).
Effects of LA on intestinal histology
The intestines of crayfish fed on diets supplied with LA levels in comparison to those reared in the LA0 are shown in Fig. 6. The intestines of crayfish in the LA0, LA13, LA16, and LA19 treatments displayed typical, orderly-organized, and tightly arranged structures of the intestinal lumen, epithelium, lamina propria, submucosa, and muscularis layer with no inflammatory signs recorded.
Figure 6.

Representative photomicrographs of the intestines of red swamp crayfish fed diets supplemented with various levels of l-ascorbic acid (LA) for 56 days. Tissue sections were stained with H & E stain and observed under light microscopy with (X 10), scale bar = 20 µm. IL: Intestinal lumen; E: Epithelium; LP: Lamina propria; Sm: Submucosa layer; M: muscularis layer.
Discussion
The beneficial effects of vitamin C (l-ascorbic acid; LA) as a dietary supplement have been proven in aquaculture, with promising results on the health of fish and crustaceans9. Herein, LA effects have been investigated on growth, muscle composition, morphology indices, immunity, antioxidant capacity, and intestinal and hepatopancreatic histology of pre-adult P. clarkii. The positive effects of LA on growth have been described in various crustacean species, such as the signal crayfish (Pacifastacus leniusculus) juveniles23, Gazami crab (Portunus trituberculatus) juveniles40, whiteleg shrimp (Litopenaeus vannamei)41, giant river prawn (Macrobrachium rosenbergii) juveniles42,43, M. malcolmsonii19, and also in P. clarkii juveniles15. We found linear and quadratic increases in the growth parameters, muscle yield, CF, and SR in all LA-supplemented treatments compared to the controls, and their highest levels were found in the LA16. Vitamin C, a vital micronutrient, is required for enhancing the growth of crustaceans12, and its deficiency has led to poor feed conversion, reduced growth, inadequate molting, and high mortality rates14,24. The improvement of crayfish survival in the present study could be associated with the positive roles of LA in improving immunity, stress tolerance, and disease resistance9. In the present study, the tank FCR decreased significantly in all LA treatments compared to the control. These results were in concordance with those obtained by Kong et al.15, who found that dietary LA significantly decreased the FCR alongside increased LA supplementation in red swamp crayfish diets. The optimum dietary LA level in the present study was 380 mg/kg for maximum biomass gain of pre-adult crayfish, as described by polynomial regression. Celada et al.23 declared that a 200 mg/kg diet of l-ascorbyl-2-monophosphate-Na is recommended for P. leniusculus juveniles. Kong et al.15 reported that a 265.67 mg/kg diet is an optimal LA supplementation dose for P. clarkii juveniles. Kong et al.16 further observed that a 300.95 mg/kg diet is an optimal supplementation dose of l-ascorbyl-2-monophosphate for red swamp crayfish.
Herein, dietary LA significantly increased the CP % in the crayfish muscles in all LA-supplemented treatments compared with those fed on the LA-free diet. The enhancement of CP% in the crayfish muscles may be associated with the dietary roles of LA in the improvement of protein synthesis and amino acid storage, as previously described in the muscles of freshwater prawns19. Reports showed that LA could also increase the CP % in the body of P. clarkii juveniles15,16. However, our results showed that LA-supplementation decreased CL (%) in the crayfish muscles compared to controls. These results differ from those described in P. trituberculatus juveniles40, M. malcolmsonii19, and P. clarkii juveniles15, as these studies reported that dietary LA increased the lipid % in the body of these crustacean species. Indeed, the differences may be linked to several factors, such as LA factors (doses, sources, forms), crayfish factors (crayfish species, initial weights, sizes), experimental conditions, duration of the feeding trial, or others.
Hematology and hemolymph biochemical indices represent crucial bio-indicators for evaluating the health and nutritional state of invertebrates44. Total protein (TP) is an important parameter for determining the functional status of crayfish45. Moreover, it was previously reported that higher TP, GLO, and ALB contents indicate a vital immuno-competent capacity27,46. Results revealed that the TP, ALB, and GLO content significantly elevated with increasing dietary LA levels compared to the controls, indicating that the LA could boost the body immunity of the treated crayfish. ALT and AST are transaminases that contribute to protein metabolism and are sensitive markers for examining hepatopancreatic functions27,47. In aquatic organisms, it was found that increased AST and ALT enzyme activities denote dysfunction, necrosis, malfunction, tissue degeneration, and change of protein metabolism in the hepatopancreas48,49. Serum ALT and AST activity levels had an inverse relationship with dietary LA, suggesting that high LA in crayfish diets may induce hepatopancreatic injury.
Lysozyme (LZM) is a critical enzyme that exerts antibacterial activities due to its lytic effects on bacterial cell walls32,50. It also has a crucial immune function in red swamp crayfish51. Respiratory burst activity (RBA) is an essential process that helps hemocytes to get rid of the foreign bacteria from the crayfish hemolymph by phagocytosis by generating reactive oxygen species33. Hence, RBA can be regarded as an essential bioindicator to reflect the innate immunity of crustaceans. Herein, it was observed that dietary LA boosted LYZ activity and RBA compared with those reared in the LA0. These results, thereby, reflect the roles of LA in enhancing the immune defense of crustaceans, as previously reported in the giant tiger prawn (Penaeus monodon)52 and red swamp crayfish15. In finfish species, reports also showed that dietary LA could increase LYZ and RBA activity53,54.
In aquatic organisms, SOD, CAT, GPx, and reduced GSH are effective antioxidant enzymatic defense mechanisms against free radicals, and they can protect organisms against oxidative stress damage55,56. MDA is regarded as the end-product of lipid peroxidation57 and an essential marker for assessing oxidative damage58. In the present study, CAT, SOD, GPx, and reduced GSH content in the hepatopancreatic tissues of crayfish fed on LA-supplied diets were increased in response to the dietary LA supplemental levels. On the other hand, hepatopancreatic MDA concentrations were decreased with regard to dietary LA supplementation levels. These results suggest the capability of dietary LA to boost the antioxidant capacity of the treated crayfish, reflecting the potent antioxidant properties of dietary LA59. Hence, LA can be considered an effective antioxidant and potent free radical scavenger. Previous research has proved the roles of dietary LA in improving the antioxidant capacity of many crustacean species, such as L. vannamei60, M. malcolmsonii19, and red swamp crayfish15. Similar findings were also reported in several finfish species61,62.
Histological analysis is a practical tool for evaluating the nutritional status and health conditions of finfish and crustaceans63,64. The present study showed that hepatopancreatic tissues exhibited well-organized glandular structures and tubular lumens in the LA0, LA13, and LA16 treatments with tightly arranged and intact hepatopancreatic tubules. In addition, the hepatopancreatic cells were easily recognized and reasonably uniform in shape and size. This means that dietary LA up to 1600 mg/kg level could be safe and optimum for the typical structures of the hepatopancreatic tissues. On the other hand, LA 1900 mg/kg resulted in prominent epithelial vacuolation, mildly degenerated hepatopancreatic cells, degenerated hepatopancreatic tubules, lumen dilatation, and enlarged hepatopancreatic nucleus. This result was also confirmed by the ALT and AST enzyme activity levels in the present study and their relationship with dietary LA levels. However, the intestines of crayfish in all treatments displayed typical, orderly-organized, and tightly arranged structures of the intestinal lumen, epithelium, lamina propria, submucosa, and muscularis layer, with no inflammatory signs recorded. This indicates that dietary LA did not negatively affect the histology of the crayfish intestines. To the best of our knowledge, no previous results were published on the dietary effects of LA on the histology of red swamp crayfish. Nonetheless, it was reported that dietary supplementation with a 100 mg/kg diet of magnesium l-ascorbyl-2-phosphate improved the histological structure of the hepatopancreas of P. monodon juveniles65. In finfish, Ibrahim et al.62 found that dietary LA improved Nile tilapia's hepatopancreatic histoarchitecture and intestinal histomorphology. Moreover, Yusuf et al.66 declared that dietary LA maintained the integrity of the liver morphology of largemouth bass. The positive effects obtained in the present study may be linked with the antioxidant effects of LA, which help to protect the tissues of the treated crayfish from the adverse effects of free radicals.
Conclusions and future perspectives
The present study's findings revealed that supplemental LA significantly improved growth, feed utilization, survival, enhanced immunity, and hepatopancreatic antioxidant capacity and maintained the normal guts of pre-adult red swamp crayfish. Besides, the treatment fed a diet with 1600 mg/kg LA induced the best non-specific immunity and antioxidant capacity. The polynomial regression analysis revealed that 380 mg/kg of the analyzed LA was optimal for the maximum biomass gain of pre-adult red swamp crayfish. To put it briefly, LA could be considered a crucial micronutrient for enhancing the aquafeed formulated for the crayfish industry. In a theoretical laboratory-based experiment, the aforementioned findings present reference data for improving the ration formulation and health status of crayfish culture. However, the impacts of dietary LA on disease resistance and intestinal microbiome at molecular and nutrigenomic levels warrant further investigations and research studies.
Acknowledgements
All authors gratefully acknowledge the hard work supported by Staff members of WorldFish Center, Egypt, for their continuous help and support while running the present study.
Author contributions
H.S.K.: data curation, methodology, investigation, writing—review and editing. H.O.A., N.E.: conceptualization, methodology. M.F.E.B.: investigation, methodology, conceptualization, validation. A.M.E.-N.: methodology, validation, investigation. A.A.A.H.: investigation, conceptualization. A.A.K.: validation, conceptualization, investigation. A.M.A.R.: formal analysis, writing. M.K.: validation, investigation. D.A.-K.: conceptualization, investigation. M.A.-T.: formal analysis, writing—review and editing. H.M.R.A.-L.: data curation, writing—original draft, writing—review and editing.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB).
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Hala S. Khalil, Email: halasaber2011@yahoo.com
Hany M. R. Abdel-Latif, Email: hmhany@alexu.edu.eg
References
- 1.Ali A, et al. Morphometric, histochemical, and ultrastructural analysis of the reproductive system and spermatogenic stages of male blue crab (Callinectessapidus rathbun, 1896) J. Mar. Sci. Eng. 2021;9:1105. doi: 10.3390/jmse9101105. [DOI] [Google Scholar]
- 2.Maulu S, Nawanzi K, Abdel-Tawwab M, Khalil HS. Fish nutritional value as an approach to children's nutrition. Front. Nutr. 2021 doi: 10.3389/fnut.2021.780844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Alhoshy M, et al. Nutrigenomics in crustaceans: Current status and future prospects. Fish Shellfish Immunol. 2022;129:1–12. doi: 10.1016/j.fsi.2022.08.056. [DOI] [PubMed] [Google Scholar]
- 4.Adam AH, et al. Effect of dietary bile acids: Growth performance, immune response, genes expression of fatty acid metabolism, intestinal, and liver morphology of striped catfish (Pangasianodon hypophthalmus) Aquac. Rep. 2023;29:101510. doi: 10.1016/j.aqrep.2023.101510. [DOI] [Google Scholar]
- 5.El-Ouny YM, et al. Effect of fishmeal replacement with dried red wigglers (Eisenia fetida) worm meal on growth and feed utilization, production efficiency, and serum biochemistry in Nile tilapia (Oreochromis niloticus) fingerlings. Aquac. Rep. 2023;29:101518. doi: 10.1016/j.aqrep.2023.101518. [DOI] [Google Scholar]
- 6.Gherardi F. Crayfish invading Europe: The case study of Procambarus clarkii. Mar. Freshw. Behav. Physiol. 2006;39:175–191. doi: 10.1080/10236240600869702. [DOI] [Google Scholar]
- 7.Ibrahim AM, Khalil MT, Mubark M. Ecological studies on the exotic crayfishes Procambarus clarkii and Procambarus zonangulus in the River Nile. Int. J. Ecol. Environ. Sci. 1997;23:217–228. [Google Scholar]
- 8.Hamdi SA, El-Monem S. Processing, products and marketing of the red swamp crawfish Procambarus clarkii (Crustacea, Decapoda) Egypt. J. Exp. Biol. (Zool.) 2006;2:93–98. [Google Scholar]
- 9.Dawood MAO, Koshio S. Vitamin C supplementation to optimize growth, health and stress resistance in aquatic animals. Rev. Aquac. 2018;10:334–350. doi: 10.1111/raq.12163. [DOI] [Google Scholar]
- 10.Shiau, S.-Y. In Ascorbic Acid In Aquatic Organisms 87–95 (2000).
- 11.Dabrowski K. Ascorbic acid status in the early life of whitefish (Coregonus lavaretus L.) Aquaculture. 1990;84:61–70. doi: 10.1016/0044-8486(90)90300-C. [DOI] [Google Scholar]
- 12.Merchie G, Lavens P, Sorgeloos P. Optimization of dietary vitamin C in fish and crustacean larvae: A review. Aquaculture. 1997;155:165–181. doi: 10.1016/S0044-8486(97)00115-4. [DOI] [Google Scholar]
- 13.Gadient M, Schai E. Leaching of various vitamins from shrimp feed. Aquaculture. 1994;124:201–205. doi: 10.1016/0044-8486(94)90378-6. [DOI] [Google Scholar]
- 14.He H, Lawrence AL. Vitamin C requirements of the shrimp Penaeus vannamei. Aquaculture. 1993;114:305–316. doi: 10.1016/0044-8486(93)90305-I. [DOI] [Google Scholar]
- 15.Kong F, et al. Effect of dietary vitamin C on the growth performance, nonspecific immunity and antioxidant ability of red swamp crayfish (Procambarus clarkii) Aquaculture. 2021;541:736785. doi: 10.1016/j.aquaculture.2021.736785. [DOI] [Google Scholar]
- 16.Kong F, Yu H, Wang X, Abouel Azm FR, Tan Q. Optimal dietary l-ascorbyl-2-monophosphate inclusion improved the growth performance and nonspecific immunity of red swamp crayfish (Procambarus clarkii) Aquac. Nutr. 2021;27:2355–2364. doi: 10.1111/anu.13368. [DOI] [Google Scholar]
- 17.Waagbø R. Water-soluble vitamins in fish ontogeny. Aquac. Res. 2010;41:733–744. doi: 10.1111/j.1365-2109.2009.02223.x. [DOI] [Google Scholar]
- 18.Xu J-Y, Wang T-T, Wang Y-F, Peng Y. Effect of combined fish meal: Soybean meal ratio, vitamin C and fish oil supplementations in diet on the growth and reproduction of red swamp crayfish, Procambarus clarkii (Crustacea: Decapoda) Aquac. Res. 2010;41:e252–e259. doi: 10.1111/j.1365-2109.2010.02517.x. [DOI] [Google Scholar]
- 19.Asaikkutti A, Bhavan PS, Vimala K, Karthik M, Cheruparambath P. Effect of different levels dietary vitamin C on growth performance, muscle composition, antioxidant and enzyme activity of freshwater prawn, Macrobrachium malcolmsonii. Aquac. Rep. 2016;3:229–236. doi: 10.1016/j.aqrep.2016.04.002. [DOI] [Google Scholar]
- 20.Lavens, et al. Supplementation of ascorbic acid 2-monophosphate during the early postlarval stages of the shrimp Penaeus vannamei. Aquac. Nutr. 1999;5:205–209. doi: 10.1046/j.1365-2095.1999.00106.x. [DOI] [Google Scholar]
- 21.Council NR. Nutrient Requirements of Fish and Shrimp. National Academies Press; 2011. [Google Scholar]
- 22.Dabrowski K. Administration of gulonolactone does not evoke ascorbic acid synthesis in teleost fish. Fish Physiol. Biochem. 1991;9:215–221. doi: 10.1007/BF02265142. [DOI] [PubMed] [Google Scholar]
- 23.Celada JD, et al. Effects of vitamin C inclusion in practical diets on survival and growth of juvenile crayfish (Pacifastacus leniusculus Dana, Astacidae) from the onset of exogenous feeding. Aquac. Nutr. 2013;19:110–116. doi: 10.1111/j.1365-2095.2012.00949.x. [DOI] [Google Scholar]
- 24.Shiau S-Y, Hsu T-S. Vitamin C requirement of grass shrimp, Penaeus monodon, as determined with l-ascorbyl-2-monophosphate. Aquaculture. 1994;122:347–357. doi: 10.1016/0044-8486(94)90343-3. [DOI] [Google Scholar]
- 25.Wang X, Kim K-W, Bai SC, Huh M-D, Cho B-Y. Effects of the different levels of dietary vitamin C on growth and tissue ascorbic acid changes in parrot fish (Oplegnathus fasciatus) Aquaculture. 2003;215:203–211. doi: 10.1016/S0044-8486(02)00042-X. [DOI] [Google Scholar]
- 26.AOAC . Official Methods of Analysis. 13. Association of Analytical Chemists; 2012. [Google Scholar]
- 27.Li S, et al. Effects of dietary carbohydrate/lipid ratios on non-specific immune responses, antioxidant capacity, hepatopancreas and intestines histology, and expression of TLR-MAPK/NF-κB signaling pathway-related genes of Procambarus clarkii. Fish Shellfish Immunol. 2022;124:219–229. doi: 10.1016/j.fsi.2022.04.005. [DOI] [PubMed] [Google Scholar]
- 28.Davis DA, Robinson EH. Estimation of the dietary lipid requirement level of the white cayfish Procambarus acutus acutus. J. World Aquac. Soc. 1986;17:37–43. doi: 10.1111/j.1749-7345.1986.tb00552.x. [DOI] [Google Scholar]
- 29.Reitman S, Frankel S. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol. 1957;28:56–63. doi: 10.1093/ajcp/28.1.56. [DOI] [PubMed] [Google Scholar]
- 30.Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 31.Doumas BT, Ard Watson W, Biggs HG. Albumin standards and the measurement of serum albumin with bromcresol green. Clin. Chim. Acta. 1971;31:87–96. doi: 10.1016/0009-8981(71)90365-2. [DOI] [PubMed] [Google Scholar]
- 32.Ellis AE. Lysozyme assays. Tech. Fish Immunol. 1990;1:101–103. [Google Scholar]
- 33.Song Y-L, Hsieh Y-T. Immunostimulation of tiger shrimp (Penaeus monodon) hemocytes for generation of microbicidal substances: Analysis of reactive oxygen species. Dev. Compar. Immunol. 1994;18:201–209. doi: 10.1016/0145-305X(94)90012-4. [DOI] [PubMed] [Google Scholar]
- 34.Aebi H. Methods in Enzymology. Academic Press; 1984. pp. 121–126. [DOI] [PubMed] [Google Scholar]
- 35.McCord JM, Fridovich I. Superoxide dismutase: An enzymic function for erythrocuprein (hemocuprein) J. Biol. Chem. 1969;244:6049–6055. doi: 10.1016/S0021-9258(18)63504-5. [DOI] [PubMed] [Google Scholar]
- 36.Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979;95:351–358. doi: 10.1016/0003-2697(79)90738-3. [DOI] [PubMed] [Google Scholar]
- 37.Ke PJ, Cervantes E, Robles-Martinez C. Determination of thiobarbituric acid reactive substances (TBARS) in fish tissue by an improved distillation–spectrophotometric method. J. Sci. Food Agric. 1984;35:1248–1254. doi: 10.1002/jsfa.2740351117. [DOI] [Google Scholar]
- 38.Bancroft JD, Gamble M. The hematoxylin and eosin. In: Suvarna SK, Layton C, Bancroft JD, editors. Theory and Practice of Histological Techniques. 7. Churchill Livingstone; 2013. pp. 179–220. [Google Scholar]
- 39.Zeitoun IH, Ullrey DE, Magee WT, Gill JL, Bergen WG. Quantifying nutrient requirements of fish. J. Fish. Res. Board Canada. 1976;33:167–172. doi: 10.1139/f76-019. [DOI] [Google Scholar]
- 40.Hou Y, Ji M, Zhang W, Huo Y, Zhou Q. Dietary vitamin C requirement of juvenile swimming crab (Portunus trituberculatus) Chin. J. Anim. Nutr. 2015;27:3772–3781. [Google Scholar]
- 41.Niu J, et al. Nutrient values of dietary ascorbic acid (l-ascorbyl-2-polyphosphate) on growth, survival and stress tolerance of larval shrimp, Litopenaeus vannamei. Aquac. Nutr. 2009;15:194–201. doi: 10.1111/j.1365-2095.2008.00583.x. [DOI] [Google Scholar]
- 42.Asaikkutti A, Bhavan PS, Vimala K, Karthik M. Effect of different levels of dietary vitamin C on growth performance, muscle composition, antioxidant and enzyme activity of Macrobrachium rosenbergii. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2018;88:477–486. doi: 10.1007/s40011-016-0772-5. [DOI] [Google Scholar]
- 43.D'Abramo LR, Moncreiff CA, Holcomb FP, Labrenty Montanez J, Buddington RK. Vitamin C requirement of the juvenile freshwater prawn, Macrobrachium rosenbergii. Aquaculture. 1994;128:269–275. doi: 10.1016/0044-8486(94)90316-6. [DOI] [Google Scholar]
- 44.Arnold, J. E. Schalm's Veterinary Hematology 1233–1240 (2022).
- 45.Sladkova SV, Kholodkevich SV. Total protein in hemolymph of crawfish Pontastacus leptodactylus as a parameter of the functional state of animals and a biomarker of quality of habitat. J. Evol. Biochem. Physiol. 2011;47:160–167. doi: 10.1134/S0022093011020058. [DOI] [PubMed] [Google Scholar]
- 46.Abdel-Latif HMR, et al. Growth performance, antioxidant activities, and immunological responses of hapa-reared thinlip mullet (Liza ramada) juveniles fed on diets supplemented with spirulina (Arthrospira platensis) Fish Shellfish Immunol. 2022;130:359–367. doi: 10.1016/j.fsi.2022.09.026. [DOI] [PubMed] [Google Scholar]
- 47.Khalil HS, et al. Metabolic growth, plankton selectivity, haemato-biochemical and intestinal morphometry of Nile tilapia (Oreochromis niloticus) fed a lysine-deficient diet in earthen ponds. Aquac. Rep. 2022;24:101122. doi: 10.1016/j.aqrep.2022.101122. [DOI] [Google Scholar]
- 48.Bruslé J, Anadon GG. Fish Morphology. Routledge; 2017. pp. 77–93. [Google Scholar]
- 49.Zaki MAA, et al. Assessment of zootechnical parameters, intestinal digestive enzymes, haemato-immune responses, and hepatic antioxidant status of Pangasianodon hypophthalmus fingerlings reared under different stocking densities. Aquac. Int. 2023 doi: 10.1007/s10499-023-01092-w. [DOI] [Google Scholar]
- 50.Kord MI, et al. The immunostimulatory effects of commercial feed additives on growth performance, non-specific immune response, antioxidants assay, and intestinal morphometry of Nile tilapia, Oreochromis niloticus. Front. Physiol. 2021 doi: 10.3389/fphys.2021.627499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Zhang H-W, Sun C, Sun S-S, Zhao X-F, Wang J-X. Functional analysis of two invertebrate-type lysozymes from red swamp crayfish, Procambarus clarkii. Fish Shellfish Immunol. 2010;29:1066–1072. doi: 10.1016/j.fsi.2010.08.023. [DOI] [PubMed] [Google Scholar]
- 52.Lee M-H, Shiau S-Y. Dietary vitamin C and its derivatives affect immune responses in grass shrimp, Penaeus monodon. Fish Shellfish Immunol. 2002;12:119–129. doi: 10.1006/fsim.2001.0357. [DOI] [PubMed] [Google Scholar]
- 53.Liang X-P, Li Y, Hou Y-M, Qiu H, Zhou Q-C. Effect of dietary vitamin C on the growth performance, antioxidant ability and innate immunity of juvenile yellow catfish (Pelteobagrus fulvidraco Richardson) Aquac. Res. 2017;48:149–160. doi: 10.1111/are.12869. [DOI] [Google Scholar]
- 54.Verlhac V, Obach A, Gabaudan J, SchÜEp W, Hole R. Immunomodulation by dietary vitamin C and glucan in rainbow trout (Oncorhynchus mykiss) Fish Shellfish Immunol. 1998;8:409–424. doi: 10.1006/fsim.1998.0148. [DOI] [Google Scholar]
- 55.Santos-Sánchez NF, Salas-Coronado R, Villanueva-Cañongo C, Hernández-Carlos B. Antioxidant compounds and their antioxidant mechanism. Antioxidants. 2019;10:1–29. [Google Scholar]
- 56.Winston GW, Di Giulio RT. Prooxidant and antioxidant mechanisms in aquatic organisms. Aquat. Toxicol. 1991;19:137–161. doi: 10.1016/0166-445X(91)90033-6. [DOI] [Google Scholar]
- 57.Gaweł S, Wardas M, Niedworok E, Wardas P. Malondialdehyde (MDA) as a lipid peroxidation marker. Wiad Lek. 2004;57:453–455. [PubMed] [Google Scholar]
- 58.Nair, V., O'Neil, C. L. & Wang, P. G. Encyclopedia of Reagents for Organic Synthesis (2008).
- 59.Arrigoni O, De Tullio MC. Ascorbic acid: Much more than just an antioxidant. Biochim. Biophys. Acta Gen. Subj. 2002;1569:1–9. doi: 10.1016/S0304-4165(01)00235-5. [DOI] [PubMed] [Google Scholar]
- 60.Chen S-J, et al. Effect of l-ascorbyl-2-polyphosphate supplementation on growth performance, body composition, antioxidative capacity and salinity stress tolerance of juvenile Pacific white shrimp, Litopenaeus vannamei. Aquac. Res. 2017;48:4608–4622. doi: 10.1111/are.13284. [DOI] [Google Scholar]
- 61.Chen Y-J, et al. Dietary vitamin C requirement and its effects on tissue antioxidant capacity of juvenile largemouth bass, Micropterus salmoides. Aquaculture. 2015;435:431–436. doi: 10.1016/j.aquaculture.2014.10.013. [DOI] [Google Scholar]
- 62.Ibrahim RE, et al. Influence of vitamin C feed supplementation on the growth, antioxidant activity, immune status, tissue histomorphology, and disease resistance in Nile tilapia, Oreochromis niloticus. Aquac. Rep. 2020;18:100545. doi: 10.1016/j.aqrep.2020.100545. [DOI] [Google Scholar]
- 63.Eissa E-SH, et al. Nano-selenium impacts on growth performance, digestive enzymes, antioxidant, immune resistance and histopathological scores of Nile tilapia, Oreochromis niloticus against Aspergillus flavus infection. Aquac. Int. 2023 doi: 10.1007/s10499-023-01230-4. [DOI] [Google Scholar]
- 64.Khalil HS, Mansour AT, Goda AMA, Omar EA. Effect of selenium yeast supplementation on growth performance, feed utilization, lipid profile, liver and intestine histological changes, and economic benefit in meagre, Argyrosomus regius, fingerlings. Aquaculture. 2019;501:135–143. doi: 10.1016/j.aquaculture.2018.11.018. [DOI] [Google Scholar]
- 65.Catacutan MR, Lavilla-Pitogo CR. l-ascorbyl-2-phosphate Mg as source of vitamin C for juvenile Penaeus monodon. Israeli J. Aquac.-Bamidgeh. 1994;46:40–47. [Google Scholar]
- 66.Yusuf A, et al. Growth and metabolic responses of juvenile largemouth bass (Micropterus salmoides) to dietary vitamin c supplementation levels. Aquaculture. 2021;534:736243. doi: 10.1016/j.aquaculture.2020.736243. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.