

Abstract
Tumor angiogenesis plays vital roles in the growth and metastasis of cancer. RNA methylation is one of the most common modifications and is widely observed in eukaryotes and prokaryotes. Accumulating studies have revealed that RNA methylation affects the occurrence and development of various tumors. In recent years, RNA methylation has been shown to play an important role in regulating tumor angiogenesis. In this review, we mainly elucidate the mechanisms and functions of RNA methylation on angiogenesis and progression in several cancers. We then shed light on the role of RNA methylation-associated factors and pathways in tumor angiogenesis. Finally, we describe the role of RNA methylation as potential biomarker and novel therapeutic target.
Keywords: RNA methylation, Tumor angiogenesis, Cancer progression, MiRNA
Introduction
Cancers represent complex ecosystems comprising tumor cells and a multitude of non-cancerous cells, embedded in an altered extracellular matrix [1]. Angiogenesis is the process by which blood vessels are formed through the proliferation and migration of endothelial cells on the basis of original blood vessels; indeed, the vasculature is much more than a passive, homogeneous conduit for oxygen and nutrient supply [2]. Under physiological conditions, angiogenesis plays significant roles in wound healing, bone repair and regeneration, and the transportation of substances that are needed by the human body [3–5]. Moreover, angiogenesis acts an essential role in tumor metastasis and expansion, atherogenesis and inflammatory diseases under pathological conditions [6–9]. In most cases, the progression of a small mass of cancerous cells to a life-threatening tumor depends upon the initiation of angiogenesis and involves the dysregulation of the angiogenic balance [10].
After the discovery of the first RNA demethylase (fat mass and obesity-associated protein, FTO) more than 10 years ago, researchers gradually elucidated that RNA methylation is a reversible process [11, 12]. This discovery also made RNA methylation research a point focus for scholars. With the development of RNA methylation immunoprecipitation sequencing technology, an increasing number of RNA methylation sites have been identified [13, 14]. RNA methylation participates in the regulation of RNA splicing [15–17] and protein synthesis [18, 19]. To date, over 100 different types of RNA modifications have been identified [20], and N6-methyladenosine (m6A) is the most common RNA modification that is observed in mRNAs [21]. (Fig. 1). In human cancer, abnormal m6A modification has been reported to affect tumor proliferation, migration and invasion [22].
Fig. 1.

The molecular mechanism of m6A modification. m6A is installed by “writers” (METTL3/14, WTAP, RBM15/15B, VIRMA, METTL16 and ZC3H13), removed by “erasers” (FTO, ALKBH5, and ALKBH3, termed “erasers”), and recognized by “readers” (YTHDC1/2, YTHDF1/2/3, IGF2BP1/2/3, HNRPNPA2B1, HNRNPC, and eIF3, termed “readers”)
At present, with increasing research on tumor angiogenesis, researchers have come to understand that there is a relationship between RNA methylation and tumor angiogenesis [23, 24]. Currently, the association between RNA methylation and tumor angiogenesis and the effect of this association on tumor progression remain unclear. In this review, we summarize the role of RNA methylation in tumor angiogenesis and describe the effects of RNA methylation on some types of tumors and the mechanisms underlying their functions. Furthermore, we propose that RNA methylation has clinical value as a cancer biomarker and therapeutic target.
Function and mechanism of RNA methylation in regulating Tumor angiogenesis
In 1971, Folkman first suggested that angiogenesis was required for the development and growth of solid tumors beyond the size of 1‑2 mm3 [25]. In addition, tumor tissues are characterized by high angiogenic capacity. The RhoA and PI3K-Akt pathways, which are common signal transduction pathways, play important roles in the development and progression of cancer, especially in the regulation of TGFβ induced epithelial mesenchymal transition (EMT), tumor progression and angiogenesis [26]. TGFβ, which is the target gene of methyltransferase-like protein 14(METTL14), is regulated by the upregulation or knockout of METTL14. METTL14 modifies TGFβ through the RhoA and PI3K-Akt pathways, which are considered to be related to tumor angiogenesis and tumor progression [27]. The Wnt/Fzd/β- catenin signalling pathway is a common pathway through which many factors [28] and various enzyme proteins to regulate tumor angiogenesis [29, 30]. For example, the BCL9 protein, which is a transcriptional Wnt/β-catenin cofactor, is the angiogenesis promoting element of the Wnt pathway in colorectal cancer (CRC) [31]. RNA methylation recognition factors affect the Wnt/β-catenin pathway in an m6A dependent manner [32], which may ultimately affect on tumor angiogenesis. The HIF-1α/VEGF/VEGFR pathway is currently considered to be related to angiogenesis and tumor progression in a variety of tumors [33–35]. Among the components of this pathway, the vascular endothelial growth factor (VEGF) family is considered to be an important regulator of tumor angiogenesis, and VEGF and its receptor VEGFR are involved in the regulation of tumor angiogenesis and tumor growth [36]. The expression of VEGF and hypoxia inducible factor HIF-1 α have the interwovenness; the latter binds to hypoxia response elements in the promoter of the VEGF gene to participate in the regulation of VEGF expression [37]. RNA methylation molecules, such as methyltransferase-like protein 3(METTL3) and insulin growth factor-2 binding protein 3(IGF2BP3), have been shown to potentially influence the expression of HIF-1α via RNA methylation [38, 39]. Therefore, RNA methylation may affect tumor angiogenesis and tumor progression through the HIF-1 α/ VEGF /VEGFR pathway.
TGFβ promotes tumor growth and metastasis by inducing factors associated with angiogenesis and promoting EMT [40]. Furthermore, RNA methylation plays an important role in the regulation of TGFβ, which has been proven to play a key role in tumor progression. In support of this finding, hypoxia (which is known to promote cancer progression, angiogenesis and metastasis) changes the levels of m6A writers, erasers and readers, resulting in a decrease in m6A levels and an increase in TGFβ1 expression in cancer cells. Some studies have noted that the expression of PDGF ( TGFβ important effectors of tumor progression) [41] and high mobility group A2 (HMGA2) [42] is regulated by changes in m6A levels. In addition, the expression of connective tissue growth factor (CTGF) [43], which is a direct target of angiogenesis-promoting medium and the TGFβ signalling pathway, is significantly reduced in METTL14/ALKBH5-silenced cancer cells [27].
CD34 is another tumor marker that is involved in angiogenesis and has been used as a quantitative indicator of microvessel density (MVD) [44]. An association was observed between low FTO expression and high CD34 expression in intrahepatic cholangiocarcinoma (ICC) [45]. The upregulation or downregulation of FTO-related genes may affect the expression of some relevant oncogenes in tumor cells by affecting mRNA methylation; for example, chemokine CCL19 expression is downregulated in tumor tissues after the knockdown of FTO genes [45]. The chemokine CCL19 has been demonstrated to inhibit angiogenesis in colorectal cancer through a CC-chemokine receptor 7 (CCR7)-dependent manner [46]. These results suggest that FTO inhibition might promote tumor angiogenesis [45]. METTL14 affects tumor angiogenesis in a manner that is related t to TRAF1 [47]. In addition, METTL3, YTHDF3 and IGF2BP3 affect tumor angiogenesis by regulating the expression of VEGF.
Regulatory effects of m6A modifiers on angiogenesis in cancers
The m6A modification needs to be catalysed by the methyltransferase complex, which mainly consists of METTL3 and METTL14 [48, 49] and their cofactors, such as wilms tumor 1 associated protein (WTAP) [50], RNA-binding motif 15/15B (RBM15/15B) [51], Cbl proto-oncogene like 1 (CBLL1) [52], Vir-like m6 A methyltransferase associated (VIRMA) [53], zinc finger CCCH-type containing 13 (ZC3H13) [54]and METTL 16 [55]. These factors are considered to be the writers and play an essential role in RNA methylation.
Among these writers, METTL3 mediates a methylation process that is involved in transcriptional regulation, protein, phosphorylation and tumor angiogenesis in tumor tissues [56]. Conversely, METTL3 inhibition or selective knockdown has been proven to reduce the expression of specific targets that are related to tumor angiogenesis, thereby significantly repressing tumor angiogenesis [57–60]. Similar to METTL3, METTL14 has been proven to participate in the regulation of RNA methylation. It has been suggested that silencing METTL14 may inhibit tumor angiogenesis [27, 47, 61]. For example, studies have reported that tumor necrosis factor receptor (TNFR) associated factor 1 (TRAF1), which is a signalling adaptor that was first recognized as a part of the TNFR2 signalling complex, plays various roles in human disease [62, 63]. TRAF1 overexpression significantly enhances angiogenesis, whereas reduced TRAF1 expression inhibits angiogenesis [47]. Both effects are mediated via the AKT/mTOR/HIF1a/VEGFA pathway. METTL14-mediated m6A modification promotes the stability of the TRAF1 mRNA in an IGF2BP2-dependent manner, thereby significantly promoting tumor angiogenesis [47].
RNA methylation is a dynamic and reversible modification process, and the FTO gene and alkb homologue 5 (ALKBH5) are considered to be involved in m6A demethylation modification [64]. Both of these molecules are called erasers. In addition, ALKBH3 has recently been proven to participate in m6A demethylation, especially in tRNA demethylation [65]. Screening m6 A levels by silencing the demethylase ALKBH5 inhibits cancer growth and angiogenesis. In ALKBH5-knockdown cells, the expression of some genes that are related to angiogenesis as well as cell cycle progression is decreased, which in turn inhibits angiogenesis and tumor progression [27]. FTO, which is the first m6A demethylase that was identified, belongs to the superfamily of Fe (II)-and 2-oxoglutarate-dependent dioxygenases and regulates integrin signalling pathways, inflammatory signalling pathways, epidermal growth factor receptor (EGFR) signalling pathways, angiogenesis and pyrimidine metabolism pathways [45]. When FTO is knocked down, the expression of angiogenesis inhibitors in tumor tissues is reduced, thus promoting tumor angiogenesis [45].
“Readers” are a class of variable RNA-binding proteins that have been shown to specifically select, recognize m6A sites and convey information, thus helping establish an efficient and orderly m6A regulatory network. The existing reports about readers primarily focus on three main categories. The first category includes the members of the YT521-B homology (YTH) domain family, including YTHDF1/2/3 and YTHDC1/2. These molecules have conserved m6A-binding domains that bind to RNA containing m6A, and they are the most important readers [66]; Second, the members of the heterogeneous nuclear ribonucleoprotein (HNRNP) family also play an important role in acting as readers. HNRNPA2/B1 is capable of recognizing m6A on a subset of primary microRNA (pri-miRNA) transcripts and interacts with drosha ribonuclease III (DROSHA) and DiGeorge syndrome critical region 8 (DGCR8), thus facilitating pri-miRNA processing [67]; Third, insulin-like growth factor 2 mRNA-binding proteins (IGF2BPs, including IGF2BP1/2/3) that identify m6A and promote mRNA stability and translation in an m6A-dependent manner are also involved in RNA methylation [68]. Among these proteins, knockdown of the IGF2BP3 gene inhibits hypoxia-induced angiogenesis in vivo by downregulating HIF1A, which inhibits angiogenesis mainly by reducing the expression of pro-angiogenic factor (VEGF) in tumor tissues [39, 69]. Moreover, lower YTHDF2 protein levels are significantly associated with more multinodular tumor and microvascular infiltration, higher TNM and Barcelona Clinic Liver Cancer (BCLC) staging classification, and shorter overall and recurrence-free survival [70]. Mechanistically, YTHDF2 mediates the decay of m6A-containing interleukin 11 (IL11) and serpin family E member 2 (SERPINE2) mRNAs, which are responsible for the disruption of vascular normalization. Therefore, a reduction in hypoxia-sensitive YTHDF2 could reprogram the m6 A-edited transcriptome and promote the development of hepatocellular carcinoma (HCC) [70]. YTHDF3 promotes the interaction of cancer cells with brain endothelial cells and astrocytes, blood-brain barrier extravasation, angiogenesis and growth. YTHDF3 expression promotes angiogenic brain metastasis. The vascular density in metastatic tumor tissues is significantly reduced after YTHDF3 knockdown, and the expression of VEGFA and other genes is positively correlated with the expression of YTHDF3. When both genes are simultaneously knocked down, the angiogenic ability of tumor tissues is further reduced(Table 1) [71].
Table 1.
The functions of m6A enzymes in the angiogenesis of various cancers
| Type | Molecule | Role in tumors | Tumor type | Mechanism | References |
|---|---|---|---|---|---|
| Writers | METTL3 | Oncogene | Bladder cancer | Promote angiogenesis via modulating TEK and VEGF-A | [72] |
| Oncogene | GC | Promote angiogenesis via stimulating m6a modification of HDGF mRNA | [58] | ||
| Oncogene | HCC | Promote angiogenesis via modulating Hippo pathway | [57] | ||
| Oncogene | RCC | Promote angiogenesis via modulating HIF-2α | [73] | ||
| METTL14 | Oncogene | Breast cancer | Promote angiogenesis via modulating VEGFA and RhoA and PI3K-Akt pathways | [27] | |
| Erasers | FTO | Suppressor gene | ICC | FTO expression is inversely correlated with MVD. | [45] |
| ALKBH5 | Oncogene | Breast cancer | Promote angiogenesis via modulating VEGFA and RhoA and PI3K-Akt pathways | [27] | |
| Readers | IGF2BP2 | Oncogene | Lung cancer | Promote angiogenesis via modulating HNF4G/IGF2BP2/TK1 axis | [74] |
| IGF2BP3 | Oncogene | GC | Promote angiogenesis via enhancing HDGF mRNA stability | [58] | |
| Oncogene | CRC | Promote angiogenesis via modulating VEGF | [39] | ||
| Oncogene | GBM | Promote angiogenesis via modulating PI3K/MAPK pathways | [75] | ||
| YTHDF1 | Oncogene | GC | Promote angiogenesis via modulating Wnt/β-catenin pathway | [32] | |
| YTHDF2 | Suppressor gene | HCC | Suppress angiogenesis via downregulating IL11 and SERPINE2 | [70] | |
| YTHDF3 | Oncogene | Breast cancer | Promote angiogenesis via modulating VEGFA and EGFR | [71] |
Effects of RNA methylation on angiogenesis in different cancers
Bladder cancer is among the top ten most common cancer types in the world, with approximately 550,000 new cases annually [76]. METTL3-mediated methylation may be involved in transcriptional regulation, protein, phosphorylation and angiogenesis in bladder cancer. For example, there are abundant m6A peaks in the TEK gene as well as VEGF-A gene, which are associated with tumor angiogenesis [77, 78]. In contrast, up- or down-regulation of METTL3 in bladder cancer cells can enhance or inhibit the transcriptions and protein expression of TEK and VEGF-A, which are components of the PI3K/AKT pathway respectively, thereby affecting the angiogenesis of bladder cancer via RNA methylation [72] (Fig. 2).
Fig. 2.
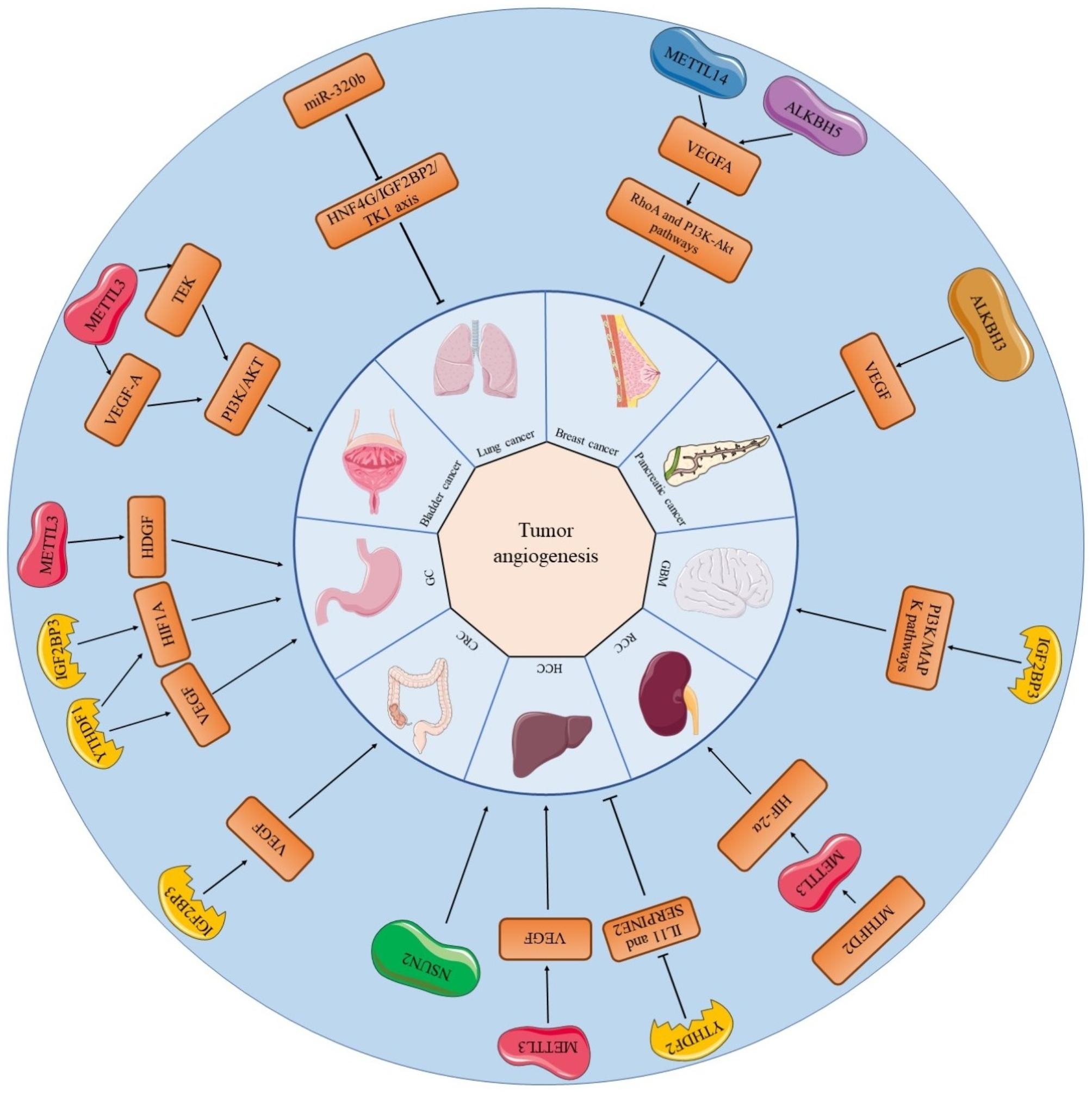
RNA methylation regulators related to angiogenesis in different cancers and their related mechanisms. Angiogenesis in some cancers is associated with more than one RNA methylation regulators. The mechanism of tumor angiogenesis varies from one tumor to another
Gastric cancer (GC) is the fifth most common cancer and the third most common cause of cancer-related death worldwide [79]. In gastric cancer, the upregulation or knockdown of METTL3 is followed by an increase or decrease respectively in m6A levels [58]. Mechanistically, P300-mediated H3K27 acetylation in the promoter region of METTL3 induces METTL3 transcription, which stimulates the m6A modification of hepatoma-derived growth factor (HDGF) mRNA; then the m6A reader IGF2BP3 directly recognizes and binds to the m6A site on HDGF mRNA and enhances HDGF mRNA stability. Secreted HDGF promotes tumor angiogenesis, while nuclear HDGF activates glucose transporter type 4 (GLUT4) and enolase 2 (ENO2) expression, which is followed by increased glycolysis in GC cells. In METTL3 overexpressing gastric cancer tumor tissues, MVD, as assessed by staining for CD31 (a marker of angiogenesis), is significantly increased compared to controls [58]. In hypoxic environments, hypoxic tumor cells undergo multiple alterations at the molecular, cellular and phenotypic levels (e.g., increased migration, invasion and angiogenesis), which contribute to their survival at primary and secondary sites or escape from unfavorable tumor environments [80, 81]. IGF2BP3 is involved in the regulation of the hypoxic responses [82, 83], which may account for the angiogenesis in cancerous tissues [39]. In gastric cancer cells that are exposed to hypoxia, deletion of IGF2BP3 suppresses VEGF expression. Studies have revealed that this effect can be achieved in gastric cancer cells by regulating HIF1A expression through IGF2BP3 binding to a specific m6A site on HIF1A mRNA [69]. HIF1A has been shown to be a major hypoxic response factor [84]. In hypoxic environment, HIF1A signalling has been proven to regulate a variety of tumorigenesis and progression, such as tumor cell survival, proliferation, metastasis, and angiogenesis [85, 86] (Fig. 2).
Although colorectal cancer (CRC) has been studied for years, it is still one of the leading causes of cancer-related death worldwide [87]. IGF2BP3 binds to and reads the m6A recognition site in VEGF mRNA in colon cancer cells, thus regulating the expression and stability of VEGF mRNA. While VEGF is an important growth factor for tumor angiogenesis, knockdown of IGF2BP3 may inhibit angiogenesis in colon cancer by regulating the reduction of VEGF expression [39] (Fig. 2).
Hepatocellular carcinoma (HCC) is the fourth leading cause of cancer-related death worldwide, despite the liver being the sixth most common site of primary cancer [88]. The concept of vasculogenic mimicry (VM) offers a new perspective on the supply of blood to tumors [89]. Existing studies have confirmed that VM is associated with angiogenesis in a variety of tumors, including HCC [90, 91]. Although VM has been proven to be induced by various extracellular ligands and microenvironments, VEGFa is considered to be the major inducer of this process in HCC cells [92]. The level of m6A in VEGFa-treated cells is also increased and positively correlated with the expression of the RNA methylation enzyme METTL3 in the cells. In contrast, when the expression of METTL3 was silenced, the level of m6A in HCC tissues was decreased, which affected VM [57]. In addition, METTL14 expression was significantly reduced in HCC and was significantly associated with microvascular infiltration [61]. Furthermore, NSUN2, which is the writer of 5-methylcystosine (m5C), has been proved to participate in the regulation of invasion, metastasis, and angiogenesis in HCC. [93] (Fig. 2).
Lung cancer remains the leading cause of cancer related deaths worldwide [94]. METTL3 promotes the growth, survival and invasion of lung cancer cells, and its deletion strongly inhibits cancer cell growth, increases apoptosis, and significantly reduces the invasive ability of cancer cells [59]. In lung cancer, the anti-cancer gene miR-320b downregulates the expression of IGF2BP2 and thymidine kinase 1 (TK1), thus suppressing angiogenesis and lung cancer growth [74] (Fig. 2).
Breast cancer is one of the most common malignancies in women throughout the world and is the major cause of most cancer-related deaths [95]. RNA methylation plays a vital role in promoting the angiogenesis and progression of breast cancer. METTL14 and ALKBH5 regulate key cell cycle- and angiogenesis-associated genes transcriptions. METTL14 or ALKBH5 overexpression leads to increased expression of target genes including VEGFA compared to vector control [27]. Angiogenesis is a major step in metastasis. The blood vessel density of metastases formed by YTHDF3 KD human breast cancer MDA-MB-231 parental cells was significantly lower than that of metastases formed by control cells. These results suggest that YTHDF3 expression facilitates angiogenic brain metastasis. Overexpression of YTHDF3 increases the invasive and angiogenic potential of cells in vitro and promotes breast cancer metastasis [71]. In addition, in breast cancer tissues, FTO overexpression was significantly correlated with tumor size, peritumor lymphovascular infiltration, and lymph node metastasis [96] (Fig. 2).
Glioblastomas (GBMs) are heterogeneous and invariably lethal tumors [97]. In glioma cells, upregulation of IGF2BP3 expression promotes tumor proliferation, invasion, migration and angiogenesis, and is also significantly associated with lower survival rates [75] (Fig. 2).
Renal cell carcinomas (RCCs) are kidney tumors that arise from the epithelial layer of the nephron [98]. Methylenetetrahydrofolate dehydrogenase 2 (MTHFD2) plays a critical role in controlling global m6A methylation levels, including the m6A methylation levels of HIF-2α mRNA, which results in enhanced translation of HIF-2α. In addition, VEGFA, which is related to tumor angiogenesis, is a transcript that is targeted by HIF-2α [73] (Fig. 2).
Pancreatic cancer is a malignancy with a poor prognosis and high mortality rate [99]. PCA-1 has been shown to be identical to ALKBH3, which is the demethylase of m1A modification [100]. Previous studies indicated that PCA-1/ALKBH3 silencing reduces VEGF expression in human pancreatic cancer and that PCA-1/ALKBH3 downregulation significantly decreases the number of microvessels in tumors [101] (Fig. 2).
Effect of RNA methylation-associated miRNAs on Tumor angiogenesis
MicroRNAs (miRNAs) are small endogenous RNAs that regulate gene-expression at the posttranscriptional level [102]. Relevant reports indicate that m6A modification participates in regulating the biogenesis of miRNA [103, 104]. Among these miRNA targets, some are involved in the regulation of tumor angiogenesis [91].
Endothelial cells(ECs) play a vital role in tumor angiogenesis [105]. METTL3 depletion in ECs decreases mature angiogenic miRNAs let-7e-5p and the miR-17-92 cluster, and increases the expression of their common target, Tsp1 (thrombospondin 1). Conversely, METTL3 increases the expression of let-7e-5p and miR-17-92 cluster and reduces protein levels of Tsp1 in ECs [106]. Therefore, miR-17-92 regulates tumor angiogenesis via m6A modification.
Previous studies have indicated that zinc ribbon domain-containing 1-antisense 1 (ZNRD1-AS1) is described as an oncogene in several types of cancer [107, 108]. ZNRD1-AS1 overexpression inhibits cell proliferation, migration and angiogenesis. YTHDC2 promotes ZNRD1-AS1 stability in malignant lung cells via m6A modification. Furthermore, the miR-942/tensin1 (TNS1) axis was demonstrated to be the downstream regulatory signaling pathway of ZNRD1-AS1. ZNRD1-AS1 promotes malignant lung cell proliferation, migration, and angiogenesis via the miR-942/TNS1 axis and is positively regulated by the m6A reader YTHDC2 [109].
The expression of circ-CCT3 is remarkably upregulated in HCC and associated with poor prognosis. Functionally and mechanistically, circ-CCT3 promotes the angiogenesis of HUVEC by the sponge activity on miR-378a-3p and improve FLT1 expression, indicating its potential tumor promoter role in liver cancer development. Furthermore, the m6 A regulator (ALKBH5 and METTL3) could bind and regulate m6 A-modification of circ-CCT3. In summary, Circ-CCT3 promotes angiogenesis via miR-378a-3p-FLT1 axis in HCC and is under m A-modification mediated by ALKBH5 and METTL3 [110].
In METTL14-deficient cells, the expression of miR126 is reduced, since it is downstream target of METTL14 and regulated in an m6A-dependent manner, and it is involved in metastasis and vascular infiltration of HCC. Through the study, miR-126 was found to be significantly reduced in the vascular infiltration positive group and slightly reduced in the liver transplantation recurrence group [61].
Relevant research indicates that knocking down the expression of METTL3 reduces the expression of miR-143-3p in cells. Conversely, when METTL3 was overexpressed, it can in turn promote the expression of miR-143-3p in cells. Angiogenesis is significantly increased in cultures that are transfected with miR-143-3p, and overexpression of miR-143-3p can increase VEGFA expression in lung cancer cells, thereby regulating angiogenesis in lung cancer tissues [111]. In addition, the expression of regulators associated with tumor metastasis or angiogenesis, such as Vasohibin-1 (VASH1), is also affected in cells transfected with miR-143-3p. One study showed that miR-143-3p might be involved in negatively regulating the expression of VASH1, which in turn was involved in promoting cancer cell dissemination and angiogenesis [112].
In CRC tissues, circ3823 is significantly highly expressed, and m6A modification has been proven to be involved in regulating its degradation. It has been demonstrated that circ3823 may promote the proliferation, metastasis and angiogenesis of colorectal cancer through the circ3823/miR-30c-5p/TCF7 axis [113]. Another study showed that the overexpression of miR-320b can inhibit cell invasion and angiogenesis in vitro and suppress angiogenesis and tumor growth in vivo. Hepatocyte nuclear factor 4 gamma (HNF4G) is a direct target gene of miR-320b, which has been proven to upregulate IGF2BP2 expression [74]. Furthermore, it has been reported that IGF2BP2 can recognize the m6A modification of the TK1 gene to increase TK1 expression [68], thus promoting angiogenesis.
In summary, these findings suggest that various miRNAs exert effects that regulate tumor angiogenesis via mechanisms that are related to RNA methylation. This discovery may provide distinguished indicators and drug intervention targets for cancer treatment.
Regulation of Tumor angiogenesis by Hippo/YAP1 pathway-related RNA methylation
In general, yes-associated protein (YAP) and transcriptional co-activator with PDZ-binding motif (TAZ) are inhibited by the Hippo pathway in normal quiescent adherent cells, and this process occurs in the cytoplasm [114]. The Hippo pathway is an important pathway that plays a crucial role in the malignant progression of cancer [115]. Relevant studies have indicated that various types of cancers, including breast, lung, pancreatic, liver, colorectal, gastric cancer and glioma, are accompanied by increased YAP/TAZ levels and activity [116–123]. Furthermore, numerous studies have noted that the Hippo/YAP1 pathway is closely related to RNA methylation and proteins associated with RNA methylation, and the methylation of RNAs that are associated with this pathway may regulate the pathway [56, 123–126]. In addition, recent studies have proposed that the Hippo/YAP1 pathway is inextricably linked to tumor angiogenesis [127, 128]. There is a question: does RNA methylation participate in regulating tumor angiogenesis through the Hippo/YAP1 pathway? The answer is yes. Currently, numerous relevant studies indicate that Hippo/YAP1 pathway-related RNA methylation plays a regulatory role in tumor angiogenesis. The role of the Hippo pathway in m6A-mediated VM formation was investigated, and the level of m6A modification of YAP1 mRNA was significantly reduced. This study indicates that YAP1 promotes tumor vasculature formation and malignant progression in an m6A-dependent manner in vitro. YAP1 is suppressed by the downregulation of METTL3 after a decrease in m6A, and upregulated by the overexpression of ALKBH5 (an important m6A demethylase) and the induction of VEGFa after an increase in m6A. Furthermore, m6A has been proven to alter the expression of the YAP1 protein by increasing translation efficiency, which may trigger the degradation of YAP1 pre-mRNA and the degradation of mature mRNA in HCC [57]. Additionally, accumulating studies have confirmed that the m6A demethylase ALKBH5 inhibits tumor growth, metastasis, invasion, and tumor angiogenesis in NSCLC by reducing YTHDFs-mediated YAP1 expression and inhibiting miR-107/LATS2-mediated YAP1 activity. This mechanism indicates that ALKBH5 may reduce the m6A level of YAP1 precursor mRNA in NSCLC, thus affecting the Hippo/YAP1 pathway [124].
Effects of RNA methylation on the blood-brain barrier and angiogenesis associated with Tumor metastasis
The blood–brain barrier (BBB) is a continuous endothelial membrane within brain microvessels that are composed of sealed cell-to-cell contacts, and is sheathed by mural vascular cells and perivascular astrocyte end-feet [129]. Currently, the main sources of metastatic brain cancer are lung cancer and breast cancer [130]. Tumor cells that have entered the bloodstream need to penetrate the BBB to gain access to the brain parenchyma [131]. Therefore, the development of brain metastatic carcinoma is usually associated with abnormalities in the blood-brain barrier, a process that is dependent on abnormal angiogenesis. For example, miR-143-3p expression is upregulated in brain metastatic cancer tissues compared to primary cancer tissues [111]. Inhibition of vasopressor-1 (VASH1) expression by targeting the three binding sites that control its 3’UTR enhances the invasiveness and angiogenesis of lung cancer in an in vitro BBB model. The m6A methyltransferase METTL3 increases splicing of the precursor miR-143-3p to promote its biogenesis. Conversely, transfection with miR-143-3p inhibitors blocks angiogenesis as well as VEGFA expression in tumor tissues [111].
YTHDF3 contains a C-terminal YTH structural domain that specifically binds to m6A-containing RNAs. It has been suggested that overexpression of the m6A reader YTHDF3 is clinically associated with brain metastasis in breast cancer patients. YTHDF3 promotes the interaction of cancer cells with brain endothelial cells and astrocytes, blood-brain barrier extravasation, angiogenesis and growth. Mechanistically, YTHDF3 enhanced the translation of m6A-enriched transcripts of ST6GALNAC5, GJA1 and EGFR, all of which are associated with brain metastasis. Notably, all of these key brain transfer gene transcripts are enriched in the m6A peak. This study also demonstrates the role of YTHDF3 in facilitating the translation of key brain transfer transcripts and that the process is dependent on the binding of YTHDF3 to m6A-modified mRNAs [71].
Effects of RNA methylation on Tumor angiogenesis and Tumor drug resistance
Drug resistance remains a major challenge to the curative treatment of various cancers [132]. Accumulating evidence has revealed that RNA methylation regulates multiple anticancer drug resistance, including drug transport and metabolism, target receptors, cancer stemness, DNA damage repair and cell death [21, 133–136]. In addition, the variation contributes to drug resistance by regulating DNA damage repair, downstream adaptive responses (apoptosis, autophagy, and oncogenic bypass signaling), cell stemness, the tumor immune microenvironment, and exosomal non-coding RNAs [137].
In previous studies, FOXO3 was identified as a key transcription factor in multiple oncogenic pathways, and it is involved in regulating cell cycle arrest [138], apoptosis [139] and autophagy [140]. Sorafenib is a multitarget drug that inhibits cancer cell proliferation and angiogenesis [141]. Recent studies have noted that FOXO3 is a direct target of METTL3 in HCC cells under intratumor environmental conditions. Silencing METTL3 at the RNA and protein levels significantly reduces the expression of FOXO3. This study also indicates that METTL3-mediated m6A modification promotes FOXO3 RNA stability in a YTHDF1-dependent manner. The reduction in METTL3-m6A function enhances autophagy and angiogenesis in HCC through the METTL3/FOXO3 axis in vivo, thus enhancing sorafenib resistance [142].
EGFR plays a significant role in angiogenesis and drug resistance in CRC [143]. The anti-EGFR antibody cetuximab has been used for CRC therapy for several years [144]. As mentioned previously, FTO and YTHDF3 were proved to influence on the expression of EGFR [45, 71]. Additionally, the m6A reader hnRNPA2B1 plays a vital role in the transcriptional activity of Wnt signalling in CRC via regulation of TCF7L2 mRNA stability, which may contribute to cetuximab resistance [145]. METTL3 enhances the translation efficiency of EGFR, followed by rebound activation of RAF/MEK/ERK, resulting in acquired PLX4032 resistance in melanoma [146]. Besides, YTHDF1 and YTHDF2 affect cancer by binding to the m6A sites in the 3′-UTR of EGFR transcription and contributed to aberrant activities of downstream signal pathways [147, 148].
As a multitarget receptor tyrosine kinase (RTK) inhibitor, sunitinib plays an important role in the treatment of RCC [149]. Previous studies revealed that TRAF1 overexpression promotes sunitinib resistance by regulating apoptosis and angiogenic pathways in a METTL14-dependent manner [47].
Protein arginine methyltransferase 5 (PRMT5) is highly expressed in multiple types of cancer and reported to decreased the breast cancer cell response to doxorubicin [150, 151]. Doxorubicin treatment markedly induces RNA m6A methylation in breast cancer cells and tumor tissues. However, PRMT5 inhibits doxorubicin-induced RNA m6A methylation by enhancing the nuclear translocation of ALKBH5 to the partner AlkB homolog 7 (ALKBH7). ALKBH5 removes the m6A methylation from BRCA1 mRNA and maintains the stability and function of BRCA1, which reduces the sensitivity of cells to doxorubicin [152].
These findings reveal that RNA methylation has immense potential for regulating drug resistance.
The potential of RNA methylation in clinical applications
An increasing number of studies have revealed that RNA methylation is very common in cancer [20]. To illustrate, METTL3, which is an RNA methylation enzyme, is inextricably linked to the process of RNA methylation in tumor cells. Currently, an increasing number of studies point to METTL3 as a biomarker that has a definite relationship with tumor prognosis, metastasis, diagnosis, and drug resistance [153–157]. For example, it has been demonstrated that the mRNA levels of METTL3 are significantly overexpressed in GC tissues and that patients with high METTL3 expression have a shorter median survival time. In addition, elevated expression of METTL3 was significantly associated with TNM stage and vessel invasion of tumors in gastric cancer patients [154]. Moreover, a relevant study also demonstrates that METTL3 expression is an independent prognostic factor and effective predictor in human patients with GC [58].
Moreover, relevant studies indicate that IGF2BP3 is not only a prognostic biomarker but also a diagnostic molecule [158]. An increasing number of studies have shown that the expression of IGF2BP3, as determined by immunohistochemistry, has prognostic significance in a number of cancers, including RCC, CRC, oophoroma, mammary carcinoma, pancreatic ductal adenocarcinoma, cervical carcinoma, and GBM [75, 159–164]. IGF2BP3 has also been shown to be associated with aggressive and advanced carcinomas [165–170]. For example, IGF2BP3 tends to exert oncogenic effects in most malignant gliomas in which IGF2BP3 is overexpressed and plays a role in promoting proliferation, invasion, migration, angiogenesis, and chemoresistance. In addition, IGF2BP3 was specifically highly expressed in glioma cells, enhancing its role as a diagnostic marker. Moreover, the median survival rate of patients that overexpress this protein was lower, indicating its potential as a prognostic indicator for disease evolution [75].
In addition to the clinical application of the m6A modifications described above, m6A methylation regulators have also been described as efficacious therapeutic targets for anticancer drugs in clinical treatment. Recently, studies have confirmed that therapeutically targeting METTL3 may offer an alternative to anti-vascular therapy for bladder cancer [56]. In addition, the m6A reader IGF2BP3 regulates cell cycle and angiogenesis in colon cancer. This study may provide a potential therapeutic target for colon cancer [39]. Furthermore, glioma cells become more sensitive to Taxol, temozolomide, and doxorubicin after the IGF2BP3 gene is knocked down, suggesting a role for IGF2BP3 in chemotherapy resistance [75].
In summary, m6A-related enzymes are not only used as markers of tumor prognosis but also act as drug targets to enhance the sensitivity of tumor treatments. Some clinical trials of RNA methylation in clinical applications are as follows (Table 2).
Table 2.
Clinical trials of the RNA methylation in clinical applications. (Data source: United States clinical trial database https://beta.clinicaltrials.gov/)
| Study title | Application phase | Disease | Clinical Trial No. |
|---|---|---|---|
| Oral Administration of STC-15 in Subjects With Advanced Malignancies | Phase 1 |
Advanced Cancer Advanced Solid Tumor |
NCT05584111 |
| The Red Blood Cells Based Blood Test for Lung Cancer EARLY Detection | Not Applicable | Lung Cancer | NCT05380999 |
| Impact of 5-Fluorouracil (5-FU) on Cancer Cells From Liver Metastases of Colon Cancer (FLUORIB) | Not Applicable |
Cancer of Colon Metastasis to Liver |
NCT04274790 |
| The Role of m6A RNA Modification in Moyamoya Disease | Not Applicable | Moyamoya Disease | NCT04696094 |
Conclusions
In this review, we summarized the progress of research on the role of RNA modification in tumor angiogenesis. We evaluated the biological functions of RNA modification in altering drug resistance and activating signalling pathways. There are various pathways that are involved in this regulation, and they exist in a variety of cancers. In addition, a variety of miRNA molecules that are related to RNA methylation are also involved in tumor angiogenesis and tumor progression. RNA methylation also affects tumor invasion and the degree of vascular invasion by affecting the BBB. Furthermore, RNA methylation related molecules may be used as biomarkers to measure tumor progression and predict prognosis. More importantly, studying the effect of RNA methylation on tumor angiogenesis is beneficial for identifying novel therapeutic drugs to control cancer progression.
Despite the abovementioned encouraging advances, there are still multiple challenges and difficulties associated with the application of RNA methylation in clinical practice. It should be noticed that such applications focus on a single RNA modification and its corresponding regulators [171]. We need more systematic studies to investigate the relationship between RNA methylation and tumor angiogenesis. In addition, accumulating evidence has indicated that RNA modifying proteins((RMPs) has significant potential as pharmacological targets or diagnostic markers [172]. However, the interactions between different RNA modifications and modifiers are only just beginning to be elucidated, leaving some doubt about the clinical application of RMPs. To date, the clinical applications of METTL3 inhibitors have been systematically reviewed [173]. For example, STM2457, which is a highly potent and selective first-in-class catalytic inhibitor of METTL3, has been proven to decrease AML growth and increase differentiation and apoptosis [174]. However, not all of these inhibitors have demonstrated acceptable target potency or enzyme selectivity and the clinical applications of many other modifiers still require further research [175]. Additionally, the effects of RNA methylation on some cancers remain unknown.
In brief, RNA methylation plays an essential role in the regulation of tumor angiogenesis and cancer progression. Although RNA methylation shows great potential for clinical application, there is still substantial work to be done before it can be applied to clinical practice due to current technical limitations.
Acknowledgements
Not applicable.
Abbreviations
- ALKBH
Alkb homologue
- BBB
Blood–brain barrier
- BCLC
Barcelona Clinic Liver Cancer
- CBLL1
Cbl proto-oncogene like 1
- CCR7
CC-chemokine receptor 7
- CRC
Colorectal cancer
- CTGF
Connective tissue growth factor
- EC
Endothelial cell
- EGFR
Epidermal growth factor receptor
- EMT
Epithelial mesenchymal transition
- ENO2
Enolase 2
- FTO
Fat mass and obesity-associated protein
- GBM
Glioblastoma
- GC
Gastric cancer
- GLUT4
Glucose transporter type 4
- HCC
Hepatocellular carcinoma
- HDGF
Hepatoma-derived growth factor
- HIF
Hypoxia inducible factor
- HMGA2
High mobility group A2
- HNF4G
Hepatocyte nuclear factor 4 gamma
- HNRNP
Heterogeneous nuclear ribonucleoprotein
- ICC
Intrahepatic cholangiocarcinoma
- IGF2BPs
Insulin growth factor-2 binding proteins
- IL11
Interleukin 11
- m5c
5-methylcystosine
- m6A
N6-methyladenosine
- METTL
Methyltransferase-like
- miRNA
MicroRNA
- MTHFD2
Methylenetetrahydrofolate dehydrogenase 2
- MVD
Microvessel density
- PRMT5
Protein arginine methyltransferase 5
- RBM
RNA-binding motif
- RCC
Renal cell carcinoma
- RTK
Receptor tyrosine kinase
- SERPINE2
Serpin family E member 2
- TAZ
Transcriptional co-activator with PDZ-binding motif
- TK1
Thymidine kinase 1
- TNFR
Tumor necrosis factor receptor
- TNS1
Tensin1
- TRAF
Tumor necrosis factor receptor-associated factor
- VASH1
Vasohibin-1
- VEGF
Vascular endothelial growth factor
- VIRMA
Vir-like m6 A methyltransferase associated
- VM
Vasculogenic mimicry
- WTAP
Wilms Tumor 1 associated protein
- YAP
Yes-associated protein
- YTHDF
YT521-B homology domain-containing protein family
- ZC3H13
Zinc finger CCCH-type containing 13
- ZNRD1-AS1
Zinc ribbon domain-containing 1-antisense 1
Authors’ contributions
ZQS provided direction and guidance throughout the preparation of this manuscript. MYH, HFS and QBZ finished the manuscript and designed the figures. ZQS, JHH and WTY reviewed and made significant revisions to the manuscript. HFS and JBL collected the related papers. All authors read and approved the final manuscript.
Funding
This study was supported by The National Natural Science Foundation of China (82173055, 81972663, U2004112), The Science Project of Henan Natural Science Foundation (212300410074, 202300410446), The Youth Talent Innovation Team Support Program of Zhengzhou University (32320290), The Provincial and Ministry co-constructed key projects of Henan medical science and technology (SBGJ202102134, SBGJ202102121), Key scientific and technological research projects of Henan Provincial Department of Science and Technology (212102310117), Henan Provincial Health Commission and Ministry of Health Co-construction Project, and Henan Provincial Health and Health Commission Joint Construction Project (LHGJ20200158), Henan Province young and middle-aged health science and technology innovation leading talent project (YXKC2022016), Henan Province Medical Affairs Technology Promotion Project (SYJS2022109).
Availability of data and materials
Not applicable.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
All authors have approved to publish this manuscript.
Competing interests
All authors declare no conflict of interest.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Junhong Hu, Email: hujunhong@zzu.edu.cn.
Weitang Yuan, Email: yuanweitang@zzu.edu.cn.
Zhenqiang Sun, Email: fccsunzq@zzu.edu.cn.
References
- 1.de Visser KE, Joyce JA. The evolving Tumor microenvironment: from cancer initiation to metastatic outgrowth. Cancer Cell. 2023;41:374–403. doi: 10.1016/j.ccell.2023.02.016. [DOI] [PubMed] [Google Scholar]
- 2.Eelen G, Treps L, Li X, Carmeliet P. Basic and Therapeutic aspects of Angiogenesis updated. Circ Res. 2020;127:310–29. doi: 10.1161/CIRCRESAHA.120.316851. [DOI] [PubMed] [Google Scholar]
- 3.Rizov M, Andreeva P, Dimova I. Molecular regulation and role of angiogenesis in reproduction. Taiwan J Obstet Gynecol. 2017;56:127–32. doi: 10.1016/j.tjog.2016.06.019. [DOI] [PubMed] [Google Scholar]
- 4.Augustine R, Prasad P, Khalaf IMN. Therapeutic angiogenesis: from conventional approaches to recent nanotechnology-based interventions. Mater Sci Eng C Mater Biol Appl. 2019;97:994–1008. doi: 10.1016/j.msec.2019.01.006. [DOI] [PubMed] [Google Scholar]
- 5.Guerra A, Belinha J, Jorge RN. Modelling skin wound healing angiogenesis: a review. J Theor Biol. 2018;459:1–17. doi: 10.1016/j.jtbi.2018.09.020. [DOI] [PubMed] [Google Scholar]
- 6.Weidner N, Semple JP, Welch WR, Folkman J. Tumor angiogenesis and Metastasis–correlation in invasive breast carcinoma. N Engl J Med. 1991;324:1–8. doi: 10.1056/NEJM199101033240101. [DOI] [PubMed] [Google Scholar]
- 7.Cao Y. Tumor angiogenesis and molecular targets for therapy. Front Biosci (Landmark Ed) 2009;14:3962–73. doi: 10.2741/3504. [DOI] [PubMed] [Google Scholar]
- 8.Sluimer JC, Daemen MJ. Novel concepts in atherogenesis: angiogenesis and hypoxia in Atherosclerosis. J Pathol. 2009;218:7–29. doi: 10.1002/path.2518. [DOI] [PubMed] [Google Scholar]
- 9.Lee HJ, Hong YJ, Kim M. Angiogenesis in chronic inflammatory skin disorders. Int J Mol Sci 2021, 22. [DOI] [PMC free article] [PubMed]
- 10.Lawler J. Counter regulation of Tumor angiogenesis by vascular endothelial growth factor and thrombospondin-1. Semin Cancer Biol. 2022;86:126–35. doi: 10.1016/j.semcancer.2022.09.006. [DOI] [PubMed] [Google Scholar]
- 11.Jia G, Fu Y, Zhao X, Dai Q, Zheng G, Yang Y, Yi C, Lindahl T, Pan T, Yang YG, He C. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat Chem Biol. 2011;7:885–7. doi: 10.1038/nchembio.687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Han Z, Niu T, Chang J, Lei X, Zhao M, Wang Q, Cheng W, Wang J, Feng Y, Chai J. Crystal structure of the FTO protein reveals basis for its substrate specificity. Nature. 2010;464:1205–9. doi: 10.1038/nature08921. [DOI] [PubMed] [Google Scholar]
- 13.Meyer KD, Saletore Y, Zumbo P, Elemento O, Mason CE, Jaffrey SR. Comprehensive analysis of mRNA methylation reveals enrichment in 3’ UTRs and near stop codons. Cell. 2012;149:1635–46. doi: 10.1016/j.cell.2012.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Dominissini D, Moshitch-Moshkovitz S, Schwartz S, Salmon-Divon M, Ungar L, Osenberg S, Cesarkas K, Jacob-Hirsch J, Amariglio N, Kupiec M, et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature. 2012;485:201–6. doi: 10.1038/nature11112. [DOI] [PubMed] [Google Scholar]
- 15.Mendel M, Delaney K, Pandey RR, Chen KM, Wenda JM, Vågbø CB, Steiner FA, Homolka D, Pillai RS. Splice site m(6)a methylation prevents binding of U2AF35 to inhibit RNA splicing. Cell. 2021;184:3125–3142e3125. doi: 10.1016/j.cell.2021.03.062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Xiao W, Adhikari S, Dahal U, Chen YS, Hao YJ, Sun BF, Sun HY, Li A, Ping XL, Lai WY, et al. Nuclear m(6)a reader YTHDC1 regulates mRNA splicing. Mol Cell. 2016;61:507–19. doi: 10.1016/j.molcel.2016.01.012. [DOI] [PubMed] [Google Scholar]
- 17.Achour C, Bhattarai DP, Groza P, Román ÁC, Aguilo F. METTL3 regulates breast cancer-associated alternative splicing switches. Oncogene. 2023;42:911–25. doi: 10.1038/s41388-023-02602-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Berulava T, Buchholz E, Elerdashvili V, Pena T, Islam MR, Lbik D, Mohamed BA, Renner A, von Lewinski D, Sacherer M, et al. Changes in m6A RNA methylation contribute to Heart Failure progression by modulating translation. Eur J Heart Fail. 2020;22:54–66. doi: 10.1002/ejhf.1672. [DOI] [PubMed] [Google Scholar]
- 19.Sendinc E, Valle-Garcia D, Dhall A, Chen H, Henriques T, Navarrete-Perea J, Sheng W, Gygi SP, Adelman K, Shi Y. PCIF1 catalyzes m6Am mRNA methylation to regulate Gene expression. Mol Cell. 2019;75:620–630e629. doi: 10.1016/j.molcel.2019.05.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Yang B, Wang JQ, Tan Y, Yuan R, Chen ZS, Zou C. RNA methylation and cancer treatment. Pharmacol Res. 2021;174:105937. doi: 10.1016/j.phrs.2021.105937. [DOI] [PubMed] [Google Scholar]
- 21.Ma S, Chen C, Ji X, Liu J, Zhou Q, Wang G, Yuan W, Kan Q, Sun Z. The interplay between m6A RNA methylation and noncoding RNA in cancer. J Hematol Oncol. 2019;12:121. doi: 10.1186/s13045-019-0805-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Shen W, Pu J, Zuo Z, Gu S, Sun J, Tan B, Wang L, Cheng J, Zuo Y. The RNA demethylase ALKBH5 promotes the progression and angiogenesis of Lung cancer by regulating the stability of the LncRNA PVT1. Cancer Cell Int. 2022;22:353. doi: 10.1186/s12935-022-02770-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Fang H, Sun Q, Zhou J, Zhang H, Song Q, Zhang H, Yu G, Guo Y, Huang C, Mou Y, et al. M(6)a methylation reader IGF2BP2 activates endothelial cells to promote angiogenesis and Metastasis of lung adenocarcinoma. Mol Cancer. 2023;22:99. doi: 10.1186/s12943-023-01791-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Zhang H, Zhou J, Li J, Wang Z, Chen Z, Lv Z, Ge L, Xie G, Deng G, Rui Y, et al. N6-Methyladenosine promotes translation of VEGFA to accelerate angiogenesis in Lung Cancer. Cancer Res. 2023;83:2208–25. doi: 10.1158/0008-5472.CAN-22-2449. [DOI] [PubMed] [Google Scholar]
- 25.Folkman J. Tumor angiogenesis: therapeutic implications. N Engl J Med. 1971;285:1182–6. doi: 10.1056/NEJM197111182852108. [DOI] [PubMed] [Google Scholar]
- 26.Edlund S, Landström M, Heldin CH, Aspenström P. Transforming growth factor-beta-induced mobilization of actin cytoskeleton requires signaling by small GTPases Cdc42 and RhoA. Mol Biol Cell. 2002;13:902–14. doi: 10.1091/mbc.01-08-0398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Panneerdoss S, Eedunuri VK, Yadav P, Timilsina S, Rajamanickam S, Viswanadhapalli S, Abdelfattah N, Onyeagucha BC, Cui X, Lai Z, et al. Cross-talk among writers, readers, and erasers of m(6)a regulates cancer growth and progression. Sci Adv. 2018;4:eaar8263. doi: 10.1126/sciadv.aar8263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kumar A, Cherukumilli M, Mahmoudpour SH, Brand K, Bandapalli OR. ShRNA-mediated knock-down of CXCL8 inhibits Tumor growth in colorectal liver Metastasis. Biochem Biophys Res Commun. 2018;500:731–7. doi: 10.1016/j.bbrc.2018.04.144. [DOI] [PubMed] [Google Scholar]
- 29.Kumaradevan S, Lee SY, Richards S, Lyle C, Zhao Q, Tapan U, Jiangliu Y, Ghumman S, Walker J, Belghasem M, et al. c-Cbl expression correlates with human Colorectal Cancer survival and its Wnt/β-Catenin suppressor function is regulated by Tyr371 phosphorylation. Am J Pathol. 2018;188:1921–33. doi: 10.1016/j.ajpath.2018.05.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Yang P, Yu D, Zhou J, Zhuang S, Jiang T. TGM2 interference regulates the angiogenesis and apoptosis of Colorectal cancer via Wnt/β-catenin pathway. Cell Cycle. 2019;18:1122–34. doi: 10.1080/15384101.2019.1609831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mosimann C, Hausmann G, Basler K. Beta-catenin hits chromatin: regulation of wnt target gene activation. Nat Rev Mol Cell Biol. 2009;10:276–86. doi: 10.1038/nrm2654. [DOI] [PubMed] [Google Scholar]
- 32.Pi J, Wang W, Ji M, Wang X, Wei X, Jin J, Liu T, Qiang J, Qi Z, Li F, et al. YTHDF1 promotes gastric carcinogenesis by Controlling translation of FZD7. Cancer Res. 2021;81:2651–65. doi: 10.1158/0008-5472.CAN-20-0066. [DOI] [PubMed] [Google Scholar]
- 33.Zhang PC, Liu X, Li MM, Ma YY, Sun HT, Tian XY, Wang Y, Liu M, Fu LS, Wang YF, et al. AT-533, a novel Hsp90 inhibitor, inhibits Breast cancer growth and HIF-1α/VEGF/VEGFR-2-mediated angiogenesis in vitro and in vivo. Biochem Pharmacol. 2020;172:113771. doi: 10.1016/j.bcp.2019.113771. [DOI] [PubMed] [Google Scholar]
- 34.Dewangan J, Srivastava S, Mishra S, Divakar A, Kumar S, Rath SK. Salinomycin inhibits Breast cancer progression via targeting HIF-1α/VEGF mediated Tumor angiogenesis in vitro and in vivo. Biochem Pharmacol. 2019;164:326–35. doi: 10.1016/j.bcp.2019.04.026. [DOI] [PubMed] [Google Scholar]
- 35.Jiang S, Gao Y, Yu QH, Li M, Cheng X, Hu SB, Song ZF, Zheng QC. P-21-activated kinase 1 contributes to Tumor angiogenesis upon photodynamic therapy via the HIF-1α/VEGF pathway. Biochem Biophys Res Commun. 2020;526:98–104. doi: 10.1016/j.bbrc.2020.03.054. [DOI] [PubMed] [Google Scholar]
- 36.Hicklin DJ, Ellis LM. Role of the vascular endothelial growth factor pathway in Tumor growth and angiogenesis. J Clin Oncol. 2005;23:1011–27. doi: 10.1200/JCO.2005.06.081. [DOI] [PubMed] [Google Scholar]
- 37.Ahn GO, Seita J, Hong BJ, Kim YE, Bok S, Lee CJ, Kim KS, Lee JC, Leeper NJ, Cooke JP, et al. Transcriptional activation of hypoxia-inducible factor-1 (HIF-1) in myeloid cells promotes angiogenesis through VEGF and S100A8. Proc Natl Acad Sci U S A. 2014;111:2698–703. doi: 10.1073/pnas.1320243111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Yang Z, Quan Y, Chen Y, Huang Y, Huang R, Yu W, Wu D, Ye M, Min Z, Yu B. Knockdown of RNA N6-methyladenosine methyltransferase METTL3 represses Warburg effect in Colorectal cancer via regulating HIF-1α. Signal Transduct Target Ther. 2021;6:89. doi: 10.1038/s41392-021-00473-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Yang Z, Wang T, Wu D, Min Z, Tan J, Yu B. RNA N6-methyladenosine reader IGF2BP3 regulates cell cycle and angiogenesis in colon Cancer. J Exp Clin Cancer Res. 2020;39:203. doi: 10.1186/s13046-020-01714-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Massagué J. How cells read TGF-beta signals. Nat Rev Mol Cell Biol. 2000;1:169–78. doi: 10.1038/35043051. [DOI] [PubMed] [Google Scholar]
- 41.Fischer AN, Fuchs E, Mikula M, Huber H, Beug H, Mikulits W. PDGF essentially links TGF-beta signaling to nuclear beta-catenin accumulation in hepatocellular carcinoma progression. Oncogene. 2007;26:3395–405. doi: 10.1038/sj.onc.1210121. [DOI] [PubMed] [Google Scholar]
- 42.Thuault S, Valcourt U, Petersen M, Manfioletti G, Heldin CH, Moustakas A. Transforming growth factor-beta employs HMGA2 to elicit epithelial-mesenchymal transition. J Cell Biol. 2006;174:175–83. doi: 10.1083/jcb.200512110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kwon S, Munroe X, Crawley SC, Lee HY, Spong S, Bradham D, Gum JR, Jr, Sleisenger MH, Kim YS. Expression of connective tissue growth factor in Pancreatic cancer cell lines. Int J Oncol. 2007;31:693–703. [PubMed] [Google Scholar]
- 44.Nanashima A, Abo T, Murakami G, Matsumoto A, Tou K, Takeshita H, Kunizaki M, Hidaka S, Sakamoto I, Hayashi H, et al. Intrahepatic cholangiocarcinoma: relationship between Tumor imaging enhancement by measuring attenuation and clinicopathologic characteristics. Abdom Imaging. 2013;38:785–92. doi: 10.1007/s00261-012-9974-3. [DOI] [PubMed] [Google Scholar]
- 45.Rong ZX, Li Z, He JJ, Liu LY, Ren XX, Gao J, Mu Y, Guan YD, Duan YM, Zhang XP, et al. Downregulation of Fat Mass and Obesity Associated (FTO) promotes the progression of Intrahepatic Cholangiocarcinoma. Front Oncol. 2019;9:369. doi: 10.3389/fonc.2019.00369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Xu Z, Zhu C, Chen C, Zong Y, Feng H, Liu D, Feng W, Zhao J, Lu A. CCL19 suppresses angiogenesis through promoting miR-206 and inhibiting Met/ERK/Elk-1/HIF-1α/VEGF-A pathway in Colorectal cancer. Cell Death Dis. 2018;9:974. doi: 10.1038/s41419-018-1010-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Chen Y, Lu Z, Qi C, Yu C, Li Y, Huan W, Wang R, Luo W, Shen D, Ding L, et al. N(6)-methyladenosine-modified TRAF1 promotes sunitinib resistance by regulating apoptosis and angiogenesis in a METTL14-dependent manner in renal cell carcinoma. Mol Cancer. 2022;21:111. doi: 10.1186/s12943-022-01549-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Bokar JA, Shambaugh ME, Polayes D, Matera AG, Rottman FM. Purification and cDNA cloning of the AdoMet-binding subunit of the human mRNA (N6-adenosine)-methyltransferase. RNA. 1997;3:1233–47. [PMC free article] [PubMed] [Google Scholar]
- 49.Liu J, Yue Y, Han D, Wang X, Fu Y, Zhang L, Jia G, Yu M, Lu Z, Deng X, et al. A METTL3-METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat Chem Biol. 2014;10:93–5. doi: 10.1038/nchembio.1432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Wu L, Wu D, Ning J, Liu W, Zhang D. Changes of N6-methyladenosine modulators promote Breast cancer progression. BMC Cancer. 2019;19:326. doi: 10.1186/s12885-019-5538-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Stevens TA, Meech R. BARX2 and estrogen receptor-alpha (ESR1) coordinately regulate the production of alternatively spliced ESR1 isoforms and control Breast cancer cell growth and invasion. Oncogene. 2006;25:5426–35. doi: 10.1038/sj.onc.1209529. [DOI] [PubMed] [Google Scholar]
- 52.Jiang X, Liu B, Nie Z, Duan L, Xiong Q, Jin Z, Yang C, Chen Y. The role of m6A modification in the biological functions and Diseases. Signal Transduct Target Ther. 2021;6:74. doi: 10.1038/s41392-020-00450-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Yue Y, Liu J, Cui X, Cao J, Luo G, Zhang Z, Cheng T, Gao M, Shu X, Ma H, et al. VIRMA mediates preferential m(6)a mRNA methylation in 3’UTR and near stop codon and associates with alternative polyadenylation. Cell Discov. 2018;4:10. doi: 10.1038/s41421-018-0019-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Wen J, Lv R, Ma H, Shen H, He C, Wang J, Jiao F, Liu H, Yang P, Tan L, et al. Zc3h13 regulates nuclear RNA m(6)a methylation and mouse embryonic stem cell Self-Renewal. Mol Cell. 2018;69:1028–1038e1026. doi: 10.1016/j.molcel.2018.02.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Warda AS, Kretschmer J, Hackert P, Lenz C, Urlaub H, Höbartner C, Sloan KE, Bohnsack MT. Human METTL16 is a N(6)-methyladenosine (m(6)A) methyltransferase that targets pre-mRNAs and various non-coding RNAs. EMBO Rep. 2017;18:2004–14. doi: 10.15252/embr.201744940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Jin D, Guo J, Wu Y, Du J, Yang L, Wang X, Di W, Hu B, An J, Kong L, et al. M(6)a mRNA methylation initiated by METTL3 directly promotes YAP translation and increases YAP activity by regulating the MALAT1-miR-1914-3p-YAP axis to induce NSCLC drug resistance and Metastasis. J Hematol Oncol. 2021;14:32. doi: 10.1186/s13045-021-01048-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Qiao K, Liu Y, Xu Z, Zhang H, Zhang H, Zhang C, Chang Z, Lu X, Li Z, Luo C, et al. RNA m6A methylation promotes the formation of vasculogenic mimicry in hepatocellular carcinoma via Hippo pathway. Angiogenesis. 2021;24:83–96. doi: 10.1007/s10456-020-09744-8. [DOI] [PubMed] [Google Scholar]
- 58.Wang Q, Chen C, Ding Q, Zhao Y, Wang Z, Chen J, Jiang Z, Zhang Y, Xu G, Zhang J, et al. METTL3-mediated m(6)a modification of HDGF mRNA promotes gastric cancer progression and has prognostic significance. Gut. 2020;69:1193–205. doi: 10.1136/gutjnl-2019-319639. [DOI] [PubMed] [Google Scholar]
- 59.Lin S, Choe J, Du P, Triboulet R, Gregory RI. The m(6)a methyltransferase METTL3 promotes translation in Human Cancer cells. Mol Cell. 2016;62:335–45. doi: 10.1016/j.molcel.2016.03.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Cai X, Wang X, Cao C, Gao Y, Zhang S, Yang Z, Liu Y, Zhang X, Zhang W, Ye L. HBXIP-elevated methyltransferase METTL3 promotes the progression of Breast cancer via inhibiting Tumor suppressor let-7 g. Cancer Lett. 2018;415:11–9. doi: 10.1016/j.canlet.2017.11.018. [DOI] [PubMed] [Google Scholar]
- 61.Ma JZ, Yang F, Zhou CC, Liu F, Yuan JH, Wang F, Wang TT, Xu QG, Zhou WP, Sun SH. METTL14 suppresses the metastatic potential of hepatocellular carcinoma by modulating N(6) -methyladenosine-dependent primary MicroRNA processing. Hepatology. 2017;65:529–43. doi: 10.1002/hep.28885. [DOI] [PubMed] [Google Scholar]
- 62.Edilova MI, Abdul-Sater AA, Watts TH. TRAF1 signaling in Human Health and Disease. Front Immunol. 2018;9:2969. doi: 10.3389/fimmu.2018.02969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Kim CM, Jeong JH, Son YJ, Choi JH, Kim S, Park HH. Molecular basis for TANK recognition by TRAF1 revealed by the crystal structure of TRAF1/TANK complex. FEBS Lett. 2017;591:810–21. doi: 10.1002/1873-3468.12584. [DOI] [PubMed] [Google Scholar]
- 64.Shi H, Wei J, He C. Where, when, and how: context-dependent functions of RNA methylation writers, readers, and Erasers. Mol Cell. 2019;74:640–50. doi: 10.1016/j.molcel.2019.04.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Ueda Y, Ooshio I, Fusamae Y, Kitae K, Kawaguchi M, Jingushi K, Hase H, Harada K, Hirata K, Tsujikawa K. AlkB homolog 3-mediated tRNA demethylation promotes protein synthesis in cancer cells. Sci Rep. 2017;7:42271. doi: 10.1038/srep42271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Haussmann IU, Bodi Z, Sanchez-Moran E, Mongan NP, Archer N, Fray RG, Soller M. M(6)a potentiates Sxl alternative pre-mRNA splicing for robust Drosophila sex determination. Nature. 2016;540:301–4. doi: 10.1038/nature20577. [DOI] [PubMed] [Google Scholar]
- 67.Liu N, Dai Q, Zheng G, He C, Parisien M, Pan T. N(6)-methyladenosine-dependent RNA structural switches regulate RNA-protein interactions. Nature. 2015;518:560–4. doi: 10.1038/nature14234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Huang H, Weng H, Sun W, Qin X, Shi H, Wu H, Zhao BS, Mesquita A, Liu C, Yuan CL, et al. Recognition of RNA N(6)-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat Cell Biol. 2018;20:285–95. doi: 10.1038/s41556-018-0045-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Jiang L, Li Y, He Y, Wei D, Yan L, Wen H. Knockdown of m6A reader IGF2BP3 inhibited Hypoxia-Induced Cell Migration and Angiogenesis by regulating Hypoxia Inducible Factor-1α in Stomach Cancer. Front Oncol. 2021;11:711207. doi: 10.3389/fonc.2021.711207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Hou J, Zhang H, Liu J, Zhao Z, Wang J, Lu Z, Hu B, Zhou J, Zhao Z, Feng M, et al. YTHDF2 reduction fuels inflammation and vascular abnormalization in hepatocellular carcinoma. Mol Cancer. 2019;18:163. doi: 10.1186/s12943-019-1082-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Chang G, Shi L, Ye Y, Shi H, Zeng L, Tiwary S, Huse JT, Huo L, Ma L, Ma Y, et al. YTHDF3 induces the translation of m(6)A-Enriched gene transcripts to promote Breast Cancer brain Metastasis. Cancer Cell. 2020;38:857–871e857. doi: 10.1016/j.ccell.2020.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Wang G, Dai Y, Li K, Cheng M, Xiong G, Wang X, Chen S, Chen Z, Chen J, Xu X, et al. Deficiency of Mettl3 in Bladder Cancer stem cells inhibits Bladder Cancer Progression and Angiogenesis. Front Cell Dev Biol. 2021;9:627706. doi: 10.3389/fcell.2021.627706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Green NH, Galvan DL, Badal SS, Chang BH, LeBleu VS, Long J, Jonasch E, Danesh FR. MTHFD2 links RNA methylation to metabolic reprogramming in renal cell carcinoma. Oncogene. 2019;38:6211–25. doi: 10.1038/s41388-019-0869-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Ma YS, Shi BW, Guo JH, Liu JB, Yang XL, Xin R, Shi Y, Zhang DD, Lu GX, Jia CY, et al. microRNA-320b suppresses HNF4G and IGF2BP2 expression to inhibit angiogenesis and Tumor growth of Lung cancer. Carcinogenesis. 2021;42:762–71. doi: 10.1093/carcin/bgab023. [DOI] [PubMed] [Google Scholar]
- 75.Suvasini R, Shruti B, Thota B, Shinde SV, Friedmann-Morvinski D, Nawaz Z, Prasanna KV, Thennarasu K, Hegde AS, Arivazhagan A, et al. Insulin growth factor-2 binding protein 3 (IGF2BP3) is a glioblastoma-specific marker that activates phosphatidylinositol 3-kinase/mitogen-activated protein kinase (PI3K/MAPK) pathways by modulating IGF-2. J Biol Chem. 2011;286:25882–90. doi: 10.1074/jbc.M110.178012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Richters A, Aben KKH, Kiemeney L. The global burden of urinary Bladder cancer: an update. World J Urol. 2020;38:1895–904. doi: 10.1007/s00345-019-02984-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Mazzieri R, Pucci F, Moi D, Zonari E, Ranghetti A, Berti A, Politi LS, Gentner B, Brown JL, Naldini L, De Palma M. Targeting the ANG2/TIE2 axis inhibits Tumor growth and Metastasis by impairing angiogenesis and disabling rebounds of proangiogenic myeloid cells. Cancer Cell. 2011;19:512–26. doi: 10.1016/j.ccr.2011.02.005. [DOI] [PubMed] [Google Scholar]
- 78.Goel HL, Mercurio AM. VEGF targets the tumour cell. Nat Rev Cancer. 2013;13:871–82. doi: 10.1038/nrc3627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Smyth EC, Nilsson M, Grabsch HI, van Grieken NC, Lordick F. Gastric cancer. Lancet. 2020;396:635–48. doi: 10.1016/S0140-6736(20)31288-5. [DOI] [PubMed] [Google Scholar]
- 80.Schito L. Hypoxia-dependent angiogenesis and Lymphangiogenesis in Cancer. Adv Exp Med Biol. 2019;1136:71–85. doi: 10.1007/978-3-030-12734-3_5. [DOI] [PubMed] [Google Scholar]
- 81.Vaupel P, Multhoff G. Fatal Alliance of Hypoxia-/HIF-1α-Driven microenvironmental traits promoting Cancer Progression. Adv Exp Med Biol. 2020;1232:169–76. doi: 10.1007/978-3-030-34461-0_21. [DOI] [PubMed] [Google Scholar]
- 82.Xueqing H, Jun Z, Yueqiang J, Xin L, Liya H, Yuanyuan F, Yuting Z, Hao Z, Hua W, Jian L, Tiejun Y. IGF2BP3 may contributes to lung tumorigenesis by regulating the alternative splicing of PKM. Front Bioeng Biotechnol. 2020;8:679. doi: 10.3389/fbioe.2020.00679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Park H, Kim H, Ha E, Yoon S, Kim MJ, Hong M, Leem KH, Hong SJ, Yang J, Chung JH. Panax ginseng increases hypoxia-induced down-regulated cellular response related genes in human neuroblastoma cells, SK-N-MC. Neurol Res. 2007;29(Suppl 1):78–87. doi: 10.1179/016164107X172338. [DOI] [PubMed] [Google Scholar]
- 84.Masoud GN, Li W. HIF-1α pathway: role, regulation and intervention for cancer therapy. Acta Pharm Sin B. 2015;5:378–89. doi: 10.1016/j.apsb.2015.05.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Garg N, Kumar P, Gadhave K, Giri R. The dark proteome of cancer: intrinsic disorderedness and functionality of HIF-1α along with its interacting proteins. Prog Mol Biol Transl Sci. 2019;166:371–403. doi: 10.1016/bs.pmbts.2019.05.006. [DOI] [PubMed] [Google Scholar]
- 86.Ciccone V, Terzuoli E, Donnini S, Giachetti A, Morbidelli L, Ziche M. Stemness marker ALDH1A1 promotes Tumor angiogenesis via retinoic acid/HIF-1α/VEGF signalling in MCF-7 Breast cancer cells. J Exp Clin Cancer Res. 2018;37:311. doi: 10.1186/s13046-018-0975-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Gupta R, Bhatt LK, Johnston TP, Prabhavalkar KS. Colon cancer stem cells: potential target for the treatment of Colorectal cancer. Cancer Biol Ther. 2019;20:1068–82. doi: 10.1080/15384047.2019.1599660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Llovet JM, Kelley RK, Villanueva A, Singal AG, Pikarsky E, Roayaie S, Lencioni R, Koike K, Zucman-Rossi J, Finn RS. Hepatocellular carcinoma. Nat Rev Dis Primers. 2021;7:6. doi: 10.1038/s41572-020-00240-3. [DOI] [PubMed] [Google Scholar]
- 89.Maniotis AJ, Folberg R, Hess A, Seftor EA, Gardner LM, Pe’er J, Trent JM, Meltzer PS, Hendrix MJ. Vascular channel formation by human Melanoma cells in vivo and in vitro: vasculogenic mimicry. Am J Pathol. 1999;155:739–52. doi: 10.1016/S0002-9440(10)65173-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Sun H, Yao N, Cheng S, Li L, Liu S, Yang Z, Shang G, Zhang D, Yao Z. Cancer stem-like cells directly participate in vasculogenic mimicry channels in triple-negative Breast cancer. Cancer Biol Med. 2019;16:299–311. doi: 10.20892/j.issn.2095-3941.2018.0209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Luo Q, Wang J, Zhao W, Peng Z, Liu X, Li B, Zhang H, Shan B, Zhang C, Duan C. Vasculogenic mimicry in carcinogenesis and clinical applications. J Hematol Oncol. 2020;13:19. doi: 10.1186/s13045-020-00858-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Bai XL, Zhang Q, Ye LY, Liang F, Sun X, Chen Y, Hu QD, Fu QH, Su W, Chen Z, et al. Myocyte enhancer factor 2 C regulation of hepatocellular carcinoma via vascular endothelial growth factor and Wnt/β-catenin signaling. Oncogene. 2015;34:4089–97. doi: 10.1038/onc.2014.337. [DOI] [PubMed] [Google Scholar]
- 93.Sun Z, Xue S, Zhang M, Xu H, Hu X, Chen S, Liu Y, Guo M, Cui H. Aberrant NSUN2-mediated m(5)C modification of H19 lncRNA is associated with poor differentiation of hepatocellular carcinoma. Oncogene. 2020;39:6906–19. doi: 10.1038/s41388-020-01475-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Toumazis I, Bastani M, Han SS, Plevritis SK. Risk-based Lung cancer screening: a systematic review. Lung Cancer. 2020;147:154–86. doi: 10.1016/j.lungcan.2020.07.007. [DOI] [PubMed] [Google Scholar]
- 95.Liang Y, Zhang H, Song X, Yang Q. Metastatic heterogeneity of Breast cancer: molecular mechanism and potential therapeutic targets. Semin Cancer Biol. 2020;60:14–27. doi: 10.1016/j.semcancer.2019.08.012. [DOI] [PubMed] [Google Scholar]
- 96.Xu Y, Ye S, Zhang N, Zheng S, Liu H, Zhou K, Wang L, Cao Y, Sun P, Wang T. The FTO/miR-181b-3p/ARL5B signaling pathway regulates cell migration and invasion in Breast cancer. Cancer Commun (Lond) 2020;40:484–500. doi: 10.1002/cac2.12075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Broekman ML, Maas SLN, Abels ER, Mempel TR, Krichevsky AM, Breakefield XO. Multidimensional communication in the microenvirons of glioblastoma. Nat Rev Neurol. 2018;14:482–95. doi: 10.1038/s41582-018-0025-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Hsieh JJ, Purdue MP, Signoretti S, Swanton C, Albiges L, Schmidinger M, Heng DY, Larkin J, Ficarra V. Renal cell carcinoma. Nat Rev Dis Primers. 2017;3:17009. doi: 10.1038/nrdp.2017.9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Cai J, Chen H, Lu M, Zhang Y, Lu B, You L, Zhang T, Dai M, Zhao Y. Advances in the epidemiology of Pancreatic cancer: Trends, risk factors, screening, and prognosis. Cancer Lett. 2021;520:1–11. doi: 10.1016/j.canlet.2021.06.027. [DOI] [PubMed] [Google Scholar]
- 100.Konishi N, Nakamura M, Ishida E, Shimada K, Mitsui E, Yoshikawa R, Yamamoto H, Tsujikawa K. High expression of a new marker PCA-1 in human prostate carcinoma. Clin Cancer Res. 2005;11:5090–7. doi: 10.1158/1078-0432.CCR-05-0195. [DOI] [PubMed] [Google Scholar]
- 101.Yamato I, Sho M, Shimada K, Hotta K, Ueda Y, Yasuda S, Shigi N, Konishi N, Tsujikawa K, Nakajima Y. PCA-1/ALKBH3 contributes to Pancreatic cancer by supporting apoptotic resistance and angiogenesis. Cancer Res. 2012;72:4829–39. doi: 10.1158/0008-5472.CAN-12-0328. [DOI] [PubMed] [Google Scholar]
- 102.Lu TX, Rothenberg ME. MicroRNA. J Allergy Clin Immunol. 2018;141:1202–7. doi: 10.1016/j.jaci.2017.08.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Alarcón CR, Lee H, Goodarzi H, Halberg N, Tavazoie SF. N6-methyladenosine marks primary microRNAs for processing. Nature. 2015;519:482–5. doi: 10.1038/nature14281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Alarcón CR, Goodarzi H, Lee H, Liu X, Tavazoie S, Tavazoie SF. HNRNPA2B1 is a mediator of m(6)A-Dependent Nuclear RNA Processing events. Cell. 2015;162:1299–308. doi: 10.1016/j.cell.2015.08.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Wang X, Chen Z, Xu J, Tang S, An N, Jiang L, Zhang Y, Zhang S, Zhang Q, Shen Y, et al. SLC1A1-mediated cellular and mitochondrial influx of R-2-hydroxyglutarate in vascular endothelial cells promotes Tumor angiogenesis in IDH1-mutant solid tumors. Cell Res. 2022;32:638–58. doi: 10.1038/s41422-022-00650-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Chamorro-Jorganes A, Sweaad WK, Katare R, Besnier M, Anwar M, Beazley-Long N, Sala-Newby G, Ruiz-Polo I, Chandrasekera D, Ritchie AA, et al. METTL3 regulates angiogenesis by modulating let-7e-5p and miRNA-18a-5p expression in endothelial cells. Arterioscler Thromb Vasc Biol. 2021;41:e325–37. doi: 10.1161/ATVBAHA.121.316180. [DOI] [PubMed] [Google Scholar]
- 107.Yin X, Zhang K, Wang J, Zhou X, Zhang C, Song X, Wu Z, Du J, Chen Q, Zhang S, Deng W. RNA polymerase I subunit 12 plays opposite roles in cell proliferation and migration. Biochem Biophys Res Commun. 2021;560:112–8. doi: 10.1016/j.bbrc.2021.04.091. [DOI] [PubMed] [Google Scholar]
- 108.Guo L, Wen J, Han J, Jiang J, Xie S, Feng X, Wei B, Dai J, Zhang K, Qi J, et al. Expression quantitative trait loci in long non-coding RNA ZNRD1-AS1 influence Cervical cancer development. Am J Cancer Res. 2015;5:2301–7. [PMC free article] [PubMed] [Google Scholar]
- 109.Wang J, Tan L, Yu X, Cao X, Jia B, Chen R, Li J. lncRNA ZNRD1-AS1 promotes malignant lung cell proliferation, migration, and angiogenesis via the miR-942/TNS1 axis and is positively regulated by the m(6)a reader YTHDC2. Mol Cancer. 2022;21:229. doi: 10.1186/s12943-022-01705-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Liu H, Jiang Y, Lu J, Peng C, Ling Z, Chen Y, Chen D, Tong R, Zheng S, Wu J. M(6)A-modification regulated circ-CCT3 acts as the sponge of miR-378a-3p to promote hepatocellular carcinoma progression. Epigenetics. 2023;18:2204772. doi: 10.1080/15592294.2023.2204772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Wang H, Deng Q, Lv Z, Ling Y, Hou X, Chen Z, Dinglin X, Ma S, Li D, Wu Y, et al. N6-methyladenosine induced mir-143-3p promotes the brain Metastasis of Lung cancer via regulation of VASH1. Mol Cancer. 2019;18:181. doi: 10.1186/s12943-019-1108-x. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 112.Nieuwenhuis J, Adamopoulos A, Bleijerveld OB, Mazouzi A, Stickel E, Celie P, Altelaar M, Knipscheer P, Perrakis A, Blomen VA, Brummelkamp TR. Vasohibins encode tubulin detyrosinating activity. Science. 2017;358:1453–6. doi: 10.1126/science.aao5676. [DOI] [PubMed] [Google Scholar]
- 113.Guo Y, Guo Y, Chen C, Fan D, Wu X, Zhao L, Shao B, Sun Z, Ji Z. Circ3823 contributes to growth, Metastasis and angiogenesis of Colorectal cancer: involvement of miR-30c-5p/TCF7 axis. Mol Cancer. 2021;20:93. doi: 10.1186/s12943-021-01372-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Dupont S, Morsut L, Aragona M, Enzo E, Giulitti S, Cordenonsi M, Zanconato F, Le Digabel J, Forcato M, Bicciato S, et al. Role of YAP/TAZ in mechanotransduction. Nature. 2011;474:179–83. doi: 10.1038/nature10137. [DOI] [PubMed] [Google Scholar]
- 115.Shao DD, Xue W, Krall EB, Bhutkar A, Piccioni F, Wang X, Schinzel AC, Sood S, Rosenbluh J, Kim JW, et al. KRAS and YAP1 converge to regulate EMT and Tumor survival. Cell. 2014;158:171–84. doi: 10.1016/j.cell.2014.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Kim DH, Kim SH, Lee OJ, Huang SM, Kwon JL, Kim JM, Kim JY, Seong IO, Song KS, Kim KH. Differential expression of yes-associated protein and phosphorylated yes-associated protein is correlated with expression of Ki-67 and phospho-ERK in colorectal adenocarcinoma. Histol Histopathol. 2013;28:1483–90. doi: 10.14670/HH-28.1483. [DOI] [PubMed] [Google Scholar]
- 117.Cordenonsi M, Zanconato F, Azzolin L, Forcato M, Rosato A, Frasson C, Inui M, Montagner M, Parenti AR, Poletti A, et al. The Hippo transducer TAZ confers cancer stem cell-related traits on Breast cancer cells. Cell. 2011;147:759–72. doi: 10.1016/j.cell.2011.09.048. [DOI] [PubMed] [Google Scholar]
- 118.Zanconato F, Cordenonsi M, Piccolo S. YAP/TAZ at the roots of Cancer. Cancer Cell. 2016;29:783–803. doi: 10.1016/j.ccell.2016.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Cheng H, Zhang Z, Rodriguez-Barrueco R, Borczuk A, Liu H, Yu J, Silva JM, Cheng SK, Perez-Soler R, Halmos B. Functional genomics screen identifies YAP1 as a key determinant to enhance treatment sensitivity in Lung cancer cells. Oncotarget. 2016;7:28976–88. doi: 10.18632/oncotarget.6721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Diep CH, Zucker KM, Hostetter G, Watanabe A, Hu C, Munoz RM, Von Hoff DD, Han H. Down-regulation of Yes Associated protein 1 expression reduces cell proliferation and clonogenicity of Pancreatic cancer cells. PLoS ONE. 2012;7:e32783. doi: 10.1371/journal.pone.0032783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Han SX, Bai E, Jin GH, He CC, Guo XJ, Wang LJ, Li M, Ying X, Zhu Q. Expression and clinical significance of YAP, TAZ, and AREG in hepatocellular carcinoma. J Immunol Res. 2014;2014:261365. doi: 10.1155/2014/261365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Da CL, Xin Y, Zhao J, Luo XD. Significance and relationship between yes-associated protein and survivin expression in gastric carcinoma and precancerous lesions. World J Gastroenterol. 2009;15:4055–61. doi: 10.3748/wjg.15.4055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Chen Y, Ling Z, Cai X, Xu Y, Lv Z, Man D, Ge J, Yu C, Zhang D, Zhang Y, et al. Activation of YAP1 by N6-Methyladenosine-modified circCPSF6 drives malignancy in Hepatocellular Carcinoma. Cancer Res. 2022;82:599–614. doi: 10.1158/0008-5472.CAN-21-1628. [DOI] [PubMed] [Google Scholar]
- 124.Jin D, Guo J, Wu Y, Yang L, Wang X, Du J, Dai J, Chen W, Gong K, Miao S, et al. M(6)a demethylase ALKBH5 inhibits Tumor growth and Metastasis by reducing YTHDFs-mediated YAP expression and inhibiting miR-107/LATS2-mediated YAP activity in NSCLC. Mol Cancer. 2020;19:40. doi: 10.1186/s12943-020-01161-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Han Z, Wang X, Xu Z, Cao Y, Gong R, Yu Y, Yu Y, Guo X, Liu S, Yu M et al. ALKBH5 regulates cardiomyocyte proliferation and heart regeneration by demethylating the mRNA of YTHDF1. Theranostics 2021, 11:3000–3016. [DOI] [PMC free article] [PubMed]
- 126.Yuan Y, Yan G, He M, Lei H, Li L, Wang Y, He X, Li G, Wang Q, Gao Y, et al. ALKBH5 suppresses Tumor progression via an m(6)A-dependent epigenetic silencing of pre-miR-181b-1/YAP signaling axis in osteosarcoma. Cell Death Dis. 2021;12:60. doi: 10.1038/s41419-020-03315-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Shen Y, Wang X, Liu Y, Singhal M, Gürkaşlar C, Valls AF, Lei Y, Hu W, Schermann G, Adler H, et al. STAT3-YAP/TAZ signaling in endothelial cells promotes Tumor angiogenesis. Sci Signal. 2021;14:eabj8393. doi: 10.1126/scisignal.abj8393. [DOI] [PubMed] [Google Scholar]
- 128.Xu S, Zhang H, Chong Y, Guan B, Guo P. YAP promotes VEGFA expression and Tumor Angiogenesis though Gli2 in human renal cell carcinoma. Arch Med Res. 2019;50:225–33. doi: 10.1016/j.arcmed.2019.08.010. [DOI] [PubMed] [Google Scholar]
- 129.Sweeney MD, Sagare AP, Zlokovic BV. Blood-brain barrier breakdown in Alzheimer Disease and other neurodegenerative disorders. Nat Rev Neurol. 2018;14:133–50. doi: 10.1038/nrneurol.2017.188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Quail DF, Joyce JA. The Microenvironmental Landscape of Brain tumors. Cancer Cell. 2017;31:326–41. doi: 10.1016/j.ccell.2017.02.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Liebner S, Dijkhuizen RM, Reiss Y, Plate KH, Agalliu D, Constantin G. Functional morphology of the blood-brain barrier in health and Disease. Acta Neuropathol. 2018;135:311–36. doi: 10.1007/s00401-018-1815-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Guo C, Liu J, Zhou Q, Song J, Zhang Z, Li Z, Wang G, Yuan W, Sun Z. Exosomal noncoding RNAs and Tumor Drug Resistance. Cancer Res. 2020;80:4307–13. doi: 10.1158/0008-5472.CAN-20-0032. [DOI] [PubMed] [Google Scholar]
- 133.Li B, Jiang J, Assaraf YG, Xiao H, Chen ZS, Huang C. Surmounting cancer drug resistance: new insights from the perspective of N(6)-methyladenosine RNA modification. Drug Resist Updat. 2020;53:100720. doi: 10.1016/j.drup.2020.100720. [DOI] [PubMed] [Google Scholar]
- 134.Liu L, Li H, Hu D, Wang Y, Shao W, Zhong J, Yang S, Liu J, Zhang J. Insights into N6-methyladenosine and programmed cell death in cancer. Mol Cancer. 2022;21:32. doi: 10.1186/s12943-022-01508-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Qin S, Mao Y, Wang H, Duan Y, Zhao L. The interplay between m6A modification and non-coding RNA in cancer stemness modulation: mechanisms, signaling pathways, and clinical implications. Int J Biol Sci. 2021;17:2718–36. doi: 10.7150/ijbs.60641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Qu F, Tsegay PS, Liu Y. N(6)-Methyladenosine, DNA repair, and Genome Stability. Front Mol Biosci. 2021;8:645823. doi: 10.3389/fmolb.2021.645823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Liu Z, Zou H, Dang Q, Xu H, Liu L, Zhang Y, Lv J, Li H, Zhou Z, Han X. Biological and pharmacological roles of m(6)a modifications in cancer drug resistance. Mol Cancer. 2022;21:220. doi: 10.1186/s12943-022-01680-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Yusuf I, Zhu X, Kharas MG, Chen J, Fruman DA. Optimal B-cell proliferation requires phosphoinositide 3-kinase-dependent inactivation of FOXO transcription factors. Blood. 2004;104:784–7. doi: 10.1182/blood-2003-09-3071. [DOI] [PubMed] [Google Scholar]
- 139.Jonsson H, Allen P, Peng SL. Inflammatory arthritis requires Foxo3a to prevent Fas ligand-induced neutrophil apoptosis. Nat Med. 2005;11:666–71. doi: 10.1038/nm1248. [DOI] [PubMed] [Google Scholar]
- 140.Mammucari C, Milan G, Romanello V, Masiero E, Rudolf R, Del Piccolo P, Burden SJ, Di Lisi R, Sandri C, Zhao J, et al. FoxO3 controls autophagy in skeletal muscle in vivo. Cell Metab. 2007;6:458–71. doi: 10.1016/j.cmet.2007.11.001. [DOI] [PubMed] [Google Scholar]
- 141.Cheng AL, Kang YK, Chen Z, Tsao CJ, Qin S, Kim JS, Luo R, Feng J, Ye S, Yang TS, et al. Efficacy and safety of sorafenib in patients in the Asia-Pacific region with advanced hepatocellular carcinoma: a phase III randomised, double-blind, placebo-controlled trial. Lancet Oncol. 2009;10:25–34. doi: 10.1016/S1470-2045(08)70285-7. [DOI] [PubMed] [Google Scholar]
- 142.Lin Z, Niu Y, Wan A, Chen D, Liang H, Chen X, Sun L, Zhan S, Chen L, Cheng C, et al. RNA m(6) a methylation regulates sorafenib resistance in Liver cancer through FOXO3-mediated autophagy. Embo j. 2020;39:e103181. doi: 10.15252/embj.2019103181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Wang Y, Liu J, Ma J, Sun T, Zhou Q, Wang W, Wang G, Wu P, Wang H, Jiang L, et al. Exosomal circRNAs: biogenesis, effect and application in human Diseases. Mol Cancer. 2019;18:116. doi: 10.1186/s12943-019-1041-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Cunningham D, Humblet Y, Siena S, Khayat D, Bleiberg H, Santoro A, Bets D, Mueser M, Harstrick A, Verslype C, et al. Cetuximab monotherapy and cetuximab plus irinotecan in irinotecan-refractory metastatic Colorectal cancer. N Engl J Med. 2004;351:337–45. doi: 10.1056/NEJMoa033025. [DOI] [PubMed] [Google Scholar]
- 145.Liu H, Li D, Sun L, Qin H, Fan A, Meng L, Graves-Deal R, Glass SE, Franklin JL, Liu Q, et al. Interaction of lncRNA MIR100HG with hnRNPA2B1 facilitates m(6)A-dependent stabilization of TCF7L2 mRNA and Colorectal cancer progression. Mol Cancer. 2022;21:74. doi: 10.1186/s12943-022-01555-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Bhattarai PY, Kim G, Poudel M, Lim SC, Choi HS. METTL3 induces PLX4032 resistance in Melanoma by promoting m(6)A-dependent EGFR translation. Cancer Lett. 2021;522:44–56. doi: 10.1016/j.canlet.2021.09.015. [DOI] [PubMed] [Google Scholar]
- 147.Huang X, Zhu L, Wang L, Huang W, Tan L, Liu H, Huo J, Su T, Zhang M, Kuang M, et al. YTHDF1 promotes intrahepatic cholangiocarcinoma progression via regulating EGFR mRNA translation. J Gastroenterol Hepatol. 2022;37:1156–68. doi: 10.1111/jgh.15816. [DOI] [PubMed] [Google Scholar]
- 148.Zhong L, Liao D, Zhang M, Zeng C, Li X, Zhang R, Ma H, Kang T. YTHDF2 suppresses cell proliferation and growth via destabilizing the EGFR mRNA in hepatocellular carcinoma. Cancer Lett. 2019;442:252–61. doi: 10.1016/j.canlet.2018.11.006. [DOI] [PubMed] [Google Scholar]
- 149.Molina AM, Lin X, Korytowsky B, Matczak E, Lechuga MJ, Wiltshire R, Motzer RJ. Sunitinib objective response in metastatic renal cell carcinoma: analysis of 1059 patients treated on clinical trials. Eur J Cancer. 2014;50:351–8. doi: 10.1016/j.ejca.2013.08.021. [DOI] [PubMed] [Google Scholar]
- 150.Chiang K, Zielinska AE, Shaaban AM, Sanchez-Bailon MP, Jarrold J, Clarke TL, Zhang J, Francis A, Jones LJ, Smith S, et al. PRMT5 is a critical Regulator of Breast Cancer stem cell function via histone methylation and FOXP1 expression. Cell Rep. 2017;21:3498–513. doi: 10.1016/j.celrep.2017.11.096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Wang Z, Kong J, Wu Y, Zhang J, Wang T, Li N, Fan J, Wang H, Zhang J, Ling R. PRMT5 determines the sensitivity to chemotherapeutics by governing stemness in Breast cancer. Breast Cancer Res Treat. 2018;168:531–42. doi: 10.1007/s10549-017-4597-6. [DOI] [PubMed] [Google Scholar]
- 152.Wu Y, Wang Z, Han L, Guo Z, Yan B, Guo L, Zhao H, Wei M, Hou N, Ye J, et al. PRMT5 regulates RNA m6A demethylation for doxorubicin sensitivity in Breast cancer. Mol Ther. 2022;30:2603–17. doi: 10.1016/j.ymthe.2022.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Zhao Z, Cai Q, Zhang P, He B, Peng X, Tu G, Peng W, Wang L, Yu F, Wang X. N6-Methyladenosine RNA methylation Regulator-Related Alternative Splicing (AS) Gene signature predicts Non-small Cell Lung Cancer Prognosis. Front Mol Biosci. 2021;8:657087. doi: 10.3389/fmolb.2021.657087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Yue B, Song C, Yang L, Cui R, Cheng X, Zhang Z, Zhao G. METTL3-mediated N6-methyladenosine modification is critical for epithelial-mesenchymal transition and Metastasis of gastric cancer. Mol Cancer. 2019;18:142. doi: 10.1186/s12943-019-1065-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Li T, Hu PS, Zuo Z, Lin JF, Li X, Wu QN, Chen ZH, Zeng ZL, Wang F, Zheng J, et al. METTL3 facilitates Tumor progression via an m(6)A-IGF2BP2-dependent mechanism in colorectal carcinoma. Mol Cancer. 2019;18:112. doi: 10.1186/s12943-019-1038-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Li J, Xie H, Ying Y, Chen H, Yan H, He L, Xu M, Xu X, Liang Z, Liu B, et al. YTHDF2 mediates the mRNA degradation of the Tumor suppressors to induce AKT phosphorylation in N6-methyladenosine-dependent way in Prostate cancer. Mol Cancer. 2020;19:152. doi: 10.1186/s12943-020-01267-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Guo W, Tan F, Huai Q, Wang Z, Shao F, Zhang G, Yang Z, Li R, Xue Q, Gao S, He J. Comprehensive Analysis of PD-L1 expression, Immune infiltrates, and m6A RNA methylation regulators in esophageal squamous cell carcinoma. Front Immunol. 2021;12:669750. doi: 10.3389/fimmu.2021.669750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Findeis-Hosey JJ, Xu H. The use of insulin like-growth factor II messenger RNA binding protein-3 in diagnostic pathology. Hum Pathol. 2011;42:303–14. doi: 10.1016/j.humpath.2010.06.003. [DOI] [PubMed] [Google Scholar]
- 159.Hoffmann NE, Sheinin Y, Lohse CM, Parker AS, Leibovich BC, Jiang Z, Kwon ED. External validation of IMP3 expression as an Independent prognostic marker for metastatic progression and death for patients with clear cell renal cell carcinoma. Cancer. 2008;112:1471–9. doi: 10.1002/cncr.23296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Chen HM, Lin CC, Chen WS, Jiang JK, Yang SH, Chang SC, Ho CL, Yang CC, Huang SC, Chao Y et al. Insulin-like growth factor 2 mRNA-Binding protein 1 (IGF2BP1) is a Prognostic Biomarker and Associated with Chemotherapy responsiveness in Colorectal Cancer. Int J Mol Sci 2021, 22. [DOI] [PMC free article] [PubMed]
- 161.Bi R, Shen X, Zhang W, Cheng Y, Feng Z, Cai X, Yang W. Clear cell carcinomas of the ovary: a mono-institutional study of 73 cases in China with an analysis of the prognostic significance of clinicopathological parameters and IMP3 expression. Diagn Pathol. 2016;11:17. doi: 10.1186/s13000-016-0467-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Walter O, Prasad M, Lu S, Quinlan RM, Edmiston KL, Khan A. IMP3 is a novel biomarker for triple negative invasive mammary carcinoma associated with a more aggressive phenotype. Hum Pathol. 2009;40:1528–33. doi: 10.1016/j.humpath.2009.05.005. [DOI] [PubMed] [Google Scholar]
- 163.Schaeffer DF, Owen DR, Lim HJ, Buczkowski AK, Chung SW, Scudamore CH, Huntsman DG, Ng SS, Owen DA. Insulin-like growth factor 2 mRNA binding protein 3 (IGF2BP3) overexpression in pancreatic ductal adenocarcinoma correlates with poor survival. BMC Cancer. 2010;10:59. doi: 10.1186/1471-2407-10-59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Lu D, Yang X, Jiang NY, Woda BA, Liu Q, Dresser K, Mercurio AM, Rock KL, Jiang Z. IMP3, a new biomarker to predict progression of cervical intraepithelial neoplasia into invasive cancer. Am J Surg Pathol. 2011;35:1638–45. doi: 10.1097/PAS.0b013e31823272d4. [DOI] [PubMed] [Google Scholar]
- 165.Xu H. IMP3: a diagnostic and prognostic biomarker in malignant Melanoma. Expert Rev Mol Diagn. 2008;8:557–8. doi: 10.1586/14737159.8.5.557. [DOI] [PubMed] [Google Scholar]
- 166.Köbel M, Xu H, Bourne PA, Spaulding BO, Shih Ie M, Mao TL, Soslow RA, Ewanowich CA, Kalloger SE, Mehl E, et al. IGF2BP3 (IMP3) expression is a marker of unfavorable prognosis in ovarian carcinoma of clear cell subtype. Mod Pathol. 2009;22:469–75. doi: 10.1038/modpathol.2008.206. [DOI] [PubMed] [Google Scholar]
- 167.Sun C, Zheng X, Sun Y, Yu J, Sheng M, Yan S, Zhu Q, Lan Q. Identification of IGF2BP3 as an adverse prognostic biomarker of Gliomas. Front Genet. 2021;12:743738. doi: 10.3389/fgene.2021.743738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Shi R, Yu X, Wang Y, Sun J, Sun Q, Xia W, Dong G, Wang A, Gao Z, Jiang F, Xu L. Expression profile, clinical significance, and biological function of insulin-like growth factor 2 messenger RNA-binding proteins in non-small cell Lung cancer. Tumour Biol. 2017;39:1010428317695928. doi: 10.1177/1010428317695928. [DOI] [PubMed] [Google Scholar]
- 169.Lin CY, Chen ST, Jeng YM, Yeh CC, Chou HY, Deng YT, Chang CC, Kuo MY. Insulin-like growth factor II mRNA-binding protein 3 expression promotes Tumor formation and invasion and predicts poor prognosis in oral squamous cell carcinoma. J Oral Pathol Med. 2011;40:699–705. doi: 10.1111/j.1600-0714.2011.01019.x. [DOI] [PubMed] [Google Scholar]
- 170.Jiang Z, Chu PG, Woda BA, Rock KL, Liu Q, Hsieh CC, Li C, Chen W, Duan HO, McDougal S, Wu CL. Analysis of RNA-binding protein IMP3 to predict Metastasis and prognosis of renal-cell carcinoma: a retrospective study. Lancet Oncol. 2006;7:556–64. doi: 10.1016/S1470-2045(06)70732-X. [DOI] [PubMed] [Google Scholar]
- 171.Chen HM, Li H, Lin MX, Fan WJ, Zhang Y, Lin YT, Wu SX. Research Progress for RNA modifications in physiological and pathological angiogenesis. Front Genet. 2022;13:952667. doi: 10.3389/fgene.2022.952667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Orsolic I, Carrier A, Esteller M. Genetic and epigenetic defects of the RNA modification machinery in cancer. Trends Genet. 2023;39:74–88. doi: 10.1016/j.tig.2022.10.004. [DOI] [PubMed] [Google Scholar]
- 173.Xu P, Ge R. Roles and drug development of METTL3 (methyltransferase-like 3) in anti-tumor therapy. Eur J Med Chem. 2022;230:114118. doi: 10.1016/j.ejmech.2022.114118. [DOI] [PubMed] [Google Scholar]
- 174.Yankova E, Blackaby W, Albertella M, Rak J, De Braekeleer E, Tsagkogeorga G, Pilka ES, Aspris D, Leggate D, Hendrick AG, et al. Small-molecule inhibition of METTL3 as a strategy against myeloid Leukaemia. Nature. 2021;593:597–601. doi: 10.1038/s41586-021-03536-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Berdasco M, Esteller M. Towards a druggable epitranscriptome: compounds that target RNA modifications in cancer. Br J Pharmacol. 2022;179:2868–89. doi: 10.1111/bph.15604. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.