

Abstract
Recently, psychedelics have emerged as promising therapeutics for numerous neuropsychiatric disorders. While their potential in the clinic has yet to be fully elucidated, understanding their molecular and biological mechanisms is imperative as these compounds are becoming widely used both in therapeutic and recreational contexts. This review examines the current understanding of basic biology, pharmacology, and structural biology in an attempt to reveal both the knowns and unknowns within the field.
Subject terms: Pharmacology, Neuroscience
Introduction
Psychedelic drugs have been used for millennia by indigenous peoples and over the past century by others for recreational, spiritual and therapeutic purposes [1, 2]. In this review, we will refer to psychedelic drugs as those which induce a lysergic acid diethylamide (LSD)-like effect in humans via activation of 5-HT2A serotonin (5-hydroxtytryptamine; 5-HT) receptors [2]. As mentioned by others in this series, psychedelic drugs have had increasing interest over the past decade because small-scale clinical trials have shown that drugs like psilocybin and LSD have robust, rapid and enduring therapeutic actions for depression and anxiety [3–8]. Psychedelic drugs have also been suggested to have potential utility for many other conditions including chronic pain, cluster and migraine headaches, and obsessive-compulsive disorder among many others [2]. As definitive Phase III trials have not yet been concluded, there are currently no FDA-approved uses of psychedelic drugs for any condition. At the federal level, psychedelics remain Schedule I drugs; however, many have been decriminalized in several municipalities and the state of Oregon in the US despite the lack of evidence for their utility. Here we will focus on the postulated mechanisms of action of psychedelic drugs and provide a perspective on their potential as transformative neurotherapeutics.
Psychedelic drug pharmacology and molecular mechanisms of action
It is well established that the mind-altering actions of psychedelic drugs are due principally to the activation of 5-HT2A receptors. Initial evidence came from studies in the 1950s which noted the structural similarities between LSD and 5-HT [9] (Fig. 1). LSD was initially described as a serotonergic antagonist [10, 11] although later studies by Aghajanian et al. showed it is a serotonergic agonist with potent actions at raphe neurons [12]. Later studies showed that LSD has actions at dopamine [13] and other biogenic amine receptors [14]. Biochemical evidence for the actions of LSD at 5-HT receptors came from radioligand binding studies, which demonstrated that LSD interacts with a 5-HT2-like serotonin receptor [15]. These studies led, ultimately, to the discovery of the relatively selective radioligand [125I]-DOI, which preferentially interacts with the high-affinity agonist state of 5-HT2 receptors [16, 17]. More recent studies have shown that LSD is a high-affinity agonist for more than 20 biogenic amine receptors [18] including virtually all 5-HT, dopamine and noradrenergic receptors.
Fig. 1. Overview of the 5-HT (5-HT1-7) family of receptors and their canonical coupling pathways.

The inhibitory Gi/o is shown in red, the excitatory Gq is shown in green, the excitatory Gs is shown in blue, and the arrestin transducer pathway is shown in yellow. Lines pointing to which receptor families canonically interact with each transducer are shown. PDB accession codes are present for each structure in the figure. The different classes of 5-HT2A agonists are also shown (both hallucinogenic and non-hallucinogenic). The tryptamines are shown in gray (representative compound is the endogenous serotonin (5-HT) and non-psychedelic), the phenethylamines are shown in pink (representative compound is dopamine and non-psychedelic), and the ergolines are shown in tan (representative compound is the ergoline backbone). The very bottom is an artistic rendering of PDB accession 6WHA (25-CN-NBOH bound 5-HT2A heterotrimeric complex) present in a lipid bilayer.
Studies in mice and rats showed that psychedelics induce a variety of behaviors including, most notably, the head-twitch response (HTR) [19]. The HTR was later demonstrated to be induced by other 5-HT2 agonists like quipazine [20], which had been developed as a potential antidepressant [21]. More definitive evidence for the involvement of 5-HT2 receptors in the actions of psychedelic drugs came from studies performed by Glennon et al. who showed a direct correlation between the ability of drugs to interact with cortical 5-HT2A receptors labeled with [3H]-Ketanserin and the induction of HTR [22]. Although several non-psychedelic drugs induce the HTR in mice and rats, to date there are no bona fide psychedelic drugs that are negative in this assay [2]. Furthermore, Halberstadt et al. have recently shown a correlation between HTR and potency of many psychedelic compounds [23]. Psychedelic drugs were later observed to potently activate other 5-HT2-family receptors including the 5-HT2C [24] and 5-HT2B [25] (Fig. 1).
Data from human studies using the relatively selective 5-HT2A antagonist ketanserin [26] are more definitive. Initial studies with psilocybin showed that pretreatment with ketanserin blocked most [27–30] but not all of psilocybin’s psychoactive actions in human volunteers [31, 32]. Notably, the psychedelic actions of psilocybin were abolished by pretreatment with ketanserin, although some effects on cognition were apparently 5-HT1A mediated [31, 32]. For other psychedelic drugs, the data published to date show that ketanserin pretreatment blocks its psychedelic effects in humans [33, 34]. Given the fact that ketanserin has several off-target actions at other biogenic amine receptors (e.g., 5-HT2C antagonism, α1-adrenergic antagonism) use of more selective 5-HT2A antagonists such as pimavanserin [35], which has an appreciable affinity for 5-HT2C receptors might be suitable for human studies while M100907 (which is quite selective) is available for studies with rodents [36] (Fig. 2).
Fig. 2. Interactions with the widely used antagonists ketanserin, M100907, and pimavanserin against the 5-HT, dopamine, and alpha-adrenergic receptor families.
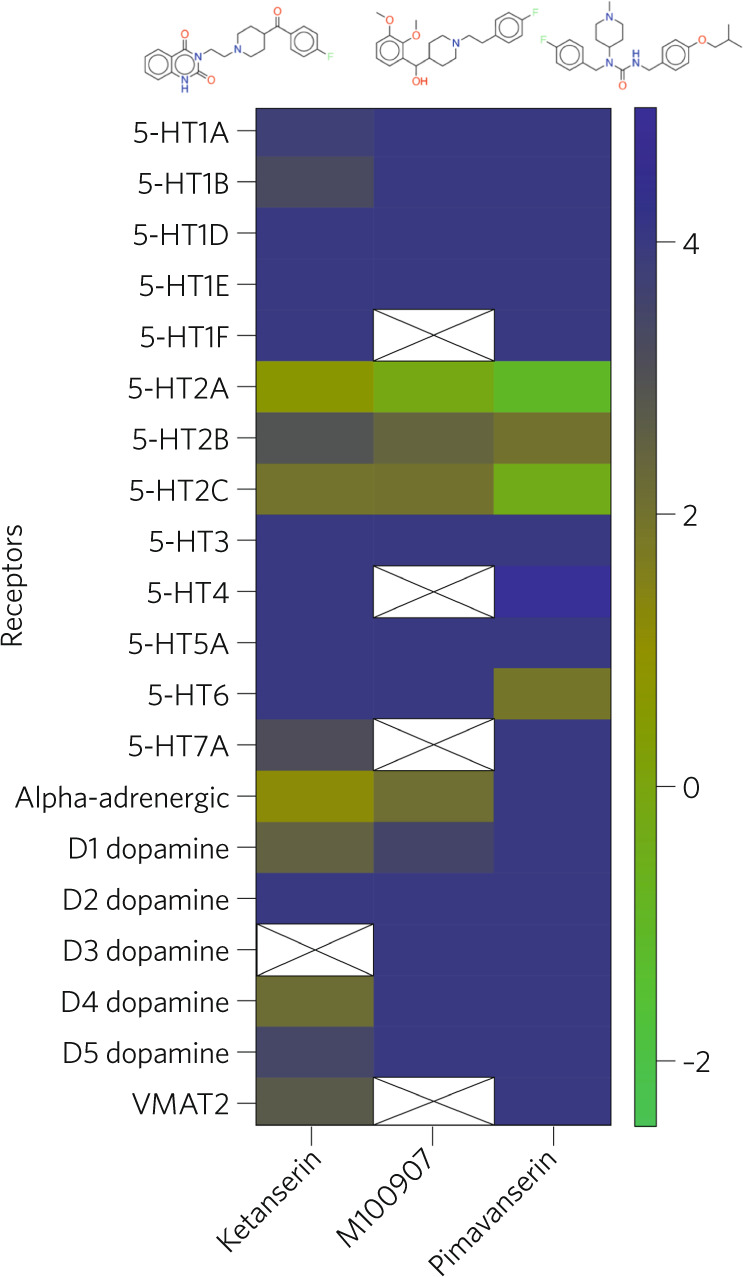
The heatmap is representative of radioligand binding affinities, with green being more potent and blue being less potent. Data obtained from KiDatabase (https://pdsp.unc.edu/databases/kidb.php).
Moreover, the molecular details regarding psychedelic drug actions at the 5-HT2A receptor have been characterized. For instance, both x-ray [37] and cryogenic electron microscopic (cryo-EM) studies of psychedelic [38] and non-psychedelic [39] drugs interacting with 5-HT2A receptors have been obtained (Fig. 3). R-69, which was found to be non-hallucinogenic in rodent models, interacts with the 5-HT2A receptor through the canonical salt-bridge (present in all aminergic receptors) D1553.32 and is positionally facing TM5 to interact with S2425.46 (Fig. 3) in which the indole nitrogen of 5-HT is observed to interact based on other 5-HT receptor structures [39]. Subsequently, many interactions are recapitulated with the potent hallucinogen 25-CN-NBOH, but the N-benzyl portion of the compound sterically “pushes” down on the toggle switch tryptophan (W3366.48) potentially playing a role in signaling bias and/or activation efficacy of the receptor (Fig. 3) [38]. In a direct comparison with hallucinogenic vs non-hallucinogenic molecules, LSD and lisuride crystal structures are available [38, 40]. Notably, the lisuride structure has an intrusive lipid in the orthosteric pocket pushing the position of the compound slightly compared to the LSD structure. While there is some evidence of lipids interacting with the orthosteric site in the 5-HT2A receptor, it remains unclear whether the lipid present in this structure is functionally relevant or an artifact of crystallization [40].
Fig. 3. Structures showing the molecular interactions over the entirety of the 5-HT2 family.

Subsets reveal specific interactions mentioned in the text. The top blue panel is representative of structures of 5-HT2A shown in cartoon representation. PDB Accession codes: receptor bound with R-69 and the mini-Gq heterotrimer – 7RAN, receptor bound with 25-CN-NBOH and the mini-Gq heterotrimer – 6WHA, receptor-bound LSD crystallization construct – 6WGT, receptor-bound Lisuride crystallization construct – 7WC7. The 5HT2A receptor is shown in cornflower blue for all the structures while in the heterotrimeric-bound structures, the mini-Gαq is shown in salmon, the β subunit is shown in green, the γ subunit is shown in yellow, and the stabilizing single-chain-Fab-16 (scFv16) is shown in orange. The stick representations showcase the respective ligands in the orthosteric pocket with specific interactions between the receptor and ligand highlighted, while the intrusive lipid is shown in the sphere representation for the lisuride structure. The bottom left panel is a representative structure of 5-HT2B bound to LSD. PDB Accession codes: Arrestin coupled – 7SRS, Gq coupled – 7SRR, and Transducer-free – 7SRQ. In this panel, the receptor is shown in dark green. For the arrestin coupled structure (top) β-arr1 is shown as a light sea green, with the respective stabilizing Fabs shown as the other colors. The mini-Gq couple heterotrimer coupled structure (middle) has a similar color as the 5-HT2A structures with the mini-Gαq shown in salmon, the β subunit shown in green, the γ subunit shown in yellow, and the stabilizing scFv16 shown in orange. The transducer-free complex (solved by cryo-EM), or receptor alone (bottom) is shown in dark green. The left box showcases LSD occupying the orthoseric pocket and highlights specific interactions with the receptor. The right box showcases the differences in the activation of TM6 between the various transducer-coupled states. Upon ligand activation, TM5/6 undergo a conformational change from the inactive state, in which the intracellular side of the helices are packed closer to the core of the receptor, to the active state characterized by the swinging outward and the ligand-activated state presented in the figure. The bottom right panel is a representative structure of the 5-HT2C receptor. PDB Accession codes: receptor bound with psilocin – 8DPG and the mini-Gq heterotrimer and the receptor bound with lorcaserin and the mini-Gq heterotrimer – 8DPF. The black boxes represent the respective ligands in the orthosteric site showcased as sticks with specific interaction(s) in the receptor also shown. The 5-HT2C receptor is shown in olive, the mini-Gαq is shown in salmon, the β subunit is shown in green, the γ subunit is shown in yellow, and the stabilizing single-chain-Fab-16 (scFv16) is shown in orange.
In addition, we also have details regarding the molecular requirements for specifying both G protein and arrestin-ergic signaling downstream of the closely related 5-HT2B receptor [41] (Fig. 3). Utilizing LSD, Cao et al. were able to structurally resolve the 5-HT2B receptor in the arrestin bound state, the Gq bound state, and the receptor without any transducer (transducer-free) by cryo-EM (Fig. 3) [41]. Remarkably, few changes within the orthosteric pocket and ligand binding mode occur between these states. However, it was noticed that TM6 was pushed farther out in the arrestin complex compared to the Gq-coupled state, while the transducer-free exhibited the smallest conformational change compared to the inactive state (Fig. 3). These movements are correlated with the position of the toggle switch tryptophan (W6.48) and could indicate a potential molecular mechanism for signaling bias.
Finally, the active state structures for the 5-HT2C receptors have also been solved with the classical psychedelic psilocin as well as the 5-HT2C selective agonist lorcaserin [42]. One key difference in the orthosteric site between 5-HT2A and 5-HT2C is at 5.46 (S242 and A222, respectively). Examining the psilocin structure, one could postulate that due to the potential loss of an H-bond on the indole N in 5-HT2C, the active conformation of psilocin in the orthosteric pocket will be slightly altered in the 5-HT2A receptor (Fig. 3) [42].
With regard to receptor signaling, it is well established from studies done nearly 40 years ago that 5-HT2A receptors couple to Gαq and modulate phosphoinositide hydrolysis (Fig. 4) [43–45]. This leads to both the mobilization of intracellular calcium via IP3-mediated release, activation of protein kinase C [46] and activation of Ca++ channels (Fig. 4) [47]. 5-HT2A receptors apparently are desensitized via direct phosphorylation [48] and subsequent arrestin binding via the third intracellular loop of the 5-HT2A receptor [49] and likely other sites including the C-terminal region [41, 50]. 5-HT2A receptors are also phosphorylated by RSK2 ribosomal S6 kinase [51], which exerts a tonic brake, or a reduction in the population of receptors available of ligand potentiated secondary messenger transduction, on 5-HT2A signaling (Fig. 4) [52]. Notably, RSK2-mediated phosphorylation alters the signaling properties of 5-HT2A receptors [53, 54]. Signaling via both Gq [55] and arrestin [56] appears essential for the actions of some psychedelic drugs in vivo.
Fig. 4. Representative signaling cascades and complexes of the 5-HT2A receptor.

Shown in the box is the canonical Gαq signaling pathway leading to spinogenesis and the potentially therapeutic mechanism of psychedelics. Other signaling mechanisms represented in the figure: the phosphorylation of the receptor by RSK2 leading to a tonic break on receptor signaling; interaction of the 5-HT2A receptor with PSD-95, which has been found to be essential for signaling; β-arrestin recruitment to the receptor and the canonical clathrin-mediated endocytosis of the receptor; and interaction between the receptor and caveolin-1.
In terms of interactions with other transducer proteins, most studies have indicated that 5-HT2A receptors selectively activate Gq-family proteins [38, 57]—at least in transfected cells in vitro. In vivo studies with Gq hetereozygote mice revealed that the behavioral actions of psychedelics are attenuated [55]. In addition, electrophysiological studies have shown that the 5-HT2A-mediated excitation of cortical neurons is insensitive to pertussis toxin [58]. As well, quite recent studies have shown that this excitation in 5-HT2A-identified neurons is abolished by pre-incubation with a selective Gq inhibitor in studies in mice [59]. In support of this hypothesis, quite recent studies have shown that the ability of psychedelic drugs to induce rapid firing of 5-HT2A receptors on identified neurons requires Gq activation [59]. By contrast, one report suggested that in rat renal mesangial cells 5-HT2A receptors may couple to a Gi-like protein [60]. Similarly, others have reported potential coupling to Gi-like proteins in the brain in vivo [61]. Taken together, these findings indicate that 5-HT2A receptors primarily couple to Gq-like G proteins and definitive studies are needed to clarify the role and potential relevance of interactions with other G protein transducers.
5-HT2A receptors are also found in multi-protein complexes in neurons in vivo and these interactions are essential for many of the actions of psychedelic drugs in vitro and in vivo. Thus, collaborative studies first demonstrated that the MAP1A microtubule-associated protein interacts with 5-HT2A receptors in vitro and in vivo [62] in intracellular vesicles. 5-HT2A receptors are also complexed with a number of PDZ-domain-containing proteins including PSD-95 [63, 64] and kalirin-7 (Fig. 4) [65]. The interactions with PSD-95 (Fig. 5) are essential for the biochemical and behavioral effects of psychedelics [64], while interactions with kalirin-7 are involved in psychedelic drug-induced spine formation [65]. Finally, 5-HT2A receptors also form complexes with caveolin-1 [66]. This interaction with caveolin-1 (CAV1) is essential for optimal 5-HT2A-mediated signaling in vitro [66] and in vivo [67]. Genetic deletion of CAV1 attenuates the signaling and behavioral actions of psychedelic drugs in vivo [67]. These results are consistent with models suggesting that 5-HT2A receptors exist in large multi-protein complexes in neurons in vivo and that these interactions are essential for many of the effects of psychedelic drugs (Fig. 5).
Fig. 5. Representative structures of the various possible interacting proteins with 5-HT2 receptors.
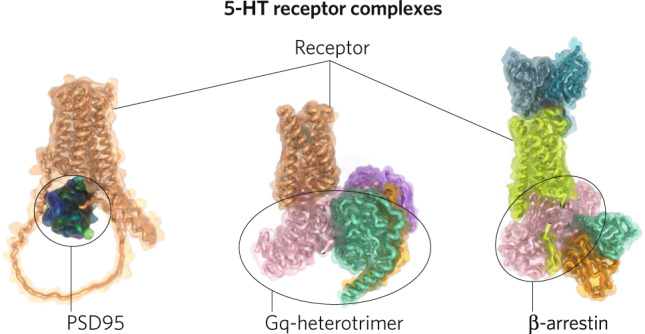
The left is the Alphafold-multimer-v2 created structure [130] of 5-HT2A and truncated PSD-95 complex. The middle is the 5-HT2A Gq complex (PDB Accession 6WHA) and the left is the 5-HT2B βarr1 complex (PDB Accession 7SRS).
Following 5-HT2A receptor activation, induction of spines and dendritic processes rapidly occurs [68]. The Rac guanine nucleotide exchange factor (RacGEF) kalirin-7 is essential for the immediate phase of this process [68], while TrkB activation may also play a role in more long-term actions [69]. The effects of psychedelics on spine formation occur within 30 min [68] and may be maintained for several weeks after a single administration in mice [70]. There has been increasing speculation that the potential therapeutic actions of psychedelic drugs may be mediated via the enhancement of spinogenesis and neuronal plasticity in cortical neurons [2], although definitive studies are lacking.
One of the complications encountered upon investigating the role of psychedelics in vivo is the fact that rodent and human 5-HT2A receptors differ significantly in pharmacology—particularly for tryptamines and ergolines [71, 72]. In this regard, it has been shown by our lab and others that a single amino acid Ser2425.46 where it is Ala2425.46 in rodents [38, 71, 72]. This Ser242Ala mutation greatly accelerates the dissociation rate of LSD at the 5-HT2A receptor [38] and decreases the affinity and potency of a variety of psychedelic and non-psychedelic ergolines and tryptamines [71, 72].
In addition, several non-synonymous single-nucleotide polymorphisms (SNPs) for the 5-HT2A receptor have been identified (see https://gnomad.broadinstitute.org/gene/ENSG00000102468?dataset=gnomad_r2_1; see ref. [73]). Of these, several have been reported to affect the agonist binding affinities, potencies and efficacies of several psychedelic and non-psychedelic 5-HT2A agonists and antagonists [48, 74, 75]. Importantly, the effects of various SNPs on 5-HT2A function were drug-specific and there were no SNPs that uniformly affected agonist or antagonist potency, affinity or efficacy [48, 74, 75]. Taken together, these results indicate that naturally occurring 5-HT2A receptor variants can have significant and unpredictable effects on drug actions.
In addition to the on-target actions of psychedelic drugs at 5-HT2A receptors, all known psychedelic drugs active in humans have substantial activity at other GPCRs [2]. LSD, for example, is a potent agonist at essentially every serotonin, dopamine and noradrenergic receptor in the brain [76]. Radioligand binding assays have also suggested a robust polypharmacological profile for all tested psychedelic drugs [77] although these studies do not clarify whether psychedelic drugs are agonists or antagonists at these receptors. While there are indications that the therapeutic actions require 5-HT2A receptor activation [69, 78, 79], currently, there is no definitive evidence in support of this mechanism, and indeed, at least one study has suggested that these effects are independent of 5-HT2A activation [80].
Perhaps the most consequential off-target actions of psychedelic drugs are at the 5-HT2B serotonin receptor. All tested psychedelic drugs are potent 5-HT2B agonists [76, 81, 82] or bind with appreciable affinity to 5-HT2B receptors [77]. We [83] and others [84] demonstrated that drugs that induce valvular heart disease in humans after chronic administration are potent 5-HT2B agonists [85]. This includes many anti-migraine drugs (e.g., ergotamine; [85]), drugs used in treating Parkinson’s Disease [85] as well as illicit drugs such as MDMA and MDA [86]. Indeed chronic administration of all of these drugs in humans is associated with clinically significant valvulopathy in as many as 30% of patients [87–91]. Taken together, these findings suggest that chronic treatment with psychedelic drugs, as could occur with so-called “microdosing”, may have unanticipated and serious side effects.
Interactions with other receptors could be important for both the therapeutic actions as well as other potential side effects of psychedelic drugs. As previously mentioned, one study has suggested the therapeutic actions of psilocybin are 5-HT2A-independent in mice [80].
Unanswered questions
How do psychedelic and non-psychedelic drugs differentially interact with 5-HT2A receptors?
It is now well known that a number of non-psychedelic medications and their active metabolites including fenfluramine and norfenfluramine [83], several ergot derivatives used in treating Parkinson’s disease and migraine headaches including ergotamine, lisuride, pergolide and bromocriptine [92], the trazodone metabolite m-CPP [93] and other drugs are potent 5-HT2A agonists. Importantly, these medications are devoid of psychedelic drug-like actions in humans at typical therapeutic doses although hallucinations have occasionally been reported when large doses of fenfluramine [94] or m-CPP [95] have been administered. In addition, bromocriptine and other ergots when used at therapeutic doses in Parkinson’s disease occasionally induce hallucinations [96]. Finally, Br-LSD, which is a weak partial agonist of human 5-HT2A receptors, is devoid of psychedelic actions in humans [97]. Importantly, several newly synthesized 5-HT2A agonists with antidepressant drug-like actions are devoid of psychedelic drug-like actions in mice including TBG [98], the lumateperone derivative IHCH-7113 [40] and the tetrahydropyridine R-69 [99]. Taken together, these reports indicate that activation of 5-HT2A receptors per se is insufficient to induce a psychedelic experience. It is currently unknown why some 5-HT2A agonists are psychedelic and further structural biology studies could elucidate these distinctions. As well, given the complicated polypharmacology of known psychedelic drugs [100], it is conceivable that actions at other molecular targets are essential for their effects. Thus, for instance, even though ketanserin blocks psychedelic actions of LSD and psilocybin in humans [27, 34], ketanserin has potent actions at other 5-HT2-family receptors, adrenergic receptors and the vesicular monoamine transporter [26, 101–103], to name but a few. Accordingly, the ability of ketanserin to block the actions of psychedelics in humans does not by itself provide unequivocal evidence for the causal role of 5-HT2A receptors in their actions. Studies with more selective and potent 5-HT2A antagonists could clarify this issue.
Do psychedelic and non-psychedelic 5-HT2A agonists have differential signaling downstream of 5-HT2A receptors?
It has been known since the 1980s that various 5-HT2A agonists differ in their signaling processes. Thus, Felder et al. [104] reported that some 5-HT2A agonists might activate arachidonic acid (AA) release independent of Gq-mediated activation of phospholipase C. These findings have been validated and extended by others [105–107], although whether a drug is apparently psychedelic or not is irrelevant to whether or not it also activated AA release. Subsequently, Gonzales-Maeso et al. reported that psychedelic 5-HT2A agonists differ from non-psychedelic drugs by selective activation of a pertussis toxin-sensitive pathway involving Src activation [61]. This group has also reported that a heterodimeric complex between mGluR2 metabotropic glutamate and 5-HT2A receptors may be responsible for these actions [108–110]. Others have provided data suggesting the complex as such is not essential for the actions of psychedelics at 5-HT2A receptors [111]. This group [111] also reported that 5-HT2A receptors coupled efficiently to Gq pathways but not to Gi signaling. One potential approach to test the hypothesis that Gq and not Gi signaling is essential for the actions of psychedelics in vivo would be to use a chemogenetic approach. Thus, for instance, expressing Gq- and Gi-DREADDs [112] in 5-HT2A neurons and stimulating them with deschloroclozapine [113] or alternative ligands [114] and observing potential behavioral actions could address these issues.
Are the putative therapeutic actions of psychedelic drugs mediated by 5-HT2A receptor activation?
Currently, there are no selective 5-HT2A agonists which are approved for use in humans and the available psychedelic drugs have a complex polypharmacologic profile. Moreover, a rich area to explore is how this polypharmacological profile can be modulated to treat various disorders. Although no studies have been performed in humans, the available preclinical data support, with some exceptions, actions at 5-HT2A receptors are essential for their therapeutic drug-like actions in rodents [40, 79, 98, 115] . As previously mentioned, there is some evidence that 5-HT2A activation by psilocybin is insufficient to induce an antidepressant-like response [80] and there have also been reports that some of LSD’s actions are mediated by 5-HT5A- serotonin [116] and D2-dopamine [117] receptors. The availability of selective 5-HT5A- [118] and D2 antagonists should prove invaluable for determining the role of these off-target actions for psychedelic actions. Finally, to definitively address this question, human studies utilizing 5-HT2A antagonists prior to psychedelic drug administration or the use of selective 5-HT2A agonists in human trials will be essential.
How are the putative therapeutic actions of psychedelics mediated?
Current hypotheses suggest that psychedelics may exert their therapeutic drug-like actions via enhancing synaptic plasticity [2]. These findings are based largely on prior studies demonstrating that psychedelic drugs induce both rapid [68, 69] and sustained [70] augmentation of dendritic spine formation and plasticity in cortical neurons in vitro and in vivo, consistent with their expression at dendritic spines [119, 120]. It has long been known that conventional antidepressant drugs [121, 122] as well as novel antidepressants like ketamine require spine formation for their therapeutic drug-like actions [123, 124]. This pathway requires various post-synaptic density proteins [68] as well as the activation of TrkB receptors by BDNF [125]. With regard to this, psychedelic drugs have long been known to enhance BDNF levels [126] and TrkB inhibition can attenuate the actions of psychedelic drugs on spine formation [69].
Discussion and directions for future basic research on psychedelics
As is clear from the foregoing, there are many potentially significant areas for continued research on psychedelics. These include determining: (1) how psychedelic and non-psychedelic 5-HT2A agonists differentially interact with the receptor; (2) how signal transduction downstream of 5-HT2A receptors differs among various psychedelic and non-psychedelic agonists; (3) whether on- or off-target actions of psychedelics are essential for their putative therapeutic actions; and (4) how are the potential therapeutic actions of psychedelic drugs mediated. Central to these unanswered questions is the overarching question of whether non-psychedelic 5-HT2A agonists may retain at least some of the therapeutic actions of psychedelic drugs.
As mentioned previously, a number of drugs that do not induce a psychedelic drug-like action in rodents have been recently discovered including TBG [98], IHCH-7113 [40] and R-69 [39]. To this list, we may also add the legacy compound Ariadne [127], Br-LSD [128] and, in some cases, lisuride [129] which have all shown efficacy in preliminary clinical trials and reports. Such medications have the potential to transform our treatment of many neuropsychiatric conditions.
Author contributions
BLR and RHG jointly wrote the paper and created the figures.
Funding
Work in the Roth lab is supported by grants from the NIH and DARPA to BLR as well as the Michael Hooker Distinguished Professorship.
Competing interests
BLR is a member of the Scientific Advisory Boards of Septerna Pharmaceuticals, Escient Pharmaceuticals and Onsero, Inc. As well, BLR is a scientific co-founder of Onsero and is listed as an inventor on patents related to the research in this review article. RHG declares no conflicts.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Nichols DE. Psychedelics. Pharm Rev. 2016;68:264–355. doi: 10.1124/pr.115.011478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.McClure-Begley TD, Roth BL. The promises and perils of psychedelic pharmacology for psychiatry. Nat Rev Drug Discov. 2022;21:463–73. doi: 10.1038/s41573-022-00421-7. [DOI] [PubMed] [Google Scholar]
- 3.Griffiths RR, Johnson MW, Carducci MA, Umbricht A, Richards WA, Richards BD, et al. Psilocybin produces substantial and sustained decreases in depression and anxiety in patients with life-threatening cancer: a randomized double-blind trial. J Psychopharmacol. 2016;30:1181–97. doi: 10.1177/0269881116675513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ross S, Bossis A, Guss J, Agin-Liebes G, Malone T, Cohen B, et al. Rapid and sustained symptom reduction following psilocybin treatment for anxiety and depression in patients with life-threatening cancer: a randomized controlled trial. J Psychopharmacol. 2016;30:1165–80. doi: 10.1177/0269881116675512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Davis AK, Barrett FS, May DG, Cosimano MP, Sepeda ND, Johnson MW, et al. Effects of psilocybin-assisted therapy on major depressive disorder: a randomized clinical trial. JAMA Psychiatry. 2021;78:481–9. 10.1001/jamapsychiatry.2020.3285. [DOI] [PMC free article] [PubMed]
- 6.Carhart-Harris R, Giribaldi B, Watts R, Baker-Jones M, Murphy-Beiner A, Murphy R, et al. Trial of psilocybin versus escitalopram for depression. N Engl J Med. 2021;384:1402–11. doi: 10.1056/NEJMoa2032994. [DOI] [PubMed] [Google Scholar]
- 7.Holze F, Gasser P, Muller F, Dolder PC, Liechti ME. Lysergic acid diethylamide-assisted therapy in patients with anxiety with and without a life-threatening illness: a randomized, double-blind, placebo-controlled Phase II study. Biol Psychiatry. 2023;93:215–23. 10.1016/j.biopsych.2022.08.025. [DOI] [PubMed]
- 8.Goodwin GM, Aaronson ST, Alvarez O, Arden PC, Baker A, Bennett JC, et al. Single-dose psilocybin for a treatment-resistant episode of major depression. N Engl J Med. 2022;387:1637–48. doi: 10.1056/NEJMoa2206443. [DOI] [PubMed] [Google Scholar]
- 9.Wooley DW, Shaw E. A biochemical and pharmacological suggestion about certain mental disorders. Proc Natl Acad Sci USA. 1954;40:228–31. doi: 10.1073/pnas.40.4.228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Gaddum JH, Hameed KA. Drugs which antagonize 5-hydroxytryptamine. Br J Pharm. 1954;9:240–8. doi: 10.1111/j.1476-5381.1954.tb00848.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Gaddum JH, Khan A, Hathway DE, Stephens FF. Quantitative studies of antagonists for 5-hydroxytryptamine. Q J Exp Physiol. 1955;40:49–74. doi: 10.1113/expphysiol.1955.sp001097. [DOI] [PubMed] [Google Scholar]
- 12.Aghjanian GK, Foote WE, Sheard MH. Lysergic acid diethylamide: sensitive neuronal units in the midbrain raphe. Science. 1968;161:706–8. doi: 10.1126/science.161.3842.706. [DOI] [PubMed] [Google Scholar]
- 13.Kelly PH, Iversen LL. LSD as an agonist at mesolimbic dopamine receptors. Psychopharmacologia. 1975;45:221–4. doi: 10.1007/BF00429064. [DOI] [PubMed] [Google Scholar]
- 14.Geyer MA, Gordon J, Adams LM. Behavioral effects of xylamine-induced depletions of brain norepinephrine: interaction with LSD. Pharm Biochem Behav. 1985;23:619–25. doi: 10.1016/0091-3057(85)90427-7. [DOI] [PubMed] [Google Scholar]
- 15.Peroutka SJ, Snyder SH. Multiple serotonin receptors: differential binding of [3H]5-hydroxytryptamine, [3H]lysergic acid diethylamide and [3H]spiroperidol. Mol Pharm. 1979;16:687–99. [PubMed] [Google Scholar]
- 16.Glennon RA, Seggel MR, Soine WH, Herrick-Davis K, Lyon RA, Titeler M. 125I-2,5-dimethoxy-4-iodophenyl-2-aminopropane (DOI): an iodinated radioligand that specifically labels the agonist high affinity state of the 5HT2 serotonin receptor. J Med Chem. 1988;31:5–7. doi: 10.1021/jm00396a003. [DOI] [PubMed] [Google Scholar]
- 17.Johnson MP, Hoffman AJ, Nichols DE, Mathis CA. Binding to the serotonin 5-HT2 receptor by the enantiomers of 125I-DOI. Neuropharmacology. 1987;26:1803–6. doi: 10.1016/0028-3908(87)90138-9. [DOI] [PubMed] [Google Scholar]
- 18.Kroeze WK, Sassano MF, Huang XP, Lansu K, McCorvy JD, Giguère PM, et al. PRESTO-Tango as an open-source resource for interrogation of the druggable human GPCRome. Nat Struct Mol Biol. 2015;22:362–9. doi: 10.1038/nsmb.3014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Corne SJ, Pickering RW, Warner BT. A method for assessing the effects of drugs on the central actions of 5-hydroxytryptamine. Br J Pharm Chemother. 1963;20:106–20. doi: 10.1111/j.1476-5381.1963.tb01302.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Malick JB, Doren E, Barnett A. Quipazine-induced head-twitch in mice. Pharm Biochem Behav. 1977;6:325–9. doi: 10.1016/0091-3057(77)90032-6. [DOI] [PubMed] [Google Scholar]
- 21.Rodriguez R, Pardo EG. Quipazine, a new type of antidepressant agent. Psychopharmacologia. 1971;21:89–100. doi: 10.1007/bf00404000. [DOI] [PubMed] [Google Scholar]
- 22.Glennon RA, Titler M, McKenney JD. Evidence for 5-HT2 involvement in the mechanism of action of hallucinogenic agents. Life Sci. 1984;35:2505–11. doi: 10.1016/0024-3205(84)90436-3. [DOI] [PubMed] [Google Scholar]
- 23.Halberstadt AL, Chatha M, Klein AK, Wallach J, Brandt SD. Correlation between the potency of hallucinogens in the mouse head-twitch response assay and their behavioral and subjective effects in other species. Neuropharmacology. 2020;167:107933. doi: 10.1016/j.neuropharm.2019.107933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Sanders-Bush E, Burris KD, Knoth K. Lysergic acid diethylamide and 2,5-dimethoxy-4-methylamphetamine are partial agonists at serotonin receptors linked to phosphoinositide hydrolysis. J Pharmacol Exp Ther. 1988;246:924–8. [PubMed] [Google Scholar]
- 25.Sard H, Kumaran G, Morency C, Roth BL, Toth BA, He P, et al. SAR of psilocybin analogs: discovery of a selective 5-HT(2C) agonist. Bioorg Med Chem Lett. 2005;15:4555–9. [DOI] [PubMed]
- 26.Leysen JE, Niemegeers CJE, Van Nueten JM, Laduron PM. [3H]-ketanserin (R 41 468) a selective 3H-ligand for serotonin2 receptor binding sites. Mol Pharmacol. 1982;21:301–14. [PubMed] [Google Scholar]
- 27.Vollenweider FX, Vollenweider-Scherpenhuyzen MF, Babler A, Vogel H, Hell D. Psilocybin induces schizophrenia-like psychosis in humans via a serotonin-2 agonist action. Neuroreport. 1998;9:3897–902. doi: 10.1097/00001756-199812010-00024. [DOI] [PubMed] [Google Scholar]
- 28.Quednow BB, Kometer M, Geyer MA, Vollenweider FX. Psilocybin-induced deficits in automatic and controlled inhibition are attenuated by ketanserin in healthy human volunteers. Neuropsychopharmacology. 2012;37:630–40. 10.1038/npp.2011.228. [DOI] [PMC free article] [PubMed]
- 29.Kometer M, Schmidt A, Bachmann R, Studerus E, Seifritz E, Vollenweider FX. Psilocybin biases facial recognition, goal-directed behavior, and mood state toward positive relative to negative emotions through different serotonergic subreceptors. Biol Psychiatry. 2012;72:898–906. doi: 10.1016/j.biopsych.2012.04.005. [DOI] [PubMed] [Google Scholar]
- 30.Kometer M, Schmidt A, Jancke L, Vollenweider FX. Activation of serotonin 2A receptors underlies the psilocybin-induced effects on alpha oscillations, N170 visual-evoked potentials, and visual hallucinations. J Neurosci. 2013;33:10544–51. doi: 10.1523/JNEUROSCI.3007-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Bernasconi F, Schmidt A, Pokorny T, Kometer M, Seifritz E, Vollenweider FX. Spatiotemporal brain dynamics of emotional face processing modulations induced by the serotonin 1A/2A receptor agonist psilocybin. Cereb Cortex. 2014;24:3221–31. doi: 10.1093/cercor/bht178. [DOI] [PubMed] [Google Scholar]
- 32.Pokorny T, Preller KH, Kraehenmann R, Vollenweider FX. Modulatory effect of the 5-HT1A agonist buspirone and the mixed non-hallucinogenic 5-HT1A/2A agonist ergotamine on psilocybin-induced psychedelic experience. Eur Neuropsychopharmacol. 2016;26:756–66. doi: 10.1016/j.euroneuro.2016.01.005. [DOI] [PubMed] [Google Scholar]
- 33.Barrett FS, Preller KH, Herdener M, Janata P, Vollenweider FX. Serotonin 2A receptor signaling underlies LSD-induced alteration of the neural response to dynamic changes in music. Cereb Cortex. 2018;28:3939–50. doi: 10.1093/cercor/bhx257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Preller KH, Burt JB, Ji JL, Schleifer CH, Adkinson BD, Stämpfli P, et al. Changes in global and thalamic brain connectivity in LSD-induced altered states of consciousness are attributable to the 5-HT2A receptor. Elife. 2018;7e35082. 10.7554/eLife.35082. [DOI] [PMC free article] [PubMed]
- 35.Abbas A, Roth B. Pimavanserin tartrate: a 5-HT2A inverse agonist with potential for treating various neuropsychiatric disorders. Expert Opin Pharmacother. 2008;9:3251–9. doi: 10.1517/14656560802532707. [DOI] [PubMed] [Google Scholar]
- 36.Sorensen SM, Kehne JH, Fadayel GM, Humphreys TM, Ketteler HJ, Sullivan CK, et al. Characterization of the 5-HT2 antagonist MDL 100907 as a putative atypical antipsychotic: behavioral, electrophysiological and neurochemical studies. J Pharmacol Exp Ther. 1993;266:684–91. [PubMed] [Google Scholar]
- 37.Wacker D, Wang S, McCorvy JD, Betz RM, Venkatakrishnan AJ, Levit A, et al. Crystal structure of an LSD-bound human serotonin receptor. Cell. 2017;168:377–89.e312. doi: 10.1016/j.cell.2016.12.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kim K, Che T, Panova O, DiBerto JF, Lyu J, Krumm BE, et al. Structure of a hallucinogen activated gq-coupled 5-HT2A serotonin receptor. Cell. 2020;182:1574–88.e1519. doi: 10.1016/j.cell.2020.08.024.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kaplan AL, Confair DN, Kim K, Barros-Álvarez X, Rodriguiz RM, Yang Y, et al. Bespoke library docking for 5-HT2A receptor agonists with antidepressant activity. Nature. 2022;610:582–91. doi: 10.1038/s41586-022-05258-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Cao D, Yu J, Wang H, Luo Z, Liu X, He L, et al. Structure-based discovery of nonhallucinogenic psychedelic analogs. Science. 2022;375:403–11. doi: 10.1126/science.abl8615. [DOI] [PubMed] [Google Scholar]
- 41.Cao C, Barros-Álvarez X, Zhang S, Kim K, Dämgen MA, Panova O, et al. Signaling snapshots of a serotonin receptor activated by the prototypical psychedelic LSD. Neuron. 2022;110:3154–67.e3157. doi: 10.1016/j.neuron.2022.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Gumpper RH, Fay JF, Roth BL. Molecular insights into the regulation of constitutive activity by RNA editing of 5HT(2C) serotonin receptors. Cell Rep. 2022;40:111211. doi: 10.1016/j.celrep.2022.111211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Roth BL, Nakaki T, Chuang DM, Costa E. Aortic recognition sites for serotonin (5HT) are coupled to phospholipase C and modulate phosphatidylinositol turnover. Neuropharmacology. 1984;23:1223–5. doi: 10.1016/0028-3908(84)90244-2. [DOI] [PubMed] [Google Scholar]
- 44.Conn PJ, Sanders-Bush E. Selective 5-HT2 antagonists inhibit serotonin-stimulated phosphatidylinositol metabolism in cerebral cortex. Neuropharmacology. 1984;23:993–6. doi: 10.1016/0028-3908(84)90017-0. [DOI] [PubMed] [Google Scholar]
- 45.Leysen JE, De Chaffoy De Courcelles D, De Clerck F, Niemegeers CJE, Van Nueten JM. Serotonin-S2 receptor binding sites and functional correlates. Neuropharmacology. 1984;23:1493–501. doi: 10.1016/0028-3908(84)90093-5. [DOI] [PubMed] [Google Scholar]
- 46.Roth BL, Nakaki T, Chuang DM, Costa E. 5-Hydroxytryptamine2 receptors coupled to phospholipase C in rat aorta: modulation of phosphoinositide turnover by phorbol ester. J Pharm Exp Ther. 1986;238:480–5. [PubMed] [Google Scholar]
- 47.Nakaki T, Roth BL, Chuang DM, Costa E. Phasic and tonic components in 5-HT2 receptor-mediated rat aorta contraction: participation of Ca++ channels and phospholipase C. J Pharm Exp Ther. 1985;234:442–6. [PubMed] [Google Scholar]
- 48.Gray JA, Compton-Toth BA, Roth BL. Identification of two serine residues essential for agonist-induced 5-HT2A receptor desensitization. Biochemistry. 2003;42:10853–62. doi: 10.1021/bi035061z. [DOI] [PubMed] [Google Scholar]
- 49.Gelber EI, Kroeze WK, Willins DL, Gray JA, Sinar CA, Hyde EG, et al. Structure and function of the third intracellular loop of the 5-hydroxytryptamine(2A) receptor: the third intracellular loop is alpha-helical and binds purified arrestins. J Neurochem. 1999;72:2206–14. doi: 10.1046/j.1471-4159.1999.0722206.x. [DOI] [PubMed] [Google Scholar]
- 50.Gray J, Bhatnagar A, Gurevich V, Roth B. The interaction of a constitutively active arrestin with the arrestin-insensitive 5-HT2A receptor induces agonist-independent internalization. Mol Pharmacol. 2003;63:961–72. doi: 10.1124/mol.63.5.961. [DOI] [PubMed] [Google Scholar]
- 51.Strachan RT, Sheffler DJ, Willard B, Kinter M, Kiselar JG, Roth BL. Ribosomal S6 kinase 2 directly phosphorylates the 5-HT2A serotonin receptor thereby modulating 5-HT2A signaling. J Biol Chem. 2009;284:5557–73. 10.1074/jbc.M805705200. [DOI] [PMC free article] [PubMed]
- 52.Sheffler DJ, Kroeze WK, Garcia BG, Deutch AY, Hufeisen SJ, Leahy P, et al. p90 ribosomal S6 kinase 2 exerts a tonic brake on G protein-coupled receptor signaling. Proc Natl Acad Sci USA. 2006;103:4717–22. doi: 10.1073/pnas.0600585103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Strachan RT, Allen JA, Sheffler DJ, Roth BL. p90 Ribosomal S6 kinase 2, a novel GPCR kinase, is required for growth factor-mediated attenuation of GPCR signaling. Biochemistry. 2010;49:2657–71. doi: 10.1021/bi901921k. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Strachan RT, Sciaky N, Cronan MR, Kroeze WK, Roth BL. Genetic deletion of p90 ribosomal S6 kinase 2 alters patterns of 5-hydroxytryptamine 2A serotonin receptor functional selectivity. Mol Pharm. 2010;77:327–38. doi: 10.1124/mol.109.061440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Garcia EE, Smith RL, Sanders-Bush E. Role of G(q) protein in behavioral effects of the hallucinogenic drug 1-(2,5-dimethoxy-4-iodophenyl)-2-aminopropane. Neuropharmacology. 2007;52:1671–7. doi: 10.1016/j.neuropharm.2007.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Rodriguiz RM, Nadkarni V, Means CR, Pogorelov VM, Chiu YT, Roth BL, et al. LSD-stimulated behaviors in mice require beta-arrestin 2 but not beta-arrestin 1. Sci Rep. 2021;11:17690. doi: 10.1038/s41598-021-96736-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Lucaites VL, Nelson DL, Wainscott DB, Baez M. Receptor subtype and density determine the coupling repertoire of the 5-HT2 receptor subfamily. Life Sci. 1996;59:1081–95. doi: 10.1016/0024-3205(96)00423-7. [DOI] [PubMed] [Google Scholar]
- 58.Zhang JY, Ashby CR, Jr, Wang RY. Effect of pertussis toxin on the response of rat medial prefrontal cortex cells to the iontophoresis of serotonin receptor agonists. J Neural Transm Gen Sect. 1994;95:165–72. doi: 10.1007/BF01271563. [DOI] [PubMed] [Google Scholar]
- 59.Schmitz GP, Chiu YT, König GM, Kostenis E, Roth BL, Herman MA. Psychedelic compounds directly excite 5-HT2A layer 5 pyramidal neurons in the prefrontal cortex through a 5-HT2A Gq-mediated activation mechanism. bioRxiv [Preprint]. 2022 2022.2011.2015.516655. 10.1101/2022.11.15.516655.
- 60.Garnovskaya MN, Nebigil CG, Arthur JM, Spurney RF, Raymond JR. 5-Hydroxytryptamine2A receptors expressed in rat renal mesangial cells inhibit cyclic AMP accumulation. Mol Pharm. 1995;48:230–7. [PubMed] [Google Scholar]
- 61.González-Maeso J, Weisstaub NV, Zhou M, Chan P, Ivic L, Ang R, et al. Hallucinogens recruit specific cortical 5-HT(2A) receptor-mediated signaling pathways to affect behavior. Neuron. 2007;53:439–52. doi: 10.1016/j.neuron.2007.01.008. [DOI] [PubMed] [Google Scholar]
- 62.Cornea-Hébert V, Watkins KC, Roth BL, Kroeze WK, Gaudreau P, Leclerc N, et al. Similar ultrastructural distribution of the 5-HT2A serotonin receptor and microtubule-associated protein MAP1A in cortical dendrites of adult rat. Neuroscience. 2002;113:23–35. doi: 10.1016/s0306-4522(02)00146-x. [DOI] [PubMed] [Google Scholar]
- 63.Xia Z, Gray J, Compton-Toth B, Roth B. A direct interaction of PSD-95 with 5-HT2A serotonin receptors regulates receptor trafficking and signal transduction. J Biol Chem. 2003;278:21901–8. doi: 10.1074/jbc.M301905200. [DOI] [PubMed] [Google Scholar]
- 64.Abbas AI, Yadav PN, Yao WD, Arbuckle MI, Grant SG, Caron MG, et al. PSD-95 is essential for hallucinogen and atypical antipsychotic drug actions at serotonin receptors. J Neurosci. 2009;29:7124–36. doi: 10.1523/JNEUROSCI.1090-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Jones KA, Srivastava DP, Allen JA, Strachan RT, Roth BL, Penzes P. Rapid modulation of spine morphology by the 5-HT2A serotonin receptor through kalirin-7 signaling. Proc Natl Acad Sci USA. 2009;106:19575–80. 10.1073/pnas.0905884106. [DOI] [PMC free article] [PubMed]
- 66.Bhatnagar A, Sheffler D, Kroeze W, Compton-Toth B, Roth B. Caveolin-1 interacts with 5-HT2A serotonin receptors and profoundly modulates the signaling of selected G alpha(q)-coupled protein receptors. J Biol Chem. 2004;279:34614–23. doi: 10.1074/jbc.M404673200. [DOI] [PubMed] [Google Scholar]
- 67.Allen JA, Yadav PN, Setola V, Farrell M, Roth BL. Schizophrenia risk gene CAV1 is both pro-psychotic and required for atypical antipsychotic drug actions in vivo. Transl Psych. 2011;1:e33. doi: 10.1038/tp.2011.35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Jones KA, Srivastava DP, Allen JA, Strachan RT, Roth BL, Penzes P. Rapid modulation of spine morphology by the 5-HT2A serotonin receptor through kalirin-7 signaling. Proc Natl Acad Sci USA. 2009;106:19575–80. doi: 10.1073/pnas.0905884106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Ly C, Greb AC, Cameron LP, Wong JM, Barragan EV, Wilson PC, et al. Psychedelics promote structural and functional neural plasticity. Cell Rep. 2018;23:3170–82. doi: 10.1016/j.celrep.2018.05.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Shao LX, Liao C, Gregg I, Davoudian PA, Savalia NK, Delagarza K, et al. Psilocybin induces rapid and persistent growth of dendritic spines in frontal cortex in vivo. Neuron. 2021;109:2535–44.e2534. doi: 10.1016/j.neuron.2021.06.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Johnson MP, Loncharich RJ, Baez M, Nelson DL. Species variations in transmembrane region V of the 5-hydroxytryptamine type 2A receptor alter the structure-activity relationship of certain ergolines and tryptamines. Mol Pharm. 1994;45:277–86. [PubMed] [Google Scholar]
- 72.Johnson MP, Baez M, Kursar JD, Nelson DL. Species differences in 5-HT2A receptors: cloned pig and rhesus monkey 5-HT2A receptors reveal conserved transmembrane homology to the human rather than rat sequence. Biochim Biophys Acta. 1995;1236:201–6. doi: 10.1016/0005-2736(95)00073-c. [DOI] [PubMed] [Google Scholar]
- 73.Karczewski KJ, Francioli LC, Tiao G, Cummings BB, Alföldi J, Wang Q, et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature. 2020;581:434–43. doi: 10.1038/s41586-020-2308-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Davies MA, Setola V, Strachan RT, Sheffler DJ, Salay E, Hufeisen SJ, et al. Pharmacologic analysis of non-synonymous coding h5-HT2A SNPs reveals alterations in atypical antipsychotic and agonist efficacies. Pharmacogenomics J. 2006;6:42–51. doi: 10.1038/sj.tpj.6500342. [DOI] [PubMed] [Google Scholar]
- 75.Schmitz GP, Jain MK, Slocum ST, Roth BL. 5-HT(2A) SNPs alter the pharmacological signaling of potentially therapeutic psychedelics. ACS Chem Neurosci. 2022;13:2386–98. doi: 10.1021/acschemneuro.1c00815. [DOI] [PubMed] [Google Scholar]
- 76.Kroeze WK, Sassano MF, Huang XP, Lansu K, McCorvy JD, Giguère PM, et al. PRESTO-Tango as an open-source resource for interrogation of the druggable human GPCRome. Nat Struct Mol Biol. 2015;22:362–9. doi: 10.1038/nsmb.3014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Ray TS. Psychedelics and the human receptorome. PLoS One. 2010;5:e9019. 10.1371/journal.pone.0009019. [DOI] [PMC free article] [PubMed]
- 78.Cameron LP, Patel SD, Vargas MV, Barragan EV, Saeger HN, Warren HT, et al. 5-HT2ARs mediate therapeutic behavioral effects of psychedelic tryptamines. ACS Chem Neurosci. 2023;14:351–8. 10.1021/acschemneuro.2c00718. [DOI] [PMC free article] [PubMed]
- 79.de la Fuente Revenga M, Zhu B, Guevara CA, Naler LB, Saunders JM, Zhou Z, et al. Prolonged epigenomic and synaptic plasticity alterations following single exposure to a psychedelic in mice. Cell Rep. 2021;37:109836. doi: 10.1016/j.celrep.2021.109836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Hesselgrave N, Troppoli TA, Wulff AB, Cole AB, Thompson SM. Harnessing psilocybin: antidepressant-like behavioral and synaptic actions of psilocybin are independent of 5-HT2R activation in mice. Proc Natl Acad Sci USA. 2021;118:e2022489118. 10.1073/pnas.2022489118. [DOI] [PMC free article] [PubMed]
- 81.Sard H, Kumaran G, Morency C, Roth BL, Toth BA, He P, et al. SAR of psilocybin analogs: discovery of a selective 5-HT 2C agonist. Bioorg Med Chem Lett. 2005;15:4555–9. doi: 10.1016/j.bmcl.2005.06.104. [DOI] [PubMed] [Google Scholar]
- 82.Huang XP, Setola V, Yadav PN, Allen JA, Rogan SC, Hanson BJ, et al. Parallel functional activity profiling reveals valvulopathogens are potent 5-hydroxytryptamine(2B) receptor agonists: implications for drug safety assessment. Mol Pharm. 2009;76:710–22. doi: 10.1124/mol.109.058057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Rothman RB, Baumann MH, Savage JE, Rauser L, McBride A, Hufeisen SJ, et al. Evidence for possible involvement of 5-HT(2B) receptors in the cardiac valvulopathy associated with fenfluramine and other serotonergic medications. Circulation. 2000;102:2836–41. doi: 10.1161/01.cir.102.23.2836. [DOI] [PubMed] [Google Scholar]
- 84.Fitzgerald LW, Burn TC, Brown BS, Patterson JP, Corjay MH, Valentine PA, et al. Possible role of valvular serotonin 5-HT(2B) receptors in the cardiopathy associated with fenfluramine. Mol Pharm. 2000;57:75–81. [PubMed] [Google Scholar]
- 85.Roth BL. Drugs and valvular heart disease. N Engl J Med. 2007;356:6–9. doi: 10.1056/NEJMp068265. [DOI] [PubMed] [Google Scholar]
- 86.Setola V, Hufeisen SJ, Grande-Allen KJ, Vesely I, Glennon RA, Blough B, et al. 3,4-Methylenedioxymethamphetamine (MDMA, “Ecstasy”) induces fenfluramine-like proliferative actions on human cardiac valvular interstitial cells in vitro. Mol Pharm. 2003;63:1223–9. doi: 10.1124/mol.63.6.1223. [DOI] [PubMed] [Google Scholar]
- 87.Connolly HM, McGoon MD. Obesity drugs and the heart. Curr Probl Cardiol. 1999;24:745–92. doi: 10.1016/s0146-2806(99)90013-0. [DOI] [PubMed] [Google Scholar]
- 88.Zanettini R, Antonini A, Gatto G, Gentile R, Tesei S, Pezzoli G. Valvular heart disease and the use of dopamine agonists for Parkinson’s disease. N Engl J Med. 2007;356:39–46. doi: 10.1056/NEJMoa054830. [DOI] [PubMed] [Google Scholar]
- 89.Montastruc F, Montastruc G, Vigreux P, Bruneval P, Guilbeau-Frugier C, Cron C, et al. Valvular heart disease in a patient taking 3,4-methylenedioxymethamphetamine (MDMA, ‘Ecstasy’) Br J Clin Pharm. 2012;74:547–8. doi: 10.1111/j.1365-2125.2012.04252.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Droogmans S, Cosyns B, D’haenen H, Creeten E, Weytjens C, Franken PR, et al. Possible association between 3,4-methylenedioxymethamphetamine abuse and valvular heart disease. Am J Cardiol. 2007;100:1442–5. doi: 10.1016/j.amjcard.2007.06.045. [DOI] [PubMed] [Google Scholar]
- 91.Cavero I, Guillon JM. Safety pharmacology assessment of drugs with biased 5-HT(2B) receptor agonism mediating cardiac valvulopathy. J Pharm Toxicol Methods. 2014;69:150–61. doi: 10.1016/j.vascn.2013.12.004. [DOI] [PubMed] [Google Scholar]
- 92.Newman-Tancredi A, Cussac D, Quentric Y, Touzard M, Verrièle L, Carpentier N, et al. Differential actions of antiparkinson agents at multiple classes of monoaminergic receptor. III. Agonist and antagonist properties at serotonin, 5-HT(1) and 5-HT(2), receptor subtypes. J Pharm Exp Ther. 2002;303:815–22. doi: 10.1124/jpet.102.039883. [DOI] [PubMed] [Google Scholar]
- 93.Willins DL, Meltzer HY. Direct injection of 5-HT2A receptor agonists into the medial prefrontal cortex produces a head-twitch response in rats. J Pharm Exp Ther. 1997;282:699–706. [PubMed] [Google Scholar]
- 94.Griffith JD, Nutt JG, Jasinski DR. A comparison of fenfluramine and amphetamine in man. Clin Pharm Ther. 1975;18:563–70. doi: 10.1002/cpt1975185part1563. [DOI] [PubMed] [Google Scholar]
- 95.Koreen AR, Lieberman JA, Alvir J, Chakos M. The behavioral effect of m-chlorophenylpiperazine (mCPP) and methylphenidate in first-episode schizophrenia and normal controls. Neuropsychopharmacology. 1997;16:61–68. doi: 10.1016/S0893-133X(96)00160-1. [DOI] [PubMed] [Google Scholar]
- 96.Parkes JD, Marsden CD, Donaldson I, Galea-Debono A, Walters J, Kennedy G, et al. Bromocriptine treatment in Parkinson’s disease. J Neurol Neurosurg Psychiatry. 1976;39:184–93. doi: 10.1136/jnnp.39.2.184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Knoll J, Vizi ES. Cross-tolerance between para-bromo-methamphetamine (V-111) and LSD-25. Pharmacology. 1970;4:278–86. doi: 10.1159/000136148. [DOI] [PubMed] [Google Scholar]
- 98.Cameron LP, Tombari RJ, Lu J, Pell AJ, Hurley ZQ, Ehinger Y, et al. A non-hallucinogenic psychedelic analogue with therapeutic potential. Nature. 2021;589:474–9. doi: 10.1038/s41586-020-3008-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Levit Kaplan A, Strachan RT, Braz JM, Craik V, Slocum S, Mangano T, et al. Structure-based design of a chemical probe set for the 5-HT5A serotonin receptor. J Med Chem. 2022;65:4201–17. doi: 10.1021/acs.jmedchem.1c02031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Kwan AC, Olson DE, Preller KH, Roth BL. The neural basis of psychedelic action. Nat Neurosci. 2022;25:1407–19. doi: 10.1038/s41593-022-01177-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Casey AB, Cui M, Booth RG, Canal CE. “Selective” serotonin 5-HT(2A) receptor antagonists. Biochem Pharm. 2022;200:115028. doi: 10.1016/j.bcp.2022.115028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Leysen JE, Gommeren W, Van Gompel P, Wynants J, Janssen PAJ. Non-serotonergic [3H]-ketanserin binding sites in striatal membranes are associated with a dopac release system on dopaminergic nerve endings. Eur J Pharm. 1987;134:373–5. doi: 10.1016/0014-2999(87)90373-6. [DOI] [PubMed] [Google Scholar]
- 103.Roth BL, McLean S, Zhu XZ, Chuang DM. Characterization of two [3H]ketanserin recognition sites in rat striatum. J Neurochem. 1987;49:1833–8. doi: 10.1111/j.1471-4159.1987.tb02444.x. [DOI] [PubMed] [Google Scholar]
- 104.Felder CC, Kanterman RY, Ma AL, Axelrod J. Serotonin stimulates phospholipase A2 and the release of arachidonic acid in hippocampal neurons by a type 2 serotonin receptor that is independent of inositolphospholipid hydrolysis. Proc Natl Acad Sci USA. 1990;87:2187–91. doi: 10.1073/pnas.87.6.2187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Berg KA, Maayani S, Goldfarb J, Clarke WP. Pleiotropic behavior of 5-HT2A and 5-HT2C receptor agonists. Ann N Y Acad Sci. 1998;861:104–10. doi: 10.1111/j.1749-6632.1998.tb10180.x. [DOI] [PubMed] [Google Scholar]
- 106.Berg KA, Maayani S, Goldfarb J, Scaramellini C, Leff P, Clarke WP. Effector pathway-dependent relative efficacy at serotonin type 2A and 2C receptors: evidence for agonist-directed trafficking of receptor stimulus. Mol Pharm. 1998;54:94–104. [PubMed] [Google Scholar]
- 107.Kurrasch-Orbaugh DM, Watts VJ, Barker EL, Nichols DE. Serotonin 5-hydroxytryptamine 2A receptor-coupled phospholipase C and phospholipase A2 signaling pathways have different receptor reserves. J Pharm Exp Ther. 2003;304:229–37. doi: 10.1124/jpet.102.042184. [DOI] [PubMed] [Google Scholar]
- 108.González-Maeso J, Ang RL, Yuen T, Chan P, Weisstaub NV, López-Giménez JF, et al. Identification of a serotonin/glutamate receptor complex implicated in psychosis. Nature. 2008;452:93–7. doi: 10.1038/nature06612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Moreno JL, Holloway T, Albizu L, Sealfon SC, Gonzalez-Maeso J. Metabotropic glutamate mGlu2 receptor is necessary for the pharmacological and behavioral effects induced by hallucinogenic 5-HT2A receptor agonists. Neurosci Lett. 2011;493:76–9. doi: 10.1016/j.neulet.2011.01.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Moreno JL, Muguruza C, Umali A, Mortillo S, Holloway T, Pilar-Cuéllar F, et al. Identification of three residues essential for 5-hydroxytryptamine 2A-metabotropic glutamate 2 (5-HT2A.mGlu2) receptor heteromerization and its psychoactive behavioral function. J Biol Chem. 2012;287:44301–19. doi: 10.1074/jbc.M112.413161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Delille HK, Becker JM, Burkhardt S, Bleher B, Terstappen GC, Schmidt M, et al. Heterocomplex formation of 5-HT2A-mGlu2 and its relevance for cellular signaling cascades. Neuropharmacology. 2012;62:2184–91. doi: 10.1016/j.neuropharm.2012.01.010. [DOI] [PubMed] [Google Scholar]
- 112.Armbruster BN, Li X, Pausch MH, Herlitze S, Roth BL. Evolving the lock to fit the key to create a family of G protein-coupled receptors potently activated by an inert ligand. Proc Natl Acad Sci USA. 2007;104:5163–8. doi: 10.1073/pnas.0700293104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Nagai Y, Miyakawa N, Takuwa H, Hori Y, Oyama K, Ji B, et al. Deschloroclozapine, a potent and selective chemogenetic actuator enables rapid neuronal and behavioral modulations in mice and monkeys. Nat Neurosci. 2020;23:1157–67. 10.1038/s41593-020-0661-3. [DOI] [PubMed]
- 114.Zhang S, Gumpper RH, Huang XP, Liu Y, Krumm BE, Cao C, et al. Molecular basis for selective activation of DREADD-based chemogenetics. Nature. 2022;612:354–62. 10.1038/s41586-022-05489-0. [DOI] [PubMed]
- 115.Yu B, Becnel J, Zerfaoui M, Rohatgi R, Boulares AH, Nichols CD. Serotonin 5-hydroxytryptamine(2A) receptor activation suppresses tumor necrosis factor-alpha-induced inflammation with extraordinary potency. J Pharm Exp Ther. 2008;327:316–23. doi: 10.1124/jpet.108.143461. [DOI] [PubMed] [Google Scholar]
- 116.Grailhe R, Waeber C, Dulawa SC, Hornung JP, Zhuang X, Brunner D, et al. Increased exploratory activity and altered response to LSD in mice lacking the 5-HT(5A) receptor. Neuron. 1999;22:581–91. doi: 10.1016/s0896-6273(00)80712-6. [DOI] [PubMed] [Google Scholar]
- 117.Marona-Lewicka D, Thisted RA, Nichols DE. Distinct temporal phases in the behavioral pharmacology of LSD: dopamine D2 receptor-mediated effects in the rat and implications for psychosis. Psychopharmacology (Berl) 2005;180:427–35. doi: 10.1007/s00213-005-2183-9. [DOI] [PubMed] [Google Scholar]
- 118.Zhang S, Chen H, Zhang C, Yang Y, Popov P, Liu J, et al. Inactive and active state structures template selective tools for the human 5-HT5A receptor. Nat Struct Mol Biol. 2022;29:677–87. doi: 10.1038/s41594-022-00796-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Miner LA, Backstrom JR, Sanders-Bush E, Sesack SR. Ultrastructural localization of serotonin2A receptors in the middle layers of the rat prelimbic prefrontal cortex. Neuroscience. 2003;116:107–17. doi: 10.1016/s0306-4522(02)00580-8. [DOI] [PubMed] [Google Scholar]
- 120.Xia Z, Hufeisen SJ, Gray JA, Roth BL. The PDZ-binding domain is essential for the dendritic targeting of 5-HT(2A) serotonin receptors in cortical pyramidal neurons in vitro. Neuroscience. 2003;122:907–20. doi: 10.1016/s0306-4522(03)00589-x. [DOI] [PubMed] [Google Scholar]
- 121.Moda-Sava RN, Murdock MH, Parekh PK, Fetcho RN, Huang BS, Huynh TN, et al. Sustained rescue of prefrontal circuit dysfunction by antidepressant-induced spine formation. Science. 2019;364:eaat8078. 10.1126/science.aat8078. [DOI] [PMC free article] [PubMed]
- 122.Hajszan T, MacLusky NJ, Leranth C. Short-term treatment with the antidepressant fluoxetine triggers pyramidal dendritic spine synapse formation in rat hippocampus. Eur J Neurosci. 2005;21:1299–303. doi: 10.1111/j.1460-9568.2005.03968.x. [DOI] [PubMed] [Google Scholar]
- 123.Coyle JT, Duman RS. Finding the intracellular signaling pathways affected by mood disorder treatments. Neuron. 2003;38:157–60. doi: 10.1016/s0896-6273(03)00195-8. [DOI] [PubMed] [Google Scholar]
- 124.Duman RS. Neuropharmacology in the next millennium: promise for breakthrough discoveries [editorial] Neuropsychopharmacology. 1999;20:97–8. doi: 10.1016/S0893-133X(98)00105-5. [DOI] [PubMed] [Google Scholar]
- 125.Liu RJ, Lee FS, Li XY, Bambico F, Duman RS, Aghajanian GK. Brain-derived neurotrophic factor Val66Met allele impairs basal and ketamine-stimulated synaptogenesis in prefrontal cortex. Biol Psychiatry. 2012;71:996–1005. doi: 10.1016/j.biopsych.2011.09.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Gewirtz JC, Chen AC, Terwilliger R, Duman RC, Marek GJ. Modulation of DOI-induced increases in cortical BDNF expression by group II mGlu receptors. Pharm Biochem Behav. 2002;73:317–26. doi: 10.1016/s0091-3057(02)00844-4. [DOI] [PubMed] [Google Scholar]
- 127.Cunningham MJ, Bock HA, Serrano IC, Bechand B, Vidyadhara DJ, Bonniwell EM, et al. Pharmacological mechanism of the non-hallucinogenic 5-HT(2A) agonist ariadne and analogs. ACS Chem Neurosci. 2022;14:119–35. doi: 10.1021/acschemneuro.2c00597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Karst M, Halpern JH, Bernateck M, Passie T. The non-hallucinogen 2-bromo-lysergic acid diethylamide as preventative treatment for cluster headache: an open, non-randomized case series. Cephalalgia. 2010;30:1140–4. doi: 10.1177/0333102410363490. [DOI] [PubMed] [Google Scholar]
- 129.Hougaku H, Matsumoto M, Hata R, Handa N, Imaizumi M, Sugitani Y, et al. [Therapeutic effect of lisuride maleate on post-stroke depression] Nihon Ronen Igakkai Zasshi. 1994;31:52–9. doi: 10.3143/geriatrics.31.52. [DOI] [PubMed] [Google Scholar]
- 130.Jumper J, Evans R, Pritzel A, Green T, Figurnov M, Ronneberger O, et al. Highly accurate protein structure prediction with AlphaFold. Nature. 2021;596:583–9. doi: 10.1038/s41586-021-03819-2. [DOI] [PMC free article] [PubMed] [Google Scholar]