

Abstract
Transmembrane AMPA receptor regulatory proteins (TARPs) are claudin-like proteins that tightly regulate AMPA receptors (AMPARs) and are fundamental for excitatory neurotransmission. We used cryo-electron microscopy (cryo-EM) to reconstruct the 36 kDa TARP subunit γ2 to 2.3 Å and reveal the structural diversity of TARPs. Our data reveals critical motifs that distinguish TARPs from claudins and define how sequence variations within TARPs differentiate subfamilies and their regulation of AMPARs.
Information transfer in the brain occurs at specialized cellular junctions known as synapses, which act as neuronal communication hubs1. Most synapses are glutamatergic, where a pre-synaptic neuron releases glutamate (Glu), and a post-synaptic neuron receives Glu. AMPARs in the post-synaptic membrane bind Glu and initiate depolarization of the post-synaptic neuron through their Glu-gated cation channels1,2. TARPs are auxiliary subunits that regulate the trafficking, gating kinetics, and pharmacology of AMPARs2,3.
TARP regulatory subunits tightly regulate AMPAR function in the post-synaptic membrane, which is a critical aspect of the brain’s ability to fine tune information processing1–3. There are six TARP subtypes (TARPγ2, γ3, γ4, γ5, γ7, γ8), split into type-I (TARPγ2, γ3, γ4, γ8) and type-II (TARP0γ5, γ7) families. Generally, TARPs increase the conductance of AMPARs, but type-I TARPs slow desensitization and deactivation kinetics, while type-II TARPs appear to have a negative effect on gating when compared to type-I TARPs2. Furthermore, structural differences between TARPs in the same class underlie sensitivity to certain classes of drugs targeted to AMPAR-TARP complexes. Since the first TARP was identified a quarter century ago (TARPγ2, also known as stargazin)4, TARPs have been recognized as an indispensable component of synaptic function1,2. Yet, the structural details of how TARPs regulate AMPARs remain ambiguous.
Cryo-EM studies of TARP subunits have advanced our understanding of TARP structure in the context of AMPAR complexes, but intermediate resolution has historically precluded de novo building of TARP structures5–14. X-ray crystallography structures of TARP homologs, such as claudins, have been indispensable for modeling TARPs15. Claudins are cellular junction proteins that form paracellular barriers between epithelial and endothelial cells and are functionally distinct from TARPs16. The reliance on claudin structures for TARP modeling has hampered identification of distinct structural features that 1) differentiate TARPs from claudins and 2) explain the regulatory potential of TARPs for AMPARs. Here, we use cryo-EM to determine the structure of the prototypical TARP, TARPγ2. We identify new motifs in TARPγ2 that distinguish TARP classes from one another and further differentiate TARPs from Claudins. These structural features likely underlie modulatory effects exhibited by TARPs on AMPAR gating.
We reconstructed the 3D architecture of TARPγ2 to an overall resolution of 2.3 Å (2.0 Å – 2.5 Å locally; Extended Data Fig. 1). Our data enables us to build most of the transmembrane domain (TMD) and extracellular domain (ECD) de novo (Fig. 1a). The high resolution of our reconstruction enables identification of multiple distinct structural features in the TARPγ2 extracellular domain (ECD), which sits atop its tetraspanin transmembrane (TM) helical bundle comprised of transmembrane (TM) helices TM1–4 (Fig. 1a). The ECD is comprised of a five-stranded β-sheet and a single extracellular helix (ECH) that immediately precedes TM2. A previously identified disulfide bridge (DSB) between b3 (C67) and b4 (C77) strands in the ECD stabilizes the TARPγ2 ECD (Fig. 1b) and is conserved across all TARPs and the TARP-like claudins.
Figure 1. Structure of TARPγ2.

a) Cryo-EM map of TARPγ2, colored rainbow from N-terminus, NT (blue) to C-terminus, CT (red). b) Extracellular portion of the TARPγ2 model showing the β3-β4 DSB, loop anchor DSB, and TARP cleat. c) Cartoon schematic of TARPγ2 structure highlighting key structural features that rigidify the entire ECD atop the tetraspanin TMD, colored as in panel a.
What makes TARPγ2, and all TARPs unique from claudins? We identify two new moieties in our reconstruction of TARPγ2 that distinguish TARPs from claudins. First, a π-π-π stack secures the TARPγ2 ECD atop the TARPγ2 TMD (Fig. 1b). This is formed by H60 (from β2), Y32 (TM1-β1 loop), and W178 (TM4). We term this the TARP cleat motif because it helps to fasten the ECD to the TMD. We also identified a second DSB in the ECD. This DSB, the loop anchor DSB, anchors the β1-β2 loop onto the b-sheet (Fig. 1b). The loop anchor DSB is made between C40 in the β1–0β2 loop and C68 on b3. All together, these motifs rigidify the structure of TARPγ2 by providing additional structural interactions within the ECD and between the ECD and TMD (Fig. 1c).
How conserved are these motifs? The TARP cleat motif is conserved in all TARPs and the TARP-like subunit germline specific gene 1-like (GSG1L) (Fig. 2a) but absent from all claudins (Extended Data Fig. 2). We also tested for conservation of the cleat motif through AlphaFold217 structure prediction. This suggests that the TARP cleat motif is present in all mammalian TARPs (Extended Data Fig. 3a). Interestingly, while the TARP cleat motif is conserved in all TARPs, the loop anchor DSB is not (Fig. 2a). Structure prediction in AlphaFold2 (Extended Data Fig. 3b) also points to the loop anchor DSB being conserved in type-I TARPs but not in type-II TARPs. Thus, while our structure pointed us to look at the conservation of the cleat motif and loop anchor DSB, this was already predicted by AlphaFold2 (Extended Data Fig. 3c).
Figure 2. Conservation of structural features among TARP family members.

a) Multiple sequence alignment demonstrating the relative conservation of the TARP Cleat Motif, β3-β4 DSB, and Loop Anchor DSB between TARP family members. Loop Anchor DSB is unique to type-I and excluded from type-II TARPs. b) Alignment of TARPγ2 structure with other TARP family members (TARPγ3, PDB: 8C2H; TARPγ5, PDB: 7RZ5; TARPγ8, PDB: 8AYN; GSG1L, PDB: 7RZ9). c) Zoomed in view of TARP extracellular domains illustrating differing orientations in the β1-β2 loops. d) View of the TARP cleat motif illustrating conservation among all TARP family members. e) Model of predicted β1-β2 loop orientations between type-I and type-II TARPs illustrating distinct potential contacts between TARP subtypes and AMPARs.
Surprisingly, the TARP cleat motif and loop anchor DSB are within previous TARP structures but not identified. Previously determined structures of TARPs are overall like our structure of TARPγ2 (Fig. 2b), and the loop anchor DSB is within structures of TARPγ318 and TARPg811,12,19, and even previously published structures of TARPγ26. However, it is absent, as expected, in the structure of the type-II TARP, TARPγ520,21 (Fig. 2c) and the TARP-like subunit GSG1L7,20 (Fig. 2c). In contrast, the TARP cleat motif is conserved in all TARPγ3, γ5, and γ8 subunit structures as well as GSG1L11,18,20 (Fig. 2d). Thus, we suggest expanding the type-II family of TARPs to include the GSG1L subunit. We hypothesize that these structural details and their conservation were previously missed because of a lack of structural resolution.
The dichotomy in β1-β2 loop organization between type-I and type-II TARPs has significant functional implications. For example, type-II TARPs lack the loop anchor DSB and have been observed to directly interact with AMPAR subunits that are in the A and C positions when they occupy the “X” auxiliary subunit site7,20 (Fig. 2e). However, we expect that this is not possible for type-I TARPs in the X site given the presence of the loop anchor DSB, which locks in the β1-β2 loop in an orientation away from the A and C AMPAR subunit positions. However, if a type-I TARP occupies the “Y” TARP position (Fig. 2e), modulation of the AMPAR at subunit positions B or D by the β1-β2 loop is likely possible despite the loop anchor DSB, and is supported by observations in cryo-EM studies of type-I TARPs in complex with AMPARs18. Given the extreme conformational changes associated with AMPAR gating, the stark difference in the presence or absence of the loop anchor DSB within type-I TARPs versus type-II TARPs potentially explains differences in electrophysiology experiments between chimeric constructs of the β1-β2 loop in type-I and type-II TARPs.
The TARP cleat motif plays a significant role in distinguishing TARPs from claudins. Both TARPs and claudins share the same overall structural fold (i.e., tetraspanin with a five-stranded extracellular β-sheet). However, claudins have strong oligomerization properties, where they self-oligomerize to form paracellular barriers. A similar phenomenon has not been reported for TARP proteins. We hypothesize that the TARP cleat motif plays a role in preventing oligomerization in TARPs, enabling their complexation with AMPARs and other synaptic proteins.
In sum, we report the structure of TARPγ2, and how the newly identified structural features may account for critical functional differences between TARPs that tune AMPAR function throughout the central nervous system. In addition, we precisely define how TARPs are differentiated from claudins, which may explain the critical point of divergence between the structurally related proteins that are functionally distinct. Our findings provide a new framework for future studies to understand the function of TARPs and new foundations to target TARPs in structure-based drug design against AMPAR-related neurological disorders.
Methods
Construct design, protein expression, and purification
Mouse TARPγ2 was covalently fused to the rat AMPAR subunit GluA2, expressed, and purified as described in the preprint Hale, et al. Biorxiv 2023 (BIORXIV/2023/569057).
Cryo-EM Sample Preparation and Data Collection
Cryo-EM samples were prepared and collected as described in the preprint Hale, et al. Biorxiv 2023 (BIORXIV/2023/569057).
Image Processing
The initial stages of cryo-EM sample preparation were carried out as in the preprint Hale, et al. Biorxiv 2023 (BIORXIV/2023/569057). After generation of a 2.80 Å AMPAR-TARPγ2 local map (Extended Data Fig. 1a), symmetry expansion was used to refine the structure of TARPγ2. To achieve this, we applied C4 symmetry to the AMPAR-TARP particles (Extended Data Fig. 1a). We masked one TARPγ2 in the AMPAR-TARPγ2, then inverted this mask, and subtracted the inverted mask from all particle images. We then used the subtracted particle images, coupled with the original TARPγ2 mask (non-inverted) applied to the complete AMPAR-TARPγ2 complex cryo-EM map reference to refine the final cryo-EM reconstruction of TARPγ2 (Extended Data Fig. 1b).
Model building, refinement, and structural analysis
Coot22 was used to build a polyalanine chain into TARPγ2 map. Bulky resides from sequence information were used to anchor the building. A previously determined structure of TARPγ2 (pdb 5WEO) and a structure predicted from AlphaFold2 (AlphaFold Protein Structure Database, #AF-O88602) were used as reference. Isolde23 and Phenix24 were used to refine the model. Quality of the model was assessed with MolProbity25. Visualizations and domain measurements were performed in ChimeraX26. Software was compiled and accessed via the SBGrid Consortium27.
Sequence Analysis
All sequence alignments were done with ClustalW28 and analyzed in Jalview29.
Structure Prediction
TARP structure predictions of TARPγ2, γ3, γ4, γ5, γ7, γ8 of human, rat, mouse species were used from AlphaFold217. For each TARP subunit structure prediction, the respective amino acids corresponding to the cleat motif and disulfide bridge were determined. Cleat motif measurements were taken by calculating the distance between the Cα’s of histidine to tyrosine and Cα’s of tyrosine to tryptophan. Calculations were performed using the Biopython.PDB package.
AlphaFold2 accession numbers of models: AF-Q9Y698, AF-A0JNG9, AF-O88602, AF-Q71RJ2, AF-Q9JJV5, AF-Q0VD05, AF-O60359, AF-Q8VHX0, AF-A0A3Q1LKG2, AF-Q9JJV4, AF-Q8VHW9, AF-Q9UBN1, AF-E1BEI3, AF-Q8VHW4, AF-Q8VHW8, AF-Q9UF02, AF-E1BIG3, AF-P62956, AF-P62957, AF-P62955, AF-Q8WXS5, AF-F1MV40, AF-Q8VHW2, AF-Q8VHW5.
Extended Data
Extended Data Figure 1. Details of TARPγ2 data processing workflow.
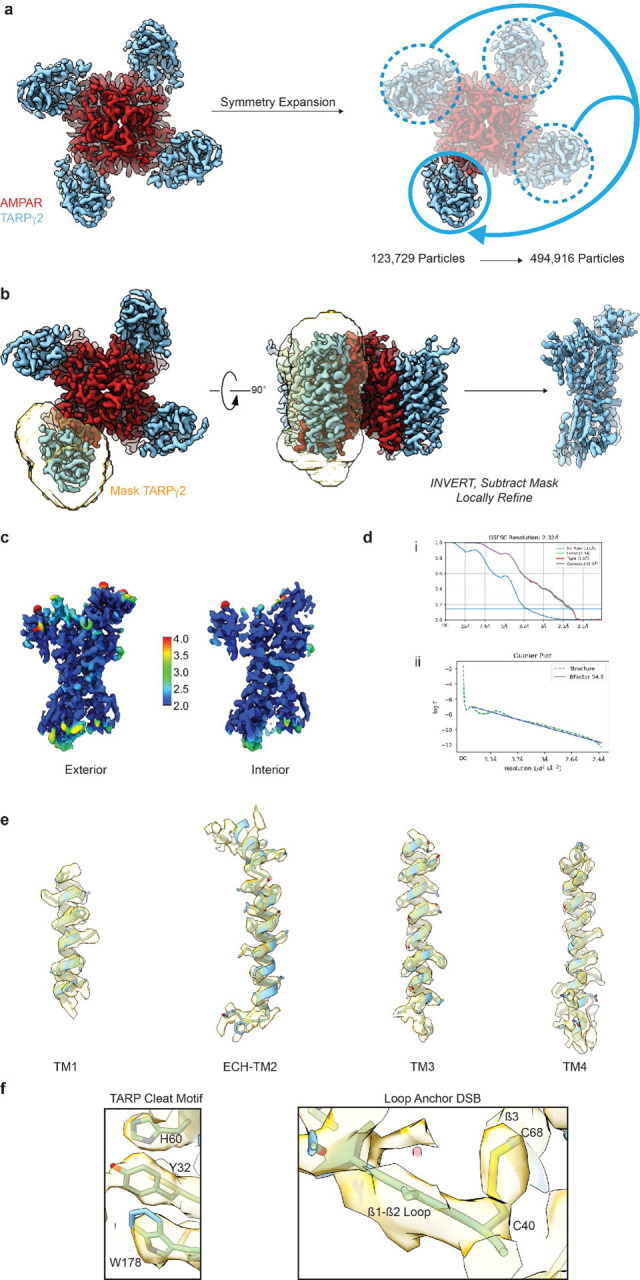
a) Symmetry expansion of the GluA2-TARPγ2 assembly (from Hale et al., 2023, BioRxiv). b) Masking scheme for isolating symmetry-expanded TARPγ2. c) TARPγ2 cryo-EM map colored by local resolution right: surface of TARPγ2 reconstruction, left: cutaway showing resolution inside the map. d) Gold Standard Fourier Shell Correlation and Guinier Plots for TARPγ2. e) Model fit to cryo-EM map of the four TARPγ2 TM helices. f) Cryo-EM map around the TARP Cleat motif and the Loop Anchor DSB.
Extended Data Figure 1 2. Multiple sequence alignment of TARPs, GSG1L and Claudins.

Multiple sequence alignments of TARPs, GSG1L and all members of the Claudin family. The TARPs and GSG1L are distinguished from Claudins by the presence of the TARP cleat motif while the β3-β4 DSB is conserved among both TARPs and Claudins.
Extended Data Figure 3. AlphaFold structure prediction of TARPs.
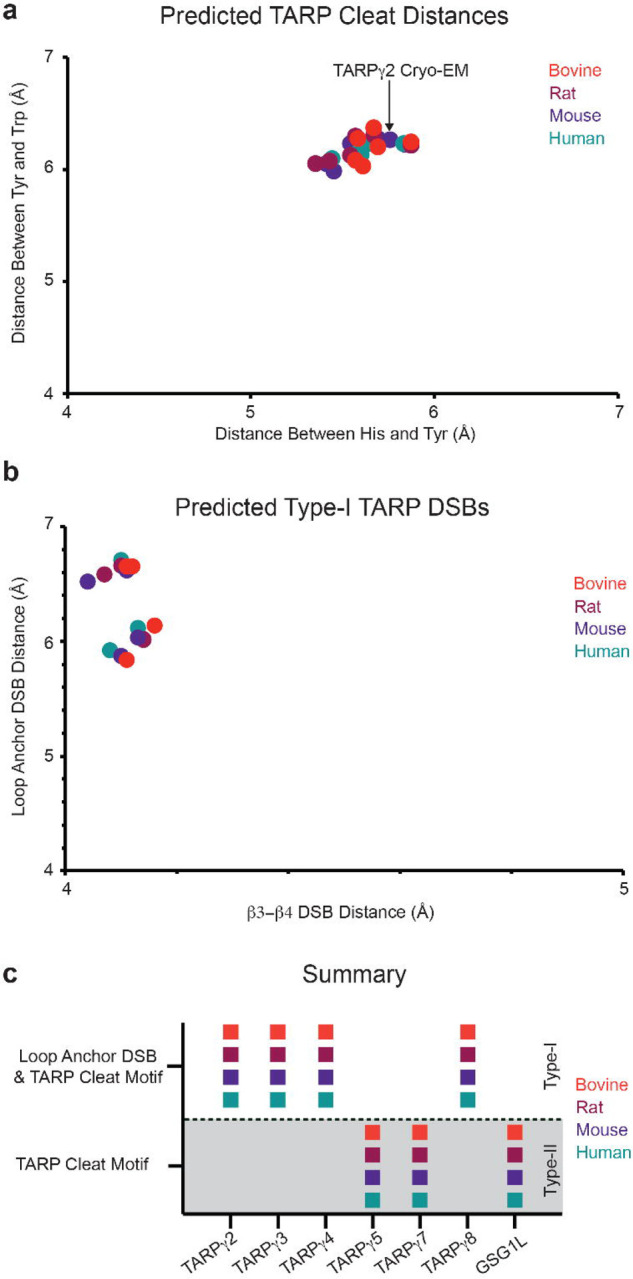
a) Conservation of TARP cleat residues in bovine, rat, mouse, and human TARPs. TARPγ2 from this study is pointed out. b) Loop anchor DSB vs. β3-β4 DSB distances. The TARP cleat is predicted to be present in all TARPs. Type-II TARPs are excluded from panel b because the loop anchor DSB is predicted to be absent in type-II TARPs. These findings are summarized in panel c.
Cryo-EM data collection, refinement and validation statistics
| TARPγ2 (EMDB-xxxx) (PDB xxxx) |
|
|---|---|
| Data collection and processing | |
| Magnification | 130,000x |
| Voltage (kV) | 300 |
| Electron exposure (e−/Å2) | 40 |
| Defocus range (μm) | −1.0 – 2.0 |
| Pixel size (Å) | 0.93 |
| Symmetry imposed | C1 |
| Initial particle images (no.) | 123,729 |
| Final particle images (no.) | 494,916 |
| Map resolution (Å) | 2.32 |
| FSC = 0.143 | |
| Map resolution range (Å) | 2 – 4 |
| Refinement | |
| Initial model used (PDB code) | N/A |
| Model resolution (Å) | 2.3 |
| FSC = 0.143 | |
| Model resolution range (Å) | |
| Map sharpening B factor (Å2) | −54.8 |
| Model composition | |
| Non-hydrogen atoms | 2678 |
| Protein residues | 172 |
| Ligands | 0 |
| B factors (Å2) | |
| Protein | 0.00/27.91/5.8 |
| Ligand | N/A |
| R.m.s. deviations | |
| Bond lengths (Å) | 0.013 |
| Bond angles (°) | 1.726 |
| Validation | |
| MolProbity score | 0.71 |
| Clashscore | 0 |
| Poor rotamers (%) | 1.44 |
| Ramachandran plot | |
| Favored (%) | 97.56 |
| Allowed (%) | 1.83 |
| Disallowed (%) | 0.61 |
Acknowledgements
We thank members of the Twomey and Huganir labs for insightful discussions. All cryo-EM data was collected at the Beckman Center for Cryo-EM at Johns Hopkins with assistance from D. Sousa and D. Ding.
Funding
E.C.T is supported by the Searle Scholars Program (Kinship Foundation #22098168) and the Diana Helis Henry Medical Research Foundation (#142548). R.L.H. is supported by National Institutes of Health (NIH) grants R01 NS036715 and R01 MH112152. W.D.H. is supported by NIH grant K99 MH132811.
Footnotes
Conflict of Interest
R.L.H. is scientific cofounder and Scientific Advisory Board (SAB) member of Neumora Therapeutics and SAB member of MAZE Therapeutics.
Data Availability
All cryo-EM reconstructions will be deposited into the Electron Microscopy Data Bank (EMDB) upon publication. All micrographs from the IS-1 and IS-2 datasets will be deposited into the Electron Microscopy Public Image Archive (EMPIAR) upon publication. All structural models generated from cryo-EM will be deposited in the Protein Data Bank upon publication.
References
- 1.Diering G. H. & Huganir R. L. The AMPA Receptor Code of Synaptic Plasticity. Neuron 100, 314–329 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hansen K. B. et al. Structure, Function, and Pharmacology of Glutamate Receptor Ion Channels. Pharmacol Rev 73, 298–487 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Twomey E. C., Yelshanskaya M. V. & Sobolevsky A. I. Structural and functional insights into transmembrane AMPA receptor regulatory protein complexes. J Gen Physiol 151, 1347–1356 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Letts V. A. et al. The mouse stargazer gene encodes a neuronal Ca2+-channel gamma subunit. Nat Genet 19, 340–347 (1998). [DOI] [PubMed] [Google Scholar]
- 5.Twomey E. C., Yelshanskaya M. V., Grassucci R. A., Frank J. & Sobolevsky A. I. Elucidation of AMPA receptor-stargazin complexes by cryo-electron microscopy. Science 353, 83–86 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Twomey E. C., Yelshanskaya M. V., Grassucci R. A., Frank J. & Sobolevsky A. I. Channel opening and gating mechanism in AMPA-subtype glutamate receptors. Nature 549, 60–65 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Twomey E. C., Yelshanskaya M. V., Grassucci R. A., Frank J. & Sobolevsky A. I. Structural Bases of Desensitization in AMPA Receptor-Auxiliary Subunit Complexes. Neuron 94, 569–580.e5 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Twomey E. C., Yelshanskaya M. V., Vassilevski A. A. & Sobolevsky A. I. Mechanisms of Channel Block in Calcium-Permeable AMPA Receptors. Neuron 99, 956–968.e4 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Zhao Y., Chen S., Swensen A. C., Qian W.-J. & Gouaux E. Architecture and subunit arrangement of native AMPA receptors elucidated by cryo-EM. Science 364, 355–362 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Yu J. et al. Hippocampal AMPA receptor assemblies and mechanism of allosteric inhibition. Nature 594, 448–453 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zhang D. et al. Modulatory mechanisms of TARP γ8-selective AMPA receptor therapeutics. Nat Commun 14, 1659 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Herguedas B. et al. Mechanisms underlying TARP modulation of the GluA1/2-γ8 AMPA receptor. Nat Commun 13, 734 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chen S. et al. Activation and Desensitization Mechanism of AMPA Receptor-TARP Complex by Cryo-EM. Cell 170, 1234–1246.e14 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zhao Y., Chen S., Yoshioka C., Baconguis I. & Gouaux E. Architecture of fully occupied GluA2 AMPA receptor-TARP complex elucidated by cryo-EM. Nature 536, 108–111 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Suzuki H. et al. Crystal Structure of a Claudin Provides Insight into the Architecture of Tight Junctions. Science (2014). [DOI] [PubMed] [Google Scholar]
- 16.Zihni C., Mills C., Matter K. & Balda M. S. Tight junctions: from simple barriers to multifunctional molecular gates. Nat Rev Mol Cell Biol 17, 564–580 (2016). [DOI] [PubMed] [Google Scholar]
- 17.Jumper J. et al. Highly accurate protein structure prediction with AlphaFold. Nature 596, 583–589 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Zhang D. et al. Structural mobility tunes signalling of the GluA1 AMPA glutamate receptor. Nature 1–6 (2023) doi: 10.1038/s41586-023-06528-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zhang D., Watson J. F., Matthews P. M., Cais O. & Greger I. H. Gating and modulation of a hetero-octameric AMPA glutamate receptor. Nature 594, 454–458 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Klykov O., Gangwar S. P., Yelshanskaya M. V., Yen L. & Sobolevsky A. I. Structure and desensitization of AMPA receptor complexes with type II TARP γ5 and GSG1L. Mol Cell 81, 4771–4783.e7 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gangwar S. P. et al. Modulation of GluA2-γ5 synaptic complex desensitization, polyamine block and antiepileptic perampanel inhibition by auxiliary subunit cornichon-2. Nat Struct Mol Biol 30, 1481–1494 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Emsley P. & Cowtan K. Coot: model-building tools for molecular graphics. Acta Cryst D 60, 2126–2132 (2004). [DOI] [PubMed] [Google Scholar]
- 23.Croll T. I. ISOLDE: a physically realistic environment for model building into low-resolution electron-density maps. Acta Crystallogr D Struct Biol 74, 519–530 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Liebschner D. et al. Macromolecular structure determination using X-rays, neutrons and electrons: recent developments in Phenix. Acta Cryst D 75, 861–877 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Williams C. J. et al. MolProbity: More and better reference data for improved all-atom structure validation. Protein Science 27, 293–315 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Pettersen E. F. et al. UCSF ChimeraX: Structure visualization for researchers, educators, and developers. Protein Science 30, 70–82 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Morin A. et al. Collaboration gets the most out of software. eLife 2, e01456 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Larkin M. A. et al. Clustal W and Clustal X version 2.0. Bioinformatics 23, 2947–2948 (2007). [DOI] [PubMed] [Google Scholar]
- 29.Procter J. B. et al. Alignment of Biological Sequences with Jalview. Methods Mol Biol 2231, 203–224 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All cryo-EM reconstructions will be deposited into the Electron Microscopy Data Bank (EMDB) upon publication. All micrographs from the IS-1 and IS-2 datasets will be deposited into the Electron Microscopy Public Image Archive (EMPIAR) upon publication. All structural models generated from cryo-EM will be deposited in the Protein Data Bank upon publication.