

Abstract
We evaluated changes in passive mechanical properties in cardiac tissues during rat pregnancy. Left and right ventricular free walls were dissected from hearts of nonpregnant, late-pregnant, and postpartum rats. Mechanical experiments in ventricular strips were done by stretch–release cycles using a step motor. The results show that during pregnancy, there is cardiac hypertrophy associated with (1) an increase in myocyte size, particularly of augmented myocyte length, (2) a decrease in passive tension developed by the myocardial walls, and (3) a decrease in both elastic modulus and hysteresis. All changes observed during rat pregnancy were reversed during postpartum. In conclusion, a heart with less ventricular rigidity could contribute to facilitating the ventricular filling in conditions of a greater circulating volume characteristic of pregnancy.
Keywords: Cardiac hypertrophy, Pregnancy, Passive tension
Introduction
Titin, also known as connectin, is a giant protein and the major contributor to the development of myocardial passive tension during ventricular diastole [1–5]. Titin spans from the Z disc to the M-line of the sarcomere. The I-band segment of titin functions as a molecular spring. Two classes of titin have been found in cardiac tissues and are referred to as N2B and N2BA [1, 6]. The I-band of both isoforms contains a PEVK region [segment rich in proline (P), glutamine (E), valine (V), and lysine (K) residues], a unique sequence (N2B/N2A), and immunoglobulin (Ig) domains [7]. The passive tension that the cardiac tissues develop depends on the expression levels of titin isoforms. The cardiac myocytes that only express N2BA titin isoform develop less passive tension than myocytes expressing N2B titin, while coexpression of both titin isoforms in the same sarcomere allows passive tension to develop to intermediate levels [6, 8].
Passive tension alterations have been found in nonfunctional cardiac hypertrophy, and these changes have been associated with variations in the expression of titin isoforms. Recently, it was reported that cardiac tissues of patients with dilated cardiomyopathy develop less passive tension than normal patients; this change was associated with an increase in the N2BA titin isoform expression [9, 10]. The increase in N2BA titin isoform was correlated with an increase in the final diastolic and systolic volumes. Another protein that contributes to the development of passive tension during muscle stretch is collagen, a main component of the extracellular matrix. In pathologic cardiac hypertrophy, collagen type I content increases, inducing an increase in ventricular stiffness [11–13].
Although several investigations associated with pathologic cardiac hypertrophy exist, there is little information about the mechanisms associated with functional cardiac hypertrophy, especially those that occur during pregnancy. During pregnancy there are dramatic increases in sex-hormone levels [14] and large hemodynamic changes including increase in circulation volume, cardiac output, stroke volume, and heart rate, and a decrease in the peripheral vascular resistance [15–17]. These changes require an adaptation of heart mechanical properties. Here, we evaluated the passive mechanical properties in heart ventricular tissues of nonpregnant, pregnant, and postpartum rats.
Methods
Animals
Animal care and experimental procedures were approved by the Ethics Committee of the University of Colima according to the “Guide for the Care and Use of Laboratory Animals” (US Department of Health, NIH). Female Sprague-Dawley rats were separated into three experimental groups: nonpregnant (NP, diestrus, 250 ± 4 g, n = 8), late-pregnant (LP, 18–21 days, 305 ± 13 g, n = 8), and postpartum (PP, 7 day, 271 ± 6 g, n = 8). Under anesthesia, hearts were extirpated and weighed for use in the following experimental procedures.
Myocyte size
Myocytes were isolated from rat cardiac ventricular tissues by digesting their extracellular matrix with collagenase and protease, according to a protocol previously published [18]. The myocyte area and length were measured with a microscope (Carl Zeiss, Goettingen, Germany) equipped with digital camera and Image 1.6 software (National Institute of Health, USA).
Passive mechanical properties
Muscle strips (length ≈ 4 mm, width ≈ 2 mm, and thickness ≈ 0.5 mm) of right (SRV) and left (SLV) ventricular free walls were dissected in oxygenated Tyrode solution (in mmol/L: NaCl 125, KCl 5.4, MgCl2 1.05, NaHCO3 24, NaH2PO4 0.42, CaCl2 1.8, and dextrose 11; pH = 7.4) supplemented with protease inhibitors. The resting tension of strips was adjusted to 500 mg [19], and then passive tension was recorded by imposing a ramp stretch and release [stretch was 10% of strip initial length (L i) to a velocity of 0.2 L i/s; sarcomere approximate length of work in our experiments was in the range of 1.9–2.2 µm] [20]. Tension recordings were analyzed by means of stress–strain relationship [21]. The stress–strain curves were fitted with the Magid and Law equation to estimate the elastic modulus of cardiac tissues [22]. The energy cost during the hysteresis process was obtained by the area under the stretch and release curves [23]. All experiments were performed at 37°C.
Statistics
Data are expressed as mean ± SE. Differences between groups (NP, LP, and PP) were estimated by the one-factor variance analysis, and multiple comparisons between means were obtained using Bonferroni test. A P ≤ 0.05 was considered statistically significant. Analysis was performed with Minitab software.
Results
Heart weight and cardiac myocyte size
During the late phase of rat pregnancy, there was a significant increase (about 15%) in heart-to-body weight ratio (see Table 1). Seven days postpartum, there was a decrease in the heart-to-body weight ratio, confirming reversibility. Also, in this study we explored changes in myocyte size induced by pregnancy. Myocyte area and length were different in both ventricles. During pregnancy (LP), myocytes from right and left ventricle free wall significantly increased both their area and length as compared with nonpregnant (NP) and postpartum (PP) groups (Table 1). In contrast, the cell width was similar in all cases. These data demonstrate that during rat pregnancy the heart develops eccentric cardiac hypertrophy, which is almost fully recovered after partum.
Table 1.
Cardiac myocyte morphology and heart mass in nonpregnant, late-pregnant, and postpartum rats
| Experimental groups | |||
|---|---|---|---|
| NP | LP | PP | |
| Myocyte total area (µm2) | |||
| RV | 2,461.1 ± 50.10 | 3,050.8 ± 40.50* | 2,741.2 ± 57.60 |
| LV | 2,484.3 ± 40.10 | 2,831.9 ± 38.70* | 2,683.8 ± 47.90 |
| Myocyte length (µm) | |||
| RV | 110.1 ± 1.00 | 133.4 ± 0.90* | 121.2 ± 0.90 |
| LV | 102.2 ± 0.80 | 114.9 ± 1.10* | 108.5 ± 1.10 |
| Heart mass (mg)/body weight (g) | 3.69 ± 0.05 | 4.34 ± 0.10* | 3.47 ± 0.09 |
RV Right ventricle, LV left ventricle, NP nonpregnant, LP late-pregnant, PP postpartum
* P < 0.05
Passive tension measurements
Figures 1 and 2 show that passive tension developed by SRV and SLV was significantly lower in the LP group than in the NP and PP groups (P < 0.05), and PP samples recovered their ability to generate. At 10% of L i, SRV developed 9.02 ± 0.57 mN/mm2, and SLV generated 7.48 ± 0.44 mN/mm2 in NP; while in LP generation of force was decreased to 5.39 ± 0.48 mN/mm2 in SRV, and SLV produced only 4.09 ± 0.61 mN/mm2. In PP, passive tension recovered to practically original levels as SRV elicited 8.60 ± 0.66 mN/mm2 and SLV produced 7.46 ± 0.84 mN/mm2 (P > 0.05) of passive tension. In all groups, the SLV developed lower passive tension than SRV as shown in Figs. 1 and 2.
Fig. 1.

Passive tension in strips of right ventricle from different groups. a Nonpregnant (NP) versus late-pregnant (LP), and b late-pregnant (LP) versus postpartum (PP). Values correspond to mean ± SE, n = 16 per group
Fig. 2.

Passive tension in strips of left ventricle from different groups. a Nonpregnant (NP) versus late-pregnant (LP), and b late-pregnant (LP) versus postpartum (PP). Values correspond to mean ± SE, n = 16 per group
Modulus of elasticity
To estimate cardiac tissue rigidity, the elastic modulus of each group was calculated using stress–strain curves fitted to the Magid and Law equation [22]:
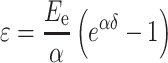 |
1 |
where ε is stress, δ strain, E e elastic modulus or Young modulus, and α corresponds to deformation rate. After taking logarithms, Eq. 1 becomes [when exp(σδ) >> 1]:
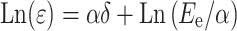 |
2 |
In SRV, the elastic moduli decreased from 31.1 ± 0.016 mN/mm2 in NP to 19.0 ± 0.015 mN/mm2 in LP and returned to 31.5 ± 0.014 mN/mm2 in PP. In SLV, the elastic moduli also decreased from 25.5 ± 0.015 mN/mm2 in NP to 14.8 ± 0.014 mN/mm2 in LP and returned to 25.4 ± 0.016 mN/mm2 in PP. These results indicate that the rigidity of cardiac tissues is less prominent in late-pregnant than in nonpregnant animals and returns to normal in postpartum conditions. In all conditions, the rigidity of SLV was smaller than SRV.
Hysteresis
Cardiac muscle exhibits the hysteresis phenomenon, where the developed tension during stretch is higher than during release. The hysteresis process is related to loss of elastic energy measured in joules. In line with a loss in rigidity during pregnancy, hysteresis was also decreased in the hypertrophied cardiac tissues of LP animals (Figs. 3, 4), both in SRV (NP = 154 ± 22 J, LP = 79 ± 21 J, PP = 172 ± 25 J) and SLV (NP = 98.7 ± 7 J, LP = 88 ± 3 J, PP = 103 ± 5 J). Energy cost during the hysteresis in SRV was higher than SLV.
Fig. 3.

Hysteresis curves in strips of right ventricle from different groups. a Nonpregnant (NP) versus late-pregnant (LP), and b late-pregnant (LP) versus postpartum (PP). Values correspond to mean ± SE, n = 16 per group
Fig. 4.
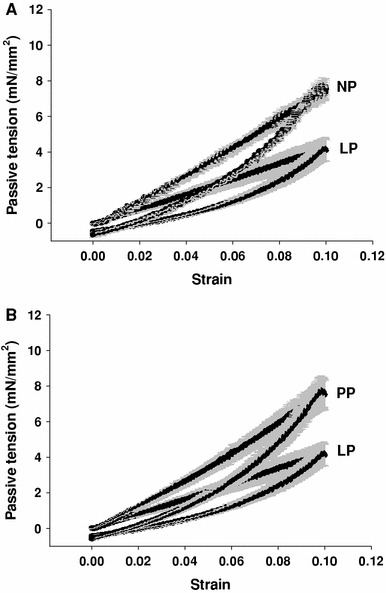
Hysteresis curves in strips of left ventricle from different groups. a Nonpregnant (NP) versus late-pregnant (LP), and b late-pregnant (LP) versus postpartum (PP). Values correspond to mean ± SE, n = 16 per group
Discussion
This study shows that rats, similar to mice [18], develop cardiac hypertrophy during pregnancy. We now show that pregnancy induces a reduction in passive tension, rigidity, and hysteresis as measured in muscle strips from free ventricular walls of the hypertrophied heart. The changes that cardiac tissues underwent during pregnancy were reversed in postpartum.
There are few reports about pregnancy-related physiological heart hypertrophy, therefore, physiological changes during pregnancy and molecular mechanisms associated with hypertrophy are not well understood. In late-pregnant mice, an increase in heart weight but a decrease in the heart-to-body weight ratio was observed [18]. In 16-day-pregnant rats, there was no significant change in heart-to-body weight when compared to nonpregnant rats (estrus cycle stage unknown) [24], while in this study we found that in late-pregnant rats (18–21 days) the heart-to-body weight ratio increased when compared to nonpregnant (diestrus) and postpartum animals.
Heart hypertrophy is a compensatory mechanism activated during pregnancy to improve the heart’s pumping capacity in response to an increased demand. At the same time, it is accepted that cardiac hypertrophy is associated with an increase in myocyte size [18, 25–27]. Our data also support this idea and demonstrate that the increase is due to an increase in myocyte length, explaining the eccentric cardiac hypertrophy observed in pregnancy. Eghbali et al. [18] also demonstrated eccentric cardiac hypertrophy in pregnant mice using echocardiography techniques.
Passive properties are an important factor in cardiac muscle function because part of the diastolic wall tension determines the extent of heart ventricular filling and its subsequent stroke volume. Physiological hypertrophy in pregnant rats is associated with a reduction in passive tension and rigidity of cardiac tissues. In pathological hypertrophy of dilated cardiomyopathy (DCM), the changes seem to differ depending on the species since an increase in passive tension is observed in birds (turkeys) and dogs, but in humans passive tension is reduced [9, 10, 13].
Both alterations were associated with variations in titin isoform expression levels [12, 13]. To test whether changes in passive tension observed during pregnancy may be related to an increase in the expression of N2BA titin, the heart samples were analyzed with SDS-PAGE (3–10% acrylamide gradient gels) and then Western blotting analysis was performed using primary antibodies specific to N2BA (X105–X106) and N2B titin (X150–X151), generously provided by Dr. Labeit (European Molecular Biology Laboratory, Heidelberg, Germany). The experiments performed in our laboratory did not show alterations in the expression level of titin isoforms during pregnancy. These results are similar to observations reported in functional cardiac hypertrophy induced by exercise in humans [28]. Nevertheless, the decrease in cardiac tissue passive tension in pregnancy could be regulated by an increase in the phosphorylation of cardiac titins. Protein kinase A-dependent phosphorylation of titin has been reported to contribute to a decrease in passive tension modulating the cardiac diastolic function. Recent experiments in humans also suggest that the protein kinase G-dependent phosphorylation of titin regulates changes in passive tension [29–32].
Another protein that regulates the ventricular passive tension is collagen, the main component of the extracellular matrix. In the heart, two collagen isoforms predominate, types I and III. Collagen type I forms a rigid structure that shows low distensibility, whereas collagen type III forms an elastic fine network. In pathologic cardiac hypertrophy, there is an increase in ventricular passive tension associated with an increase in collagen I content [11–13]. In rodents, collagen shows a greater contribution to passive tension than titin [33]. In the functional cardiac hypertrophy during pregnancy, a decrease in passive tension also could be explained by an increase in collagen type III, but this requires future experiments. Preliminary data from our laboratory in late pregnancy show that there is an increase in the amount of collagen in the heart (data not unpublished).
Our experiments show that passive tension of the hypertrophied ventricular tissues decreases during rat pregnancy. Furthermore, the ventricular elastic modulus indicates that cardiac walls are less stiff in pregnant rats than in PP and NP animals. In the rat, as in the mouse, it is clear that cardiac hypertrophy in late pregnancy results from an increase in cell and myofibril length, leading to an increase in ventricular volume and eccentric cardiac hypertrophy. Our observation of a parallel decrease in ventricular rigidity supports the idea that this diminution in rigidity could be the compensatory mechanism that operates in the heart during pregnancy to handle the blood volume overload that this physiological condition demands.
Another interesting variable is the hysteresis that originates when the tissues undergo cycles of stretch–release, a measurement of the energy lost as heat during each cycle [23, 34]. In pregnancy, we observed an increase in the heart rate and an increase in stretch–release cycles leading to the possibility of greater heat production. The data reveal a diminution of cardiac-tissue hysteresis during pregnancy, thereby diminishing loss of energy as heat in the subsequent cycle. Thus, during pregnancy the heart has a greater efficiency as compared with nonpregnancy and postpartum.
In conclusion, during pregnancy the longitudinal growth of myocytes leads to eccentric cardiac hypertrophy with less ventricular passive tension, less stiffness, and less hysteresis. Furthermore, all changes observed in pregnant rats were reversed during postpartum. Finally additional experiments are needed to elucidate the molecular mechanism that modulates the decrease in passive tension in cardiac hypertrophy during pregnancy.
Acknowledgments
This work was supported by UC-Mexus (2003 to L.T., J.M.), CONACyT-México (# 42065 M to J.M.), FRABA-UdeC (to J.M.), and CONACyT-México Doctoral Fellowship (#143228 to A. V.-O.).
Footnotes
J. L. Marin deceased: 16 November 2007.
References
- 1.Cazorla O, Freiburg A, Helmes M, Centner T, McNabb M, Wu Y, Trombitas K, Labeit S, Granzier H. Differential expression of cardiac titin isoforms and modulation of cellular stiffness. Circ Res. 2000;86:59–67. doi: 10.1161/01.res.86.1.59. [DOI] [PubMed] [Google Scholar]
- 2.Freiburg A, Trombitas K, Hell W, Cazorla O, Fougerousse F, Centner T, Kolmerer B, Witt C, Beckmann J-S, Gregorio C-C, Granzier H, Labeit S. Series of exon-skipping events in the elastic spring region of titin as the structural basis for myofibrillar elastic diversity. Circ Res. 2000;86:1114–1121. doi: 10.1161/01.res.86.11.1114. [DOI] [PubMed] [Google Scholar]
- 3.Labeit S, Gautel M, Lakey A, Trinick J. Towards a molecular understanding of titin. EMBO J. 1992;11:1711–1716. doi: 10.1002/j.1460-2075.1992.tb05222.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Squire J-M. Architecture and function in the muscle sarcomere. Curr Opin Struct Biol. 1997;7:247–257. doi: 10.1016/S0959-440X(97)80033-4. [DOI] [PubMed] [Google Scholar]
- 5.Trinick J, Tskhovrebova L. Titin: a molecular control freak. Trends Cell Biol. 1999;9:377–380. doi: 10.1016/S0962-8924(99)01641-4. [DOI] [PubMed] [Google Scholar]
- 6.Trombitas K, Wu Y, Labeit D, Labeit S, Granzier H. Cardiac titin isoforms are coexpressed in the half-sarcomere and extend independently. Am J Physiol Heart Circ Physiol. 2001;281:H1793–H1799. doi: 10.1152/ajpheart.2001.281.4.H1793. [DOI] [PubMed] [Google Scholar]
- 7.Lewinter M-M. Titin isoforms in heart failure are there benefits to supersizing? Circulation. 2004;110:109–111. doi: 10.1161/01.CIR.0000137284.17083.93. [DOI] [PubMed] [Google Scholar]
- 8.Granzier H-l, Labeit S. Cardiac titin: an adjustable multi-functional spring. J Physiol. 2002;41:335–342. doi: 10.1113/jphysiol.2001.014381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Nagueh S-F, Shah G, Wu Y, Torre-Amione G, King N-M, Lahmers S, Witt C-C, Becker K, Labeit S, Granzier H-l. Altered titin expression, myocardial stiffness, and left ventricular function in patients with dilated cardiomyopathy. Circulation. 2004;10:155–162. doi: 10.1161/01.CIR.0000135591.37759.AF. [DOI] [PubMed] [Google Scholar]
- 10.Neagoe C, Kulke M, Del Monte F, Gwathmey J-K, De Tombe P-P, Hajjar R-J, Linke W-A. Titin isoform switch in ischemic human heart disease. Circulation. 2002;106:1333–1341. doi: 10.1161/01.CIR.0000029803.93022.93. [DOI] [PubMed] [Google Scholar]
- 11.Marijianowski M-M, Teeling P, Mann J, Becker A-E. Dilated cardiomyopathy is associated with an increase in the type I/type III collagen ratio: a quantitative assessment. J Am Coll Cardiol. 1995;25:1263–1272. doi: 10.1016/0735-1097(94)00557-7. [DOI] [PubMed] [Google Scholar]
- 12.Wu Y, Bell S-P, Trombitas K, Witt C-C, Labeit S, Lewinter M-M, Granzier H. Changes in titin isoform expression in pacing-induced cardiac failure give rise to increased passive muscle stiffness. Circulation. 2002;106:1384–1389. doi: 10.1161/01.CIR.0000029804.61510.02. [DOI] [PubMed] [Google Scholar]
- 13.Wu Y, Tobias A-H, Bell K, Barry W, Helmes M, Trombitas K, Tucker R, Campbell K-B, Granzier H-l. Cellular and molecular mechanisms of systolic and diastolic dysfunction in an avian model of dilated cardiomyopathy. J Mol Cell Cardiol. 2004;37:111–119. doi: 10.1016/j.yjmcc.2004.04.010. [DOI] [PubMed] [Google Scholar]
- 14.LaPolt P-S, Matt D-W, Judd H-L, Lu J-K. The relation of ovarian steroid levels in young female rats to subsequent estrous cyclicity and reproductive function during aging. Biol Reprod. 1986;35:1131–1139. doi: 10.1095/biolreprod35.5.1131. [DOI] [PubMed] [Google Scholar]
- 15.Katz R, Karliner J-S, Resnik R. Effects of a natural volume overload state (pregnancy) on left ventricular performance in normal human subjects. Circulation. 1978;58:434–441. doi: 10.1161/01.cir.58.3.434. [DOI] [PubMed] [Google Scholar]
- 16.Poppas A, Shroff S-G, Korcarz C-E, Hibbard JU, Berger D-S, Lindheimer M-D, Lang R-M. Serial assessment of the cardiovascular system in normal pregnancy. Role of arterial compliance and pulsatile arterial load. Circulation. 1997;95:2407–2415. doi: 10.1161/01.cir.95.10.2407. [DOI] [PubMed] [Google Scholar]
- 17.Spaanderman ME, Meertens M, Van Bussel M, Ekhart T-H, Peeters L-l. Cardiac output increases independently of basal metabolic rate in early human pregnancy. Am J Physiol Heart Circ Physiol. 2000;278:H1585–H1588. doi: 10.1152/ajpheart.2000.278.5.H1585. [DOI] [PubMed] [Google Scholar]
- 18.Eghbali M, Deva R, Alioua A, Minosyan T-Y, Ruan H, Wang Y, Toro L, Stefani E. Molecular and functional signature of heart hypertrophy during pregnancy. Circ Res. 2005;96:1208–1216. doi: 10.1161/01.RES.0000170652.71414.16. [DOI] [PubMed] [Google Scholar]
- 19.Cameron N-E, Cotter M-A, Robertson S. Contractile properties of cardiac papillary muscle in streptozotocin-diabetic rats and the effects of aldose reductase inhibition. Diabetologia. 1989;32:365–370. doi: 10.1007/BF00277260. [DOI] [PubMed] [Google Scholar]
- 20.Granzier H-L, Irving T-C. Passive tension in cardiac muscle: contribution of collagen, titin, microtubules, and intermediate filaments. Biophys J. 1995;68:1027–1044. doi: 10.1016/S0006-3495(95)80278-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Muñiz J, Del Rio J, Huerta M, Marin J-L. Effects of sprint and endurance training on passive stress–strain relation of fast- and slow-twitch skeletal muscle in Wistar rat. Acta Physiol Scand. 2001;173:207–212. doi: 10.1046/j.1365-201X.2001.00875.x. [DOI] [PubMed] [Google Scholar]
- 22.Magid A, Law D-J. Myofibrils bear most of the resting tension in frog skeletal muscle. Science. 1985;230:1280–1282. doi: 10.1126/science.4071053. [DOI] [PubMed] [Google Scholar]
- 23.Minajeva A, Kulke M, Fernandez J-M, Linke W-A. Unfolding of titin domains explains the viscoelastic behavior of skeletal myofibrils. Biophys J. 2001;80:1442–1451. doi: 10.1016/S0006-3495(01)76116-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Gonzalez A-M, Osorio J-C, Manlhiot C, Gruber D, Homma S, Mital S. Hypertrophy signaling during peripartum cardiac remodeling. Am J Physiol Heart Circ Physiol. 2007;293:H3008–H3013. doi: 10.1152/ajpheart.00401.2007. [DOI] [PubMed] [Google Scholar]
- 25.Chen Y, Torry R-J, Baumbach G-l, Tomanek R-J. Proportional arteriolar growth accompanies cardiac hypertrophy induced by volume overload. Am J Physiol Heart Circ Physiol. 1994;267:H2132–H2137. doi: 10.1152/ajpheart.1994.267.6.H2132. [DOI] [PubMed] [Google Scholar]
- 26.Natali A, Turner D, Harrison S, White E. Regional effects of voluntary exercise on cell size and contraction-frequency responses in rat cardiac myocytes. Exp Biol. 2001;204:1191–1199. doi: 10.1242/jeb.204.6.1191. [DOI] [PubMed] [Google Scholar]
- 27.Wang X, Ren B, Liu S, Sentex E, Tappia P-S, Dhalla N-S. Characterization of cardiac hypertrophy and heart failure due to volume overload in the rat. J Appl Physiol. 2003;94:752–763. doi: 10.1152/japplphysiol.00248.2002. [DOI] [PubMed] [Google Scholar]
- 28.Natali A-J, Wilson L-A, Pechham M, Turner D-L, Harrison S-M, White E. Different regional effects of voluntary exercise on the mechanical and electrical properties of rat ventricular myocytes. J Physiol. 2002;541:863–875. doi: 10.1113/jphysiol.2001.013415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Fukuda N, Wu Y, Nair P, Granzier H-L. Phosphorylation of titin modulates passive stiffness of cardiac muscle in a titin isoform-dependent manner. J Gen Physiol. 2005;125:257–271. doi: 10.1085/jgp.200409177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Yamasaki R, Wu Y, McNabb M, Greaser M, Labeit S, Granzier H. Protein kinase A phosphorylates titin’s cardiac-specific N2B domain and reduces passive tension in rat cardiac myocytes. Circ Res. 2002;90:1181–1188. doi: 10.1161/01.RES.0000021115.24712.99. [DOI] [PubMed] [Google Scholar]
- 31.Fukuda N, Granzier H-L, Ishiwata S, Kurihara S. Physiological functions of the giant elastic protein titin in mammalian striated muscle. J Physiol Sci. 2008;58:151–159. doi: 10.2170/physiolsci.RV005408. [DOI] [PubMed] [Google Scholar]
- 32.Krüger M, Kötter S, Grützner A, Lang P, Andresen C, Redfield M-M, Butt E, dos Remedios C-G, Linke W-A. Protein kinase G modulates human myocardial passive stiffness by phosphorylation of the titin springs. Circ Res. 2009;104:87–94. doi: 10.1161/CIRCRESAHA.108.184408. [DOI] [PubMed] [Google Scholar]
- 33.Wu Y, Cazorla O, Labeit D, Labeit S, Granzier H. Changes in titin and collagen underlie diastolic stiffness diversity of cardiac muscle. J Mol Cell Cardiol. 2000;32:2151–2162. doi: 10.1006/jmcc.2000.1281. [DOI] [PubMed] [Google Scholar]
- 34.Helmes M, Trombitas K, Centner T, Kellermayer M, Labeit S, Linke W-A, Granzier H. Mechanically driven contour-length adjustment in rat cardiac titin’s unique N2B sequence: titin is an adjustable spring. Circ Res. 1999;84:1339–1352. doi: 10.1161/01.res.84.11.1339. [DOI] [PubMed] [Google Scholar]