

Abstract
Recent studies have suggested that increased oxidative stress is a potential etiology in Parkinson’s disease (PD). In this study, we investigated whether acupuncture regulates antioxidants in the striatum (ST) of a PD mouse model. Male C57BL/6 mice were administered 30 mg/kg of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) intraperitoneally once a day for 5 days and given acupuncture stimulation at SI3 or GB34 (Yanglingquan) was for 12 consecutive days. Dopaminergic neuronal survival in the nigrostriatal pathway and DJ-1 expression in the ST was evaluated by immunostaining, and the activities of superoxide dismutase (SOD) and catalase (CAT) in the ST was by enzyme-linked immunosorbent assay. MPTP administration induced dopaminergic neuronal death in the nigrostriatal pathway, which was suppressed by acupuncture stimulation at GB34. MPTP administration also suppressed DJ-1 expression and SOD and CAT activities in the ST, which were restored by acupuncture stimulation at GB34. These results indicate that the neuroprotective effect of acupuncture stimulation is due to regulation of the antioxidants.
Keywords: Parkinson’s disease; Acupuncture; Antioxidant; DJ-1; Superoxide dismutase; Catalase; 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine
Introduction
Parkinson’s disease (PD) is a representative neurodegenerative disorder that is accompanied by symptoms such as rigidity, postural instability, akinesia, bradykinesia, and tremor [1]. From an etiological point of view, PD is generally caused by dopaminergic neuronal degeneration in the nigrostriatal pathway, and it is well known that it is influenced by several factors including environmental conditions and genetics. Moreover, recent studies have suggested that increased oxidative stress is a potential etiology in PD [2].
To induce PD-like symptoms and neuronal death, a few neurotoxins that restrain complex I of the mitochondrial respiratory chain are used. Among them, 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) is well recognized for causing the traits of sporadic PD [3]. When MPTP is absorbed by an organism, it passes through the brain because it can penetrate the blood–brain-barrier. In the brain, monoamine oxidase B converts MPTP to 1-methyl-4-phenylpyridinium (MPP+), which is a toxic metabolite that subsequently permeates into dopaminergic neurons through dopamine transporters. The MPP+ in the dopaminergic neurons deters the complex I activation, which induces oxidative stress and apoptosis in sequence, leading to development of PD symptoms [4]. Oxidative stress also plays a significant role in the onset and progress of PD [5]; therefore, suppression of oxidative stress can alleviate PD symptoms and protect against dopaminergic neuronal death in PD patients and animal models.
Acupuncture has been broadly applied to treat various diseases [6], and there have been several reports of electroacupuncture being able to enhance antioxidants. For example, Kim et al. showed that 100-Hz electroacupuncture at GB34 (Yanglingquan) and GB39 enhances cytochrome c oxidase [7], while Lv et al. reported that 100-Hz electroacupuncture alleviates nuclear factor-E2-related factor-2 and heme oxygenase-1 [8], and Wang et al. suggested that electroacupuncture stimulation at ST36 and SP6 increases glutathione concentration and superoxide dismutase (SOD) activity [9]. Taken together, these studies indicate that 100-Hz electroacupuncture stimulation enhances antioxidant activities in the striatum (ST) of MPTP-administrated mice, but it is still not clear whether manual acupuncture stimulation can also enhance them.
The GB34 acupoint has been used to cure a variety of diseases in lower limbs, muscles, and the gall bladder, as well as paralysis, and recent studies have confirmed the effects of acupuncture stimulation at GB34 on PD animal models [10]. For example, acupuncture stimulation at GB34 suppressed MPTP-induced behavioral impairment and dopaminergic neuronal destruction in the substantia nigra (SN) and ST by regulating the PI3K-Akt pathway [11] and enhancing synaptic dopamine availability [12]. However, although several studies have shown the effects of acupuncture on dopaminergic neuronal degeneration in PD mouse models, it is still not clear if manual acupuncture at GB34 alleviates MPTP-induced oxidative stress in the ST. Therefore, this study investigated the potential for acupuncture stimulation to improve motor function, prevent dopaminergic neuronal death, and alleviate the levels of anti-oxidants including DJ-1, SOD, and catalase (CAT) in the ST of MPTP-treated mice.
Materials and methods
Animals
This study was conducted with the approval of the Pusan National University Institutional Animal Care and Use Committee. We purchased 8-week-old male C57BL/6 mice weighing 20–23 g from Orientbio Inc. (Seongnam, Korea) and housed them at room temperature (22 ± 2 °C) under a standard 12-h light/dark cycle (lights on at 07:00 h) with food and water provided ad libitum.
Group classification
Mice were randomly placed into four groups of 16: a saline-injected group (saline), a MPTP injected group (MPTP), a MPTP-injected plus acupuncture at SI3 (Houxi) acupoint group (MPTP + SI3), and a MPTP-injected plus acupuncture at GB34 acupoint group (MPTP + GB34).
MPTP injection
Except for the saline group, all of the mice were injected with MPTP-HCl (30 mg/kg of free base; Sigma, St. Louis, MO, USA) in sterilized normal saline (vehicle) intraperitoneally at 24-h intervals for 5 days. The mice in the saline group were injected with vehicle intraperitoneally following the same schedule [13].
Acupuncture treatment
Two hours after MPTP injection, mice in the MPTP + SI3 and MPTP + GB34 groups received acupuncture stimulation at SI3 (MPTP + SI3 group) or GB34 (MPTP + GB34 group). SI3 is located at the ulnar end of the distal palmar crease proximal to the 4th metacarpal phalangeal joint at the junction of the red and white skin, and GB34 is located at the point at which lines from the anterior borders to the head of the fibula intersect. Mice in the MPTP + SI3 group were lightly immobilized, after which stainless-steel acupuncture needles (0.18 mm × 8 mm; Dongbang acupuncture, Boryeng, Republic of Korea) were inserted to a depth of 2 mm at the SI3 acupoint, turned at a rate of two revolutions per second for 30 s, and then immediately removed. The acupuncture stimulation was performed from the left to right acupoint in sequence (treatment lasting 60 s). Mice in the MPTP + GB34 group underwent the same procedure, except they received acupuncture stimulation at GB34. Mice in the saline and the MPTP groups were also lightly immobilized for 60 s without acupuncture, then returned to their cage. This procedure continued at 24-h intervals for 12 consecutive days.
Behavioral test (pole test)
A modified version of the method established by Abe et al. [14] was applied for the pole test. Mice were positioned head-downwards near the top of a rough-surfaced wood pole (1 cm in diameter and 55 cm in height), after which the time taken to arrive at the bottom of the pole was measured. The test was repeated three times at 30-s time intervals, and behavioral changes were evaluated based on the average of the three times. The test was performed 1 day before the first MPTP administration (day 0), 2 h after the fifth acupuncture stimulation on day 5, and 2 h after the last acupuncture stimulation on day 12.
Immunofluorescent staining
After the pole test was conducted on day 12, mice (n = 6 at each group) were anesthetized with isoflurane (JW Pharmaceutical, Seoul, Republic of Korea) and perfused with 4% paraformaldehyde in 0.1 M phosphate buffer. The brain was then quickly removed and postfixed in 4% paraformaldehyde solution for 48 h, after which it was washed in 0.1 M phosphate buffer, then immersed in 30% sucrose solution for storage at 4 °C until sectioning. The brains were cut to a thickness of 30 μm using a Leica CM3050S cryostat (Leica Microsystems, Wetzlar, Germany). The SN sections located between AP −3.08 and −3.28 mm from the bregma and the ST sections located between AP +0.48 and +0.68 mm from the bregma were incubated with 1% H2O2 in 0.05 M phosphate-buffered saline for 15 min, followed by 0.3% Triton X-100 and 3% normal blocking serum in PBS at room temperature for 1 h, then stained overnight at room temperature using a primary antibody. An anti-tyrosine hydroxylase (TH, 1:1000; Santa Cruz Biotechnology, Santa Cruz, CA, USA), an anti-caspase-3 antibody (1:1000; Abcam, Cambridge, UK) and an anti-DJ-1 antibody (1:100; Cell Signaling Technology, Danvers, MA, USA) were used as a primary antibody. On the following day, the sections were reacted with anti-rabbit Alexa-488 IgG for 1 h at room temperature. The tissues were then mounted on gelatin-coated slides, air-dried, dehydrated, and coverslipped.
Histological features were captured using an Axio Observer Z1 microscope (Carl ZEISS, Oberkochen, Germany). The number of TH-positive neuronal cells in the SN was counted to evaluate the survival of dopaminergic neurons. An independent observer who was not aware of the expected test results manually counted the TH-positive neurons bilaterally in five continuous SN sections. The number of cells was counted and confirmed three times to validate the data. To evaluate the changes in dopaminergic neuronal fibers in the ST, the mean values of optical density (OD) of the TH, DJ-1, and caspase-3 in the ST were determined using Image-Pro Plus 6.0 (Media Cybernetics, Silver Spring, MD, USA).
Western blot
To confirm immunofluorescent data, we performed Western-blot analysis. Mice (n = 4 at each group) were sacrificed with CO2 gas on day 12, then ST tissues were dissected rapidly and kept at −80 °C until use. The tissues were homogenized with protease inhibitor and RIPA buffer, centrifuged for 20 min at 4 °C at 12,000 rpm, and the supernatants were separated. Equal amounts of protein (30 µg) from each sample were separated on 12% SDS–polyacrylamide gels and transferred to nitrocellulose membranes. The membrane was blocked with 5% bovine serum albumin in Tris-buffered saline containing 0.1% Tween-20 for 1 h at room temperature and incubated overnight with anti-DJ-1 (Abcam), anti-caspase-3, anti-phosphorylated AKT and anti-AKT (Cell Signaling Technology) antibodies that were diluted 1:1000 in blocking solution antibody at 4 °C. Then the membrane was washed and incubated with horseradish peroxidase-conjugated secondary anti-rabbit antibody (1:2000, Santa Cruz Biotechnology) at room temperature for 1 h. After washing the membrane, bands were detected using the enhanced chemiluminescence detection kit (Thermo Scientific, Rockford, IL, USA). Then, these blots were re-probed with rabbit monoclonal anti-β-actin antibody (1:1000; Santa Cruz Biotechnology) as a loading control for all experiments. Quantification of immunoreactivity corresponding to the total bands was performed by densitometric analysis using an Image Quant LAS 4000 (Fujifilm, Tokyo, Japan).
Assessment of superoxide dismutase and catalase in mouse brain
Mice (n = 6 at each group) were sacrificed and their STs were immediately enucleated after completion of the pole test on day 12. Collected ST tissues were homogenized and centrifuged, then the supernate was collected and stored at −80 °C until assayed. The activities of SOD and CAT in the ST were detected using enzyme-linked immunosorbent assay (ELISA) kits (Cusabio Biotech, Wuhan, China) according to the manufacturer’s instructions. Briefly, after 100 μl of sample per well was added and incubated for 2 h at 37 °C, the liquid of each well was removed. Then 100 μl of biotin-antibody was added to each well and incubated for 1 h at 37 °C. After each well was aspirated and washed, 100 μl of horseradish peroxidase-avidin was added and incubated for 1 h at 37 °C. After being washed, 90 μl of 3,3′,5,5′-tetramethylbenzidine substrate was added, incubated for 3 min at 37 °C with light protection, 50 μl of stop solution was added to each well. Immediately the SOD and the CAT activities were measured spectrofluorometrically using a multi-well plate reader, and the value of subtracted readings at 540 nm from the readings at 450 nm was used.
Statistical analysis
All of data were expressed as the means ± the standard errors of mean. All data were analyzed by one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test using Prism 5 for Windows (GraphPad Software Inc., La Jolla, CA, USA). A p < 0.05 was considered to indicate significance.
Results
Effects of acupuncture on MPTP-induced behavioral impairment
The pole test was conducted to verify the efficacy of acupuncture on behavioral impairment by MPTP [14]. No significant differences were observed in the descending time of the mice before MPTP administration (day 0). On day 5, an ANOVA revealed a significant difference on the descending times among the groups [F(3,23) = 6.446, p < 0.01]. The descending times in all of MPTP-administrated groups were significantly increased relative to the vehicle-administrated (saline) group (p < 0.05). On day 12, an ANOVA revealed a significant difference on the descending times among the groups [F(3,23) = 6.929, p < 0.01]. The descending times in the MPTP (12.25 ± 1.32 s) and the MPTP + SI3 (11.75 ± 1.25 s) groups were still significantly higher than in the saline group (7.60 ± 0.75 s, p < 0.05 at each group), but that in the MPTP + GB34 group (7.25 ± 0.63 s) was significantly decreased compared to those in the MPTP and MPTP + SI3 groups (p < 0.05 at each group). There was no significant difference between the saline and the MPTP + GB34 groups (Fig. 1).
Fig. 1.
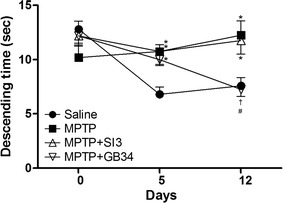
Pole test results. Acupuncture stimulation at GB34 alleviated 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced behavioral impairment on day 12. Saline saline injected group, MPTP MPTP injected group, MPTP + SI3 MPTP injected plus acupuncture at SI3 (Houxi) acupoint group, MPTP + GB34 MPTP injected plus acupuncture at GB34 (Yanglingquan) acupoint group. Data shown are the means ± the standard errors of means. *p < 0.05 versus the saline group, # p < 0.05 versus the MPTP group and † p < 0.05 versus the MPTP + SI3 group
Neuroprotective effects of acupuncture on MPTP-induced neuronal death in the SN and the ST
TH is a standard marker to identify dopaminergic neurons [15]. The neuroprotective effects of acupuncture on MPTP-induced neuronal death were evaluated by counting the number of TH-positive neurons in the SN and measuring the OD in the ST. An ANOVA revealed a significant effect on the TH-positive neurons in the SN [F(3,23) = 7.953, p < 0.001] and the ST [F(3,23) = 8.131, p < 0.001] among the groups. The numbers in the MPTP (72.12 ± 3.15%) and the MPTP + SI3 (72.39 ± 5.14%) groups were significantly lower than in the saline group (100 ± 6.47%, p < 0.01 at each group). However, the number in the MPTP + GB34 group (89.57 ± 4.63%) was significantly higher than in the MPTP and the MPTP + SI3 groups (p < 0.05 at each group), and there was no significant difference between the saline and the MPTP + GB34 groups.
The OD in the ST of mice was significantly lower in the MPTP (53.13 ± 4.96%, p < 0.001) and the MPTP + SI3 (62.64 ± 9.48%, p < 0.01) groups compared with the saline group (100.00 ± 5.05%). Conversely, that in the MPTP + GB34 group was significantly higher than in the MPTP group (79.06 ± 8.33%, p < 0.05), and there was no significant difference between the saline and the MPTP + GB34 groups (Fig. 2).
Fig. 2.
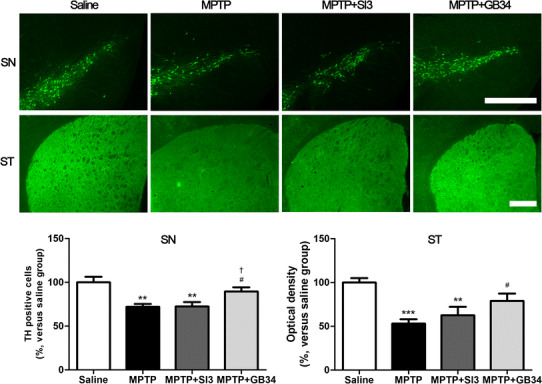
Effects of acupuncture stimulation on MPTP-induced dopaminergic neuronal death in the substantia nigra (SN) and the striatum (ST). MPTP administration induced dopaminergic neuronal death, whereas acupuncture stimulation at GB34 prevented it. Scale bar 200 µm. Data are shown as the means ± the standard errors of means. **p < 0.01 and ***p < 0.001 versus the saline group and # p < 0.05 versus the MPTP group and † p < 0.05 versus the MPTP + SI3 group
Change in caspase-3 expression in the ST
To confirm the level of MPTP-induced apoptosis, we investigated the caspase-3 expression in the ST because the caspase-3 is an indicator of apoptosis [16]. ANOVA revealed a significant difference on the caspase-3 expression among the groups [F(3,23) = 9.053, p < 0.001]. When compared with caspase-3 expression in the ST of mice in the saline group (100.00 ± 6.95%), the expression levels in the MPTP (129.00 ± 5.48%) and the MPTP + SI3 (117.00 ± 5.25%) groups were significantly increased (p < 0.01 and p < 0.05, respectively). However, the expression in the MPTP + GB34 group (93.58 ± 4.21%) was significantly decreased compared to the levels in the MPTP (p < 0.001) and the MPTP + SI3 (p < 0.05) groups, and there was no significant difference between the saline and the MPTP + GB34 groups (Fig. 3).
Fig. 3.
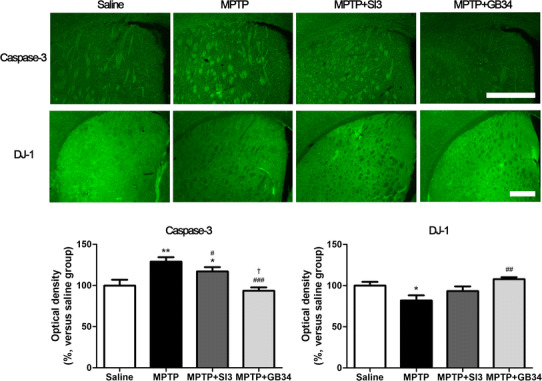
Changes in caspase-3 and DJ-1 expressions in the striatum in response to MPTP and acupuncture stimulations. MPTP administration increased caspase-3 expression and suppressed DJ-1 expression; however, acupuncture stimulation at GB34 alleviated them. Scale bar 200 µm. Data are shown as the means ± the standard errors of means. *p < 0.05 and **p < 0.01 versus the saline group, # p < 0.05, ## p < 0.01 and ### p < 0.001 versus the MPTP group, and † p < 0.05 versus the MPTP + SI3 group
Change in DJ-1 expression in the ST
We investigated DJ-1 expression in the ST because it is a PD-related protein [17] as well as an anti-oxidant [18]. ANOVA revealed a significant difference on the DJ-1 expression among the groups [F(3,23) = 5.420, p < 0.01]. When compared with the DJ-1 expression in the ST of mice in the saline group (100.00 ± 4.70%), significant reductions were found in the MPTP (82.02 ± 6.20%, p < 0.05). Conversely, that in MPTP + GB34 group was significantly higher than that in the MPTP group (107.80 ± 2.51%, p < 0.01), and there was no significant difference between the saline and the MPTP + GB34 groups (Fig. 3).
Confirmation of caspase-3 and DJ-1 expressions by Western-blot analysis
To verify the reliability of the immunofluorescent data, Western-blot analysis was performed. The results of triplicate Western blots for caspase-3 and DJ-1 were consistent with those of immunofluorescence staining (Fig. 4).
Fig. 4.
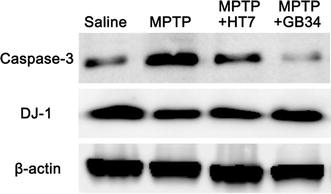
Confirmation of caspase-3 and DJ-1 expressions in the striatum by Western-blot analysis. The results of triplicate Western blots for caspase-3 and DJ-1 were consistent with those of immunofluorescence staining
Changes in SOD and CAT activities in the ST
Decreased endogenous antioxidants are risk factors causing early degeneration of dopaminergic neurons [8]. To confirm whether acupuncture regulates anti-oxidants in the ST, the activities of SOD and CAT were measured by ELISA. ANOVA revealed a significant difference on the SOD [F(3,23) = 6.156, p < 0.01] and the CAT [F(3,23) = 5.229, p < 0.01] expressions among the groups. When compared with the SOD activity in the ST of mice in the saline group (100.00 ± 7.20%), the activities in the MPTP (65.94 ± 6.36%) and the MPTP + SI3 (73.66 ± 7.47%) groups were significantly decreased (p < 0.05 at each group). However, that in the MPTP + GB34 group (92.98 ± 4.25%) was significantly higher than those in the MPTP and the MPTP + SI3 groups (p < 0.05 at each group), and there was no significant difference between the saline and the MPTP + GB34 groups (Fig. 5).
Fig. 5.
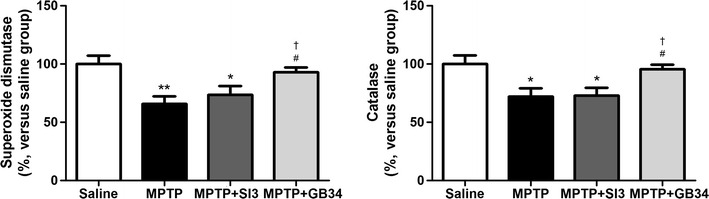
Changes in superoxide dismutase and catalase activities in the striatum. MPTP administration suppressed the activities, which were restored by acupuncture stimulation at GB34. *p < 0.05 and **p < 0.01 versus the saline group, # p < 0.05 versus the MPTP group, and † p < 0.05 versus the MPTP + SI3 group
Similar results were observed for CAT activity. When compared with the saline group (100.00 ± 7.44%), significant reductions in CAT activity were found in the MPTP (72.11 ± 7.13%, p < 0.05) and the MPTP + SI3 (73.00 ± 6.54%, p < 0.05) groups. On the other hand, the activity in the MPTP + GB34 group was significantly higher than in the MPTP and the MPTP + SI3 groups (95.51 ± 3.97%, p < 0.05 at each group). There was no significant difference between the saline and the MPTP + GB34 groups (Fig. 5).
Discussion
Previous studies have shown that acupuncture stimulation at GB34 suppresses behavioral impairment and dopaminergic neuronal death from MPTP toxicity. This present study also demonstrated that acupuncture stimulation improved MPTP-induced behavioral impairment, protected against dopaminergic neuronal death and suppressed caspase-3 expression. In addition, it could increase the level of DJ-1 and the activities of SOD and CAT in the ST.
DJ-1, also known as PD protein 7, has been recognized as a major protein in the pathogenesis of PD. Previous studies have shown that DJ-1 inhibits the aggregation of α-synuclein by regulating chaperone activity under oxidative conditions [17], while it protects dopaminergic neurons in the nigrostriatal pathway against MPTP-induced oxidative stress [18], and that the inhibition of DJ-1 causes dopaminergic neuronal degeneration via increases in reactive oxygen species [19]. Based on these results, DJ-1 plays a crucial role in the pathogenesis of PD by regulating oxidative stress [20]. In the present study, MPTP administration suppressed DJ-1 expression in the ST, which was alleviated by acupuncture stimulation at GB34. These results indicate that the enhancement of DJ-1 expression by acupuncture stimulation at GB34 influences the neuroprotective effect of the acupuncture stimulation.
A recent study showed that modulation of the Akt pathway is essential for DJ-1 activity, which protects dopaminergic neurons in the nigrostriatal pathway against MPTP toxicity [18]. Interestingly, previous studies have shown that the phosphatidylinositol 3-kinase (PI3K)/Akt signaling pathway plays a central role in the mechanism underlying acupuncture-induced benefits. For example, acupuncture increased Akt phosphorylation in the brain after traumatic brain injury [21] and electroacupuncture ameliorated acute renal injury in lipopolysaccharide-stimulated rabbits [22] and exerted an anti-apoptotic effect on cerebral ischemia–reperfusion injury [23] through modulating PI3K/Akt pathway. Moreover, Kim et al. reported that PI3K-Akt signaling pathway mediates acupuncture-induced dopaminergic neuronal protection in the brains of MPTP-treated mice [11]. Therefore, the increase of DJ-1 observed in this study may have been caused by the increase of Akt phosphorylation induced by acupuncture stimulation at GB34.
MPTP is widely used for PD research. Following administration, it is metabolized into 1-methyl-4-phenylpyridinum by monoamine oxidase B, which leads to increased oxidative stress that causes neuronal death [24], as well as interruption of antioxidant activities including those of SOD and CAT [25]. The combination of increased oxidative stress and decreased endogenous antioxidants is a major risk factor causing early degeneration of dopaminergic neurons [8]. In the present study, MPTP administration suppressed the activities of SOD and CAT, which is assumed to contribute to neuronal death. Conversely, acupuncture stimulation at GB34 alleviated the activities, indicating that the enhanced activities of SOD and CAT following acupuncture stimulation play an important role in preventing the MPTP-induced dopaminergic neuronal death in the ST. Moreover, DJ-1 can directly modulate SOD activity and may influence CAT activity [26]. Therefore, it is possible that the increase of DJ-1 induced by acupuncture stimulation at GB34 may influence the enhancement of SOD and CAT activities in the ST of MPTP-treated mice. However, additional study is needed to confirm this.
Conclusions
MPTP administration caused behavioral impairment, dopaminergic neuronal death in the nigrostriatal pathway, reduction of DJ-1 expression, and suppression of SOD and CAT activities in the ST. These changes were alleviated by acupuncture stimulation at GB34, indicating that the neuroprotective effect of acupuncture stimulation is due to regulation of the antioxidants. Taken together, these findings indicate that acupuncture stimulation at GB34 can be an alternative therapy for PD.
Acknowledgements
This work was supported by a National Research Foundation of Korea (NRF) grant funded by the Korean government (MSIP) (No. NRF-2014R1A5A2009936).
Compliance with ethical standards
Conflict of interest
The authors declare that they have no conflicts of interest.
Footnotes
Yukyung Lee and Gaeun Choi contributed equally to this work.
References
- 1.Kim D, Jeon H, Ryu S, Koo S, Ha KT, Kim S. Proteomic analysis of the effect of Korean red ginseng in the striatum of a Parkinson’s disease mouse model. PLoS One. 2016;11:e0164906. doi: 10.1371/journal.pone.0164906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kim GH, Kim JE, Rhie SJ, Yoon S. The role of oxidative stress in neurodegenerative diseases. Exp Neurobiol. 2015;24:325–340. doi: 10.5607/en.2015.24.4.325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Dawson TM, Dawson VL. Molecular pathways of neurodegeneration in Parkinson’s disease. Science. 2003;302:819–822. doi: 10.1126/science.1087753. [DOI] [PubMed] [Google Scholar]
- 4.Zuo L, Motherwell MS. The impact of reactive oxygen species and genetic mitochondrial mutations in Parkinson’s disease. Gene. 2013;532:18–23. doi: 10.1016/j.gene.2013.07.085. [DOI] [PubMed] [Google Scholar]
- 5.Jiang T, Sun Q, Chen S. Oxidative stress: a major pathogenesis and potential therapeutic target of antioxidative agents in Parkinson’s disease and Alzheimer’s disease. Prog Neurobiol. 2016;147:1–19. doi: 10.1016/j.pneurobio.2016.07.005. [DOI] [PubMed] [Google Scholar]
- 6.Bae CH, Kim DS, Jun YL, Kwon S, Park HJ, Hahm DH, Lee H, Kim ST. Proteomic analysis of the effect of acupuncture on the suppression of kainic acid-induced neuronal destruction in mouse hippocampus. Evid Based Complement Alternat Med. 2013;2013:436315. doi: 10.1155/2013/436315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kim ST, Moon W, Chae Y, Kim YJ, Lee H, Park HJ. The effect of electroaucpuncture for 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced proteomic changes in the mouse striatum. J Physiol Sci. 2010;60:27–34. doi: 10.1007/s12576-009-0061-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Lv E, Deng J, Yu Y, Wang Y, Gong X, Jia J, Wang X. Nrf2-ARE signals mediated the anti-oxidative action of electroacupuncture in an MPTP mouse model of Parkinson’s disease. Free Radic Res. 2015;49:1296–1307. doi: 10.3109/10715762.2015.1067696. [DOI] [PubMed] [Google Scholar]
- 9.Wang H, Pan Y, Xue B, Wang X, Zhao F, Jia J, Liang X, Wang X. The antioxidative effect of electro-acupuncture in a mouse model of Parkinson’s disease. PLoS One. 2011;6:e19790. doi: 10.1371/journal.pone.0019790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kwon S, Seo BK, Kim S. Acupuncture points for treating Parkinson’s disease based on animal studies. Chin J Integr Med. 2016;22:723–727. doi: 10.1007/s11655-016-2525-y. [DOI] [PubMed] [Google Scholar]
- 11.Kim SN, Kim ST, Doo AR, Park JY, Moon W, Chae Y, Yin CS, Lee H, Park HJ. Phosphatidylinositol 3-kinase/Akt signaling pathway mediates acupuncture-induced dopaminergic neuron protection and motor function improvement in a mouse model of Parkinson’s disease. Int J Neurosci. 2011;121:562–569. doi: 10.3109/00207454.2011.591515. [DOI] [PubMed] [Google Scholar]
- 12.Kim SN, Doo AR, Park JY, Bae H, Chae Y, Shim I, Lee H, Moon W, Lee H, Park HJ. Acupuncture enhances the synaptic dopamine availability to improve motor function in a mouse model of Parkinson’s disease. PLoS One. 2011;6:e27566. doi: 10.1371/journal.pone.0027566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Jun YL, Bae CH, Kim D, Koo S, Kim S. Korean red ginseng protects dopaminergic neurons by suppressing the cleavage of p35 to p25 in a Parkinson’s disease mouse model. J Ginseng Res. 2015;39:148–154. doi: 10.1016/j.jgr.2014.10.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Abe K, Taguchi K, Wasai T, Ren J, Utsunomiya I, Shinohara T, Miyatake T, Sano T. Biochemical and pathological study of endogenous 1-benzyl-1,2,3,4-tetrahydroisoquinoline-induced parkinsonism in the mouse. Brain Res. 2001;907:134–138. doi: 10.1016/S0006-8993(01)02573-2. [DOI] [PubMed] [Google Scholar]
- 15.White RB, Thomas MG. Moving beyond tyrosine hydroxylase to define dopaminergic neurons for use in cell replacement therapies for Parkinson’s disease. CNS Neurol Disord Drug Targets. 2012;11:340–349. doi: 10.2174/187152712800792758. [DOI] [PubMed] [Google Scholar]
- 16.Florentin A, Arama E. Caspase levels and execution efficiencies determine the apoptotic potential of the cell. J Cell Biol. 2012;196:513–527. doi: 10.1083/jcb.201107133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Zhou W, Zhu M, Wilson MA, Petsko GA, Fink AL. The oxidation state of DJ-1 regulates its chaperone activity toward α-synuclein. J Mol Biol. 2006;356:1036–1048. doi: 10.1016/j.jmb.2005.12.030. [DOI] [PubMed] [Google Scholar]
- 18.Aleyasin H, Rousseaux MW, Marcogliese PC, Hewitt SJ, Irrcher I, Joselin AP, Parsanejad M, Kim RH, Rizzu P, Callaghan SM, Slack RS, Mak TW, Park DS. DJ-1 protects the nigrostriatal axis from the neurotoxin MPTP by modulation of the AKT pathway. Proc Natl Acad Sci USA. 2010;107:3186–3191. doi: 10.1073/pnas.0914876107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zhou W, Freed CR. DJ-1 up-regulates glutathione synthesis during oxidative stress and inhibits A53T α-synuclein toxicity. J Biol Chem. 2005;280:43150–43158. doi: 10.1074/jbc.M507124200. [DOI] [PubMed] [Google Scholar]
- 20.Ariga H, Takahashi-Niki K, Kato I, Maita H, Niki T, Iguchi-Ariga SM. Neuroprotective function of DJ-1 in Parkinson’s disease. Oxid Med Cell Longev. 2013;2013:683920. doi: 10.1155/2013/683920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Li X, Chen C, Yang X, Wang J, Zhao ML, Sun H, Zhang S, Tu Y. Acupuncture improved neurological recovery after traumatic brain injury by activating BDNF/TrkB pathway. Evid Based Complement Alternat Med. 2017;2017:8460145. doi: 10.1155/2017/8460145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Yu JB, Shi J, Zhang Y, Gong LR, Dong SA, Cao XS, Wu LL, Wu LN. Electroacupuncture ameliorates acute renal injury in lipopolysaccharide-stimulated rabbits via induction of HO-1 through the PI3K/Akt/Nrf2 pathways. PLoS One. 2015;10:e0141622. doi: 10.1371/journal.pone.0141622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Xue X, You Y, Tao J, Ye X, Huang J, Yang S, Lin Z, Hong Z, Peng J, Chen L. Electro-acupuncture at points of Zusanli and Quchi exerts anti-apoptotic effect through the modulation of PI3K/Akt signaling pathway. Neurosci Lett. 2014;558:14–19. doi: 10.1016/j.neulet.2013.10.029. [DOI] [PubMed] [Google Scholar]
- 24.Bove J, Perier C. Neurotoxin-based models of Parkinson’s disease. Neuroscience. 2012;211:51–76. doi: 10.1016/j.neuroscience.2011.10.057. [DOI] [PubMed] [Google Scholar]
- 25.Tsai SJ, Kuo WW, Liu WH, Yin MC. Antioxidative and anti-inflammatory protection from carnosine in the striatum of MPTP-treated mice. J Agric Food Chem. 2010;58:11510–11516. doi: 10.1021/jf103258p. [DOI] [PubMed] [Google Scholar]
- 26.Xu XM, Moller SG. ROS removal by DJ-1: Arabidopsis as a new model to understand Parkinson’s disease. Plant Signal Behav. 2010;5:1034–1036. doi: 10.4161/psb.5.8.12298. [DOI] [PMC free article] [PubMed] [Google Scholar]