

Abstract
The milk-producing hormone prolactin (PRL) increases the transcellular intestinal calcium absorption by enhancing apical calcium uptake through voltage-dependent L-type calcium channel (Cav) 1.3. However, the redundancy of apical calcium channels raised the possibility that Cav1.3 may operate with other channels, especially transient receptor potential vanilloid family calcium channels (TRPV) 5 or 6, in an interdependent manner. Herein, TRPV5 knockdown (KD), TRPV5/TRPV6, TRPV5/Cav1.3, and TRPV6/Cav1.3 double KD, and TRPV5/TRPV6/Cav1.3 triple KD Caco-2 monolayers were generated by transfecting cells with small interfering RNAs (siRNA). siRNAs downregulated only the target mRNAs, and did not induce compensatory upregulation of the remaining channels. After exposure to 600 ng/mL PRL, the transcellular calcium transport was increased by ~2-fold in scrambled siRNA-treated, TRPV5 KD and TRPV5/TRPV6 KD monolayers, but not in TRPV5/Cav1.3, TRPV6/Cav1.3 and TRPV5/TRPV6/Cav1.3 KD monolayers. The results suggested that Cav1.3 was the sole apical channel responsible for the PRL-stimulated transcellular calcium transport in intestine-like Caco-2 monolayer.
Keywords: Calcium absorption, Small interfering RNA (siRNA), Transcellular transport, Triple knockdown, Voltage-dependent calcium channel (Cav)
Introduction
Prolactin (PRL) has been shown to be a calcium-regulating hormone since it could stimulate transcellular calcium absorption in intestinal epithelium of rats as well as in intestine-like Caco-2 monolayers [1–4]. Transcellular calcium transport is crucial during high calcium demand, such as pregnancy, lactation, or inadequate calcium intake [5]. It is a three-step process consisting of (1) apical calcium entry via transient receptor potential vanilloid Ca2+ channels (TRPV) 5, TRPV6 and voltage-dependent L-type calcium channel (Cav) 1.3, (2) cytoplasmic diffusion in a calbindin-D9k-bound form, and (3) basolateral extrusion via plasma membrane Ca2+-ATPase (PMCA) and Na+/Ca2+ exchanger (NCX) [6–9]. In general, Cav1.3 mediates intestinal calcium absorption under certain conditions, e.g., in the presence of depolarization state induced by glucose absorption [7, 8].
Recently, PRL was found to rapidly enhance transcellular calcium absorption in a non-genomic manner via Cav1.3, but not TRPV6 [9]. Pituitary-grafted rats with hyperprolactinemia also exhibited upregulation of L-type calcium channels [10]. However, the redundancy of apical calcium channels raised the possibility that the PRL-stimulated transcellular calcium transport may still require the presence of TRPV5 and/or TRPV6. Alternatively, one channel may be upregulated to compensate for the absence of the others. For example, TRPV5 knockout mice with hypercalciuria responded to negative calcium balance by upregulating TRPV6 expression in the small intestine to increase calcium absorption [11]. Indeed, it was not known whether calcium entry through TRPV5 contributed to the PRL-stimulated calcium flux in the intestine. Since TRPV5 and TRPV6, known to share 75% homology at the amino acid level and exhibit a similar ion permeation sequence for divalent cations (Ca2+ > Sr2+ ≈ Ba2+ > Mn2+), were co-expressed in the intestinal epithelial cells [12, 13], it was necessary to investigate the participation of TRPV5 in the PRL-stimulated calcium transport. Furthermore, whether Cav1.3 is the sole calcium channel required for PRL action was still not known.
Therefore, the objectives of the present study were (1) to examine the involvement of TRPV5 in the PRL-stimulated calcium absorption in intestinal epithelial monolayer, (2) to investigate whether an absence of one calcium channel led to upregulation of the others, and (3) to provide evidence that Cav1.3 was the sole calcium channel which mediated the PRL-stimulated calcium transport. Human colorectal adenocarcinoma cell line, Caco-2 cells, were used in the present study as they are easy to manipulate genetically. Despite being derived from the colon and having relatively low basal calcium flux compared to the small intestine [14], confluent Caco-2 monolayer has been widely used in the studies of drugs or hormones that enhance calcium absorption since it possesses characteristics of the small intestine, including the presence of the brush border, expression of sucrase-isomaltase enzymes, and expression of the transcellular calcium transporters [15–17]. Moreover, Caco-2 monolayer also expresses functional PRL receptors and responds well to PRL by increasing transcellular calcium flux by ~2-fold, similar to that observed in the rat duodenum [14].
Materials and methods
Cell culture
Caco-2 cells (ATCC No. HTB-37) were grown in Dulbecco’s modified Eagle medium (DMEM; Sigma, St. Louis, MO, USA) supplemented with 15% fetal bovine serum (FBS; Gibco, Grand Island, NY, USA), 1% l-glutamine (Gibco), 1% non-essential amino acid (Sigma), 100 U/mL penicillin/streptomycin (Sigma), and 0.25 μL/mL amphotericin B (Sigma). Cells were later propagated in 25-cm2 T-flask (Corning, NY, USA) under a humidified atmosphere containing 5% CO2 at 37°C, and subcultured as described in the ATCC protocol. Confluent Caco-2 monolayers were prepared by seeding cells (5.0 × 105 cells/cm2) on a polyester Snapwell with 12 mm diameter and 0.4 μm pore size (Corning). Culture medium was changed daily after 48 h of seeding.
Small interfering RNA transfection
Small interfering RNA oligonucleotides targeted for human TRPV5, TRPV6, and Cav1.3 as well as scrambled sequences were designed by siRNA Target Designer (version 1.51; Promega, Madison, WI, USA), as shown in Table 1. All oligonucleotides were synthesized by T7 RiboMax Express RNAi System (Promega) according to the manufacturer’s instruction. At day 12 after seeding of Caco-2 cells on a Snapwell, in vitro transfection was performed with 10 μg/mL polyethyleneimine (PEI) and 1 μmol/mL siRNA molecules to generate TRPV5 knockdown (KD), TRPV5/TRPV6, TRPV5/Cav1.3, TRPV6/Cav1.3 double KD, and TRPV5/TRPV6/Cav1.3 triple KD monolayers. At day 14 (i.e., 48 h after transfection), the Snapwell was mounted in an Ussing chamber with an exposed surface area of 1.13 cm2 to measure the electrical parameters and calcium fluxes, as previously described [18]. Efficiency of siRNA was evaluated by quantitative real-time PCR (qRT-PCR). Recent investigations showed that conventional siRNA KD successfully abolished the functions of TRPV5 and TRPV6 channels, as determined by patch-clamp technique [19, 20]. This KD protocol was approved by the Institutional Biosafety Committee of the Mahidol University.
Table 1.
siRNA oligonucleotides used in KD study
| Targets | siRNA oligonucleotides |
|---|---|
| TRPV5 siRNA |
5′-GGCACUUGAAUCUUGGACU-3′ 5′-AAAGUCCAAGAUUCAAGUGCC-3′ |
| TRPV6 siRNA |
5′-GGGAAACACAGUGUUACAC-3′ 5′-GTGUAACACUGUGUUUCCC-3′ |
| Cav1.3 siRNA |
5′-GAGCACCUUUGACAAUUUC-3′ 5′-AAGAAAUUGUCAAAGGTGCUC-3′ |
| Scramble siRNA |
5′-GGCGCAAUAAAGCAAGACC-3′ 5′-AAGGUCUUGCUUUAUUGCCC-3′ |
TRPV5/6 Transient receptor potential vanilloid family Ca2+ channels 5/6, Ca v 1.3 voltage-dependent L-type Ca2+ channel 1.3
Quantitative real-time PCR and sequencing
Expression levels of TRPV5, TRPV6 and Cav1.3 were quantified by a real-time PCR (model MiniOpticon; Bio-Rad) as described previously [9]. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH), a housekeeping gene, served as a control for normalization. Sense and antisense primers used for qRT-PCR are presented in Table 2. PCR reaction was performed with iQ SYBR Green SuperMix (Bio-Rad). Relative expression of TRPV5, TRPV6 or Cav1.3 over GAPDH was calculated from the threshold cycle (C t) values by using 2∆Ct method. Besides melting curve analysis, PCR products were also visualized on 1.5% agarose gel stained with 1.0 μg/mL ethidium bromide. After electrophoresis, PCR bands were extracted by HiYield Gel/PCR DNA Extraction kit (Real Biotech Corporation, Taipei, Taiwan), and were sequenced by ABI Prism 3100 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA). qRT-PCR experiments were performed in triplicate.
Table 2.
Homo sapiens oligonucleotide sequences used in the qRT-PCR experiments
| Name | Accession no. | Primer (forward/reverse) | Product length (bp) |
|---|---|---|---|
| TRPV5 | NM_019841 |
5′–CACTGTTATTGATGCACCTGC–3′ 5′–CCATCATGGCGATGAACA–3′ |
120 |
| TRPV6 | AF365928 |
5′–TCTGACTGCGTGTTCTCAC–3′ 5′–ACATTCCTTGGCGTTCAT–3′ |
144 |
| Cav1.3 | NM_000720 |
5′–TGATCCAAGTGGAGCAGTCA–3′ 5′–GTGTGAAAGTCCGGTAGGAGA–3′ |
113 |
| GAPDH | NM_002046 |
5′–CTGGTAAAGTGGATATTGTTG–3′ 5′–GAGGCTGTTGTCATACTTCTC–3′ |
359 |
TRPV5/6 Transient receptor potential vanilloid family Ca2+ channels 5/6, Ca v 1.3 voltage-dependent L-type Ca2+ channel 1.3, GAPDH glyceraldehyde-3-phosphate dehydrogenase
Bathing solution for Ussing chamber technique
The bathing solution for Ussing chamber experiments contained (in mmol/L) 118 NaCl, 4.7 KCl, 1.1 MgCl2, 1.25 CaCl2, 23 NaHCO3, 12 d-glucose and 2 mannitol (all purchased from Sigma). The solution was continuously gassed with humidified 5% CO2 in 95% O2, maintained at 37°C, pH 7.4, and had an osmolality of 290–293 mmol/kg water. Distilled water used in the present work had a resistance higher than 18.3 MΩ cm and a free ionized calcium concentration of <2.5 nmol/L.
Measurement of electrical parameters
Three electrical parameters, i.e., potential (voltage) difference (PD), short-circuit current (I sc), and transepithelial resistance (TER), were determined as described previously [14]. In brief, a pair of Ag/AgCl electrodes connected to agar bridges (3.0 mol/L KCl per 4% agar) was located near each surface of the mounted Snapwell for measurement of PD. The other ends of the PD-sensing electrodes were connected to a pre-amplifier (model EVC-4000; World Precision Instruments, Sarasota, FL, USA). Another pair of Ag/AgCl electrodes connected in series to the EVC-4000 current-generating unit was placed at the end of each hemichamber to supply I sc, which is external current to nullify PD. TER was calculated from Ohm’s equation. Fluid resistance was automatically subtracted by the EVC-4000 system.
Calcium flux measurement
Calcium fluxes were determined by the method of Thongon et al. [9]. In brief, after a 20-min incubation in the Ussing chamber, the bathing solution was changed with a fresh one. The solution on one side contained 45Ca (initial specific activity of 5 mCi/mL, final specific activity of ~450–500 mCi/mol; Amersham, Buckinghamshire, UK). Unidirectional flux of calcium (J H→C, nmol h−1 cm−2) from the hot side (H) to the cold side (C) was calculated with Eqs. 1 and 2.
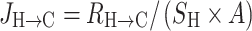 |
1 |
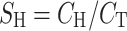 |
2 |
where R H→C is the rate of tracer appearance in the cold side (cpm/h), S H the specific activity in the hot side (cpm/nmol), A the surface area of Snapwell (cm2), C H is a radioactivity in the hot side (cpm), and C T is the total calcium in the hot side (nmol). 45Ca radioactivity in counts per minute (cpm) was analyzed by liquid scintillation spectrophotometer (model Tri-Carb 3100; Packard, Meriden, CT, USA). Total calcium concentration in the hot side was determined by atomic absorption spectrophotometer (model SpectrAA-300; Varian Techtron, Springvale, Australia). In the absence of transepithelial calcium gradient, i.e., bathing solution in both hemichambers contained equal calcium concentration of 1.25 mmol/L, the measured calcium fluxes represented the transcellular active calcium transport [9, 14]. In some experiments, Caco-2 monolayer was exposed on the basolateral side to 600 ng/mL recombinant human PRL (rhPRL; purity >97%; catalog no. 682-PL; R&D Systems, Minneapolis, MN, USA), which is the maximal effective concentration reported by Jantarajit et al. [14].
Statistic analysis
Unless otherwise specified, results are expressed as mean ± SE. Multiple sets of data were compared by one-way analysis of variance (ANOVA) with Dunnett’s multiple comparison test. The level of significance for all statistical tests was P < 0.05. Data were analyzed by GraphPad Prism (version 4.0 for Mac OS X; GraphPad Software, San Diego, CA, USA).
Results
Expression of TRPV5, TRPV6 and Cav1.3 in TRPV5 KD monolayer
Since it was not known whether TRPV5 mediated the PRL-stimulated transcellular calcium transport, we generated TRPV5 KD monolayers to demonstrate the significance of TRPV5 for such PRL action. After being treated with PEI, scrambled siRNA, or TRPV5 siRNA, Caco-2 cells exhibited a stable expression of GAPDH (Fig. 1a), indicating that GAPDH could be used as a control transcript. mRNA expression of TRPV5, but not TRPV6 or Cav1.3, markedly decreased in TRPV5 KD cells by ~17-fold (Fig. 1b–e). The results suggested that TRPV5 siRNA was effective in suppressing TRPV5 expression without causing compensatory upregulation of TRPV6 and Cav1.3.
Fig. 1.
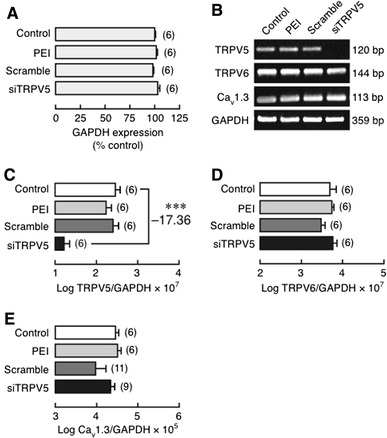
a Expression of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA in the control, PEI-treated, scrambled siRNA (Scramble)-transfected and TRPV5 siRNA (siTRPV5)-transfected Caco-2 monolayers. GAPDH expression of the control group was adjusted to 100%, while those of other groups were relative to the control group. b Representative electrophoretic bands of TRPV5, TRPV6 and Cav1.3 expression in the control, PEI, Scramble and siTRPV5 groups. c–e Expression of TRPV5, TRPV6 and Cav1.3 in TRPV5 KD Caco-2 monolayer as demonstrated by qRT-PCR. Expression levels were normalized by GAPDH expression. Values are presented as log mean ± SE. ***P < 0.001 compared with the control group. Value of fold difference was also presented along with the statistical symbol. Numbers in parentheses represent the number of independent experiments. Each experiment was performed in triplicate
PRL-enhanced calcium transport in TRPV5 KD monolayer
After being exposed to 600 ng/mL rhPRL, both control and TRPV5 KD monolayers manifested an approximately twofold increase in the transcellular calcium transport (Fig. 2). The basal calcium transport was not affected by transfecting agent PEI, scrambled siRNA, or TRPV5 siRNA. The results, therefore, suggested that TRPV5 may not be required for the PRL-enhanced transcellular calcium transport.
Fig. 2.
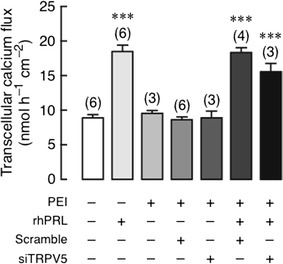
Transcellular calcium transport in Caco-2 monolayer transfected with TRPV5 siRNA (siTRPV5) with or without 600 ng/mL rhPRL exposure. Scrambled siRNA was used as a negative control. Monolayer was bathed on both sides with 1.25 mmol/L calcium-containing solution. ***P < 0.001 compared with the control group. Numbers in parentheses represent the number of independent experiments
Expression of TRPV5, TRPV6 and Cav1.3 in Caco-2 monolayers after combinatorial KD
To exclude the possibility that, in response to PRL, the remaining two channels were upregulated to compensate for the absence of the other channels after single KD of TRPV5, TRPV6, or Cav1.3, we generated Caco-2 monolayers with double KD of TRPV5/TRPV6, TRPV5/Cav1.3, or TRPV6/Cav1.3, or triple KD of TRPV5/TRPV6/Cav1.3. We first demonstrated that, in a control monolayer, exposure to 600 ng/mL rhPRL for 60 min did not alter TRPV5, TRPV6, and Cav1.3 expression (Fig. 3a). Moreover, GAPDH expression was stable in both double and triple KD monolayers (Fig. 3b). TRPV5 mRNA expressions in TRPV5/TRPV6, TRPV5/Cav1.3, and triple KD monolayers were decreased by ~24-, ~16-, and ~18-fold, respectively, whereas no significant change was found in TRPV6/Cav1.3 KD monolayer (Fig. 3c, d). Similarly, TRPV6 and Cav1.3 mRNA expressions were downregulated only in the presence of their target siRNA molecules (Fig. 3c, e, f). These findings indicated that there was no compensatory expression of the remaining channel when mRNAs of the two channels were disrupted. In other words, the lack of expression of one channel could not be rescued by expression of the others.
Fig. 3.
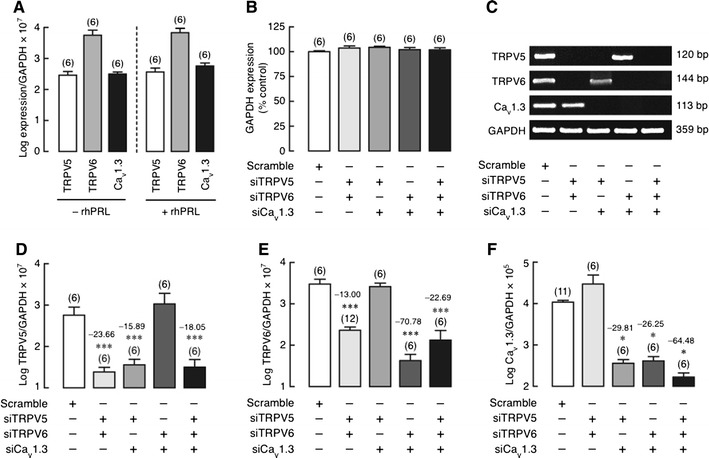
a Expression of TRPV5, TRPV6, and Cav1.3 mRNA in Caco-2 monolayer directly exposed for 60 min to 600 ng/mL rhPRL. b GAPDH expression in TRPV5/TRPV6, TRPV5/Cav1.3, TRPV6/Cav1.3 double KD, and TRPV5/TRPV6/Cav1.3 triple KD monolayers. GAPDH expression in scrambled siRNA-treated group was adjusted to 100%. c Representative electrophoretic bands of TRPV5, TRPV6, and Cav1.3 mRNA expression in double and triple KD monolayers. d–f Expression of TRPV5, TRPV6, and Cav1.3 mRNA in double and triple KD Caco-2 monolayers as demonstrated by qRT-PCR. Expression levels were normalized by GAPDH expression. Data are presented as log mean ± SE. *P < 0.05, ***P < 0.001 compared with the scrambled siRNA-treated group. Values of fold differences were also presented along with the statistical symbols. Numbers in parentheses represent the number of independent experiments. Each experiment was performed in triplicate. siTRPV5, siTRPV6, and siCav1.3 mean siRNAs against TRPV5, TRPV6, and Cav1.3 mRNA, respectively
PRL-enhanced calcium transport in Caco-2 monolayers after combinatorial KD of TRPV5, TRPV6 and Cav1.3
In TRPV5/TRPV6 double KD monolayer, 600 ng/mL rhPRL was able to stimulate the transcellular calcium transport, similar to that observed in the control and PEI-treated monolayers (Fig. 4). However, in TRPV5/Cav1.3, TRPV6/Cav1.3 double KD, and triple KD monolayers, the PRL-stimulated transcellular calcium transport was completely abolished (Fig. 4). Such combinatorial KD had no effect on the basal calcium transport, PD, I sc, or the PRL-induced decrease in TER (Figs. 4, 5), the latter of which indirectly represented an increase in the paracellular permeability by PRL [9, 18, 21]. The results corroborated that Cav1.3 was the sole calcium channels required for the PRL-stimulated transcellular calcium transport across the intestinal epithelium.
Fig. 4.
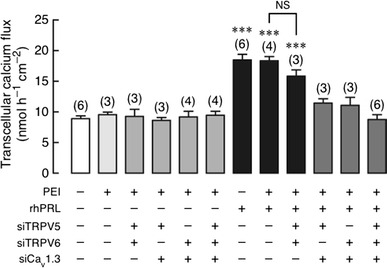
Transcellular calcium transport in TRPV5/TRPV6, TRPV5/Cav1.3, TRPV6/Cav1.3 double KD, and TRPV5/TRPV6/Cav1.3 triple KD monolayers directly exposed to 600 ng/mL rhPRL. PEI was a transfecting agent. Monolayer was bathed on both sides with 1.25 mmol/L calcium-containing solution. ***P < 0.001 compared with the scrambled siRNA-treated group (negative control). Numbers in parentheses represent the number of independent Snapwells. siTRPV5, siTRPV6, and siCav1.3 mean siRNAs against TRPV5, TRPV6, and Cav1.3 mRNA, respectively. NS Not significant
Fig. 5.
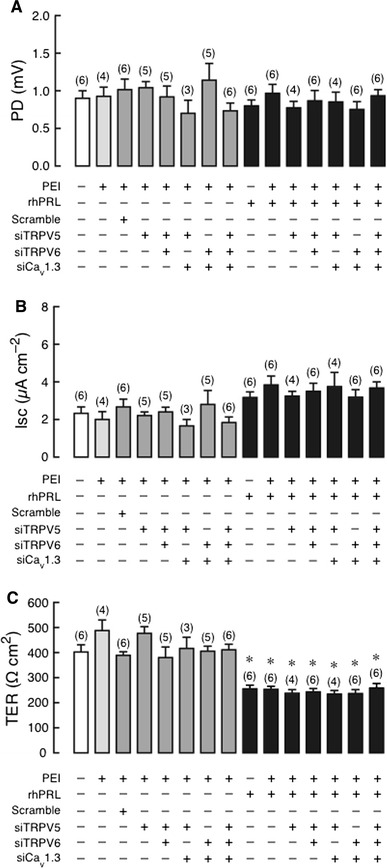
a Transepithelial potential difference (PD), b short-circuit current (I sc), and c transepithelial resistance (TER) in TRPV5 KD, TRPV5/TRPV6, TRPV5/Cav1.3, TRPV6/Cav1.3 double KD, and TRPV5/TRPV6/Cav1.3 triple KD monolayers directly exposed to 600 ng/mL rhPRL. PEI was a transfecting agent. The apical side had negative voltage with respect to the basolateral side. *P < 0.05 compared with the scrambled siRNA-treated group. Numbers in parentheses represent the number of independent Snapwells. siTRPV5, siTRPV6, and siCav1.3 mean siRNAs against TRPV5, TRPV6, and Cav1.3 mRNA, respectively. The absence of changes in PRL actions in KD monolayers indicated that these calcium channels were not required for the PRL-induced increase in paracellular permeability
Discussion
A significant amount of maternal calcium (~200–400 mg/day) is lost during pregnancy and lactation for fetal growth and milk production, thus resulting in severe negative calcium balance and osteopenia [22]. In lactation, PRL is not only an important hormone for lactogenesis but also for the regulation of overall calcium metabolism in pregnancy and lactation, in part, by stimulating renal calcium reabsorption and transcellular calcium absorption in the small intestine, thereby ameliorating stress on calcium metabolism [2, 5]. Previous investigations in duodenal epithelial cells showed that PRL enhanced the transcellular calcium transport by increasing the apical (brush-border) calcium uptake [23]. The present study further demonstrated that Cav1.3 was the sole apical calcium channel for the PRL-stimulated transcellular calcium transport in Caco-2 monolayer, which is an accepted model for calcium absorption study [9, 15].
Three calcium channels, namely TRPV5, TRPV6, and Cav1.3, were co-expressed on the apical membrane of duodenal, jejunal, and colonic epithelial cells [24, 25]. For the past decade, TRPV6 has been postulated to be the principal calcium channel for apical calcium entry in the small intestine, whereas TRPV5, with its expression being much lower than that of TRPV6 in both duodenum [26] and Caco-2 cells (Fig. 3a), was thought to play a minor role. However, physiological significance of TRPV6 became doubtful when TRPV6 null mice were found to be normocalcemic and responded to 1,25(OH)2D3 by increasing transcellular calcium transport, similar to that observed in the wild-type mice [27, 28]. Moreover, Morgan and co-workers [8] suggested that TRPV5 and TRPV6 could not have been involved in the glucose-stimulated calcium absorption since both channels normally operated at negative membrane potentials below −90 mV (hyperpolarization), whereas postprandial glucose-rich luminal condition usually led to cell depolarization. Therefore, in the presence of luminal glucose, Cav1.3 must have been responsible for the intestinal calcium absorption [7, 8].
Nevertheless, intestinal TRPV5 and TRPV6 could be important for the enhancement of transcellular calcium transport in certain stimulating conditions, such as chronic metabolic acidosis, and long-term exposure to estrogen or 1,25(OH)2D3 [26, 29–31]. Indeed, TRPV6 expression was markedly upregulated after 1,25(OH)2D3 administration to vitamin D-deficient mice [32]. A number of hormones or local factors, such as parathyroid hormone, arginine vasopressin, prostaglandin E2, and calcitonin, were also found to activate TRPV5 and/or TRPV6 by raising intracellular cAMP or cGMP levels [33]. It is possible that TRPV5, TRPV6, and Cav1.3 may differentially respond to various stimuli or extracellular environments. In other words, these channels have their own unique properties and can function independently of each other. Such a hypothesis was confirmed by the present findings that neither TRPV5 nor Cav1.3 was upregulated when TRPV6 expression was suppressed by siRNA. Nevertheless, other alternative mechanisms for apical calcium entry might exist, since TRPV5/TRPV6/Cav1.3 triple KD Caco-2 monolayer still exhibited transcellular calcium flux with a magnitude comparable to that of normal monolayer. Possible alternative pathways included the vesicular transport and voltage-dependent T-type calcium channels, the latter of which was reported to mediate the testosterone-stimulated calcium transport in renal tubular epithelium [34]. Alternatively, since siRNA-induced calcium channel KD did not totally suppress mRNA expression of target calcium channels (Fig. 3d–f), it was possible that a small number of calcium channels were sufficient to allow calcium entry under a non-stimulated condition. Feedback regulation of calcium influx could also explain the unaltered basal calcium transport after triple KD, which should lead to a decrease in the intracellular calcium concentration (Cai). Low Cai has been postulated to promote activities of TRPV5 and TRPV6 channels [35, 36], thereby leading to an increase in calcium influx.
Our previous investigation provided evidence that stimulatory effect of PRL on transepithelial calcium transport was abolished by Cav1.3 KD or L-type calcium channel blockers, nifedipine and verapamil, in the apical solution, but not in the basolateral solution [9]. Cav1.3 thus appeared to be responsible for apical calcium entry in PRL-exposed intestinal epithelium [9], but participation of TRPV5 and compensatory TRPV6 upregulation after Cav1.3 KD were still possible and were not excluded experimentally. Herein, we further elucidated that only Cav1.3 was involved in the PRL-stimulated transcellular calcium transport across the intestinal epithelium. Interdependent or interactive functions of TRPV5, TRPV6, and Cav1.3 in the presence of PRL could also be ruled out because enhanced transcellular calcium transport was still observed in TRPV5/TRPV6 double KD Caco-2 monolayer.
Molecular mechanism by which PRL stimulates intestinal calcium absorption is poorly understood. Recent studies in duodenal epithelium and Caco-2 monolayer substantiated pivotal roles of phosphoinositide 3-kinase (PI3K) and protein kinase C (PKC), especially PKCζ isozyme, in the intracellular PRL signaling [14]. Panspecific PKC inhibitor (GF109203X) was shown to diminish the PRL-accelerated apical calcium uptake [9]. Although Cav1.3 could be phosphorylated by several PKC isozymes, e.g., PKCβII [7], it is not known whether PRL-activated PKC directly phosphorylated Cav1.3. Since Na+ and glucose uptake through sodium-dependent glucose transporter 1 (SGLT1) at the apical membrane is coupled with Cav1.3 opening, which requires cell depolarization induced by SGLT1-mediated Na+ entry [8, 37], it is also possible that PRL indirectly activates Cav1.3 by acting through SGLT1, known to be expressed in Caco-2 cells [38].
Besides the transcellular calcium transport, PRL could also stimulate calcium transport across the paracellular route [5]. The observed decrease in TER by PRL in the present study confirmed an increase in paracellular permeability, which could facilitate the paracellular calcium transport [9, 18, 21]. Due to its non-saturable passive nature, paracellular calcium transport could become substantial after ingestion of calcium-rich meals, or in the presence of a transepithelial calcium gradient [6]. Moreover, the presence of PRL-induced decrease in TER in TRPV5/TRPV6/Cav1.3 KD monolayer suggested that the PRL-enhanced paracellular permeability did not involve the functions of these three calcium channels, although perijunctional cytoskeletal rearrangement, an initial step to increase paracellular permeability, may be controlled, in part, by apical calcium entry [37, 39].
In conclusion, we used siRNA-based combinatorial KD technique to elucidate that the apical calcium entry step in the PRL-stimulated transcellular calcium transport in the intestine-like Caco-2 monolayer occurred solely via Cav1.3. Although intestinal epithelial cells expressed several calcium channels, i.e., TRPV5, TRPV6, and Cav1.3, there was no redundancy in their functions, at least during PRL exposure, since the PRL-stimulated calcium transport was totally diminished after Cav1.3 siRNA transfection. The present results, therefore, provided corroborative evidence for better understanding of the cellular mechanism of PRL in intestinal epithelial cells, and could explain the mechanism by which PRL stimulated intestinal calcium absorption in pregnant and lactating animals.
Acknowledgments
This research was supported by grants from the Mahidol University Postdoctoral Fellowship Program (to L. Nakkrasae), the Faculty of Science, Mahidol University (SCY52-02 and SCR52-01 to N. Charoenphandhu), the Commission on Higher Education, and the Thailand Research Fund (RSA5180001 to N. Charoenphandhu, and RTA5080008 to N. Krishnamra).
Conflict of interest statement
The authors declare no conflict of interest.
References
- 1.Boass A, Lovdal JA, Toverud SU. Pregnancy- and lactation-induced changes in active intestinal calcium transport in rats. Am J Physiol Gastrointest Liver Physiol. 1992;263:G127–G134. doi: 10.1152/ajpgi.1992.263.1.G127. [DOI] [PubMed] [Google Scholar]
- 2.Charoenphandhu N, Nakkrasae LI, Kraidith K, Teerapornpuntakit J, Thongchote K, Thongon N, Krishnamra N. Two-step stimulation of intestinal Ca2+ absorption during lactation by long-term prolactin exposure and suckling-induced prolactin surge. Am J Physiol Endocrinol Metab. 2009;297:E609–E619. doi: 10.1152/ajpendo.00347.2009. [DOI] [PubMed] [Google Scholar]
- 3.Krishnamra N, Taweerathitam P. Acute effect of prolactin on active calcium absorption in rats. Can J Physiol Pharmacol. 1995;73:1185–1189. doi: 10.1139/y95-168. [DOI] [PubMed] [Google Scholar]
- 4.Krishnamra N, Wirunrattanakij Y, Limlomwongse L. Acute effects of prolactin on passive calcium absorption in the small intestine by in vivo perfusion technique. Can J Physiol Pharmacol. 1998;76:161–168. doi: 10.1139/cjpp-76-2-161. [DOI] [PubMed] [Google Scholar]
- 5.Charoenphandhu N, Krishnamra N. Prolactin is an important regulator of intestinal calcium transport. Can J Physiol Pharmacol. 2007;85:569–581. doi: 10.1139/Y07-041. [DOI] [PubMed] [Google Scholar]
- 6.Hoenderop JG, Nilius B, Bindels RJ. Calcium absorption across epithelia. Physiol Rev. 2005;85:373–422. doi: 10.1152/physrev.00003.2004. [DOI] [PubMed] [Google Scholar]
- 7.Morgan EL, Mace OJ, Affleck J, Kellett GL. Apical GLUT2 and Cav1.3: regulation of rat intestinal glucose and calcium absorption. J Physiol. 2007;580:593–604. doi: 10.1113/jphysiol.2006.124768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Morgan EL, Mace OJ, Helliwell PA, Affleck J, Kellett GL. A role for Cav1.3 in rat intestinal calcium absorption. Biochem Biophys Res Commun. 2003;312:487–493. doi: 10.1016/j.bbrc.2003.10.138. [DOI] [PubMed] [Google Scholar]
- 9.Thongon N, Nakkrasae LI, Thongbunchoo J, Krishnamra N, Charoenphandhu N. Enhancement of calcium transport in Caco-2 monolayer through PKCζ-dependent Cav1.3-mediated transcellular and rectifying paracellular pathways by prolactin. Am J Physiol Cell Physiol. 2009;296:C1373–C1382. doi: 10.1152/ajpcell.00053.2009. [DOI] [PubMed] [Google Scholar]
- 10.Charoenphandhu N, Wongdee K, Teerapornpuntakit J, Thongchote K, Krishnamra N. Transcriptome responses of duodenal epithelial cells to prolactin in pituitary-grafted rats. Mol Cell Endocrinol. 2008;296:41–52. doi: 10.1016/j.mce.2008.09.025. [DOI] [PubMed] [Google Scholar]
- 11.Hoenderop JG, van Leeuwen JP, van der Eerden BC, Kersten FF, van der Kemp AW, Merillat AM, Waarsing JH, Rossier BC, Vallon V, Hummler E, Bindels RJ. Renal Ca2+ wasting, hyperabsorption, and reduced bone thickness in mice lacking TRPV5. J Clin Invest. 2003;112:1906–1914. doi: 10.1172/JCI19826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hoenderop JG, Hartog A, Stuiver M, Doucet A, Willems PH, Bindels RJ. Localization of the epithelial Ca2+ channel in rabbit kidney and intestine. J Am Soc Nephrol. 2000;11:1171–1178. doi: 10.1681/ASN.V1171171. [DOI] [PubMed] [Google Scholar]
- 13.Zhuang L, Peng JB, Tou L, Takanaga H, Adam RM, Hediger MA, Freeman MR. Calcium-selective ion channel, CaT1, is apically localized in gastrointestinal tract epithelia and is aberrantly expressed in human malignancies. Lab Invest. 2002;82:1755–1764. doi: 10.1097/01.lab.0000043910.41414.e7. [DOI] [PubMed] [Google Scholar]
- 14.Jantarajit W, Thongon N, Pandaranandaka J, Teerapornpuntakit J, Krishnamra N, Charoenphandhu N. Prolactin-stimulated transepithelial calcium transport in duodenum and Caco-2 monolayer are mediated by the phosphoinositide 3-kinase pathway. Am J Physiol Endocrinol Metab. 2007;293:E372–E384. doi: 10.1152/ajpendo.00142.2007. [DOI] [PubMed] [Google Scholar]
- 15.Nakane M, Ma J, Rose AE, Osinski MA, Wu-Wong JR. Differential effects of vitamin D analogs on calcium transport. J Steroid Biochem Mol Biol. 2007;103:84–89. doi: 10.1016/j.jsbmb.2006.07.004. [DOI] [PubMed] [Google Scholar]
- 16.Yee S. In vitro permeability across Caco-2 cells (colonic) can predict in vivo (small intestinal) absorption in man—fact or myth. Pharm Res. 1997;14:763–766. doi: 10.1023/A:1012102522787. [DOI] [PubMed] [Google Scholar]
- 17.Zweibaum A, Triadou N, Kedinger M, Augeron C, Robine-Leon S, Pinto M, Rousset M, Haffen K. Sucrase-isomaltase: a marker of foetal and malignant epithelial cells of the human colon. Int J Cancer. 1983;32:407–412. doi: 10.1002/ijc.2910320403. [DOI] [PubMed] [Google Scholar]
- 18.Thongon N, Nakkrasae LI, Thongbunchoo J, Krishnamra N, Charoenphandhu N. Prolactin stimulates transepithelial calcium transport and modulates paracellular permselectivity in Caco-2 monolayer: mediation by PKC and ROCK pathways. Am J Physiol Cell Physiol. 2008;294:C1158–C1168. doi: 10.1152/ajpcell.00020.2008. [DOI] [PubMed] [Google Scholar]
- 19.Irnaten M, Blanchard-Gutton N, Harvey BJ. Rapid effects of 17β-estradiol on epithelial TRPV6 Ca2+ channel in human T84 colonic cells. Cell Calcium. 2008;44:441–452. doi: 10.1016/j.ceca.2008.02.007. [DOI] [PubMed] [Google Scholar]
- 20.Irnaten M, Blanchard-Gutton N, Praetorius J, Harvey BJ. Rapid effects of 17β-estradiol on TRPV5 epithelial Ca2+ channels in rat renal cells. Steroids. 2009;74:642–649. doi: 10.1016/j.steroids.2009.02.002. [DOI] [PubMed] [Google Scholar]
- 21.Greger R. Epithelial transport. In: Greger R, Windhorst U, editors. Comprehensive human physiology: from cellular mechanisms to integration. 1. Berlin: Springer; 1996. pp. 1217–1232. [Google Scholar]
- 22.Prentice A. Calcium in pregnancy and lactation. Annu Rev Nutr. 2000;20:249–272. doi: 10.1146/annurev.nutr.20.1.249. [DOI] [PubMed] [Google Scholar]
- 23.Charoenphandhu N, Limlomwongse L, Krishnamra N. Prolactin directly enhanced Na+/K+- and Ca2+-ATPase activities in the duodenum of female rats. Can J Physiol Pharmacol. 2006;84:555–563. doi: 10.1139/y05-161. [DOI] [PubMed] [Google Scholar]
- 24.Hoenderop JG, Vennekens R, Muller D, Prenen J, Droogmans G, Bindels RJ, Nilius B. Function and expression of the epithelial Ca2+ channel family: comparison of mammalian ECaC1 and 2. J Physiol. 2001;537:747–761. doi: 10.1113/jphysiol.2001.012917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Peng JB, Chen XZ, Berger UV, Weremowicz S, Morton CC, Vassilev PM, Brown EM, Hediger MA. Human calcium transport protein CaT1. Biochem Biophys Res Commun. 2000;278:326–332. doi: 10.1006/bbrc.2000.3716. [DOI] [PubMed] [Google Scholar]
- 26.Van Cromphaut SJ, Dewerchin M, Hoenderop JG, Stockmans I, Van Herck E, Kato S, Bindels RJ, Collen D, Carmeliet P, Bouillon R, Carmeliet G. Duodenal calcium absorption in vitamin D receptor-knockout mice: functional and molecular aspects. Proc Natl Acad Sci USA. 2001;98:13324–13329. doi: 10.1073/pnas.231474698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Benn BS, Ajibade D, Porta A, Dhawan P, Hediger M, Peng JB, Jiang Y, Oh GT, Jeung EB, Lieben L, Bouillon R, Carmeliet G, Christakos S. Active intestinal calcium transport in the absence of transient receptor potential vanilloid type 6 and calbindin-D9k . Endocrinology. 2008;149:3196–3205. doi: 10.1210/en.2007-1655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kutuzova GD, Sundersingh F, Vaughan J, Tadi BP, Ansay SE, Christakos S, DeLuca HF. TRPV6 is not required for 1α, 25-dihydroxyvitamin D3-induced intestinal calcium absorption in vivo. Proc Natl Acad Sci USA. 2008;105:19655–19659. doi: 10.1073/pnas.0810761105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Charoenphandhu N, Tudpor K, Pulsook N, Krishnamra N. Chronic metabolic acidosis stimulated transcellular and solvent drag-induced calcium transport in the duodenum of female rats. Am J Physiol Gastrointest Liver Physiol. 2006;291:G446–G455. doi: 10.1152/ajpgi.00108.2006. [DOI] [PubMed] [Google Scholar]
- 30.Van Abel M, Hoenderop JG, Dardenne O, St Arnaud R, Van Os CH, Van Leeuwen HJ, Bindels RJ. 1,25-dihydroxyvitamin D3-independent stimulatory effect of estrogen on the expression of ECaC1 in the kidney. J Am Soc Nephrol. 2002;13:2102–2109. doi: 10.1097/01.ASN.0000022423.34922.2A. [DOI] [PubMed] [Google Scholar]
- 31.Weber K, Erben RG, Rump A, Adamski J. Gene structure and regulation of the murine epithelial calcium channels ECaC1 and 2. Biochem Biophys Res Commun. 2001;289:1287–1294. doi: 10.1006/bbrc.2001.6121. [DOI] [PubMed] [Google Scholar]
- 32.Song Y, Peng X, Porta A, Takanaga H, Peng JB, Hediger MA, Fleet JC, Christakos S. Calcium transporter 1 and epithelial calcium channel messenger ribonucleic acid are differentially regulated by 1,25 dihydroxyvitamin D3 in the intestine and kidney of mice. Endocrinology. 2003;144:3885–3894. doi: 10.1210/en.2003-0314. [DOI] [PubMed] [Google Scholar]
- 33.den Dekker E, Hoenderop JG, Nilius B, Bindels RJ. The epithelial calcium channels, TRPV5 and TRPV6: from identification towards regulation. Cell Calcium. 2003;33:497–507. doi: 10.1016/S0143-4160(03)00065-4. [DOI] [PubMed] [Google Scholar]
- 34.Couchourel D, Leclerc M, Filep J, Brunette MG. Testosterone enhances calcium reabsorption by the kidney. Mol Cell Endocrinol. 2004;222:71–81. doi: 10.1016/j.mce.2004.05.001. [DOI] [PubMed] [Google Scholar]
- 35.Lambers TT, Mahieu F, Oancea E, Hoofd L, de Lange F, Mensenkamp AR, Voets T, Nilius B, Clapham DE, Hoenderop JG, Bindels RJ. Calbindin-D28K dynamically controls TRPV5-mediated Ca2+ transport. EMBO J. 2006;25:2978–2988. doi: 10.1038/sj.emboj.7601186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Niemeyer BA, Bergs C, Wissenbach U, Flockerzi V, Trost C. Competitive regulation of CaT-like-mediated Ca2+ entry by protein kinase C and calmodulin. Proc Natl Acad Sci USA. 2001;98:3600–3605. doi: 10.1073/pnas.051511398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Mace OJ, Morgan EL, Affleck JA, Lister N, Kellett GL. Calcium absorption by Cav1.3 induces terminal web myosin II phosphorylation and apical GLUT2 insertion in rat intestine. J Physiol. 2007;580:605–616. doi: 10.1113/jphysiol.2006.124784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Mahraoui L, Rodolosse A, Barbat A, Dussaulx E, Zweibaum A, Rousset M, Brot-Laroche E. Presence and differential expression of SGLT1, GLUT1, GLUT2, GLUT3 and GLUT5 hexose-transporter mRNAs in Caco-2 cell clones in relation to cell growth and glucose consumption. Biochem J. 1994;298(Pt 3):629–633. doi: 10.1042/bj2980629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Pérez M, Barber A, Ponz F. Modulation of intestinal paracellular permeability by intracellular mediators and cytoskeleton. Can J Physiol Pharmacol. 1997;75:287–292. doi: 10.1139/cjpp-75-4-287. [DOI] [PubMed] [Google Scholar]