

Abstract
This review provides a summary of the physiological significance of the TRPV2 ion channel. While TRPV2 was initially characterized as a noxious heat sensor, we found that TRPV2 can also act as a mechanosensor in embryonic neurons or adult myenteric neurons. Here, we summarize the newly characterized functions of TRPV2, including the research progress that has been made toward our understanding of TRPV2 physiology, and discuss other recent data pertaining to TRPV2. It is thought that TRPV2 may be an important drug target based on its broad expression patterns and important physiological roles. The possible associations between diseases and TRPV2 are also discussed.
Keywords: TRPV2, Mechanical stimulus, Axon outgrowth, Mechanosensor, Thermosensor
Introduction
Members of the transient receptor potential (TRP) superfamily are widely expressed throughout numerous tissues. The channels are activated by various stimuli such as temperature, light, chemical and physical stimuli. TRP channels were originally identified in Drosophila photoreceptors [1], from which the first channel was cloned [2]. The TRP channels are expressed in a range of species from yeast to humans. Most TRP channels act as polymodal receptors. For example, some TRP channels expressed in the nociceptive sensory neurons of the dorsal root ganglion (DRG) generate pain [3]. Such TRP channels represent leading targets for the cure of acute and chronic pain [3–5]. Given this importance, the recent advances in our understanding of TRPV1 and TRPA1 have been impressive; these include the characterization of channel structure [6, 7], identification of ligands, and modulation of channels and intracellular signaling related to pain [3]. Beyond pain sensation, TRP channels have other important functions, or are involved in pathological processes such as diabetes, cancer and other genetic disorders [8]. TRP channels have a common membrane structure with six transmembrane segments (S1–S6) containing a TRP domain and a loop between S5 and S6, which defines the pore and selectivity filter (Fig. 1a) [9]. Similar to other voltage-gated potassium channels, TRP tetramers form the functional non-selective cation channels (Fig. 1b). The N- and C-terminal cytoplasmic domains are important for channel gating [9]. TRP channels are classified by their amino acid sequence identity and similarity, but not by their functional roles. The mammalian TRP channel superfamily is classified into six subfamilies with high sequence similarity (Fig. 1c). The six subfamilies [10] are TRPV (vanilloid), TRPA (ANKTM1), TRPC (canonical), TRPM (melastatin), TRPML (mucolipin), and TRPP (polycystin). A seventh family, TRPN (NompC) is present in invertebrates and some vertebrates, although this subfamily is absent in mammals [9]. In this review, we focus on the unique characteristics of TRPV2 as a mechanosensor, thermosensor and lipid sensor.
Fig. 1.
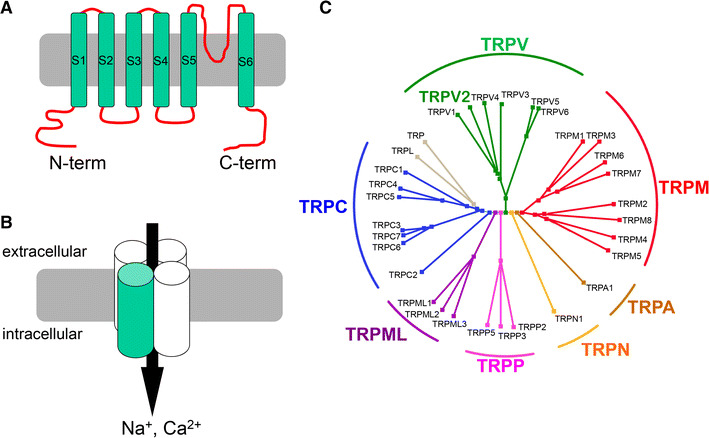
The overview of TRP channels. a The TRP channels have six transmembrane topology in their monomer molecules. The pore region is located between S5 and S6. b Tetrameric channels form the functional TRP channels. c The classification of TRP channels modified from another review [10]
TRPV2 ion channel
The first identified member of the TRPV family was osmotic avoidance abnormal family member 9 (OSM-9) from Caenorhabditis elegans (C. elegans) [9, 11]. This protein was shown to have specific functions for mechanosensation, olfaction and olfactory adaptation. C. elegans has five TRPV channels including OSM-9. In Drosophila, the TRPV channels Nanchung and Inactive are involved in sensory perception [12]. In 1997, TRPV1 was the first TRPV family member identified in mammals using a Ca2+-imaging-based expression cloning method [13]. TRPV1 was shown to be highly expressed in sensory neurons, and pungent compounds such as capsaicin could evoke TRPV1 activation [13, 14]. Based on these studies, it was hypothesized that TRPV1 was related to pain sensation and, indeed, TRPV1KO mice showed impaired pain sensation [15]. TRPV1 is highly expressed in sensory neurons with small diameters which give rise to unmyelinated C-fibers. Given the burning qualities of capsaicin-induced pain, it suggested that capsaicin and heat may evoke painful responses through a common molecular mechanism. As hypothesized, TRPV1 can be activated by heat at >43 °C, a temperature threshold that is consistent with that of heat-evoked pain [13].
Another molecule with 49 % shared identity to TRPV1 was also isolated and later named TRPV2 [3, 16, 17]. In contrast to TRPV1, TRPV2 is not activated by vanilloids such as capsaicin, protons or thermal stimuli near 43 °C; this is likely attributed to the structure of the ankyrin repeat domains of TRPV2 which are clearly different from those of TRPV1 [18]. However, TRPV2 can be activated by higher temperatures with a threshold of >52 °C. TRPV2 currents showed similar properties to TRPV1 such as an outwardly rectifying I–V relationship and relatively high Ca2+ permeability (P Ca/P Na = 2.9). While translocation of TRPV2 from the cytosol to the plasma membrane by IGF-1 stimulation has been reported [17], this phenomenon is under debate [19]. In contrast to the expression of TRPV1, TRPV2 is expressed in medium- to large-diameter neurons which give rise to myelinated Aδ- or Aβ-fibers in the DRG [16]. The temperatures at which TRPV2 is activated are more noxious than those that activate TRPV1. This may explain the expression of TRPV2 in myelinated fibers. However, TRPV2KO mice did not display phenotypes which would suggest altered heat sensation [20]. Therefore, further research is required to determine the importance of TRPV2 in the detection of noxious heat stimuli using TRPV2KO mice. TRPV2 expression was not only found in sensory neurons but also in motor neurons and in many non-neuronal tissues [9, 21–23]. These observations indicate that TRPV2 has numerous physiological roles in addition to nociception. For example, we demonstrated that TRPV2 activation regulates the formation of neural circuits in a membrane stretch-dependent manner [21].
Regulation of axon outgrowth by TRPV2 mechanosensitivity
We have previously demonstrated that TRPV2 expression begins at embryonic day 10 (E10) in mouse DRG sensory and motor neurons [21]. Given that mouse embryos are maintained in a constant temperature environment, we hypothesized that TRPV2 has other functions during the developmental process. Therefore, we examined embryonic stage-specific events which might involve TRPV2, such as cell proliferation, apoptosis, cell migration, and axon outgrowth. While TRPV2 was not found to be related to cell proliferation, apoptosis or cell migration, we did identify a role for TRPV2 in the enhancement of axon outgrowth in developing neurons [21]. We further sought to identify the activators of TRPV2 within developing neurons. Since it was suggested that TRPV2 can be activated by membrane stretch in cardiomyocytes [24], we hypothesized that membrane stretch resulting from axon outgrowth might endogenously activate TRPV2. To examine this hypothesis, we prepared HEK293T cells expressing mouse TRPV2 and applied artificial membrane stretch (+3 %) to the cells using a pulse-motor-driven stretch machine called STREX [25] during Ca2+-imaging experiments. The membrane stretch-evoked increase in intracellular Ca2+ ([Ca2+]i) was observed only in transfected (TRPV2-expressing) cells, indicating that TRPV2 can be activated by weak membrane stretch. Consistent with the HEK293T results, most embryonic DRG and motor neurons showed a stretch-evoked [Ca2+]i increase, whereas dominant-negative TRPV2 significantly inhibited this increase [21]. Thus, we demonstrated that developing neurons can sense membrane stretch via TRPV2, and the subsequent activation enhances axon outgrowth (Fig. 2). Moreover, we showed that Ca2+ influx through TRPV2 is necessary for enhancing axon outgrowth. We also demonstrated regulation of axon outgrowth by TRPV2 in vivo using chick embryos [21]. More recently, it has been reported that phosphorylation of TRPV2 by NGF-ERK signaling resulted in the activation of intracellular, but not membranous, TRPV2 as well as enhanced axon outgrowth [26].
Fig. 2.
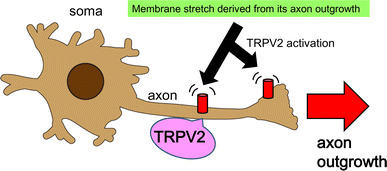
TRPV2 activation by membrane stretch promotes axon outgrowth in developing sensory and motor neurons. TRPV2 is expressed in both axon shafts and growth cones as shown by red images. The TRPV2 is activated by membrane stretch derived from endogenous axon outgrowth shown by black arrows. The activation of TRPV2 caused high Ca2+ influx and further axon outgrowth
Mechanosensitivity of TRPV2 regulates intestinal motility
We also analyzed the expression of TRPV2 in the mouse intestine [22]. TRPV2-immunoreactive (IR) cells were co-stained with the neuronal marker PGP9.5; however, TRPV2-IR was not observed in GFAP (glial fibrillary acidic protein)-IR enteroglia or KIT-IR ICCs (interstitial cells of Cajal). These results indicated that TRPV2 was expressed in neurons, but not in enteroglia or ICCs. About half of the PGP9.5-IR sensory neurons were TRPV2-positive. Furthermore, most of the nNOS (neuronal nitric oxide synthase)-IR neurons were TRPV2 positive. It has been reported that nNOS-IR neurons are inhibitory circular muscle motor neurons, inhibitory longitudinal muscle motor neurons and descending interneurons. In the inhibitory motor neurons, even slight pressure could evoke TRPV2-mediated current responses, suggesting that enteric neurons expressing TRPV2 might be responsive to weak intestinal distortion. These results strongly suggested that intestinal TRPV2 [22] is also mechanosensitive, similar to its role in the DRG and spinal cord [21]. We examined nitric oxide (NO) production in specimens from the small intestine that were stimulated by a TRPV2 agonist using an enzymatic Griess reaction, and confirmed that TRPV2 activation induces NO release from inhibitory motor neurons which relaxes intestinal tone (Fig. 3) [22]. Furthermore, we also revealed that TRPV2 contributes to transit within the gastrointestinal tract in vivo. Thus, TRPV2 might be a good molecular target for developing a cure for irritable bowel syndrome.
Fig. 3.
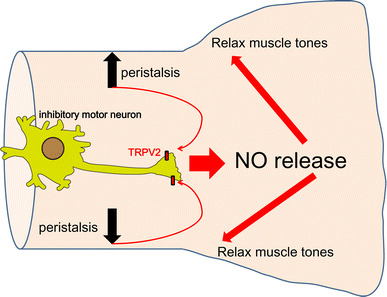
TRPV2 activation by peristalsis relaxes intestinal muscle tone. The inhibitory motor neurons (yellow) express TRPV2 in the intestine (beige) as shown by red images. The TRPV2 is activated by membrane stretch derived from peristalsis shown by black arrows. The activation of TRPV2 caused high Ca2+ influx and NO release. The NO release relaxes muscle tone, and contributes to transit within the gastrointestinal tract
TRPV2 in glial and immune cells
In addition to neurons, glial cells are also important for the maintenance of brain function. Astrocytes provide metabolic support and eliminate waste products, such as neurotransmitters, from the extracellular synaptic space [27]. These glia also regulate blood flow, depending on neuronal activity [28, 29]. Astrocytes are also essential for bidirectional communication with neurons, and thus can modulate neuronal activity [30–33]. We have shown that astrocytes in the brain express functional TRPV2 [23], and astrocytic TRPV2 is activated by lysophosphatidylcholine (LPC), a known endogenous lipid ligand of TRPV2. In a separate study we demonstrated that another thermo-TRP member, TRPV4, was only expressed in a specific subtype of astrocytes [32, 34]. This astrocytic TRPV4 responded to arachidonic acid, and modulated neuronal excitability through the release of gliotransmitters [32]. During the generation of arachidonic acid at postsynaptic sites, LPC is simultaneously generated, which could affect the excitation of astrocytes through TRPV2 activation. Therefore, astrocytic TRPV2 might be activated in response to increased lipid metabolism at synaptic sites.
Microglial cells also express TRPV2 [35]. Microglia are present throughout the central nervous system (CNS), and are activated by numerous pathogens. The characteristics of microglial cells are very similar to those of macrophages. Using TRPV2KO mice, it was clearly shown that TRPV2 is involved in the phagocytosis of macrophages [36]. It has also been reported that an inflammatory peptide, fMetLeuPhe, could induce membrane-translocation of TRPV2, and TRPV2 is expressed in immune-related tissues such as the spleen [9, 36, 37]. TRPV2 expression is also found in mast cells and lymphocytes [38–40]. One study of mast cells revealed that physical stimuli which lead to cellular degranulation are associated with TRPV2 activation and Ca2+ entry [39]. Thus, these studies highlight the importance of TRPV2 function in numerous immune cell types.
Circulatory function of TRPV2
TRPV2 is also expressed in endothelial cells and the smooth muscle of arteries and veins [41, 42]. It has been reported that hypotonic swelling of aortic myocytes induced Ca2+ influx through TRPV2 activation [43]. Furthermore, it was recently reported that TRPV2-deficient neonatal cardiomyocytes did not form intercalated discs and showed an extracellular Ca2+-dependent increase in intracellular Ca2+ and insulin-like growth factor (IGF-1) secretion in response to stretch stimulation [44]. Moreover, the elimination of TRPV2 from the mouse heart resulted in severe cardiac dysfunction through myocardial conduction defects [44]. Hence, TRPV2 is a critical determinant in the maintenance of cardiac structure and function. Several lines of evidence indicate that TRPV2 acts as a mechanosensor within circulatory organs [43]. However, the mechanisms involved in TRPV2 channel opening by membrane stretch or hypotonic swelling have not been identified. Ankyrin repeats, which are present at the N-terminal region of TRPV2, are thought to be important for mechanosensory function based on the loss of TRPV4 heat-sensitivity by the deletion of its ankyrin repeats [45]. Ankyrin repeats interact with several cytoskeletal proteins [18]; therefore, the interaction between TRPV2 and the cytoskeleton may be necessary for mechanosensory function. Additionally, it is also possible that membrane stretch or cellular swelling produces an endogenous lipid ligand that activates TRPV2. As described above, LPC is generated during arachidonic acid formation and, when membrane stretch occurs, it induces the generation of arachidonic acid. Therefore, it is highly possible that LPC affects TRPV2 activation [23].
TRPV2 in the pancreas
TRPV2 is highly expressed in MIN6 cells, a β-type insulinoma cell line. Under unstimulated conditions, TRPV2 is mainly localized within the cytoplasm. In contrast, application of exogenous insulin induces the translocation of TRPV2 from the cytoplasm to the plasma membrane [46]. This translocation evokes Ca2+ entry through TRPV2, resulting in insulin secretion. TRPV2 is also expressed in mouse β-cells prepared from pancreatic islets, but not in α-cells [46]. In addition to insulin, glucose can also induce the translocation of TRPV2 from the cytoplasm to the plasma membrane, and knockdown of TRPV2 reduces insulin secretion. In β-cells, the expression level of TRPV2 in the plasma membrane is also affected by α-Klotho, a known regulator of calcium homeostasis [47]. It was shown that α-Klotho regulates the translocation of TRPV2 from the cytoplasm to the plasma membrane in β-cells, and acts upstream of insulin secretion [48]. These results indicate that pancreatic TRPV2 is an important key regulator of glucose homeostasis.
Conclusion
In this review, we focused on the physiological importance of TRPV2 ion channels as mechanosensors, thermosensors and lipid sensors. TRPV2 is broadly expressed in numerous organ systems (Table 1) where it displays tissue-specific physiological functions. For example, TRPV2 is involved in axon outgrowth, consistent with its cellular localization in the axon shafts and growth cones in embryonic DRG sensory and spinal motor neurons. We have previously demonstrated that TRPV2 has two distinct roles, depending on developmental stage: during embryonic development it regulates axon outgrowth as a mechanosensor [21] and in adults it regulates thermosensation and/or nociception where it functions as a heat sensor [16]. This was the first characterization of an embryonic stage-specific physiological role for a thermo-TRP channel. We also reported that TRPV2 functions as a mechanosensor in the intestine where it contributes to transit within the gastrointestinal tract [22]. Combined, these results demonstrate that the same channel can have many distinct functions depending on its location. As described above, TRPV2 can also act as a lipid sensor, therefore further identification of novel physiological roles for TRPV2 will be dependent on its pattern of expression. Recently, many reports have indicated that TRPV2 is involved in the disease progression of bladder or prostate cancer [49, 50]. It was found that there was a significant relationship between the overexpression of TRP genes, including TRPV2, and the survival of patients with glioblastoma [51]. These results demonstrate that TRP channels contribute to the progression and survival of glioblastoma patients. Thus, future studies into the physiological function of TRPV2 may reveal new mechanisms behind several diseases.
Table 1.
Summary of TRPV2 function and related disease
| Organ | Cells | Function | Related disease |
|---|---|---|---|
| Brain | Neuron, astrocyte | Synaptic and glial transmission | Depression |
| DRG | Neuron | Nociception | Pain |
| Heart | Myocyte | Myocardial conduction | Cardiac hypertrophy |
| Pancreas | β-Cells | Insulin secretion | Diabetes |
| Intestine | Myenteric neuron | Intestinal motility | Irritable bowel syndrome |
| Spleen | Macrophage, mast cell, lymphocyte | Immune response | Immunodeficiency |
| Urinary bladder | Epithelial cell | Sensing stretch | Bladder cancer |
| Prostate | Epithelial cell | LPC receptor | Prostate cancer |
| Bone | Osteoclast | Calcium oscillation | Cancer |
| Muscle | Skeletal and cardiac muscle cells | Sensing stretch | Muscular dystrophy |
| Blood vessel | Smooth muscle, endothelial cell | Blood pressure control | Cardiomyopathy |
Acknowledgments
The author would like to acknowledge the following funding sources: Grants-in-Aid for Scientific Research (KAKENHI Project No. 15H05934 <Thermal Biology>, 21200012, 20399554, 24111507 + 26111702 <Brain Environment>, 26117502 <glial assembly>, 15H03000) from the Ministry of Education, Culture, Sports, Science and Technology, Japan; a grant from Uehara Memorial Foundation; a grant from Takeda Science Foundation, Tokyo, Japan; a grant from the Sumitomo Foundation; a grant from the Brain Science Foundation; a grant from Narishige Neuroscience Research Foundation; a grant from Salt Science Research Foundation No.14C2; and a grant from the Ichiro Kanehara Foundation.
References
- 1.Minke B. Drosophila mutant with a transducer defect. Biophys Struct Mech. 1977;3:59–64. doi: 10.1007/BF00536455. [DOI] [PubMed] [Google Scholar]
- 2.Montell C, Rubin GM. Molecular characterization of the Drosophila trp locus: a putative integral membrane protein required for phototransduction. Neuron. 1989;2:1313–1323. doi: 10.1016/0896-6273(89)90069-X. [DOI] [PubMed] [Google Scholar]
- 3.Tominaga M, Caterina MJ. Thermosensation and pain. J Neurobiol. 2004;61:3–12. doi: 10.1002/neu.20079. [DOI] [PubMed] [Google Scholar]
- 4.Levine JD, Alessandri-Haber N. TRP channels: targets for the relief of pain. Biochim Biophys Acta. 2007;1772:989–1003. doi: 10.1016/j.bbadis.2007.01.008. [DOI] [PubMed] [Google Scholar]
- 5.Chung MK, Jung SJ, Oh SB. Role of TRP channels in pain sensation. Adv Exp Med Biol. 2011;704:615–636. doi: 10.1007/978-94-007-0265-3_33. [DOI] [PubMed] [Google Scholar]
- 6.Cao E, Liao M, Cheng Y, Julius D. TRPV1 structures in distinct conformations reveal activation mechanisms. Nature. 2013;504:113–118. doi: 10.1038/nature12823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Paulsen CE, Armache JP, Gao Y, Cheng Y, Julius D. Structure of the TRPA1 ion channel suggests regulatory mechanisms. Nature. 2015;520:511–517. doi: 10.1038/nature14367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Nilius B. TRP channels in disease. Biochim Biophys Acta. 2007;1772:805–812. doi: 10.1016/j.bbadis.2007.02.002. [DOI] [PubMed] [Google Scholar]
- 9.Peralvarez-Marin A, Donate-Macian P, Gaudet R. What do we know about the transient receptor potential vanilloid 2 (TRPV2) ion channel? FEBS J. 2013;280:5471–5487. doi: 10.1111/febs.12302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fleig A, Penner R. The TRPM ion channel subfamily: molecular, biophysical and functional features. Trends Pharmacol Sci. 2004;25:633–639. doi: 10.1016/j.tips.2004.10.004. [DOI] [PubMed] [Google Scholar]
- 11.Colbert HA, Smith TL, Bargmann CI. OSM-9, a novel protein with structural similarity to channels, is required for olfaction, mechanosensation, and olfactory adaptation in Caenorhabditis elegans . J Neurosci Off J Soc Neurosci. 1997;17:8259–8269. doi: 10.1523/JNEUROSCI.17-21-08259.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kim J, Chung YD, Park DY, Choi S, Shin DW, et al. A TRPV family ion channel required for hearing in Drosophila . Nature. 2003;424:81–84. doi: 10.1038/nature01733. [DOI] [PubMed] [Google Scholar]
- 13.Caterina MJ, Schumacher MA, Tominaga M, Rosen TA, Levine JD, et al. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature. 1997;389:816–824. doi: 10.1038/39807. [DOI] [PubMed] [Google Scholar]
- 14.Tominaga M, Caterina MJ, Malmberg AB, Rosen TA, Gilbert H, et al. The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron. 1998;21:531–543. doi: 10.1016/S0896-6273(00)80564-4. [DOI] [PubMed] [Google Scholar]
- 15.Caterina MJ, Leffler A, Malmberg AB, Martin WJ, Trafton J, et al. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science. 2000;288:306–313. doi: 10.1126/science.288.5464.306. [DOI] [PubMed] [Google Scholar]
- 16.Caterina MJ, Rosen TA, Tominaga M, Brake AJ, Julius D. A capsaicin-receptor homologue with a high threshold for noxious heat. Nature. 1999;398:436–441. doi: 10.1038/18906. [DOI] [PubMed] [Google Scholar]
- 17.Kanzaki M, Zhang YQ, Mashima H, Li L, Shibata H, et al. Translocation of a calcium-permeable cation channel induced by insulin-like growth factor-I. Nat Cell Biol. 1999;1:165–170. doi: 10.1038/11086. [DOI] [PubMed] [Google Scholar]
- 18.Jin X, Touhey J, Gaudet R. Structure of the N-terminal ankyrin repeat domain of the TRPV2 ion channel. J Biol Chem. 2006;281:25006–25010. doi: 10.1074/jbc.C600153200. [DOI] [PubMed] [Google Scholar]
- 19.Cohen MR, Huynh KW, Cawley D, Moiseenkova-Bell VY. Understanding the cellular function of TRPV2 channel through generation of specific monoclonal antibodies. PLoS One. 2013;8:e85392. doi: 10.1371/journal.pone.0085392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Park U, Vastani N, Guan Y, Raja SN, Koltzenburg M, et al. TRP vanilloid 2 knock-out mice are susceptible to perinatal lethality but display normal thermal and mechanical nociception. J Neurosci Off J Soc Neurosci. 2011;31:11425–11436. doi: 10.1523/JNEUROSCI.1384-09.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Shibasaki K, Murayama N, Ono K, Ishizaki Y, Tominaga M. TRPV2 enhances axon outgrowth through its activation by membrane stretch in developing sensory and motor neurons. J Neurosci Off J Soc Neurosci. 2010;30:4601–4612. doi: 10.1523/JNEUROSCI.5830-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Mihara H, Boudaka A, Shibasaki K, Yamanaka A, Sugiyama T, et al. Involvement of TRPV2 activation in intestinal movement through nitric oxide production in mice. J Neurosci Off J Soc Neurosci. 2010;30:16536–16544. doi: 10.1523/JNEUROSCI.4426-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Shibasaki K, Ishizaki Y, Mandadi S. Astrocytes express functional TRPV2 ion channels. Biochem Biophys Res Commun. 2013;441:327–332. doi: 10.1016/j.bbrc.2013.10.046. [DOI] [PubMed] [Google Scholar]
- 24.Muraki K, Iwata Y, Katanosaka Y, Ito T, Ohya S, et al. TRPV2 is a component of osmotically sensitive cation channels in murine aortic myocytes. Circ Res. 2003;93:829–838. doi: 10.1161/01.RES.0000097263.10220.0C. [DOI] [PubMed] [Google Scholar]
- 25.Mochizuki T, Sokabe T, Araki I, Fujishita K, Shibasaki K, et al. The TRPV4 cation channel mediates stretch-evoked Ca2+ influx and ATP release in primary urothelial cell cultures. J Biol Chem. 2009;284:21257–21264. doi: 10.1074/jbc.M109.020206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Cohen MR, Johnson WM, Pilat JM, Kiselar J, DeFrancesco-Lisowitz A, et al. Nerve growth factor regulates transient receptor potential vanilloid 2 via extracellular signal-regulated kinase signaling to enhance neurite outgrowth in developing neurons. Mol Cell Biol. 2015;35:4238–4252. doi: 10.1128/MCB.00549-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kettenmann H, Ransom BR (eds) (2005) Neuroglia, 3rd edn. Oxford University Press, New York, pp 135–148
- 28.Takano T, Tian GF, Peng W, Lou N, Libionka W, et al. Astrocyte-mediated control of cerebral blood flow. Nat Neurosci. 2006;9:260–267. doi: 10.1038/nn1623. [DOI] [PubMed] [Google Scholar]
- 29.Mulligan SJ, MacVicar BA. Calcium transients in astrocyte endfeet cause cerebrovascular constrictions. Nature. 2004;431:195–199. doi: 10.1038/nature02827. [DOI] [PubMed] [Google Scholar]
- 30.Piet R, Jahr CE. Glutamatergic and purinergic receptor-mediated calcium transients in Bergmann glial cells. J Neurosci. 2007;27:4027–4035. doi: 10.1523/JNEUROSCI.0462-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sakatani S, Seto-Ohshima A, Shinohara Y, Yamamoto Y, Yamamoto H, et al. Neural-activity-dependent release of S100B from astrocytes enhances kainate-induced gamma oscillations in vivo. J Neurosci. 2008;28:10928–10936. doi: 10.1523/JNEUROSCI.3693-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Shibasaki K, Ikenaka K, Tamalu F, Tominaga M, Ishizaki Y. A novel subtype of astrocytes expressing TRPV4 (transient receptor potential vanilloid 4) regulates neuronal excitability via release of gliotransmitters. J Biol Chem. 2014;289:14470–14480. doi: 10.1074/jbc.M114.557132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Shibasaki K, Sugio S, Takao K, Yamanaka A, Miyakawa T, et al. TRPV4 activation at the physiological temperature is a critical determinant of neuronal excitability and behavior. Pflugers Arch. 2015;467:2495–2507. doi: 10.1007/s00424-015-1726-0. [DOI] [PubMed] [Google Scholar]
- 34.Shibasaki K, Suzuki M, Mizuno A, Tominaga M. Effects of body temperature on neural activity in the hippocampus: regulation of resting membrane potentials by transient receptor potential vanilloid 4. J Neurosci. 2007;27:1566–1575. doi: 10.1523/JNEUROSCI.4284-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hassan S, Eldeeb K, Millns PJ, Bennett AJ, Alexander SP, et al. Cannabidiol enhances microglial phagocytosis via transient receptor potential (TRP) channel activation. Br J Pharmacol. 2014;171:2426–2439. doi: 10.1111/bph.12615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Link TM, Park U, Vonakis BM, Raben DM, Soloski MJ, et al. TRPV2 has a pivotal role in macrophage particle binding and phagocytosis. Nat Immunol. 2010;11:232–239. doi: 10.1038/ni.1842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Nagasawa M, Nakagawa Y, Tanaka S, Kojima I. Chemotactic peptide fMetLeuPhe induces translocation of the TRPV2 channel in macrophages. J Cell Physiol. 2007;210:692–702. doi: 10.1002/jcp.20883. [DOI] [PubMed] [Google Scholar]
- 38.Zhang D, Spielmann A, Wang L, Ding G, Huang F, et al. Mast-cell degranulation induced by physical stimuli involves the activation of transient-receptor-potential channel TRPV2. Physiol Res Acad Sci Bohemoslov. 2012;61:113–124. doi: 10.33549/physiolres.932053. [DOI] [PubMed] [Google Scholar]
- 39.Stokes AJ, Shimoda LM, Koblan-Huberson M, Adra CN, Turner H. A TRPV2-PKA signaling module for transduction of physical stimuli in mast cells. J Exp Med. 2004;200:137–147. doi: 10.1084/jem.20032082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Saunders CI, Kunde DA, Crawford A, Geraghty DP. Expression of transient receptor potential vanilloid 1 (TRPV1) and 2 (TRPV2) in human peripheral blood. Mol Immunol. 2007;44:1429–1435. doi: 10.1016/j.molimm.2006.04.027. [DOI] [PubMed] [Google Scholar]
- 41.Park KS, Kim Y, Lee YH, Earm YE, Ho WK. Mechanosensitive cation channels in arterial smooth muscle cells are activated by diacylglycerol and inhibited by phospholipase C inhibitor. Circ Res. 2003;93:557–564. doi: 10.1161/01.RES.0000093204.25499.83. [DOI] [PubMed] [Google Scholar]
- 42.Fantozzi I, Zhang S, Platoshyn O, Remillard CV, Cowling RT, et al. Hypoxia increases AP-1 binding activity by enhancing capacitative Ca2+ entry in human pulmonary artery endothelial cells. Am J Physiol Lung Cell Mol Physiol. 2003;285:L1233–L1245. doi: 10.1152/ajplung.00445.2002. [DOI] [PubMed] [Google Scholar]
- 43.Muraki K, Iwata Y, Katanosaka Y, Ito T, Ohya S, et al. TRPV2 is a component of osmotically sensitive cation channels in murine aortic myocytes. Circ Res. 2003;93:829–838. doi: 10.1161/01.RES.0000097263.10220.0C. [DOI] [PubMed] [Google Scholar]
- 44.Katanosaka Y, Iwasaki K, Ujihara Y, Takatsu S, Nishitsuji K, et al. TRPV2 is critical for the maintenance of cardiac structure and function in mice. Nat Commun. 2014;5:3932. doi: 10.1038/ncomms4932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Watanabe H, Vriens J, Suh SH, Benham CD, Droogmans G, et al. Heat-evoked activation of TRPV4 channels in a HEK293 cell expression system and in native mouse aorta endothelial cells. J Biol Chem. 2002;277:47044–47051. doi: 10.1074/jbc.M208277200. [DOI] [PubMed] [Google Scholar]
- 46.Hisanaga E, Nagasawa M, Ueki K, Kulkarni RN, Mori M, et al. Regulation of calcium-permeable TRPV2 channel by insulin in pancreatic beta-cells. Diabetes. 2009;58:174–184. doi: 10.2337/db08-0862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Imura A, Tsuji Y, Murata M, Maeda R, Kubota K, et al. alpha-Klotho as a regulator of calcium homeostasis. Science. 2007;316:1615–1618. doi: 10.1126/science.1135901. [DOI] [PubMed] [Google Scholar]
- 48.Lin Y, Sun Z. Antiaging gene Klotho enhances glucose-induced insulin secretion by up-regulating plasma membrane levels of TRPV2 in MIN6 beta-cells. Endocrinology. 2012;153:3029–3039. doi: 10.1210/en.2012-1091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Caprodossi S, Lucciarini R, Amantini C, Nabissi M, Canesin G, et al. Transient receptor potential vanilloid type 2 (TRPV2) expression in normal urothelium and in urothelial carcinoma of human bladder: correlation with the pathologic stage. Eur Urol. 2008;54:612–620. doi: 10.1016/j.eururo.2007.10.016. [DOI] [PubMed] [Google Scholar]
- 50.Monet M, Lehen’kyi V, Gackiere F, Firlej V, Vandenberghe M, et al. Role of cationic channel TRPV2 in promoting prostate cancer migration and progression to androgen resistance. Cancer Res. 2010;70:1225–1235. doi: 10.1158/0008-5472.CAN-09-2205. [DOI] [PubMed] [Google Scholar]
- 51.Alptekin M, Eroglu S, Tutar E, Sencan S, Geyik MA, et al. Gene expressions of TRP channels in glioblastoma multiforme and relation with survival. Tumour Biol J Int Soc Oncodevelop Biol Med. 2015;36:9209–9213. doi: 10.1007/s13277-015-3577-x. [DOI] [PubMed] [Google Scholar]