

Abstract
Long noncoding RNAs (lncRNAs) have been shown to play a critical role in cancer development and progression. LncRNA metastasis-associated lung adenocarcinoma transcript 1 (MALAT-1) is a kidney cancer-associated onco-lncRNA involved in the progression of renal cell carcinoma (RCC). However, the pathological role of lncRNA MALAT-1 in RCC proliferation and metastasis remains poorly understood. This study was designed to investigate the biological role and mechanism of MALAT-1 in RCC proliferation and metastasis. The experiments were performed in human tissues, renal carcinoma cell lines, and nude mice. The expression of lncRNA MALAT-1, Livin mRNA, and the Livin protein was determined by quantitative real-time PCR (qRT-PCR) or a Western blot. The interaction between MALAT-1 and Livin was evaluated by RNA pull-down and RNA binding protein immunoprecipitation (RIP). Cell viability and apoptosis in RCC cell lines were detected using CCK-8 and TUNEL assays. LncRNA MALAT-1 and the Livin protein were highly expressed in RCC tissues, as well as in RCC 786-O and Caki-1 cell lines. MALAT-1 interference contributed to an increase in cell apoptosis and a reduction in the cell viability of 786-O and Caki-1 cells. The increase in apoptosis by si-MALAT-1 was reversed by overexpression of Livin. The RIP results showed that MALAT-1 promoted the expression of the Livin protein in 786-O and Caki-1 cells by enhancing the stability of the protein. Furthermore, the volume of si-MALAT-1-786-O cell xenograft was significantly suppressed. These data indicate that lncRNA MALAT-1-mediated promotion of RCC proliferation and metastasis may be due to the upregulation of the expression of Livin.
Electronic supplementary material
The online version of this article (doi:10.1007/s12576-016-0486-8) contains supplementary material, which is available to authorized users.
Keywords: LncRNA MALAT-1, Renal cell carcinoma, Cell proliferation and metastasis, Livin
Introduction
Malignant kidney tumors account for approximately 2 % of all cancers, and more than 273,000 new cases of this disease are diagnosed worldwide each year [1], among which renal cell carcinoma (RCC) accounts for more than 85 % of all cases. Although the treatment of RCC has improved greatly in recent years, the disease is still associated with substantial morbidity and mortality. In 2011, more than 275,000 women diagnosed with kidney cancer died, most of which occurred in developing countries, with death due to regional or distant metastasis [2, 3]. It is becoming increasingly clear that the molecular mechanism of RCC involves a comprehensive biological process that may include aberrant expression of protein-coding and noncoding genes [4]. Therefore, the discovery of new and effective prognostic markers and therapeutic strategies has the potential to improve the treatment of RCC.
The findings of various studies have pointed to the involvement of long noncoding RNAs (lncRNAs), which are more than 200nt RNA, in tumorigenesis. Metastasis-associated lung adenocarcinoma transcript 1 (MALAT-1), also known as noncoding nuclear-enriched abundant transcript 2, is a nuclear lncRNA, with a length of 8.7 kb, which is expressed by chromosome 11q13 [5]. Various studies have presented evidence that MALAT-1 may be an RCC-associated onco-lncRNA [6, 7]. For example, Zhang et al. found that RCC patients with higher levels of MALAT-1 had advanced clinical features and a shorter overall survival time than those with lower MALAT-1 expression [8]. Moreover, MALAT-1 silencing was shown to restrict RCC cell proliferation and invasion [9]. Although recent research explored the potential mechanism by which MALAT-1 promoted RCC aggression and tumorigenesis [8, 9], the exact mechanism remains unknown.
Livin, also known as ML-IAP, is a member of the inhibitor of apoptosis protein (IAP) family and was initially linked to malignant melanoma [10]. The IAP family is composed of structurally related proteins, some of which are thought to contribute to the development and therapeutic resistance of cancers [11]. Livin is a powerful predictor of tumor progression and patient survival in hepatocellular carcinoma [12], nonsmall cell lung cancer [13], colorectal [14], ovarian, and breast cancers [15, 16]. Livin has also been linked to RCC by encoding negative regulatory proteins that prevent cell apoptosis [10]. The relative overexpression of Livin in RCC indicates that it may represent a therapeutic target to increase the apoptotic sensitivity of kidney cancer cells [17]. As both Livin and MALAT-1 are onco-molecules, it is possible that there is a relationship between them.
The aim of the present study was to investigate whether lncRNA MALAT-1 was efficacious in promoting RCC proliferation and metastasis, with a particular focus on its upregulation of the expression of Livin. Our results may provide evidence for lncRNA MALAT-1 as a candidate for RCC therapy.
Materials and methods
Tissue samples
Matched fresh samples of tumor tissue and adjacent noncancerous tissue were obtained from 30 patients with clear cell RCC that has been confirmed by pathological diagnosis attending local hospital between 2013 and 2015 and were immediately frozen in liquid nitrogen. All the samples were stored at −80 °C until quantitative real-time PCR (qRT-PCR) or a Western-blot analysis was performed. The study was approved by the Medical Ethics Committee of the First People’s Hospital of Hangzhou. Written informed consent was obtained from all the participants.
Cell culture
Normal human HK-2 kidney cells and two RCC cell lines, 786-O and Caki-1, were purchased from the American Type Culture Collection (ATCC, USA) and routinely maintained in Dulbecco’s modified Eagle’s medium (DMEM, Gibco), with 10 % fetal bovine serum (Gibco), 100 U/ml of penicillin, and 100 mg/ml of streptomycin sulfate. All the cells were cultured in a humidified 5 % CO2 incubator at 37 °C and used when they were in the logarithmic growth phase.
RNA extraction and qRT-PCR
qRT-PCR was performed to detect lncRNA MALAT-1 and Livin mRNA expression. Total RNA was extracted from tissue samples using TRIzol reagent (Invitrogen, Carlsbad, CA, USA), according to the manufacturer’s instructions. RNA was reverse transcribed using the SuperScript First Strand cDNA System (Invitrogen). The PCR reaction with cDNA product and a SYBR Premix Ex Taq II kit (Takara, Dalian City, Liaoning Province, China) was performed on an Applied Biosystems 7500 Fast Real-Time PCR system (Applied Biosystems Life Technologies, Carlsbad, CA, USA), according to the manufacturer’s protocol. The relative level of MALAT-1 and Livin mRNA was calculated by the 2−ΔΔCt method and normalized to β-actin. The following primers for MALAT-1, Livin and GAPDH were used:
MALAT-1: forward: 5′-GAGTTCTAATTCTTTTTACTGCTCAATC-3′, reverse 5′- AGAGCAGA GCAGCGTAGAGC -3′
Livin: forward: 5′-CCAGGGTGGGCCCCGGGGGTCA-3′, reverse: 5′-GCGGGGTCCACAGCG CTCCTGC-3′.
GAPDH: forward: 5′-AACTTTGGCATTGTGGAAGG-3′, reverse: 5′-ACACATTGGGGGTAGG AACA-3′.
Protein extraction from fresh-frozen tissue and Western-blot analyses
Proteins were extracted from tumorous and nontumorous kidney specimens. Briefly, tissue sections were homogenized in lysis buffer, supplemented with protease inhibitors. Lysates were centrifuged at 10,000 × g for 15 min at 4 °C to collect supernatants. The total proteins in the supernatant were determined using a BCA Protein Assay kit (Thermo Scientific Pierce). Equal amounts of proteins were then separated by 12 % SDS–polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes. After blocking using a blocking solution, the membranes were exposed to monoclonal antibodies against Livin at 4 °C overnight and then incubated with horseradish peroxidase-labeled secondary antibody for 2 h under room temperature. The target gene was visualized by enhanced chemiluminescence (Beyotime, China), according to the manufacturer’s specifications.
RNA oligonucleotides and transfection
The siRNA sequences targeting human MALAT-1 (si-MALAT-1) or negative control RNA (si-control) were designed and produced by Genepharma (Shanghai, China). For overexpression of Livin and MALAT-1, RCC cells were transfected with pcDNA-Livin, and recombinant adenovirus-MALAT-1 was transfected with a pcDNA empty plasmid or Ad-GFP as a control (Invitrogen). All the transfection procedures were performed using Lipo2000 (QIAGEN) regent for 24 h and cells were collected for subsequent analysis.
Cell viability assay
Stable 786-O and Caki-1 cells (2 × 103 cells/well) were seeded in 96-well culture plates and incubated for 24 h in 100 μl of a culture medium. After incubation, 10 μl of CCK-8 solution (Seven Sea Biology Ltd. CO, Shanghai, China) were added to each well, and the plate was incubated for 4 h in a humidified atmosphere at 37 °C. Finally, the viability of the cells was measured at 405 nm using an ELISA reader (BioTek, Winooski, VT, USA), according to the manufacturer’s instructions. All the experiments were performed in triplicate, and the average value was then calculated.
TdT-mediated dUTP nick end labeling (TUNEL) analysis
A TUNEL analysis, a terminal deoxynucleotidyl transferase (TdT)-based end-labeling assay for DNA strand breaks [18], was performed to detect RCC cell apoptosis. In brief, the cells were fixed on slides using 4 % neutral buffered formalin. After washing with a PBS buffer, the slides were exposed to an equilibration buffer for 10 min at 37 °C and incubated with TdT reaction buffer for 30 min at 37 °C. A termination reaction buffer was then added to the samples. Stain was developed with DAB solution. The percentage of TUNEL-positive cells was calculated by dividing the number of apoptotic cells by the number of total cells in seven positive stainings.
RNA pull-down
The cells were harvested by trypsinization and incubated with a nuclear isolation buffer. The cell lysate was then incubated with streptavidin agarose Dynabeads (Invitrogen) at room temperature for 30 min, with continuous shaking. After movement of unbound proteins, the generated protein-coupled beads were blocked by adding 5 μg of recombinant L. braziliensis α-tubulin for 20 min. Then the proteins of the protein-coupled beads were interacted with biotinylated RNA in a pull-down 1 × buffer for 20 min. This protein–RNA complex was analyzed by a Western blot.
RNA binding protein immunoprecipitation (RIP)
RIP was used to evaluate the integration of MALAT-1 and Livin. A number of 1 × 105 cells/ml were fixed with 0.3 % formaldehyde and incubated with glycine before re-suspending in RIP buffer for 30 min. For RNA immunoprecipitation, antibodies to Livin were added to the samples, and they were incubated overnight at 4 °C. Precipitated RNAs were isolated using TRIzol RNA extraction reagent and analyzed using reverse transcription PCR. The retrieved MALAT-1 data are presented as an agarose gel image.
Tumor transplantation
All animal experiments were performed and conducted in accordance with the guidelines of the Provincial Hospital Affiliated to Shandong University. Six female athymic BALB/c nude mice (Shanghai, China) were injected with si-MALAT-1-transfected 786-O cells into the subcutaneous tissue of the right flank, and six nude mice were transplanted with si-control-transfected 786-O cells as a control. Tumor growth was monitored by two-dimensional measurements using electronic calipers starting from the tenth day after tumor transplantation (once every 4 days for a total of 30 days). The tumor volume was calculated using the formula a × b/2,
Statistical analysis
SPSS version 18.0 software (SPSS Inc., Chicago, IL, USA) was used for the analysis. All the experiments were repeated at least three times. Data are expressed as the mean ± SD. The Mann–Whitney U test was used to determine differences of two groups, and overall survival was estimated using the Kaplan–Meier method and log-rank test. Findings were considered significant when p < 0.05.
Results
LncRNA MALAT-1 and Livin protein were highly expressed in RCC
We performed qRT-PCR and a Western-blot analysis to detect the ectopic expression of MALAT-1 and Livin in 30 fresh clinical samples of RCC tumor tissues and paired adjacent noncancerous tissues. The results showed that compared to adjacent noncancerous tissues, RCC tumor tissues expressed higher expression levels of MALAT-1 (Fig. 1a). The level of the Livin protein was elevated in RCC tissues (Fig. 1b). However, there was no significant difference in Livin mRNA expression between the RCC tumor tissues and adjacent noncancerous tissues (Fig. 1b). Interestingly, there was a significant correlation between the expression levels of MALAT-1 and Livin protein in RCC cell lines (Fig. 1c).
Fig. 1.
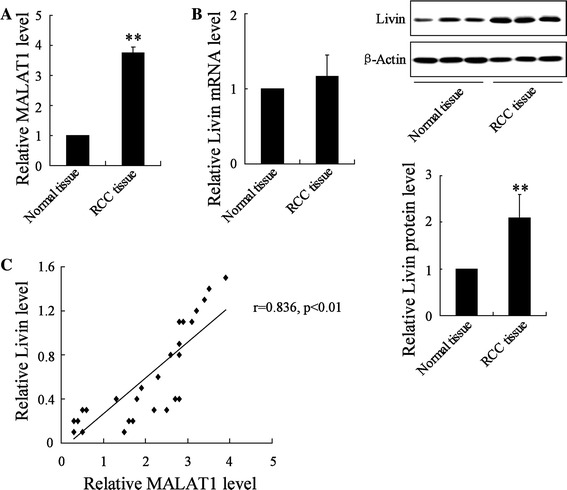
Determination of lncRNA MALAT-1 and Livin expression in RCC tissues. a The expression of MALAT-1 in RCC tumor tissues and noncancerous tissues was evaluated by qRT-PCR. b Livin mRNA and protein expression levels in RCC tumor tissues and adjacent normal tissues were determined by qRT-PCR and Western-blot analysis, respectively. c Correlation analysis between relative expression of Livin protein and MALAT1. **p < 0.01 vs. si-control
LncRNA MALAT-1 and Livin protein expression levels were significantly elevated in RCC cell lines
To investigate the involvement of MALAT-1 and Livin in RCC, we next performed cell experiments with two RCC cell lines, 786-O and Caki-1. We first confirmed the abnormal expression of lncRNA MALAT-1 and Livin protein rather than Livin mRNA using qRT-PCR and Western-blot analyses in 786-O and Caki-1 cells compared to that of human regular renal tubular epithelial cell HK-2, which was used as a control. The results were in keeping with Fig. 1 and revealed upregulation of MALAT-1 (Fig. 2a) and the Livin protein (Fig. 2b) in RCC cells compared to that of HK-2 cells. Relative expression level of Livin mRNA showed no significant differences among these three cell lines (Fig. 2b).
Fig. 2.
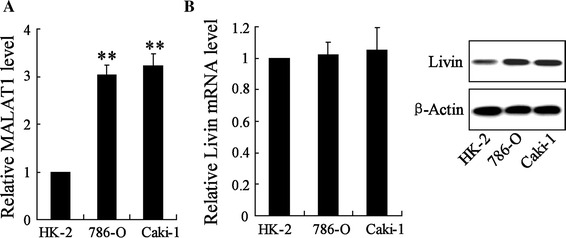
Expression of lncRNA MALAT-1 and Livin in RCC cell lines. a The expression of MALAT-1 in RCC cell lines (786-O and Caki-1) was measured by qRT-PCR, with that in regular renal cell (HK-2) as control. b Expression of Livin mRNA and protein were assessed in786-O, Caki-1, and HK-2. **p < 0.01 vs. HK-2 cells
LncRNA MALAT-1 knockdown significantly suppressed cell proliferation and markedly promoted apoptosis in RCC cell lines
To determine the involvement of MALAT-1 in RCC cell survival, we examined cell viability and cell apoptosis in 786-O and Caki-1 cells following MALAT-1 silencing. In the 786-O cells, the si-MALAT-1 treatment contributed to significant downregulation of MALAT-1. Furthermore, knockdown of MALAT-1 significantly reduced cell viability, whereas it induced an increase in the numbers of apoptotic cells (Fig. 3a). Caki-1 cells showed similar responses to si-MALAT-1, with significant downregulation of MALAT-1, reduced cell viability, and enhanced cell apoptosis (Fig. 3b).
Fig. 3.
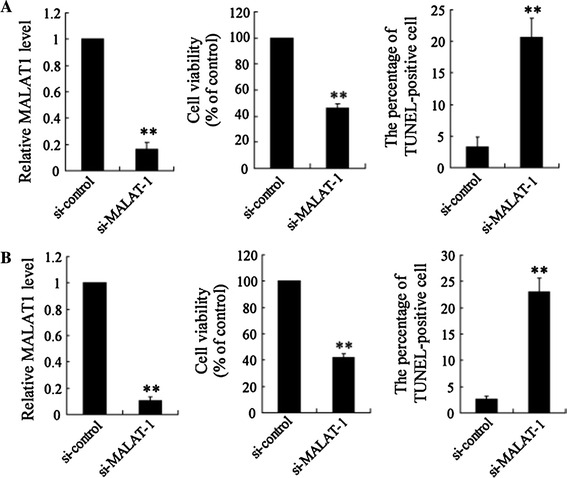
LncRNA MALAT-1 interference affected cell viability and apoptosis in RCC 786-O and Caki-1 cell lines. SiRNA was used to inhibit MALAT-1 expression to investigate its biological role in RCC cell viability and apoptosis. 786-O (a) and Caki-1 (b) cells were transfected with si-MALAT-1 or an si-control, respectively. Relative cell viability was assessed by a CCK-8 assay. Cell nuclei were visualized by DAPI staining, and DNA strand breaks in apoptotic cells were detected by a TUNEL assay. Data are presented as the mean ± SD. **p < 0.01 vs. si-control
MALAT-1 regulated and increased Livin levels by enhancing the stability of the protein
To evaluate the functional interaction between MALAT-1 and Livin, we determined the relative expression of Livin mRNA and the Livin protein in 786-O and Caki-1 cells transfected with si-MALAT-1. The results revealed no differences in the expression level of Livin mRNA in the cells transfected with si-MALAT-1 compared to those transfected with the si-control (Fig. 4a), whereas the expression of the Livin protein was dramatically decreased in the cells transfected with si-MALAT-1 as compared to that in the cells transfected with the si-control (Fig. 4b).
Fig. 4.
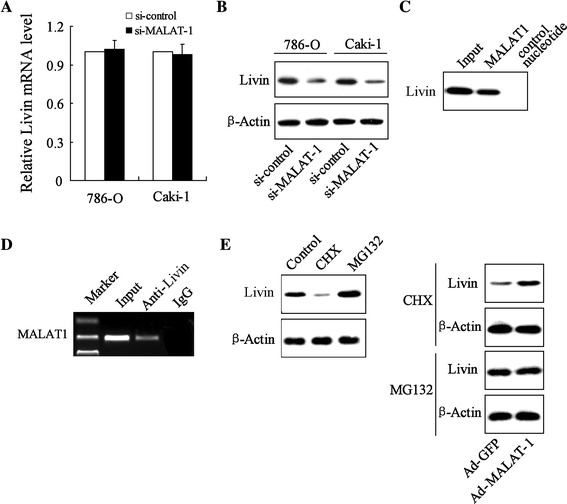
MALAT-1 elevated Livin levels by enhancing the stability of the Livin protein. 786-O and Caki-1 cells were transfected with si-MALAT-1 or an si-control, respectively. Livin mRNA (a) and Livin protein (b) levels in the 786-O and Caki-1 cell lines were detected by qRT-PCR and a Western-blot analysis, respectively. c An RNA pull-down assay was performed, as described in the experimental procedures. MALAT-1 or antisense RNA (NC) was incubated with cell extracts, and the Livin protein was assayed by Western blot. d RIP experiments were performed using a Livin antibody or nonspecific IgG, and specific primers were used to detect MALAT-1. MALAT-1-overexpressed 786-O cells were treated with CHX and MG132. After incubation for 4 h, Livin protein expression was detected by a Western-blot analysis (e)
We further explored the mechanism underlying the effect of MALAT-1 in increasing the expression of the Livin protein rather than Livin mRNA in RCC cells. In the RNA pull-down assay, the Livin protein was pulled down by MALAT-1 (Fig. 4c). The results of the RIP showed that the Livin antibody pulled down the MALAT-1 promotor (Fig. 4d). These data point to the specificity of the interaction between MALAT-1 and the Livin protein. CHX, a protein synthesis inhibitor, dramatically decreased Livin protein expression and MG132, a potent proteasome inhibitor, enhanced Livin expression. In contrast, in cells transfected with Ad-MALAT-1, the CHX treatment caused upregulation of Livin, and MG132 exerted no effect on Livin expression (Fig. 4e; Supplementary Fig. 1). These data point to the prostability of Livin by MALAT-1 in RCC cells.
Livin overexpression reversed the decreased cell viability and elevated apoptosis in RCC cell lines induced by lncRNA MALAT-1 interference
To further explore the roles of the interaction of lncRNA MALAT-1 and Livin in the cell survival of RCC cell lines, we examined cell viability and cell apoptosis in the 786-O and Caki-1 cells after transfection with si-MALAT-1 or the si-control. The data demonstrated that lncRNA MALAT-1 interference decreased cell viability and accelerated cell apoptosis in both RCC cell lines (Fig. 5a, b). However, Livin overexpression reversed the reduction of cell viability and increase in cell apoptosis.
Fig. 5.
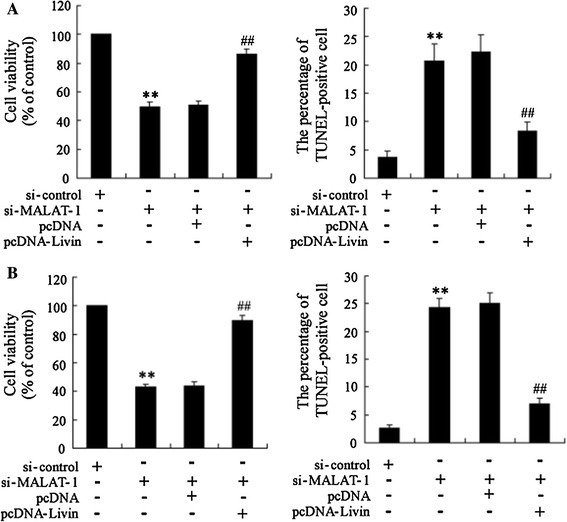
LncRNA MALAT-1 interference reduced cell viability and expedited apoptosis in RCC cell lines, and Livin overexpression reversed these processes. The relative cell viability and cell apoptosis of 786-O (a) and Caki-1 (b) cells was assessed. **p < 0.01 vs. si-control; ##p < 0.01 vs. si-MALAT-1 + pcDNA
LncRNA MALAT-1 interference inhibited tumor growth in a subcutaneous 786-O xenograft model
To determine the antitumor activity of lncRNA MALAT-1 knockdown in vivo, we assessed its effect in a nude mouse xenograft model of 786-O cells. Cultured 786-O cells were stably transfected with si-MALAT-1 or si-control and then subcutaneously injected into the right flank of the mice. After 30-day tumor growth, relative expression of MALAT1 in tumor xenograft was significantly attenuated (Fig. 6a); also, MALAT-1 knockdown resulted in smaller tumor size than si-control transfection 786-O cell. Moreover, during 30-day tumor growth, the tumor volume was measured every 4 days from day 10 after tumor transplantation. The data revealed that si-MALAT-1-786-O xenograft was smaller than si-control 786-O xenograft, with a significant difference emerging at 18 days (Fig. 6).
Fig. 6.
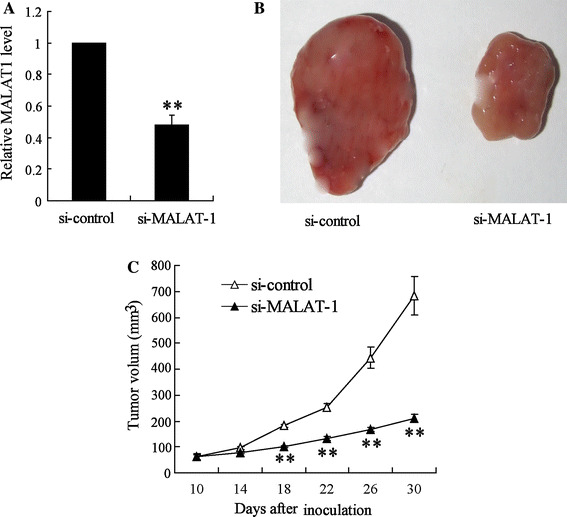
Effects of lncRNA MALAT-1 interference on the tumor growth of nude mice bearing 786-O xenograft tumors. Si-MALAT1 stable transfection 786-O cell line was transplanted into nude mice and the mice were sacrificed after 30 days. a Relative expression of MALAT1 in tumor xenograft. b Morphology of tumor xenograft. c The change in tumor volume was determined every 4 days during the tumor growth. Statistical analysis was performed by a Student’s t test, and all measurement data are expressed as the mean volume of xenograft tumors ± SEM. **p < 0.01 vs. si-control
Discussion
RCC is the most common form of adult kidney cancer, representing between 80 and 90 % of kidney cancers, and accounting for 23 % of all adult malignancies globally [19]. The incidence of RCC has increased in recent years [1, 19]. Although therapeutic surgery and drugs have improved outcomes, the prognosis of metastatic RCC is very poor, with 5-year survival rates of less than 10 % [20]. Recent studies have suggested that the identification of genes associated with the progression of prostate cancer is important for understanding the disease, and this is considered a major goal in cancer research. The present study found that MALAT-1 was associated with the Livin protein and contributed to the survival of RCC.
MALAT-1 has been shown to be a prognostic marker in patients with stage I lung cancer [21]. The MALAT-1 transcript, a lncRNA, is associated with human lung adenocarcinoma metastasis and dysregulated in many types of cancers in humans [22]. MALAT-1 has been shown to be overexpressed in many other tumors [23]. Studies reported that higher expression levels of MALAT-1 in human RCC tissues were associated with reduced patient survival [8, 9]. In the present study, lncRNA MALAT-1 and the Livin protein were upregulated in RCC tissues. As both Livin and MALAT-1 are onco-molecules, we focused on the functional interaction of MALAT-1 with Livin in RCC cell survival.
Tumor cell survival is essential for tumor progression, with survival depending on the apoptosis and proliferation of tumor cells [24]. The findings of the present study suggested that lncRNA MALAT-1 silencing significantly inhibited cell proliferation and promoted apoptosis in both RCC cell lines (786-O and Caki-1) and that MALAT-1 was a tumor promotor, as reported in other studies [25, 26]. The data also indicated that Livin was downregulated in response to MALAT-1 silencing. Livin is one of eight known human inhibitors abundantly expressed in fetal liver [24]. Livin alone was detected in various cancer cell lines and in specific tumor tissue cells, including RCC, but substantially lower levels or almost non-detectable levels were found in corresponding normal tissue, suggesting that its expression was largely tumor-specific [27, 28]. Recent studies suggested that targeted inhibition of Livin may present a novel tumor-specific therapeutic strategy [17].
The present study is the first to confirm the functional interaction between MALAT-1 and Livin. As shown in Fig. 4, the results of RNA pull-down and RIP revealed a direct interaction between MALAT-1 and Livin protein. Also, MALAT-1 silencing exerts an effect on expression of Livin mRNA. These data pointed to a direct interaction between MALAT-1 and the Livin protein. The results also suggested that CHX, a protein synthesis inhibitor, suppressed the expression of the Livin protein, whereas MG132, a proteinase inhibitor, enhanced its expression. Moreover, we found that the MG132 treatment exerted no effect on the expression of Livin when cellular MALAT-1 was overexpressed (Supplementary Fig. 1). These data confirmed that MALAT-1 caused upregulation of the Livin protein via lncRNA–protein interaction and protein stabilization. The targeted inhibition of Livin could provide a novel therapeutic strategy to lower the apoptotic threshold and tumor growth potential of RCC cells and to improve the apoptotic sensitivity and other therapeutic responses of all Livin-expressing RCC cell lines. A functional interaction was observed between MALAT-1 and Livin in si-MALAT-1 RCC cells cotransfected with pcDNA-Livin, and this interaction enhanced cell proliferation and suppressed cell apoptosis in comparison to that of si-MALAT-1. Overall, the findings indicate that upregulation of the Livin protein by MALAT-1 may be of great significance in enhancing tumor cell proliferation.
Conclusions
In summary, protumor lncRNA MALAT-1 promoted Livin protein expression by enhancing the stability of the protein. This interaction between MALAT-1 and Livin was essential for RCC cell viability and cell apoptosis. These results identify an important role for lncRNA MALAT-1 in the inhibition of RCC progression and suggest that it may be a promising candidate in RCC therapy.
Authors' contributions
Shaoan Chen is the first author of this paper and carried out the most designing work. Xunbo Jin is the corresponding author. Chuan Liu is also corresponding author, and participated in its design and helped to draft the manuscript. Ying Zhao performed the statistical analysis. Pengpeng Ma and Bin Li carried out the part of experiments. Pengpeng Ma also helped to draft the manuscript. Shaobo Jiang, Hui Xiong, Zheng Wang and Hanbo Wang participated in literature retrieval. All authors read and approved the final manuscript.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplemental Fig. 1 The signaling interaction of MALAT1 overexpression and Livin expression in presence of CHX and MG132 (TIFF 473 kb)
Acknowledgments
We are extremely grateful for Prof. Chuan Liu of the Second Affiliated Hospital of Chongqing Medical University for his technical guidance and support.
Funding
This study was supported by Natural Science Foundation of China (81300629) and the Natural Science Fund of Chongqing (cstc2013jcyjA0437).
Contributor Information
Xunbo Jin, Email: jinxbslyy@163.com.
Chuan Liu, Email: chuan_liucc@163.com.
References
- 1.Zhou S, Wang J, Zhang Z. An emerging understanding of long noncoding RNAs in kidney cancer. J Cancer Res Clin Oncol. 2014;140:1989–1995. doi: 10.1007/s00432-014-1699-y. [DOI] [PubMed] [Google Scholar]
- 2.Schiffman M, Wentzensen N, Wacholder S, Kinney W, Gage JC, Castle PE. Human papillomavirus testing in the prevention of cervical cancer. J Natl Cancer Inst. 2011;103:368–383. doi: 10.1093/jnci/djq562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011;61:69–90. doi: 10.3322/caac.20107. [DOI] [PubMed] [Google Scholar]
- 4.Maher ER. Genomics and epigenomics of renal cell carcinoma. Semin Cancer Biol. 2013;23:10–17. doi: 10.1016/j.semcancer.2012.06.003. [DOI] [PubMed] [Google Scholar]
- 5.Raitoharju E, Lyytikainen LP, Levula M, Oksala N, Mennander A, Tarkka M, et al. miR-21, miR-210, miR-34a, and miR-146a/b are up-regulated in human atherosclerotic plaques in the Tampere Vascular Study. Atherosclerosis. 2011;219:211–217. doi: 10.1016/j.atherosclerosis.2011.07.020. [DOI] [PubMed] [Google Scholar]
- 6.Kuiper RP, Schepens M, Thijssen J, van Asseldonk M, van den Berg E, Bridge J, et al. Upregulation of the transcription factor TFEB in t(6;11)(p21;q13)-positive renal cell carcinomas due to promoter substitution. Hum Mol Genet. 2003;12:1661–1669. doi: 10.1093/hmg/ddg178. [DOI] [PubMed] [Google Scholar]
- 7.Davis IJ, Hsi BL, Arroyo JD, Vargas SO, Yeh YA, Motyckova G, et al. Cloning of an Alpha-TFEB fusion in renal tumors harboring the t(6;11)(p21;q13) chromosome translocation. Proc Natl Acad Sci USA. 2003;100:6051–6056. doi: 10.1073/pnas.0931430100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zhang HM, Yang FQ, Chen SJ, Che J, Zheng JH. Upregulation of long non-coding RNA MALAT1 correlates with tumor progression and poor prognosis in clear cell renal cell carcinoma. Tumour Biol J Int Soc Oncodev Biol Med. 2015;36:2947–2955. doi: 10.1007/s13277-014-2925-6. [DOI] [PubMed] [Google Scholar]
- 9.Hirata H, Hinoda Y, Shahryari V, Deng G, Nakajima K, Tabatabai ZL, et al. Long noncoding RNA MALAT1 promotes aggressive renal cell carcinoma through Ezh2 and interacts with miR-205. Cancer Res. 2015;75:1322–1331. doi: 10.1158/0008-5472.CAN-14-2931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Crnkovic-Mertens I, Muley T, Meister M, Hartenstein B, Semzow J, Butz K, et al. The anti-apoptotic Livin gene is an important determinant for the apoptotic resistance of non-small cell lung cancer cells. Lung Cancer. 2006;54:135–142. doi: 10.1016/j.lungcan.2006.07.018. [DOI] [PubMed] [Google Scholar]
- 11.Chung SW, Chen YH, Perrella MA. Role of Ets-2 in the regulation of heme oxygenase-1 by endotoxin. J Biol Chem. 2005;280:4578–4584. doi: 10.1074/jbc.M409125200. [DOI] [PubMed] [Google Scholar]
- 12.Ding C, Yang Z, Lv Z, Du C, Xiao H, Peng C, et al. Long non-coding RNA PVT1 is associated with tumor progression and predicts recurrence in hepatocellular carcinoma patients. Oncol Lett. 2015;9:955–963. doi: 10.3892/ol.2014.2730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Yang YR, Zang SZ, Zhong CL, Li YX, Zhao SS, Feng XJ. Increased expression of the lncRNA PVT1 promotes tumorigenesis in non-small cell lung cancer. Int J Clin Exp Pathol. 2014;7:6929–6935. [PMC free article] [PubMed] [Google Scholar]
- 14.Takahashi Y, Sawada G, Kurashige J, Uchi R, Matsumura T, Ueo H, et al. Amplification of PVT-1 is involved in poor prognosis via apoptosis inhibition in colorectal cancers. Br J Cancer. 2014;110:164–171. doi: 10.1038/bjc.2013.698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Guan Y, Kuo WL, Stilwell JL, Takano H, Lapuk AV, Fridlyand J, et al. Amplification of PVT1 contributes to the pathophysiology of ovarian and breast cancer. Clin Cancer Res Off J Am Assoc Cancer Res. 2007;13:5745–5755. doi: 10.1158/1078-0432.CCR-06-2882. [DOI] [PubMed] [Google Scholar]
- 16.Crnkovic-Mertens I, Wagener N, Semzow J, Grone EF, Haferkamp A, Hohenfellner M, et al. Targeted inhibition of Livin resensitizes renal cancer cells towards apoptosis. Cell Mol Life Sci CMLS. 2007;64:1137–1144. doi: 10.1007/s00018-007-6510-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Wagener N, Crnkovic-Mertens I, Vetter C, Macher-Goppinger S, Bedke J, Grone EF, et al. Expression of inhibitor of apoptosis protein Livin in renal cell carcinoma and non-tumorous adult kidney. Br J Cancer. 2007;97:1271–1276. doi: 10.1038/sj.bjc.6604028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Li X, Darzynkiewicz Z. Labelling DNA strand breaks with BrdUTP. Detection of apoptosis and cell proliferation. Cell Prolif. 1995;28:571–579. doi: 10.1111/j.1365-2184.1995.tb00045.x. [DOI] [PubMed] [Google Scholar]
- 19.Badimon L, Vilahur G, Padro T. Nutraceuticals and atherosclerosis: human trials. Cardiovasc Ther. 2010;28:202–215. doi: 10.1111/j.1755-5922.2010.00189.x. [DOI] [PubMed] [Google Scholar]
- 20.Aiello RJ, Bourassa PA, Lindsey S, Weng W, Natoli E, Rollins BJ, et al. Monocyte chemoattractant protein-1 accelerates atherosclerosis in apolipoprotein E-deficient mice. Arterioscler Thromb Vasc Biol. 1999;19:1518–1525. doi: 10.1161/01.ATV.19.6.1518. [DOI] [PubMed] [Google Scholar]
- 21.Ji P, Diederichs S, Wang W, Boing S, Metzger R, Schneider PM, et al. MALAT-1, a novel noncoding RNA, and thymosin beta4 predict metastasis and survival in early-stage non-small cell lung cancer. Oncogene. 2003;22:8031–8041. doi: 10.1038/sj.onc.1206928. [DOI] [PubMed] [Google Scholar]
- 22.Lin R, Maeda S, Liu C, Karin M, Edgington TS. A large noncoding RNA is a marker for murine hepatocellular carcinomas and a spectrum of human carcinomas. Oncogene. 2007;26:851–858. doi: 10.1038/sj.onc.1209846. [DOI] [PubMed] [Google Scholar]
- 23.Andreou I, Sun X, Stone PH, Edelman ER, Feinberg MW. miRNAs in atherosclerotic plaque initiation, progression, and rupture. Trends Mol Med. 2015;21:307–318. doi: 10.1016/j.molmed.2015.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Seneviratne A, Hulsmans M, Holvoet P, Monaco C. Biomechanical factors and macrophages in plaque stability. Cardiovasc Res. 2013;99:284–293. doi: 10.1093/cvr/cvt097. [DOI] [PubMed] [Google Scholar]
- 25.Qi Y, Ooi HS, Wu J, Chen J, Zhang X, Tan S, et al. MALAT1 long ncRNA promotes gastric cancer metastasis by suppressing PCDH10. Oncotarget. 2016;7:12693–12703. doi: 10.18632/oncotarget.7281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Jin C, Yan B, Lu Q, Lin Y, Ma L. Reciprocal regulation of Hsa-miR-1 and long noncoding RNA MALAT1 promotes triple-negative breast cancer development. Tumour Biol J Int Soc Oncodev Biol Med. 2016;37:7383–7394. doi: 10.1007/s13277-015-4605-6. [DOI] [PubMed] [Google Scholar]
- 27.Treguer K, Heinrich EM, Ohtani K, Bonauer A, Dimmeler S. Role of the microRNA-17-92 cluster in the endothelial differentiation of stem cells. J Vasc Res. 2012;49:447–460. doi: 10.1159/000339429. [DOI] [PubMed] [Google Scholar]
- 28.Chan YC, Roy S, Khanna S, Sen CK. Downregulation of endothelial microRNA-200b supports cutaneous wound angiogenesis by desilencing GATA binding protein 2 and vascular endothelial growth factor receptor 2. Arterioscler Thromb Vasc Biol. 2012;32:1372–1382. doi: 10.1161/ATVBAHA.112.248583. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplemental Fig. 1 The signaling interaction of MALAT1 overexpression and Livin expression in presence of CHX and MG132 (TIFF 473 kb)