

Abstract
Bilobalide, a unique constituent of Ginkgo biloba, has been reported to potentiate population spikes in hippocampal CA1 pyramidal cells and to protect the brain against cell death. In this study, the effects of bilobalide on synaptic transmission and its plasticity in rat hippocampal subfields were electrophysiologically investigated. Bilobalide (50 μM) significantly potentiated the input–output relationship at Schaffer collateral (SC)-CA1 synapses but not at medial perforant path (MPP)-dentate gyrus (DG), lateral perforant path (LPP)-DG, or mossy fiber (MF)-CA3 synapses. Facilitative effects of bilobalide on synaptic plasticity were only observed at MPP-DG synapses, in which the induction of long-term depression was blocked in the presence of bilobalide. However, no effect on synaptic plasticity was observed at SC-CA1 synapses. These results suggest that bilobalide has differential effects on synaptic efficacy in each hippocampal subfield.
Keywords: Hippocampus, Bilobalide, Synaptic transmission, Plasticity, LTP, LTD
Introduction
Pharmacological therapies for memory improvement are urgently needed to remedy memory disorders. Among the brain regions related to memory function, the hippocampus has been intensively studied as a brain region that plays a crucial role in memory [1, 2], and hippocampal synaptic plasticity such as that related to long-term potentiation (LTP) is widely regarded as a candidate for the biological basis of memory [3]. Since animal studies have shown that the enhancement of LTP was associated with improved learning performance [4, 5] and that the experience of learning itself evokes hippocampal LTP [6], identifying chemicals that facilitate hippocampal synaptic responses and plasticity seems a useful strategy for developing effective anti-dementia drugs.
Ginkgo biloba extract has traditionally been used as a Chinese herbal medicine. EGb761®, an extract of Gingko biloba, has been reported to improve cognitive function in Alzheimer’s disease [7], decrease the number of errors in the radial maze [8], increase latency in the passive avoidance task in aged mice [9], reduce the threshold of population spikes, and potentiate the magnitude of LTP [10]. Moreover, the administration of EGb761® protects the brain against ischemia-induced cell death [11]. Although these results suggest the possibility of using Ginkgo biloba extract as an anti-dementia drug, the property of each constituent of the extract should be analyzed in order to develop an optimally effective anti-dementia drug.
Ginkgolides A, B, C, J, and bilobalide (GA, GB, GC, GJ, and BB), which are also known as terpene trilactones (TTL), are unique components of Ginkgo biloba. TTL are believed to play important roles in the biologically active properties of the extract [12]. In this study, we focused on bilobalide, which can be isolated from Ginkgo biloba leaves. Bilobalide reportedly increases the magnitude of population spikes and the amplitude of secondary population spikes by paired-pulse stimulation at hippocampal CA1 synapses [13] in addition to protecting the brain against cell death [11]; however, the effects of bilobalide on synaptic transmission and plasticity at hippocampal synapses are still unclear. In the present study, we isolated and purified bilobalide from Gingko biloba leaves and compared the effects of bilobalide on synaptic transmission and plasticity among rat hippocampal subfields.
Materials and methods
Isolation of bilobalide
Isolation of 9-tert-butyl-8,9-dihydroxy-dihydro-furo[2,3-b]furo[3′,2′:2,3]cyclopenta[1,2-c]furan-2,4,7-trione, bilobalide (BB), is as follows. The green leaves of female Ginkgo biloba (250 g) were frozen in liquid N2 and crushed with a mortar. The crushed leaves and EtoAc (800 mL) were added to a 2-L flask and kept in a water bath with a reflux condenser at 80°C for 1 h. After being cooled to room temperature, the solution was filtered using a glass filter covered with Celite 545. The filtrate was washed with H2O, then with saturated aqueous Na2SO3. After removing the solvent using a rotary evaporator, the organic layer was stirred with activated charcoal (25 g, decolorizing; Sigma Aldrich) in EtOAc for 1 h at room temperature. After filtration and evaporation, the organic layer was stirred with activated charcoal (15 g) in MeOH for 1 h at room temperature. After filtration and evaporation, a rough extract (803 mg) was obtained. The rough extract in a minimum amount of EtOAc was loaded onto a silica gel (60 spherical, acidic) for column chromatography (Kanto Chemicals). The column was then eluted with hexane/EtOAc solvent mixtures. The fraction collected at hexane/EtOAc = 5/1 → 3/1 contained ginkgolic acids. The fraction collected at hexane/EtOAc = 1/1 contained pure bilobalide (126 mg) and then a ginkgolide mixture (74 mg). Bilobalide was identified with TLC (silica gel 60 F254 plates produced by Merck), 1H NMR (recorded on a JEOL Lambda 300 spectrometer in deuterium solvents using MeOH as the internal standard), and ESI-MS (recorded on a JEOL JMS-T100LC instrument): colorless crystal; R f = 0.67 (silica gel, hexane/EtOAc = 1/2); 1H NMR (300 MHz, MeOH-d 4) δ 6.30 (1H, s, H12), 5.17 (1H, s, H10), 5.04 (1H, t, J = 7.0 Hz, H6), 3.03 (1H, d, J = 18.0 Hz, H1α), 2.80 (1H, d, J = 18.0 Hz, H1β), 2.62 (1H, dd, J = 7.0, 6.6 Hz, H7α), 2.29 (1H, dd, J = 6.7, 6.6 Hz, H7β), 1.13 (9H, s, t-Bu); ESI-MS (m/z) calcd for C15H18O8Na [M+Na]+ 349.09, found 349.11.
In order to efficiently remove impurities such as ginkgolic acids, we optimized the purification procedure by using activated charcoal for the extraction step, as described above. Filtration and/or shaking of the extract with charcoal are normally carried out to remove impurities [14]. We have improved this procedure by stirring the obtained rough Ginkgo extract with activated charcoal and found that washing the extract twice with charcoal gave more efficient results. In this way, ca. 100–150 mg of bilobalide were isolated from 250 g of wet leaves in 1 day.
Preparations
Hippocampal slices (400 μm thick) were obtained from 3-week-old male Wistar rats. The details of the procedures have been described elsewhere [15]. In short, the rats were deeply anesthetized using pentobarbital sodium, and their brains were removed quickly. Each hippocampus was chilled with ice-cold artificial cerebrospinal fluid (ACSF) and cut using a microslicer (DTK-1000; Dosaka, Kyoto, Japan). The slices were incubated for more than 1 h in ACSF at 32°C. The ASCF contained (in mM) 125 NaCl, 2.5 KCl, 2 CaCl2, 1 MgCl2, 26 NaHCO3, 1.25 NaH2PO4, and 10 d-glucose, saturated with 95% O2 and 5% CO2 (pH adjusted to 7.4). All procedures were performed in accordance with the Japanese Physiological Society’s Guiding Principles for the Care and Use of Animals in the Field of Physiological Sciences.
Electrophysiological recordings
Each slice was then transferred to a recording chamber on the stage of a BX51WI upright microscope (Olympus, Tokyo, Japan) and held down with a U-shaped platinum wire and several nylon threads. The slices were continuously perfused with control ACSF at a rate of 1.5–2.0 mL/min, and the ACSF temperature was maintained at 32 ± 1°C during each experimental session using a TC-324B inline heater (Warner Instrument, Hamden, CT, USA). A tungsten concentric bipolar electrode (Unique Medical, Tokyo, Japan) was placed onto presynaptic fibers (medial perforant path, MPP; lateral perforant path, LPP; mossy fibers, MF; or Schaffer collaterals, SC). A glass microelectrode filled with control medium was placed onto the middle molecular layer (for MPP-DG recording), outer molecular layer (for LPP-DG recording), stratum lucidum (for MF-CA3 recording), or the stratum radiatum (for SC-CA1 recording) to record extracellular field excitatory postsynaptic potentials (fEPEP). Selective stimulation of MF was examined by applying the group II metabotropic glutamate receptor (mGluR) agonist DCG-IV (1 μM) for 10 min after the recordings [16, 17], and the data for fEPSP that decreased by >70% after DCG-IV application were used for analysis. Electrical stimulation was controlled using an electronic stimulator and isolator (Master-8 and ISO-Flex; AMP Instruments, Jerusalem, Israel). The frequency of test pulse application was 0.1 Hz (duration 100 μs), and the intensity was set to evoke fEPSP with an initial gradient of about 50% of the maximal value. Stable fEPSP were recorded for at least 10 min with the test pulses under the normal ACSF conditions. Then, input–output relationships were recorded [18, 19] by changing the intensity of the test pulse from minimal to maximal intensity to evoke fEPSP. After that, bilobalide (50 μM) was applied, and the stable fEPSP were recorded for at least 10 min. Then, the input–output relationships were recorded again. To induce synaptic plasticity, high frequency stimulation (HFS, 100 Hz, 1 s) was applied after stable fEPSP had been recorded for 10 min in the presence or absence of bilobalide. The magnitudes of the fEPSP were evaluated by comparing the responses observed 50–60 min after HFS with those seen at the baseline. Bilobalide was perfused for at least 10 min before and 10 min after HFS. In the case of MPP-DG synapses, the concentration of CaCl2 was increased from 2 to 2.5 mM and that of MgCl2 was decreased from 1 to 0.5 mM. To separate the MPP-DG and LPP-DG synaptic responses, paired-pulse stimuli (50 ms interstimulus interval) were applied before the baseline recordings because it was reported that MPP-DG synapses show paired-pulse depression (PPD) and LPP-DG synapses show paired-pulse facilitation (PPF) [20]. The fEPSP were amplified through a CEZ-2300 patch-clamp amplifier (current-clamp mode) (Nihon-Kohden, Tokyo, Japan) and filtered at 1 Hz (low cut) and 1 kHz (high cut). The pCLAMP system (version 8; Axon Instruments, Foster City, CA, USA) was used for data acquisition and analysis. Bilobalide dissolved in dimethyl sulfoxide (DMSO) (200 mM stock solution) was added to the perfused ACSF at a final concentration of 50 μM (0.025% DMSO), and strychnine (STR) dissolved in DMSO (10 mM stock solution) was added to the perfused ACSF at a final concentration of 2 μM (0.02% DMSO). Medium containing bilobalide or strychnine was made daily.
Materials
Picrotoxin (PTX), strychnine, and DMSO were obtained from Sigma (St. Louis, MO, USA), (2S,2′R,3′R)-2-(2′,3′-dicarboxycyclopropyl)glycine (DCG-IV) was purchased from Tocris (Ellisville, MO, USA), and all other chemicals were obtained from Wako Chemicals (Tokyo, Japan).
Statistics
The relative amplitude of fEPSP is shown in the text and figures as the mean ± SE. Statistical analyses were performed with two-tailed t tests for paired data or non-paired data, using SAS (version 9; SAS Institute, Cary, NC, USA). Significance was established at p < 0.05, except for when Levene’s test of the equality of variances was used (p < 0.2). When variances were found to be unequal, t tests were performed using the Satterthwaite correction.
Results
First, to examine whether bilobalide affected synaptic transmission at SC-CA1 synapses, we compared the input–output relationships between experiments conducted in the absence and presence of bilobalide by recording the magnitudes of presynaptic fiber volleys and fEPSP (Fig. 1a). As a result, we found that the slopes of each linear regression were significantly different between the two conditions [F (2, 33) = 5.64, p < 0.05]. Additionally, the averaged minimal magnitude of the presynaptic fiber volley evoking the maximal fEPSP in the presence of bilobalide was also significantly smaller than that seen in the absence of bilobalide [CTL 0.96 ± 0.04, BB 0.73 ± 0.09; t (4) = 3.03, p < 0.05; n = 5]. The finding that activation by a smaller presynaptic fiber volley caused a larger fEPSP in the presence of bilobalide demonstrates the facilitative effect of bilobalide on synaptic transmission at SC-CA1 synapses.
Fig. 1.
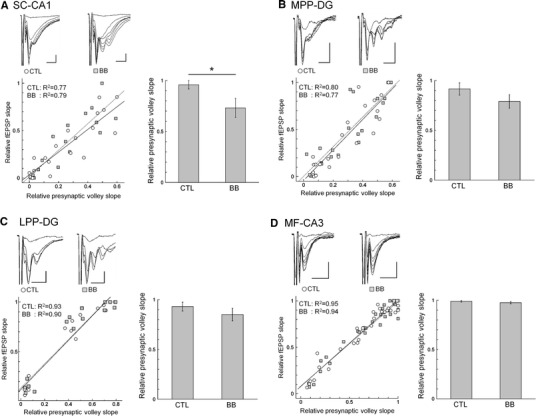
The effects of bilobalide on hippocampal synaptic transmission. Left Relative fEPSP slopes were plotted against the relative presynaptic volley slopes at SC-CA1 (a), MPP-DG (b), LPP-DG (c), and MF-CA3 (d) synapses. Linear regressions were calculated under each condition (solid line control conditions, dotted line bilobalide conditions). Inset The typical waveforms in the absence (left) and presence (right) of bilobalide. Scales 0.5 mV, 5 ms. Right Averaged minimal magnitude of the presynaptic fiber volley required to evoke the maximal fEPSP. *p < 0.05
On the other hand, the MPP-DG synapses showed no difference in their input–output relationships between the absence and presence of bilobalide [F (2, 38) = 0.39, n.s.; Fig. 1b], and the minimal presynaptic fiber volley value required to evoke fEPSP in the presence of bilobalide did not significantly differ from that seen under the control conditions [CTL 0.92 ± 0.06, BB 0.79 ± 0.07; t (6) = 1.68, n.s.; n = 7]. Likewise, at LPP-DG synapses, there were no significant differences in the linear regression slopes [F (2, 40) = 0.03, n.s.; Fig. 1c] or the minimal presynaptic fiber volley values required to evoke the maximal fEPSP [CTL 0.93 ± 0.04, BB 0.85 ± 0.06; t (4) = 1.06, n.s.; n = 7] regardless of the presence or absence of bilobalide. Also, at MF-CA3 synapses, the linear regression slopes in the absence and presence of bilobalide [F (2, 68) = 0.19, n.s.; Fig. 1d] and the minimal of presynaptic fiber volley value required to evoke for maximal fEPSP were not significantly different [CTL 0.99 ± 0.01, BB 0.96 ± 0.01; t (5) = 0.91, n.s.; n = 6]. Taken together, a facilitative effect of bilobalide on synaptic transmission was only detected at SC-CA1 synapses among the hippocampal subfields.
Next, we investigated the effects of bilobalide on plasticity at hippocampal synapses. At SC-CA1 synapses, the relative fEPSP slope at 50–60 min after HFS was significantly potentiated in both the absence of bilobalide [1.43 ± 0.15; t (7) = 3.11, p < 0.05, n = 8; Fig. 2a] and the presence of bilobalide [1.34 ± 0.10; t (6) = 3.39, p < 0.05; n = 7]. However, the magnitude of LTP in the presence of bilobalide did not differ from that seen under the control conditions [t (13) = 0.47, n.s.]. Thus, no effect of bilobalide on synaptic plasticity at SC-CA1 synapses was detected.
Fig. 2.
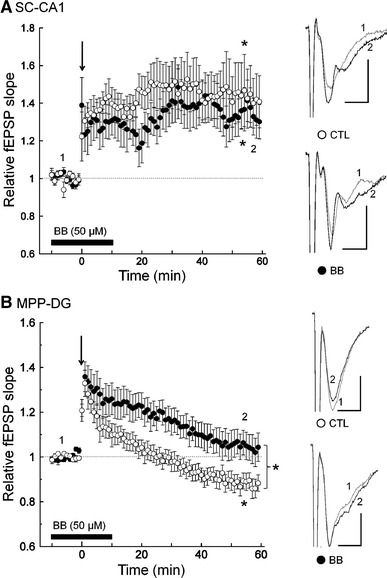
The effects of bilobalide on hippocampal synaptic plasticity. a HFS (arrow 100 Hz, 1 s) induced LTP at SC-CA1 synapses. The black bar shows the timing of the application of bilobalide. The magnitudes of potentiation under the bilobalide conditions did not differ from those seen under the control conditions. Insets Typical waveforms of synaptic responses before (1) and 60 min after (2) the HFS. Scales 0.5 mV, 5 ms. *p < 0.05. b HFS induced LTD under the control conditions at MPP-DG synapses, and bilobalide blocked the induction of LTD. Insets Typical waveforms of synaptic responses before (1) and 60 min after (2) the HFS. Scales 0.5 mV, 5 ms. *p < 0.05
At MPP-DG synapses, which have almost identical LTP induction properties to SC-CA1 synapses [20, 21], HFS induced long-term depression (LTD) under our control conditions [0.88 ± 0.04; t (9) = 3.01, p < 0.05, n = 10; Fig. 2b]. On the other hand, HFS induced neither LTD nor LTP in the presence of bilobalide [1.05 ± 0.06; t (7) = 0.80, n.s., n = 8], and the relative fEPSP slope at 50–60 min after HFS in the presence of bilobalide was significantly larger than that seen in the absence of bilobalide [t (16) = 2.46, p < 0.05]. These results show that bilobalide has a facilitative effect on synaptic plasticity at MPP-DG synapses; i.e., that the synaptic responses were rescued from LTD.
Since bilobalide antagonizes GABAA receptors and glycine receptors [22], we examined the possibility that the blockade of these systems is involved in the facilitative effects of bilobalide on the plasticity of MPP-DG synapses. When the GABAA receptor antagonist PTX (50 μM) was applied, HFS did not induce either LTD or LTP [1.10 ± 0.07; t (9) = 1.21, n.s.; n = 10] (Fig. 3), which was similar to the facilitative effects of bilobalide (Fig. 2). Note that the co-application of PTX and bilobalide resulted in further potentiation of synaptic responses; i.e., LTP was induced by HFS in the presence of both PTX and bilobalide [1.35 ± 0.08; t (8) = 4.01, p < 0.01; n = 9], and the synaptic responses measured at 50–60 min after the application of HFS were significantly larger than those seen in the control [t (11.7) = 5.52, p < 0.01], bilobalide [t (15) = 3.07, p < 0.01], and PTX [t (17) = 2.53, p < 0.05] conditions, demonstrating that neither PTX nor bilobalide completely inhibited each other’s facilitative effects. On the other hand, HFS did induce LTD in the presence of the glycine receptor antagonist strychnine (2 μM) [0.84 ± 0.06; t (4) = 2.81, p < 0.05, n = 5] (Fig. 3), suggesting that blockade of glycinergic system is not involved in the facilitative effects of bilobalide on HFS-induced MPP-DG synaptic plasticity.
Fig. 3.
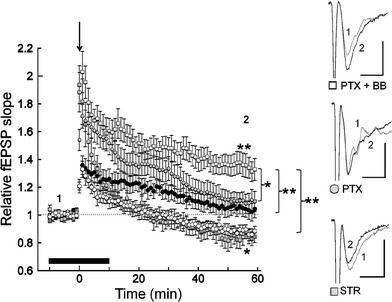
The effects of GABAa receptors and glycine receptors on MPP-DG synaptic plasticity. Picrotoxin (PTX) blocked the induction of the LTD observed under the control conditions (CTL). The potentiation magnitude observed under the picrotoxin conditions did not differ from that seen under the bilobalide (BB) conditions. HFS induced LTP under the picrotoxin with bilobalide (PTX + BB) conditions. HFS also induced LTD under the strychnine (STR) conditions. The black bar shows the timing of the application of picrotoxin, picrotoxin with bilobalide, or strychnine. The findings obtained under the control (CTL) conditions (open circles) and the bilobalide (BB) conditions (filled circles) were the same as those shown in Fig. 2b. Insets Typical waveforms of the synaptic responses observed before (1) and 60 min after (2) the HFS. Scales 0.5 mV, 5 ms. *p < 0.05, **p < 0.01
Discussion
In the present study, we compared the effects of bilobalide on synaptic transmission and plasticity among rat hippocampal subfields for the first time, and bilobalide was demonstrated to have facilitative effects on synaptic transmission and plasticity in hippocampal CA1 and MPP-DG synapses, respectively.
By recording input–output relationships, we found that bilobalide had facilitative effects on SC-CA1 synaptic transmission; i.e., that lower presynaptic activation caused larger post-synaptic activation in the presence of bilobalide than that seen under the control conditions, and this facilitative effect was only seen in CA1 synapses (Fig. 1a). Since bilobalide has been reported to increase neuronal activity by antagonizing GABAa receptors [13], its facilitative effect might be caused by the bilobalide-induced inhibition of GABAergic modulation. However, we cannot rule out the possibility that bilobalide has targets other than the GABAergic system because no facilitative effect on baseline responses was observed at other hippocampal subfields (Fig. 1b–d) despite the activation of GABA and glutamic acid decarboxylase (GAD) in all subfields [23]. The elucidation of the targets of bilobalide (it probably shows specificity for SC-CA1 synapses) is necessary to clarify the cellular mechanisms of its facilitative effects on baseline synaptic responses.
The chronic administration of EGb761® reportedly protects the brain against ischemia-induced cell death [11]. Since the synaptic responses of SC-CA1 synapses remained stable for more than 1 h under our experimental conditions, it is not plausible to suggest that the facilitative effects of bilobalide on SC-CA1 synaptic responses observed in this study (Fig. 1a) were due to the prevention of CA1 neuron death. However, for the purpose of memory improvement over a longer time scale, the in vivo application of bilobalide might be a useful anti-dementia treatment due to its ability to prevent the death of hippocampal CA1 neurons (i.e., to prevent cell death in one of the most vulnerable areas of the brain).
As for its effects on synaptic plasticity, it had no significant effect on synaptic plasticity at CA1 synapses (Fig. 2a) but did facilitate synaptic plasticity at MPP-DG synapses; i.e., the synaptic responses were rescued from LTD (Fig. 2b). The finding that bilobalide had inhibitory effects on the GABAergic system was expected because it was reported that GABAergic modulation of tetraethylammonium-induced synaptic plasticity was increased in DG compared with that observed in CA1 synapses [15]. The similarity between the effects of PTX and bilobalide on MPP-DG synaptic plasticity (Fig. 3) in our experimental conditions are consistent with the idea that bilobalide has inhibitory effects on the GABAergic system at MPP-DG synapses, although we cannot rule out the possibility that bilobalide also affects other target(s) because of the larger facilitative effects observed under the PTX plus bilobalide conditions than under the PTX or bilobalide alone conditions (Fig. 3). Bilobalide also blocks glycine receptors [22], but bilobalide-induced modulation of the glycinergic system does not seem to be involved in the facilitative effects of bilobalide because HFS-induced LTD at MPP-DG synapses was not rescued by the presence of strychnine (Fig. 3). Further studies are needed to identify all targets of bilobalide, which will hopefully lead to the elucidation of the mechanisms responsible for not only its facilitative effects on synaptic plasticity but also its population spike amplifying [13] and neuroprotective effects [24].
In the presence of GABAA receptor blockers (picrotoxin and bicuculline), LTP was reportedly induced at DG synapses [20, 25–28], and LTP was induced, even in the absence of picrotoxin [25], by HFS involving strong stimulation (10 impulses at 100 Hz, twice the test intensity) and a much higher extracellular Ca2+ concentration (4 mM CaCl2). In the present study, HFS (1 train of 100 Hz for 1 s) induced LTD in the absence of bilobalide at MPP-DG synapses, perhaps due to the intermediate amount of Ca2+ influx that occurred under our experimental conditions.
Generally, the effects of Gingko biloba on cognitive function have been investigated using a mixture of extracts including flavonoids and terpene trilactones (ginkgolide A, B, C, J, M, and bilobalide), and little information is available about the individual effects of each component. As for its effectiveness on inhibitory systems, bilobalide inhibits GABAA receptors more potently than ginkgolide B [22], and ginkgolide B, C, and M are significantly more potent than ginkgolide A and J for blocking glycine receptors [22]. Since it was recently reported that bilobalide was detected in the brain after the intravenous administration of bilobalide [29] and that the concentration of bilobalide was much higher than those of ginkgolides A and B after the oral administration of EGb761® [30], the facilitative effects of bilobalide on synaptic transmission and synaptic plasticity suggest that it is a candidate molecule that could be used to improve cognitive function in clinical situations.
Acknowledgment
This study was supported by the 2009 Sophia University-wide Collaborative Research Grant (to T.O. and T.U.).
Abbreviations
- ACSF
Artificial cerebrospinal fluid
- BB
Bilobalide
- DG
Dentate gyrus
- DMSO
Dimethyl sulfoxide
- ESI-MS
Electrospray ionization-mass spectrometry
- EtOAc
Ethyl acetate
- fEPSP
Field excitatory postsynaptic potentials
- LPP
Lateral perforant path
- LTD
Long-term depression
- LTP
Long-term potentiation
- MeOH
Methyl alcohol
- MPP
Medial perforant path
- NMR
Nuclear magnetic resonance
- PPD
Paired-pulse depression
- PPF
Paired-pulse facilitation
- PTX
Picrotoxin
- STR
Strychnine
- TLC
Thin-layer chromatography
References
- 1.Zola-Morgan S, Squire LR, Amaral DG. Human amnesia and the medial temporal region: enduring memory impairment following a bilateral lesion limited to field CA1 of the hippocampus. J Neurosci. 1986;6:2950–2967. doi: 10.1523/JNEUROSCI.06-10-02950.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Corkin S. What’s new with the amnesic patient H.M.? Nat Rev Neurosci. 2002;3:153–160. doi: 10.1038/nrn726. [DOI] [PubMed] [Google Scholar]
- 3.Bliss TVP, Collingridge GL. A synaptic model of memory: long-term potentiation in the hippocampus. Nature. 1993;361:31–39. doi: 10.1038/361031a0. [DOI] [PubMed] [Google Scholar]
- 4.Manabe T, Noda Y, Mamiya T, Katagiri H, Houtani T, Nishi M, Noda T, Takahashi T, Sugimoto T, Nabeshima T, Takeshima H. Facilitation of long-term potentiation and memory in mice lacking nociception receptors. Nature. 1998;394:577–581. doi: 10.1038/29073. [DOI] [PubMed] [Google Scholar]
- 5.Tang YP, Shimizu E, Dube GR, Rampon C, Kerchner GA, Zhuo M, Liu G, Tsien JZ. Genetic enhancement of learning and memory in mice. Nature. 1999;401:63–69. doi: 10.1038/43432. [DOI] [PubMed] [Google Scholar]
- 6.Whitlock JR, Heynen AJ, Shuler MG, Bear MF. Learning induces long-term potentiation in the hippocampus. Science. 2006;313:1093–1097. doi: 10.1126/science.1128134. [DOI] [PubMed] [Google Scholar]
- 7.Weinmann S, Roll S, Schwarzbach C, Vauth C, Willich SN. Effects of Ginkgo biloba in dementia: systematic review and meta-analysis. BMC Geriatr. 2010;10:14. doi: 10.1186/1471-2318-10-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Winter JC. The effects of an extract of Ginkgo biloba, EGb 761, on cognitive behavior and longevity in the rat. Physiol Behav. 1998;63:425–433. doi: 10.1016/S0031-9384(97)00464-2. [DOI] [PubMed] [Google Scholar]
- 9.Stoll S, Scheuer K, Pohl O, Müller WE. Ginkgo biloba extract (EGb 761) independently improves changes in passive avoidance learning and brain membrane fluidity in the aging mouse. Pharmacopsychiatry. 1996;29:144–149. doi: 10.1055/s-2007-979561. [DOI] [PubMed] [Google Scholar]
- 10.Williams B, Watanabe CMH, Schultz PG, Rimbach G, Krucker T. Age-related effects of Ginkgo biloba extract on synaptic plasticity and excitability. Neurobiol Aging. 2004;25:955–962. doi: 10.1016/j.neurobiolaging.2003.10.008. [DOI] [PubMed] [Google Scholar]
- 11.Chandrasekaran K, Mehrabian Z, Spinnewyn B, Drieu K, Fiskum G. Neuroprotective effects of bilobalide, a component of the Ginkgo biloba extract (EGb 761), in gerbil global brain ischemia. Brain Res. 2001;922:282–292. doi: 10.1016/S0006-8993(01)03188-2. [DOI] [PubMed] [Google Scholar]
- 12.Strømgaard K, Nakanishi K. Chemistry and biology of terpene trilactones from Ginkgo biloba . Angew Chem Int Ed. 2004;43:1640–1658. doi: 10.1002/anie.200300601. [DOI] [PubMed] [Google Scholar]
- 13.Sasaki K, Oota I, Wada K, Inomata K, Ohshika H, Haga M. Effects of bilobalide, a sesquiterpene in Ginkgo biloba leaves, on population spikes in rat hippocampal slices. Comp Biochem Physiol C Pharmacol Toxicol Endocrinol. 1999;124:315–321. doi: 10.1016/S0742-8413(99)00082-1. [DOI] [PubMed] [Google Scholar]
- 14.Kuria KAM, Chepkwony H, Govaerts C, Roets E, Busson R, de Witte P, Zupko I, Hoornaert G, Quirynen L, Maes L, Janssens L, Hoogmartens J, Laekeman G. The antiplasmodial activity of isolates from Ajuga remota . J Nat Prod. 2002;65:789–793. doi: 10.1021/np0104626. [DOI] [PubMed] [Google Scholar]
- 15.Suzuki E, Okada T. Regional differences in GABAergic modulation for TEA-induced synaptic plasticity in rat hippocampal CA1, CA3 and dentate gyrus. Neurosci Res. 2007;59:183–190. doi: 10.1016/j.neures.2007.06.1472. [DOI] [PubMed] [Google Scholar]
- 16.Toth K, Suares G, Lawrence JJ, Philips-Tansey E, McBain CJ. Differential mechanisms of transmission at three types of mossy fiber synapse. J Neurosci. 2000;20:8279–8289. doi: 10.1523/JNEUROSCI.20-22-08279.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nicoll RA, Schmitz D. Synaptic plasticity at hippocampal mossy fibre synapses. Nat Rev Neurosci. 2005;6:863–876. doi: 10.1038/nrn1786. [DOI] [PubMed] [Google Scholar]
- 18.Reeves TM, Kao CQ, Phillips LL, Bullock MR, Povlishock JT. Presynaptic excitability changes following traumatic brain injury in the rat. J Neurosci Res. 2000;60:370–379. doi: 10.1002/(SICI)1097-4547(20000501)60:3<370::AID-JNR12>3.0.CO;2-B. [DOI] [PubMed] [Google Scholar]
- 19.Winegar BD, MacIver MB. Isoflurane depresses hippocampal CA1 glutamate nerve terminals without inhibiting fiber volleys. BMC Neurosci. 2006;7:5. doi: 10.1186/1471-2202-7-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Colino A, Malenka RC. Mechanisms underlying induction of long-term potentiation in rat medial and lateral perforant paths in vitro. J Neurophysiol. 1993;69:1150–1159. doi: 10.1152/jn.1993.69.4.1150. [DOI] [PubMed] [Google Scholar]
- 21.Bramham CR, Milgram NW, Srebro B. Activation of AP5-sensitive NMDA receptors is not required to induce LTP of synaptic transmission in the lateral perforant path. Eur J Neurosci. 1991;3:1300–1308. doi: 10.1111/j.1460-9568.1991.tb00062.x. [DOI] [PubMed] [Google Scholar]
- 22.Ivic L, Sands TTJ, Fishkin N, Nakanishi K, Kriegstein AR, Strømgaard K. Terpene trilactones from Ginkgo biloba are antagonists of cortical glycine and GABAA receptors. J Biol Chem. 2003;278:49279–49285. doi: 10.1074/jbc.M304034200. [DOI] [PubMed] [Google Scholar]
- 23.Hörtnagl H, Berger ML, Sperk G, Pifl C. Regional heterogeneity in the distribution of neurotransmitter markers in the rat hippocampus. Neuroscience. 1991;45:261–272. doi: 10.1016/0306-4522(91)90224-C. [DOI] [PubMed] [Google Scholar]
- 24.Kiewert C, Kumar V, Hildmann O, Rueda M, Hartmann J, Naik RS, Klein J. Role of GABAergic antagonism in the neuroprotective effects of bilobalide. Brain Res. 2007;1128:70–78. doi: 10.1016/j.brainres.2006.10.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hanse E, Gustafsson B. Long-term potentiation and field EPSPs in the lateral and medial perforant paths in the dentate gyrus in vitro: a comparison. Eur J Neurosci. 1992;4:1191–1201. doi: 10.1111/j.1460-9568.1992.tb00144.x. [DOI] [PubMed] [Google Scholar]
- 26.Chen C, Magee JC, Marcheselli V, Hardy M, Bazan NG. Attenuated LTP in hippocampal dentate gyrus neurons of mice deficient in the PAF receptor. J Neurophysiol. 2001;85:384–390. doi: 10.1152/jn.2001.85.1.384. [DOI] [PubMed] [Google Scholar]
- 27.Murray HJ, O’Connor JJ. A role for monomeric G-proteins in synaptic plasticity in the rat dentate gyrus in vitro. Brain Res. 2004;1000:85–91. doi: 10.1016/j.brainres.2003.11.044. [DOI] [PubMed] [Google Scholar]
- 28.Wu J, Harney S, Rowan MJ, Anwyl R. Involvement of group I mGluRs in LTP induced by strong high frequency stimulation in the dentate gyrus in vitro. Neurosci Lett. 2008;436:235–238. doi: 10.1016/j.neulet.2008.03.027. [DOI] [PubMed] [Google Scholar]
- 29.Madgula VLM, Avula B, Yu YB, Wang YH, Tchantchou F, Fisher S, Luo Y, Khan IA, Khan SI. Intestinal and blood–brain barrier permeability of ginkgolides and bilobalide: in vitro and in vivo approaches. Planta Med. 2010;76:599–606. doi: 10.1055/s-0029-1240601. [DOI] [PubMed] [Google Scholar]
- 30.Ude C, Paulke A, Nöldner M, Schubert-Zsilavecz M, Wurglics M. Plasma and brain levels of terpene trilactones in rats after an oral single dose of standardized Ginkgo biloba extract EGb 761® . Planta Med. 2011;77:259–264. doi: 10.1055/s-0030-1250286. [DOI] [PubMed] [Google Scholar]