

Abstract
The hippocampus and associated structures are responsible for episodic memory in humans. In rodents, the most prominent behavioral correlate of hippocampal neural activity is place coding, which is thought to underlie spatial navigation. While episodic memory is considered to be unique to humans in a restricted context, it has been proposed that the same neural circuitry and algorithms that enable spatial coding and navigation also support episodic memory. Here we review the recent progress in neural circuit mechanisms of hippocampal activity by introducing several topics: (1) cooperation and specialization of the bilateral hippocampi, (2) the role of synaptic plasticity in gamma phase-locking of spikes and place cell formation, (3) impaired goal-related activity and oscillations in a mouse model of mental disorders, and (4) a prefrontal-thalamo-hippocampal circuit for goal-directed spatial navigation.
Keywords: Hippocampus, Place cells, Navigation, Oscillations, Left–right asymmetry, Synaptic plasticity
Introduction
The hippocampus is crucial for episodic memory in humans [1–5]. Episodic memory is claimed to be unique to humans, which allows an individual to mentally travel back in time to remember personal experiences in the context of both time and space [6–8]. There has been controversy over whether other species have a similar ability to memorize their personal episodes and whether their hippocampus mediates it [3, 4, 9–19]. In rodents, the most prominent behavioral correlate of hippocampal principal neuron activity is place coding [20]. Individual position-tuned cells (place cells) fire in particular locations in the environment [20]. As an ensemble, place cells provide a representation of space [21], which is thought to support spatial memory and navigation. Recently, it has been proposed that the mechanisms of declarative (episodic and semantic) memory have evolved from the mechanisms of spatial navigation [22–24].
Temporal coordination in the hippocampus is secured by various oscillations [25], especially theta [26–33], gamma [34], and sharp wave-ripple complexes (SPW-R) [35]. Consistent with anatomical, morphological, and molecular properties [36], accumulating evidence indicates that there are sub-region- and sub-layer-specific functional properties in the hippocampus. Hippocampal CA1 and CA3 areas have distinct activity dynamics, which may support distinct computational roles in the circuitry [29, 37–46]. Even within the same sub-region (e.g., CA1, CA2, CA3), principal neurons are not homogenous. Pyramidal neurons in the dorsal and ventral hippocampus show distinct neuronal representations and network dynamics [47–51]. CA1 pyramidal cells show a gradient in the proximal–distal (CA3 to subiculum) direction in terms of place coding [52] and propensity of emitting spike bursts [53]. In CA1 and CA2 areas, deep and superficial pyramidal cells have distinct connectivities and functions [54–63]. Further, numerous parameters (firing rate of individual neurons, magnitude of synchrony, probability of spike transmission, synaptic strength) in the same sub-regions in the hippocampus are hugely heterogeneous, and follow skewed distributions with a heavy tail [64–67], suggesting that the skewed (typically log-normal) distributions are fundamental to structural and functional organization in the circuitry [68].
Here we review recent progress in the neural circuit mechanisms of hippocampal activity by introducing several topics presented at the Annual Meeting of the Physiological Society of Japan in 2016 [69–72]. The topics cover a wide range of research areas; (1) cooperation and specialization of the bilateral hippocampi, (2) the role of synaptic plasticity in gamma phase-locking of spikes and place cell formation, (3) impaired goal-related activity and oscillations in a mouse model of mental disorders, and (4) a prefrontal-thalamo-hippocampal circuit for goal-directed spatial navigation.
Cooperation and specialization of the bilateral hippocampi in rodents
Left–right asymmetry of brain functions in mammals with higher cognitive abilities, such as primates, is a well-accepted concept [73]. A recent report has shown that dogs have functional brain laterality similar to primates [74]. The brain area in which asymmetry is most well studied in rodents is the hippocampus. The hippocampus is comprised of laminar structures in which projections from different brain areas terminate in distinct layers [75]. In the CA1, the pyramidal cell layer is sandwiched by the stratum oriens (str ori) and stratum radiatum (str rad). The former and the latter harbor the basal and the apical dendrite synapses of the CA1 pyramidal neurons, respectively. While both layers receive CA3 pyramidal axon projections, the str ori contain more synapses from the contralateral CA3 than the ipsilateral CA3. On the other hand, ipsilateral CA3–CA1 projections are more numerous in the str rad [76]. The CA1 areas have one more distal layer than the str rad, termed as the stratum lacunosum moleculare (str l-m), and here the CA1 pyramidal neurons receive direct inputs from the entorhinal cortex. The majority of entorhical-CA1 projections are ipsilateral [77]. However, the molecular/cellular basis of mammalian brain laterality is still largely unknown. Kawakami et al. reported that NR2B subunits of NMDA receptors are asymmetrically distributed between the left and right sides of the hippocampus [78]. The asymmetry was discovered by Kawakami and Ito using an NR2B receptor-specific antagonist. When excitatory postsynaptic currents (EPSCs) of ipsilateral projections from CA3–CA1 were compared between the left and right hippocampus, sensitivity to the NR2B receptor antagonist Ro 25-6981 was greater on the left side in the str rad. Interestingly, this asymmetry is the opposite in the str ori. The physiological data were then supported by quantitative analysis of postsynaptic proteins. After the commissural axons and their synapses were denervated by surgery, the synaptic NR2B proteins were more abundant on the left side in the str rad, and on the right side in the str ori. So, Shinohara proposed a hypothesis that the presynaptic side of hippocampal CA3–CA1 projection (i.e., CA3) might be a main determinant factor for synaptic properties.
This hypothesis was later confirmed by our anatomical paper [79]. Using GFP-expressing lentivirus as an anterograde tracer, Shinohara et al. showed that the synapses receiving innervation from the left CA3 are smaller and dominant in NR2B densities in the str rad. On the other hand, the synapses innervated by right CA3 pyramidal neurons are larger and exhibit higher densities of GluR1 subunits of AMPA receptors. As most of the other ionic glutamate receptors are expressed in higher numbers in wider synapses [80], the excitatory postsynaptic currents (EPSCs) evoked by right CA3 inputs are likely to be greater than those evoked by the left CA3. Given that ipsilateral innervations are slightly more numerous than contralateral in CA3–CA1 excitatory connections [79], the EPSCs in CA1 pyramidal neurons might be greater in right hippocampi than in the left one if the activities are comparable between the left and right CA3 pyramidal neurons.
One legitimate question for this asymmetry would be: How is this asymmetry formed—innately coded by the genetic programming or acquired by animals’ postnatal experience? The experiment using natural mutant inversus viscerum (iv) clearly demonstrated that the former is the case. iv has a point mutation in axonemal dynein in cilia during fetal development [81], and shows randomized laterality of the body axis. Regardless of the laterality (left-sided or right-sided) of the heart and other visceral organs, the mutant mice lacked the left–right asymmetry of the CA3–CA1 NR2B receptor appositions, which are observed in wild-type mice [78]. This result means that the genes that form hippocampal laterality are expressed downstream of left/right-dynein (lrd) [82]. As handedness of mice is reported to be 50–50 [83], the hippocampal CA3–CA1 asymmetry appears to be independent of mouse handedness.
Then, does postnatal experience have any effects on rodent hippocampal asymmetry? To answer this question, experience-dependent modulation of hippocampal local field potentials (LFPs) was investigated, since measurement of LFP is likely to be the most direct method to evaluate neural population dynamics. Bilateral LFPs were recorded simultaneously from dorsal CA1 areas of rats using multi-channel silicon probes (Fig. 1a) [84]. After recording, followed by calibration and compensation of the gains between the two probes, left vs. right CA1 LFPs were compared.
Fig. 1.
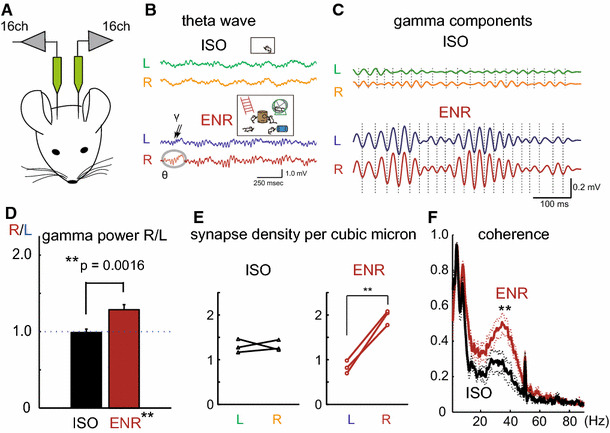
a Schematic drawing of the experiment [84]. Rats were anesthetized with urethane, and multichannel silicon probes were inserted into bilateral dorsal parts of CA1. b Representative left (L) and right (R) hippocampal LFPs recorded from the middle layer of CA1 str rad during theta periods. Upper traces and lower traces are from ISO and ENR rats, respectively. c Magnified view of theta-associated gamma oscillations. Gamma oscillations components are extracted. d Right–left ratios (R/L) of gamma powers averaged from individual ISO and ENR rats (N = 10 for both animals). e The synapse densities of CA1 pyramidal neuron spines of the middle layers of CA1 str rad observed by electron microscopy. Each dot indicates synapse densities in individual animals. f Interhemispheric coherence of bilateral LFPs in the str rad during theta oscillation periods
Hippocampal LFPs are largely classified into two states, theta states and non-theta states, with the former manifesting brain online states (during active exploration and REM sleep) and the latter representing offline states and appearing during slow-wave sleep, consummatory behavior, and quiet waking [85]. Here, we focused on the gamma oscillations, because the gamma oscillations are accompanied by the theta status, observed during animals’ attentive and vigilant period, and are believed to reflect synaptic inputs [86]. Shinohara et al. analyzed rats reared in an enriched environment (ENR) and an isolated condition (ISO) for 3–4 weeks immediately after weaning, respectively. They detected similar bilateral synchronized theta activities in both ISO and ENR rats (Fig. 1a, b) [84]. Strikingly, the powers of accompanying gamma oscillations were greatly enhanced in ENR (Fig. 1c). In the comparison between left and right gamma powers in individual rats, the right gamma powers were significantly greater than the left in most of the subjects (Fig. 1d), while the left–right gamma power ratios were balanced in ISO rats. This right-sided dominance of gamma powers is observed in a wide area of the CA1, but is greatest in the middle layer of the str rad. Consecutive anatomical analysis of these areas revealed that synapse densities, but not synapse size, are significantly larger on the right, and almost double that of the left side in ENR rats (Fig. 1e). In contrast, in ISO rats, the bilateral synapse densities are nearly comparable. Interestingly, the average synapse densities on the left are slightly smaller than those observed in ISO rats.
Next, they analyzed the temporal relationship of LFP between the left and right sides of CA1 areas. In addition to synchronized theta activities, the interhemispheric coherence of gamma oscillations was elevated in the CA1 str rad and str l-m in ENR-reared rats (Fig. 1f). Given the higher gamma powers on the right side under ENR rearing, environmental stimuli appear to promote bilateral hippocampal cooperation, and also facilitate functional specialization of left and right hippocampal hemispheres. Since enhanced gamma powers and interhemispheric coherence are both abolished by chronic administration of a low dose of ketamine (NMDA receptor antagonist), ENR-rearing effects appear to be mediated by NMDA receptor-dependent synaptic plasticity.
Here we have introduced several molecular/functional differences observed between the left and right hippocampus in rodents. With accumulating evidence of a left–right asymmetry in the rodent brain, more studies are needed to address whether the left and right hippocampus have distinct functions, and if bilateral cooperation in the hippocampus is needed for animals’ cognitive abilities.
The role of synaptic plasticity in slow gamma phase-locking and place cell formation in the hippocampal CA1 area during novel experience
Gamma oscillations, a type of neural oscillations present in many brain regions, including the hippocampal–entorhinal circuits, often regulate the timing of neuronal firing so that neurons fire at specific phases of ongoing gamma oscillations [34]. This phenomenon, phase-locking to gamma oscillations, is implicated in information transfer between neuronal populations in various cognitive functions such as learning and memory [87]. In the rodent hippocampal CA1 area, at least two types of gamma oscillations are present with distinct frequency ranges: slow gamma (25–50 Hz) and fast gamma (65–140 Hz) oscillations [88]. These two types of gamma oscillations are thought to be derived from structures synaptically upstream of the CA1, the CA3 area and the entorhinal cortex, respectively. The synchronization of gamma oscillations between the CA1 area and the upstream structures dynamically changes during memory-associated behavior [89–92]. Moreover, phase-locking of CA1 principal cells to these gamma oscillations is modulated through particular behaviors [90, 93]. Thus, the phase-locking to gamma oscillations may dynamically regulate information flow in hippocampal–entorhinal circuits.
Despite such a proposed key role, the molecular/cellular mechanisms establishing the gamma phase locking are still not well understood. As the balance of excitatory and inhibitory synaptic inputs is thought to be important for the gamma phase-locking [34, 94], synaptic plasticity may control the gamma phase-locking as a mechanism of modulating synaptic strength. To test this hypothesis, Kitanishi et al. blocked synaptic plasticity in CA1 pyramidal cells and monitored the firing of these neurons together with gamma oscillations in freely behaving rats [95]. They injected either an adeno-associated viral (AAV) vector expressing GFP-fused GluR1-c-tail or a control AAV into the dorsal CA1 area, and monitored the neuronal activity via chronically implanted tetrodes. GluR1-c-tail is a dominant-negative mutant that blocks long-term potentiation (LTP) at Shaffer-CA1 synapses by interfering with synaptic delivery of GluR1-containing AMPA receptors [95–97].
When rats explored a novel open field, the power of slow gamma oscillations increased in both GluR1-c-tail-expressing and control CA1 areas [95]. The increased power returned to baseline levels as the environment became familiar. This transient increase in slow gamma power suggests that novel experience strengthens synaptic input coming from the CA3 area at the gamma frequency range irrespective of the LTP blockade in the CA1 area. By contrast, GluR1-c-tail impairs phase-locking of CA1 principal cells to slow gamma, but not to fast gamma, oscillations in the novel open field [95]. The result suggests that GluR1-dependent synaptic plasticity facilitates entrainment of CA1 principal cells by slow gamma oscillations during novel experience. The intact fast gamma phase-locking is surprising because GluR1-c-tail distributes in both dendritic segments receiving CA3 inputs and entorhinal inputs. Dividing the fast gamma frequency range into two sub-bands [98] and analyzing entrainment by gamma oscillations recorded from distinct dendritic segments separately [99] may reveal effects on the phase-locking associated with entorhinal inputs.
How is the slow gamma phase-locking linked to place cell activity? Place cells are the hippocampal neurons that fire whenever animals traverse particular locations of an environment [20]. The spatially selective firing pattern of a place cell rapidly emerges when animals encounter a novel environment [100], and is stable for days or even months [101, 102]. This rapid formation and temporal stability may reflect the acquisition and storage of spatial information. When GluR1-c-tail is expressed in the CA1 area, the formation of fine place cell activity in a novel environment is slower, while repeated exposure to the same environment gradually sharpens the place cell activity to the level of controls [95]. Thus, GluR1-dependent synaptic plasticity required for slow gamma phase-locking also contributes to rapid formation of place cell activity during novel experience. Figure 2 summarizes a series of novelty-induced events and the role of synaptic plasticity: the strengthening of slow gamma oscillations originating from the CA3 area rapidly induces GluR1-dependent synaptic plasticity in CA3–CA1 synapses, which in turn establishes the phase-locking to slow gamma oscillations and the place cell activity as the CA1 output to the entorhinal cortex.
Fig. 2.
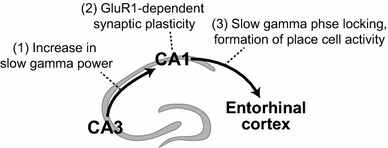
Schematic showing the proposed information flow during novel experience regulated by GluR1-dependent synaptic plasticity
Goal-related place cell activity and network dynamics are impaired in mouse models of mental disorders
The hippocampus is important for spatial memory, and place cell activity in the hippocampus is thought to support this function [3, 21]. However, how hippocampal neurons contribute to goal-directed navigation during spatial tasks is not yet clear. One possible idea is that the goal location is represented differently from other off-target areas by the neurons. Several experiments have tried to detect the effect of goal-directed behaviors on place fields: some studies reported skewed firing field distribution of the place cells in animals during goal-related tasks. Place fields were accumulated at the goal while rats seek a fixed reward zone [103–106] or swam in an annular water maze to find a hidden platform [107]. When the goal location was changed, this goal-related activity was reorganized to represent new goal locations [104, 106, 107]. In these experiments, animals seek a fixed, unmarked goal zone and were rewarded at the place. Using a slightly different task, Hok and colleagues found another effect of goal-directed behavior on place fields [108]. In their experiments, animals also seek a fixed, unmarked goal zone to release a food pellet, but were rewarded at an unfixed location, because the reward site depends on the bouncing of the pellet (Fig. 3a). They found that place cells fire at goal location in addition to their main place fields (Fig. 3b). Although there are differences in the goal-related activity among experiments, these results suggest that the hippocampus overrepresents behaviorally significant regions in space. This reward-related activity predicted task performance [62, 104], indicating a functional link between the activity at the goal zone and the animal’s spatial memory performance. Noteworthy, Dupret et al. reported that accumulation of place fields was observed when goals were unmarked and the animal needed to remember the reward locations, but such reorganization of place fields did not occur when goals were visually guided [104]. The result indicates that such accumulation of place fields occurs when animals use a cognitive map to locate hidden rewards [104]. This may be the reason why some previous studies failed to see place field accumulation at the goal while searching for rewards [109–112].
Fig. 3.
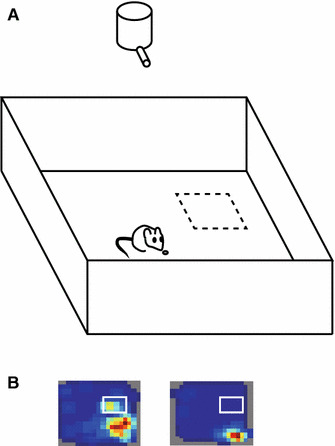
Goal-related activity in the hippocampus. a The testing box for the place preference task. Animals were required to stay at least 1 s in the unmarked goal zone (dashed square), after which time a food pellet was dropped from the overhead dispenser. b Example place field map with putative goal-zone activity (left) and without goal-zone activity (right)
Although place cell activity was found in all regions of the hippocampal trisynaptic circuit, CA1, CA3, and the dentate gyrus, the place field accumulation at the goal was observed in the CA1, but not in the CA3 [104]. Moreover, within the CA1, representation of the reward zone by deep, but not superficial, pyramidal cells predicts the performance of a goal-oriented learning task [62]. These results suggest functional specialization within the hippocampus for spatial navigation; while the CA3 provides task-invariant representation of space, the CA1 place representations are adapted to task requirements.
The origin of the goal-related activity in the CA1 is not yet known. The medial prefrontal cortex (mPFC), known as a site of action evaluation and selection [113], is a candidate for the source of goal-related information. However, inhibition of the mPFC does not abolish goal-related activity in the CA1 [114], indicating that the mPFC is not essential for goal-related firing patterns in the CA1. Another possible source of the goal-related activity is dopaminergic input, which conveys a signal of reward expectation [115]. The hippocampus receives input from two major sources of dopamine signals, the substantia nigra and the ventral tegmental area [116], and these dopaminergic inputs have been shown to enhance long-term potentiation in the hippocampus, thereby facilitating learning [117, 118].
Learning impairment is a key clinical feature of several mental disorders, and several mouse models of mental illnesses have been reported to exhibit learning abnormalities [119–121]. As the hippocampus plays a key role in spatial navigation and several forms of learning [24], it would be intriguing to investigate the activity of the hippocampus during spatial tasks in an animal model of mental disorders. Several studies have reported impairment of hippocampal activity in mouse models of mental disorders. Disrupted-in-Schizophrenia 1 (DISC1) is a strong susceptibility gene for schizophrenia and other mental illnesses [122–124]. Hayashi et al. used a place preference task (Fig. 3a) [108, 125] and found a performance deficit in dominant-negative DISC1 mutant mice (DISC1-DN-Tg-PrP mice [126]) in the task and impaired goal-related activity in the hippocampus [121]. Abnormalities in dopaminergic neurons in the DISC1 mutant mice [126] may underlie both behavioral and neurophysiological deficits in the mice.
In addition to neuronal representation, network oscillations, which are remarkably preserved in mammalian evolution [127], can contribute to neural information processing. It has been reported that theta band and SPW-R oscillations are also impaired in mouse models of mental disorders. SPW-Rs, which arise from the excitatory recurrent system of the CA3 region during consummatory behaviors and non-REM sleep, represent a synchronous population pattern in the CA3–CA1-subicular complex and entorhinal cortex [35]. The spike content of SPW-Rs is temporally coordinated to replay fragments of waking neuronal sequences in a compressed manner [35]. It has been suggested that SPW-Rs assist in transferring the hippocampal compressed representation to distributed circuits to support memory consolidation [35, 85, 128, 129]. Suh et al. found increased occurrence and intensity of SPW-Rs and abnormality in SPW-R-associated replay events in Calcineurin knockout mice [130], which have been shown to exhibit behavioral and cognitive abnormalities [131]. Similar SPW-R abnormalities are found in transgenic mice expressing DISC1-DN under a CaMKIIalpha promoter [132, 133]. Long-range synchrony of neuronal activity by network oscillations may facilitate effective communication between distributed brain regions [134]. Df(16)A1 ± mice, which model a microdeletion on human chromosome 22 (22q11.2) that has been established as a genetic risk factor for schizophrenia, showed impaired spatial working memory task performance and reduced hippocampal–prefrontal synchrony by theta oscillations [120]. Not only behavioral and structural phenotyping but also neurophysiological characterization of animal models would be a great help in understanding the pathophysiology of mental disorders.
A prefrontal-thalamo-hippocampal circuit for goal-directed spatial navigation
The ability to explore geometric space is essential for the survival of animals. Virtually all animals, from insects to mammals, are able to come back home after exploration of the environment, which is necessary for stable settling. Each species of animals thus developed its own strategy for navigation by making use of various cues available in the environment as well as by monitoring their own movements. For instance, desert ants are known to have the ability to estimate the direction and distance of the nest location after a long exploration. A number of studies suggest that this ability is primarily based on a navigation strategy called path integration, in which the brain computes a summation of its own movements during exploration to keep track of the relationship between the animal’s position and the nest location [135, 136]. On the other hand, sea turtles or pigeons have an ability to return home after thousands of kilometers of navigation, which is thought to be achieved by their special sense for the earth’s magnetic field [137–139].
Rats are one of such animals that have excellent navigation ability. Many lines of evidence suggest that rats are able to use positional relationships of the environment, or a map, for navigation as humans do [21, 140]. For instance, rats can find a novel shortcut to the destination [140], or are able to correctly estimate the goal location even when navigation is started from different positions [141, 142]. This idea was further enforced by the discovery of place cells in the rat hippocampus that fire selectively at a particular location in the environment [20].
It is, however, still largely unclear how place cells can be used for goal-directed navigation. This is because, while place cells may be useful for the animal to identify its instantaneous position, these cells do not fire once the animal leaves the firing fields of the cells, which raises the question of how these cells can be used to plan a route to a destination that is somewhere distant from the animal’s position. Several studies have provided a clue to this problem. It was discovered that the activity of place cells not only represents the animal’s current position but is also influenced by the animal’s trajectories during navigation. For example, the firing rates, but not the firing locations, of place cells change depending on the animal’s past or future trajectory [143–145], and brief sequences of place-cell activity reflect the animal’s next movement direction [146, 147].
If the activity of place cells reflects the animal’s intended route in navigation, how can these cells receive such information about action plans? A key experiment was performed by Ito and colleagues [148]. They recorded the activity of neurons while the animal performed an alternation task in a modified T-maze. The study showed that the trajectory-dependent rate change in place cells is derived from activity in the mPFC, a crucial brain area for action planning and decision-making [149–151]. Although there is essentially no major anatomical connection from mPFC to the hippocampus, this information transfer is mediated by the thalamic nucleus reuniens (NR) that receives inputs from mPFC and gives rise to inputs in area CA1 of the hippocampus [152–154] (Fig. 4). Consistent with the anatomical projections of NR neurons that do not make synapses with CA3 neurons, the trajectory-dependent rate change is significantly smaller in place cells in CA3 compared with those in CA1. Ito et al. further demonstrated that neurotoxic lesions or optogenetic silencing of NR neurons resulted in a significant reduction of trajectory-dependent rate change in CA1 place cells. Finally, the analysis of error trials on the T-maze alternation task revealed that trajectory-dependent rate change in the mPFC-NR-CA1 circuit is crucial to represent the animal’s intended routes rather than past trajectories, indicating a key role for the circuit in route planning.
Fig. 4.
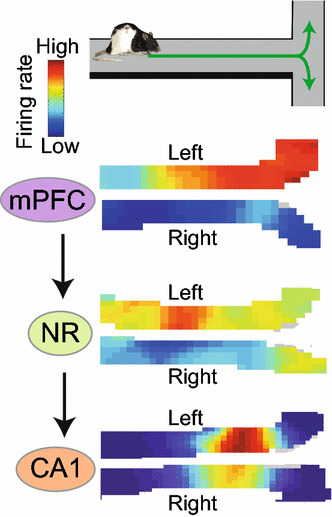
When the animal makes a choice between right- and left-oriented trajectories on the central stem of a T-maze, place cells in the hippocampal CA1 change their firing rates, but not firing location, depending on the next route. This trajectory information is provided by the medial prefrontal cortex (mPFC) through the thalamic nucleus reuniens (NR). Note that neurons in mPFC or NR do not exhibit location-selective firing as place cells do, but their firing rates change depending on the future trajectory
It is thus now clear that the hippocampus is a part of the action planning system, including prefrontal brain regions. The study by Ito et al. is, however, still the beginning of research toward the understanding of navigation circuits. For instance, while several studies have reported activity change in mPFC and the striatum around the goal location [155, 156], how can such information be used to estimate a direction and distance of the destination during the route planning process? Decades of research on the hippocampus and associated parahippocampal regions have clarified the spatial representation system in the brain [157]. A next key step will be to understand how the brain makes use of such spatial representations for action planning. Further investigation of interactions between the hippocampus and prefrontal brain regions will provide new insights into the relationship between the brain’s spatial representation system and behaviors.
Conclusions
The Nobel Prize in Physiology or Medicine 2014 was awarded to John O’Keefe, May-Britt Moser, and Edward I. Moser “for their discoveries of cells that constitute a position system in the brain”. Given the award, one might think that the most important discovery in the hippocampal formation has already been made, and it would be better to do research in other brain regions to discover something new. This sober idea can be true. However, we believe that the hippocampus is still an excellent, probably one of the best, brain region to investigate for discovering the fundamental principles of the brain. Thanks to the wealth of the accumulated knowledge from anatomical, molecular biological, physiological, pharmacological, and behavioral studies, it might be more feasible to untangle the complicated relationships between molecules, structure, plasticity, network oscillations, neuronal representation, and behaviors in the hippocampus than in other brain regions. In this review paper, we introduced examples of such efforts in studying (1) molecular mechanisms and experience-dependent development of the left–right asymmetry of brain functions, (2) the relationship between synaptic plasticity, local field potential, and neuronal representation, (3) the pathophysiology of mental disorders, and (4) neural circuit mechanisms of goal-directed navigation. Brief summaries of each respective section are as follows:
Here, we review recent papers demonstrating the distinct circuitry/function of left–right hippocampal hemispheres in rodent CA3–CA1 projections, and discuss that both genetic and postnatal factors play an important roles in the laterality. The CA1 pyramidal neurons innervated by the right CA3 have larger excitatory synapses and contain more glutamate receptors, implying a greater likelihood of CA1 pyramidal neuron excitation by the right CA3 than by the left CA3. Moreover, given that ipsilateral CA3–CA1 inputs are more numerous than contralateral innervations, the right CA1 synapses in the str rad tend to be more strongly excited than those of the left CA1 if the activities of the CA3 are comparable between the left and the right side. In addition, through environmental stimulation during adolescence, the powers of gamma oscillations in the CA1 become greater on the right side.
Synaptic plasticity in the CA1 area mediates two dynamic changes in neuronal firing during novel experiences, (1) phase-locking to slow gamma oscillations, and (2) rapid formation of place cell activity. These functions of synaptic plasticity may in turn enhance the information flow through the CA3–CA1–entorhinal pathway.
During spatial tasks, hippocampal CA1 cells represent goal locations in a unique way, which may support goal-directed navigation. This characteristic goal representation is impaired in DISC1 mutant mice. Furthermore, several mouse models of mental disorders exhibit impaired sharp-wave ripples and theta-mediated hippocampal-prefrontal synchrony. These impairments may underlie the cognitive/learning impairments in mental disorders.
While decades of neuroscience research have identified major brain circuits that represent the animal’s instantaneous position, such information alone is not sufficient for goal-directed navigation. The interactions between the prefrontal action planning system and the hippocampus allow hippocampal place cells to represent the animal’s intended movements on top of the information about its current position, likely a key mechanism for the brain to estimate the animal’s next position in the environment for route planning.
While all of these studies were performed in rodents, this is not simply due to available experimental tools. Accumulating evidence suggests that the hippocampus of humans and rodents show significant functional similarity. For example, left–right asymmetry of hippocampal function has been reported in both humans and rodents [73, 158, 159]. Theta oscillations, gamma oscillations, and SPW-Rs in the hippocampus are preserved between humans and rodents [127]. Similar to the hippocampus of rodents, the human hippocampus has place cells and is involved in spatial navigation [159, 160]. Collectively, such prominent similarity suggests that the reviewed studies here will help us to understand the fundamental network mechanisms of the human hippocampus and our brain.
Acknowledgements
We thank Dr. Tom Macpherson for critical reading of the manuscript. TK is supported by grants from Takeda Science Foundation, the Sumitomo Foundation, SEI Group CSR Foundation, Narishige Neuroscience Research Foundation. HTI is supported by the Max Planck Society, the Behrens-Weise Foundation, JST PRESTO program. YS is supported by RIKEN intramural fund, RIKEN Special Postdoctoral Researchers Program, JSPS KAKENHI (23590273, 26282222). KM is supported by JSPS KAKENHI (16H04656, 16H01279, 15H06549), Daiichi Sankyo Foundation of Life Science, the Naito Foundation, Mochida Memorial Foundation for Medical and Pharmaceutical Research, Astellas Foundation for Research on Metabolic Disorders, SEI Group CSR Foundation, the Asahi Glass Foundation, Hokuto Foundation for Bioscience, Toray Science Foundation, Life Science Foundation of Japan, Shimadzu Science Foundation, the Tokyo Biochemical Research Foundation, Brain Science Foundation, the Ichiro Kanehara Foundation for the Promotion of Medical Science & Medical Care. TH is supported by JSPS KAKENHI (16H06568, 16K14579, 15H04275), Daiichi Sankyo Foundation of Life Science, Takeda Science Foundation, Smoking Research Foundation, the Kato Memorial Trust for Nambyo Research.
Compliance with ethical standards
Conflict of interest
The authors declare that they have no conflicts of interest.
Footnotes
T. Kitanishi, H. T. Ito, Y. Hayashi, Y. Shinohara, K. Mizuseki and T. Hikida contributed equally to this review.
Contributor Information
Kenji Mizuseki, Phone: +81-6-6645-3715, Email: mizuseki.kenji@med.osaka-cu.ac.jp.
Takatoshi Hikida, Phone: +81-75-366-7458, Email: hikida@tk.med.kyoto-u.ac.jp.
References
- 1.Scoville WB, Milner B. Loss of recent memory after bilateral hippocampal lesions. J Neurol Neurosurg Psychiatry. 1957;20:11–21. doi: 10.1136/jnnp.20.1.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Vargha-Khadem F, Gadian DG, Watkins KE, Connelly A, Van PW, Mishkin M. Differential effects of early hippocampal pathology on episodic and semantic memory. Science. 1997;277:376–380. doi: 10.1126/science.277.5324.376. [DOI] [PubMed] [Google Scholar]
- 3.Squire LR. Memory and the hippocampus: a synthesis from findings with rats, monkeys, and humans. Psychol Rev. 1992;99:195–231. doi: 10.1037/0033-295X.99.2.195. [DOI] [PubMed] [Google Scholar]
- 4.Cohen NJ, Eichenbaum H. Memory, amnesia, and the hippocampal system. Cambridge: MIT Press; 1993. [Google Scholar]
- 5.Milner B, Squire LR, Kandel ER. Cognitive neuroscience and the study of memory. Neuron. 1998;20:445–468. doi: 10.1016/S0896-6273(00)80987-3. [DOI] [PubMed] [Google Scholar]
- 6.Tulving E. Episodic and semantic memory. In: Tulving E, Donaldson W, editors. Organization of memory. New York: Academic Press; 1972. [Google Scholar]
- 7.Tulving E. Multiple memory systems and consciousness. Hum Neurobiol. 1987;6:67–80. [PubMed] [Google Scholar]
- 8.Tulving E. Episodic memory: from mind to brain. Annu Rev Psychol. 2002;53:1–25. doi: 10.1146/annurev.psych.53.100901.135114. [DOI] [PubMed] [Google Scholar]
- 9.Babb SJ, Crystal JD. Episodic-like memory in the rat. Curr Biol. 2006;16:1317–1321. doi: 10.1016/j.cub.2006.05.025. [DOI] [PubMed] [Google Scholar]
- 10.Clayton NS, Bussey TJ, Dickinson A. Can animals recall the past and plan for the future? Nat Rev Neurosci. 2003;4:685–691. doi: 10.1038/nrn1180. [DOI] [PubMed] [Google Scholar]
- 11.Clayton NS, Dickinson A. Episodic-like memory during cache recovery by scrub jays. Nature. 1998;395:272–274. doi: 10.1038/26216. [DOI] [PubMed] [Google Scholar]
- 12.Crystal JD. Episodic-like memory in animals. Behav Brain Res. 2010;215:235–243. doi: 10.1016/j.bbr.2010.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Eichenbaum H, Cohen NJ. Can we reconcile the declarative memory and spatial navigation views on hippocampal function? Neuron. 2014;83:764–770. doi: 10.1016/j.neuron.2014.07.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Eichenbaum H, Dudchenko P, Wood E, Shapiro M, Tanila H. The hippocampus, memory, and place cells: is it spatial memory or a memory space? Neuron. 1999;23:209–226. doi: 10.1016/S0896-6273(00)80773-4. [DOI] [PubMed] [Google Scholar]
- 15.Ergorul C, Eichenbaum H. The hippocampus and memory for “what,” “where,” and “when”. Learn Mem. 2004;11:397–405. doi: 10.1101/lm.73304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hoffman ML, Beran MJ, Washburn DA. Memory for “what”, “where”, and “when” information in rhesus monkeys (Macaca mulatta) J Exp Psychol Anim Behav Process. 2009;35:143–152. doi: 10.1037/a0013295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Allen TA, Fortin NJ. The evolution of episodic memory. Proc Natl Acad Sci USA. 2013;110(Suppl 2):10379–10386. doi: 10.1073/pnas.1301199110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Wood ER, Dudchenko PA, Eichenbaum H. The global record of memory in hippocampal neuronal activity. Nature. 1999;397:613–616. doi: 10.1038/16564. [DOI] [PubMed] [Google Scholar]
- 19.Roberts WA, Feeney MC, Macpherson K, Petter M, McMillan N, Musolino E. Episodic-like memory in rats: is it based on when or how long ago? Science. 2008;320:113–115. doi: 10.1126/science.1152709. [DOI] [PubMed] [Google Scholar]
- 20.O’Keefe J, Dostrovsky J. The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Res. 1971;34:171–175. doi: 10.1016/0006-8993(71)90358-1. [DOI] [PubMed] [Google Scholar]
- 21.O’Keefe J, Nadel L. The hippocampus as a cognitive map. Oxford: Clarendon Press; 1978. [Google Scholar]
- 22.Buzsáki G. Theta rhythm of navigation: link between path integration and landmark navigation, episodic and semantic memory. Hippocampus. 2005;15:827–840. doi: 10.1002/hipo.20113. [DOI] [PubMed] [Google Scholar]
- 23.Pastalkova E, Itskov V, Amarasingham A, Buzsáki G. Internally generated cell assembly sequences in the rat hippocampus. Science. 2008;321:1322–1327. doi: 10.1126/science.1159775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Buzsáki G, Moser EI. Memory, navigation and theta rhythm in the hippocampal–entorhinal system. Nat Neurosci. 2013;16:130–138. doi: 10.1038/nn.3304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Buzsáki G. Rhythms of the Brain. New York: Oxford University Press; 2006. [Google Scholar]
- 26.Buzsáki G. Theta oscillations in the hippocampus. Neuron. 2002;33:325–340. doi: 10.1016/S0896-6273(02)00586-X. [DOI] [PubMed] [Google Scholar]
- 27.O’Keefe J, Recce ML. Phase relationship between hippocampal place units and the EEG theta rhythm. Hippocampus. 1993;3:317–330. doi: 10.1002/hipo.450030307. [DOI] [PubMed] [Google Scholar]
- 28.Skaggs WE, McNaughton BL, Wilson MA, Barnes CA. Theta phase precession in hippocampal neuronal populations and the compression of temporal sequences. Hippocampus. 1996;6:149–172. doi: 10.1002/(SICI)1098-1063(1996)6:2<149::AID-HIPO6>3.0.CO;2-K. [DOI] [PubMed] [Google Scholar]
- 29.Dragoi G, Buzsáki G. Temporal encoding of place sequences by hippocampal cell assemblies. Neuron. 2006;50:145–157. doi: 10.1016/j.neuron.2006.02.023. [DOI] [PubMed] [Google Scholar]
- 30.Mizuseki K, Sirota A, Pastalkova E, Buzsáki G. Theta oscillations provide temporal windows for local circuit computation in the entorhinal–hippocampal loop. Neuron. 2009;64:267–280. doi: 10.1016/j.neuron.2009.08.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mizuseki K, Buzsáki G. Theta oscillations decrease spike synchrony in the hippocampus and entorhinal cortex. Philos Trans R Soc Lond B Biol Sci. 2014;369:20120530. doi: 10.1098/rstb.2012.0530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Wang Y, Romani S, Lustig B, Leonardo A, Pastalkova E. Theta sequences are essential for internally generated hippocampal firing fields. Nat Neurosci. 2015;18:282–288. doi: 10.1038/nn.3904. [DOI] [PubMed] [Google Scholar]
- 33.Stark E, Eichler R, Roux L, Fujisawa S, Rotstein HG, Buzsáki G. Inhibition-induced theta resonance in cortical circuits. Neuron. 2013;80:1263–1276. doi: 10.1016/j.neuron.2013.09.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Buzsáki G, Wang XJ. Mechanisms of gamma oscillations. Annu Rev Neurosci. 2012;35:203–225. doi: 10.1146/annurev-neuro-062111-150444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Buzsáki G. Hippocampal sharp wave-ripple: a cognitive biomarker for episodic memory and planning. Hippocampus. 2015;25:1073–1188. doi: 10.1002/hipo.22488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Cembrowski MS, Bachman JL, Wang L, Sugino K, Shields BC, Spruston N. Spatial gene-expression gradients underlie prominent heterogeneity of CA1 pyramidal neurons. Neuron. 2016;89:351–368. doi: 10.1016/j.neuron.2015.12.013. [DOI] [PubMed] [Google Scholar]
- 37.Mizuseki K, Royer S, Diba K, Buzsáki G. Activity dynamics and behavioral correlates of CA3 and CA1 hippocampal pyramidal neurons. Hippocampus. 2012;22:1659–1680. doi: 10.1002/hipo.22002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Barnes CA, McNaughton BL, Mizumori SJ, Leonard BW, Lin LH. Comparison of spatial and temporal characteristics of neuronal activity in sequential stages of hippocampal processing. Prog Brain Res. 1990;83:287–300. doi: 10.1016/S0079-6123(08)61257-1. [DOI] [PubMed] [Google Scholar]
- 39.Lee I, Yoganarasimha D, Rao G, Knierim JJ. Comparison of population coherence of place cells in hippocampal subfields CA1 and CA3. Nature. 2004;430:456–459. doi: 10.1038/nature02739. [DOI] [PubMed] [Google Scholar]
- 40.Lee I, Rao G, Knierim JJ. A double dissociation between hippocampal subfields: differential time course of CA3 and CA1 place cells for processing changed environments. Neuron. 2004;42:803–815. doi: 10.1016/j.neuron.2004.05.010. [DOI] [PubMed] [Google Scholar]
- 41.Leutgeb S, Leutgeb JK, Treves A, Moser MB, Moser EI. Distinct ensemble codes in hippocampal areas CA3 and CA1. Science. 2004;305:1295–1298. doi: 10.1126/science.1100265. [DOI] [PubMed] [Google Scholar]
- 42.Leutgeb JK, Leutgeb S, Treves A, Meyer R, Barnes CA, McNaughton BL, Moser MB, Moser EI. Progressive transformation of hippocampal neuronal representations in “morphed” environments. Neuron. 2005;48:345–358. doi: 10.1016/j.neuron.2005.09.007. [DOI] [PubMed] [Google Scholar]
- 43.Vazdarjanova A, Guzowski JF. Differences in hippocampal neuronal population responses to modifications of an environmental context: evidence for distinct, yet complementary, functions of CA3 and CA1 ensembles. J Neurosci. 2004;24:6489–6496. doi: 10.1523/JNEUROSCI.0350-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Lee I, Knierim JJ. The relationship between the field-shifting phenomenon and representational coherence of place cells in CA1 and CA3 in a cue-altered environment. Learn Mem. 2007;14:807–815. doi: 10.1101/lm.706207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Alvernhe A, Van CT, Save E, Poucet B. Different CA1 and CA3 representations of novel routes in a shortcut situation. J Neurosci. 2008;28:7324–7333. doi: 10.1523/JNEUROSCI.1909-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Leutgeb S, Leutgeb JK. Pattern separation, pattern completion, and new neuronal codes within a continuous CA3 map. Learn Mem. 2007;14:745–757. doi: 10.1101/lm.703907. [DOI] [PubMed] [Google Scholar]
- 47.Kjelstrup KB, Solstad T, Brun VH, Hafting T, Leutgeb S, Witter MP, Moser EI, Moser MB. Finite scale of spatial representation in the hippocampus. Science. 2008;321:140–143. doi: 10.1126/science.1157086. [DOI] [PubMed] [Google Scholar]
- 48.Royer S, Sirota A, Patel J, Buzsáki G. Distinct representations and theta dynamics in dorsal and ventral hippocampus. J Neurosci. 2010;30:1777–1787. doi: 10.1523/JNEUROSCI.4681-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Patel J, Fujisawa S, Berenyi A, Royer S, Buzsáki G. Traveling theta waves along the entire septotemporal axis of the hippocampus. Neuron. 2012;75:410–417. doi: 10.1016/j.neuron.2012.07.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Patel J, Schomburg EW, Berenyi A, Fujisawa S, Buzsáki G. Local generation and propagation of ripples along the septotemporal axis of the hippocampus. J Neurosci. 2013;33:17029–17041. doi: 10.1523/JNEUROSCI.2036-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Strange BA, Witter MP, Lein ES, Moser EI. Functional organization of the hippocampal longitudinal axis. Nat Rev Neurosci. 2014;15:655–669. doi: 10.1038/nrn3785. [DOI] [PubMed] [Google Scholar]
- 52.Henriksen EJ, Colgin LL, Barnes CA, Witter MP, Moser MB, Moser EI. Spatial representation along the proximodistal axis of CA1. Neuron. 2010;68:127–137. doi: 10.1016/j.neuron.2010.08.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Jarsky T, Mady R, Kennedy B, Spruston N. Distribution of bursting neurons in the CA1 region and the subiculum of the rat hippocampus. J Comp Neurol. 2008;506:535–547. doi: 10.1002/cne.21564. [DOI] [PubMed] [Google Scholar]
- 54.Deguchi Y, Donato F, Galimberti I, Cabuy E, Caroni P. Temporally matched subpopulations of selectively interconnected principal neurons in the hippocampus. Nat Neurosci. 2011;14:495–504. doi: 10.1038/nn.2768. [DOI] [PubMed] [Google Scholar]
- 55.Mizuseki K, Diba K, Pastalkova E, Buzsáki G. Hippocampal CA1 pyramidal cells form functionally distinct sublayers. Nat Neurosci. 2011;14:1174–1181. doi: 10.1038/nn.2894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Slomianka L, Amrein I, Knuesel I, Sorensen JC, Wolfer DP. Hippocampal pyramidal cells: the reemergence of cortical lamination. Brain Struct Funct. 2011;216:301–317. doi: 10.1007/s00429-011-0322-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Kohara K, Pignatelli M, Rivest AJ, Jung HY, Kitamura T, Suh J, Frank D, Kajikawa K, Mise N, Obata Y, Wickersham IR, Tonegawa S. Cell type-specific genetic and optogenetic tools reveal hippocampal CA2 circuits. Nat Neurosci. 2014;17:269–279. doi: 10.1038/nn.3614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Lee SH, Marchionni I, Bezaire M, Varga C, Danielson N, Lovett-Barron M, Losonczy A, Soltesz I. Parvalbumin-positive basket cells differentiate among hippocampal pyramidal cells. Neuron. 2014;82:1129–1144. doi: 10.1016/j.neuron.2014.03.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Valero M, Cid E, Averkin RG, Aguilar J, Sanchez-Aguilera A, Viney TJ, Gomez-Dominguez D, Bellistri E, de la Prida LM. Determinants of different deep and superficial CA1 pyramidal cell dynamics during sharp-wave ripples. Nat Neurosci. 2015;18:1281–1290. doi: 10.1038/nn.4074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Oliva A, Fernández-Ruiz A, Buzsáki G, Berényi A. Role of hippocampal CA2 region in triggering sharp-wave ripples. Neuron. 2016 doi: 10.1016/j.neuron.2016.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Maroso M, Szabo GG, Kim HK, Alexander A, Bui AD, Lee SH, Lutz B, Soltesz I. Cannabinoid control of learning and memory through HCN channels. Neuron. 2016;89:1059–1073. doi: 10.1016/j.neuron.2016.01.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Danielson NB, Zaremba JD, Kaifosh P, Bowler J, Ladow M, Losonczy A. Sublayer-specific coding dynamics during spatial navigation and learning in hippocampal area CA1. Neuron. 2016;91:652–665. doi: 10.1016/j.neuron.2016.06.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Stark E, Roux L, Eichler R, Senzai Y, Royer S, Buzsáki G. Pyramidal cell–interneuron interactions underlie hippocampal ripple oscillations. Neuron. 2014;83:467–480. doi: 10.1016/j.neuron.2014.06.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Mizuseki K, Buzsáki G. Preconfigured, skewed distribution of firing rates in the hippocampus and entorhinal cortex. Cell Rep. 2013;4:1010–1021. doi: 10.1016/j.celrep.2013.07.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Sayer RJ, Friedlander MJ, Redman SJ. The time course and amplitude of EPSPs evoked at synapses between pairs of CA3/CA1 neurons in the hippocampal slice. J Neurosci. 1990;10:826–836. doi: 10.1523/JNEUROSCI.10-03-00826.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Ikegaya Y, Sasaki T, Ishikawa D, Honma N, Tao K, Takahashi N, Minamisawa G, Ujita S, Matsuki N. Interpyramid spike transmission stabilizes the sparseness of recurrent network activity. Cereb Cortex. 2013;23:293–304. doi: 10.1093/cercor/bhs006. [DOI] [PubMed] [Google Scholar]
- 67.Omura Y, Carvalho MM, Inokuchi K, Fukai T. A lognormal recurrent network model for burst generation during hippocampal sharp waves. J Neurosci. 2015;35:14585–14601. doi: 10.1523/JNEUROSCI.4944-14.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Buzsáki G, Mizuseki K. The log-dynamic brain: how skewed distributions affect network operations. Nat Rev Neurosci. 2014;15:264–278. doi: 10.1038/nrn3687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Shinohara Y. Cooperation and specialization of the bilateral hippocampi in rodents. J Physiol Sci. 2016;66:S45. [Google Scholar]
- 70.Kitanishi T. Synaptic plasticity controls hippocampal circuit activity. J Physiol Sci. 2016;66:S45. [Google Scholar]
- 71.Hayashi Y, Sawa A, Hikida T. Impaired hippocampal activity at the goal zone on the place preference task in a DISC1 mouse model. J Physiol Sci. 2016;66:S45. doi: 10.1016/j.neures.2015.10.005. [DOI] [PubMed] [Google Scholar]
- 72.Ito H. A prefrontal-thalamo-hippocampal circuit for goal-directed spatial navigation. J Physiol Sci. 2016;66:S45. doi: 10.1007/s12576-016-0442-7. [DOI] [PubMed] [Google Scholar]
- 73.Toga AW, Thompson PM. Mapping brain asymmetry. Nat Rev Neurosci. 2003;4:37–48. doi: 10.1038/nrn1009. [DOI] [PubMed] [Google Scholar]
- 74.Andics A, Gábor A, Gácsi M, Faragó T, Szabó D, Miklósi Á. Neural mechanisms for lexical processing in dogs. Science. 2016;353:1030–1032. doi: 10.1126/science.aaf3777. [DOI] [PubMed] [Google Scholar]
- 75.Andersen P, Morris R, Amaral D, Bliss T, O’Keefe J. The hippocampus book. Oxford: Oxford University Press; 2006. [Google Scholar]
- 76.Shinohara Y, Hosoya A, Yahagi K, Ferecskó AS, Yaguchi K, Sík A, Itakura M, Takahashi M, Hirase H. Hippocampal CA3 and CA2 have distinct bilateral innervation patterns to CA1 in rodents. Eur J Neurosci. 2012;35:702–710. doi: 10.1111/j.1460-9568.2012.07993.x. [DOI] [PubMed] [Google Scholar]
- 77.van Groen T, Miettinen P, Kadish I. The entorhinal cortex of the mouse: organization of the projection to the hippocampal formation. Hippocampus. 2003;13:133–149. doi: 10.1002/hipo.10037. [DOI] [PubMed] [Google Scholar]
- 78.Kawakami R, Shinohara Y, Kato Y, Sugiyama H, Shigemoto R, Ito I. Asymmetrical allocation of NMDA receptor ε2 subunits in hippocampal circuitry. Science. 2003;300:990–994. doi: 10.1126/science.1082609. [DOI] [PubMed] [Google Scholar]
- 79.Shinohara Y, Hirase H, Watanabe M, Itakura M, Takahashi M, Shigemoto R. Left–right asymmetry of the hippocampal synapses with differential subunit allocation of glutamate receptors. Proc Natl Acad Sci USA. 2008;105:19487–19502. doi: 10.1073/pnas.0807461105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Shinohara Y, Hirase H. Size and receptor density of glutamatergic synapses: a viewpoint from left–right asymmetry of CA3–CA1 connections. Front Neuroanat. 2009;3:10. doi: 10.3389/neuro.05.010.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Kawakami R, Dobi A, Shigemoto R, Ito I. Right isomerism of the brain in inversus viscerum mutant mice. PLoS One. 2008;3:1945. doi: 10.1371/journal.pone.0001945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Supp DM, Witte DP, Potter SS, Brueckner M. Mutation of an axonemal dynein affects left–right asymmetry in inversus viscerum mice. Nature. 1997;389:963–966. doi: 10.1038/40140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Collins RL. On the inheritance of handedness. I. Laterality in inbred mice. J Hered. 1968;59:9–12. doi: 10.1093/oxfordjournals.jhered.a107656. [DOI] [PubMed] [Google Scholar]
- 84.Shinohara Y, Hosoya A, Hirase H. Experience enhances gamma oscillations and formation of interhemispheric asymmetry in the hippocampus. Nat Commun. 2013;4:1652. doi: 10.1038/ncomms2658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Buzsáki G. Hippocampal sharp waves: their origin and significance. Brain Res. 1986;398:242–252. doi: 10.1016/0006-8993(86)91483-6. [DOI] [PubMed] [Google Scholar]
- 86.Bragin A, Jandó G, Nádasdy Z, Hetke J, Wise K, Buzsáki G. Gamma (40–100 Hz) oscillation in the hippocampus of the behaving rat. J Neurosci. 1995;15:47–60. doi: 10.1523/JNEUROSCI.15-01-00047.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Fell J, Axmacher N. The role of phase synchronization in memory processes. Nat Rev Neurosci. 2011;12:105–118. doi: 10.1038/nrn2979. [DOI] [PubMed] [Google Scholar]
- 88.Colgin LL, Denninger T, Fyhn M, Hafting T, Bonnevie T, Jensen O, Moser MB, Moser EI. Frequency of gamma oscillations routes flow of information in the hippocampus. Nature. 2009;462:353–357. doi: 10.1038/nature08573. [DOI] [PubMed] [Google Scholar]
- 89.Montgomery SM, Buzsáki G. Gamma oscillations dynamically couple hippocampal CA3 and CA1 regions during memory task performance. Proc Natl Acad Sci USA. 2007;104:14495–14500. doi: 10.1073/pnas.0701826104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Kemere C, Carr MF, Karlsson MP, Frank LM. Rapid and continuous modulation of hippocampal network state during exploration of new places. PLoS One. 2013;8:e73114. doi: 10.1371/journal.pone.0073114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Bieri KW, Bobbitt KN, Colgin LL. Slow and fast gamma rhythms coordinate different spatial coding modes in hippocampal place cells. Neuron. 2014;82:670–681. doi: 10.1016/j.neuron.2014.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Yamamoto J, Suh J, Takeuchi D, Tonegawa S. Successful execution of working memory linked to synchronized high-frequency gamma oscillations. Cell. 2014;157:845–857. doi: 10.1016/j.cell.2014.04.009. [DOI] [PubMed] [Google Scholar]
- 93.Chen Z, Resnik E, McFarland JM, Sakmann B, Mehta MR. Speed controls the amplitude and timing of the hippocampal gamma rhythm. PLoS One. 2011;6:e21408. doi: 10.1371/journal.pone.0021408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Bartos M, Vida I, Jonas P. Synaptic mechanisms of synchronized gamma oscillations in inhibitory interneuron networks. Nat Rev Neurosci. 2007;8:45–56. doi: 10.1038/nrn2044. [DOI] [PubMed] [Google Scholar]
- 95.Kitanishi T, Ujita S, Fallahnezhad M, Kitanishi N, Ikegaya Y, Tashiro A. Novelty-induced phase-locked firing to slow gamma oscillations in the hippocampus: requirement of synaptic plasticity. Neuron. 2015;86:1265–1276. doi: 10.1016/j.neuron.2015.05.012. [DOI] [PubMed] [Google Scholar]
- 96.Hayashi Y, Shi SH, Esteban JA, Piccini A, Poncer JC, Malinow R. Driving AMPA receptors into synapses by LTP and CaMKII: requirement for GluR1 and PDZ domain interaction. Science. 2000;287:2262–2267. doi: 10.1126/science.287.5461.2262. [DOI] [PubMed] [Google Scholar]
- 97.Shi S, Hayashi Y, Esteban JA, Malinow R. Subunit-specific rules governing AMPA receptor trafficking to synapses in hippocampal pyramidal neurons. Cell. 2001;105:331–343. doi: 10.1016/S0092-8674(01)00321-X. [DOI] [PubMed] [Google Scholar]
- 98.Belluscio MA, Mizuseki K, Schmidt R, Kempter R, Buzsáki G. Cross-frequency phase–phase coupling between theta and gamma oscillations in the hippocampus. J Neurosci. 2012;32:423–435. doi: 10.1523/JNEUROSCI.4122-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Schomburg EW, Fernández-Ruiz A, Mizuseki K, Berényi A, Anastassiou CA, Koch C, Buzsáki G. Theta phase segregation of input-specific gamma patterns in entorhinal–hippocampal networks. Neuron. 2014;84:470–485. doi: 10.1016/j.neuron.2014.08.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Wilson MA, McNaughton BL. Dynamics of the hippocampal ensemble code for space. Science. 1993;261:1055–1058. doi: 10.1126/science.8351520. [DOI] [PubMed] [Google Scholar]
- 101.Thompson LT, Best PJ. Long-term stability of the place-field activity of single units recorded from the dorsal hippocampus of freely behaving rats. Brain Res. 1990;509:299–308. doi: 10.1016/0006-8993(90)90555-P. [DOI] [PubMed] [Google Scholar]
- 102.Ziv Y, Burns LD, Cocker ED, Hamel EO, Ghosh KK, Kitch LJ, El Gamal A, Schnitzer MJ. Long-term dynamics of CA1 hippocampal place codes. Nat Neurosci. 2013;16:264–266. doi: 10.1038/nn.3329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Markus EJ, Qin YL, Leonard B, Skaggs WE, McNaughton BL, Bames CA. Interactions between location and task affect the spatial and directional firing of hippocampal neurons. J Neurosci. 1995;15:7079–7094. doi: 10.1523/JNEUROSCI.15-11-07079.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Dupret D, O’Neill J, Pleydell-Bouverie B, Csicsvari J. The reorganization and reactivation of hippocampal maps predict spatial memory performance. Nat Neurosci. 2010;13:995–1002. doi: 10.1038/nn.2599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Breese CR, Hampson RE, Deadwyler SA. Hippocampal place cells: stereotypy and plasticity. J Neurosci. 1989;9:1097–1111. doi: 10.1523/JNEUROSCI.09-04-01097.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Kobayashi T, Nishijo H, Fukuda M, Bures J, Ono T. Task-dependent representations in rat hippocampal place neurons. J Neurophysiol. 1997;78:597–613. doi: 10.1152/jn.1997.78.2.597. [DOI] [PubMed] [Google Scholar]
- 107.Hollup SA, Molden S, Donnett JG, Moser MB, Moser EI. Accumulation of hippocampal place fields at the goal location in an annular watermaze task. J Neurosci. 2001;21:1635–1644. doi: 10.1523/JNEUROSCI.21-05-01635.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Hok V, Lenck-Santini PP, Roux S, Save E, Muller RU, Poucet B. Goal-related activity in hippocampal place cells. J Neurosci. 2007;27:472–482. doi: 10.1523/JNEUROSCI.2864-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Trullier O, Shibata R, Mulder AB, Wiener SI. Hippocampal neuronal position selectivity remains fixed to room cues only in rats alternating between place navigation and beacon approach tasks. Eur J Neurosci. 1999;11:4381–4388. doi: 10.1046/j.1460-9568.1999.00839.x. [DOI] [PubMed] [Google Scholar]
- 110.Speakman A, O’Keefe J. Hippocampal complex spike cells do not change their place fields if the goal is moved within a cue controlled environment. Eur J Neurosci. 1990;2:544–555. doi: 10.1111/j.1460-9568.1990.tb00445.x. [DOI] [PubMed] [Google Scholar]
- 111.O’Keefe J. Place units in the hippocampus of the freely moving rat. Exp Neurol. 1976;51:78–109. doi: 10.1016/0014-4886(76)90055-8. [DOI] [PubMed] [Google Scholar]
- 112.Zinyuk L, Kubik S, Kaminsky Y, Fenton AA, Bures J. Understanding hippocampal activity by using purposeful behavior: place navigation induces place cell discharge in both task-relevant and task-irrelevant spatial reference frames. Proc Natl Acad Sci USA. 2000;97:3771–3776. doi: 10.1073/pnas.97.7.3771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Euston DR, Gruber AJ, McNaughton BL. The role of medial prefrontal cortex in memory and decision making. Neuron. 2012;76:1057–1070. doi: 10.1016/j.neuron.2012.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Hok V, Chah E, Save E, Poucet B. Prefrontal cortex focally modulates hippocampal place cell firing patterns. J Neurosci. 2013;33:3443–3451. doi: 10.1523/JNEUROSCI.3427-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Schultz W. Predictive reward signal of dopamine neurons. J Neurophysiol. 1998;80:1–27. doi: 10.1152/jn.1998.80.1.1. [DOI] [PubMed] [Google Scholar]
- 116.Scatton B, Simon H, Le Moal M, Bischoff S. Origin of dopaminergic innervation of the rat hippocampal formation. Neurosci Lett. 1980;18:125–131. doi: 10.1016/0304-3940(80)90314-6. [DOI] [PubMed] [Google Scholar]
- 117.Lisman JE, Grace AA. The hippocampal-VTA loop: controlling the entry of information into long-term memory. Neuron. 2005;46:703–713. doi: 10.1016/j.neuron.2005.05.002. [DOI] [PubMed] [Google Scholar]
- 118.McNamara CG, Tejero-Cantero A, Trouche S, Campo-Urriza N, Dupret D. Dopaminergic neurons promote hippocampal reactivation and spatial memory persistence. Nat Neurosci. 2014;17:1658–1660. doi: 10.1038/nn.3843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Tabuchi K, Blundell J, Etherton MR, Hammer RE, Liu X, Powell CM, Südhof TC. A neuroligin-3 mutation implicated in autism increases inhibitory synaptic transmission in mice. Science. 2007;318:71–76. doi: 10.1126/science.1146221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Sigurdsson T, Stark KL, Karayiorgou M, Gogos JA, Gordon JA. Impaired hippocampal-prefrontal synchrony in a genetic mouse model of schizophrenia. Nature. 2010;464:763–767. doi: 10.1038/nature08855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Hayashi Y, Sawa A, Hikida T. Impaired hippocampal activity at the goal zone on the place preference task in a DISC1 mouse model. Neurosci Res. 2016;106:70–73. doi: 10.1016/j.neures.2015.10.005. [DOI] [PubMed] [Google Scholar]
- 122.St Clair D, Blackwood D, Muir W, Carothers A, Walker M, Spowart G, Gosden C, Evans HJ. Association within a family of a balanced autosomal translocation with major mental illness. Lancet. 1990;336:13–16. doi: 10.1016/0140-6736(90)91520-K. [DOI] [PubMed] [Google Scholar]
- 123.Millar JK, Wilson-Annan JC, Anderson S, Christie S, Taylor MS, Semple CA, Devon RS, St Clair DM, Muir WJ, Blackwood DH, Porteous DJ. Disruption of two novel genes by a translocation co-segregating with schizophrenia. Hum Mol Genet. 2000;9:1415–1423. doi: 10.1093/hmg/9.9.1415. [DOI] [PubMed] [Google Scholar]
- 124.Blackwood DH, Fordyce A, Walker MT, St Clair DM, Porteous DJ, Muir WJ. Schizophrenia and affective disorders–cosegregation with a translocation at chromosome 1q42 that directly disrupts brain-expressed genes: clinical and P300 findings in a family. Am J Hum Genet. 2001;69:428–433. doi: 10.1086/321969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Rossier J, Kaminsky Y, Schenk F, Bures J. The place preference task: a new tool for studying the relation between behavior and place cell activity in rats. Behav Neurosci. 2000;114:273–284. doi: 10.1037/0735-7044.114.2.273. [DOI] [PubMed] [Google Scholar]
- 126.Niwa M, Jaaro-Peled H, Tankou S, Seshadri S, Hikida T, Matsumoto Y, Cascella NG, Kano S, Ozaki N, Nabeshima T, Sawa A. Adolescent stress-induced epigenetic control of dopaminergic neurons via glucocorticoids. Science. 2013;339:335–339. doi: 10.1126/science.1226931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Buzsáki G, Logothetis N, Singer W. Scaling brain size, keeping timing: evolutionary preservation of brain rhythms. Neuron. 2013;80:751–764. doi: 10.1016/j.neuron.2013.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Girardeau G, Benchenane K, Wiener SI, Buzsáki G, Zugaro MB. Selective suppression of hippocampal ripples impairs spatial memory. Nat Neurosci. 2009;12:1222–1223. doi: 10.1038/nn.2384. [DOI] [PubMed] [Google Scholar]
- 129.Ego-Stengel V, Wilson MA. Disruption of ripple-associated hippocampal activity during rest impairs spatial learning in the rat. Hippocampus. 2010;20:1–10. doi: 10.1002/hipo.20707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Suh J, Foster DJ, Davoudi H, Wilson MA, Tonegawa S. Impaired hippocampal ripple associated replay in a mouse model of schizophrenia. Neuron. 2013;80:484–493. doi: 10.1016/j.neuron.2013.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Miyakawa T, Leiter LM, Gerber DJ, Gainetdinov RR, Sotnikova TD, Zeng H, Caron MG, Tonegawa S. Conditional calcineurin knockout mice exhibit multiple abnormal behaviors related to schizophrenia. Proc Natl Acad Sci USA. 2003;100:8987–8992. doi: 10.1073/pnas.1432926100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Hikida T, Hikida T, Jaaro-Peled H, Seshadri S, Oishi K, Hookway C, Kong S, Wu D, Xue R, Andrade M, Tankou S, Mori S, Gallagher M, Ishizuka K, Pletnikov M, Kida S, Sawa A. Dominant-negative disc1 transgenic mice display schizophrenia-associated phenotypes detected by measures translatable to humans. Proc Natl Acad Sci USA. 2007;104:14501–14506. doi: 10.1073/pnas.0704774104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Altimus C, Harrold J, Jaaro-Peled H, Sawa A, Foster DJ. Disordered ripples are a common feature of genetically distinct mouse models relevant to schizophrenia. Mol Neuropsychiatry. 2015;1:52–59. doi: 10.1159/000380765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Engel AK, Fries P, Singer W. Dynamic predictions: oscillations and synchrony in top-down processing. Nat Rev Neurosci. 2001;2:704–716. doi: 10.1038/35094565. [DOI] [PubMed] [Google Scholar]
- 135.Muller M, Wehner R. Path integration in desert ants, Cataglyphis fortis . Proc Natl Acad Sci USA. 1988;85:5287–5290. doi: 10.1073/pnas.85.14.5287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Wittlinger M, Wehner R, Wolf H. The ant odometer: stepping on stilts and stumps. Science. 2006;312:1965–1967. doi: 10.1126/science.1126912. [DOI] [PubMed] [Google Scholar]
- 137.Walcott C. Pigeon homing: observations, experiments and confusions. J Exp Biol. 1996;199:21–27. doi: 10.1242/jeb.199.1.21. [DOI] [PubMed] [Google Scholar]
- 138.Lohmann KJ, Lohmann CM, Putman NF. Magnetic maps in animals: nature’s GPS. J Exp Biol. 2007;210:3697–3705. doi: 10.1242/jeb.001313. [DOI] [PubMed] [Google Scholar]
- 139.Wu LQ, Dickman JD. Neural correlates of a magnetic sense. Science. 2012;336:1054–1057. doi: 10.1126/science.1216567. [DOI] [PubMed] [Google Scholar]
- 140.Tolman EC. Cognitive maps in rats and men. Psychol Rev. 1948;55:189–208. doi: 10.1037/h0061626. [DOI] [PubMed] [Google Scholar]
- 141.Morris RG. Spatial localization does not require the presence of local cues. Learn Motiv. 1981;12:239–260. doi: 10.1016/0023-9690(81)90020-5. [DOI] [Google Scholar]
- 142.Eichenbaum H, Stewart C, Morris RG. Hippocampal representation in place learning. J Neurosci. 1990;10:3531–3542. doi: 10.1523/JNEUROSCI.10-11-03531.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Wood ER, Dudchenko PA, Robitsek RJ, Eichenbaum H. Hippocampal neurons encode information about different types of memory episodes occurring in the same location. Neuron. 2000;27:623–633. doi: 10.1016/S0896-6273(00)00071-4. [DOI] [PubMed] [Google Scholar]
- 144.Frank LM, Brown EN, Wilson M. Trajectory encoding in the hippocampus and entorhinal cortex. Neuron. 2000;27:169–178. doi: 10.1016/S0896-6273(00)00018-0. [DOI] [PubMed] [Google Scholar]
- 145.Ferbinteanu J, Shapiro ML. Prospective and retrospective memory coding in the hippocampus. Neuron. 2003;40:1227–1239. doi: 10.1016/S0896-6273(03)00752-9. [DOI] [PubMed] [Google Scholar]
- 146.Johnson A, Redish AD. Neural ensembles in CA3 transiently encode paths forward of the animal at a decision point. J Neurosci. 2007;27:12176–12189. doi: 10.1523/JNEUROSCI.3761-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Pfeiffer BE, Foster DJ. Hippocampal place-cell sequences depict future paths to remembered goals. Nature. 2013;497:74–79. doi: 10.1038/nature12112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Ito HT, Zhang SJ, Witter MP, Moser EI, Moser MB. A prefrontal-thalamo-hippocampal circuit for goal-directed spatial navigation. Nature. 2015;522:50–55. doi: 10.1038/nature14396. [DOI] [PubMed] [Google Scholar]
- 149.Tanji J, Shima K, Mushiake H. Concept-based behavioral planning and the lateral prefrontal cortex. Trends Cogn Sci. 2007;11:528–534. doi: 10.1016/j.tics.2007.09.007. [DOI] [PubMed] [Google Scholar]
- 150.Gold JI, Shadlen MN. The neural basis of decision making. Annu Rev Neurosci. 2007;30:535–574. doi: 10.1146/annurev.neuro.29.051605.113038. [DOI] [PubMed] [Google Scholar]
- 151.Sul JH, Kim H, Huh N, Lee D, Jung MW. Distinct roles of rodent orbitofrontal and medial prefrontal cortex in decision making. Neuron. 2010;66:449–460. doi: 10.1016/j.neuron.2010.03.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Herkenham M. The connections of the nucleus reuniens thalami: evidence for a direct thalamo-hippocampal pathway in the rat. J Comp Neurol. 1978;177:589–610. doi: 10.1002/cne.901770405. [DOI] [PubMed] [Google Scholar]
- 153.Wouterlood FG, Saldana E, Witter MP. Projection from the nucleus reuniens thalami to the hippocampal region: light and electron microscopic tracing study in the rat with the anterograde tracer Phaseolus vulgaris-leucoagglutinin. J Comp Neurol. 1990;296:179–203. doi: 10.1002/cne.902960202. [DOI] [PubMed] [Google Scholar]
- 154.Vertes RP, Hoover WB, Szigeti-Buck K, Leranth C. Nucleus reuniens of the midline thalamus: link between the medial prefrontal cortex and the hippocampus. Brain Res Bull. 2007;71:601–609. doi: 10.1016/j.brainresbull.2006.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Hok V, Save E, Lenck-Santini PP, Poucet B. Coding for spatial goals in the prelimbic/infralimbic area of the rat frontal cortex. Proc Natl Acad Sci USA. 2005;102:4602–4607. doi: 10.1073/pnas.0407332102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Howe MW, Tierney PL, Sandberg SG, Phillips PE, Graybiel AM. Prolonged dopamine signalling in striatum signals proximity and value of distant rewards. Nature. 2013;500:575–579. doi: 10.1038/nature12475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Moser EI, Kropff E, Moser MB. Place cells, grid cells, and the brain’s spatial representation system. Annu Rev Neurosci. 2008;31:69–89. doi: 10.1146/annurev.neuro.31.061307.090723. [DOI] [PubMed] [Google Scholar]
- 158.ShinoharaY Hosoya A, Yamasaki N, Ahmed H, Hattori S, Eguchi M, Yamaguchi S, Miyakawa T, Hirase H, Shigemoto R. Right-hemispheric dominance of spatial memory in split-brain mice. Hippocampus. 2012;22:117–121. doi: 10.1002/hipo.20886. [DOI] [PubMed] [Google Scholar]
- 159.Maguire EA, Burgess N, Donnett JG, Frackowiak RS, Frith CD, O’Keefe J. Knowing where and getting there: a human navigation network. Science. 1998;280:921–924. doi: 10.1126/science.280.5365.921. [DOI] [PubMed] [Google Scholar]
- 160.Ekstrom AD, Kahana MJ, Caplan JB, Fields TA, Isham EA, Newman EL, Fried I. Cellular networks underlying human spatial navigation. Nature. 2003;425:184–188. doi: 10.1038/nature01964. [DOI] [PubMed] [Google Scholar]